b cell epitopes - cbs · b cell epitopes tuesday, 9 june 2009. technical university of denmark -...
TRANSCRIPT

Technical University of Denmark - DTUDepartment of systems biology
CE
NT
ER
FOR
BIO
LOG
ICA
L SE
QU
EN
CE
AN
ALY
SIS
B cell Epitopes
Tuesday, 9 June 2009

Technical University of Denmark - DTUDepartment of systems biology
CE
NT
ER
FOR
BIO
LOG
ICA
L SE
QU
EN
CE
AN
ALY
SIS
Antibody Effect
Virus or Toxin Antibodies
Tuesday, 9 June 2009

Technical University of Denmark - DTUDepartment of systems biology
CE
NT
ER
FOR
BIO
LOG
ICA
L SE
QU
EN
CE
AN
ALY
SIS
Antibody Effect
Virus or Toxin Antibodies
Tuesday, 9 June 2009

Technical University of Denmark - DTUDepartment of systems biology
CE
NT
ER
FOR
BIO
LOG
ICA
L SE
QU
EN
CE
AN
ALY
SIS
Antibodies
Tuesday, 9 June 2009

Technical University of Denmark - DTUDepartment of systems biology
CE
NT
ER
FOR
BIO
LOG
ICA
L SE
QU
EN
CE
AN
ALY
SIS
Antibodies
Tuesday, 9 June 2009

Technical University of Denmark - DTUDepartment of systems biology
CE
NT
ER
FOR
BIO
LOG
ICA
L SE
QU
EN
CE
AN
ALY
SIS
Antibody - Antigen interactionAntigen
Antibody
Tuesday, 9 June 2009

Technical University of Denmark - DTUDepartment of systems biology
CE
NT
ER
FOR
BIO
LOG
ICA
L SE
QU
EN
CE
AN
ALY
SIS
Antibody - Antigen interaction
Fab
Antigen
Antibody
Tuesday, 9 June 2009

Technical University of Denmark - DTUDepartment of systems biology
CE
NT
ER
FOR
BIO
LOG
ICA
L SE
QU
EN
CE
AN
ALY
SIS
Antibody - Antigen interaction
Fab
Antigen
Epitope
Antibody
Tuesday, 9 June 2009

Technical University of Denmark - DTUDepartment of systems biology
CE
NT
ER
FOR
BIO
LOG
ICA
L SE
QU
EN
CE
AN
ALY
SIS
Antibody - Antigen interaction
Fab
Antigen
Paratope
Antibody
Tuesday, 9 June 2009
Technical University of Denmark - DTUDepartment of systems biology
CE
NT
ER
FOR
BIO
LOG
ICA
L SE
QU
EN
CE
AN
ALY
SIS
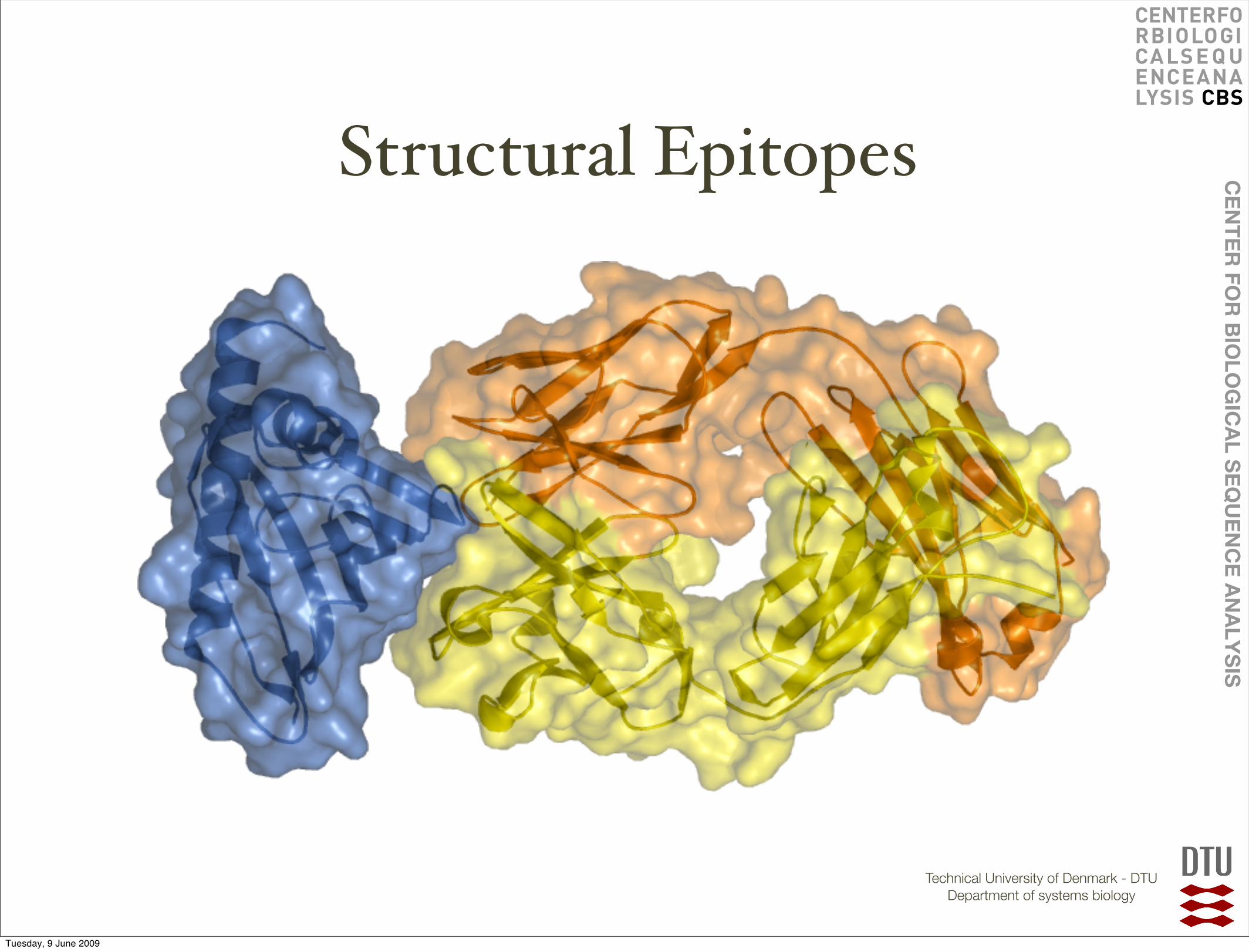
Structural Epitopes
Tuesday, 9 June 2009

Technical University of Denmark - DTUDepartment of systems biology
CE
NT
ER
FOR
BIO
LOG
ICA
L SE
QU
EN
CE
AN
ALY
SIS
Discontinuous Epitopes
Tuesday, 9 June 2009

Technical University of Denmark - DTUDepartment of systems biology
CE
NT
ER
FOR
BIO
LOG
ICA
L SE
QU
EN
CE
AN
ALY
SIS
Discontinuous Epitopes
Tuesday, 9 June 2009

Technical University of Denmark - DTUDepartment of systems biology
CE
NT
ER
FOR
BIO
LOG
ICA
L SE
QU
EN
CE
AN
ALY
SIS
Discontinuous Paratopes
Tuesday, 9 June 2009

Technical University of Denmark - DTUDepartment of systems biology
CE
NT
ER
FOR
BIO
LOG
ICA
L SE
QU
EN
CE
AN
ALY
SIS
Variable regionsAlpha-carbon trace of the structure of the heavy chain and
light chain variable regions of a typical antibody. The framework regions of both chains are shown in grey whilst
the complementarity determining regions (CDRs) are coloured individually, i.e.
Heavy chainCDR 1 = Light blue
CDR 2 = CeriseCDR 3 = Yellow
Light ChainCDR 1 = Red
CDR 2 = GreenCDR 3 = Blue
CDR RegionsCDR = complementarity determining region
http://212.219.234.139/html/anti_alpha.html
Tuesday, 9 June 2009

Technical University of Denmark - DTUDepartment of systems biology
CE
NT
ER
FOR
BIO
LOG
ICA
L SE
QU
EN
CE
AN
ALY
SIS
B-Cells
B-lymphocytes each displaying a unique B-cell receptor
Stem Cell
Precurser B-lymphocytes
Gene rearrangements
Tuesday, 9 June 2009
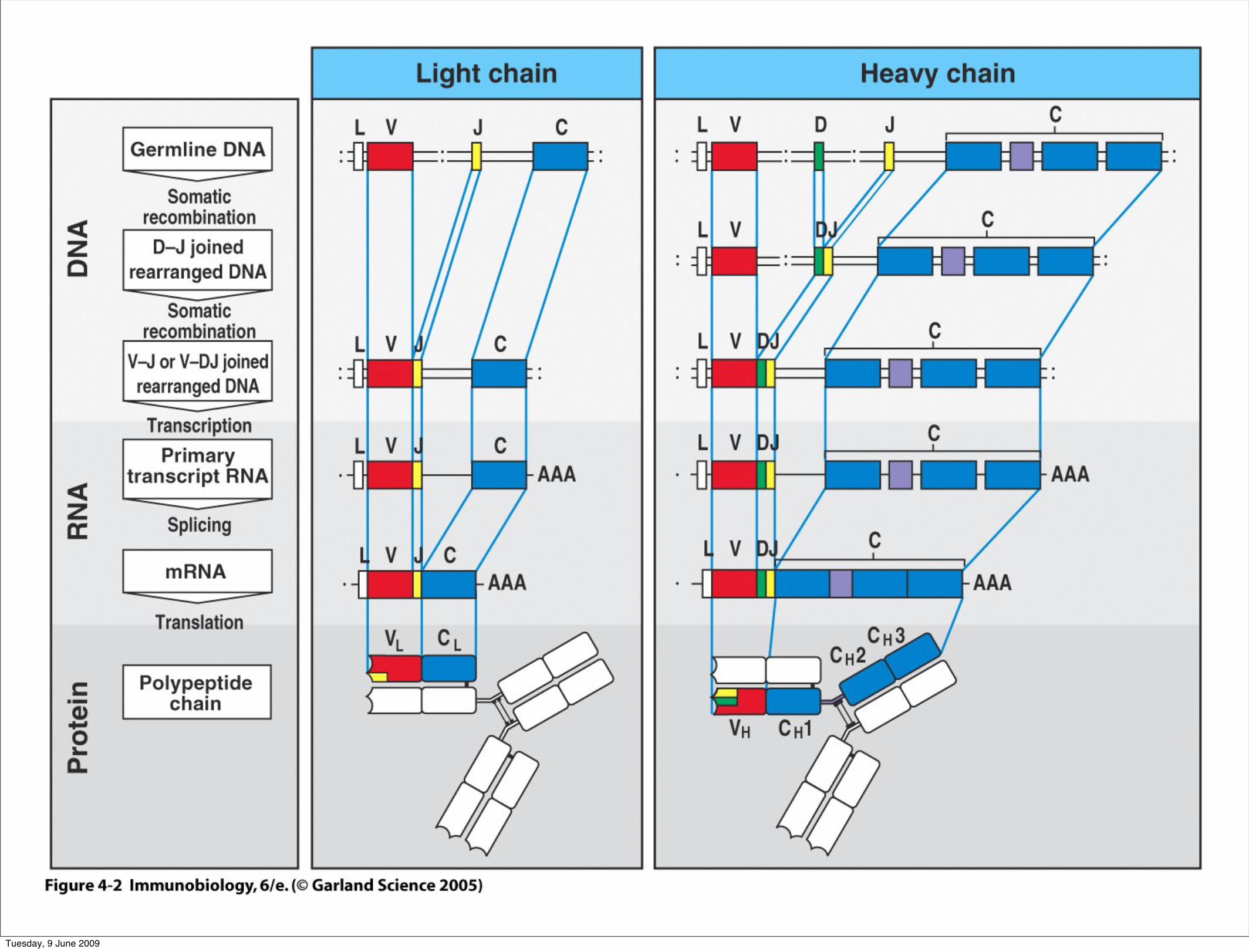
Figure 4-2
Tuesday, 9 June 2009

Figure 4-3
Tuesday, 9 June 2009

Figure 4-4
Tuesday, 9 June 2009
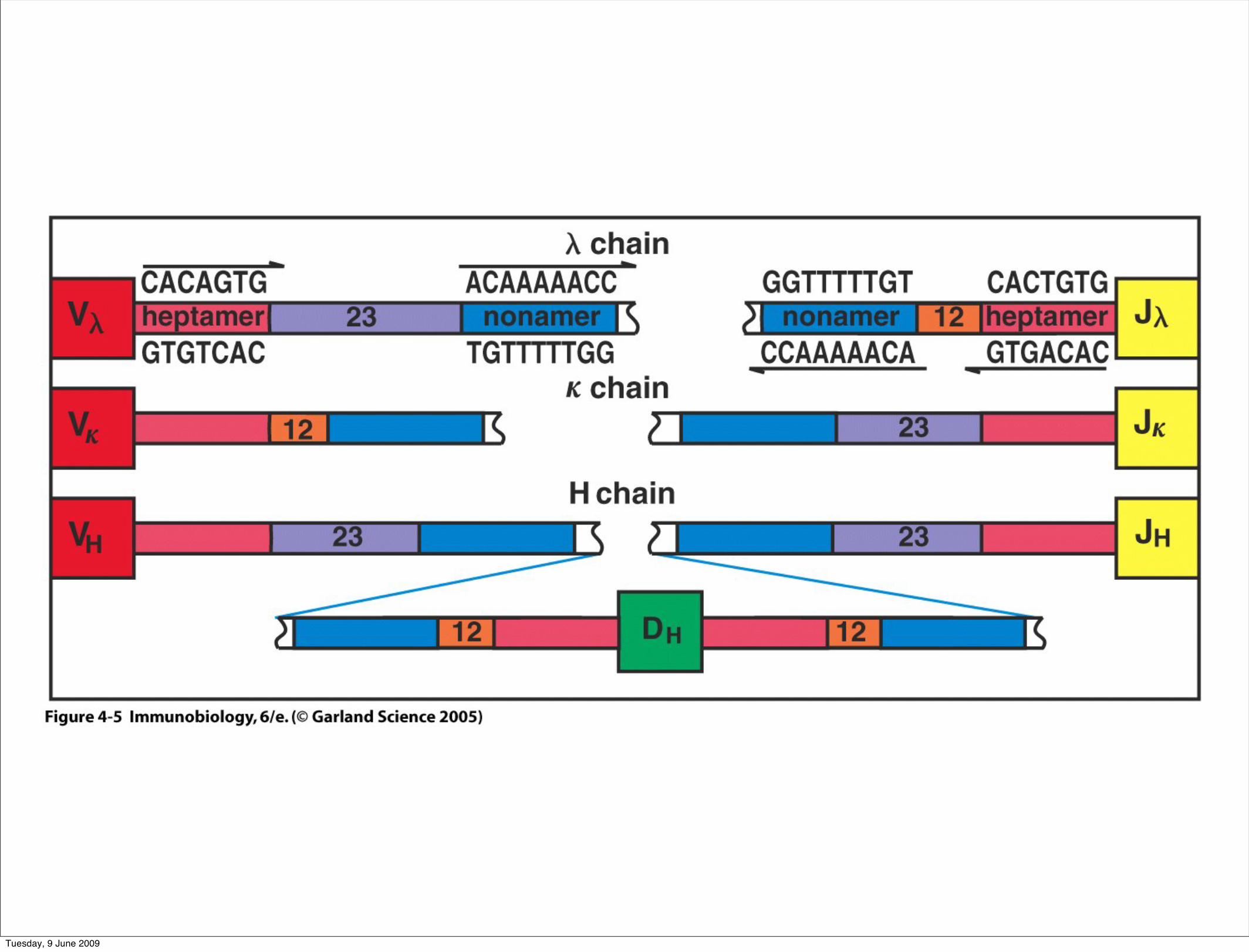
Figure 4-5
Tuesday, 9 June 2009

Figure 4-6
Tuesday, 9 June 2009

Technical University of Denmark - DTUDepartment of systems biology
CE
NT
ER
FOR
BIO
LOG
ICA
L SE
QU
EN
CE
AN
ALY
SIS
P-nucleotide introduction
Tuesday, 9 June 2009

Technical University of Denmark - DTUDepartment of systems biology
CE
NT
ER
FOR
BIO
LOG
ICA
L SE
QU
EN
CE
AN
ALY
SIS
N-nucleotide introduction
Tuesday, 9 June 2009

Technical University of Denmark - DTUDepartment of systems biology
CE
NT
ER
FOR
BIO
LOG
ICA
L SE
QU
EN
CE
AN
ALY
SIS
Somatic Hypermutations
© 2001 by Garland Publishing
Tuesday, 9 June 2009

Technical University of Denmark - DTUDepartment of systems biology
CE
NT
ER
FOR
BIO
LOG
ICA
L SE
QU
EN
CE
AN
ALY
SIS
B-Cell ActivationNo AffinityLow AffinityNo Affinity
High Affinity
Somatic Hypermutations
Memory B-cells
Plasma cells
Tuesday, 9 June 2009

Technical University of Denmark - DTUDepartment of systems biology
CE
NT
ER
FOR
BIO
LOG
ICA
L SE
QU
EN
CE
AN
ALY
SIS
B-Cell Activation
B Cell
T Helper Cell
Class II MHC
Bound Peptide
TCR
Tuesday, 9 June 2009

Technical University of Denmark - DTUDepartment of systems biology
CE
NT
ER
FOR
BIO
LOG
ICA
L SE
QU
EN
CE
AN
ALY
SIS
Cartoon by Eric Reits
Tuesday, 9 June 2009

Technical University of Denmark - DTUDepartment of systems biology
CE
NT
ER
FOR
BIO
LOG
ICA
L SE
QU
EN
CE
AN
ALY
SIS
Monoclonal Abs
•Hybridomas
•Phage Display
Tuesday, 9 June 2009

Technical University of Denmark - DTUDepartment of systems biology
CE
NT
ER
FOR
BIO
LOG
ICA
L SE
QU
EN
CE
AN
ALY
SIS
Hybridomas
http://www.britannica.com/
Tuesday, 9 June 2009

Technical University of Denmark - DTUDepartment of systems biology
CE
NT
ER
FOR
BIO
LOG
ICA
L SE
QU
EN
CE
AN
ALY
SIS
Phage Display
Tuesday, 9 June 2009

Technical University of Denmark - DTUDepartment of systems biology
CE
NT
ER
FOR
BIO
LOG
ICA
L SE
QU
EN
CE
AN
ALY
SIS
Phage Display
http://www.bio.anl.gov/highthroughputbiology/images/phage_display1.jpg
Tuesday, 9 June 2009

Technical University of Denmark - DTUDepartment of systems biology
CE
NT
ER
FOR
BIO
LOG
ICA
L SE
QU
EN
CE
AN
ALY
SIS
Identification of germ-line genes
Why?
As bookkeeping and reporting tool
For detection of potentially important residues in binding affinity.
Tuesday, 9 June 2009

Technical University of Denmark - DTUDepartment of systems biology
CE
NT
ER
FOR
BIO
LOG
ICA
L SE
QU
EN
CE
AN
ALY
SIS
Joinsolver
Tuesday, 9 June 2009

Technical University of Denmark - DTUDepartment of systems biology
CE
NT
ER
FOR
BIO
LOG
ICA
L SE
QU
EN
CE
AN
ALY
SIS
Vbase
Tuesday, 9 June 2009

Technical University of Denmark - DTUDepartment of systems biology
CE
NT
ER
FOR
BIO
LOG
ICA
L SE
QU
EN
CE
AN
ALY
SIS
Vquest
Tuesday, 9 June 2009

Technical University of Denmark - DTUDepartment of systems biology
CE
NT
ER
FOR
BIO
LOG
ICA
L SE
QU
EN
CE
AN
ALY
SIS
VDJsolver
Tuesday, 9 June 2009

Technical University of Denmark - DTUDepartment of systems biology
CE
NT
ER
FOR
BIO
LOG
ICA
L SE
QU
EN
CE
AN
ALY
SIS
Questions to be addressed
•Can multiple D genes be inserted?
•Violation of 12/23 rule
•Can D genes be inserted backwards?
•Is there a D gene preference?
•Is there a reading frame preference for D genes?
•If yes, is it part of the gene rearrangement?
Tuesday, 9 June 2009

Technical University of Denmark - DTUDepartment of systems biology
CE
NT
ER
FOR
BIO
LOG
ICA
L SE
QU
EN
CE
AN
ALY
SIS
Data sets
•6329 clonally unrelated rearrangements.
•1968 un-mutated functional
•3707 mutated functional
•274 un-mutated non-functional
•380 mutated non-functional
Tuesday, 9 June 2009

Technical University of Denmark - DTUDepartment of systems biology
CE
NT
ER
FOR
BIO
LOG
ICA
L SE
QU
EN
CE
AN
ALY
SIS
SequencesSequencesSequences Permutated sequencesPermutated sequencesPermutated sequences
Distance from heptamer to gene end
No. ofseq
No. withP % with P No. of
seqNo. with
P % with P p-value
VH geneVH geneVH geneVH geneVH geneVH geneVH geneVH gene
1 1448 474 32.7 1635 103 6.3 <10-5
2 1027 48 4.7 1068 65 6.1 0.091
3 762 53 7.0 612 36 5.9 0.245
JH geneJH geneJH geneJH geneJH geneJH geneJH geneJH gene
1 324 60 18.5 350 23 6.6 <10-5
2 184 2 1.0 209 3 1.4 0.560
3 219 8 3.7 250 14 5.6 0.220
5’ end of D gene 5’ end of D gene 5’ end of D gene 5’ end of D gene 5’ end of D gene 5’ end of D gene 5’ end of D gene 5’ end of D gene
1 519 128 24.7 619 54 8.7 <10-5
2 343 31 9.0 347 26 7.5 0.275
3 474 25 5.3 454 17 3.7 0.168
3’ end of D gene3’ end of D gene3’ end of D gene3’ end of D gene3’ end of D gene3’ end of D gene3’ end of D gene3’ end of D gene
1 616 86 14.0 684 58 8.5 0.001
2 266 30 11.3 276 24 8.7 0.195
3 460 5 1.1 485 9 1.9 0.241
P nucleotides
Tuesday, 9 June 2009

Technical University of Denmark - DTUDepartment of systems biology
CE
NT
ER
FOR
BIO
LOG
ICA
L SE
QU
EN
CE
AN
ALY
SIS
SequencesSequencesSequences Permutated sequencesPermutated sequencesPermutated sequences
Distance from heptamer to gene end
No. ofseq
No. withP % with P No. of
seqNo. with
P % with P p-value
VH geneVH geneVH geneVH geneVH geneVH geneVH geneVH gene
1 1448 474 32.7 1635 103 6.3 <10-5
2 1027 48 4.7 1068 65 6.1 0.091
3 762 53 7.0 612 36 5.9 0.245
JH geneJH geneJH geneJH geneJH geneJH geneJH geneJH gene
1 324 60 18.5 350 23 6.6 <10-5
2 184 2 1.0 209 3 1.4 0.560
3 219 8 3.7 250 14 5.6 0.220
5’ end of D gene 5’ end of D gene 5’ end of D gene 5’ end of D gene 5’ end of D gene 5’ end of D gene 5’ end of D gene 5’ end of D gene
1 519 128 24.7 619 54 8.7 <10-5
2 343 31 9.0 347 26 7.5 0.275
3 474 25 5.3 454 17 3.7 0.168
3’ end of D gene3’ end of D gene3’ end of D gene3’ end of D gene3’ end of D gene3’ end of D gene3’ end of D gene3’ end of D gene
1 616 86 14.0 684 58 8.5 0.001
2 266 30 11.3 276 24 8.7 0.195
3 460 5 1.1 485 9 1.9 0.241
P nucleotides
Tuesday, 9 June 2009

Technical University of Denmark - DTUDepartment of systems biology
CE
NT
ER
FOR
BIO
LOG
ICA
L SE
QU
EN
CE
AN
ALY
SIS
SequencesSequencesSequences Permutated sequencesPermutated sequencesPermutated sequences
Distance from heptamer to gene end
No. ofseq
No. withP % with P No. of
seqNo. with
P % with P p-value
VH geneVH geneVH geneVH geneVH geneVH geneVH geneVH gene
1 1448 474 32.7 1635 103 6.3 <10-5
2 1027 48 4.7 1068 65 6.1 0.091
3 762 53 7.0 612 36 5.9 0.245
JH geneJH geneJH geneJH geneJH geneJH geneJH geneJH gene
1 324 60 18.5 350 23 6.6 <10-5
2 184 2 1.0 209 3 1.4 0.560
3 219 8 3.7 250 14 5.6 0.220
5’ end of D gene 5’ end of D gene 5’ end of D gene 5’ end of D gene 5’ end of D gene 5’ end of D gene 5’ end of D gene 5’ end of D gene
1 519 128 24.7 619 54 8.7 <10-5
2 343 31 9.0 347 26 7.5 0.275
3 474 25 5.3 454 17 3.7 0.168
3’ end of D gene3’ end of D gene3’ end of D gene3’ end of D gene3’ end of D gene3’ end of D gene3’ end of D gene3’ end of D gene
1 616 86 14.0 684 58 8.5 0.001
2 266 30 11.3 276 24 8.7 0.195
3 460 5 1.1 485 9 1.9 0.241
P nucleotides
Tuesday, 9 June 2009

Technical University of Denmark - DTUDepartment of systems biology
CE
NT
ER
FOR
BIO
LOG
ICA
L SE
QU
EN
CE
AN
ALY
SIS
How many types of D genes?
•Conventional D genes
• Identified in 81% of sequences unmutated sequences, 64% of mutated sequences
•Inverted D genes
•Long inverted D genes can not be excluded
•Two or more D genes
Tuesday, 9 June 2009

Technical University of Denmark - DTUDepartment of systems biology
CE
NT
ER
FOR
BIO
LOG
ICA
L SE
QU
EN
CE
AN
ALY
SIS
D gene usage27 conventional D genes, 34 known alleles
Tuesday, 9 June 2009

Technical University of Denmark - DTUDepartment of systems biology
CE
NT
ER
FOR
BIO
LOG
ICA
L SE
QU
EN
CE
AN
ALY
SIS
D-gene usage and JH gene
•JH proximal D genes more often recombined to JH4 than JH6 and JH distal D genes more often to JH6
Tuesday, 9 June 2009

Technical University of Denmark - DTUDepartment of systems biology
CE
NT
ER
FOR
BIO
LOG
ICA
L SE
QU
EN
CE
AN
ALY
SIS
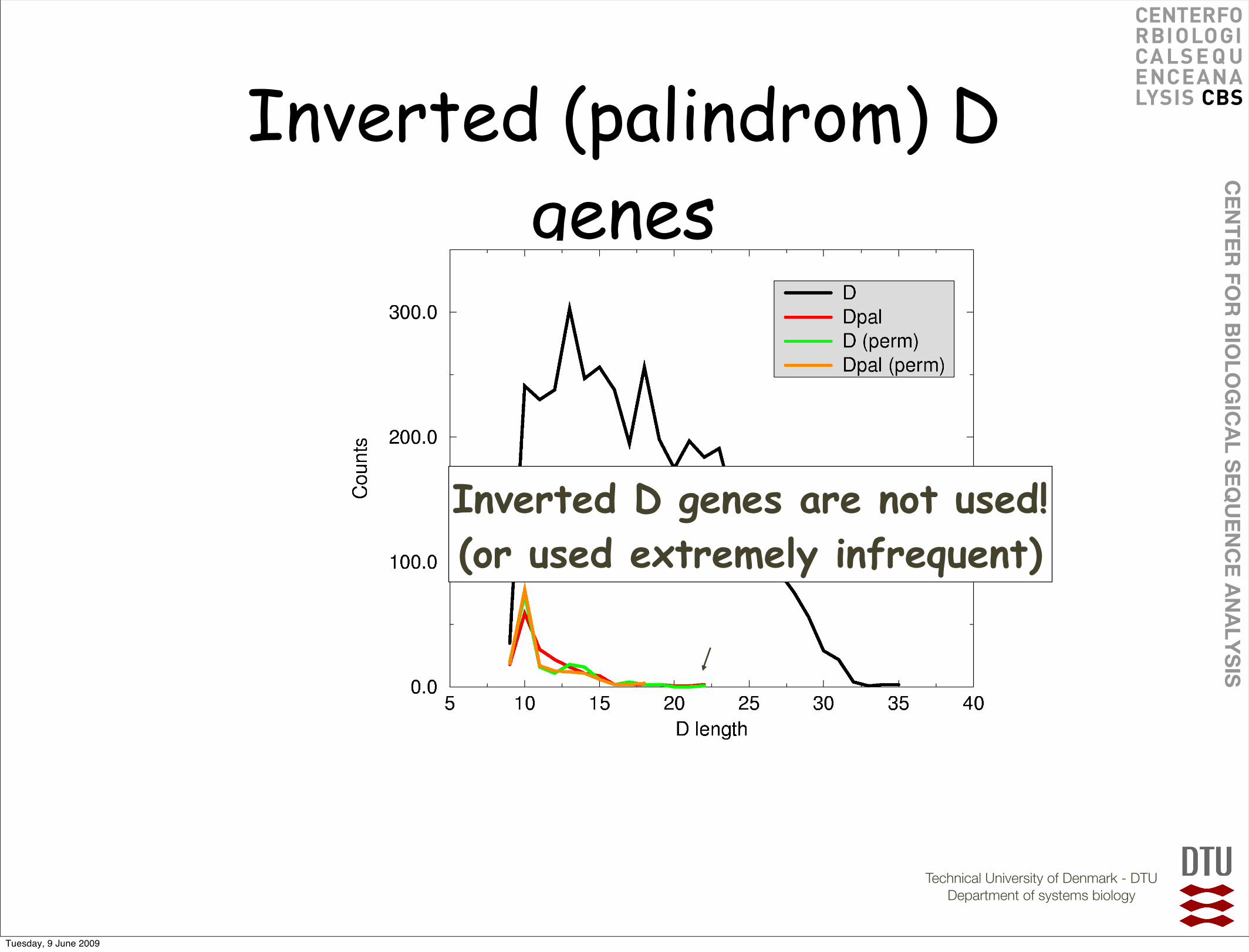
Inverted (palindrom) D genes
Tuesday, 9 June 2009
Technical University of Denmark - DTUDepartment of systems biology
CE
NT
ER
FOR
BIO
LOG
ICA
L SE
QU
EN
CE
AN
ALY
SIS
Inverted (palindrom) D genes
Inverted D genes are not used!(or used extremely infrequent)
Tuesday, 9 June 2009

Technical University of Denmark - DTUDepartment of systems biology
CE
NT
ER
FOR
BIO
LOG
ICA
L SE
QU
EN
CE
AN
ALY
SIS
Multiple D genes
•65 sequences with two D genes
•Average length of shortest D genes: 11.6bp
•Average length of longest D genes: 18.8bp
•Average length of D genes in permuted sequences: 11.3bp
•Average length of D genes in normal sequences: 17.8bp
• => multiple D genes are not present!!!
Longest-D Shortest-DV-gene J-gene
Tuesday, 9 June 2009

Technical University of Denmark - DTUDepartment of systems biology
CE
NT
ER
FOR
BIO
LOG
ICA
L SE
QU
EN
CE
AN
ALY
SIS
D gene reading frames
• The recombination mechanism utilizes each D gene reading frame at same frequency
Reading Frame StopStopStop HydrophilicHydrophilicHydrophilic HydrophobicHydrophobicHydrophobic
Gene P NP P NP P NP
D2-2*01 RIL**YQLLC (1) 6.5 34.7 GYCSSTSCYA (2) 61.2 32.6 DIVVVPAAM (3) 32.2 32.6
D2-2*02 RIL**YQLLY (1) 11.3 46.7 GYCSSTSCYT (2) 55.0 20.0 DIVVVPAAI (3) 33.8 33.3
D2-2*03 WIL**YQLLC (1) 0.0 50.0 GYCSSTSCYA (2) 66.7 50.0 DIVVVPAAM (3) 33.3 0.0
D2-8*01 RILY*WCMLY (1) 2.4 42.9 GYCTNGVCYT (2) 68.3 28.6 DIVLMVYAI (3) 29.3 28.6
D2-8*02 RILYWWCMLY (1) 0.0 0.0 GYCTGGVCYT (2) 88.9 0.0 DIVLVVYAI (3) 11.1 100
D2-15*01 RIL*WW*LLL (1) 1.5 32.5 GYCSGGSCYS (2) 70.8 37.5 DIVVVVAAT (3) 27.7 30.0
D2-21*01 SILWW*LLF (1) 8.3 50.0 AYCGGDCYS (2) 58.3 25.0 HIVVVIAI (3) 33.3 25.0
D2-21*02 SILWW*LLF (1) 0.0 54.5 AYCGGDCYS (2) 78.0 18.2 HIVVVTAI (3) 22.0 27.3
Total - 10.8 33.6 - 62.2 32.4 - 26.9 34.0
Tuesday, 9 June 2009

Technical University of Denmark - DTUDepartment of systems biology
CE
NT
ER
FOR
BIO
LOG
ICA
L SE
QU
EN
CE
AN
ALY
SIS
D gene reading frames
• The recombination mechanism utilizes each D gene reading frame at same frequency
Reading Frame StopStopStop HydrophilicHydrophilicHydrophilic HydrophobicHydrophobicHydrophobic
Gene P NP P NP P NP
D2-2*01 RIL**YQLLC (1) 6.5 34.7 GYCSSTSCYA (2) 61.2 32.6 DIVVVPAAM (3) 32.2 32.6
D2-2*02 RIL**YQLLY (1) 11.3 46.7 GYCSSTSCYT (2) 55.0 20.0 DIVVVPAAI (3) 33.8 33.3
D2-2*03 WIL**YQLLC (1) 0.0 50.0 GYCSSTSCYA (2) 66.7 50.0 DIVVVPAAM (3) 33.3 0.0
D2-8*01 RILY*WCMLY (1) 2.4 42.9 GYCTNGVCYT (2) 68.3 28.6 DIVLMVYAI (3) 29.3 28.6
D2-8*02 RILYWWCMLY (1) 0.0 0.0 GYCTGGVCYT (2) 88.9 0.0 DIVLVVYAI (3) 11.1 100
D2-15*01 RIL*WW*LLL (1) 1.5 32.5 GYCSGGSCYS (2) 70.8 37.5 DIVVVVAAT (3) 27.7 30.0
D2-21*01 SILWW*LLF (1) 8.3 50.0 AYCGGDCYS (2) 58.3 25.0 HIVVVIAI (3) 33.3 25.0
D2-21*02 SILWW*LLF (1) 0.0 54.5 AYCGGDCYS (2) 78.0 18.2 HIVVVTAI (3) 22.0 27.3
Total - 10.8 33.6 - 62.2 32.4 - 26.9 34.0
Tuesday, 9 June 2009

Technical University of Denmark - DTUDepartment of systems biology
CE
NT
ER
FOR
BIO
LOG
ICA
L SE
QU
EN
CE
AN
ALY
SIS
VDJsolver performanceUnmutated sequences
Mutated sequences
#: p<0.01§: P<0.001
Tuesday, 9 June 2009