bacillus -bakteerien tuottamat lipopeptidit...
TRANSCRIPT

BACILLUS -BAKTEERIEN TUOTTAMAT LIPOPEPTIDIT
JAPANILAISESSA SOIJAPOHJAISESSA ELINTARVIKKEESSA
(NATTO)
Mari Juola
Pro gradu -tutkielma
Biotieteiden koulutusohjelma
Biotieteiden pääaine
Itä-Suomen Yliopiston
luonnontieteiden ja metsätieteiden tiedekunta
Helmikuu 2015

ITÄ-SUOMEN YLIOPISTO, Luonnontieteiden ja metsätieteiden tiedekunta
Biotiede
Ravitsemus- ja elintarvikebiotekniikka
JUOLA MARI K.: Bacillus -bakteerien tuottamat lipopeptidit japanilaisessa soijapohjaisessa
elintarvikkeessa (natto)
Pro gradu -tutkielma, 82 sivua, liitteet 4 kpl, 3 sivua
Tutkielman ohjaajat: Prof. Atte von Wright ja prof. Sirpa Kärenlampi
Helmikuu 2015
Avainsanat: Bacillus subtilis, natto, sykliset lipopeptidit, surfaktiinit, sytotoksisuus
Tämän tutkielman tavoitteena oli tuottaa tietoa japanilaisen soijapavuista valmistetun,
fermentoidun elintarvikkeen nimeltä natto elintarviketurvallisuudesta. Tutkimuksessa
selvitettiin fermentoinnin heräteviljelminä käytettyjen Bacillus subtilis var. natto -bakteeri-
kantojen syklisten lipopeptidien tuottokyky ja pitoisuus nattoateriassa yhteistyössä Chr. Hansen
A/G:n kanssa sekä arvioitiin tuotettujen lipopeptidien sytotoksisuutta. Tulosten pohjalta
suoritettiin toksikologinen arvio turvallisesta kerta-annoksesta nattoa.
Bacillus subtilis on hyvin yleinen, harvoin infektioita aiheuttava ympäristöbakteeri, jota
hyödynnetään elintarviketeollisuudessa usein eri tavoin. Toisinaan B. subtilis -löydöksiä
tehdään myös ruokamyrkytyksen aiheuttaneista elintarvikkeista. Lisäksi joidenkin B. subtilis -
bakteerikantojen on havaittu kykenevän tuottamaan aineenvaihduntatuotteenaan syklisiä
lipopeptidejä kuten surfaktiineja, fengysiinejä, plipastatiineja ja mykosubtiliinia.
Lipopeptidien, erityisesti surfaktiinien on todettu olevan hemolyyttisiä ja sytotoksisia, ja
bakteerikantojen käyttöturvallisuus elintarvikeketjussa on kyseenalaistettu. Euroopan
elintarviketurvallisuusvirasto EFSA edellyttääkin B. subtilis -bakteerikantoja sisältävien
elintarvikkeiden toksikologista arviointia ja seurantaa sekä heräteviljelminä käytettyjen
bakteerikantojen identifiointia.
Tutkimuksen kokeellisessa osiossa identifioitiin kolmessa nattoateriassa heräteviljelminä
toimineet mikrobikannat 16S rDNA -sekvenoinnin avulla ja analysoitiin niiden lipopeptidien
synteesi PCR- ja UHPLC-qTOF -tekniikoiden avulla. Lisäksi uutettiin bakteerien tuottamat
mahdolliset lipopeptidit sekä nattoaterioista että isolaateista ja määritettiin niiden
hemolyyttisyys naudan verisolujen ja sytotoksisuus karjun siittiöiden liikkuvuuden estotestin
avulla.
Tutkimustulokset osoittivat, että nattoaterioista eristetyt B. subtilis -bakteerikannoiksi
osoittautuneet heräteviljelmät tuottivat merkittäviä pitoisuuksia surfaktiineja. Isolaatit
osoittautuivat sekä β-hemolyyttisiksi että toksisiksi karjun siittiöille.
Tulosten perusteella arvioitiin ilman akuutteja terveyshaittoja aiheuttavaksi kerralla
nautittavaksi annokseksi nattoa 50 grammaa, mikä on yleinen Japanissa myytävä
annospakkaus. Akuutti toksisuus arvioitiin hyvin lieväksi.

UNIVERSITY OF EASTERN FINLAND, Faculty of Natural and Forest Sciences
Biosciences
Nutrition and Food Biotechnology
JUOLA MARI K.: The production of lipopeptides by Bacillus -bacteria in Japanese food
made from soya beans (natto)
Master’s thesis, 82 pages, 4 appendices, 3 pages
Supervisors: Prof. Atte von Wright and prof. Sirpa Kärenlampi
February 2015
Keywords: Bacillus subtilis, natto, cyclic lipopeptides, surfactins, cytotoxicity
The aim of this thesis was to find out the information of the food safety of a Japanese, soya
bean based, fermented food terming natto. The cyclic lipopeptide production potential of
fermentation starter strains Bacillus subtilis var. natto and their concentration in natto meal was
determined in association with Chr. Hansen A/G. Furthermore, the cytotoxicity of the
lipopeptides was tested. On the strength of the results a toxicological assessment of the safe
level of natto without any toxic effects was done.
Bacillus subtilis is very commonly found environmental bacterium, causing infections rarely.
It is widely utilized on the food technology. It has been used on traditional, soya based,
fermented food terming natto as a starter strain for several centuries. Occasionally, the findings
of B. subtilis, are being made on food poisonings. Furthermore, the ability of some B. subtilis
strains to produce cyclic lipopeptides as surfactins, fengycins, plipastatins and mycosubtilin as
metabolism products has documented. Notably the surfactins are hemolytic and cytotoxic, and
the safety for humans has challenged. European Food Safety Authority EFSA requires the
toxicological assessment of safety, follow-up studies and the identification of bacterial strains
in foods including B. subtilis.
In the experimental part, the starter strains from three natto meal samples was identificated by
16S rDNA and the synthesis of lipopeptides was analyzed by PCR and UHPLC-qTOF. The
possible lipopeptides was extracted from isolates and natto samples. The hemolytic activity on
bovine erythrocytes was detected and cytotoxicity assay by boar sperm motility inhibition was
performed.
The results indicated, that the starter strains characterized B. subtilis strains and produced
surfactins significantly. Moreover, all isolates proved to be β-hemolytic and culture extracts
were toxic to boar sperm cells.
On the strength of the results the safe level of natto without any toxic effects is 50 grams, which
is a standard package of natto in Japan. The acute toxicity of natto was assessed very low.

SISÄLTÖ
1. JOHDANTO 7
KIRJALLISUUS 8
2. BACILLUS SUBTILIS 8
2.1. Taksonomia 8
2.2. Ominaisuudet 9
2.3. Esiintyminen ja käyttö 11
2.3.1. Bacillus subtilis -bakteerin probioottikäyttö elintarvikkeissa 11
2.3.2. Bacillus subtilis -bakteerin probioottikäyttö rehuissa 12
2.3.3. Bacillus subtilis -bakteerikantojen käyttö heräteviljelminä 13
2.3.3.1. Natto 14
2.4. Bacillus subtilis var. natto 15
2.4.1. Genomi 16
2.4.2. Ominaisuudet 16
2.4.3. Poly-γ-glutamaattisynteesi 18
2.5. Bacillus subtilis infektioiden aiheuttajana 18
2.5.1. Elintarvikeperäiset infektiot 19
2.5.1.1. Probioottivälitteiset infektiot 19
2.5.2. Ruokamyrkytykset 20
2.5.2.1. B. subtilis -ryhmän bakteerien esiintyvyys elintarvikkeissa 20
2.5.2.2. Ruokamyrkytysten esiintyvyys 22
2.6. Bacillus subtilis -bakteerikantojen tuottamat lipopeptidit 26
2.6.1. Surfaktiinit 28
2.6.1.1. Synteesi 29
2.6.1.2. Ominaisuudet ja käyttö 30
2.6.2. Muut B. subtilis -ryhmän bakteerikantojen tuottamat lipopeptidit 31
2.6.2.1. Fengysiinit ja plipastatiinit 32
2.6.2.2. Mykosubtiliini 33
3. NATON ELINTARVIKETURVALLISUUSARVIOINTI 35
3.1. B. subtilis var. natto -bakteerikantojen käyttöturvallisuus 35
3.1.1. Nattoaterioiden B. subtilis var. natto -kokonaispitoisuus 35
3.1.2. Probioottien käyttöturvallisuus 38
3.2. B. subtilis var. natto -bakteerikantojen tuottamien toksiinien
aiheuttamat terveysriskit 39

3.3. Suolistokolonisaatio 43
3.4. Turvallisuuden arviointimenetelmät 44
3.4.1. Hemolyysi 45
3.4.2. Sytotoksisuus 45
3.4.3. Lipopeptidien tuottokyky 45
KOKEELLINEN OSA 46
4. TUTKIMUKSEN TAVOITTEET 46
5. AINEISTO JA MENETELMÄT 47
5.1. Bakteerikantojen eristäminen nattoaterioista 47
5.2. Tutkimuksessa käytetyt referenssibakteerikannat 48
5.3. Identifiointi 48
5.4. Lipopeptidien uuttaminen mikrobistosta 49
5.4.1. Uutto MOLP-elatusliemeen 49
5.4.1.1. Uutteen valmistaminen mikrobipesäkkeestä 49
5.4.1.2. Uutteen valmistaminen nattonäytteestä 50
5.4.2. 2-butanoliuutto 50
5.5. Hemolyysin määrittäminen 51
5.6. Sytotoksisuuden määritys karjun siittiöiden liikkuvuuden estotestillä 51
5.7. Lipopeptidien synteesikyky 52
5.7.1. Surfaktiinien synteesiin vaadittavan geenin määrittäminen 52
5.7.2. Fengysiinien, plipastatiinien ja mykosubtiliinin synteesiin
vaadittavien geenien määrittäminen 53
5.8. Mikrobien tuottamien lipopeptidien analysointi 54
6. TULOKSET 55
6.1. Identifiointi 55
6.2. Hemolyysi 55
6.3. Sytotoksisuus 56
6.4. Surfaktiinien synteesiin vaadittavan geenien esiintyminen 57
6.5. Fengysiinien, plipastatiinien ja mykosubtiliinin synteesiin
vaadittavien geenien esiintyminen 57
6.6. Nestekromatografiset tulokset 58
7. POHDINTA 60
8. LÄHTEET 64
9. LIITTEET 80

LYHENTEET
Caco-2 (Colon Carcinoma cells) Ihmisen suolistosyövästä eristetty epiteelisolulinja
EC50 (Effective Concentration 50 %) Pitoisuus, jossa puolella koe-eliöistä ilmenee koeaikana jokin
erikseen määriteltävä myrkkyvaikutus
EFSA (European Food Safety Authorization) Euroopan elintarviketurvallisuusviranomainen
GRAS (Generally Recognized as Safe) Yleisesti turvalliseksi tunnustettu
Hep-2 (Human larynx carcinoma cell line) Ihmisen kurkunpääsyövästä eristetty solulinja
HT29-16E (Human colonic epithelial goblet cell line) Ihmisen suolistosyövästä eristetty
epiteelisolulinja
LD50 (Lethal Dose 50 %) Kerta-annos, joka tappaa puolet koe-eliöistä koeaikana
M-CGH (Microarray-based comparative genomic hybridization) Mikrosirutekniikkaan
perustuva vertaileva genomien hybridisaatioanalyysi
MS (Mass Spectrometry) Massaspektrometri(a)
MS/MS (Tandem Mass Spectrometry) Kaksivaiheinen MS-analyysi
NOAEL (No Observed Adverse Effect Level) Annos, joka ei aiheuta koe-eliössä haitallista
muutosta
QPS (Qualified presumption of safety) Oletettavasti turvallinen
qTOF (Quadrupole-Time-Of-Flight) Kvadrupoli-lentoaika-(massa)analysaattori
TOF (Time-Of-Flight Mass Analysator) Lentoaika-(massa)analysaattori
UHPLC (Ultra High Performance Liquid Chromatography) Äärimmäisen suuren erotuskyvyn
nestekromatografi(a)

7
1. JOHDANTO
Osa Bacillus –suvun bakteerilajeista on todettu ruokamyrkytyksiä aiheuttaviksi patogeeneiksi.
Osaa pidetään terveydelle harmittomina, ja niitä hyödynnetään elintarvike- ja rehutuotannossa.
Eräs käytetyimmistä on Bacillus subtilis. Organismin turvallinen käyttö elintarvikkeissa on
kuitenkin kyseenalaistettu, sillä joissain ruokamyrkytystapauksissa, joiden aiheuttajaksi on
todettu tunnettu toksiineja tuottava patogeeni Bacillus cereus, näytteissä on esiintynyt
samanaikaisesti myös B. subtilis -bakteerikantoja. B. subtilis -ryhmän bakteerien on myös
havaittu itsessään kykenevän aiheuttamaan toisinaan sekä ruokamyrkytyksiä että muita
mahdollisia terveyshaittoja.
Merkittävimmäksi mahdolliseksi terveysriskien aiheuttajaksi on noussut esiin B. subtilis -
bakteerikantojen kyky tuottaa voimakkaita pinta-aktiivisia aineita kuten sytotoksisia,
hemolyyttisiä lipopeptidejä. Useilla bakteerikannoilla on havaittu lipopeptidien synteesiin
vaadittavia geenejä, ja erityisesti syklisen lipopeptidi surfaktiinin tuotto katsotaan terveydelle
mahdollisesti haitalliseksi sen korkean bioaktiivisuuden vuoksi (Mikkola ym. 2000).
Potentiaalinen surfaktiinien tuottaja on esimerkiksi perinteisessä japanilaisessa natto-nimisessä
elintarvikkeessa esiintyvä B. subtilis var. natto.
Natto valmistetaan fermentoimalla soijapapuja B. subtilis var. natto -bakteerikannan avulla, ja
lopullinen tuote sisältää organismin eläviä soluja yli 108 pmy grammassa. Perusteellista näyttöä
B. subtilis -bakteerikantojen elintarviketurvallisuudesta ei ole eikä B. subtilis var. natto
-bakteerikannan nattoateriassa tuottamista lipopeptideistä tiedetä juuri lainkaan.
Useat Bacillus -bakteerit, kuten B. subtilis, sisältyvät Euroopan Elintarviketurvallisuus-
viranomaisen (EFSA) elintarviketurvallisiksi oletettujen mikrobien nk. QPS-listalle (Qualified
presumption of safety). Euroopan ulkopuolella ei edellytetä käytön virallista
turvallisuusarviointia, mutta EFSA on ohjeistanut, että organismien lisääminen
elintarvikeketjuun vaatii toksikologisen arvioinnin ja seurannan (EFSA 2007, 2013). Lisäksi
edellytetään elintarvikkeessa käytetyn bakteerikannan identifiointia.
Tämän pro gradu -tutkielman tavoitteena on tuottaa tietoa siitä, kykenevätkö natto-aterian
valmistuksessa käytetyt bakteerikannat tuottamaan tiettyjä lipopeptidejä ja kuinka paljon sekä
arvioida näin naton elintarviketurvallisuutta.

8
KIRJALLISUUS
2. BACILLUS SUBTILIS
2.1. Taksonomia
Bacillus subtilis on Firmicutes -pääjaksoon ja Bacillus -sukuun kuuluva bakteerilaji. Bacillus
subtilis -laji jaetaan alalajeihin B. subtilis ssp. subtilis, B. subtilis ssp. spizizenii sekä B. subtilis
ssp. inaquosorum (Nakamura ym. 1999, Rooney ym. 2009, Yi ym. 2013). Bacillus subtilis ssp.
subtilis -klusterin muodostavat B. subtilis, B. pumilus ja B. licheniformis (Salkinoja-Salonen
ym. 1999). B. subtilis -ryhmän fylogenetiikka on hyvin haastavaa. Alalajit ovat fenotyypiltään
hyvin samankaltaisia ja niiden geenit ovat niin konservoituneita, että niitä on vaikea erottaa
toisistaan luotettavasti (Yi ym. 2013). Lähitaksonien luokittelu muuttuu hieman käytetyn
analyysimenetelmän mukaan.
Hyvin läheistä sukua B. subtilis -ryhmän alalajeille ovat esimerkiksi B. tequilensis, B.
mojavensis ja B. vallismortis (Roberts ym. 1994, 1996). Sen lisäksi, että B. mojavensis ja B.
vallismortis -lajit on vaikea erottaa alalajeista, myös erottamisen käytännön merkitys on
kiistanalaista. Perinteisin fenotyyppisin testein eroja ei voidakaan havaita (Rooney ym. 2009,
Logan ja de Vos 2009), ja yleisimmin käytetty 16S rRNA -geenin sekvenointi antaa usein
virheellisen tuloksen (Barbosa ym. 2005, Earl ym. 2007, Wang ym. 2007). Tarkin erotuskyky
B. subtilis –ryhmän alalajien kesken saavutetaan mikrosirutekniikkaan perustuvalla DNA-
hybridianalyysillä (M-CGH) tai sekvenoimalla gyrA tai gyrB, joten on suositeltavaa käyttää
niitä 16S rRNA –sekvenoinnin sijasta (Earl ym. 2007, Wang ym. 2007, Yi ym. 2013).
Toisilleen hyvin läheisten B. subtilis -bakteerikantojen vaikea erotettavuus on johtanut siihen,
että teollisuudessa käytetään B. subtilis -bakteerikantana käytännössä myös B. mojavensis- tai
B. vallismortis -bakteerikantoja (Euroopan Komissio 2000). Erityisesti B. vallismortis
-bakteerikannat voidaan erottaa B. subtilis ssp. subtilis -alalajista luotettavimmin sekvenoimalla
gyrA ja gyrB (Wang ym. 2007, EFSA 2013).

9
2.2. Ominaisuudet
Bacillus subtilis (Ehrenberg 1835, Koch 1872) on Gram-positiivinen, katalaasipositiivinen,
liikkuva 0,7 – 3,0 µm pituinen sauvabakteeri. Pesäkkeet kasvavat yleensä joko yksittäin tai
pareina, harvoin ketjuina. Niiden morfologia vaihtelee jopa saman bakteerikannan sisällä
kiinteistä voimaisiin tai limaisista kalvopintaisiin. Myös reunan muoto vaihtelee säännöllisistä
hyvinkin aaltoileviin ja epäsäännöllisiin. B. subtilis luokitellaan aerobiseksi, mutta se voi
lisääntyä myös anaerobisissa olosuhteissa joko nitraattia pelkistäen tai glukoosia fermentoiden
sitä stimuloivan pyruvaatin läsnä ollessa (Nakano ym. 1997, Nakano ja Zuber 1998, Rooney
ym. 2009).
Bacillus subtilis hydrolysoi kaseiinia, gelatiinia ja tärkkelystä. Fermentaatiossa organismi
tuottaa laktaattia ja asetaattia sekä glukoosin läsnä ollessa myös 2,3-butaanidiolia. Solut
kykenevät myös muodostamaan polysakkaridi- tai polyglutamiinihappokapselin kasvun
stationäärisessä vaiheessa, mutta sillä ei ole yleensä taksonomista merkitystä. B. subtilis var.
natto -bakteerikantojen syntetisoimalla solunulkoisen poly-γ-glutamiinihapon (PGA)
tuotannolla on sitä vastoin merkittävää taloudellista hyötyä. (Logan ja de Vos 2009.)
Optimikasvulämpötila on 28–30 oC vaihteluvälin ollessa yleensä 5–55 oC (Logan ja de Vos
2009). Useat B. subtilis -bakteerikannat voivat kasvaa jopa yli 56 oC:ssa (Rooney ym. 2009).
Kasvualueen pH vaihtelee arvojen 5,5 ja 8,5 välillä. Osa B. subtilis -bakteerikannoista sietää
jopa 10 % natriumkloridipitoisuutta, mutta tavallisesti kasvu heikkenee NaCl-pitoisuuden
kohotessa 6 %:iin (van der Ploeg ym. 2012). B. subtilis käyttää ravinteenaan tyypillisesti
sitraattia ja hiilenlähteinään esimerkiksi L-arabinoosia, D-glukoosia, natriumglukonaattia,
sakkaroosia, tärkkelystä ja D-ksyloosia (Pötter ym. 2001, Logan ja de Vos 2009).
Hyvin tunnusomaista B. subtilis -bakteerille on elinolosuhteiden heikentyessä esiintyvä itiöinti.
Pääosin organismin tiedetään tuottavan endosporeja, mutta myös eksosporeja on havaittu
(Hong ym. 2009). Muoto vaihtelee ellipsoidaalisista sylinterimäisiin (Logan ja de Vos 2009).
Lajintunnistuksessa itiönmuodostus korostuukin solujen ja itiöiden välisessä erottumisessa.
Merkittävä rakenteellinen ominaisuus B. subtilis -lajin itiöillä on niitä ympäröivä
monikerroksinen, vähintään 70 erilaisesta proteiinista muodostunut ulkokuori, joka
mahdollistaa vuosien, jopa vuosisatojen resistenttiyden ympäristön ääriolosuhteita vastaan
(McKenney ym. 2013).

10
Itiöt ovat metabolisesti lepotilassa ja osittain dehydratoituneita, mikä mahdollistaa niiden
säilymisen elinkykyisenä ravinteiden puuttuessa (Nguyen Thi Minh ym. 2011). Resistenttiyden
kestoa ravinteettomuutta, kuumuutta, kuivuutta, hydrostaattista painetta, suolapitoisuutta sekä
muita olosuhteita vastaan on haastavaa luotettavasti arvioida, sillä itiöt kehittyvät ja muuntuvat
olosuhteiden muutosten mukaan. Esimerkiksi B. subtilis -bakteereilla saastuneessa
elintarvikkeessa puolen tunnin ajan 100 oC:ssa kuumentamisen jälkeen 10 % itiöistä on edelleen
elinkelpoisia (Evira 2010). Takaisin vegetatiivisiksi soluiksi ne puolestaan muuntuvat hyvin
nopeasti. Itiöt voidaan tuhota autoklavoiden 20 minuuttia 121 oC lämpötilassa. Itiönmuodostus
on puolet nopeampaa ns. villin tyypin bakteerikannoilla kuin laboratoriokannoilla (Tam ym.
2006). Itiöitymistehokkuutta nostaa myös organismin kyky tuottaa useita kymmeniä eri
lipopeptidejä (Nakano ym. 1991).
Kasvuolosuhteiden heikentyessä ja usein itiönmuodostuksen yhteydessä useat B. subtilis
-bakteerikannat muodostavat myös soluja ympäristön epäedullisilta fysikokemiallisilta
tekijöiltä suojaavaa biofilmiä. Nopea itiönmuodostuskyky korreloi suoraan biofilmin
muodostuksen kanssa (Hong ym. 2009). Biofilmin rakennetta pitää koossa ja vahvistaa
solunulkoinen, pääasiassa eksopolysakkarideista ja amyloidikuidusta koostuva matriisi.
Biofilmin rakenteeseen vaikuttaa merkittävästi solujen kyky syntesoida surfaktiineja.
Surfaktiinit ovat syklisiä lipopeptidejä, jotka laskevat voimakkaasti solun pintajännitystä.
Surfaktiinia tuottavat bakteerikannat muodostavat tyypillisesti paksun, monisyisen ja
suonimaisen biofilmin, kun taas sitä tuottamattomien kantojen muodostama biofilmi on ohut,
hauras ja sileä. Myös pesäkkeiden koko on suurempi kuin surfaktiinia tuottamattomien
kantojen. Jotkut surfaktiinia tuottavat bakteerikannat myös kykenevät elämään pysyvästi
esimerkiksi ihmisen suolistossa persistoituen sinne pidemmäksi ajaksi kuin sitä tuottamattomat
kannat. (Branda ym. 2001, 2006; Barbosa ym. 2005; Tam ym. 2006.)
Suurin osa tunnetuista B. subtilis -bakteerikannoista ei ole patogeeneja, ja useat inhiboivat
patogeenisten bakteerien kasvua (Barbosa ym. 2005, Hong ym. 2008, Larsen ym. 2013).
Organismin ei yleensä ole havaittu tuottavan enterotoksiinia (Beattie ja Williams 1999,
Pedersen ym. 2002, Phelps ja McKillip 2002). Enterotoksiinituotanto on kuitenkin mahdollista
mediumin koostumuksen ollessa maltodekstriini- tai glukoosipitoinen (Rowan ym. 2001). B.
subtilis on usein vahvasti hemolyyttinen (Barbosa ym. 2005).

11
2.3. Esiintyminen ja käyttö
Bacillus subtilis on hyvin yleinen ympäristöbakteeri. Se on eristetty alun perin kuivasta heinästä
(Logan ja de Vos 2009), ja sitä esiintyykin runsaasti maaperässä, kasveissa, vesisedimentissä
sekä ilmassa. B. subtilis -bakteerin laajat kasvuominaisuudet, antimikrobisten yhdisteiden ja
biofilmin muodostamiskyky sekä aerobinen ja anaerobinen itiöinti edistävät organismin
runsasta esiintyvyyttä ympäristössä (Hong ym. 2008, Rooney ym. 2009). B. subtilis on
luonteeltaan alloktoninen, eli esiintyessään ihmisen elimistössä se on kulkeutunut sinne
elimistön ulkopuolelta joko sillä kontaminoituneen tai fermentoidun elintarvikkeen mukana. Se
voi kuitenkin kasvaa ja lisääntyä ihmisen ruoansulatuskanavassa koko elinkaarensa ajan
(Sanders ym. 2003).
B. subtilis -bakteerikantoja ja niiden itiöitä hyödynnetään laajalti elintarvike-, rehu- ja
tekstiiliteollisuudessa. Isolaatteja ja niiden avulla tuotettuja peptidejä käytetään erilaisten
entsyymien, kuten amylaasien ja ekstrasellulaaristen proteaasien tuotannossa, pinta-aktiivisten
lipopeptidien kuten surfaktiinin tuottajina, erilaisina apuaineina sekä pestisideinä. Hong ym.
(2008) havaitsivat B. subtilis var. natto -bakteerikannan tuottavan hyvin biosurfaktantti
surfaktiinia. Organismia hyödynnetään myös lääketeollisuudessa vitamiinien, antibioottien,
nukleotidien ja nukleosidien tuotannossa. B. subtilis -bakteerikantojen fermentaatioprosessissa
tuottamalla D-riboosilla voidaan voimistaa ihmis- ja eläinravinnon aromia lääketeollisuuden
sovellusten lisäksi. Etenkin B. subtilis -bakteerikantojen itiöitä käytetään heräteviljelminä
perinteisissä aasialaisissa ja afrikkalaisissa elintarvikkeissa sekä probiootteina elintarvikkeissa
ja rehuissa. (Schallmey ym. 2004, Hong ym. 2008, EFSA 2013.)
2.3.1. Bacillus subtilis -bakteerin probioottikäyttö elintarvikkeissa
B. subtilis -bakteerikantoja sisältäviä valmisteita käytetään yleisesti ihmisille ja eläimille
probiootteina. Käyttönsä luonteen vuoksi probioottien tulisi olla resistenttejä maha- ja
sappinesteelle sekä kyetä kiinnittymään limakalvolle tai epiteelisoluihin. Niiden tulisi myös
inhiboida potentiaalisten patogeenien kasvua ja kyetä vähentämään patogeenien adheesiota
pinnoille sekä hydratoimaan sappinestettä sekä in vitro että in vivo (FAO/WHO 2002).

12
Organismista valmistettujen probioottivalmisteiden on havaittu inhiboivan useiden
patogeenien, kuten Listeria monocytogenes- ja Clostridium perfringens -bakteerikantojen
kasvua. Patogeenien inhibitio vaihtelee kuitenkin suuresti ollen sekä kanta- että
substraattispesifistä (Barbosa ym. 2005, Cutting ym. 2011, Kaboré ym. 2013).
Itiöitä muodostavilla B. subtilis -bakteerikannoilla on useita merkittäviä etuja verrattuna ei-
itiöiviin mikrobeihin, kuten joihinkin Lactobacillus -bakteerikantoihin. Probioottisia B. subtilis
-bakteerikantoja hyödynnetäänkin useita eri Bacillus -bakteerikantojen itiöitä sisältävinä
itiövalmisteina. Kuivatut itiöt säilyvät erittäin hyvin huoneenlämmössä elinkyvyn
heikkenemättä. Niiden nopea germinoituminen, biofilmin muodostuskyky ja uudelleen
itiöityminen mahdollistavat koko elinkaaren aikaisen säilymisen ruuansulatuskanavan
happamassa ja osittain anaerobisessa ympäristössä. Lisäksi solujen muodostama biofilmi
suojaa niitä mahanesteen happamuudelta ja voimistaa niiden adheesiota suolistolimaan.
(Barbosa ym. 2005, Tam ym. 2006, Hong ym. 2008, 2009.)
Epäsuora käyttösovellus on myös mahdollisuus hyödyntää B. subtilis -bakteerikantojen
tuottamaa polypeptidiä, poly-γ-glutamaattia (PGA) muiden probioottien kryoprotektanttina.
Säilyvyyden parantamiseksi ei-itiöivistä maitohappobakteereista valmistetut probiootit usein
pakastetaan. Pakastaminen kuitenkin itsessään vähentää elinkykyisten mikrobien pitoisuutta
tuotteessa keskimäärin kolme kertaluokkaa. Bhatin ym. (2013) mukaan pakastaminen steriilissä
PGA-matriisissa vahvistaa erityisesti Lactobacillus paracasei -bakteerikannan solujen
elinkykyä. Ei-hemolyyttistä B. subtilis var. natto ATCC 15245 -bakteerikantaa pidetään
turvallisena PGA:n tuottajana, sillä sen genomissa ei ole havaittu esiintyvän tunnettuja entero-
tai enteritoksiineja koodaavia geenejä hblD/A, nheB, bceT ja entFM. Myöskään toksisten
sfingomyelinaasin tai fosfolipaasi A2:n tuotantoon vaadittavia geenejä sph tai piplc ei ole
havaittu. (Bhat ym. 2013.)
2.3.2. Bacillus subtilis -bakteerin probioottikäyttö rehuissa
Organismia käytetään probioottina myös tuotantoeläimille, pääasiassa siipikarjalle ja porsaille
(Euroopan komissio 2002; 2013a, b). B. subtilis ssp. subtilis -bakteerikantojen itiöitä sisältäviä
valmisteita lisätään niiden rehuun kasvun promoottoriksi sekä lisäämään eläinten
vastustuskykyä patogeenejä kohtaan.

13
Probiootin on arvioitu laskevan infektioiden määrää ja siten antibioottien käyttöä
tuotantoeläimillä, minkä seurauksena niistä saatava taloudellinen tuotto paranee. (Hong ym.
2009, Cutting 2011, Larsen ym. 2014.)
Myös uusimmat tutkimustulokset organismin probioottikäytöstä tuotantoeläinten rehussa ovat
taloudellisesti lupaavia. B. subtilis var. natto -solujen tai niiden avulla fermentoidun tuotteen
lisääminen lypsylehmien rehuun on havaittu lisäävän niiden maidontuottoa sekä maidon
sisältämän rasvan määrää (Peng ym. 2012, Sun ym. 2013). Myös pötsin somaattisten solujen
määrän lasku ja proteolyyttisten bakteerien pitoisuuden nousu sekä suoliston
kokonaisbakteerimäärän nousu viittaavat kyseisten bakteerikantojen käytön kannattavuuteen
lypsylehmien rehussa. Vasikoiden ravintoon lisätyt vegetatiiviset B. subtilis var. natto -solut
puolestaan edistävät niiden pötsin kehittymistä nopeuttaen vasikan muuttumista märehtijäksi
vähentäen näin karjanhoitokustannuksia (Sun ym. 2011).
Song ym. (2014) puolestaan havaitsivat, että B. subtilis var. natto -bakteerikantojen itiöitä
sisältävän valmisteen lisäys naudan rehuun laski ulosteen typpipitoisuutta ja vähensi
proteiinineritystä ulosteeseen. Organismi nosti näin naudan ravinnon sisältämän typen
hyödyntämisastetta. Tutkimuksessa havaittiin myös ruoansulatuskanavan mikrobipopulaation
muuttuneen merkitsevästi kahdeksan viikon probioottilisäyksen jälkeen esimerkiksi nostamalla
hyödyllisten bifidobakteerien määrää suolistossa. Lisäksi eläinravinnon lisäaineena voidaan
käyttää B. subtilis -bakteerikantojen tuottamaa PGA:a (Schallmey ym. 2004).
2.3.3. Bacillus subtilis -bakteerikantojen käyttö heräteviljelminä
Tiettyjä B. subtilis -bakteerikantojen itiöitä käytetään runsaasti heräteviljelminä useissa
perinteisissä fermentoiduissa elintarvikkeissa. Käyttökohteita ovat esimerkiksi
kaakkoisaasialaiset perinneruoat ”natto” Japanissa, ”shi” Kiinassa, ”thua-nao” Thaimaassa,
”chunk-kook-jong” Koreassa ja ”tao-si” Filippiineillä (Wei ym. 2001). Fermentoinnin aikana
lisätty organismi saa aikaan raaka-aineiden proteiinien, hiilihydraattien ja lipidien
pilkkoutumisen asetaateiksi, hiilidioksidiksi ja vedyksi. Mikrobien aikaansaamat pH-arvojen
muutokset ja metaboliatuotteiden kuten haihtuvien orgaanisten yhdisteiden muodostuminen
antavat elintarvikkeelle sille ominaisen rakenteen, aromin ja maun. (Beaumont 2002, Shrestra
ym. 2010.)

14
2.3.3.1. Natto
Natto on japanilainen, useita satoja vuosia vanha perinneruoka, joka keksittiin alun perin
vahingossa: soijapapuja pakattiin olkien sisään ja säilytettiin vähähappisissa olosuhteissa maan
alla vähintään viikon ajan. Oljissa luonnollisesti esiintyvät B. subtilis -bakteerikantojen
haihtuvat aldehydit, ketonit ja hapot aiheuttivat papuihin säilytyksenaikaisen fermentoitumisen
aikana voimakkaan, pähkinäisen aromin sekä solujen tuottama PGA papuja yhdistävän runsaan
liman muodostumisen. Teollinen valmistus alkoi vasta 1900-luvun alussa, kun japanilainen
kemisti S. Sawamura eristi ja puhdisti bakteerikantoja nattoaterioista, nimesi ne Bacillus natto
-bakteerikannoiksi, ja organismin itiösuspensiota alettiin lisätä valmistusprosessiin
tarkoituksellisesti. Nykyään teollinen valmistus on tarkoin säädettyä ja valvottua.
Valmistusprosessin yksinkertaisuuden vuoksi nattoa valmistetaan kuitenkin myös
yksityistalouksissa. (Sawamura 1906, Beaumont 2002, Nagai 2012.)
Teollinen valmistusprosessi on monivaiheinen. Pienehköt soijapavut pestään, ja niitä liotetaan
10 oC vedessä 18 tuntia. Pehmenneitä papuja höyrytetään kiertokeittimessä 2 kg paineessa
1–1,5 tunnin ajan, jolloin mikrobit kuolevat ja papujen ravintoarvo paranee proteiinien
denaturoituessa. B. subtilis var. natto -bakteerien itiöitä laimennetaan veteen siten, että
pitoisuus on keskimäärin 108 per 60 kg papuja ja suspensio lisätään höyrytettyihin papuihin.
Inokulaatin mikrobipitoisuus vaihtelee suuresti käytettävän bakteerikannan mukaan (Wei ym.
2001). Toisaalta myös valmistusprosessin olosuhteet vaikuttavat tarvittavan populaation
määrään. Pavut jaetaan polystyreenirasioihin 30–50 g annoksiin ja peitetään muovikalvolla.
Oheen laitetaan maustepakkaukset, yleensä maustekastiketta ja sinappia, ja annoksia
fermentoidaan 40–50 oC lämpötilassa korkeassa kosteuspitoisuudessa 16–18 tunnin ajan,
jolloin proteiinit hydrolysoituvat. Sen jälkeen fermentointia jatketaan alhaisemmassa
kosteuspitoisuudessa 3–10 oC lämpötilassa 8 tuntia. Lopullisen tuotteen B. subtilis -pitoisuus
on 108 – 1010 pmy/g. Valmistusprosessissa mikrobien limantuotto on oleellista. Natolle
ominainen korkeaviskositeettinen lima koostuu pääasiassa levaanimuotoisesta
eksopolysakkaridista fruktaanista sekä PGA:sta (Hara ym. 1981, Wei ym. 2001). Onnistuneena
lopputuotteena pidetään vaalean, silkkisen limakerroksen peitossa olevaa pehmeää ja
karakteristisen aromin omaavaa korkeaviskoosista massaa, joka kyetään hyvin nauttimaan
syömäpuikoilla. Valmiin naton laatu vaihtelee voimakkaasti myös liotus- ja höyrytysajan,
inokulaatin ja sen pitoisuuden sekä fermentaatioajan mukaan (Wei ym. 2001).

15
Korkean viskositeetin aikaansaamiseksi nattoa sekoitetaan sakkaroosin läsnä ollessa. Mitsuin
ym. (2011) mukaan viskositeetti laskee säilytyksen aikana, mutta bakteriofageilla
kontaminoituneen naton viskositeetti laskee jyrkästi jo sekoittamisen aikana. (Wei ym. 2001,
Schallmey ym. 2004, Kim ym. 2012, Nagai 2012.)
Fermentaatiossa tapahtuviin monimutkaisiin biokemiallisiin reaktioihin, niiden asteeseen ja
nopeuteen sekä tuotettuihin yhdisteisiin vaikuttavat voimakkaasti fermentointiprosessin eri
vaiheiden kestot sekä käytetyt bakteerikannat (Wei ym. 2001).
Natto on erittäin proteiinipitoinen elintarvike. Noin 36 % soijapavun kuivapainosta koostuu
proteiinista, ja valmis tuote sisältää noin 55–60 % vettä, 17 % proteiinia ja 10 % rasvaa (Wei
ym. 2001, Fineli 2014, USDA 2014). Nattoa nautitaan Japanissa tyypillisesti aamiaiseksi
kuumentamatta (Wei ym. 2001). Nattoa on kolmenlaista: itohiki-natto, yukiwai-natto ja hama-
natto. Suurimmat määrät tuotetaan Itohiki-nattoa, ja sana ”natto” viittaakin yleensä siihen.
Kyseessä on edullinen ja arkinen elintarvike, jonka säilyvyyttä pidetään hyvänä. Vaikka natto
nautitaan perinteisesti sipulin, merilevän tai mausteiden sekä riisin kera, sitä käytetään
Japanissa myös aromaattisena aineena valmistettaessa liha-, merilevä- tai kasvisruokia taikka
yhtenä kastikkeen ainesosana. (Wei ym. 2001.)
Nattoa pidetään Japanissa yleisesti terveellisenä ja terveysvaikutteisena elintarvikkeena, ja siitä
valmistetaan myös ravintosupplementteja (Chen ym. 2012). Naton ja B. subtilis var. natto
-bakteerikantojen vegetatiivisten solujen tuottaman fibrinolyyttisen entsyymin nattokinaasin
nauttimisen on havaittu stimuloivan immuunijärjestelmää sekä suojaavan sydän- ja
verisuonitaudeilta (Sumi ym. 2004, Cutting 2011). Myös korkeaan kalsium- ja K2-
vitamiinipitoisuuteen perustuvaa osteoporoosilta suojaavaa vaikutusta on raportoitu (Ikeda ym.
2006). Toisaalta natto sisältää terveydelle haitallisen määrän biogeenisia amiineja, erityisesti β-
fenyylietyyliamiinia ja tyramiinia (Kim ym. 2012).
2.4. Bacillus subtilis var. natto
Erityisesti naton tuottamiseen soveltuvat B. subtilis var. natto -bakteerikannat (lyhyesti B.
subtilis natto tai Bacillus natto) ovat läheistä sukua referenssibakteerikannalle B. subtilis
Marburg 168. Naton fermentaatioprosessiin soveltuu useita kymmeniä eri B. subtilis var. natto
-bakteerikantoja (Kubo ym. 2011).

16
Kaupallisessa tuotannossa käytetään kuitenkin pääasiassa kolmea, joista yli 50 vuotta sitten
eristetty B. subtilis natto Mira (kutsutaan myös nimellä Miura) -bakteerikannan markkinaosuus
on 70–80 %. Muut pääosin käytetyt bakteerikannat ovat Naruse ja Takahashi (Nagai 2012).
Fermentointiin käytetty bakteerikanta vaikuttaa naton lopulliseen mikrobipitoisuuteen
(vaihteluväli 2,6 – 5,0 ×109 pmy/g), pH-arvoon (vaihteluväli noin 7…8), viskositeettiin sekä
ammoniakkipitoisuuteen (Wei ym. 2001). B. subtilis var. natto -bakteerikannat eroavat muista
B. subtilis –bakteerikannoista pääosin korkean PGA:n tuottokyvyn, bakteerifaagispesifisyyden
sekä insertiosekvenssien suhteen. Ne ovat myös biotiiniauksotrofisia. (Hara ym. 1982; Nagai
ym. 1997, 2000; Kubo ym. 2011.)
2.4.1. Genomi
B. subtilis var. natto -bakteerikantojen genomille tunnusomaisia ovat karakteristiseen tuoksuun
ja bakteriofagi φNIT1:n herkkyyteen sekä itiön muodostukseen vaikuttavat
sekundäärimetaboliittien metaboliareittejä säätelevät geenit kuten gyrA, rpoB, purH, polC ja
groEL (Kubo ym. 2011). Korkeaviskoosisen liman muodostukseen vaaditaan myös PGA:ta
koodaava geenioperoni pgsBCAywtC (Ashiuchi ym. 2003, Qiu ym. 2004, Kimura ym. 2009).
Genomissa ei esiinny profaagia SPβ eikä skin -elementtiä, kun taas profagi PBSX esiintyy (Qiu
ym. 2004). Myöskään biotiinin synteesiin vaadittavaa bio-geeniä genomissa ei esiinny.
Insertiosekvenssien (IS) IS4Bsu1 ja IS256Bsu1 puuttuminen genomista kuvaa osaltaan myös
organismin kykyä toimia naton heräteviljelmänä. IS4Bsu1 aiheuttaa epästabiiliutta mikrobien
PGA-synteesiin. IS-elementtien esiintyminen genomissa ei sinänsä estä onnistunutta
fermentaatioprosessia, mutta niiden läsnäolo voi heikentää lopputuotteen laatua. (Inagaki. ym.
2003, Kubo ym. 2011.) Enterotoksiineja tuottavia geenejä hbl, nhe, cytK eikä bceT ei ole
havaittu (Hong ym. 2008, Bhat ym. 2013).
2.4.2. Ominaisuudet
B. subtilis var. natto -bakteerikannat tuottavat spesifisellä kasvualustalla proteaasia ja osa niistä
myös amylaasia (Inatsu ym. 2002). Mutantin B. subtilis natto Mira -bakteerikannan avulla
tuotetaan EFSA:n turvalliseksi arvioimaa lasten K2-vitamiiniöljyvalmistetta. (Nishiguchi ym.
2002, EFSA 2008). Lisäksi bakteerikannat ovat tehokkaita pinta-aktiivisten aineiden tuottajia.
Antibioottiresistenttiyttä ei ole havaittu (Hong ym. 2008).

17
Natosta eristetyille bakteerikannoille on ominaista myös runsas fibrinolyyttisen entsyymin,
nattokinaasin tuotto, jonka on havaittu inhiboivan sydän- ja verisuonisairauksia osaltaan
aiheuttavien verihyytymien muodostumista in vivo (Sumi ym. 2004). Nattokinaasin on havaittu
myös estävän valtimoiden sisäkalvon paksuuntumista, joka on seurausta suonten
endoteelisoluvaurioista (Suzuki ym. 2003). Vauriot altistavat verihyytymien muodostumiselle.
Lisäksi on saatu viitteitä organismin tuottaman surfaktiinin antitumoraalisuudesta (Schallmey
ym. 2004, Cao ym. 2009). Altistettuaan ihmisen rintasyöpäsolut surfaktiinipitoisuudelle 40
mg/l Cao ym. (2009) havaitsivat apoptoosinopeuden merkitsevää lisääntymistä ja solusyklin
pysähtymistä G2/M -faasiin.
Hongin ym. (2008) tutkimuksessa havaittiin B. subtilis var. natto -bakteerikantojen säilyttävän
elinkykynsä hyvin ruoansulatuskanavan haastavissa olosuhteissa. Ne itiöivät hyvin
mahanesteessä ja suolilimassa sekä muodostivat selviämistä merkittävästi edesauttavan
hydrofobisen biofilmin. Organismi myös germinoitui nopeasti. Lisäksi se inhiboi merkitsevästi
sekä gram-positiivisten että gram-negatiivisten patogeenien kasvua.
Vaikka B. subtilis var. natto -bakteerikantojen genomien ole havaittu sisältävän
enterotoksiineja koodaavia geenejä, organismilla esiintyy muita virulenssitekijöitä (Hong ym.
2008). Sen on todettu olevan hemolyyttinen ja tuottavan runsaasti lämpöresistenttejä toksiineja:
sytotoksista syklistä lipopeptidiä surfaktiinia sekä nekroottista neurotoksista proteiinia,
fosfolipaasi A2:a (Hong ym. 2008, EFSA 2013). Neurotoksisuuden vuoksi fosfolipaasi A2:n
tuotanto on syytä huomioida toksikologisessa arvioinnissa (Chu 1949, Matarante ym. 2004,
Hong ym. 2008). Hongin ym. (2008) mukaan B. subtilis var. natto –bakteerikanta on myös
sytotoksinen Caco-2-, Hep-2- ja HT29-16E –soluille.
Nattoaterioiden fermentointiin käytettäville bakteerikannoille on tunnusomaista myös herkkyys
spesifisille bakteriofageille. Nagain (2012) mukaan heräteviljelmiä infektoivia bakteriofageja
tunnetaan useita kymmeniä. Ne luokitellaan nykyään infektiivisyyden, lämpöresistenttiyden ja
kasvualustavaatimusten perusteella kahteen ryhmään. Ryhmään 1 sisältyy mm. tunnettu
JNDMP -fagi sekä ryhmään 2 tunnetut P-1 ja ONPA. Ryhmän 2 bakteriofagit ovat
virulentimpia ja lämpöresistentimpiä kuin ryhmään 1 sisältyvät. Ne kykenevät kontaminoimaan
kaikkia kaupallisia nattoon käytettäviä heräteviljelmiä, kun taas ryhmän 1 bakteriofagit eivät
voi kontaminoida Mira -viljelmiä. Jälkimmäiset myös vaativat kasvualustaansa magnesiumia.

18
Bakteriofagien infektoimien solujen tuottaman PGA:n viskositeetti laskee voimakkaasti pian
itiösuspension lisäämisen ja solujen germinoitumisen jälkeen heikentäen näin naton laatua
(Kimura ja Itoh 2003). Fagikontaminaatio pyritään estämään normaalein elintarvikehygienian
keinoin (Nagai 2012).
2.4.3. Poly-γ-glutamaattisynteesi
Hyvin tunnusomaista ja merkityksellistä B. subtilis var. natto -bakteerikannoille on niiden kyky
tuottaa solunulkoista poly-γ-glutamaattia (PGA) L-glutamaatin puuttuessa kasvualustasta
(Inatsu ym. 2002). PGA on glutamiinihapon polymeeri, anioninen polypeptidi, jossa kaksi
karboksyyliryhmää on polymeroitunut hiiliketjun α- ja γ -hiiliin. Naton solunulkoisen liman
sisältämä PGA koostuu 50–80 %:sti D- ja 20–50 %:sti L -muodon glutamaatista (Ashiuchi ym.
2003). Isomeerien keskinäinen suhde määräytyy mediumin mangaanipitoisuuden mukaan: mitä
enemmän siinä on mangaania, sitä suurempi osa glutamaatista on D-muodossa (Leonard ym.
1958, Nagai 2012). PGA absorboi tehokkaasti vettä ja veteen liuenneena suojaa mikrobisoluja
kylmältä sekä muodostaa fysikaalisen esteen ei-toivottujen organismien kuten bakteriofagien
pääsylle soluun (Kimura ja Itoh 2003, Kimura ym. 2009). PGA:n aktiivisuuden väheneminen
säilytyksen aikana voi aiheuttaa viskositeetin stabiloitumisen (Mitsui ym. 2011). Parhaiten
limaa erittävät ja niin ollen tehokkaimpia heräteviljelmiä ovat ne toisilleen hyvin läheiset B.
subtilis -bakteerikannat, jotka sekä tuottavat poly-γ-glutamaattia että ovat biotiinille
auksotrofisia (Kubo ym. 2011).
2.5. Bacillus subtilis infektioiden aiheuttajana
B. subtilis infektoi ihmisiä tiettävästi suhteellisen harvoin. Suurimman terveyshaitan on arvioitu
olevan vakava, paikallinen tai systeeminen infektio (Sanders ym. 2003). Tämänhetkisen tiedon
valossa näyttää siltä, että B. subtilis -bakteerin aiheuttama infektioriski on suurin
vastustuskyvyltään heikoilla henkilöillä kuten imeväisillä, immunosuppressiivisilla tai muuten
immuniteetiltaan heikentyneillä henkilöillä sekä implantin omaavilla potilailla (Logan 2011).
Immunosuppressiiviset syöpäpotilaat tai jonkin muun vaikean sairauden heikentämät henkilöt
voivat herkimmin sairastua organismin aiheuttamaan hengenvaaralliseen bakteremiaan, jolloin
mikrobit lisääntyvät potilaan verenkierrossa. B. subtilis -itiöitä sisältävät probioottivalmisteet
toimivat eräänä reittinä elimistöön.

19
Lisäksi vaikean sairauden hoitoon liittyy usein injektointeja, punkteerauksia, katetrointeja tai
vastaavia toimenpiteitä, jotka mahdollistavat ympäristöstä vaikeasti häädettävän B. subtilis
-bakteerin pääsyn potilaan kudoksiin. Organismi voi saada aikaan paikallisen infektion myös
lapsella esim. ihon lävitse tunkeutuneen tikun kautta. (de Boer ja Diderichsen 1991.)
2.5.1. Elintarvikeperäiset infektiot
Elintarvikeperäisen infektion aiheuttaa joko elintarvikkeessa esiintyvän bakteerin suuri
pitoisuus tai sen tuottama(t) toksiini(t). Elintarvikeperäiset B. subtilis -infektiot jaetaan
aiheuttamismekanismin perusteella elintarvikevälitteisiin infektioihin ja ruokamyrkytyksiin.
Elintarvikevälitteisen infektion aiheuttaa B. subtilis -bakteerikannan riittävän suuri
infektiivinen annos elintarvikkeessa tai siihen verrattavassa tuotteessa kuten probiootissa.
Ruokamyrkytys aiheutuu bakteerin tuottamasta toksiinista elintarvikkeessa ja/tai sitä nauttineen
henkilön elimistössä. (Evira 2010.)
2.5.1.1. Probioottivälitteiset infektiot
B. subtilis -bakteerikantoja sisältäviä itiövalmisteita on käytetty jo vuosia suolistosairauksien
rinnakkaishoitokeinona (Spinosa ym. 2000). Tunnettuudesta huolimatta B. subtilis -bakteerien
käyttäminen probiootteina ei ole riskitöntä. Eräässä eurooppalaisessa sairaalassa oli 1980-
luvulla B. subtilis -bakteerien aiheuttamaan bakteremiaan sairastunut kuuden vuoden aikana
kahdeksan potilasta. Puolet heistä oli kärsinyt ripulista, jota pyrittiin hoitamaan B. subtilis
-itiöprobiootilla. Potilaat olivat letkuruokinnassa ja pääosin syöpäpotilaita. Kukaan potilaista
ei menehtynyt. (Richard ym. 1988.)
Seuraavalla vuosikymmenellä immuunipuolustukseltaan heikentynyt iäkäs potilas menehtyi
bakteremiaan nautittuaan B. subtilis -bakteerin itiöitä sisältävää probioottivalmistetta. Valmiste
sisälsi liian suuren pitoisuuden itiöitä potilaan yleistilaan nähden. Germinoiduttuaan potilaan
elimistössä vegetatiivisten solujen suuri määrä sai aikaan yleistilan heikentymisen ja sitä kautta
kuoleman voimakkaista antibioottihoidosta huolimatta. Nautitut bakteerikannat eivät olleet
resistenttejä käytetyille antibiooteille. (Oggioni ym. 1998.)

20
Hyväkuntoisetkin aikuiset voivat toisinaan sairastua vakavasti B. subtilis -bakteerin
aiheuttamaan infektioon. Suuren ihmisryhmän turvallisena pitämät Herbalife®-
ravintovalmisteet ovat kasviperäisiä luontaistuotteita, jotka eivät sisällä vaikuttavana
ainesosana B. subtilis -itiöitä. Kaksi valmisteita useita vuosia nauttinutta henkilöä sai vakavan
maksavaurion vuonna 2009. Heistä vanhempi kärsi akuutista pahoinvoinnista sekä sai
sapensalpausoireita, hepatiitin ja maksakirroosin nautittuaan nestemäisiä ravintovalmisteita
kolmen vuoden ajan. Keski-ikäinen nainen puolestaan sairastui sappifibroosiin ja -kirroosiin
nautittuaan kiinteitä valmisteita kuten vitamiinitabletteja toista vuotta. Kolmen eri
ravintovalmisteen todettiin kontaminoituneen B. subtilis -bakteereilla. Myös yhdessä
avaamattomassa valmisteessa havaittiin B. subtilis -kontaminaatio. Valmisteista eristetyt
mikrobit identifioitiin 16S rRNA- ja gyrB- sekvenoinnein B. subtilis -bakteerikannoiksi. Yksi
bakteerikanta osoittautui HepG2-solujen avulla suoritetuissa hepatologisissa testeissä
maksatoksiseksi in vitro. Sappifibroosin todettiin aiheutuneen nimenomaan valmisteiden
pitkäaikaiskäytöstä. Mitään raskasmetalleja, pestisidejä tai vastaavia kontaminantteja ei
näytteistä löydetty. (Stickel ym. 2009.)
2.5.2. Ruokamyrkytykset
Yleisesti ei-patogeenina pidetyn B. subtilis -bakteerin taudinaiheuttamis- ja toksiinien
tuottokyky ja siten yhteys elintarvikevälitteisiin infektioihin on selkeä. Ruokamyrkytysten
esiintyvyys on kuitenkin matala.
2.5.2.1. B. subtilis -ryhmän bakteerien esiintyvyys elintarvikkeissa
B. subtilis -ryhmän bakteerikantojen itiölöydökset ovat tavallisimpia maitotuotteissa, erityisesti
maitojauheessa, sekä vilja- ja lihatuotteissa. Raaka-aineissa esiintyy B. subtilis -bakteerikantoja
siinä kuin useita muitakin ympäristöbakteereja. Itiöinti mahdollistaa Bacillus -bakteerien
säilymisen elintarvikeprosessien aikana. Lämpöresistentit itiöt muuntuvat vegetatiivisiksi
soluiksi hyvin nopeasti kuumennetun elintarvikkeen jäähdyttämisen, tarjoilun ja nauttimisenkin
aikana. Infektion aiheuttanut elintarvike on tutkimusten mukaan usein valmistettu astiassa,
jossa on tarjoiltu lihaa tai kalaa yhdessä viljapohjaisen komponentin kuten riisin tai jonkin
leivonnaisen kanssa. (Logan 2011.)

21
Meijerituotteet ovat hyvin tavallinen Bacillus -bakteerilähde. Jakauma on kirjavaa. Esimerkiksi
eri meijerituotteissa esiintyneistä Bacillus -lajeista B. subtilis -bakteerikantoja havaittiin
Cosentinon ym. (1997) mukaan olevan 7,4 %, B. licheniformis -bakteerikantoja 11,8 % ja B.
pumilus -bakteerikantoja 6,2 %. B. subtilis -löydöksiä tehtiin pastöroidusta maidosta,
prosessoiduista juustoista ja juustolevitteistä ja muita tarkasteltavia bakteerikantoja myös
iskukuumennetusta maidosta sekä tuorejuustosta. Kuitenkin vaikka B. subtilis -ryhmän
bakteerikantojen esiintyminen maitotuotteissa on hyvin tavallista, mikrobipitoisuuden on
oltava vähintään 107 pmy/ml, jotta merkittäviä määriä toksiinia voi maidossa muodostua
(Griffiths 1990). Cosentinon ym. (1997) suorittamassa tutkimuksessa mikrobipitoisuus oli
hyvin alhainen vaihdellen välillä 101 - 103 pmy/ml. Mataranten ym. (2004) mukaan tulisi silti
huomioida, että Bacillus -bakteeripitoisuus määritetään tavallisesti pastöroinnin jälkeen, joten
lasketaankin vain itiöt ja osa soluista. Sen vuoksi nautittavien solujen kokonaispitoisuus on
tuntematon ja niiden potentiaalinen riski aiheuttaa ruokamyrkytys saatetaan aliarvioida.
Leivässä itiöivien Bacillus -bakteerien esiintyminen on myös hyvin tavallista. Tutkittaessa eri
bakteeripitoisuuksia valkoisessa ja täysjyvävehnäleivässä havaittiin erityisesti B. subtilis
-bakteerikantojen selviävän valmistusprosessista hyvin. Tutkittavat leivät oli valmistettu ilman
säilöntäaineita tai hapattamista. Niitä säilytettiin kahden vuorokauden ajan 25–30 oC:ssa, minkä
jälkeen niistä eristettiin seuraavia bakteerikantoja: B. subtilis (70 %), B. licheniformis (24 %),
B. pumilus (2 %) ja B. cereus (2 %). Mikrobipitoisuudet olivat noin 106 pmy/ml. B. subtilis
-bakteerikannat olivat vallitsevia todennäköisesti korkean lämpöresistenttiyden ansiosta.
Pilaantuneessa leivässä havaittiin ainoastaan B. subtilis -bakteerikantoja. Leipien raaka-
aineissa, kuten hiutaleissa ja leseissä, havaittiin Bacillus -bakteerien itiöitä 0–1×102 pmy/ml.
(Rosenkvist ja Hansen 1995.)
Mataranten ym. (2004) mukaan valmistusprosessi ja sitä kautta mikrobipitoisuus valmiissa
tuotteessa sen nautintahetkellä on merkitsevä tekijä ruokamyrkytysriskiä arvioitaessa.
Tutkittaessa kahdeksan kappaletta sekä käsin että teollisesti valmistettuja eurooppalaisia
kestomakkaroita havaittiin 54 % niistä eristetyistä mikrobeista olevan B. subtilis- ja 38 %
saman ryhmän B. pumilus -bakteerikantoja. Mikrobipitoisuudet olivat kuitenkin niin pieniä,
noin 103 pmy/g, ettei pitoisuuden katsottu aiheuttavan ruokamyrkytyksen vaaraa. Tulosten
perusteella on päätelty, että kestomakkaroiden hyvin pitkä valmistusaika (vähintään 2 kk)
laskee niistä saatavan ruokamyrkytyksen riskiä. Eristetyt B. subtilis -bakteerikannat
osoittautuivat kuitenkin α-hemolyyttisiksi, ja niistä 67 %:lla ilmeni fosfolipaasi A2 -aktiivisuus.
Enterotoksiinien tuottoon vaadittavia geenejä hbl tai nhe ei havaittu.

22
Maitokarjan utaretulehdukset eli mastiitit ovat eräs maitotuotteissa esiintyvien Bacillus
-bakteerien lähde (Nieminen ym. 2007). Meijeriin tulevista utaretulehduksen sairastaneiden,
parantuneiksi todettujen lehmien maitonäytteistä on eristetty B. licheniformis- ja B. pumilus
-organismeja. Mastiittitapauksissa Bacillus -bakteerit luetaan harvinaisiksi taudinaiheuttajiksi
tai kontaminanteiksi eikä antibioottihoitoa kohdisteta niihin (Evira 2001, ETT 2012). Niemisen
ym. (2007) tutkimuksen mukaan kuusi prosenttia kotimaisista maitonäytteistä sisälsi sellaisia
B. licheniformis- ja B. pumilus -bakteerikantoja, jotka kykenivät tuottamaan lämpöresistenttiä
likenysiini A -sytotoksiinia. Koska B. licheniformis -bakteerikantojen itiöt eivät tuhoudu
pastöroitaessa ja mikrobien likenysiini A -tuotanto on mahdollista myös anaerobisesti
suolistossa, maitojauheen käyttämisen ilman huolellista kuumennusta on katsottu olevan
turvallisuusriski erityisesti vastustuskyvyltään heikon henkilön terveydelle (Mikkola ym.
2000).
Tunnetuin B. subtilis -ryhmän mikrobeja sisältävä riskielintarvike lienee kontaminoituneesta
äidinmaidonvastikejauheesta huolimattomasti valmistettu maito. Rowan ym. selvittivät vuonna
2001 julkaistussa tutkimuksessaan, kykenevätkö B. subtilis- ja B. licheniformis -bakteerikannat
tuottamaan B. cereus –bakteerikantojen aiheuttamassa ripulioireisessa ruokamyrkytyksessä
esiintyviä toksiineja hemolysiini-BL (HBL) ja enterotoksiini T:ä (BceT). Yksi B. subtilis- ja
kaksi B. licheniformis -bakteerikantojen genomeista sisälsivät toksiinituotantoon vaadittavia
hbl- ja bce –geenejä. Kyseiset bakteerikannat kykenivät metaboloimaan enterotoksiinia
äidinmaidonvastikkeesta valmistetussa maidossa sen sisältäessä maltodekstriiniä tai glukoosia.
Tutkittujen bakteerikantojen havaittiin olevan myös sytotoksisia Caco-2- ja Hep2 -solulinjoilla.
Toksisuus heikkeni kuumennuksen jälkeen. Tulokset korostavat enterotoksiinituotannon
riippuvuutta mediumin koostumuksesta sekä äidinmaidonvastikkeen huolellisen
kuumennuksen ja pikaisen tarjoilun merkitystä.
2.5.2.2. Ruokamyrkytysten esiintyvyys
Vaikka varmennettuja B. subtilis -bakteerikantojen aiheuttamia ruokamyrkytyksiä raportoidaan
harvoin, täsmällistä ja luotettavaa kokonaismäärää ei tiedetä. Syynä on mahdollisesti se, etteivät
ruokamyrkytyspotilaita hoitavat tahot aina erota infektion aiheuttanutta B. subtilis -ryhmän
bakteerikantaa B. cereus -bakteerikannoista.

23
Sekavuutta löydösten esiintyvyyden tarkastelussa aiheuttaa myös se, että ennen 1970-lukua
mitä tahansa kliinistä Bacillus -isolaattia kutsuttiin usein nimellä B. subtilis. Näin ollen pitkän
aikavälin epidemiologisiin tutkimustuloksiin on suhtauduttava varauksella. Viimeisten 30–40
vuoden aikana infektioiden aiheuttaja on nimetty raporteissa täsmällisemmin. Bakteriologian
kehitys voikin muuttaa raportoitujen tapausten suhteellista lukumäärää pitkällä aikavälillä.
Toisaalta on huomioitava immuniteetiltään heikentyneiden henkilöiden määrän jatkuva kasvu
esimerkiksi syöpäpotilaiden määrän kasvaessa ja eliniän pidentyessä. (de Boer ja Diderichsen
1991, Sanders ym. 2003, Logan 2011.)
Todennetut muiden kuin B. cereus -ryhmään kuuluvien Bacillus -bakteerien aiheuttamat
ruokamyrkytystapaukset ovat lähinnä B. subtilis -ryhmään kuuluvien, hyvin läheistä sukua sille
olevien B. licheniformis- ja B. pumilus -bakteerikantojen aiheuttamia. Elintarvikkeissa
esiintyvän B. subtilis -organismin aiheuttaman infektion edellytyksenä on pidetty Loganin
(2011) mukaan sekä huomattavan suurta mikrobipitoisuutta että bakteerikantojen mahdollista
lipopeptidituotantoa, joskin jälkimmäisen yhteys infektioihin on vielä osittain epäselvä. Riskin
muodostavat paikalliset ja yleisinfektiot immuunivasteeltaan heikkojen henkilöiden kohdalla,
kun taas organismin toksiinien tuotto on merkitsevää sekä terveillä että sairailla henkilöillä.
Infektiot ovat monimuotoisia. Tyypillistä on lyhyt, jopa 10 minuutin ja keskimäärin 2,5 tunnin
inkubaatioaika sekä rajut vatsaoireet, erityisesti raju oksentaminen ja voimakas vatsakipu.
Myös ripulia ja sappitietulehdusta on raportoitu. B. subtilis kykenee kolonisoimaan suoliston.
Immunosuppressiivisilla henkilöillä on esiintynyt vähäisesti myös bakteremiaa. Nautitun
elintarvikkeen mikrobipitoisuus vaihtelee tyypillisesti 105 ja 109 pmy/ml välillä. (Oggioni ym.
1998, Kramer ja Gilbert 1989, Logan 2011.)
Organismin aiheuttamien ruokamyrkytystapausten esiintyvyydeksi on vv. 1975–1986
raportoitu Yhdistyneissä Kuningaskunnissa 49 tapausta. Diagnosoinnin epätarkkuuden vuoksi
tieto ei ole eksakti, mutta tarkasteltuna jälkimmäisten esiintyvyystietojen kanssa osaltaan
suuntaa-antava. (Kramer & Gilbert 1989.)
Salkinoja-Salosen ym. vuonna 1999 ilmestyneeseen tutkimukseen oli koottu Suomen ja
Yhdistyneiden Kuningaskuntien alueelta yli 10 vuoden ajalta 210 B. licheniformis -bakteeri-
kantojen aiheuttaman ruokamyrkytyksen isolaattia. Niiden alkuperät olivat kirjavia: mastiittia
sairastaneiden lypsylehmien utareet, erilaiset maitotuotteet, riisi, lihapiirakat ja voileivät.

24
Yksi isolaatti oli peräisin ruokamyrkytykseen menehtyneen imeväisen nauttimasta
äidinmaidonvastikkeesta. Yksi puolestaan oli peräisin ruokamyrkytyspotilaan ulosteesta.
Organismien aiheuttama ruokamyrkytys oli ollut akuutti tai jopa puolen vuorokauden kuluttua
ruoan nauttimisesta havaittu. Elintarvikkeista tutkimuksessa eristettyjen bakteerikantojen
pitoisuus vaihteli 3×105 – 1×108 pmy/g välillä, mutta nautitun ravinnon todelliset
mikrobipitoisuudet elimistössä olivat tuntemattomia. Isolaateista valmistettujen solujen
metaboliittia, likenysiiniä sisältävien uutteiden sytotoksisuus arvioitiin solujen hemolyysin,
Corynebacterium renale -solujen kasvun inhibition ja karjun siittiöiden liikkuvuuden estotestin
avulla. Kaikista tutkituista isolaateista noin 7 % osoittautui β-hemolyyttisiksi ja 6,2 %
sytolyyttisiksi. Puolet hemolyyttisistä – eli 4,3 % kaikista – isolaateista inhiboi siittiöiden
liikkeen häiritsemällä solun energiametaboliaa kuluttamalla solujen ATP:n loppuun ja
laajentamalla akrosomit. Vaikutukset korreloivat pitoisuuden kanssa. Mastiittia sairastaneen
naudan raakamaidosta eristetyistä kolmesta isolaatista kaksi osoitti siittiötestissä samanlaisia
toksisia vasteita kuin ruokamyrkytyksen aiheuttaneesta äidinmaidonvastikejauheesta eristetty
bakteerikanta. Äidinmaidonvastikejauhe luokiteltiin imeväisen riskielintarvikkeeksi.
Vuonna 2000 äidinmaidonvastikkeessa esiintyneet B. licheniformis -bakteerikannat aiheuttivat
sitä nauttineen imeväisen kuoleman. Karjun siittiöiden liikkuvuuden estotestissä havaittiin, että
sekä eristetyt isolaatit että niistä uutettu lipopeptidi, likenysiini A olivat erittäin sytotoksisia.
Siittiöiden plasmamembraani- ja akrosomivauriot olivat selkeitä ja niiden liikkuvuus heikkeni
huomattavasti. Solujen NADH ja ATP vähenivät myös selkeästi. Likenysiini A:n toksisuus oli
kymmenkertainen itse isolaattiin nähden. Likenysiini A:n ja vertaillun kaupallisen surfaktiinin
vaikutus oli sama. (Mikkola ym. 2000.)
Vuonna 2005 kaksi pientä lasta, iältään 9 ja 12 kk, olivat nauttineet kaupallista muro-
kuivahedelmä-maitojauhevalmistetta, johon sekoitetaan ennen tarjoilua vettä. Lapset olivat
sairastuneet akuutisti useaksi päiväksi rajuun oksentamiseen ja pahoinvointiin. Myös samasta
pakkauksesta uuden erän tuotetta valmistanut ja nauttinut toisen lapsen äiti sai
pahoinvointioireita. Valmisteen maitojauheessa ja muroissa havaittiin merkitseviä määriä sekä
B. cereus- että B. subtilis -bakteerien itiöitä. B. cereus -bakteerikantojen ei havaittu tuottavan
organismin oksennusmuotoa aiheuttavaa emeettistä toksiinia, kereulidia. Näytteissä havaittiin
enterotoksiineja koodaavia geenejä hbl ja nhe, mutta potilaat eivät kärsineet ripulista. Toisen
eristetyn B. subtilis -bakteerikannan todettiin olevan hemolyyttinen. Lasten ikä, oireet ja
löydökset viittasivat siihen, että ruokamyrkytyksen syynä oli valmisteen sisältämä B. subtilis
-itiöiden korkea pitoisuus yhdessä B. cereus –itiöiden kanssa. (Duc ym. 2005.)

25
Useissa ruokamyrkytyksissä osallisena olleiden B. subtilis- ja B. mojavensis -bakteerikantojen
toksiinintuottokykyä oli tutkittu Fromin ym. tutkimuksessa, joka ilmestyi vuonna 2005.
Bakteerikantoja oli eristetty erilaisista ruokamyrkytystapauksista, joissa niiden oli todettu
olleen ainakin yhtenä osatekijänä. Tutkimukseen sisältyi 333 isolaattia, joista 37 % oli peräisin
erilaisista elintarvikkeista kuten tyhjiökypsennetyistä ruoista, elintarviketeollisuuden raaka-
aineista ja valmispizzoista. Esimerkiksi pakastetusta, marinoidusta kanasta eristetyn B. subtilis
-bakteerikannan oli arvioitu aiheuttaneen usealle sitä nauttineelle henkilölle ruokamyrkytyksen.
Oirekuvaan kuului myös ripuli osoittaen organismin lisääntyneen myös suolistossa. Lähes
viidennes bakteerikannoista oli peräisin mausteista. Kolmannes oli eristetty vesihanasta
otetusta juomavedestä, mistä ne voivat kulkeutua elintarvikkeisiin. Toksiinin tuottokyvyn
tutkimukset Vero-solutestillä osoittivat, että vain alle 2 % organismeista tuotti lämpöresistenttia
sytotoksiinia. Karjun siittiöiden liikkuvuuden estotestin ja nestekromatografi-
massaspektrofotometrin (LC-MS) avulla suoritettujen emeettisen toksiinin analyysitulosten
perusteella pääteltiin, että alle 1 % bakteerikannoista tuotti emeettistä toksiinia. Yhdeksi
emeettisen toksiinin tuottajaorganismiksi osoittautui B. subtilis. Enterotoksiinin tuottokykyä ei
ilmennyt. Ruokamyrkytystapausten matalasta esiintyvyydestä huolimatta terveysriskinä
mainitaan erityisesti hitaasti jäähdytetyt, viljapohjaiset elintarvikkeet. (From ym. 2005.)
Apetroaie-Constantin ym. (2009) tutkivat 26 eri elintarvikkeesta peräisin olevia isolaatteja 94
kpl. Näytteet oli koottu vuosiväliltä 1990–2006. Niistä 76 oli eristetty
ruokamyrkytystapauksista. Osa aterioista oli tarjottu kypsänä ja osa raakana, osa kylminä ja osa
lämpimänä. Kahdesta ruokamyrkytystapauksesta peräisin olevat kuusi isolaattia, 6 % kaikista,
osoittautui lämpöresistentteja lipopeptidejä amylosiinia sekä surfaktiinia sekä aerobisesti että
anaerobisesti tuottaviksi Bacillus -bakteerikannaksi. Viisi niistä luokiteltiin B. subtilis- ja yksi
B. mojavensis -bakteerikannaksi. Yksi amylosiinia tuottanut B. subtilis -bakteerikanta oli
peräisin Yhdistyneiden Kuningaskuntien alueelta valmisbroileriateriasta, ja se oli aiheuttanut
nauttijalleen oksennus- ja ripulioireita. Viisi ruokamyrkytystä oli peräisin Suomessa myydyistä
kurpitsa-curry -aterioista. Näistä neljän aiheutti B. subtilis ja yhden sen lähisukulainen B.
mojavensis. Valmisbroileriateriasta eristetyn B. subtilis -bakteerikannan ja yhdestä kurpitsa-
curry -ateriasta eristetyn B. mojavensis -bakteerikannan todettiin metaboloivan sekä surfaktiinia
että amylosiinia, jonka osuus oli suurempi. Amylosiini todettiin sytotoksiseksi sekä Caco-2
-solulinjan tulosten että siittiöiden liikkuvuuden estotestin perusteella, ja sen havaittiin toimivan
toksiinina hyvin samankaltaisesti kuin ituriiniryhmän lipopeptidit, kuten likenysiini.

26
Myös B. subtilis -ryhmään kuuluva B. pumilus on aiheuttanut ruokamyrkytyksiä. Oirekuva on
samankaltainen kuin B. subtilis -bakteerikantojen aiheuttamissa infektioissa. Tyypillinen
inkubaatioaika on 2–11 tuntia ja oireita ovat raju oksentaminen ja huimaus sekä vatsakivut ja
ripuli (Kramer & Gilbert 1989). Kolmen kiinalaisessa ravintolassa aterioineen asiakkaan on
Fromin ym. (2007 b) mukaan raportoitu saaneen ruokamyrkytyksen B. pumilus -bakteerikannan
tuottamasta lipopeptidi pumilasidiinista. Akuutit oireet ilmaantuivat jo aterian aikana.
Muutamia tunteja myöhemmin potilaat saivat vatsakramppeja ja ripulin, mitkä kestivät useita
viikkoja. Ruokamyrkytyksen lähteeksi jäljitettiin edellisenä päivänä keitetty ja tarjoilupäivänä
uudelleen kuumennettu riisi. Esikypsennetty riisi oli säilytetty yli yön yli +10 oC:ssa ennen
kuumennusta ja tarjoilua. Riisistä eristettiin B. pumilus -soluja noin 105 pmy/ml. Solujen
todettiin tuottaneen sytotoksista pumilasidiini A:ta. Mikrobit tuottivat sitä koko testatulla
lämpötila-alueella, 10–37 oC:ssa. Pitkä säilytysaika +10 oC:ssa mahdollisti itiöiden
germinoitumisen, mikrobien lisääntymisen ja toksiinituotannon. Organismi kasvoi myös
anaerobisesti +37 oC:ssa, mikä on yhdenmukainen havainto pitkäkestoisten suolisto-oireiden
kanssa. Potilaiden oireet korreloivat suoraan nautitun riisimäärän kanssa.
2.6. Bacillus subtilis -bakteerikantojen tuottamat lipopeptidit
Eri Bacillus -alalajit kykenevät tuottamaan yli 30 erilaista amfipaattista lipopeptidiä. Niissä on
hydrofiilinen aminohappopää, jonka aminohapot ovat vaihtelevasti D- ja L -konfiguraatioissa
peptidisidoksin muodostaen syklisen rakenteen. Aminohapporenkaaseen on liittynyt
hydrofobinen rasvahappoketju. Lipopeptidit eroavat toisistaan toisinaan hyvinkin vähän
aminohappotähteiden lukumäärän ja konfiguraatioiden, aminohappoketjun rakenteen,
rasvahappoketjun pituuden sekä sen sisältämien hydroksyyli- ja metyyliryhmien suhteen.
Bacillus -bakteerien tuottamat lipopeptidit voidaan jakaa neljään ryhmään: surfaktiineihin,
fengysiineihin, ituriineihin ja kurstakiineihin (taulukko 1). Lipopeptidien tuottokyky on
toisaalta hyvin kantaspesifistä, toisaalta sama bakteerikanta voi tuottaa useita eri lipopeptidejä,
ja vaikutukset ovat usein synergistisiä. Mikrobin kasvuolosuhteet, kuten esimerkiksi
fermentointiprosessin aikana käytetty typenlähde, mediumin pH ja muut vaikuttavat siihen,
mitä lipopeptidiä / lipopeptidejä kulloinkin tuotetaan. (Ahimou ym. 2000, Volpon ym. 2000,
Wei ym. 2001, Ongena ja Jacques 2008, Jacques 2011.)

27
Taulukko 1. Bacillus -bakteerien tuottamat lipopeptidit (Jacques 2011).
Surfaktiiniryhmä Fengysiiniryhmä Ituriiniryhmä Kurstakiiniryhmä
Surfaktiini Fengysiini A Ituriini A Kurstakiini
Pumilasidiini Fengysiini B Ituriini AL
Likenysiini Plipastatiini A Ituriini C
Esperiini Plipastatiini B Mykosubtiliini
Bamylosiini A Basillomysiini D
Basillomysiini F
Basillomysiini L tai Lc
B. subtilis on hyvin tunnettu syklisten lipopeptidien tuottaja. Lipopeptidit ovat mikrobien
sekundäärimetaboliitteja. Ne syntetoidaan ei-ribosomaalisten peptidisyntetaasien välityksellä
(nonribosomally synthesized peptides, NRPS) tai polyketidisyntetaasien ja ei-ribosomaalisten
peptidisyntetaasien hybridin (PKS/NRPS) avulla. Syntetaasit ovat useista toiminnallisista
yksiköistä, moduuleista, koostuvia megaentsyymejä, jotka katalysoivat erilaisia peptidi- tai
polyketiditransformaatioon johtavia reaktioita. Jokainen moduuli on jaettu useisiin
katalyyttisiin domeeneihin, jotka vastaavat kustakin biokemiallisesta reaktiosta. Tyypillinen
NRPS-moduuli rakentuu tavallisesti noin tuhannesta aminohappotähteestä. Moduulit sisältävät
ne operonit, joilla koodataan kunkin lipopeptidin synteesiä katalysoivat entsyymit. Kukin
moduuli on vastuussa yhden aminohapon synteesistä. NRPS-biosynteesimekanismeihin
vaikuttaa kulloisenkin mediumin koostumus, mikä vaikuttaa tuotetun lipopeptidin lopulliseen
aminohappojärjestykseen. (Ongena ja Jacques 2008.)
Lipopeptidit toimivat useiden patogeenien antagonisteina ihmisillä, eläimillä ja kasveilla
vaikeuttamalla patogeenin pääsyä soluun muuttamalla solukalvon läpäisevyyttä sekä
kolonisoimalla kasvien juurimikrobistoa tai ihmisen suolistoflooraa (Mongkolthanaruk 2012).
Sykliset lipopeptidit ovat erittäin stabiileja ja kestävät hyvin kuumentamista ja pH-muutoksia
aktiivisuuden heikentymättä (Salkinoja-Salonen ym. 1999, Cao ym. 2009).
Tässä tutkielmassa on keskitytty B. subtilis -bakteerikantojen tuottamaan surfaktiiniryhmän
surfaktiineihin, fengysiiniryhmän fengysiineihin ja plipastatiineihin sekä ituriiniryhmän
mykosubtiliiniin.

28
2.6.1. Surfaktiinit
Surfaktiiniryhmään sisältyvästä noin 20 eri lipopeptidistä B. subtilis -bakteerin tuottama
surfaktiinit ovat tutkituimpia (Duitman 2003). Rakenteeltaan surfaktiini on syklinen
heptapeptidi, jonka kiraaliseen LLDLLDL-sekvenssiin on laktonisidoksella liittynyt 13–15 hiilen
pituinen β-hydroksirasvahappoketju (kuva 1). Surfaktiinit ovat rakenteeltaan hyvin
samankaltaisia kuin B. licheniformis -bakteerikantojen tuottama likenysiini A ja tunnetun
patogeenin, B. cereus -bakteerin tuottama kereulidi sekä B. pumilus -bakteerikantojen tuottama
pumilasidiini. (Mikkola ym. 2000.)
Kuva 1. Surfaktiini C15 -molekyylirakenne. Lähde: Liu ym. 2012.
Surfaktiinit jaetaan kuuteen muotoon, A-F, asemissa 2, 4 ja 7 esiintyviin alifaattisiin ryhmiin
kuuluvien aminohappotähteiden perusteella (kuva 2). Asemissa esiintyy valiinia, leusiinia,
isoleusiinia tai alaniinia L-konfiguraatiossa. Surfaktiinien molekyylipaino on 1050 ± 100 u.
(Arima ym. 1968, Oka ym. 1993.)
HT─CH─CH2─CO─L-Glu─L-X1─D-Leu─L-X2─L-Asp─D-Leu─L-X3─O
Kuva 2. Surfaktiinimolekyylin peptidisekvenssi. Asemassa X1 ja X3 esiintyy valiinia, leusiinia
tai isoleusiinia. Asemassa X2 puolestaan esiintyy alaniinia, valiinia, leusiinia tai isoleusiinia.
Lähde: Ongena ja Jacques 2008.

29
2.6.1.1. Synteesi
Surfaktiinien synteesiä säätelevät useat tekijät. Syntetaasia koodaa kuvassa 3 esitetty srfA
-operoni, joka koostuu neljästä lineaarisesta osasta. Sen ensimmäiset osat, srfA-A ja srfA-B,
rakentuvat kumpikin kolmesta moduulista, jonka ensimmäisen moduulin geenituotteen
aktivoituun aminohappoon rasvahappoketju lisätään. srfA-C sisältää puolestaan yhden
moduulin. Viimeiseen, srfA-D -geeniin, johon ei kokonaista moduulia sisälly, on koodattu
biosynteesin aloitusta stimuloivan entsyymin synteesi. (Ongena ja Jacques 2008)
C A P
C
P C A
P
C
P C A
P
C
P E
C A
P
C
P C A
P
C
P C A
P
C
P E
C A
P
C
P
T
e
Te/
At
Glu Leu Leu Val Asp Leu Leu
srfA-A srfA-B srfA-C srfA-D
Kuva 3. Surfaktiinioperoni srfA. Kolme surfaktiinin synteesiä koodaavaa moduulia ovat srfA-
A, srfA-B ja srfA-C. Kaksi ensimmäistä moduulia rakentuvat kolmesta osasta, ja niissä A-
domeeni vastaa adenylaatin aktivaatiosta, PCP-domeeni peptidyylin kuljettajaproteiinista ja C
peptidisidoksen muodostumisesta. Kolmas moduuli on yksiosainen, ja siinä esiintyvä Te
koodaa tioesterisidosten ja usein syklisen rakenteen muodostumisesta. Substraatin
epimerisaatiosta vastaa E-kirjaimella kuvattu domeeni. Neljännen osan muodostava srfA-D
-geeni koodaa tioesteraasi/asyylitransferaasia (Te/At) ja biosynteesin aloitusta. Lähde: Ongena
ja Jacques 2008.
Peptidin biosynteesiin osallistuvat myös erilaiset säätelytekijät kuten 4´-
fosfopanteteinyylitransferaasia koodaava sfp -geeni, itiön muodostusta säätelevät proteiinit
SpoOA ja AbrB sekä srfA -operonin transkriptionaalisen aktivaattorin, comA –geenin tuottama
operonin promoottorialueeseen sitoutuva säätelytekijä ComA (Strauch ym. 1990, McLoon ym.
2011). Synteesin alkuvaiheessa merkityksellisiä ovat myös ComP- ja ComQ -proteiinit
(Nakano ym. 1991, Marahiel ym. 1993).

30
2.6.1.2. Ominaisuudet ja käyttö
Surfaktiinia on hyödynnetty teollisuudessa kymmeniä vuosia. Molekyyli karakterisoitiin
osittain jo noin 50 vuotta sitten (Arima ym. 1968). Surfaktiinimolekyylit ilmentävät liuoksessa
tyypillistä hevosen satulan muotoa, mikä saa aikaan niiden laaja-alaisen biologisen
aktiivisuuden ja tekee ne hyvin kiinnostavaksi sekä teollisuuden sovelluksiin että akateemisiin
tutkimuksiin (Peypoux ym. 1999). Peptidin korkeaa bioaktiivisuutta lisää myös
aminohapporenkaan ja rasvahappoketjun yhdistävä laktonisidos (Mikkola ym. 2000).
Surfaktiini on tunnettu erityisesti voimakkaana biosurfaktanttina, joka kykenee vähentämään
veden pintajännitystä jo 20 µM pitoisuudessa 72 mN/m:sta 27 mN/m:iin. Lisäksi sillä on hyvät
emulsifikaatio- ja vaahdottamisominaisuudet. (Peypoux ym. 1999.)
Fermentaation aikana tapahtuvassa B. subtilis -bakteerikantojen surfaktiinituotannossa
tehokkaita hiilenlähteitä ovat glukoosi, sakkaroosi ja fruktoosi. Jatkuvassa viljelmätuotannossa
määrää lisäävät myös rauta ja manganaasi. (Peypoux ym. 1999, Schallmey ym. 2004.)
Elintarviketeollisuudessa mikrobien tuottaman surfaktiinin ominaisuuksia hyödynnetään
probioottituotannossa. Surfaktiini toimii signaalimolekyylinä, joka vahvistaa biofilmin
rakennetta ja lisää näin B. subtilis -bakteerin sietokykyä ruoansulatuskanavan haastavissa
olosuhteissa (Branda ym. 2001). Ahimoun (2000) mukaan surfaktiinin tuotanto parantaa myös
organismin adheesiokykyä, sillä sen molekyylit kykenevät adsorboitumaan pinnoille ja
muokkaamaan niiden hydrofobisuutta. Hydrofobisuus vaihtelee kuitenkin bakteerikannan
mukaan.
Ravinnossa esiintyvän surfaktiinin vaikutukset ihmiselle ovat kaksijakoisia. Yhtäältä
surfaktiini laskee veren LDL-kolesterolipitoisuutta inhiboimalla verihyytymien
muodostumista, tuhoaa suolistopatogeenejä sekä kasvaimia in vitro (Oka ym. 1983, Schallmey
ym. 2004). Sen on myös havaittu tehoavan lukuisia ihmiselle ja eläimille haitallisia tai
kiusallisia sairauksia, kuten eri herpesinfektioita, suu- ja sorkkatautia sekä suolistotulehduksia,
aiheuttavia vaipallisia DNA- ja RNA-viruksia vastaan. Teho perustunee surfaktiinin kykyyn
tuhota virusvaippa muodostamalla solumembraanin lipidikerrokseen kationikanavia ja näin
invasoitua virukseen.

31
Invaasio aiheuttaa kuitenkin ristiriitaa surfaktiinin hyötyä arvioitaessa, sillä havaittu
hemolyyttisyys ja sytotoksisuus heikentää käyttösovellusten tuomia etuja ihmisten kohdalla.
Lisäksi teho on heikko runsaasti proteiinia sisältävissä elintarvikkeissa kuten soijatuotteissa.
(Vollenbroich ym. 1997b, Hong ym. 2008.)
Surfaktiini on vahvasti antiviraalinen ja kohtalaisen antibakteerinen. Sen kykyä erottaa
Mycoplasma -bakteerin solut muista bakteereista on hyödynnetty farmaseuttisten tuotteiden
laadun parantamisessa (Nissen ym. 1997, Vollenbroich ym. 1997a). Sen on havaittu myös
inhiboivan B. subtilis -bakteerikantojen tuottamaa ruoansulatusentsyymiä, virheellisesti
aktivoituessaan haimatulehdusta aiheuttavaa fosfolipaasi A2:a, ja toimivan näin tulehdusta
hillitsevänä tekijänä (Kim ym. 1998). Surfaktiinivalmisteiden potentiaalisiksi funktioiksi on
harkittu myös elinsiirtojen hylkimisreaktioiden estoa ja autoimmuunisairauksien hoitoa, sillä se
heikentää fagosytoosia inhiboimalla makrofaagien toimintaa suoraan ja välillisesti säätelevien
proteiinien toimintaa (Park ja Kim 2009).
Surfaktiinin on havaittu olevan myös tehokas perunan ja useiden vihannesten antifungaalinen
pestisidi. Sen vaikutusmekanismi perustuu usein kasvin juuristossa elävien B. subtilis -baktee-
rien aiheuttamaan systeemisen puolustusjärjestelmän stimuloimiseen (indusoitu systeeminen
resistanssi, ISR). Surfaktiini voi saada aikaan myös erilaisia entsymaattisia muutoksia kasvin
puolustusjärjestelmän metaboliassa. Vaikutus on havaittu sekä käyttämällä suoraan
tuottajabakteerikantoja että niistä uutettua surfaktiinia kohdekasvin mukaan vaihdellen.
(Ongena ja Jacques 2008.)
Surfaktiini uutetaan usein mikrobiviljelmästä yleensä klassisilla metodeilla kuten saostamalla,
uudelleenkristallisoinnilla tai uuttamalla orgaaniseen liuottimeen. Tutkimuskäyttöön peptidiä
on uutettu myös kaksifaasiuuton, ultrasuodatuksen ja kiinteäfaasiuuton avulla. (Peypoux ym.
1999.)
2.6.2. Muut B. subtilis -ryhmän bakteerikantojen tuottamat lipopeptidit
B. subtilis -ryhmän bakteerikannat tuottavat sopivissa olosuhteissa myös muita lipopeptidejä.
Organismi kykenee tuottamaan fengysiinejä, plipastatiineja, ituriineja ja mykosubtiliinia
(Jacques 2011). Lisäksi B. subtilis ja B. amyloliquefaciens kykenevät tuottamaan sekä
basillibaktiinia että basilysiiniä eli antikapsiinia (May ym. 2002, Rajavel ym. 2009).

32
Lineaarista lipopeptidiä amylosiinia kykenee tuottamaan niiden lisäksi B. mojavensis (Mikkola
2006, Apetroiae-Constantin ym. 2009). Syklisiä lipopeptidejä ovat puolestaan likenysiini A,
jota tuottavat B. licheniformis ja B. pumilus, sekä B. pumilus -bakteerikantojen tuottama
pumilasidiini A (Duc ym. 2005).
2.6.2.1. Fengysiinit ja plipastatiinit
Suurimmat B. subtilis -bakteerin tuottamat lipopeptidit ovat dekapeptidejä: fengysiiniryhmään
luokiteltuja fengysiinejä ja hyvin samankaltaisia plipastatiineja (kuva 4a). Fengysiinin
makrolaktonirenkaan kymmenen aminohappotähdettä ovat konfiguraatiossa LDLDLDLLDL, ja
siihen on kiinnittynyt 14–18 hiilen pituinen β-hydroksirasvahappoketju, kun taas plipastatiinin
rasvahappoketju sisältää 11–13 hiiliatomia (Vanittanakom ym. 1986, Duitman 2003).
Fengysiinit ja plipastatiinit eroavat toisistaan myös aminohapporenkaiden asemassa 3
sijaitsevien tyriinitähteiden stereokemian osalta (kuva 4b). Sekä fengysiinejä että plipastatiineja
on kahta muotoa, A ja B, jotka luokitellaan aseman 6 aminohapon perusteella: A-muodossa se
on D-alaniini ja B-muodossa D-valiini. (Vanittanakom ym. 1986, Volpon ym. 2000)
4a
Fengysiini A L-Glu–D-Orn–D-Tyr–D-aThr–L-Glu–D-Ala-L-Pro–L-Gln–L-Tyr–L-Ile
Fengysiini B L-Glu–D-Orn–D-Tyr–D-aThr–L-Glu–D-Val-L-Pro–L-Gln–L-Tyr–L-Ile
Plipastatiini A L-Glu–D-Orn–L-Tyr–D-aThr–L-Glu–D-Ala-L-Pro–L-Gln–L-Tyr–L-Ile
Plipastatiini B L-Glu–D-Orn–L-Tyr–D-aThr–L-Glu–D-Val-L-Pro–L-Gln–L-Tyr–L-Ile
4b
Kuva 4. Fengysiini A -molekyylin rakenne osakuvassa 4a sekä fengysiienien ja plipastatiinien
aminohapposekvenssit osakuvassa 4b. Lähteet: NaPDoS 2011, Ongena ja Jacques 2008.

33
Fengysiini syntetisoidaan fen -operonilla koodattavien NRPS-syntetaasein. Operoni koostuu
viidestä geenistä, fenA-E (Ongena ja Jacques 2008). Plipastatiinin biosynteesiin vaaditaan viisi
avointa lukuraamia sisältävä syntetaasioperoni ppsA-E (kutsuttu myös pli -operoniksi) sekä
geenit degQ ja sfp (Tsuge ym. 1999, Ongena ja Jacques 2008). Plipastatiinia B. subtilis
-bakteerikannat kykenevät syntetoimaan L-glutamaatin ollessa ainoa typenlähde (Volpon ym.
2000).
Sekä fengysiinien että plipastatiinien ominaisuuksista tunnetuimpia ovat antibakteerisuus ja
antifungaalisuus. Useiden entsyymien, kuten immuunipuolustukselle merkityksellisen, mutta
toksisen fosfolipaasi A2:n inhiboiva vaikutus on myös molemmille ominaista. Fengysiinit ja
plipastatiinit vähentävät sitä kautta tulehduksia, akuutteja yliherkkyysreaktioita ja
verisuonitukoksia. (Vanittanakom ym. 1986, Peypoux 1999, Volpon ym. 2000.)
Fengysiinillä on havaittu olevan myös antikarsinogeenisiä vaikutuksia, ja siitä valmistetulle
syöpälääkevalmisteelle onkin jätetty patenttihakemus (Vanittanakom ym. 1986, Yin ym. 2013).
Viljelykasvien taudinkestävyyden parantamiseen fengysiinistä on merkittävää hyötyä. Paras
teho on saavutettu käsittelemällä kasveja fengysiiniuutteella, joka muuttaa kasvin
puolustusjärjestelmään liittyvää metaboliaa (Ongena ja Jacques 2008). Elintarvikkeissa
esiintyvän fengysiinin haittana on kuitenkin sen hemolyyttisyys, joskin heikohko
(Vanittanakom ym. 1986).
2.6.2.2. Mykosubtiliini
Ituriiniryhmään kuuluva mykosubtiliini on kuvassa 5 esitetty syklinen heptapeptidi, jonka
aminohappopään α-aminohapoista on muodostunut pysyvä kiraalinen LDDLLDL-sekvenssi,
johon on liittynyt 14–17 hiilen pituinen alifaattinen β-aminorasvahappoketju (Peypoux ym.
1986, Maget-Dana ja Peypoux 1994).

34
5a
Mykosubtiliini L-Asn–D-Tyr–D-Asn–L-Gln–L-Pro–D-Ser–L-Asn
5b
Kuva 5. Mykosubtiliinin rakenne (5a) ja aminohapposekvenssi (5b). Lähteet: Ongena ja
Jacques 2008, NaPDoS 2011.
Mykosubtiliinisyntetaasin muodostavat proteiinit MycA, MycB ja MycC. Ne syntetoidaan
PKS-NRPS -hybridikompleksin avulla. Syntetaasia koodaava hybridioperoni mycS koostuu
neljästä avoimesta lukuraamista: fenF ja mycA-C. Myc-geenit vastaavat aminohappotähteiden
yhdistämisestä, ja fenF koodaa malonyyli-CoA -transasylaasia. Operonin insertionaalinen
mutageneesi saa aikaan mykosubtiliini-negatiivisen fenotyypin. Sponaanistikin tapahtuva
mahdollinen profagi-insertio voi aiheuttaa mutaatioita ja peittää säätelyelementtien
koodisekvenssejä. Tämä koskee myös mykosubtiliinia hyvin tuottavaa referenssikantaa B.
subtilis ATCC 6633. Fengysiiniä ja mykosubtiliinia syntetoivat operonit kykenevät siirtymään
niitä tuottavien bakteerikantojen välillä. (Duitman 1999, 2003; Ongena ja Jacques 2008.)
Mykosubtiliinin on todettu olevan hemolyyttinen, antibakteerinen ja vahvasti antifungaalinen.
Sen vaikutusta tehostaa kyky lisätä kohdesolun solukalvon huokoisuutta. (Maget-Dana ja
Peypoux 1994, Duitman 2003)

35
3. NATON ELINTARVIKETURVALLISUUSARVIOINTI
Naton fermentoinnissa käytettävät B. subtilis -klusterin bakteerikannat sisältyvät EFSA:n
elintarviketurvallisiksi arvioimien mikrobien nk. QPS-listalle (Qualified presumption of
safety). Aasialaista perinneruokaa nattoa nautitaan myös Euroopassa. Koska Euroopan
elintarviketurvallisuusviranomainen EFSA katsoo niiden lisäämisen elintarvikeketjuun
vaativan toksikologista arviointia ja virulenssitekijöiden esiintyvyyden seurantaa sekä
suosittelee käytetyn B. subtilis -bakteerikannan identifiointia (EFSA 2007, 2013; Wang ym.
2007), myös B. subtilis var. natto -bakteerikantojen käytön toksikologinen seuranta on
perusteltua, vaikkei elintarviketurvallisuusarviointia Euroopan ulkopuolella edellytetäkään.
Arvioinnin perusteena on B. subtilis -bakteerikantojen ominaisuuksien perusteella nautittujen
mikrobien kokonaispitoisuus, niiden tuottamien toksiinien, etenkin surfaktiinin tuottokyky ja
sytotoksisuus sekä suolistokolonisaatio.
3.1. B. subtilis var. natto -bakteerikantojen käyttöturvallisuus
Bacillus -bakteerien muodostaman potentiaalisen infektioriskin muodostaa niiden yleisyys,
nopea itiöityvyys ja siten elinkyvyn säilyminen kuivissa, kuumissa ja ravinneköyhissä
olosuhteissa sekä kyky mahdollisen elintarvikekontaminaation jälkeen lisääntyä kypsennetyssä
ateriassa ja tuottaa sopivissa olosuhteissa toksiineja (de Boer ja Diderichsen 1991, Apetroaie-
Constantin ym. 2009). Lisäksi toksiinit ovat lämpöresistenttejä, joten ne eivät tuhoudu
kuumentamalla elintarviketta jälkeenpäin. Nattoaterioissa esiintyvien B. subtilis var. natto
-bakteerikantojen ei tällä hetkellä tiedetä aiheuttaneen infektioita (von Wright, Itä-Suomen
Yliopisto, suullinen tiedonanto 1.10. 2013).
3.1.1. Nattoaterioiden B. subtilis var. natto -kokonaispitoisuus
Likimäärin 75 % japanilaista syö nattoa vähintään kerran viikossa ja puolet heistä keskimäärin
joka kolmas päivä (Murooka ja Yamshita 2008). Ikedan ym. (2006) mukaan japanilainen nainen
nauttii nattoa tyypillisesti yhdestä kahteen annosta eli 56–80 grammaa viikossa. Lopullisessa
tuotteessa B. subtilis -pitoisuus on korkea, 108 - 1010 pmy/g fermentointiolosuhteista riippuen,
jolloin nautittu mikrobimäärä on noin 6×109 – 8×1011 pmy per annos(Wei ym. 2001).

36
Laskennallisesti kolmen neljänneksen japanilaisen pitkäaikainen B. subtilis natto -saanto on siis
3×109 – 4×1010 pmy vuodessa, ja lähes 40 prosentin 7×109 – 1×1012 pmy vuodessa.
Yksilötasolla saantoa voi nostaa hieman mahdollinen itiöprobioottien nauttiminen sekä naton
käyttäminen aromaattisena ainesosana muissa aterioissa ja kastikkeissa (Wei ym. 2001). Tämä
tuo hieman epätarkkuutta nautittavien mikrobimäärien arviointiin etenkin selvitettäessä saantoa
yksittäisellä kyselyllä.
Infektiivinen B. subtilis -pitoisuus immuniteetiltaan heikentyneille henkilöille on vähintään 105
pmy/ml ja terveille vähintään 107 pmy/ml (Griffiths 1990, Oggioni ym. 1998, Logan 2011).
Näin ollen ateriassa esiintyvä korkean mikrobipitoisuuden voidaan epäillä aiheuttavan
potentiaalisen ruokamyrkytysriskin etenkin immunosuppressiivisille henkilöille, jotka nauttivat
vähintään kohtalaisen kokoisen aterian, mikäli organismin toksiinintuotto on merkittävää.
Japanin kansalaiset pitävät nattoa arkisena, terveellisenä ja turvallisena elintarvikkeena jo
pienille lapsille. Suhtautumista kuvastaa esimerkiksi se, että B. subtilis var. natto -mikrobi-
peräistä, fermentoitua K2-vitamiinivalmistetta annettiin Nishiguchin ym. (2002) tutkimuksessa
jo vastasyntyneille. EFSA on katsonut, että kyseinen K2-vitamiiniöljyvalmisteen nauttiminen
on turvallista myös imeväisille, ja sitä voidaan lisätä elintarvikkeisiin, myös vauvan ruokaan ja
äidinmaidonvastikkeeseen (EFSA 2008). Viitteitä sen toksisuudesta ei ole tullut esiin.
Valmisteen koe-eläimille haitattomaksi NOAEL-arvoksi (No Observed Adverse Effect Level)
on arvioitu 20 mg/kg/vrk (Ikeda ym. 2006).
Ateriakokonaisuudellakin on oma merkityksensä infektioriskin arvioinnissa. Nattoateria
nautitaan useimmin riisin kera. Aasiassa yleinen tapa on esikypsentää suuri määrä riisiä ja
säilyttää se huoneenlämmössä. Tällä pyritään minimoimaan jäähdyttämisen aikana tapahtuvaa
kokkaroitumista. Riisissä luontaisesti esiintyvät Bacillus -bakteerien itiöt voivat säilyä
keittämisen ajan ja germinoitua lämpötilan laskiessa. Vegetatiiviset Bacillus -solut lisääntyvät
hyvin nopeasti juuri hiilihydraattipitoisessa mediumissa ja kykenevät tuottamaan hyvinkin
suuria pitoisuuksia toksisia lipopeptidejä. Myös naton fermentaatioprosessin
jäähdytysvaiheessa toksiinintuotanto on mahdollista. Näin ollen yksilön nauttimien mikrobien
ja tuotettujen toksiinien kokonaismäärä voi olla hyvinkin suuri. Vastustuskyvyltään heikkojen
henkilöiden riski saada ruokamyrkytys nattoateriasta voi näin kohota. (From ym. 2007b, Logan
2011.)

37
Kaneki ym. (2001) vertasivat naton kulutusta Japanin eri hallintoalueiden välillä. Erityisesti Itä-
Japanissa nattoa nautitaan tutkijoiden mukaan runsaasti arvioituna siihen käytettävän
vuosittaisen rahamäärän perusteella, joka on 2,5-kertainen Länsi-Japaniin verrattuna.
Tutkimuksessa mitattiin seerumin K2-vitamiinipitoisuus kahdeksalta vaihdevuosi-iän
ohittaneelta naiselta heidän nautittuaan yksi 80 gramman natto-ateria. B. subtilis var. natto on
tehokas K2-vitamiinin tuottajamikrobi. Nattoa ei syöty kahteen viikkoon ennen ja jälkeen
tutkimuksen, ja verinäytteet otettiin ennen tutkimusta sekä 1, 3, 7 ja 14 vrk aterian nauttimisen
jälkeen. Seuraavana päivänä seerumin K2-vitamiinipitoisuus oli kohonnut lähes 20-kertaiseksi,
kolmantena tutkimuspäivänä se oli vielä 8-kertainen ja 14 vuorokauden kuluttuakin
kaksinkertainen lähtötilanteeseen verrattuna. Tutkimus kuvasi mikrobin tehokasta
lisääntymistä ihmisen elimistössä.
Epidemiologisen kohorttitutkimuksen Japanissa suorittivat Ikeda ym. (2006). Tutkimuksessa
seurattiin ravinnon K2-vitamiinipitoisuuden ja osteoporoosiin sairastumisen välistä yhteyttä.
Sen ohessa saatiin tietoa myös nautituista nattomääristä. Koehenkilöt olivat 20–79 -vuotiaita
naisia (n = 944), jotka jaoteltiin iän mukaan pre- ja postmenopausaalisiin ryhmiin. Seuranta-
aika oli 3 vuotta. Tutkimuksessa määriteltiin japanilaisen aikuisen nauttimaksi kerta-
annokseksi 40 g nattoa, minkä katsottiin edustavan tavanomaista määrää. Nautittava nattomäärä
arvioitiin laskemalla nautittujen 40 gramman nattopakkausten lukumäärä viikossa.
Premenopausaalisista koehenkilöistä 30 % söi 1-2 pakkausta eli 40–80 g viikossa, 22 % yli 2
pakkausta eli yli 80 g viikossa ja lähes puolet, 48 % ei lainkaan. Postmenopausaalisista
koehenkilöistä 36 % söi nattoa 40–80 g viikossa, 15 % yli 80 g viikossa ja 49 % ei lainkaan.
Keskimäärin premenopausaaliset naiset söivät nattoa lähtötilanteessa 1,4±1,8 pakkausta eli
56±72 grammaa viikossa ja seurannan aikana 1,3±1,7 pakkausta eli 52±68 grammaa viikossa.
Postmenopausaaliset söivät sitä aluksi 2,0±2,6 eli 80±104 grammaa ja seurannassa 2,1±2,7
pakkausta eli 84±108 grammaa viikossa. Iäkkäämmät naiset söivät siis nattoa lähtökohtaisesti
enemmän kuin nuoremmat. Tutkimustulokset vahvistivat aiempia tuloksia, joiden mukaan
naton nauttiminen voi suojata osteoporoosilta lisäten luuntiheyttä vaikkakin heikommin kuin
kliininen K2-vitamiinivalmiste, jonka K2-vitamiinipitoisuus on lähes satakertainen nattoon
nähden. Tutkimuksen oleellinen virhelähde on nautittujen ateriamäärien laskennan
epätarkkuus, sillä arvio perustui kyselytutkimuksen tuloksiin, sekä suuri keskihajonta. (Ikeda
ym. 2006)

38
3.1.2. Probioottien käyttöturvallisuus
Elintarvikeketjuun päätyvien probioottien arviointiin on laadittu kansainväliset ohjeet
(FAO/WHO 2002). Ne sisältävät muun muassa käytettävän bakteerikannan identifioinnin,
turvallisuusarvioinnin, toiminnallisten ominaisuuksien selvittämisen ja ohjeita tuotteen
ominaisuuksien informoinnista asiakkaalle.
Probioottien turvallisuutta arvioitaessa on merkityksellistä tuotteessa käytetyn bakteerikannan
huolellinen sekvenointi, sillä valmisteen sisältämä mikrobikoostumus saattaa poiketa
tuoteselosteessa ilmoitetusta ja luoda harhakuvia mikrobikannan toksisuudesta. Esimerkiksi
valmistajan mukaan B. subtilis -bakteerikannan itiöitä sisältävän probioottivalmisteen oli
todettu aiheuttaneen 1990-luvulla suolistotulehduksen 15-vuotiaalle munuaissairaalle lapselle
(Wallet ym. 1996). Spinosan ym. (2000) suorittamat sekvenoinnit sekä probiootti- että
infektioisolaateista osoittivat, ettei valmiste sisältänytkään väitettyä B. subtilis- vaan B. cereus
-bakteerikannalle läheistä sukua olevaa B. thuringiensis -bakteerikantaa. Samoin Greenin ym.
vuonna 1999 julkaistussa tutkimuksessa korostui huolellisen karakterisoinnin merkitys
turvallisuusarvioinnissa. Kahden yleisesti tunnetun probioottivalmisteen väitettiin
todenvastaisesti sisältävän B. subtilis -bakteerikantojen itiöitä, mutta ne olivatkin
todennäköisimmin B. pumilus- ja B. alcalophilus -bakteerikantojen itiöitä.
Akuutti- ja kroonisen toksisuustutkimuksen vähäisyyden lisäksi antimikrobisia aineita
tuottavien probioottien lääkkeenomaisen käytön on epäilty olevan riskialtista mahdollisen
antibioottiresistenttiyden leviämisen vuoksi. Erityisesti B. subtilis var. natto -bakteerikantojen
runsaasti tuottaman surfaktiinin aikaansaamaa antibioottiresistenttiyttä on pohdittu Hongin ym.
(2009) suorittaman tutkimuksen tulosten tarkastelussa. Antibioottiresistenttiyttä ei kuitenkaan
ole tutkimuksissa ilmennyt. Probioottien turvallisuuden arvioinnin ongelmana on Hongin ym.
(2009) mukaan myös akuuttitutkimuksen osalta LD50:n määrittämisen epätarkkuus.
Vaikka B. subtilis var. natto -itiövalmisteen käyttö tuotantoeläinten rehulisäaineena on arvioitu
turvalliseksi sekä tuotantoeläimille että niitä nauttiville ihmisille, tieto pitkäaikaisvaikutuksista
ihmisen elimistöön ja terveydelle on suppeaa. Tutkimukset osoittavat organismin muuttavan
eläinten suoliston mikrobikoostumusta, mutta tulosten perusteella on mahdotonta tehdä
päätelmiä käytön pitkäaikaisvaikutuksista. (Sun ym. 2011; Peng ym. 2012; Euroopan komissio
2013 a,b; Song ym. 2014.)

39
3.2. B. subtilis var. natto -bakteerikantojen tuottamien toksiinien aiheuttamat terveysriskit
B. subtilis -bakteerikantojen ei-patogeenisuus on kyseenalaistettu, ja ne voitaneenkin luokitella
opportunistisiksi patogeeneiksi. B. subtilis -bakteerien aiheuttamien ruokamyrkytysten
matalasta esiintyvyydestä huolimatta niitä sisältävien itämaisten fermentoitujen
elintarvikkeiden turvallisuusarviointi on mikrobien toksiinien tuottokyvyn vuoksi tärkeää. On
myös huomioitava, että B. subtilis -bakteerikantojen itiöt ovat lämpöresistentimpiä kuin B.
cereus –bakteerikantojen itiöt. (Apetroaie-Constantin ym. 2009.)
Ei-hemolyyttisen B. subtilis natto ATCC 15245 -bakteerikannan genomissa ei ole havaittu
esiintyvän tunnettuja enterotoksiinien tuotantoon vaadittavia eikä sfingomyelinaasin tai
fosfolipaasi A2:n tuotantoon vaadittavia geenejä (Bhat ym. 2013). Toksisuusarviointien
tulokset viitaavat siihen, että B. subtilis -bakteerikantojen infektiivisyys voisi perustua
nautittujen solujen suuren pitoisuuden lisäksi niiden hemolyyttisyyteen ja
aineenvaihduntatuotteiden, erityisesti surfaktiinin, toksisuuteen (EFSA 2011). Koska B. subtilis
var. natto omaa edellä mainittuja ominaisuuksia, terveysriskien arviointi keskittyy niiden
tarkasteluun.
Sekä kuumentamattomat että kuumennetut B. subtilis -isolaatit ovat Beattien ja Williamsin
vuonna 1999 ilmestyneessä tutkimuksessa osoittautuneet CHO-solulinjalla (Chinese Hamster
Ovarios) sytotoksisiksi. Hongin ym. (2008) suorittamassa tutkimuksessa B. subtilis var. natto
-bakteerikannan tuottama surfaktiini puolestaan osoittautui toksiseksi ihmisten Caco-2-, Hep-
2- ja HT29-16E -soluille. Organismi myös invasoitui hyvin epiteelisoluihin. Invaasio Hep-2-
soluihin oli merkitsevästi voimakkaampaa kuin tunnetusti hyvin invasoituvalla Listeria
monocytogenes -bakteerikannalla, mikä voisi Hongin ym. mukaan selittyä B. subtilis var. natto
-bakteerikannan surfaktiinituotannolla surfaktiinin rikkoessa epiteelisolun
fosfolipidimembraanin. Altistuksessa käytetty mikrobipitoisuus oli 107 – 108 pmy/ml. Marsuilla
ja kaniineilla suoritetut akuuttia ja subkroonista toksisuutta in vivo arvioivat koetulokset olivat
negatiivisia. Akuutin toksisuuden arviointikriteereinä käytettiin ruokahalun, painon ja
infektioherkkyyden muutoksia sekä kudosten patologisia muutoksia. Lisättäessä B. subtilis var.
natto -organismin itiöitä päivittäin 30 vuorokauden ajan kaniinien (n=6) rehuun suspensiona,
jonka itiöpitoisuus oli 109 pmy/ml, muutoksia niiden voinnissa ei havaittu.

40
Subkroonisen toksisuusarvioinnin kesto oli 17 vuorokautta ja arviointikriteereinä käytettiin
yleistä statusta, hematologisia ja kudosmorfologisia muutoksia sekä muutoksia ulosteessa. Koe
suoritettiin annostelemalla marsuille (n=10) oraalisesti ja kertaluonteisesti itiösuspensiota
pitoisuudeltaan 1012 pmy/ml. Seuranta-aika oli jaettu yhden viikon jaksoihin.
Mikkolan ym. (2000) karjun siittiöiden liikkuvuuden estotestin perusteella tekemien
havaintojen mukaan surfaktiini vaurioitti siittiöiden solukalvoa ja aktivoi akrosomireaktiota.
Puolet karjun siittiöistä lamaava surfaktiinipitoisuus (EC50) oli 5–7 µg/ml. Surfaktiinilla
havaitut sytotoksiset vaikutukset todettiin erilaisiksi kuin siittiöiden mitokondrioon
vaikuttavilla kereulidilla tai valinomysiinillä, rakenteellisesti toisilleen läheisillä toksiineilla,
joita tuottavat muut gram-positiiviset bakteerit, kuten B. cereus.
Surfaktiinin käyttöä patogeenisten, vaipallisten virusten torjunnassa tutkivat puolestaan
Vollenbroich ym. (1997b). Tutkimusten ohessa suoritettu nisäkässolujen
sytotoksisuusarvioinnin mukaan toksisuus voi vaikuttaa nisäkässolujen lisääntymiseen riittävän
suurella, yli 40 mM, pitoisuudella, kun altistusaika on vähintään kolme vuorokautta. Virusten
inaktivointi heikkeni mediumin proteiinipitoisuuden kasvaessa. Koska vaipallisten virusten
inaktivointi tapahtuu muutamassa minuutissa, pääteltiin, että surfaktiinia voidaan käyttää
paikallisessa virustentorjunnassa matalan proteiinipitoisuuden omaavissa tuotteissa, kuten
rokotteissa. Aiempien toksisuustutkimusten in vivo (hiirillä) perusteella oli asetettu surfaktiinin
LD50-arvoksi lihakseen annosteltuna 200 mg/kg ja oraalisesti 4000 mg/kg. Surfaktiinin
arvioitiin olevan niin lievästi toksinen, ettei sitä tarvitse täysin poistaa suun kautta nautittavista
tai lihakseen annosteltavista tuotteista. Toisaalta surfaktiini on hemolyyttinen ja inhiboi
fibriiniverihyytymiä, mikä on huomioitava lääketieteellisissä sovelluksissa.
Nishiguchi ym. (2002) tutkivat imettävien äitien nauttiman, B. subtilis natto -bakteerikannoilla
tuotetun K2-vitamiinivalmisteen vaikutusta vastasyntyneiden lasten K-vitamiinistatukseen.
Tutkimuksessa oli mukana 31 lasta, joiden äidit olivat saaneet 15 mg/vrk K2-vitamiinia
tabletteina kaksi viikkoa alkaen 14. päivästä synnytyksestä ja 46 lasta, joiden äidit eivät olleet
nauttineet sitä. Kaikki lapset saivat kaksi kertaa ensimmäisen elinviikkonsa aikana 2 mg K2-
vitamiinia siirappivalmisteena. Tutkimuksessa havaittiin, että K2-vitamiinisupplementtia
saaneiden äitien lasten syntymäpaino sekä äitien itsensä painoindeksi kohosi selvästi verrattuna
verrokkiryhmään. Maksatoksisuutta tai muita merkittäviä haittavaikutuksia ei havaittu, mutta
huomionarvoista on K2-vitamiinin erittyminen äidinmaitoon.

41
Avoimeksi kysymykseksi jää, erittyvätkö siihen myös organismin muut metaboliatuotteet kuten
toksiset lipopeptidit. Kuten aiemmin on mainittu, NOAEL-arvoksi arvioitiin 20 mg/kg/vrk.
Vuonna 2002 Kikuchi ja Hasumi arvioivat surfaktiini C:n LD50 -arvoksi yli 100 mg/paino-kg.
Tutkimus suoritettiin muutamalla 6-viikkoisella uroshiirellä, joille annettiin suonensisäisesti
suurina kerta-annoksina, suurimmillaan 100 mg/kg, surfaktiini C:tä. Altistuksen jälkeen eläimiä
seurattiin neljän viikon ajan. Hiirissä ei havaittu vireystilan, ruokahalun, painon tai
käyttäytymisen muutoksia. Tutkijat arvioivat tulosten ja pitkäaikaiskokemusten perusteella,
että yhdessä 30 g natto-aterian arvioitu noin 10 mg määrä surfaktiinia ei aiheuta ihmiselle
akuutteja terveyshaittoja.
Surfaktiinin C-muodon geno- ja kehitystoksisuustutkimustuloksia puolestaan julkaisivat
Hwang ym. vuonna 2008. Genotoksisuutta tutkittiin sekä in vitro bakteerireversiotestillä (nk.
Amesin testi) että in vivo luuydinmikrotumamenetelmällä. Bakteerireversiotesti suoritettiin
yhden Escherichia coli- ja neljän Salmonella typhimurium -bakteerikannan avulla. Mikrobeja
altistettiin viidellä eri surfaktiinipitoisuudella, tasavälein 312,5–5000 µg/alusta. Mutageenisuus
arvioitiin vähintään yhdellä toistolla käyttäen kolmea alustaa per annos. Tutkimuksessa
huomioitiin mahdollinen maksametabolia. Luuydinmikrotumamenetelmässä kahdeksan viikon
ikäisiä uroshiiriä ruokittiin 2000, 3000 ja 4000 mg/kg surfaktiiniliuoksilla kaksi kertaa yhden
vuorokauden ajan. Kummatkaan tutkimustulokset eivät antaneet viitteitä genotoksisuudesta.
Kehitystoksisuustutkimuksen tutkimusryhmät koostuivat kolmesta 14–15 raskaana olevasta
hiirestä sikiöineen. Testi suoritettiin raskausviikoilla 6–17. Kullekin ryhmälle annettiin
oraalisesti surfaktiini C -liuosta 125, 250 ja 500 mg päivittäin 12 vuorokauden ajan. Surfaktiinin
aiheuttamia emojen kuolemia, keskenmenoja, painoeroja tai kliinisiä oireita ei esiintynyt, kuten
ei sikiöepämuodostumiakaan tai muutoksia niiden ruumiinpainossa. Tutkimuksen perusteella
todettiin, että surfaktiini C:n kehitystoksisuudesta ei ole viitteitä pitoisuuden ollessa
korkeintaan 500 mg painokiloa kohti vuorokaudessa ja altistuksen 12 vuorokautta. (Hwang ym.
2008)
Hwang ym. (2009) tutkivat myös surfaktiini C:n subkroonisia vaikutuksia rottiin. Kuudesta
kahdeksaan viikon ikäiset rotat (n = 90) jaettiin kolmeen yhtä suureen (n = 30) ryhmään, joissa
kussakin oli puolet uroksia ja puolet naaraita. Rotat saivat oraalisesti surfaktiini C:tä 500, 1000
ja 2000 mg painokiloa kohti 28 vuorokauden ajan. Rottien fysiologista tilaa ja käyttäytymistä
tarkkailtiin päivittäin.

42
Kuolemia, myrkytyksiä, hematologisia tai käyttäytymismuutoksia ei havaittu tutkimusaikana
millään altistuspitoisuudella. Sitä vastoin korkein pitoisuus hidasti ruumiinpainon nousua
merkitsevästi molemmilla sukupuolilla. Syynä tähän oli oletettavasti surfaktiinin aiheuttamat
ohutsuolivauriot, mistä seurasi ravintoaineiden imeytymisen heikkeneminen. Myös maksa-
arvot ALP (alkaliinifosfataasi), ALAT (alaniiniaminotransferaasi) ja AST
(aspartaattiaminotransferaasi) kohosivat rotilla, joita altistettiin vähintään 1000 mg / paino-kg
surfaktiinia. ALP kohosi molemmilla pitoisuuksilla vähintään 50 %, jopa kaksinkertaisiksi.
ALAT kohosi noin 50 % ja AST vähintään kolmanneksen. Naarailla muutos oli pienempi kuin
uroksilla. Arvojen nousun aiheuttajaksi epäiltiin maksalaskimoissa esiintynyttä nekroosia.
Maksan massakin oli merkitsevästi suurempi vähintään 1000 mg/paino-kg saaneilla ryhmillä,
neljänneksen uroksilla ja viidenneksen naarailla. Tutkimuksen perusteella tehtiin päätelmä, että
toistuvat yli 1000 mg/paino-kg pitoisuudet surfaktiini C:tä voivat aiheuttaa maksakudokseen
rakenteellisia ja toiminnallisia muutoksia ollen näin maksatoksinen. Tutkimuksen perusteella
arvioitiin surfaktiinin NOAEL-tasoksi alle 500 mg/kg 28 vuorokauden altistusajalla.
Sytotoksisuusarvioinnissa on huomioitava myös Hwangin ym. (2009) mukaan surfaktiinilla
aiemmin havaitut veritulppien muodostumista estävät, fibrinolyyttiset ja antikoagulanttiset,
ominaisuudet in vitro tekevät siitä potentiaalisesti veritoksisen. Kyseiset ominaisuudet
vaikuttavat osaltaan systeemisten ja/tai aivoverenvuotojen syntyyn.
Tarkasteltaessa surfaktiinin sytotoksisuutta maksa- ja munuaissoluille (brl ja HEK-293)
fluoresenssimikroskopian avulla havaittiin sen aikaansaavan morfologisia muutoksia. Soluissa
esiintyi sekä apoptoottisia muutoksia että tuman tiivistymistä. Sitä vastoin MCF-7 -epiteeli-
solulinjalla merkitsevää toksisuutta ei havaittu. Arviointikriteerinä oli surfaktiinialtistuksen
jälkeinen LDH:n (laktaattidehydrogenaasi) vapautuminen. (Cao ym. 2009.)
Okan ym. (1993) mukaan nattoaterian surfaktiinipitoisuus on noin 0,35 mg/g. Keskimääräisessä
30–50 gramman kerta-annoksessa elimistöön päätyy siis 10,5–17,5 mg surfaktiinia. Ikedan ym.
vuonna 2006 julkaiseman tutkimuksen mukaan laskettuna keskimäärin 56–80 grammaa nattoa
viikossa eli 224–320 g kuukaudessa nauttivan japanilaisnaisen nattoaterioista saama
surfaktiinialtistus on 78–112 mg. Kikuchin ja Hasumin (2002) arvioiman akuuttia toksisuutta
ilmaisevan LD50-arvon, yli 100 mg/kg, mukaisesti 60 kg painava aikuinen voi nauttia
surfaktiineja ilman havaittavia haittavaikutuksia 6000 mg eli 6 g kerta-annoksen, ja 10-kiloinen
lapsi yhden gramman verran olettaen, että hänen elimistönsä toimisi kuten aikuisen.

43
Hwang ym. (2009) puolestaan päätyivät suosittelemaan surfaktiinin NOAEL-arvoksi alle 500
mg/kg/kk. Ilman havaittavia surfaktiinin aikaansaamia haittavaikutuksia 60-kiloinen aikuinen
voisi kuukauden ajan nauttia surfaktiinia korkeintaan 30000 mg, minkä hän saa noin 1700:sta
50 gramman annoksesta nattoa. Hyvin suoraviivaisesti arvioiden 10-kiloinen lapsikaan ei
kykene ylittämään NOAEL-suositusta: hänen kestokykynsä olisi näiden laskelmien perusteella
korkeintaan 5000 mg surfaktiinia eli 285 kappaletta 50 gramman ateriapakkausta kuukaudessa.
Mikrobien surfaktiinimetabolian lisäksi toksikologisessa arvioinnissa on syytä huomioida
mahdollinen muiden toksiinien, kuten muiden lipopeptidien ja fosfolipaasi A2:n tuottokyky
(Chu 1949; Matarante ym. 2004; Hong ym. 2008; EFSA 2011, 2013). Fosfolipaasi A2:n tuotto
tosin estyy yli 80-prosenttisesti 30 µM surfaktiinipitoisuudella 20 minuutissa (Kim ym. 1998).
3.3. Suolistokolonisaatio
Suolistokolonisaation merkitys naton fermentointiin käytettävien mikrobien
elintarviketurvallisuusarvioinnissa on vielä epäselvä. Jo useita vuosikymmeniä sitten on
havaittu B. subtilis -bakteerikantojen kykenevän kolonisoimaan suoliston mikrobiflooran
(Kramer ja Gilbert 1989), ja Hongin ym. (2008) mukaan B. subtilis var. natto itiöi hyvin
mahanesteessä ja suolilimassa sekä kykenee muodostamaan suolistossa biofilmin ja
invasoituvan suoliston epiteelisoluihin hyvin.
Hieman viitteitä organismin vaikutuksista suolistoon antaa Samanyan ja Yamauchin suorittama
in vivo -tutkimus (2002). He lisäsivät neljän viikon ajan kuivattuja B. subtilis var. natto -soluja
yhteensä 52 kananpojan rehuun ja tarkkailivat sen vaikutusta kananpoikien suoliston
histologiaan. Mikrobipitoisuudet olivat 108 – 1010 pmy/g eli samaa suuruusluokkaa kuin natto-
ateriassa. Rehut sisälsivät probioottia 0; 0,2; 0,5 ja 1 %. Tulokset osoittivat itiöitä saaneiden
kananpoikien suolen nukkalisäkkeiden painon nousevan sekä solujen pinta-alan ja mitoosin
lisääntyvän verrattuna kontrolliryhmään, joskaan ei merkitsevästi.

44
Song ym. (2014) puolestaan havaitsivat merkitsevän naudan suolistomikrobikoostumuksen
muuttumisen B. subtilis natto -bakteerikantojen nauttimisen tuloksena. Itiöprobiootin, jonka
mikrobipitoisuus oli 8,3×109 pmy/ml, lisäys 36 lypsylehmän rehuun kahdeksaksi päiväksi
muutti merkitsevästi suoliston mikrobipopulaation koostumusta. Huomionarvoista on
patogeenien väheneminen ja bifidobakteerien määrän lisääntyminen, joskin syy on vielä
tuntematon.
Kanekin ym. (2001) tekemien havaintojen mukaan 80 g kertaluonteisen natto-annoksen
syöneiden naisten ulosteen B. subtilis var. natto -mikrobipitoisuus vaihteli suuresti päivittäin,
ja organismi selviytyi suolistossa kohtalaisen hyvin. Vuorokauden päästä nauttimisesta
organismi havaittiin vain kolmasosalla, kahden vuorokauden kuluttua 66 %:lla ja seitsemän
vuorokauden kuluttua 66 %:lla kuudesta koehenkilöstä. Nattoa ei nautittu kahteen viikkoon
ennen ja jälkeen tutkimuksen.
3.4. Turvallisuuden arviointimenetelmät
Perinteiset turvallisuuden arviointimenetelmät, jotka perustuvat toksikologisiin eläinkokeisiin,
eivät sovellu valtaosaan elintarvikkeista eivätkä uusien elintarvikkeiden turvallisuuden
arviointiin ihmisten ja eläinten välisten metaboliaerojen ja niiden aiheuttamien tulosten
rinnastusvaikeuksien sekä eläinkokeiden eettisten ongelmien vuoksi (FAO/WHO 2002). B.
subtilis -ryhmän mikrobeja sisältävien elintarvikkeiden toksisuusarviointiin käytetään niiden
absoluuttisen määrän mittaamista suoraan elintarvikkeista ja niitä nauttineiden henkilöiden
ulosteista. Arviointia täydennetään epäsuorilla menetelmillä kuten hemolyysin tarkastelulla,
mahdollisesti tuotettujen sytotoksiinien esiintymistä mittaavilla menetelmillä
epiteelisolulinjoilla ja karjun siittiöiden liikkuvuuden estotestillä sekä toksiineja koodaavien
geenien sekvenoinnilla (From ym. 2007a). Organismin aineenvaihduntatuotteiden identifiointi
on merkityksellistä paitsi ruokamyrkytystapauksissa, joissa haetaan varmuutta siitä, ovatko
kyseessä B. subtilis -ryhmän metaboloimat lipopeptidit vai mahdolliset B. cereus -bakteerin
tuottamat toksiinit, myös sellaisen elintarvikkeen kohdalla, johon on tarkoituksellisesti lisätty
mikro-organismeja (Salkinoja-Salonen ym. 1999, Sanders ym. 2003).

45
3.4.1. Hemolyysi
B. subtilis hajottaa useiden nisäkkäiden verisoluja (Williams 1957). Elintarvikkeissa esiintyvän
hemolyysin katsotaan aiheutuvan SpoVG -geenin läsnäolosta organismin genomissa sekä
matriksista peräisin olevien solujen normaalimetaboliiteista kuten lipopeptideistä.
Hemolyyttisyys luokitellaan yhdeksi mikrobin virulenssitekijäksi. Vaikka B. subtilis -bakteeri-
kantojen hemolyysiaktiivisuus on vähäisempi kuin patogeeneiksi katsottujen organismien, se
katsotaan niin merkitykselliseksi, että se on määritettävä elintarvikkeiden probiootteina ja
fermentoinnin heräteviljelminä käytettävistä bakteerikannoista. (Matarante ym. 2004, EFSA
2011, Pan ym. 2013.)
3.4.2. Sytotoksisuus
B. subtilis -ryhmän bakteerikantojen tuottamien sytotoksisten lipopeptidien metabolaatio
voidaan havaita kvalitatiivisesti karjun siittiöiden liikkuvuuden estotestillä (Andersson ym.
1998, Beattie ja Williams 1999, Hoornstra ym. 2003). Menetelmää käytetään yleisesti
esimerkiksi Euroopan rekisteröintiasiakirjoissa arvioitaessa Bacillus -mikrobeja sisältävien
elintarvikelisäaineiden turvallisuutta (Apetroaie-Constantin ym. 2009), ja sitä suosittelee myös
Euroopan elintarviketurvallisuusvirasto (EFSA 2013).
3.4.3. Lipopeptidien tuottokyky
Lipopeptidien synteesiin vaadittavien geenien läsnäolon tunnistamiseksi mikrobien genomista
monistetaan kyseinen geenisekvenssi polymeraasiketjureaktion (PCR) avulla (Tapi ym. 2010).
Erittäin korkean erottelukyvyn omaava nestekromatografia (ultra high performance liquid
chromatography, UHPLC) on nopea ja tarkka yhdisteen sisältämien fragmenttien
erottelumenetelmä. Sen avulla B. subtilis -bakteerikantojen eri metaboliatuotteet voidaan
määrittää luotettavasti samanaikaisesti yhdestä näytteestä (Oka ym. 1993). Laitteistoon voidaan
yhdistää erilaisia detektoreja. Erittäin korkearesoluutioisella, nelinkertaisen nopeuden
omaavalla kvadrupoli-lentoaikamassaspektrometrillä qTOFMS (quadrupole Time Of Flight
Mass Spectrometer) -analysaattorilla on näytteen kvalitatiivisen ja kvantitatiivisen tiedon
tarkka ilmaisukyky (Kildgaard ym. 2014).

46
KOKEELLINEN OSA
4. TUTKIMUKSEN TAVOITTEET
Tutkimuksen tavoitteena oli arvioida kolmen perinteisen japanilaisen soijapavuista valmistetun
natto-aterian elintarviketurvallisuutta analysoimalla niiden tuottamisessa käytettyjen Bacillus
subtilis var. natto -bakteerikantojen aineenvaihduntatuotteina syntyneiden syklisten
lipopeptidien läsnäolo ja kvalitatiivinen sytotoksisuus. Koska aterioiden mikrobipitoisuus on
korkea ja niitä nautitaan usein runsaasti ja useiden vuosien ajan, tavoitteena oli lisäksi arvioida
turvallinen ilman haittavaikutuksia nautittava kerta-annos nattoa.

47
5. AINEISTO JA MENETELMÄT
Käytetyt arviointimenetelmät pohjautuvat Euroopan elintarviketurvallisuusviraston EFSA:n
voimassaoleviin ohjeisiin. Työ jakautui viiteen osaan. Natto-annospakkauksista eristettiin
fermentoinnissa heräteviljelminä käytetyt bakteerikannat, ja niiden lajimääritys suoritettiin 16S
rDNA -sekvenoinnin avulla. Hemolyyttisyys määritettiin naudan veriagarilla. Lipopeptidien
synteesiä ilmentävien geenien esiintyminen määritettiin polymeraasiketjureaktion avulla.
Bakteereista valmistettiin myös kaksi erilaista lipopeptidiuutetta, joiden sytotoksisuutta
arvioitiin karjun siittiöiden liikkuvuuden estotestillä. Lipopeptidien kvantitatiivinen analysointi
suoritettiin Tanskan teknillisessä yliopistossa UHPLC-qTOFMS -analysaattorin avulla.
5.1. Bakteerikantojen eristäminen nattoaterioista
Elintarvikenäytteinä oli neljä Japanista toimitettua 50 g annospakkausta nattoa. Kaksi näytettä
oli peräisin samasta erästä, joten niitä käsiteltiin rinnakkaisnäytteinä. Näytteiden mikrobistot
eristettiin ja viljeltiin Itä-Suomen yliopistossa valmistetuille tryptoni-soija- (TSA) ja plate
count- (PC) -yleiselatusalustoille. Laktoosia fermentoivien koliformisten bakteerien
havaitsemiseksi mikrobistot viljeltiin myös violettipuna-sappi-glukoosi- (VRBGA) ja hiivojen
ja homeiden erottamiseksi oksitetrasykliini- (OGYE) -elatusalustoille, jotka myös oli
valmistettu Itä-Suomen yliopistossa.
Näytteeksi otettiin kustakin annoksesta soijapapuja sekä niitä ympäröivää limaa yhteensä 1 g,
mihin lisättiin 4 ml peptoni-suolaliuosta. Näyte homogenoitiin Seward Stomacher® 80 -homo-
genisaattorilla käyttäen normaalia nopeutta 30 sekunnin ajan. Homogenoitua näytettä
pipetoitiin kullekin elatusalustalle 50 µl. VRBGA- ja OGYE-elatusalustoille valmistettiin
laimennossarjat -2…-4 sekä TSA- ja PC-alustoille -2…-8. Maljoja inkuboitiin +30 oC:ssa
vuorokauden ajan, minkä jälkeen TSA- ja PC-alustoilta laskettiin pesäkkeet ja niiden
perusteella mikrobitiheys pmy/g. PC-alustoilla parhaan kasvun omaavat pesäkkeet
(keskimäärin 109 pmy/ml) valittiin tutkimukseen, ja niistä tehtiin puhdasviljelmät toisen
vuorokauden inkuboinnin jälkeen. Mikrobien karkea luokittelu Bacillus -bakteereihin
suoritettiin pesäkkeiden morfologisten ominaisuuksien ja gram-värjäyksen perusteella.

48
5.2. Tutkimuksessa käytetyt referenssibakteerikannat
Tutkimuksessa käytettiin natto-näytteistä eristettyjen kolmen tutkittavan bakteerikannan lisäksi
positiivi- ja negatiivikontrolleina Itä-Suomen yliopistosta peräisin olevia
referenssibakteerikantoja sekä positiivikontrollikantana Chr. Hansen A/G:n toimittamaa
bakteerikantaa B. subtilis CHCC 16388. Kontrollibakteerikannat on esitetty taulukossa 2.
Taulukko 2. Natto-elintarvikkeessa esiintyvien Bacillus -bakteerien tuottamien lipopeptidien
toksisuusarvioinnissa käytetyt positiivi- (+) ja negatiivikontrollibakteerikannat (-). Lyhenne e.t.
merkitsee ei tutkittu.
Bakteerikanta Hemo-
lyysi
Siittiötes-
ti
Surfaktii-
nien
synteesi
Fengysii-
nien
synteesi
Plipastatii-
nien
synteesi
Mykosubtilii-
nien synteesi
B. subtilis
ATCC 6051 + + e.t. e.t. + e.t.
B. subtilis
ATCC 6633 + e.t. e.t. + e.t. +
B. subtilis
ATCC 21332 + + + e.t. e.t. e.t.
B. lichenifor-
mis ATCC
14580
- - e.t. e.t. e.t. e.t.
B. subtilis
CHCC 16388 + + + e.t. e.t. e.t.
Lactobacillus
plantarum E
71084
e.t. e.t. - - - -
5.3. Identifiointi
Natto-näytteistä A, C ja D eristetyt isolaatit identifiointiin aluksi organismien hiilenlähteisiin
perustuvalla GEN III -menetelmällä valmistajan ohjeiden mukaan (GEN III MicroPlate;
Biolog, Hayward, Kanada). Tarkempi lajimääritys suoritettiin sekvenoimalla isolaatit ns.
uuden sukupolven 16S rDNA -menetelmällä DNA eristettiin kitin avulla (NucleoSpin® Extract
II, Macherey-Nagel, Germany) sen ohjeen mukaisesti. Kukin bakteerikanta rikastettiin
inkuboimalla 1 µl bakteerimassaa 10 ml:ssa tryptoni-soija-lientä (TSB) 24 tuntia +37 oC:ssa.
Suspensio sekoitettiin koeputkisekoittajalla ja 1 ml sitä sentrifugoitiin kahden minuutin ajan
11000 rpm nopeudella. Pelletti suspensoitiin 180 µl:aan puskuriliuosta (20 mM Tris-HCl + 2
mM EDTA + 1 % Triton × 100). Suspensioon lisättiin 75 µl lysotsyymiä (50 ng/ml).

49
Näytettä inkuboitiin 1 tunnin ajan 37 oC:ssa, minkä jälkeen siihen lisättiin 25 µl proteinaasi K:ta
ja inkuboitiin vielä kahden tunnin ajan 56 oC:ssa. Eristetyn DNA:n sekvenssi AGA GTT TGA
TCM TGG CTC AG – TAC GGY TAC CTT GTT ACG AC monistettiin
polymeraasiketjureaktion (PCR) avulla käyttäen universaalialukkeita 27-f ja 1492-r (Lane ym.
1991). Saatu PCR-tuote puhdistettiin em. kitin protokollan mukaisesti, ja riittävä puhtaus sekä
DNA-saanto varmistettiin Nanodrop-spektrofotometrillä. Puhtaat PCR-tuotteet lähetettiin
sekvenoitavaksi Agovalle Saksaan.
5.4. Lipopeptidien uuttaminen mikrobistosta
Bakteerien metaboloimat lipopeptidit uutettiin kahdella eri menetelmällä, joista ensimmäinen
oli Abdel-Mawgoudin ym. (2008) sekä Ahimoun ym. (2000) käyttämien metodien mukaelma.
Metaboliatuotteet uutettiin sekä koostumukseltaan Ahimoun ym. (2000) mukaiseen MOLP-
elatusliemeen (Medium for optimal lipopeptide production) Abdel-Mawgoudin ym. (2008)
menetelmän mukaisesti että 2-butanoliin Tanskan teknillisen yliopiston professori Kristian Fog
Nielsenin henkilökohtaisen ohjeen mukaan (liite 1). Valmiit uutteet säilytettiin +7 oC:ssa.
5.4.1. Uutto MOLP-elatusliemeen
Aluksi valmistetiin elatusliemeksi MOLP (liite 2). Surfaktiini analysoitiin sekä epäsuoralla
uuttomenetelmällä näytteestä eristetyistä Bacillus -isolaateista että suoralla uutolla natto-
ateriasta Abdel-Mawgoudin ym. (2008) mukaan.
5.4.1.1. Uutteen valmistaminen mikrobipesäkkeestä
Mikrobipesäke siirrostettiin PC-alustalta 25 ml:aan brain-heart-infusion-kasvatuslientä (BHI).
Inokulaattia inkuboitiin 24 tunnin ajan 30 oC lämpötilassa 150 rpm ravistelussa (Orbital Shaker
SO1), minkä jälkeen 600 µl inokuloitiin 30 ml:aan MOLP-mediumia. Suspensiota inkuboitiin
72 tuntia 30 oC:ssa 150 rpm nopeudella ja ultrasentrifugoitiin viiden minuutin ajan 10000 g:n
voimalla 5 oC:ssa (Avanti J-301, roottori JA-30.50; Beckman Coulter, USA). Supernatantista
käytettiin 20 ml. Sen pH laskettiin 6M HCl:lla kahteen.

50
Lipopeptidien annettiin sakkautua 7 oC lämpötilassa vähintään 18 tuntia, minkä jälkeen
supernatanttia sentrifugoitiin 4100 rpm nopeudella 5 oC:ssa 25 minuutin ajan (Jouan BR4i,
Thermo Scientific). Pelletti suspensoitiin 99 % etanoliin, ja etanoli haihdutettiin 70 oC:ssa
typpivirrassa (N-EVAP™, Organomation Associates, USA). Kuivaan näytteeseen lisättiin 5 ml
dikloorimetaania (DCM), ja sitä inkuboitiin huoneenlämmössä 18 tuntia hitaasti ravistellen.
Ekstrakti suodatettiin ja uutettiin uudelleen 5 ml:aan dikloorimetaania kahdesti. DCM
haihdutettiin typpivirrassa 40 oC lämpötilassa. Surfaktiini eluoitiin 1 ml:aan 5 mM Tris-HCl
-puskuriliuosta (pH 8,5) ja säilytettiin 7 oC:ssa.
5.4.1.2. Uutteen valmistaminen nattonäytteestä
Kunkin natto-annospakkauksen soijapavuista sekä niitä yhdistävästä B. subtilis -bakteerien
tuottamasta limasta valmistettiin vertailunäyte. Sytotoksisuustestin negatiivikontrolliksi
valmistettiin uute steriilistä, maustamattomasta säilykesoijapavusta (maahantuoja Inex
Partners). Seward Stomacher®80 -homogenisaattorilla (Seward Laboratory Systems, USA)
homogenoitiin 10 g näytettä ja 25 ml peptoni-suolavettä keskisuurella nopeudella
silmämääräisesti arvioiden homogeeniseksi. Näytettä ultrasentrifugoitiin 5 minuuttia 10000 g:n
voimalla 5 oC:ssa. Ekstraktin valmistus jatkui samoin kuin eristetystä mikrobipesäkkeestä.
5.4.2. 2-butanoliuutto
PC-alustalta siirrostettua mikrobipesäkettä inkuboitiin 25 ml:ssa BHI-lientä 24 tuntia 30 oC
lämpötilassa 150 rpm nopeudella ravistellen, minkä jälkeen 10 ml:aan suspensiota lisättiin 12
ml 2-butanolia (Merck) ja ravistelua jatkettiin 15 minuutin ajan (liite 1). Sentrifugoinnin (4000
rpm, 10 min.) jälkeen 10 ml supernatanttia otettiin talteen. Jäljelle jääneeseen supernatanttiin
(10 ml) lisättiin jälleen 12 ml 2-butanolia, ja ravistelu sekä sentrifugointi toistettiin. Saatu
supernatantti yhdistettiin edelliseen, 2-butanoli haihdutettiin typpivirrassa, ja kuiva pelletti
eluoitiin 1ml:aan 5 mM Tris-HCl -puskuriliuosta (pH 8,5). Uute säilytettiin 7 oC:ssa.

51
5.5. Hemolyysin määrittäminen
Bakteerikantojen fenotyyppistä hemolyyttisyyttä tarkasteltiin naudan erytrosyyttien avulla
Euroopan elintarviketurvallisuusviraston ohjeiden mukaan (EFSA 2011). PC-alustalta
siirrostettiin pesäke naudan verilevylle (bioTRADING, Benelux B.V., Mijdrecht, Alankomaat),
ja sitä inkuboitiin 3 vrk 30 oC:ssa. Verisolujen hajoamista seurattiin vuorokauden välein.
Positiivikontrollikantoina toimivat B. subtilis -bakteerikannat ATCC 6051, ATCC 6633 ja
ATCC 21332 (Hsieh ym. 2004, Dehghan-Noudeh ym. 2005, EFSA 2011). Tutkimukset
toistettiin kahdesti.
5.6. Sytotoksisuuden määritys karjun siittiöiden liikkuvuuden estotestillä
Sytotoksisuustestit suoritettiin sekä suoraan natto-annospakkauksista että niistä eristetyistä
Bacillus-bakteerikannoista valmistetuilla surfaktiiniuutteilla Anderssonin ym. (1998)
kehittämää karjun siittiötestiä mukaillen. Tutkimukseen sisältyi kolme siittiötestiä, joissa
käytettiin vaihtelevia näytekokoonpanoja. Abdel-Mawgoudin ym. (2008) menetelmän mukaan
Bacillus -isolaateista A MOLP-elatusliemeen valmistetut surfaktiiniuutteet rinnakkais-
näytteineen testattiin kahdesti sekä isolaateista C ja D valmistetut uutteet kolmesti, joista
viimeisessä oli mukana rinnakkaisnäytteet. Suoraan natto-aterioista Abdel-Mawgoudin ym.
(2008) menetelmällä MOLP-elatusliemeen uutetut näytteet A, C ja D testattiin kahdesti. Sekä
kaupalliset surfaktiininäytteet (c = 5 µg/ml ja 10 µg/ml) että 2-butanoliin uutetut pesäkenäytteet
A, C ja D rinnakkaisnäytteineen testattiin yhden kerran.
Positiivikontrollikantoina toimivat B. subtilis ATCC 6051, B. subtilis ATCC 21332 ja B.
subtilis CHCC 16388. Lisäksi positiivikontrollinäytteenä käytettiin siittiöiden liikkuvuuden jo
nanomolaarisina pitoisuuksina estävää dimetyylisulfoksidiin (DMSO) liuotettua
valinomysiiniä (Sigma-Aldrich®, Saksa). (Mikkola ym. 2000, Hoornstra ym. 2003.)
Negatiivikontrolleina käytettiin maustamattomasta säilykesoijapavusta valmistettua uutetta
sekä kahdesti suodatettua dikloorimetaania. Positiivisena vertailunäytteenä käytettiin
kaupallista surfaktiinia S3523 (Sigma-Aldrich®, Saksa) liuotettuna 99 % etanoliin
pitoisuudeltaan 10 mg/ml liuokseksi (Mikkola ym. 2000).

52
Karjun siittiöiden liikkuvuuden estotesti suoritettiin altistamalla huoneenlämpöisillä uutteilla
huoneenlämpöistä karjun siemennestettä (Perussiemen 57, Tuomikylän karjuasema, Seinäjoki)
kolmen vuorokauden ajan. Polystyreenilinkoputkessa olevaan kevyesti ravisteltuun
siemennesteeseen (2 ml) lisättiin uutteet 10 µl/ml, 20 µl/ml ja 40 µl/ml pitoisuuksiksi.
Valinomysiinikontrollien altistuspitoisuuksina käytettiin 0,08 ng/ml; 0,2 ng/ml; 0,4 ng/ml ja
0,8 ng/ml. Kevyen ravistelun jälkeen näytteitä säilytettiin polystyreenilaatikossa
huoneenlämmössä. Siittiöiden liikkuvuus tarkistettiin päivittäin mikroskoopilla 20-kertaisella
suurennuksella (Olympus CK40, Japani) +37-asteisena.
5.7. Lipopeptidien synteesikyky
Natto-näytteistä eritettyjen B. subtilis -bakteerikantojen surfaktiinin, fengysiinin,
mykosubtiliinin ja plipastatiinin synteesikyky todennettiin monistamalla niistä synteesiin
tarvittava geenialue polymeraasiketjureaktion (PCR) avulla Tapin ym. (2010) kehittämän
menetelmän mukaan. Ohjeesta poiketen bakteerien DNA:ta ei laimennettu. Kunkin
bakteerikannan DNA eristettiin käytetyn kitin (Nucleo Spin®Extract II, Macherey-Nagel,
Saksa) menetelmäohjeen mukaan kuten kohdassa 4.2. PCR-ajoa varten valmistetut
reaktioseokset on kuvattu liitteessä 3 ja ajo-ohjelmat liitteessä 4.
5.7.1. Surfaktiinien synteesiin vaadittavan geenin määrittäminen
Surfaktiinien synteesiin vaadittavan geenin sfp tunnistamiseksi eristettiin aluksi DNA näytteistä
A, C ja D sekä positiivikontrolleista B. subtilis ATCC 21332 ja B. subtilis CHCC 16388 sekä
negatiivikontrollista Lb. plantarum E 71084 Tapin ym. (2010) menetelmän mukaan. DNA-
saannot sekä -puhtaudet absorbanssisuhteella 260/280 nm määritettiin Nanodrop ND-1000
-spektrofotometrilla (Nanodrop Technologies, USA), ja tulokset käyvät ilmi taulukosta 3.

53
Taulukko 3. Surfaktiinien synteesiin vaadittavan geenin tunnistamiseksi tutkituista
bakteerikannoista eristettyjen isolaattien DNA-pitoisuudet ja -puhtaudet. Kaikkien isolaattien
DNA-pitoisuus oli riittävä (≥ 10,0 ng/µl) sfp-geenin havaitsemiseksi.
Näyte Pitoisuus ng/µl Puhtaus (A260/280)
A 151,5 1,96
C 64,8 2,11
D 110,0 2,17
B. subtilis ATCC 21332 50,6 1,96
B. subtilis CHCC 16388 55,6 2,18
Lb. plantarum E 71084 95,0 2,06
Näytteistä monistettiin 419–431 emäsparin kokoinen DNA-sekvenssi CGC GGM TAC CGV
ATY GAG-ATB CCT TTB TWD GAA TGT CCG CC alukkeiden As1-F ja Ts2-R avulla.
PCR-tuote analysoitiin agaroosigeelielektroforeesilla 1,2 % agaroosigeelillä ajoparametreilla
80 V/1h/20 min. Monistustuotteen määrittämiseen käytettiin Gene Ruler™ 100 bp DNA Ladder
-molekyylipainomarkkeria (Fermentas).
5.7.2. Fengysiinien, plipastatiinien ja mykosubtiliinin synteesiin vaadittavien geenien
määrittäminen
Eristetyistä DNA-näytteistä määritettiin myös fengysiinin, mykosubtiliinin ja plipastatiinin
synteesiin vaadittavat geenit Tapin ym. (2010) menetelmän mukaan. Fengysiinien tuottamiseen
vaadittavan geenin fen (koko 443–455 ep) tunnistamiseksi monistettiin sekvenssiä GAA TAY
MTC GGM CGT MTK GA - GCT TTW ADK GAA TSB CCG CC alukkeilla Af2-F ja Tf1-R.
Plipastatiineja koodaavan geenin pps (koko 893–929 ep) havaitsemiseksi bakteerin DNA:ssa
puolestaan käytettiin alukkeita Ap1-F ja Tp1-R sekvenssillä AGM CAG CKS GCM ASA TCM
CC - GCK ATW WTG AAR RCC GGC GG. Mykosubtiliinin synteesiin tarvittavan geenin
myc (416–419 ep) läsnäolo analysoitiin monistamalla sekvenssiä CAK CAR GTS AAA ATY
CGM GG - CCD ASA TCA AAR AAD TTA TC alukkeiden Am1-F ja Tm1-R avulla.

54
5.8. Mikrobien tuottamien lipopeptidien analysointi
Uutteista analysoitiin lipopeptidit Tanskan Teknillisen Yliopiston toimesta Poroshell -fenyyli-
heksyylipylvään sisältävän erittäin korkean erottelukyvyn omaavan nestekromatografin (ultra
high performance liquid chromatography, UHPLC) Agilent 1290 -järjestelmän (Agilent
Technologies, Santa Clara, Kanada), ja kaksoissähkösumutuksella varustetun Agilent 6550
kvadrupoli-lentoaikamassaspektrometrin, qTOF-MS:n (quadrupole Time Of Flight Mass
Spectrometer), muodostamalla hybridianalysaattorilla (Juola ym. 2014). Ajoliuoksena toimi
asetonitriili. Kvantifiointi muodostettiin ionikromatogrammeista, jotka saatiin surfaktiinien
[M+H]+-ionien täysskannauksesta. Automaattinen tandem-massaspektrometri (MS/MS) antoi
MS/MS-spektrillä ionit m/z -arvoilla 950–1300.
Kontrollinäytteet valmistettiin 85 % asetonitriililiuokseen 0,5 grammasta soijajauhoa 50 µl:aan
S3523-surfaktiinia (Sigma-Aldrich®, Saksa) 3000; 1500; 750; 375; 188; 93,8; 46,9; 23,4 ja 0
µg/g -pitoisuuksiin. Seoksen sisältämät surfaktiinit uutettiin 2-butanoliin liitteen 1 mukaisesti
yön yli +4 oC:ssa inkuboinnin jälkeen. Kaupallisen surfaktiinin molekyylipaino oli 1036,34 u.
Eri surfaktiinien A1, B, C1 ja D suhteelliset osuudet määritettiin UHPLC-qTOF-MS -
analysaattorin avulla 200 nanometrin signaalilla. Positiivikontrollikantana käytettiin B. subtilis
CHCC 16388 -bakteerikantaa (Chr. Hansen A/S, Hørsholm, Tanska).

55
6. TULOKSET
6.1. Identifiointi
GEN III -identifikaatiotulokset osoittivat isolaattien A ja B olevan B. subtilis -bakteerikantaa ja
isolaatin C sille hyvin läheistä, B. subtilis -ryhmään sisältyvää B. vallismortis -bakteerikantaa.
Sekvenointitulosten perusteella kaikki isolaatit osoittautuivat 99 % todennäköisyydellä Bacillus
subtilis -lajeiksi. Tulokset ovat yhdenmukaisia morfologisten havaintojen kanssa, joissa
mikrobit osoittautuivat voimakkaasti PC-alustalla leviäviksi, endosporeja sisältäviksi gram-
positiivisiksi sauvabakteereiksi. Bakteerit kasvoivat myös TSA-yleisalustalla eivätkä lainkaan
spesifisillä VRBGA- tai OGYE-alustoilla. Fermentointiin katsottiin käytetyn yksittäistä
bakteerikantaa.
6.2. Hemolyysi
Kaikki isolaatit osoittautuivat sekä α- että β-hemolyyttisiksi (kuva 5). Hemin hajoamisesta
seuraava α-hemolyysi ilmenee pesäkkeen vihertymisenä, ja täydellisen hajoamisen eli β-
hemolyysin seurauksena verimalja on täysin kirkastunut pesäkkeen ympäriltä. Hemolyysi
ilmeni selkeästi jo yhden vuorokauden inkuboinnin jälkeen. Kuvasta voidaan havaita myös
bakteerikannalle ominainen pesäkkeiden voimakas leviäminen alustan pinnalla.
Kuva 5. Natto-näytteistä eristetyn Bacillus subtilis –bakteerin aiheuttama hemolyysi naudan
veriagarilla kolmen vuorokauden +30 oC:ssa inkuboinnin jälkeen. α-hemolyysin seurauksena
pesäkkeet ovat vihertyneet. Täydellisen, β-hemolyysin seurauksena kasvualusta on kirkastunut.

56
6.3. Sytotoksisuus
Sytotoksisuuden voimakkuuden kuvaamiseen käytettiin siittiöiden liikkuvuutta asteikolla -
(liikkuvuus 45–50 %) … +++ (liikkuvuus 0-10 %). Tulokset käyvät ilmi taulukosta 4. Puhtaan
siemennesteen siittiöiden liikkuvuus kunkin testin lähtötilanteessa oli 45–50 % merkiten hyvää
liikkuvuutta. Ensimmäisessä testissä käytetty siemenneste oli toimitettaessa viileää kylmän
vuodenajan vuoksi. Puolet karjun siittiöistä lamaannuttavan EC50-pitoisuuden määritykset
perustuvat testin lisäksi Tanskan teknillisen yliopiston natto-isolaateista ja kaupallisesta
surfaktiinista määrittämiin surfaktiinipitoisuuksiin, ja osoittavat, että nattoateriassa esiintyvä
jopa alle 0,2 µg/ml -surfaktiinipitoisuus on toksinen karjun siittiöille.
Taulukko 4. Natto-näytteistä ja niistä eristetyistä isolaateista valmistettujen surfaktiiniuutteiden
aikaansaama karjun siittiöiden liikkuvuuden esto ja sen perusteella määritetyt EC50-arvot.
Inhibitio on merkitty koodein - (liikkuvuus 45–50 %), + (liikkuvuus 30–40 %), ++ (liikkuvuus
15–25 %) ja +++ (liikkuvuus 0-10 %), ja e.t. merkitsee ei tutkittu.
Näyte Liikkuvuuden inhibitio
EC50 (µg/ml) MOLP-
mediumiin uutettu
2-butanoliin
uutettu
Isolaatti A +/++ +++ 0,238
Isolaatti C ++ +++ 0,140
Isolaatti D ++/+++ +++ 0,130
Natto A + e.t. e.t.
Natto C ++/+++ e.t. e.t.
Natto D +/+++ e.t. e.t.
Säilykesoijapapu - e.t. e.t.
B. subtilis ATCC 6633 +/++ e.t. e.t.
B. subtilis ATCC 6051 ++ +++ e.t.
B. subtilis ATCC 21332 ++ +++ e.t.
B. subtilis CHCC 16388 +/++ +++ 0,531
Surfaktiini S3525 5 µl/ml + +++ 0,018
Surfaktiini S3525 10
µl/ml + +++ 0,048
Valinomysiiniliuos ++/+++ ++/+++ 0,0002
DCM-suodos - - e.t.

57
6.4. Surfaktiinien synteesiin vaadittavan geenin esiintyminen
PCR ja agaroosigeelielektroforeesi osoittivat kaikkien tutkittujen isolaattien DNA:n sisältävän
surfaktiinien synteesiin vaadittavan sfp-geenin (kuva 6).
Kuva 6. Kolmesta natto-ateriasta eristettyjen Bacillus -isolaattien surfaktiinia koodaavien
geenien sfp (419-431 ep) esiintyminen bakteerien DNA:ssa. A1 ja A2 kuvaavat isolaatin A
vyöhykkeitä, C kuvaa isolaatin C ja D isolaatin D vyöhykettä. Positiivikontrollina toimineet B.
subtilis ATCC 21332 ja B. subtilis CHCC 16388 sekä negatiivikontrolli Lb. plantarum E 71084
on kuvattu bakteerikantoja vastaavin numerokoodein. Molekyylipainomarkkerina (M)
käytettiin Gene Ruler™ 100 bp DNA Ladder:a (Fermentas).
6.5. Fengysiinien, plipastatiinien ja mykosubtiliinin synteesiin vaadittavien geenien
esiintyminen
Natto-näytteistä eristettyjen B. subtilis -bakteerikantojen genomeissa ei havaittu esiintyvän
fengysiiniä, mykosubtiliinia tai plipastatiinia syntetoivien geenien läsnäoloa PCR-analyysin
perusteella. Yhteistyötahon suorittaman UHPLC-qTOF -analyysin tulokset tukivat PCR-
tutkimustuloksia (tuloksia ei esitetty).
M
A1
A2
C
D
AT
CC
21332
CH
CC
16388
M
0
E 7
10
84
500 ep

58
6.6. Nestekromatografiset tulokset
UHPLC-qTOF -analyysin tulokset osoittivat nattoaterioiden fermentoinnissa käytettyjen
mikrobien tuottaneen ainoana non-ribosomaalisena lipopeptidinä surfaktiinia (kuva 7).
Surfaktiinia esiintyi neljänä pitoisuuksiltaan hyvin vaihtelevina jakeina, A1, B, C1 ja D. Ne
havaittiin [M+H]+- ja [M+Na]+ -ioneina, joiden massa-alueen (m/z-arvo) vaihteluväli oli
1022.6756 ja 1044.6570 toleranssien mukaisesti.
Kuva 7. Natto-isolaatin A UHPLC-qTOF-MS -analysaattorilla havaitut surfaktiinifraktiot
(positiivinen sähkösumuionisointi ESI+ m/z –alueella 950-1500). Osakuvasta A voidaan
havaita surfaktiinijakeet A1, B, C1 ja D. Osakuvasta B ilmenee lipopeptidifraktioiden
havainnointi [M+H]+-ioneina sekä surfaktiini B -jakeen tarkka kromatografinen spektri.
Osakuva C osoittaa standardina käytetyn surfaktiini B -jakeen (Sigma-Aldrich®) aikaansaaman
MS-spektrin molekyylipainoltaan 1022,6756 u, ja osakuva D sille identtisen, isolaatin
tuottaman MS-spektrin. Viitearvo surfaktiinin molekyylipainolle on 1050 ± 100 u ja
standardina käytetyn surfaktiinin ilmoitettiin olevan molekyylipainoltaan 1036,34 u.
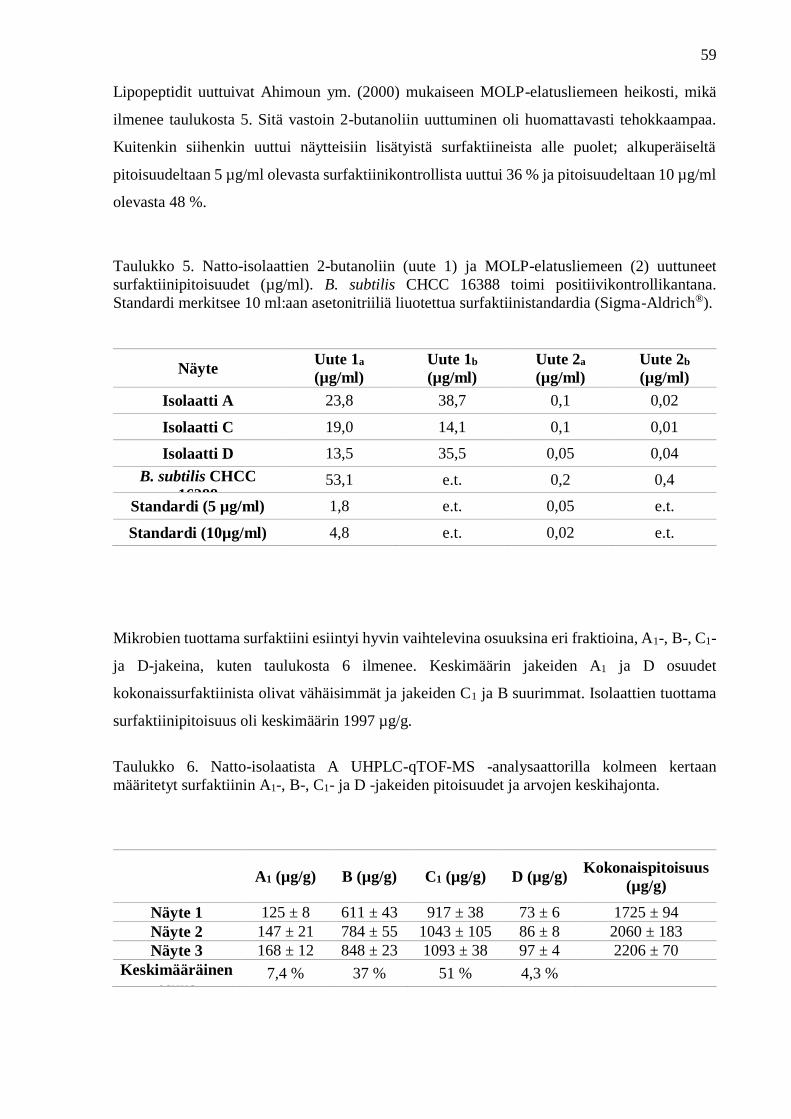
59
Lipopeptidit uuttuivat Ahimoun ym. (2000) mukaiseen MOLP-elatusliemeen heikosti, mikä
ilmenee taulukosta 5. Sitä vastoin 2-butanoliin uuttuminen oli huomattavasti tehokkaampaa.
Kuitenkin siihenkin uuttui näytteisiin lisätyistä surfaktiineista alle puolet; alkuperäiseltä
pitoisuudeltaan 5 µg/ml olevasta surfaktiinikontrollista uuttui 36 % ja pitoisuudeltaan 10 µg/ml
olevasta 48 %.
Taulukko 5. Natto-isolaattien 2-butanoliin (uute 1) ja MOLP-elatusliemeen (2) uuttuneet
surfaktiinipitoisuudet (µg/ml). B. subtilis CHCC 16388 toimi positiivikontrollikantana.
Standardi merkitsee 10 ml:aan asetonitriiliä liuotettua surfaktiinistandardia (Sigma-Aldrich®).
Näyte Uute 1a
(µg/ml)
Uute 1b
(µg/ml)
Uute 2a
(µg/ml)
Uute 2b
(µg/ml)
Isolaatti A 23,8 38,7 0,1 0,02
Isolaatti C 19,0 14,1 0,1 0,01
Isolaatti D 13,5 35,5 0,05 0,04
B. subtilis CHCC
16388 53,1 e.t. 0,2 0,4
Standardi (5 µg/ml) 1,8 e.t. 0,05 e.t.
Standardi (10µg/ml) 4,8 e.t. 0,02 e.t.
Mikrobien tuottama surfaktiini esiintyi hyvin vaihtelevina osuuksina eri fraktioina, A1-, B-, C1-
ja D-jakeina, kuten taulukosta 6 ilmenee. Keskimäärin jakeiden A1 ja D osuudet
kokonaissurfaktiinista olivat vähäisimmät ja jakeiden C1 ja B suurimmat. Isolaattien tuottama
surfaktiinipitoisuus oli keskimäärin 1997 µg/g.
Taulukko 6. Natto-isolaatista A UHPLC-qTOF-MS -analysaattorilla kolmeen kertaan
määritetyt surfaktiinin A1-, B-, C1- ja D -jakeiden pitoisuudet ja arvojen keskihajonta.
A1 (µg/g) B (µg/g) C1 (µg/g) D (µg/g) Kokonaispitoisuus
(µg/g)
Näyte 1 125 ± 8 611 ± 43 917 ± 38 73 ± 6 1725 ± 94
Näyte 2 147 ± 21 784 ± 55 1043 ± 105 86 ± 8 2060 ± 183
Näyte 3 168 ± 12 848 ± 23 1093 ± 38 97 ± 4 2206 ± 70
Keskimääräinen
osuus 7,4 % 37 % 51 % 4,3 %

60
7. POHDINTA
Tämän pro gradu -tutkielman tarkoitus oli tuottaa tietoa japanilaisen, Bacillus subtilis var. natto
-bakteerikantojen avulla fermentoidun perinteisen elintarvikkeen, naton sisältämien
lipopeptidien, erityisesti surfaktiinien pitoisuuksista ja akuutista sytotoksisuudesta in vitro, ja
arvioida tulosten pohjalta elintarvikkeen turvallisuutta.
Huolimatta nattoaterioiden sisältämistä korkeista mikrobipitoisuuksista ja samanaikaisesta
niiden tuottamiseen soveltuvien bakteerikantojen toksisten lipopeptidien tuottokyvystä
aterioiden ei tämän hetken tutkimustiedon valossa tiedetä aiheuttaneen ruokamyrkytyksiä. Sitä
vastoin niiden tuottamiseen soveltuvien bakteerikantojen lievästä toksisuudesta on hieman
viitteitä. Sen merkitys on kuitenkin vielä epäselvä. Tutkimus on vähäistä ja suppeaa, sillä
organismin aterioissa tuottamien surfaktiinien aiheuttamia haittavaikutuksia ei Japanissa
vaadita seurattavan eivätkä niiden mahdollisesti aiheuttamat infektiot tule välttämättä
tutkijoiden tietoon. Pitkäaikaisvaikutuksista väestötasolla tutkimustietoa ei ole lainkaan.
Avoimena kysymyksenä ovat myös naton nauttimisesta seuraavat mahdolliset sytofiiliset,
kuten hemolyyttiset ja hepatosellulaariset vaikutukset in vivo sekä suolistokolonisaation
merkitys.
Tutkimuksessa analysoitujen kolmen natto-aterian sisältämä B. subtilis var. natto -pitoisuus oli
tavanomaista suuruusluokkaa, 108 – 109 pmy/g. Pesäkkeet olivat yleisalustalla voimakkaasti
leviäviä, gram-positiivisia, itiöiviä sauvabakteereja. Muita organismeja ei viljelmin eikä
mikroskooppisin tarkasteluin havaittu, ja fermentointiin voidaan sekä sillä että sekvenoinnin
perusteella olettaa käytetyn yhtä B. subtilis -bakteerikantaa.
Kaikki isolaatit aiheuttivat täydellisen β-hemolyysin naudan verisoluille. Hemolyysiä
tarkasteltiin EFSA:n ohjeiden (2011) mukaisesti kolme vuorokautta, mutta se oli selkeästi
havaittavissa jo yhden vuorokauden inkuboinnin jälkeen. Vertailu positiivikontrolleihin tuki
tulosten oikeellisuutta.
Eristettyjen isolaattien genomien havaittiin sisältävän surfaktiinien synteesiin vaadittavan sfp-
geenin ja tuottavan ainoana syklisenä lipopeptidinä surfaktiinia tutkimusolosuhteissa.
Fengysiinien, plipastatiinien ja mykosubtiliinin synteesiin vaadittavia geenejä ei havaittu.

61
Tulokset eivät olleet niiltä osin täysin luotettavia, sillä ainoastaan fengysiinien
positiivikontrollibakteerikannan B. subtilis ATCC 6633 geenituotteen saattoi PCR-kuvassa
havaita, mutta sen, kuten näytteidenkin sitoutuminen oli erittäin epäspesifistä PCR-
reaktioseoksen koostumuksesta riippumatta. Plipastatiinien ja mykosubtiliinin
positiivikontrollibakteerikantojen geenituotteita ei havaittu. Tutkimus keskeytettiin tältä osin.
Tulosten perusteella ei voida luotettavasti tietää, esiintyikö tutkittujen organismien genomeissa
muita lipopeptidejä kuin surfaktiineja koodaavia geenejä. Koska mediumin koostumuksen on
havaittu vaikuttavan merkittävästi B. subtilis -bakteerikantojen lipopeptidien tuottokykyyn, se
ettei nestekromatografinen erottelu osoittanut kyseisiä lipopeptidejä tuotetun tutkituissa
aterioissa, ei takaa sitä, etteivätkö mikrobit olisi voineet erilaisessa mediumissa erilaisen
fermentointiprosessin aikana ja jälkeen niitä tuottaa.
Surfaktiinien saanto vaihteli voimakkaasti käytetyn uuttomenetelmän mukaan. Erityisesti
lipopeptidien uuttoon optimaaliseksi kehitettyyn MOLP-elatusliemeen ei surfaktiinia uuttunut
juuri lainkaan. Sitä vastoin 2-butanoliin surfaktiinit uuttuivat tehokkaammin. Saanto oli samaa
suuruusluokkaa, joskin hieman vähäisempi kuin kirjallisuudessa aiemmin esitetty. Näytteisiin
lisätyistä surfaktiineista uuttui alle puolet, 36 % ja 48 %. Kuten kirjallisuudessa on todettu,
nattoaterioissa esiintyvien lipopeptidien lopulliseen pitoisuuteen vaikuttaa voimakkaasti paitsi
fermentointiolosuhteet myös mediumina toimivan soijapapujen ja niitä yhdistävän liman
koostumus. Nattoaterioissa todellisuudessa esiintyvät surfaktiinipitoisuudet ovatkin
mahdollisesti tässä tutkimuksessa havaittua korkeampia. Koska surfaktiinit ovat solujen
aineenvaihduntatuotteita, lopulliseen pitoisuuteen vaikuttaa myös nattoaterian valmistuksen
jälkeinen säilytysaika. Tämän tutkimuksen näytteiden osalta jäi avoimeksi se, vastasiko niiden
säilytysaika lähellekään tavanomaista. Tutkitut näytteet oli hankittu vähittäismyyntiliikkeistä
hyvissä ajoin ennen viimeistä myyntipäivää. Nattoa pidetään hyvin säilyvänä tuotteena, ja sitä
voidaan säilyttää kenties pitkiäkin aikoja ennen nauttimista. On myös mahdollista, että
keskimääräinen säilytysaika on lyhyt, sillä tuotetta myydään Japanissa myös kadunkulmissa ja
nautitaan kenties hyvinkin pian. Surfaktiinipitoisuus olisikin hyvä analysoida sekä
keskimääräisen säilytysajan jälkeen että viimeisenä myyntipäivänä.
Karjun siittiöiden liikkuvuuden estotestin tulosten perusteella surfaktiinit osoittautuivat
sytotoksisiksi, sillä siittiöiden liike inhiboitui 2-butanoliin uutetuilla surfaktiineilla täydellisesti
0,13 – 0,24 µg/ml -surfaktiinipitoisuudella ollen yli 20 kertaluokkaa pienempi kuin
kirjallisuudessa esitetty, kuten Mikkolan ym. tutkimuksen (2000) mukainen.

62
Abdel-Mawgoudin (2008) menetelmän mukaan valmistettujen uutteiden havaittiin olevan
vähemmän siittiötoksisia, mutta pitoisuusmääritystulokset osoittivat surfaktiinia uuttuneen
näytteistä kyseisellä menetelmällä vain 0,2–0,3 % ja positiivikontrollibakteerikannasta B.
subtilis CHCC 16388 0,56 % 2-butanoliuutteisiin verrattuna. Pitoisuudeltaan 5 µg/ml olevasta
surfaktiinistandardista uuttui Abdel-Mawgoudin (2008) menetelmää käyttäen 1 % ja
pitoisuudeltaan 10 µg/ml olevasta 0,2 %. Voidaankin epäillä menetelmän toimivuutta.
Uuttomenetelmät poikkesivat toisistaan selvästi. MOLP haihtui pelletistä nopeasti, 2-butanoli
hyvin hitaasti. On epätodennäköistä, että 2-butanolista olisi jäänyt uutteisiin kuitenkaan jäämiä,
sillä se on haihtuva liuotin ja pellettien jatkokäsittely suoritettiin vasta niiden ollessa selkeästi
kuivia. On kuitenkin mahdollista, että 2-butanolin avulla valmistetuissa uutteissa oli joitain
epäpuhtauksia, sillä uutteita ei suodatettu toisin kuin MOLP-elatusliemeen valmistettuja
uutteita.
Surfaktiinia esiintyi 50 gramman nattoateriassa keskimäärin 100 mg, mikä ei ylitä LD50 -
suositusta ja kuvaa hyvin lievää akuuttia toksisuutta. Aterioiden surfaktiinipitoisuus oli
kirjallisuudessa esitettyä 17,5 mg:aa (Oka ym. 1993) selkeästi suurempi, mutta yhden 50
gramman nattoaterian nauttimisen voidaan tämänhetkisen tiedon valossa katsoa olevan
aikuiselle turvallista. Vaikka pitkäaikaisvaikutuksia ei tutkittu, mainittakoon, että yhdestä
isolaatista analysoitujen surfaktiinijakeiden A1, B, C1 ja D jakauma osoitti, että surfaktiini C:tä
esiintyi natossa eniten, yli puolet kokonaissurfaktiinipitoisuudesta. Juuri kyseisen jakeen
maksatoksisuudesta rotille on viitteitä. Kuitenkaan NOAEL-taso (alle 500 mg/kg) ei ylity, sillä
surfaktiini C:tä esiintyi noin 1 mg/g eli 50 mg per ateria, ja surfaktiinien kokonaispitoisuuskin
oli NOAEL-tasoa selkeästi alhaisempi. NOAEL-tason mukaisesti 60-kiloisen henkilön
oletetaan sietävän noin 3000 mg surfaktiinia kuukaudessa. Nattoa vuosikymmenien ajan
nauttivien henkilöiden osalta jatkuvan surfaktiinialtistuksen merkitys maksalle on kuitenkin
tuntematon. B. subtilis -bakteerikantojen aiheuttamasta maksatoksisuudesta ihmiselle
tunnetaan kaksi esimerkkiä, mikä on huomionarvoinen seikka, muttei riitä lainkaan
johtopäätösten tekoon.
Koska surfaktiinia tuottavat B. subtilis var. natto -bakteerikannat muodostavat tyypillisesti
paksun ja tiukan biofilmin sekä kykenevät persistoitumaan ihmisen suolistoon pidemmäksi
ajaksi kuin sitä tuottamattomat kannat, on tässä tutkielmassa tutkittujen bakteerikantojen
persistoituminen suolistoon mahdollista. Myös B. subtilis var. natto -bakteerikantojen
vaikutuksista eläinten suoliston mikrobiston muutoksiin ja ohutsuolen nukkalisäkkeeseen on
havaintoja.

63
Lisäksi bakteerikantojen aiemmin havaittu kyky invasoitua tehokkaasti suolen epiteelisoluihin
viittaa siihen, että tutkitut bakteerikannat mahdollisesti kykenevät invasoitumaan nattoa
nauttivan henkilön suoliston soluihin. Asian käytännön merkitys on kuitenkin tuntematon.
Tietoa ei ole myöskään niiden suolistossa mahdollisesti tuottamista lipopeptideistä.
Infektioiden matala esiintyvyys viittaa matalaan riskiin, mutta seurannan puuttuminen Aasiassa
voi vääristää mielikuvaa.
Immunosuppressiivisten ja vastustuskyvyltään muuten erityisen heikkojen henkilöiden tulisi
nattoa nauttiessaan huomioida paitsi sen sisältämä korkea mikrobipitoisuus ja pitkä
säilytysaika, myös tuottajaorganismien säilytyksen aikana mahdollisesti syntetisoimat toksiinit
ja aasialaisen perinnetavan mukaan hitaasti jäähdyttäen valmistettu, naton ohessa usein
nautittava riisi, joka saattaa sisältää huomattavia pitoisuuksia toksisia lipopeptidejä kuten
pumilasidiinia. Hyvän terveydentilan omaaville henkilöille riisi ei tuo merkitsevästi kohonnutta
infektioriskiä. Julkaistujen tutkimustulosten vaikutukset naton nauttimisen edullisista
luustovaikutuksista voivat teoriassa nostaa infektioriskiä iäkkäillä, heikkokuntoisilla naisilla,
mikäli nattoprobioottien kulutus nattoaterioiden ohella kasvaa merkittävästi.
Koska naton fermentaatiossa tuotetut yhdisteet ovat lopputulos soijapapuihin inokuloitujen
mikrobien pitoisuudesta, käytetyistä bakteerikannoista ja fermentointiolosuhteista, tämä
tutkielma antaa ainoastaan viitteitä elintarvikkeen toksisuudesta, joka vaikuttaa olevan hyvin
lievä. Tutkimuksen perusteella naton lyhytaikainen elintarvikekäyttö ei ole terveydelle
haitallista, mutta asiaa on seurattava ja lisätutkimukset ovat tarpeen. Toksisuusarvioinnin
pohjana on pääasiassa useita solutason vaikutuksia arvioivia tutkimuksia in vitro sekä koe-
eläinhavaintoja, eikä niitä voi suoraan soveltaa ihmiseen. Tässä tutkielmassa arvioitiin kolmen
näytteen surfaktiinituotantoa ja testit toistettiin kahdesti, joten tulokset antoivat vain viitteitä
lievästä toksisuudesta.
Naton nauttimisen kroonisia vaikutuksia kuvaava epidemiologinen näyttö puuttuu kokonaan.
Jotta saataisiin relevanttia tietoa natto-aterioiden elintarviketurvallisuudesta, olisi suositeltavaa
selvittää naton pitkäaikaisen nauttimisen vaikutukset sekä maksaan että suolistoon. Olisi hyvä
tutkia myös nattoaterioista eristettyjen bakteerikantojen biofilmin muodostuskykyä surfaktiinin
mahdollisten suolistovaikutusten arvioimiseksi. Lisäksi olisi hyvä selvittää nattoa nauttivan
imettävän äidin rintamaitoon mahdollisesti erittyvän surfaktiinin ja muiden syklisten
lipopeptidien keskimääräiset pitoisuudet ja niiden pitkäaikaiset vaikutukset esimerkiksi lapsen
maksaan, mikäli häntä ruokitaan natolla rintaruokinnan jälkeen.

64
8. LÄHTEET
Abdel-Mawgoud AM, Aboulwafa MM, Hassouna NA-H. Characterization of Surfactin
Produced by Bacillus subtilis Isolate BS5. Applied Biochemistry and Biotechnology Journal
2008; 150: 289-303.
Ahimou F, Jacques P, Deleu M. Surfactin and iturin A effects on Bacillus subtilis surface
hydrophobicity. Enzyme and Microbial Technology 2000; 27: 749-754.
Andersson MA, Mikkola R, Helin J, Andersson MC, Salkinoja-Salonen M. A novel sensitive
bioassay for detection of Bacillus cereus emetic toxin and related depsipeptide ionosphores.
Applied and Environmental Microbiology 1998; 64(4): 1338-1343.
Apetroaie-Constantin C, Mikkola R, Andersson MA, Teplova V, Suominen I, Johansson T,
Salkinoja-Salonen M. Bacillus subtilis and B. mojavensis strains connected to food poisoning
produce the heat stable toxin amylosin. Journal of Applied Microbiology 2009; 106: 1976-
1985.
Arima K, Kakinuma A, Tamura G. Surfactin, a crystalline peptidelipid surfactant produced
by Bacillus subtilis: isolation, characterization and its inhibition of fibrin clot formation.
Biochemical And Biophysical Research Communications 1968; 31(3): 488-494.
Ashiuchi M, Kamei T, Misono H. Poly-γ-glutamate synthetase of Bacillus subtilis. Journal of
Molecular Catalysis 2003; 23: 101-106.
Ashiuchi M, Misono H. Biochemistry and molecular genetics of poly-γ-glutamate synthesis.
Mini-Review. Applied microbiology and biotechnology 2002; 59(1): 9–14.
Barbosa TM, Serra CS, La Ragione RM, Woodward MJ, Henriques AO. Screening for Bacillus
Isolates in the Broiler Gastrointestinal Tract. Applied and Environmental Microbiology 2005;
968-978.
Beattie SH, Williams AG. Detection of toxigenic strains of Bacillus cereus and other Bacillus
spp. with an improved cytotoxicity assay. Letters in Applied Microbiology 1999; 28: 221–225.

65
Beaumont M. Flavouring composition prepared by fermentation with Bacillus spp.
International Journal of Food Microbiology 2002; 75(3): 189-196.
Bernheimer AW, Avigad L. Nature and Properties of a Cytolytic Agent Produced be Bacillus
subtilis. Journal of General Microbiology 1970; 61: 361-369.
Bhat AR, Irorere VU, Bartlett T, Hill D, Kedia G, Morris MR, Charalampopoulos D, Radecka
I. Bacillus subtilis natto: a non-toxic source of poly-γ-glutamic acid that could be used as a
cryoprotectant for probiotic bacteria. AMB Express 2013; 3(1): 36.
de Boer AS, Diderichsen B. On the safety of Bacillus subtilis and B. amyloliquefaciens: a
review. Applied Microbiology and Biotechnology 1991; 36(1): 1-4.
Branda SS, Gonzáles-Pastor JE, Ben-Yehuda S, Losick R, Kolter R. Fruiting body formation
by Bacillus subtilis. Proceedings of the National Academy of Sciences of the United States of
America 2001; 98(20): 11621-11626.
Branda SS, Chu F, Kearns DB, Losick R, Kolter R. A major protein component of the Bacillus
subtilis biofilm matrix. Molecular Microbiology 2006; 59(4): 1229-1238.
Cao X-H, Liao Z-Y, Wang C-L, Cai P, Yang WY, Lu M-F, Huang G-W. Purification and
antitumour activity of a lipopeptide biosurfactant produced by Bacillus natto TK-I.
Biotechnology an applied biochemistry 2009; 52: 97-106.
Cao X, Wang AH, Jiao RZ, Wang CL, Mao DZ, Yan L, Zeng B. Surfactin Induces Apoptosis
and G2/M Arrest in Human Breast Cancer MCF-7 Cells Through Cell Cycle Factor Regulation.
Cell Biochemistry and Biophysics 2009; 55(3): 163-171.
Chen K-I, Erh M-H, Su N-W, Liu W-H, Chou C-C, Cheng KC. Soyfoods and soybean products:
from traditional use to modern applications. Applied Microbiology and Biotechnology 2012;
96(1): 9-22.
Chu HP. The Lesithinase of Bacillus cereus and its Comparison with Clostridium welchii α-
toxin. Journal of General Microbiology 1949; 3(2): 255-273.

66
Cosentino S, Mulargia AF, Pisano P, Tuveri P, Palmas F. Incidence and biochemical
characteristics of Bacillus flora in Sardinian dairy products. International Journal of Food
Microbiology 1997; 38: 235-238.
Cutting SM. Bacillus probiotics. Review. Food Microbiology 2011; 28: 214-220.
Dehghan-Noudeh G, Housaindokht M, Bazzaz BS. Isolation, characterization, and
investigation of surface and hemolytic activities of a lipopeptide biosurfactant produced by
Bacillus subtilis ATCC 6633. Journal of Microbiology 2005; 43(3): 272-276.
Duc LH, Dong NA, Logan NA, Sutherland AD, Taylor J, Cutting SM. Cases of emesis
associated with bacterial contamination of an infant cereal product. International Journal of
Food Microbiology 2005; 102: 245-251.
Duitman EH, Hamoen LW, Rembold M, Venema G, Seitz H, Saenger W, Bernhard F,
Reinhardt R, Schmidt M, Ullrich C, Stein T, Leenders F, Vater J. The mycosubtilin synthetase
of Bacillus subtilis ATCC6633: a multifunctional hybrid between a peptide synthetase, an
amino transferase, and a fatty acid synthase. Proceedins of the National Academy of Sciences
of the United States of America 1999; 96(23): 13294-13299.
Duitman, E. Nonribosomal peptide synthesis in Bacillus subtilis. Väitöskirja. Groningenin
yliopisto 2003.
Earl AM, Losick R, Kolter R. Bacillus subtilis Genome Diversity. Journal of Bacteriology
2007; 1163-1170.
EFSA (European Food Safety Authority). Introduction of a Qualified Presumption of Safety
(QPS) approach for assessment of selected microorganisms referred to EFSA. The EFSA
Journal 2007; 587: 1-16.
EFSA (European Food Safety Authority). Scientific Opinion of the Panel on Dietetic Products,
Nutrition and Allergies. Vitamin K2 added for nutritional purposes in foods for particular
nutritional uses, food supplements and foods intended for the general population and Vitamin
K2 as a source of vitamin K added for nutritional purposes to foodstuffs, in the context of
Regulation (EC) No 258/97. The EFSA Journal 2008; 822: 1-31.

67
EFSA (European Food Safety Authority). Scientific Opinion on the maintenance of the list of
QPS biological agents intentionally added to food and feed. EFSA Journal 2013; 11(11): 3449.
EFSA (European Food Safety Authority). Technical Guidance on the assessment of the
toxigenic potential of Bacillus species used in animal nutrition. EFSA Journal 2011; 9(11):
2445.
Euroopan komissio 2013a. Komission täytäntöönpanoasetus (EU) N:o 306/2013. Annettu 2.
päivänä huhtikuuta 2013.
Euroopan komissio 2013b. Komission täytäntöönpanoasetus (EU) N:o 787/2013. Annettu 16.
päivänä elokuuta 2013.
Euroopan komissio. Report of the scientific committee on animal nutrition on the safety of the
enzymatic product Belfeed B1100 MP® for use as feed additive for pigs for fattening. Annettu
18. päivänä huhtikuuta 2002.
Elintarviketurvallisuusvirasto Evira. Utaretulehdusta aiheuttavien mikrobien eristäminen ja
tunnistaminen. Menetelmäohje Evira 2001/3. Viitattu 11.9.2014.
Elintarviketurvallisuusvirasto Evira. Elintarvikkeiden mikrobiologiset vaarat. Eviran julkaisuja
1/2010. Viitattu 9.1.2014.
FAO/WHO. Guidelines for the Evaluation of Probiotics in Food. Report of a Joint FAO/WHO
Working Group on Drafting Guidelines for the Evaluation of Probiotics in Food 2002. London,
Ontario, Canada. ftp://ftp.fao.org/es/esn/food/wgreport2.pdf . Viitattu 19.9.2014.
FDA. Food Additive Status List.
http://www.fda.gov/food/ingredientspackaginglabeling/foodadditivesingredients/ucm091048.
html. Viitattu 30.5.2014.
Fineli ® - elintarvikkeiden koostumustietopankki. Terveyden ja hyvinvoinnin laitoksen
ravitsemusyksikön ylläpitämä elintarvikkeiden koostumustietopankki. Viitattu 1.9.2014.

68
From C, Hormazábal V, Hardy SP, Granum PE. Cytotoxicity in Bacillus mojavensis is
abolished following loss of surfactin synthesis: implications for assessment of toxicity and food
poisoning potential. International Journal of Food Microbiology 2007a; 117(1): 43-49.
From C, Hormazábal V, Granum PE. Food poisoning associated with pumilacidin-producing
Bacillus pumilus in rice. International Journal of Food Microbiology 2007b; 115: 319-324.
From C, Pukall R, Schumann P, Hormazábal V, Granum PE. Toxin-Producing Ability among
Bacillus spp. Outside the Bacillus cereus Group. Applied and Environmental Microbiology
2005; 71(3): 1178-1183.
Green DH, Wakeley PR, Page A, Barnes A, Baccigalupi L, Ricca E, Cutting SM.
Characterization of Two Bacillus Probiotics. Applied and Environmental Microbiology 1999;
4288-4291.
Griffiths MW. Toxin production by psychotropic Bacillus spp. present in milk. Journal of Food
Protection 1990; 53: 790-792.
Hara T, Aumayr A, Fujio Y, Ueda S. Elimination of Plasmid-Linked Polyglutamate Production
by Bacillus subtilis (natto) with Acridine Orange. Applied and Environmental Microbiology.
1982; 44(6): 1456-1458.
Hong HA, Huang J-M, Khaneja R, Hiep LV, Urdaci MC, Cutting SM. The safety of Bacillus
subtilis and Bacillus indicus as food probiotics. Journal of Applied Microbiology 2008; 105:
510-520.
Hong HM, Khaneja R, Tam NMK, Cazzato A, Tan S, Urdaci M, Brisson A, Gasbarrini A,
Barnes I, Cutting SM. Bacillus subtilis isolated from the human gastrointestinal tract. Research
in Microbiology 2009; 160(2): 134–143.
Hoornstra D, Andersson MA, Mikkola R, Salkinoja-Salonen MS. A new method for in vitro
detection of microbially produced mitochondrial toxins. Toxicology in Vitro 2003; 745-751.

69
Hsieh F-C, Li M-C, Lin T-C, Kao S-S. Rapid Detection and Characterization of Surfactin-
Producing Bacillus subtilis and Closely Related Species Based on PCR. Current Microbiology.
2004; 49: 186-191.
Hwang Y-H, Park B-K, Lim J-H, Kim M-S, Song I-B, Park S-C, Yun H-I. Evaluation of
Genetic and Developmental Toxicity of Surfactin C from Bacillus subtilis BC1212. Journal of
Health Science 2008; 54(1): 101-106.
Hwang Y-H, Kim M-S, Song I-B, Park B-K, Lim J-H, Yun H-I. Subacute (28 day) Toxicity of
Surfactin C, a Lipopeptide Produced by Bacillus subtilis, in Rats. Journal of Health Science
2009; 55(3): 351-355.
Ikeda Y, Iki M, Morita A, Kajita E, Kagamimori S, Kagawa Y, Yoneshima H. Intake of
fermented soybeans, natto, is associated with reduced bone loss in postmenopausal women:
Japanese Population-Based Osteoporosis (JPOS) Study. The Journal of Nutrition 2006; 136(5):
1323-1328.
Inagaki S, Domon E, Saito A, Akutagawa H. Discrimination of Bacillus subtilis natto "strain
KA-145" using insertion sequence, IS4Bsu1. Journal of the Japanese Society for Food Science
and Technology. 2003; 50(10): 483-487.
Inatsu Y, Kimura K, Itoh Y. Characterization of Bacillus subtilis Strains Isolated from
Fermented Soybean Foods in Southeast Asia: Comparison with B. subtilis (natto) Starter
Strains. JARQ 2002; 36 (3): 169–175.
Jacques P. Surfactin and Other Lipopeptides from Bacillus spp. Kirjassa: Soberón‐Chávez G,
toim. Biosurfactants, Microbiology Monographs. Berlin Heidelberg: G. Springer-Verlag 2011,
s. 57–84.
Kaboré D, Nielsen DS, Sawadogo-Lingani H, Diawara B, Dicko MH, Jakobsen M, Thorsen L.
Inhibition of Bacillus cereus growth by bacteriocin producing Bacillus subtilis isolated from
fermented baobab seeds (maari) is substrate dependent. International Journal of Food
Microbiology 2013; 162(1): 114-119.

70
Kaneki M, Hodges, SJ, Hosoi T, Fujiwara S, Lyons A, Crean SJ, Ishida N, Nakagawa M,
Takechi M, Sano Y, Mizuno Y, Hoshino S, Miyao M, Inoue S, Horiki K, Shiraki M, Ouchi Y,
Orimo H. Japanese fermented soybean food as the major determinant of the large geographic
difference in circulating levels of vitamin K2: possible implications for hip-fracture risk.
Nutrition 2001; 17(4): 315-321.
Kikuchi T, Hasumi K. Enhancement of plasminogen activation by surfactin C: augmentation
of fibrinolysis in vitro and in vivo. Biochimica et Biophysica Acta (BBA) - Protein Structure
and Molecular Enzymology 2002; 1596(2): 234-245.
Kildgaard S, Mansson M, Dosen I, Klitgaard A, Frisvald JC, Larsen TO, Nielsen KF. Accurate
Dereplication of Bioactive Secondary Metabolites from Marine-Derived Fungi by UHPLC-
DAD-QTOFMS and a MS/HRMS Library. Marine Drugs 2014; 12: 3681-3705.
Kim B, Byun BY, Mah JH. Biogenic amine formation and bacterial contribution in Natto
products. Food Chemistry 2012; 135: 2005-2011.
Kim K, Jung SY, Lee DK, Jung JK, Park JK, Kim DK, Lee CH. Suppression of inflammatory
responses by surfactin, a selective inhibitor of platelet cytosolic phospholipase A2. Biochemical
Pharmacology 1998; 55(7): 975-985.
Kimura K, Itoh Y. Characterization of Poly-γ-Glutamate Hydrolase Encoded by a
Bacteriophage Genome: Possible Role in Phage Infection of Bacillus subtilis Encapsulated with
Poly-γ-Glutamate. Applied and Environmental Microbiology 2003; 69(5): 2491-2497.
Kimura K, Tran L-SP, Do T-H, Itoh Y. Expression of the pgsB Encoding the Poly-gamma-DL-
glutamate Synthetase of Bacillus subtilis (natto). Bioscience, biotechnology and biochemistry
2009; 73(5): 1149-1155.
Kramer JM, Gilbert RJ. Bacillus cereus and other Bacillus species. Kirjassa: Doyle MP, toim.
Foodborne bacterial pathogens. Marcel Dekker Inc. New York. 1989, s. 58-62.
Kubo Y, Rooney AP, Tsukakoshi Y, Nakagawa R, Hasegawa H, Kimura K. Phylogenetic
Analysis of Bacillus subtilis strains Applicable to Natto (Fermented Soybean) Production.
Applied and Environmental Microbiology 2011; 77(18): 6463–6469.

71
Lane DJ. 16S/23S rRNA sequencing. Kirjassa: Stackebrandt, E ja Goodfellow, M, toim.
Nucleic acid techniques in bacterial systematics. Chichester: Wiley 1991, s. 115-175.
Larsen N, Thorsen L, Kpikpi EN, Stuer-Lauridsen B, Cantor MD, Nielsen B, Brockmann E,
Derkx PMF, Jespersen L. Characterization of Bacillus spp. strains for use as probiotic additives
in pig feed. Applied Microbiology and Biotechnology 2014; 98: 1105–1118.
Leonard CG, Housewright RD, Thorne CB. Effects of some metallic ions on glutamyl
polypeptide synthesis by Bacillus subtilis. Journal of Bacteriology 1958; 76(5): 499-503.
Liu X, Ren B, Gao H, Liu M, Dai H, Song F, Yu Z, Wang S, Hu J, Kokare CR, Zhang L.
Optimization for the Production of Surfactin with a New Synergistic Antifungal Activity. PloS
one 2012; 7(5)
Logan NA. Bacillus and relatives in foodborne illness. Journal of Applied Microbiology 2011;
112: 417-429.
Logan NA, de Vos P. Genus I. Bacillus Cohn 1872, 174AL. Kirjassa: de Vos P, Garrity GM,
Jones D, Krieg NR, Ludwig W, Rainey FA, Schleifer K-H, Whitman WB, toim. Bergey’s
manual of systematic bacteriology, toinen painos, kolmas osa. 2009, s. 21-128. Springer, New
York.
Maget-Dana R, Peypoux F. Iturins, a special class of pore-forming lipopeptides: biological and
physicochemical properties. Toxicology 1994; 87(1-3): 151-174.
Marahiel MA, Nakano MM, Zuber P. Regulation of peptide antibiotic production in Bacillus.
Molecular Microbiology 1993; 7(5): 631-636.
Matarante A, Baruzzi F, Cocconcelli PS, Morea M. Genotyping and Toxigenic Potential of
Bacillus subtilis and Bacillus pumilus Strains Occurring in Industrial and Artisanal Cured
Sausages. Applied and Environmental Microbiology 2004; 5168-5176.
May JJ, Kessler N, Marahiel MA, Stubbs MT. Crystal structure of DhbE, an archetype for aryl
acid activating domains of modular nonribosomal peptide synthetases. Proceedings of the
National Academy of Sciences of the United States of America 2002; 99(19): 12120-12125.

72
McKenney PT, Adams D, Eichenberger P. The Bacillus subtilis endospore: assembly and
functions of the multilayered coat. Nature Reviews Microbiology 2013; 11: 33-44.
McLoon AL, Guttenplan SB, Kearns DB, Kolter R, Losick R. Tracing the domestication of a
biofilm-forming bacterium. Journal of Bacteriology 2011; 193(8): 2027–2034.
Mikkola R, Kolari M, Andersson MA, Helin J, Salkinoja-Salonen MS. Toxic lactonic
lipopeptide from food poisoning isolates of Bacillus licheniformis. European Journal of
Biochemistry 2000; 267: 4068-4074.
Mitsui N, Murasawa H, Sekiguchi J. Disruption of the cell wall lytic enzyme CwlO affects the
amount and molecular size of poly-γ-glutamic acid produced by Bacillus subtilis (natto). The
journal of general and applied microbiology 2011; 57(1): 35-43.
Mongkolthanaruk W. Classification of Bacillus beneficial substances related to plants, humans
and animals. Review. Journal of Microbiology and Biotechnology. 2012; 22(12): 1597-1604.
Murooka Y, Yamshita M. Traditional healthful fermented products of Japan. Review. Journal
of Industrial Microbiology & Biotechnology 2008; 35: 791–798.
Nagai T, Tran L-SP, Inatsu Y, Itoh Y. A new IS4 Family Insertion Sequence, IS4Bsu1,
Responsible for Genetic Instability of Poly-γ-Glutamic Acid Production in Bacillus subtilis.
Journal of Bacteriology 2000; 182(9): 2387-2392.
Nagai T. Bacteriophages of Bacillus subtilis (natto) and Their Contamination in Natto
Factories. Kirjassa: Kurtboke I, toim. Bacteriophages. InTech 2012, s. 95-110.
Nagai T, Itoh Y. Characterization of a Generalized Transducing Phage of Poly-(gamma)-
Glutamic Acid-Producing Bacillus subtilis and Its Application for Analysis of Tn917-LTV1
Insertional Mutants Defective in Poly-(gamma)-Glutamic Acid Production. Applied and
environmental microbiology 1997; 63(10): 4087-4089.

73
Nakamura LK, Roberts MS, Cohan FM. Relationship of Bacillus subtilis clades associated with
strains 168 and W23: a proposal for Bacillus subtilis subsp. subtilis subsp. nov, and Bacillus
subtilis subsp. spizizenii subsp. nov. International Journal of Systematic Bacteriology 1999; 49:
1211-1215.
Nakano MM, Dailly YP, Zuber P, Clark DP. Characterization of Anaerobic Fermentative
Growth of Bacillus subtilis: Identification of Fermentation End Products and Genes Required
for Growth. Journal of Bacteriology 1997; 179(21): 6749.
Nakano MM, Magnuson R, Myers A, Curry J, Grossman AD, Zuber P. srfA Is an Operon
Required for Surfactin Production, Competence Development, and Efficient Sporulation in
Bacillus subtilis. Journal of Bacteriology 1991; 173(5): 1770-1778.
Nakano MM, Zuber P. Anaerobic growth of a "strict aerobe" (Bacillus subtilis). Annual review
of microbiology 1998; 52: 165-190.
Natural Domain Product Seeker NaPDoS. Jensenlab. USA 2011.
http://napdos.ucsd.edu/pathway_templates/fengycin_info.html
Nguyen Thi Minh H, Durand A, Loison P, Perrier-Cornet J-M, Gervais P. Effect of sporulation
on the resistance of Bacillus subtilis spores to heat and high pressure. Applied Microbiology
and Biotechnology 2011; 90: 1409-1417.
Nieminen T, Rintaluoma N, Andersson M, Taimisto A-M, Ali-Vehmas T, Seppälä A, Priha O,
Salkinoja-Salonen M. Toxinogenic Bacillus pumilus and Bacillus licheniformis from mastitic
milk. Veterinary Microbiology 2007; 124: 329-339.
Nishiguchi T, Yamashita M, Maeda M, Matsuyama K, Kanavama N, Terao T. Improvement of
vitamin K status of breastfeeding infants with maternal supplement of vitamin K2 (MK40).
Seminars in Thrombosis and hemostasis 2002; 28(6): 533-538.
Nishito Y, Osana Y, Hachiya T, Popendorf K, Toyoda A, Fujiyama A, Itaya M, Sakakibara Y.
Whole genome assembly of a natto producing strain Bacillus subtilis natto from very short read
data. BMC Genomics 2010; 11: 243.

74
Nissen E, Vater J, Pauli G, Vollenbroich D. Application of surfactin for mycoplasma
inactivation in virus stocks. In vitro cellular & developmental biology. Animal. 1997; 33: 414-
415.
Oggioni MR, Pozzi G, Valensin PE, Galieni P, Bigazzi C. Recurrent septicemia in an
immunocompromised patient due to probiotic strains of Bacillus subtilis. Journal of Clinical
Microbiology 1998; 36(1): 325-326.
Oka K, Hirano T, Homma M, Ishii H, Murakami K, Mogami S, Motizuki A, Morita H, Takeya
K, Itokawa H. Satisfactory separation and MS-MS spectrometry of six surfactins isolated from
Bacillus subtilis natto. Chemical & pharmaceutical bulletin 1993; 41(5): 1000–1002.
Ongena M, Jacques P. Bacillus lipopeptides: versatile weapons for plant disease biocontrol.
Trends of Microbiology 2008; 16: 115-125.
Pan X, Chen X, Su X, Feng Y, Tao Y, Dong Z. Involvement of SpoVG in hemolysis by Bacillus
subtilis. Biochemical and Biophysical Research Communications 2014; 443: 899-904.
Park, SY. & Kim, YH. Surfactin inhibits immunostimulatory function of macrophages through
blocking NK-κB, MAPK and Akt pathway. International Immunopharmacology 2009; 9: 886-
893.
Pedersen PB, Bjørnvad ME, Rasmussen MD, Petersen JN. Cytotoxic potential of industrial
strains of Bacillus sp. Regulatory Toxicology and Pharmacology: RTP. 2002; 36(2):155-161.
Peng H, Wang JQ, Kang HY, Dong SH, Sun P, Bu DP Zhou LY. Effect of feeding Bacillus
subtilis natto fermentation product on milk production and composition, blood metabolites and
rumen fermentation in early lactation dairy cows. Journal of Animal Physiology and Animal
Nutrition 2012; 96(3): 506-512.
Peypoux F, Pommier MT, Marion D, Ptak M, Das BC, Michel G. Revised structure of
mycosubtilin, a peptidolipid antibiotic from Bacillus subtilis. The Journal of antibiotics 1986;
39(5): 636-641.

75
Peypoux F, Bonmatin J-M, Labbe H, Grangemard I, Das BC, Ptak M, Wallach J, Michel G.
[Ala4]Surfactin, a Novel Isoform from Bacillus subtilis Studied by Mass and NMR
Spectroscopies. European Journal of Biochemistry 1994; 224(1): 89-96.
Peypoux F, Bonmatin J-M, Wallach J. Recent trends in the biochemistry of surfactin. Applied
and Environmental Microbiology 1999; 51: 553-563.
Phelps RJ, McKillip JL. Enterotoxin Production in Natural Isolates of Bacillaceae outside the
Bacillus cereus Group. Applied and Environmental Microbiology 2002; 68(6): 3147-3151.
van der Ploeg R, Mäder U, Homuth G, Schaffer M, Denham EL, Monteferrante CG, Miethke
M, Marahiel MA, Harwood CR, Winter T, Hecker M, Antelmann H, van Dijl JM.
Environmental Salinity Determines the Specificity and Need for Tat-Dependent Secretion of
the YwbN Protein in Bacillus subtilis. PLoS one 2011; 6(3): 18140.
Pyörälä S. Harvinaiset utaretulehduksen aiheuttajat. Eläintautien torjuntayhdistys ETT.
Helsingin yliopisto 2012. Viitattu 11.9.2014.
Pötter M, Oppermann-Sanio FB, Steinbüchel A. Cultivation of Bacteria Producing Polyamino
Acids with Liquid Manure as Carbon and Nitrogen Source. Applied and Environmental
Microbiology 2001: 617-622.
Qiu D, Fujita K, Sakuma Y, Tanaka T, Ohashi Y, Ohshima H, Tomita M, Itaya M. Comparative
Analysis of Physical Maps of Four Bacillus subtilis (natto) Genomes. Applied and
Environmental Microbiology 2004; 6247-6256.
Rajavel M, Mitra A, Gopal B. Role of Bacillus subtilis BacB in the synthesis of bacilysin. The
journal of biological chemistry 2009; 284(46): 31882-31892.
Richard V, van der Auwera P, Snoeck R., Daneau D, Meunier, F. Nosocomial bacteremia
caused by Bacillus species. European journal of clinical microbiology & infectious diseases
1988; 7(6): 783-785.

76
Roberts MS, Nakamura LK, Cohan FM. Bacillus mojavensis sp. nov., Distinguishable from
Bacillus subtilis by Sexual Isolation, Divergence in DNA Sequence, and Differences in Fatty
Acid Composition. International Journal of Systematic Bacteriology 1994; 44(2): 256-264.
Roberts MS, Nakamura LK, Cohan FM. Bacillus vallismortis sp. nov., a Close Relative of
Bacillus subtilis, Isolated from Soil in Death Valley, California. International Journal of
Systematic Bacteriology 1996; 46(2): 470-475.
Rooney A, Price N, Ehrherdt C, Swezey JL, Bannan JD. Phylogeny and molecular taxonomy
of the Bacillus subtilis species complex and description of Bacillus subtilis subsp. inaquosorum
subsp. nov. International Journal of Systematic and Evolutionary Microbiology 2009; 59(10):
2429-2436.
Rosenkvist H, Hansen Å. Contamination profiles and characterization of Bacillus species in
wheat bread and raw materials for bread production. International Journal of Food
Microbiology 1995; 26: 353-363.
Rowan NJ, Deans K, Anderson JG, Gemmell CG, Hunter IS, Chaithong T. Putative Virulence
factor Expression by Clinical and Food Isolates of Bacillus spp. after Growth in Reconstituted
Infant Milk Formulae. Applied and Environmental Microbiology 2001; 3873-3881.
Salkinoja-Salonen MS, Vuorio R, Andersson MA, Kämpfer P, Andersson MC, Honkanen-
Buzalski T, Scoging AC. Toxigenic Strains of Bacillus Licheniformis Related to Food
Poisoning. Applied and Environmental Microbiology 1999; 65(10): 4637-4645.
Samanya M, Yamauchi KE. Histological alterations of intestinal villi in chickens fed dried
Bacillus subtilis var. natto. Comparative Biochemistry and Physiology Part A: Molecular &
Integrative Physiology 2002; 133(1): 95-104.
Sanders ME, Morelli L, Tompkins TA. Sporeformers as Human Probiotics: Bacillus,
Sporolactobacillus and Brevibacillus. Comprehensive Reviews in Food Science and Food
Safety 2003; 2: 101-110.
Sawamura S. 1906. On the micro-organisms of natto. Bulletin of the college of agriculture,
Tokyo Imperial University 1906; 7(1): 107-110.

77
Schallmey M, Singh A, Ward OP. Developments in the use of Bacillus species for industrial
production. Canadian journal of microbiology 2004; 50(1): 1-17.
Shrestha AK, Dahal NR, Ndungutse V. Bacillus Fermentation of Soybean: A Review. Journal
of Food Science and Technology 2010; 6: 1-9.
Song DJ, Kang HY, Peng H, Bu DP. Effect of Feeding Bacillus subtilis natto on Hindgut
Fermentation and Microbiota of Holstein Dairy Cows. Asian-Australasian Journal of Animal
Sciences 2014; 27(4): 495-502.
Spinosa MR, Wallet F, Courcol RJ, Oggioni MR. The trouble in tracing opportunistic
pathogens: cholangitis due to Bacillus in a French hospital caused by a strain related to an Italian
probiotic? Microbial Ecology in Health and Disease 2000; 12: 99-101.
Stickel F, Droz S, Patsenker E, Bögli-Stuber K, Aebi B, Leib SL. Severe hepatotoxicity
following ingestion of Herbalife® nutritional supplements contaminated with Bacillus subtilis.
Journal of Hepatology 2009; 50(1): 111-117.
Strauch M, Webb V, Spiegelman G, Hoch JA. The SpoOA protein of Bacillus subtilis is a
repressor of the abrB gene. Genetics 1990; 87: 1801-1805.
Sumi H, Yanagisawa Y, Yatagai C, Saito J. Natto Bacillus as an Oral Fibrinolytic Agent:
Nattokinase Activity and the Ingestion Effect of Bacillus subtilis natto. Food Science and
Technology 2004; 10(1): 17-20.
Sun P, Wang J-Q, Zhang H-T. Effects of supplementation of Bacillus subtilis natto Na and N1
strains on rumen development in dairy calves. Animal Feed Science and Technology 2011; 164
(3-4): 154-160.
Sun P, Wang JQ, Deng LF. Effects of Bacillus subtilis natto on milk production, rumen
fermentation and ruminal microbiome of dairy cows. Cambridge Journals 2013; 7(2): 216-222.
Suzuki Y, Kondo K, Ichise H, Tsukamoto Y, Urano T, Umemura K. Dietary Supplementation
With Fermented Soybeans Suppresses Intimal Thickening. Nutrition 2003; 19: 261-264.

78
Tam NKM, Uyen NQ, Hong HA, Duc LH, Hoa TT, Serra CR, Henriques AO, Cutting SM. The
Intestinal Life Cycle of Bacillus subtilis and Close Relatives. Journal of Bacteriology 2006;
188(7): 2692-2700.
Tapi A, Chollet-Imbert M., Scherens B, Jacques P. New approach for the detection of non-
ribosomal peptide synthetase genes in Bacillus strains by polymerase chain reaction. Applied
Microbiology and Biotechnology Journal 2010; 85: 1521-1531.
Tsuge K, Ano T, Hirai M, Nakamura Y, Shoda M. The Genes degQ, pps, and lpa-8(sfp) Are
Responsible for Conversion of Bacillus subtilis 168 to Plipastatin Production. Antimicrobioal
Agents and Chemotherapy 1999; 43(9): 2183-2192.
USDA National Nutrient Database for Standard Reference Release 26. Statistics Report 16113,
Natto.
http://ndb.nal.usda.gov/ndb/foods/show/4833?format=Stats&reportfmt=pdf&pdfQvs=%7B%
7D. Luettu 30.5.2014.
Vanittanakom N, Loeffler W, Koch U, Jung G. Fengycin - a novel antifungal lipopeptide
antibiotic produced by Bacillus subtilis f-29-3. The Journal of Antibiotics. 1986; 39(7): 888-
901.
Vollenbroich D, Pauli G, Özel M, Vater J. Antimycoplasma Properties and Application in Cell
Culture of Surfactin, a Lipopeptide Antibiotic from Bacillus subtilis. Applied and
Environmental Microbiology 1997a; 63(1): 44-49.
Vollenbroich D, Özel M, Vater J, Kamp RM, Pauli G. Mechanism of Inactivation of Enveloped
Viruses by the Biosurfactant Surfactin from Bacillus subtilis. Biologicals 1997b; 25: 289-297.
Volpon L, Besson F, Lancelin J-M. NMR structure of antibiotics plipastatins A and B from
Bacillus subtilis inhibitors of phospholipase A2. FEBS Letters 2000; 485(1): 76-80.
Wallet F, Crunelle V, Roussel-Delvallez M, Fruchart A, Saunier P, Courcol RJ. Bacillus subtilis
as a cause of cholangitis in polycystic kidney and liver disease. American Journal of
Gastroenterology 1996; 91(7): 1477-1478.

79
Wang LT, Lee FL, Tai CJ, Kasai H. Comparison of gyrB gene sequences, 16S rRNA gene
sequences and DNA-DNA hybridization in the Bacillus subtilis group. International Journal of
Systematic and Evolutionary Microbiology 2007; 57: 1846-1850.
Wei Q, Wolf-Hall C, Chang KC. Natto Characteristics as Affected by Steaming Time, Bacillus
Strain, and Fermentation Time. Journal of Food Science 2001; 66(1): 167-172.
Williams GR. Haemolytic Material from Aerobic Sporing Bacilli. Journal of general
Microbiology 1957; 16: 16-21.
Yi H, Chun J, Cha C-J. Genomic insights into the taxonomic status of the three subspecies of
Bacillus subtilis. Systematic and Applied Microbiology 2013; 13: 160-164.
Yin H, Guo C, Wang Y, Liu D, Lv Y, Lv F, Lu Z. Fengycin inhibits the growth of the human
lung cancer cell line 95D through reactive oxygen species production and mitochondria-
dependent apoptosis. Anti-cancer drugs 2013; 24(6): 587-598.

80
9. LIITTEET
LIITE 1
Liite 1. Lipopeptidien uutto 2-butanoliin.
10 ml
suspensiota
12 ml 2-
butanolia
150 rpm 15 min.
Mikrobipesäkkeen
nestekasvatus 25 ml:ssa BHI-
lientä 24 h 30 oC 150 rpm
Sentrifugointi
4000 rpm 10 min.
10 ml
supernatanttia
12 ml 2-
butanolia
150 rpm 15 min.
Sentrifugointi
4000 rpm 10 min.
10 ml
supernatanttia
10 ml
supernatanttia
Supernatanttien
yhdistäminen
2-butanolin
haihduttami
nen N2-
virrassa
Pelletin
eluointi 1
ml:aan 5
mM Tris-
HCl (pH
8,5)

81
LIITE 2
Liite 2. Medium of optimal lipopeptide production (MOLP) –elatusliemen koostumus Ahimoun
ym. (2000) mukaan. Elatusliemen pH säädettiin 5 M natriumhydroksidilla arvoon 7 ja
autoklavoitiin.
Ainesosa Pitoisuus
Peptoni 30 g/l
Sakkaroosi 20 g/l
Hiivauute 7 g/l
Kaliumdivetyfosfaatti (KH2PO4) 1,9 g/l
Kuparisulfaatti (CuSO4) 0,001 mg/l
Kidevedellinen Ferrikloridi (FeCl3·6H2O) 0,005 mg/l
Natriumolybdaatti (Na2MoO4) 0,0047 mg/l
Kaliumjodidi (KI) 0,002 mg/l
Kidevedellinen magnaanisulfaatti (MnSO4·H2O) 3,6 mg/l
Kidevedellinen magnesiumsulfaatti (MgSO4·7H2O) 0,921 mg/l
Kidevedellinen sinkkisulfaatti (ZnSO4·7H2O) 0,14 mg/l
Vetyboraatti (H3BO3) 0,01 mg/l
Kidevedellinen sitraatti (C6H8O7·H2O) 10,9 mg/l

82
LIITE 3
Liite 3. Lipopeptidien synteesikykyä määrittävien geenien tutkimiseksi valmistettu PCR-
reaktioseos. Kokonaistilavuus 50 µl / näyte.
Ainesosa Pitoisuus (surfaktiinit ja
fengysiinit)
Pitoisuus (plipastatiinit ja
mykosubtiliini)
Nukleaasivapaa vesi
5 × SYBR Green -puskuriliuos 1 × 1 ×
MgCl2+ -ionit 1,5 mM 1,5 mM
Aluke 1 12 ng / µl 12 ng / µl
Aluke 2 12 ng / µl 12 ng / µl
Nukleotidit (dNTP) 0,2 mM 0,4 mM
Go Taq Flexi -polymeraasi 1,25 U 2,5 U
Kopioitava DNA ≥ 10 ng / µl ≥ 10 ng / µl
LIITE 4
Liite 4. Tutkittuja lipopeptidejä koodaavien geenien määrittämisessä käytetyt PCR-ajo-
ohjelmat. Kaikille näytteille käytettiin kolmen minuutin 94 oC alkudenaturaatiota, 30 sykliä, ja
ajot päätettiin 4 oC:een.
Lipopeptidit Denaturaatio Alukkeiden
kiinnittyminen Ekstensio Loppuekstensio
Syklejä
kpl
Surfaktiinit 94 oC / 1 min 43 oC / 30 s 72 oC / 45 s 72 oC / 10 min 30
Fengysiinit 94 oC/ 1 min 45 oC / 30 s 72 oC / 45 s 72 oC / 10 min 25
Mykosubtiliini 94 oC/ 1 min 45 oC / 30 s 72 oC / 45 s 72 oC / 10 min 25
Plipastatiinit 94 oC/ 1 min 58 oC / 30 s 72 oC / 75 s 72 oC / 10 min 30