bedeutung von peroxisomen im metabolismus von ... · tierärztliche hochschule hannover zur...
TRANSCRIPT




Bibliografische Informationen der Deutschen Bibliothek
Die Deutsche Bibliothek verzeichnet diese Publikation in der Deutschen
Nationalbibliografie;
Detaillierte bibliografische Daten sind im Internet über http://dnb.ddb.de abrufbar.
1. Auflage 2012
© 2012 by Verlag: Deutsche Veterinärmedizinische Gesellschaft Service GmbH,
Gießen
Printed in Germany
ISBN 978-3-86345-0
Verlag: DVG Service GmbH
Friedrichstraße 17
35392 Gießen
0641/24466
www.dvg.net
75-5

Tierärztliche Hochschule Hannover
Zur Bedeutung von Peroxisomen im Metabolismus von
degenerativen und neoplastischen Lebererkrankungen beim Hund
INAUGURAL – DISSERTATION
zur Erlangung des Grades einer Doktorin der Veterinärmedizin
- Doctor medicinae veterinariae -
(Dr. med. vet.)
vorgelegt von
Julia Schüttler
geb. Borchert
Bad Kreuznach
Hannover 2012

Wissenschaftliche Betreuung: Univ. Prof. Dr. F.-J. Kaup
Deutsches Primatenzentrum, Göttingen
Abteilung Infektionspathologie
1. Gutachter: Univ. Prof. Dr. F.-J. Kaup
Tierärztliche Hochschule Hannover
Deutsches Primatenzentrum, Göttingen
2. Gutachterin: Prof. Dr. Marion Hewicker-Trautwein
Tierärztliche Hochschule Hannover
Institut für Pathologie
Tag der mündlichen Prüfung: 27.04.2012

Meinem Vater, meiner Mutter
und Raimund


Inhaltsverzeichnis
Inhaltsverzeichnis
1 Einleitung ...................................................................................................... 11
2 Literaturübersicht .......................................................................................... 14
2.1 Das Peroxisom.............................................................................................. 14
2.1.1 Morphologie .................................................................................................. 15
2.1.2 Funktion ........................................................................................................ 18
2.1.3 Interaktion mit Mitochondrien ........................................................................ 29
2.2 Kontrolle der Peroxisomenzahl ..................................................................... 30
2.2.1 Biogenese und Proliferation .......................................................................... 31
2.2.2 Abnahme der Peroxisomenzahl durch Autophagie ...................................... 37
2.3 Hepatopathien des Hundes .......................................................................... 39
2.3.1 Hepatozelluläre Degeneration ...................................................................... 42
2.3.2 Verhalten der Peroxisomen bei degenerativen Veränderungen der
Hepatozyten .................................................................................................. 48
2.3.3 Hepatitiden .................................................................................................... 49
2.3.4 Lebertumoren ............................................................................................... 51
3 Eigene Untersuchungen ............................................................................... 57
3.1 Material und Methoden ................................................................................. 57
3.1.1 Einleitung ...................................................................................................... 57
3.1.2 Patienten ....................................................................................................... 57
3.1.3 Lichtmikroskopische Untersuchung .............................................................. 63
3.1.4 Präparation für die Transmissionselektronenmikroskopie ............................. 63
3.1.5 Auswertung und Dokumentation ................................................................... 65
3.1.6 Statistik ......................................................................................................... 68
3.2 Ergebnisse .................................................................................................... 68
3.2.1 Zusammensetzung der Patientengruppen .................................................... 69
3.2.2 Veränderungen der labordiagnostischen Leberwerte ................................... 72
3.2.3 Lichtmikroskopische Untersuchungsergebnisse ........................................... 76

Inhaltsverzeichnis
3.2.4 Elektronenmikroskopische Untersuchungsergebnisse .................................. 95
4 Diskussion .................................................................................................. 123
4.1 Lichtmikroskopische Untersuchungen ........................................................ 123
4.2 Elektronenmikroskopische Untersuchung ................................................... 127
4.2.1 Quantitative Erfassung der Peroxisomen/Hepatozyt- Fläche ...................... 128
4.2.2 Quantitative Erfassung der Peroxisomen/Hepatozyt- Fläche/Alter ............. 133
4.2.3 Quantitative Erfassung der Peroxisomen/Hepatozyt- Fläche/Rasse........... 133
4.2.4 Morphologische Untersuchungen ............................................................... 134
4.3 Beeinflussung der Peroxisomenzahl durch Medikamente .......................... 140
5 Zusammenfassung ..................................................................................... 142
6 Summary .................................................................................................... 145
7 Literaturverzeichnis ..................................................................................... 148
8 Abbildungsverzeichnis ................................................................................ 166
9 Tabellenverzeichnis .................................................................................... 172
10 Anhang ....................................................................................................... 174
10.1 Protokolle für die Histologie ........................................................................ 174
10.1.1 Fixierlösungen............................................................................................. 174
10.1.2 Färbungen .................................................................................................. 176
10.1.3 Laborprotokolle für Einbettungen ................................................................ 181
10.2 Anhangstabellen ......................................................................................... 182

Abkürzungsverzeichnis
Abkürzungsverzeichnis
ALT Alaninaminotransferase
ALKP Alkalische Phosphatase
AST Aspartataminotransferase
ATG autophagy-related genes
ß-Oxidation Beta-Oxidation
bzw. beziehungsweise
ca. circa
CuZn SOD Kupfer-Zink Superoxiddismutase
d Tag
DNA Desoxyribonucleic acid
Elmi Elektronenmikroskopie
ggr. geringgradig
GLDH Glutamatdehydrogenase
GS Gallensäuren
HWZ Halbwertzeit
h Stunde
H2O2 Wasserstoffperoxid
hgr. hochgradig
IBD Inflammatory bowel disease
IL Interleukin
IFN γ Interferon gamma

Abkürzungsverzeichnis
JRT Jack Russel Terrier
KGW Körpergewicht
m/mk männlich/männlich kastriert
MDV Mitochondria derived vesicles
MAPL Mitochondria-anchored protein ligase
mgr. mittelgradig
M. Morbus
NASH non alcoholic steatohepatitis
nm Nanometer
NO Stickstoff
PD/PU Polydipsie/Polyurie
PEX Peroxin
PTS Peroxisomal targeting signal
PPO Peroxisomenproliferatoren
PPARs Peroxisome proliferator activated receptors
Px Peroxisom
rER raues (rough) Endoplasmatisches Retikulum
RHT Rauhaardackel
ROS Reactive oxygen species
sER glattes (smooth) Endoplasmatisches Retikulum
s Sekunde
SOD Superoxiddismutase
TGF β Transforming growth factor

Abkürzungsverzeichnis
TNFα Tumor necrosis factor alpha
vak. vakuolär
VLDL Very long density lipoprotein
WHWT Westhighland White Terrier
w/wk weiblich/weiblich kastriert
µm² Quadratmikrometer


1 Einleitung 11
1 Einleitung
Peroxisomen haben eine außerordentliche Bedeutung erlangt, da sie Gegenstand
zahlreicher wissenschaftlicher Untersuchungen in der Humanmedizin sind. Die
Untersuchungsergebnisse zeigen, dass die Peroxisomen an zahlreichen humanen
kongenitalen Erkrankungen beteiligt sind. „Lorenzo´s oil“ ist der Titel eines Filmes,
der sehr eindrücklich das Leben eines Jungen erzählt, der an einer unerklärlichen
Erkrankung leidet. Gleichgewichtsstörungen und letztlich schwere Hirnschädigungen
sind Folgen dieser Erkrankung, bei der es sich um die „Adrenoleukodystrophie“
handelt. Intensive wissenschaftliche Untersuchungen der letzten Jahrzehnte führten
zu der Erkenntnis, dass kleine Zellorganellen, Peroxisomen, die Ursache dieser
Erkrankung sind. Das Zellweger Syndrom und die „Refsum disease“ sind weitere
kongenitale peroxisomale Erkrankungen. Durch die Entdeckung der Peroxisomen
1954 durch RHODIN und die weiteren wissenschaftlichen Untersuchungen des
belgischen Wissenschaftlers DE DUVE (1966) erlangte diese kleine Organelle eine
immer größere Bedeutung. Ihre Beteiligung innerhalb komplexer physiologischer
Vorgänge, wie Einbindung in den Wasserstoffperoxidstoffwechsel, die ß-Oxidation
langer bis sehr langer Fettsäuren, die Synthese komplexer Substrate wie
Cholesterine, Plasmalogene und Gallensäuren, sowie die Fähigkeit sich in
Anwesenheit sogenannter Peroxisomenproliferatoren zu vermehren, machten sie zu
einem spannenden Forschungsgebiet, vor allem in der Humanmedizin. Während sich
die ersten Arbeiten mit den kongenitalen Erkrankungen der Peroxisomen befassten,
folgten durch DE CRAEMER (1995) wissenschaftliche Untersuchungen zum
Verhalten der Peroxisomen bei erworbenen Erkrankungen, wie Steatose, Hepatitis
und Zirrhose, hervorgerufen durch verschiedene Noxen wie Alkohol, Viren und
diverse Medikamente. Weitere Untersuchungen folgten zum Verhalten der
Peroxisomen bei cholestatischen Prozessen der Leber, Hepatomen und
extrahepatischen Tumoren mit oder ohne Lebermetastasen.
Die wissenschaftlichen Erkenntnisse über Peroxisomen in der Veterinärmedizin sind
bis dato sehr gering. LAGING et al. (1988) untersuchten vergleichend das Verhalten

12 1 Einleitung
der Peroxisomen in Leber und Niere von Hunden der Rasse Beagle und Dalmatiner
bei purinreicher und purinarmer Ernährung. Sie kamen zu der Erkenntnis, dass
Dalmatiner unabhängig von der Ernährung größere und zahlreichere Peroxisomen
haben als Beagle. Eine weitere Untersuchung von CENTER und Mitarbeitern (1993)
befasste sich mit dem Verhalten der hepatischen Peroxisomen in an Lipidose
erkrankten Katzen. CENTER et al. (1993) vermuteten dabei, dass eine Abnahme der
Peroxisomen mit dem Krankheitsbild der hepatischen Lipidose einhergeht. Die ersten
elektronenmikroskopischen Daten zur Morphometrie der hepatozellulären
Peroxisomen des Hundes wurden 1973 erhoben. Dies erschien notwendig, da der
Hund immer häufiger für pharmakologische und toxikologische Studien als Modell für
humane Lebererkrankungen herangezogen wurde (HESS et al. 1973).
Elektronenmikroskopische und biochemische Untersuchungen aus der
Humanmedizin zum Verhalten von Peroxisomen bei tumorösen und degenerativen
Erkrankungen der Leber (HRUBAN 1979; DE CRAEMER et al. 1993) ergaben, dass
bei degenerativen Erkrankungen, z.B. aufgrund von hepatischer Lipidose/Steatose
und entzündlichen Prozessen, im Rahmen eines erhöhten oxidativen Stresses für die
Hepatozyten, die Zahl der Peroxisomen ansteigt. Tumoröse Veränderungen der
Leber, seien sie nun primären oder sekundären Ursprunges, führten dagegen in der
Mehrzahl der Fälle zu einer Verminderung der Peroxisomen. Weiterhin haben
Untersuchungen gezeigt, dass unter dem Einfluss des Tumornekrosefaktors die
Bildung von Radikalen erheblich ansteigt. In einem solchen Fall ist die
Kapazitätsgrenze der Peroxisomen erreicht, und es kommt zu einem Abbau dieser
Organelle infolge zunehmender Zellzerstörung (YASMINEH et al. 1991). Es kann
deshalb vermutet werden, dass die Anzahl der Peroxisomen in einem Verhältnis zum
Grad der Malignität zu sehen ist. Weiterhin kann vermutet werden, dass
therapeutische Maßnahmen, welche zu einer Peroxisomenvervielfältigung führen
über eine Beeinflussung des oxidativen Stresses und Veränderungen bei der
Energiegewinnung einen Einfluss auf die Leberzelle haben könnten. Angesichts
dieser Informationen aus der Humanmedizin ist es Ziel dieser Arbeit, das Verhalten
der hepatozellulären Peroxisomen im Metabolismus von Lebererkrankungen des
Hundes elektronenmikroskopisch zu untersuchen. Dabei wurde insbesondere auf

1 Einleitung 13
quantitative und qualitative Unterschiede zwischen degenerativen und entzündlichen
Veränderungen einerseits und malignen Tumoren der Leber andererseits
eingegangen, um zu überprüfen, ob die von CRAEMER et al. (1993) bzw. HRUBAN
(1979) gewonnenen Erkenntnisse auch auf den Hund übertragbar sind.

14 2 Literaturübersicht
2 Literaturübersicht
2.1 Das Peroxisom
Peroxisomen kommen sowohl im Pflanzen- als auch im Tierreich vor. Es wurde von
ihrem Vorkommen in Amphibien, Insekten, Schnecken, Würmern und Protozoen
berichtet (MASTERS u. CRANE 1995).
Die Peroxisomen wurden erstmals 1954 durch RHODIN in Nierentubuluszellen von
Ratten entdeckt, er gab diesen Zellorganellen zunächst den Namen „Microbodies“
(RHODIN 1954). Im Jahre 1956 wurden Zellorganellen, die den bereits
beschriebenen „Microbodies“ der Niere sehr ähnlich waren, in Hepatozyten von
Rattenlebern gefunden (Abbildung 2-1) (LAZAROW u. FUJIKI 1985).
Weitere Aufklärung haben in den 60iger Jahren die biomedizinischen Experimente
von DE DUVE und BAUDHUIN (1966) erbracht, in denen Peroxisomen von
Rattenlebern isoliert und deren enzymatische Ausstattung näher differenziert wurde.
Diese biochemischen Erkenntnisse führten dazu, dass der morphologisch geprägte
Name „Microbody“ der funktionellen Bezeichnung „Peroxisom“ wich.
Die Isolierung der Peroxisomen und die anschließende zentrifugale Fraktionierung
ihrer einzelnen Komponenten, wie das peroxisomale Core oder Nukleoid, die
peroxisomale Matrix und die peroxisomale Membran, ermöglichten eine detaillierte
Aufschlüsselung der enzymatischen Ausstattung der Peroxisomen. Darüber hinaus
diente die Elektronenmikroskopie, insbesondere die zytochemische Färbung und die
immunhistochemische Elektronenmikroskopie, der Aufklärung der Funktion, der Art
und der Lokalisation der einzelnen peroxisomalen Enzyme und der Aufklärung der
verschiedenartigen morphologischen Erscheinungsbilder (MASTERS u. CRANE
1995).
Im Folgenden wird vor allem auf den Aufbau der Peroxisomen und deren
enzymatische Ausstattung im Hinblick auf die verschiedenen Funktionen
eingegangen. Außerdem wird kurz das Peroxisom im Zusammenspiel mit anderen

2 Literaturübersicht 15
Zellorganellen betrachtet. Im Anschluss werden die Mechanismen zur Regulation der
Peroxisomenzahl in der Zelle besprochen, bevor ein allgemeiner Überblick zu den
Hepatopathien des Hundes folgt.
Abbildung 2-1 Aufbau einer idealisierten tierischen Zelle (LÖFFLER et al. 2007)
2.1.1 Morphologie
Peroxisomen sind dynamische Organellen, die in Größe und Aussehen variieren
können. Darüber hinaus sind sie in der Lage, ihre räumliche Position in der Zelle
entlang des Zytoskelettes zu verändern (SCHRADER et al. 2003). Im Gegensatz zu
den Lysosomen, die sich eher perikanalikulär aufhalten, folgt die Lage der
Peroxisomen im Leberläppchen keinem festgelegten Muster (NOVIKOFF u.

16 2 Literaturübersicht
GOLDFISCHER 1969). In gesunden Hepatozyten liegen die Peroxisomen homogen
im Hepatozyten verteilt vor und das Auftreten von Peroxisomen in Gruppen ist üblich
(ROELS et al. 1983).
Peroxisomen können eine sphärische bis längliche Gestalt haben (Abbildung 2-2),
sich aber auch in Form eines tubulär-retikulären Netzwerkes darstellen, welches
häufig mit Fetttropfen assoziiert ist (SCHRADER 2001).
Abbildung 2-2 Elektronenmikroskopische Darstellung eines Peroxisoms (Pfeil) mit Marginalplatte und Nukleoid. (P596-07/K1979), Ultradünnschnitt, Transmissionselektronenmikroskopie, Gerätevergr. 16.000x).
Peroxisomen besitzen im Zentrum einen dunklen homogenen Kern, der kristalline
Strukturen enthält (BERNHARD u. ROUILLER 1956) und nach BAUDHUIN et al.
(1965) als Kern mit „amorpher und fein granulärer Struktur“ beschrieben wird. Der
Kern, auch Nukleoid genannt, ist in Abhängigkeit zur Schnittebene des
elektronenmikroskopischen Präparates nicht in allen Peroxisomen der Leber
sichtbar. Ebenso abhängig von der Schnittebene ist das Erscheinungsbild des
Nukleoids: feingranulär oder polytubulär (DE DUVE u. BAUDHUIN 1966). Im

2 Literaturübersicht 17
Längsschnitt haben die Nukleoide eine längliche rechteckige Gestalt, während sie im
Querschnitt rund erscheinen. Sie liegen frei in der Matrix der Peroxisomen. Der
Umfang der Nukleoide beträgt 0,69-0,74 µm (LAGING et al. 1988). Allgemein ist das
Auftreten der Nukleoide in Leber und Nieren verbunden mit der Anwesenheit von
Uratoxidase, so dass Organismen ohne Nukleoide auch nicht über das Enzym
Uratoxidase verfügen. Es gibt aber Abweichungen davon, so dass Nieren von Ratten
und Hepatozyten des Menschen keine Uratoxidaseaktivität zeigen, jedoch über
Nukleoide verfügen und einige Nichtvertebraten, große Mengen an Uratoxidase
beherbergen, aber keine entsprechend hohe Anzahl an Nukleoiden aufweisen
(MASTERS u. CRANE 1995).
Die Membran der Peroxisomen besteht aus einer trilamellären Struktur, die eine
Dicke von 4,5 bis 8 nm aufweist (HRUBAN et al. 1966). Damit ist sie deutlich dünner
als die Membran von Lysosomen (MASTERS u. CRANE 1995).
Die Peroxisomen des Hundes sind 0,3-0,9 µm im Diameter und somit etwas kleiner
als die Peroxisomen der Ratte. Die Peroxisomen in der Leber von Dalmatinern sind
im Vergleich mit denen von Beagle deutlich größer und weisen eine höhere Dichte
auf (LAGING et al. 1988). Peroxisomen des Hundes enthalten kristalloide Nukleoide,
die aus 3-8 parallel verlaufenden Linien bestehen (SHNITKA 1966). Das Nukleoid
des Peroxisoms beim Hund ist Sitz der Urikase (BAUDHUIN et al. 1965; HAYASHI et
al. 1976; CHEVILLE 1994a). Peroxisomen in Zellen der kaninen Perianaldrüse
(KUHN 1968) sowie der Leber und der Niere (HRUBAN u. RECHCIGL 1969)
besitzen eine Marginalplatte, in Form einer elektronendichten Verdickung unter der
Membran, die durchschnittlich 0,25-0,39 µm lang ist (LAGING et al. 1988). In diesem
Teil des Peroxisoms ist die L-alpha-Hydroxysäureoxidase B lokalisiert (ZAAR 1992).
Die Peroxisomen des Hundes nehmen nur ca. 3,9 % des Zytoplasmas ein, obwohl
ca. doppelt so viele in einer Zelle vorhanden sind wie Mitochondrien, die 24 % des
zytoplasmatischen Volumens ausmachen (HESS et al. 1973).
Aufgrund ihrer charakteristischen morphologischen Eigenschaften sollen
Peroxisomen auch ohne spezielle Färbung eindeutig von den häufig räumlich
benachbarten Mitochondrien zu unterscheiden sein (BERNHARD u. ROUILLER

18 2 Literaturübersicht
1956). Eine Abgrenzung von Lysosomen ist ebenfalls möglich, da Peroxisomen
kleiner sind, ein homogeneres Erscheinungsbild besitzen, häufig zentral ein Nukleoid
aufweisen und eng mit dem Endoplasmatischen Retikulum assoziiert sind (SHNITKA
1966). Darüberhinaus sind Lysosomen in der Lage Substrate zu inkorporieren, eine
Eigenschaft, die den Peroxisomen fehlt (SHNITKA 1966).
Im Rahmen eines schnellen Zellwachstums können die Peroxisomen sehr
verschiedenartige und komplexe Formen annehmen. Dieses Phänomen kann
beispielsweise durch eine partielle Hepatektomie, Stimulation mit
Wachstumsfaktoren, Zugabe freier Fettsäuren oder freier Radikale provoziert werden
(SCHRADER et al. 1998a; SCHRADER et al. 1999).
2.1.2 Funktion
Die Peroxisomen sind in ein großes Spektrum an Aufgaben und Funktionen in der
eukaryontischen Zelle involviert (PLATTA u. ERDMANN 2007). Sie sind besonders
an Oxidations- und Detoxifikationsvorgängen beteiligt (CHEVILLE 1994e). Die
Aufgaben der Peroxisomen variieren je nach Organismus, Gewebe und der
Umgebung, in der sie sich befinden. Daher verfügen sie in den einzelnen Geweben
über unterschiedliche Enzymausstattungen (MANNAERTS u. VAN VELDHOVEN
1992) mit über 50 verschiedenen Enzymen (DE DUVE u. BAUDHUIN 1966). Davon
sind die meisten ausschließlich in Peroxisomen lokalisiert, während einige außerdem
in Mitochondrien vorkommen (WANDERS u. WATERHAM 2006). Trotz der großen
enzymatischen Variation gilt jedoch das Auftreten von Oxidase und Katalase in den
Peroxisomen als konstant und dient somit ihrer Charakterisierung und Identifizierung
(MASTERS u. CRANE 1995).
Mehr als siebzig Prozent der peroxisomalen Enzyme befinden sich in der Matrix.
Einige wenige Enzyme sind im Nukleoid lokalisiert und der verbleibende Anteil ist in
der einfachen Membran der Peroxisomen angesiedelt (MANNAERTS u. VAN
VELDHOVEN 1992).
Das Aufgabenspektrum ist sehr komplex und umfasst den Peroxidmetabolismus, die
ß-Oxidation sowie die Biosynthese von Etherphospholipiden (Plasmalogene), welche

2 Literaturübersicht 19
besonders im Nervensystem zu finden sind. Die Synthese von Cholesterol,
Gallensäuren, Leukotrienen, Prostaglandinen und der Metabolismus von Xenobiotika
obliegt ebenfalls den Peroxisomen. Cholesterol ist ein wichtiger Bestandteil von
Membranen und dient als Komponente von Lipoproteinen. Darüber hinaus ist es an
der Synthese von Gallensäuren und Steroiden beteiligt (MASTERS u. CRANE 1995).
Da Peroxisomen auch am Abbau insbesondere von sehr langkettigen Fettsäuren
beteiligt sind, haben ihre Abwesenheit oder das Fehlen eines ihrer Enzyme, wie
Katalase oder Oxidase, metabolische Konsequenzen. Zahlreiche angeborene
Erkrankungen, darunter das Zellweger Syndrom, lassen Peroxisomen in den
Körperzellen missen und führen dadurch zu schweren körperlichen Anomalien, die
mit einem Überleben über das Säuglingsalter hinaus nicht vereinbar sind. Ein
anderes Beispiel ist der Urikasedefekt des Dalmatiners, bei dem das dunkle Zentrum
des Peroxisomes, als Sitz der Urikase definiert, beim Dalmatiner deutlich kleiner
ausgeprägt ist als bei anderen Hunderassen. Das klinische Bild ist durch deutlich
erhöhte Harnsäureaussscheidung charakterisiert mit der Gefahr der Uratsteinbildung
in den harnbildenden und –ableitenden Wegen (CHEVILLE 1994a).
Im Folgenden werden die wichtigsten Funktionen und Aufgaben der Peroxisomen
näher erläutert.
2.1.2.1 Peroxidmetabolismus
Eine der wichtigsten Funktionen der Peroxisomen ist die Neutralisierung von freien
Radikalen mit Hilfe enzymatischer Mechanismen, die im Folgenden besprochen
werden.
Oxidativer Stress ist ein Zustand der Imbalanz zwischen prooxidativ wirkenden
Komponenten (Oxidantien) und antioxidativ wirkenden Abwehrmechanismen
(Antioxidantien), der durch einen Überschuss an freien Radikalen gekennzeichnet ist
(DJORDJEVIC 2004). Der Redox Status ist dabei ein übergeordneter Begriff, der
beide Mechanismen zusammenfasst (WEBB u. TWEDT 2008).

20 2 Literaturübersicht
Freie Radikale wirken prooxidativ, da sie ein oder mehrere ungepaarte Elektronen
aufweisen (WEBB u. TWEDT 2008). Zu dieser Gruppe gehören das Superoxid-
Radikal O2-, das Hydrogen Radikal H•, das Hydroxyl Radikal OH• und das Hydrogen
Peroxid Radikal H2O2 (DJORDJEVIC 2004). Diese sind in der Lage, unterschiedliche
Biomoleküle wie Lipide, Proteine und Nukleinsäuren (DJORDJEVIC 2004)
anzugreifen und in ihrer Funktion schwer zu beeinträchtigen (LÖFFLER et al. 2007).
Die meisten freien Radikale stammen vom Sauerstoff ab, dann bezeichnet man sie
auch als reaktive Sauerstoffverbindungen (ROS=reactive oxygen species). Die
Radikalbildung ist unweigerlich an die mitochondriale Energieproduktion gekoppelt
(GEROK 1995; MANDELKER 2008; WEBB u. TWEDT 2008). Die Reduktion des
Sauerstoffes während der mitochondrialen Atmungskette erzeugt freie Radikale wie
das Superoxidradikal (MANDELKER 2008). Die zweite Gruppe der Radikale stammt
von Stickstoff ab und wird als Gruppe reaktiver Nitrogenverbindungen (RNS)
bezeichnet (MANDELKER 2008). Der gesunde Organismus wandelt ein Viertel des
eingeatmeten Sauerstoffes zu Radikalen um. Bei Krankheit werden sogar drei Viertel
des inspiratorischen Sauerstoffes in Radikale umgesetzt (DJORDJEVIC 2004).
Freie Radikale sind an der Pathogenese von zahlreichen Krankheitsprozessen
beteiligt, beeinflussen das Immunsystem negativ und beschleunigen den
Alterungsprozess der Zelle (DJORDJEVIC 2004; MANDELKER 2008; CAMOES et
al. 2009). So können zahlreiche unterschiedliche Ursachen wie Toxämien,
Infektionen, hypoxisch-ischämische Zustände, Hyperglykämien, die Verstoff-
wechslung von Xenobiotika, Hyperlipidämien, Hyperproteinämien, Neoplasien,
Entzündungen sowie Immunreaktionen und erhöhte metabolische Raten oxidativen
Stress auslösen. Vor allem bei alternden Geweben kann ein zu niedriger ATP
Gehalt, bedingt durch eine ungenügende mitochondriale Produktion, zu erhöhtem
oxidativen Stress führen (MANDELKER 2008). Außerdem entstehen freie Radikale
bei Absorption von Strahlung, dem Metabolismus von Ethanol in Hepatozyten und
Lipidperoxidation von ungesättigten Fettsäuren (DJORDJEVIC 2004). Der Vorgang
der Lipidperoxidation wird dabei selbst auch durch freie Radikale in Gang gesetzt
und dann durch sekundär gebildete Radikale aufrecht erhalten, welche das
umliegende Gewebe schädigen und so eine Kettenreaktion auslösen. Von der

2 Literaturübersicht 21
Lipidperoxidation sind besonders die sehr empfindlichen, mehrfach ungesättigten
Fettsäuren als Bestandteil von Zellmembranen und intrazellulären Organellen
betroffen (DJORDJEVIC 2004).
Die Folge der Lipidperoxidation ist eine Zunahme der Membranpermeabiltät, so dass
die Integrität von Zellorganellen wie Peroxisomen, Mitochondrien und Lysosomen
herabgesetzt wird. Darüber hinaus werden Enzyme und Rezeptoren inaktiviert.
Letztlich kann die Peroxidation zur Zerstörung aller Lipidmembranen führen
(HARMAN 1956). Mit der Zerstörung von Zellorganellen, wie z.B. den Mitochondrien,
beginnt der Alterungsprozess der Zelle, da die Energieproduktion abnimmt, vermehrt
freie Radikale anfallen und der oxidative Stress zunimmt, was wiederum zu einer
weiteren Zellschädigung führt (WALLACE 2001). Zusammenfassend kann also der
oxidative Schaden, verursacht durch freie Radikale, entweder die Ursache aber auch
die Folge mitochondrialer Dysfunktion sein (MANDELKER 2008).
Der Entstehung von Tumoren liegen Mutationen und unkontrollierte
Zellproliferationen zu Grunde. Die chronische Exposition durch oxidativen Stress
kann Mutationen der DNA zur Folge haben, die zur Expression modifizierter Gene
führen und somit das Wachstum von Tumoren unterstützen. Daher wird dem
oxidativen Stress auch eine Beteiligung an der Karzinogenese zugesprochen
(DJORDJEVIC 2004; KLAUNIG u. KAMENDULIS 2004).
Antioxidantien
Antioxidative Mechanismen wirken den freien Radikalen entgegen und können in
enzymatische und nicht enzymatische Prozesse eingeteilt werden. Im Folgenden
werden nur die enzymatischen Antioxidantien besprochen, da diese in Peroxisomen
vorkommen. Zu dieser Gruppe gehören die Katalase, als ein Bestandteil der
Peroxisomen, die Glutathionperoxidase und die Superoxiddismutase (SOD) (GEROK
1995; SCHRADER u. FAHIMI 2004). Glutathionperoxidasen sind wichtige
Bestandteile des antioxidativen Schutzsystems aller Zellen, kommen aber auch im
Extrazellularraum vor (LÖFFLER et al. 2007). Das Enzym ist in der Matrix und in den
Mitochondrien lokalisiert und kann als ein alternatives Enzym zur Katalase betrachtet
werden. Es ist in der Lage, organische Peroxide, wie Lipidperoxide (CHEVILLE

22 2 Literaturübersicht
1994b; DJORDJEVIC 2004; LÖFFLER et al. 2007) oder Wasserstoffperoxide
abzubauen, wenn die Katalaseaktivität ausgeschaltet ist, wie z.B. im Falle einer
Endotoxämie oder des Ischämie-Reperfusionssyndroms (SINGH et al. 1994).
Die Superoxiddismutase (SOD) existiert in verschiedenen Varianten mit
unterschiedlichen Metallionen im aktiven Bereich des Enzyms. CuZn SOD findet sich
im Zytoplasma, im Kern und in Peroxisomen von Säugetieren (DJORDJEVIC 2004).
Weiterhin existiert eine SOD im Serum und anderen extrazellulären Flüssigkeiten
(MARKLUND 1980). Eine erhöhte Aktivität der SOD wird über Zytokine wie IFN γ
induziert, während eine verminderte Aktivität über TNFα und TGF β eingeleitet wird
(MARKLUND 1992). Die SOD unterstützt die Beseitigung von Superoxidanion- und
Hydroxyperoxidradikalen, indem das weniger reaktionsfähige Wasserstoffperoxid
und molekularer Sauerstoff entsteht (GEROK 1995). Das Wasserstoffperoxid dient
dann wiederum der Katalase als Substrat (MASTERS u. CRANE 1995;
ANGERMULLER et al. 2009).
Peroxisomen verfügen sowohl über Oxidasen, die Sauerstoff zu Wasserstoffperoxid
reduzieren können, als auch über Katalasen, die Wasserstoffperoxid zu Wasser
reduzieren (BAUDHUIN et al. 1965; DE DUVE u. BAUDHUIN 1966; SHNITKA 1966).
Die Enzyme Katalase und einige Oxidasen sind in der Matrix lokalisiert (BAUDHUIN
et al. 1965). Es existieren verschiedene Oxidasen, die ein spezielles Endprodukt und
zusätzlich Wasserstoffperoxid produzieren. Zu den peroxisomalen Oxidasen gehören
die L-α-Hydroxyacid-Oxidasen A und B. Hydroxyacid-Oxidase A ist in der Matrix und
Hydroxyacid-Oxidase B ist in der Marginalplatte lokalisiert. Beide Enzyme finden sich
insbesondere in Leber und Niere. Eine weitere Oxidase, die D-Aminoacid-Oxidase,
ist außer in den Peroxisomen auch frei im Zytosol der Zelle vorhanden (MASTERS u.
CRANE 1995).
Die Produktion und Reduktion der Wasserstoffperoxide mittels Katalase sowie die
Entgiftung anderer reaktiver Sauerstoffradikale schützt Zellen vor oxidativen
Schädigungen (TERLECKY et al. 2006). Die Katalase ist ein ubiquitäres Enzym
(SHARMA et al. 1989), das 40 % des peroxisomalen Proteins ausmacht (DE DUVE
u. BAUDHUIN 1966) und eine besonders hohe Aktivität in der Leber aufweist. Außer

2 Literaturübersicht 23
in Peroxisomen ist das Enzym auch in Mitochondrien lokalisiert, eine für die Zelle
günstige Lage, da hier viele Wasserstoffperoxid produzierende Enzyme ansässig
sind (SHARMA et al. 1989).
Die Katalase unterstützt sowohl katalytische als auch peroxidative Reaktionen.
Beide Reaktionen benötigen Wasserstoffperoxid. Im katalytischen Fall kommt ein
zweites Wasserstoffperoxid zur Reaktion dazu und es entsteht Wasser und
Sauerstoff. In der peroxidativen Reaktion reagiert Wasserstoff mit weiteren
Wasserstoffdonatoren, wie Alkoholen, Phenolen, Nitriten, Formaldehyd und primären
Aminen (MANNAERTS u. VAN VELDHOVEN 1992; GEROK 1995; MASTERS u.
CRANE 1995).
Katalytische Reaktion: H2O2 + H2O 2H2O + O2
Peroxidative Reaktion: H2O2 + RH2 2H2O + R
Peroxiosmen verfügen über zahlreiche weitere Enzyme. Eine umfassende Übersicht
bieten Kapitel 2 („Enzymology“) und Kapitel 3 („Intraparticulate organization of
peroxisomal proteins-methodology and topology“) des Buches „The peroxisome: a
vital organelle“ von Colin Masters und Denis Crane (MASTERS u. CRANE 1995).
Rolle der Mitochondrien bei oxidativem Stress
Mitochondrien werden ursächlich durch Hypoxie, freie Radikale und Toxinbelastung
geschädigt (VIEIRA u. KROEMER 1999).
Neben den vielseitigen, für die Zelle nützlichen Aufgaben, tragen die Mitochondrien
aber auch selbst zur Radikalbildung bei (ROTH 1997). O2- und H2O2 reagieren häufig
mit Eisen, das sich in den Mitochondrien befindet. Dadurch kann die mitochondriale
DNA geschädigt werden. Die Folge dieser Schädigung ist eine Verstärkung des
oxidativen Stresses aufgrund Expression veränderter Proteine, die verantwortlich für
die Elektronentransportkette sind. Es entsteht ein Circulus vitiosus mit verstärkter

24 2 Literaturübersicht
Radikalbildung (LEE u. WEI 1997) Unter physiologischen Bedingungen werden die
Radikale durch Antioxidantien neutralisiert (ROTH 1997). Das wichtigste Antioxidans
zum Schutz der Mitochondrien ist Glutathion (MANDELKER 2008). Bei
Krankheitszuständen, z.B. Sepsis, reichen diese Mechanismen jedoch nicht aus und
erschöpfen (ROTH 1997). Oxidativer Stress der Mitochondrien kann zu einer
Aktivierung der Mitochondrienexpression führen und damit zu Erhöhung der
Organellenanzahl (MANDELKER 2008). Neben dem Enzym Glutathion schützen
erhöhte peroxisomale Katalasekonzentrationen die Mitochondrien vor Fehl-
funktionen, während niedrige Konzentrationen zu einer mitochondrialen Dysfunktion
führen können (KOEPKE et al. 2008).
2.1.2.2 ß-Oxidation
In der Pilz- und Pflanzenzelle ist dieser biochemische Vorgang ausschließlich in den
Peroxisomen lokalisiert. In eukaryontischen Zellen findet die ß-Oxidation sowohl in
Mitochondrien als auch in Peroxisomen statt, mit dem Ziel, die Lipidhomöostase
aufrechtzuerhalten (WANDERS 2004b; POIRIER et al. 2006).
Das peroxisomale ß-Oxidationssystem dient über etwa fünf hintereinander
ablaufende ß-Oxidationszyklen der Verkürzung von sehr langen und toxischen
Fettsäuren (REDDY u. MANNAERTS 1994; MASTERS u. CRANE 1995;
HASHIMOTO 1999). Kurze Fettsäuren (<C8) sind kein geeignetes Substrat für
Peroxisomen (MANNAERTS u. VAN VELDHOVEN 1992). Die peroxisomale
Oxidationsrate der Fettsäuren durch die Peroxisomen nimmt bei gleichzeitig sich
verkürzender Kettenlänge ab. Ungesättigte Fettsäuren werden dahingegen
bevorzugt von den Peroxisomen oxidiert (MASTERS u. CRANE 1995).
Die daraus hervorgehenden kurz- und mittellangen Fettsäuren erreichen dann das ß-
Oxidationssystem der Mitochondrien. Diese Verstrickung der Stoffwechselwege zeigt
das enge Verhältnis von Mitochondrien und Peroxisomen (LAZAROW 1978;
HASHIMOTO 1987; MASTERS u. CRANE 1995). Der weitaus größte Anteil der
langen Fettsäuren wird in Mitochondrien der ß-Oxidation unterzogen. Die
Begründung liegt darin, dass die mitochondriale ß-Oxidation im Hinblick auf die

2 Literaturübersicht 25
Energiegewinnung um das zweifache effektiver ist als die peroxisomale ß-Oxidation
der Peroxisomen (MANNAERTS u. VAN VELDHOVEN 1992).
Die Enzyme zur peroxisomalen ß-Oxidation befinden sich in der Matrix. Dabei
unterliegt die peroxisomale Enzymausstattung einer Dynamik, die von dem Gewebe,
der Spezies und den Rahmenbedingungen sowie dem Einfluss von Umweltfaktoren
oder Xenobiotika abhängt (MASTERS u. CRANE 1995).
Die eigentliche ß-Oxidation gesättigter Fettsäuren verläuft in vier Schritten
(Abbildung 2-3):
1) Acyl-CoA wird mittels Acyl-CoA-Dehydrogenase zu 2-trans-Enoyl-CoA
oxidiert, dabei entsteht Wasserstoffperoxid.
2) Hydratationsschritt mittels Enoyl-CoA-Hydratase mit Zwischenprodukt L-
Hydroxyl-CoA.
3) Oxidationsschritt mittels L-3-Hydroxyacyl-CoA-Dehydrogenase mit dem
Zwischenprodukt 3-Ketoacyl-CoA
4) Aus 3-Ketoacyl-CoA entsteht durch das Enzym ß-Ketoacyl-CoA-Thiolase,
mittels der thiolytischen Spaltung des Ketoacyl-CoAs, ein Acetyl-CoA und eine
um zwei C-Atome verkürzte aktivierte Fettsäure, die wieder in den ß-
Oxidationszyklus einfließen kann (LAZAROW 1978; HASHIMOTO 1987)
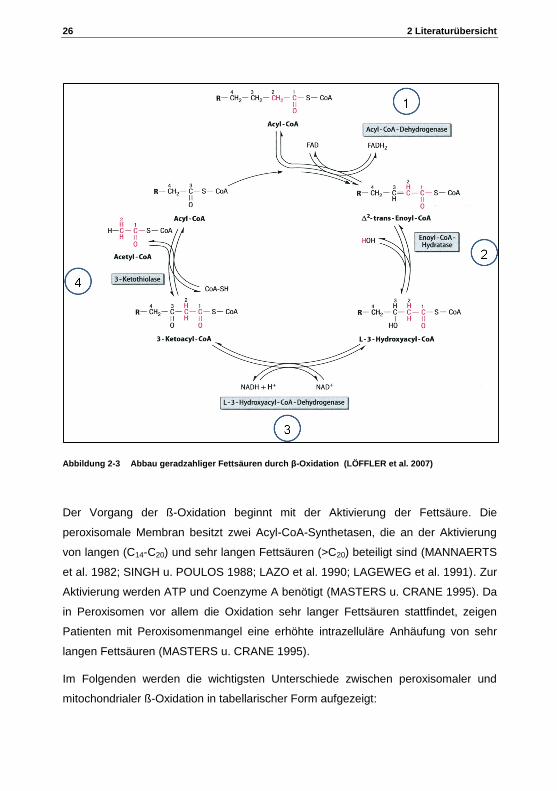
26 2 Literaturübersicht
Abbildung 2-3 Abbau geradzahliger Fettsäuren durch β-Oxidation (LÖFFLER et al. 2007)
Der Vorgang der ß-Oxidation beginnt mit der Aktivierung der Fettsäure. Die
peroxisomale Membran besitzt zwei Acyl-CoA-Synthetasen, die an der Aktivierung
von langen (C14-C20) und sehr langen Fettsäuren (>C20) beteiligt sind (MANNAERTS
et al. 1982; SINGH u. POULOS 1988; LAZO et al. 1990; LAGEWEG et al. 1991). Zur
Aktivierung werden ATP und Coenzyme A benötigt (MASTERS u. CRANE 1995). Da
in Peroxisomen vor allem die Oxidation sehr langer Fettsäuren stattfindet, zeigen
Patienten mit Peroxisomenmangel eine erhöhte intrazelluläre Anhäufung von sehr
langen Fettsäuren (MASTERS u. CRANE 1995).
Im Folgenden werden die wichtigsten Unterschiede zwischen peroxisomaler und
mitochondrialer ß-Oxidation in tabellarischer Form aufgezeigt:

2 Literaturübersicht 27
Tabelle 2-1 Unterschiede peroxisomaler und mitochondrialer β-Oxidation
Funktion
ß-Oxidation
(Peroxisom)
ß-Oxidation
(Mitochondrien)
Fettsäuretransport in die Organelle
(MANNAERTS et al. 1979; MASTERS u.
CRANE 1995)
membranständiges Transportsystem,
Carnithin unabhängig
Carnithin abhängig
Energieweiterverarbeitung (LAZAROW u.
DE DUVE 1976; MANNAERTS et al.
1979)
Abgabe von Wärme
Verknüpfung mit oxidativer
Phosphorylierung/Elektronentransportkette
Ablauf der ß-Oxidation (LAZAROW 1978) unvollständig,
lediglich Verkürzung von Fettsäuren
vollständig
Funktion der ß-Oxidation
(LAZAROW 1987; HAYASHI u. MIWA
1989; HAYASHI u. TAKAHATA 1991;
OSMUNDSEN et al. 1991)
Substratbereitstellung für anabole
Vorgänge (Cholesterin- u. Gallen-
säuresynthese, Phospholipide)
Eliminierung schlecht
verstoffwechselbarer Verbindungen
Bereitstellung von Energie für zelluläre
Abläufe
Enzymatische Ausstattung
(MASTERS u. CRANE 1995) 3 Enzyme
4 Enzyme
Energiestatus der Zelle
(MASTERS u. CRANE 1995) ATP unabhängig
ATP abhängig
steigt mit sinkendem ATP Gehalt
2.1.2.3 Weitere Funktionen der Peroxisomen
Synthese der Plasmalogene
Die Peroxisomen verfügen über eine enzymatische Ausstattung zur Synthese von
Plasmalogenen. Diese werden außer in den Peroxisomen auch im
Endoplasmatischen Retikulum hergestellt (MANNAERTS u. VAN VELDHOVEN
1992). Im menschlichen Organismus bilden die Plasmalogene 18 % der gesamten
Phospholipide (WANDERS u. WATERHAM 2006) und machen mehr als 80 % des
Phospholipidgehaltes der weißen Substanz im Gehirn aus (WANDERS 2004a).
Cholesterinsynthese
Außerdem enthalten Peroxisomen Anteile der enzymatischen Ausstattung zur
Bildung des Cholesterols (WANDERS u. WATERHAM 2006). Die Menge des in den
Peroxisomen produzierten Cholesterols ist jedoch im Vergleich zur Cholesterol-
synthese des Endoplasmatischen Retikulums sehr klein. Die physiologische Rolle

28 2 Literaturübersicht
der peroxisomalen Cholesterolsynthese ist bis jetzt unklar (MANNAERTS u. VAN
VELDHOVEN 1992).
Harnsäurestoffwechsel
Das Enzym Uratoxidase oder auch Urikase ist am Purinkatabolismus beteiligt und
findet sich in den meisten Säugetierperoxisomen (USUDA et al. 1988). Dort ist es,
wie auch die Xanthinoxidase bevorzugt, im Nukleoid lokalisiert. Es gibt jedoch auch
Vertebraten, die eine nennenswerte peroxisomale Aktivität dieses Enzyms
aufweisen, ohne Nukleoide zu besitzen (MASTERS u. CRANE 1995). Die
Xanthinoxidase verstoffwechselt Xanthin zu Harnsäure, während die Uratoxidase die
Oxidation von Urat mit Hilfe von molekularem Sauerstoff zu Allantoin katalysiert
(MASTERS u. CRANE 1995). Bei einigen Neuweltaffen und dem Menschen fehlt die
Uratoxidase in hepatozellulären Peroxisomen, sodass bei diesen Spezies Harnsäure
als Endprodukt des Purinkatabolismus ausgeschieden wird (USUDA et al. 1988;
MASTERS u. CRANE 1995). Primaten verfügen dagegen über eine gewisse
Leberuratoxidaseaktivität und zeigen daher nur niedrige Harnsäurespiegel
(MASTERS u. CRANE 1995).
Die Gicht des Menschen ist eine Erkrankung in Folge des Mangels an Uratoxidase
(MANNAERTS u. VAN VELDHOVEN 1992; HAYASHI et al. 2000). Es ist das einzige
peroxisomale Enzym, das in der veterinärmedizinischen Literatur für den Hund
beschrieben ist und befindet sich in den hepatozellulären Peroxisomen im Nukleoid
(LAGING et al. 1988). Der Dalmatiner besitzt im Vergleich zum Beagle eine höhere
Anzahl und auch größere Peroxisomen. Es handelt sich hier um einen
kompensatorischen Mechanismus, um einer erhöhten Harnsäurekonzentration im
Blut vorzubeugen. Die Unfähigkeit der Hepatozyten des Dalmatiners, aus Harnsäure
mittels der in den Peroxisomen lokalisierten Urikase Allantoin zu synthetisieren, liegt
jedoch nicht an dem bei dieser Rasse vorherrschenden Urikasemangel, sondern
aufgrund eines Transportdefektes, an einer verminderten Fähigkeit der Hepatozyten,
die Harnsäure in die Hepatozyten hineinzutransportieren (GIESECKE et al. 1985;
LAGING et al. 1988).

2 Literaturübersicht 29
2.1.3 Interaktion mit Mitochondrien
Peroxisomen sind Organellen, die nicht isoliert im Zytoplasma liegen, sondern
aufgrund metabolischer Zusammenhänge mit anderen subzellulären Strukturen, wie
den Mitochondrien, dem Endoplasmatischen Retikulum, Fetttropfen und dem Zytosol
assoziiert sind (WANDERS 2004b; POIRIER et al. 2006).
Wie bereits beschrieben, ist die ß-Oxidation ein grundlegender Mechanismus, der
ebenso wie die Thermogenese sowohl in Peroxisomen als auch in Mitochondrien
stattfindet (REDDY u. MANNAERTS 1994; WANDERS 2000, 2004b). Darüber
hinaus teilen sie sich Komponenten in der Zelle, die zur Organellenteilung notwendig
sind (SCHRADER 2006; SCHRADER u. YOON 2007). Vor kurzem wurde ein
Transportmechanismus zwischen Mitochondrium und Peroxisom entdeckt
(NEUSPIEL et al. 2008), der dazu dienen könnte, Metabolite, Lipide oder Proteine
zwischen den beiden Organellen auszutauschen (KOCH et al. 2005). Dazu befindet
sich an der äußeren mitochondrialen Membran eine Ligase MAPL (mitochondria-
anchored protein ligase), welche ein „RING-Finger Motiv“ umfasst und die
Morphologie der Mitochondrien reguliert. Diese MAPL Struktur kann dann in Form
von Vesikeln (MDV=mitochondria derived vesicles) von Mitochondrien abschnüren
und später mit Peroxisomen verschmelzen. Somit teilen sich Mitochondrium und
Peroxisom nicht nur wichtige Regularien zur Teilung, sondern sind auch über
metabolische Funktionen eng miteinander verknüpft (NEUSPIEL et al. 2008). Eine
andere Funktion der MDV´s könnte der Rücktransport von zu den Mitochondrien
fehlgeleiteten peroxisomalen Membranproteinen sein, was häufig unter
experimentellen Bedingungen beobachtet wird (SACKSTEDER et al. 2000).

30 2 Literaturübersicht
2.2 Kontrolle der Peroxisomenzahl
Die Zelle hat die Fähigkeit auf metabolischen oder umgebungsbedingten Stress zu
reagieren, indem sie die Zahl der Peroxisomen erhöht, bei Bedarf jedoch auch
überschüssige oder geschädigte Organellen entfernt (YAN et al. 2005).
Peroxisomen sind Zellorganellen, die sich sowohl über Wachstum und Teilung
vermehren, als auch aufgrund ihrer engen Assoziation mit dem Endoplasmatischen
Retikulum neu gebildet werden können (GRABENBAUER et al. 2000; SCHRADER
u. FAHIMI 2006).
Die Peroxisomen sind sehr stark in Hepatozyten vertreten. In einem Hepatozyten
finden sich bis zu 1000 Peroxisomen in enger Verbindung zum Endoplasmatischen
Retikulum (CHEVILLE 1994e). Weiterhin findet man sie in großer Zahl in renalen
Tubuluszellen und in steroidsynthetisierenden Zellen (CHEVILLE 1994e).
Die Verteilung von Peroxisomen innerhalb einer Zelle bzw. im Zellverband bedarf der
Bewegung einzelner Organellen. Für diesen Zweck benötigen die Peroxisomen der
Säugetiere auf Mikrotubuli basierende Filamente (SCHRADER et al. 2003). Die
Motilität der Organelle dient dazu, eine gleichmäßige Verteilung innerhalb der Zelle
aufrechtzuerhalten, mit anderen Peroxisomen oder anderen Zellorganellen in Kontakt
zu treten oder auch Zellkompartimente zu erreichen, die aufgrund der metabolischen
Situation Peroxisomen benötigen (SCHRADER et al. 2003). Es können zwei
Bewegungsarten unterschieden werden. Die erste Variante beinhaltet eine
langsame, nicht gerichtete Bewegung, der sich ca. 95 % der Peroxisomen bedienen.
Sie benötigt keine Mikrotubuli und ist energieunabhängig. Die zweite Möglichkeit der
Bewegung ist eine energieverbrauchende, schnelle, gerichtete Motilität, die an
Mikrotubuli gekoppelt ist (WIEMER et al. 1997). Weitere Untersuchungen der
Peroxisomenmotilität an kultivierten Zellen ergaben, dass sich elongierte oder
retikulär ausgerichtete Peroxisomen langsamer und ohne Mikrotubuli bewegen,
während sphärische Peroxisomen deutlich schneller entlang des Mikrotubulisystems
ihre Lokalisation verändern können (SCHRADER 2001).

2 Literaturübersicht 31
In eukaryontischen Zellen gibt es drei verschiedene Wege, die die Anzahl der
Peroxisomen in der Zelle beeinflussen können (YAN et al. 2005).
Einer der drei Mechanismen umfasst die Peroxisomenteilung im Rahmen der
Zellteilung mit Aufteilung der Organellen auf die Tochterzellen (VEENHUIS et al.
2003). Der zweite Mechanismus beinhaltet die Autophagie von überflüssigen oder
geschädigten Peroxisomen (FARRE u. SUBRAMANI 2004; YAN et al. 2005). Der
letzte Vorgang umfasst die übermäßige starke numerische Zunahme von
Peroxisomen in kurzer Zeit unabhängig vom Zellzyklus; dieser Vorgang wird
allgemein als Peroxisomenproliferation bezeichnet (YAN et al. 2005).
2.2.1 Biogenese und Proliferation
Die Neuentstehung der Peroxisomen war und ist ein ständiger Diskussionspunkt
(PLATTA u. ERDMANN 2007) und seit ihrer Entdeckung 1954 gibt es dazu viele
Theorien (BERNHARD u. ROUILLER 1956).
Die ersten Untersuchungen, die zur Aufklärung der Biogenese von Peroxisomen in
eukaryontischen Zellen beitrugen, wurden an Hefen und Pflanzen durchgeführt und
die daraus gewonnenen Erkenntnisse auf die Säugetierzelle übertragen (GOULD u.
VALLE 2000). Dabei zeigte sich, dass die Regulation und Transkription bestimmter
Gene, sogenannter PEX-Gene, und die daraus hervorgehenden Proteine, die
sogenannten Peroxine, maßgeblich an der Entstehung von Peroxisomen beteiligt
sind.
Bis heute sind 32 PEX Gene bekannt, die an drei verschiedenen Schlüsselpositionen
der Peroxisomentwicklung involviert sind (VAN DER ZAND et al. 2006): an der
Bildung der peroxisomalen Membran, der Peroxisomenproliferation und der
Bereitstellung und Positionierung von peroxisomalen Matrixproteinen (PLATTA u.
ERDMANN 2007).
Peroxisomenteilung
Hierbei können zwei grundlegende Teilungsvorgänge unterschieden werden. Zum
einen findet während der Zellteilung eine Teilung der vorhandenen Peroxisomen

32 2 Literaturübersicht
statt, um eine gleichbleibende Anzahl an Peroxisomen in den Tochterzellen zu
gewährleisten. Es handelt sich dann um eine konstitutive Teilung.
Zum anderen kann durch extrazelluläre Stimuli eine intrazelluläre Teilung der
Peroxisomen provoziert werden, die eine starke Vermehrung (Proliferation) zur Folge
hat. Diese Peroxisomenproliferation ist charakterisiert durch eine numerische
Zunahme, erhöhte Volumendichte, zunehmende Größe und gegebenenfalls eine
vermehrte Bereitstellung von peroxisomalen Enzymen. Dadurch wird erhöhten
metabolischen Anforderungen Rechnung getragen (SCHRADER u. FAHIMI 2006).
Peroxisomen enthalten keine DNA. Die Informationen zur Herstellung von
peroxisomalen Proteinen ist in nukleären Genen verschlüsselt (LAZAROW u. FUJIKI
1985). Biochemische Untersuchungen haben gezeigt, dass die Proteine der
Membran und der Peroxisomenmatrix an freien, im Zytosol befindlichen
Polyribosomen produziert und posttranslational in bereits bestehende Peroxisomen
transportiert werden. Hierauf begründet sich die Theorie, dass die Peroxisomen
Organellen sind, die sich wie Mitochondrien und Chloroplasten über Wachstum und
Teilung vermehren (LAZAROW u. FUJIKI 1985). Das Wachstum von Peroxisomen
wird dabei durch den Import von Membran- und Matrixproteinen gewährleistet. Bei
entsprechender Größe kommt es zur Aufteilung der Matrix und der peroxisomalen
Membran in mindestens zwei neue Peroxisomen (SCHRADER u. FAHIMI 2006).
Neuere Untersuchungen am Beispiel von Saccharomyces cerevisiae zeigen, dass
peroxisomale Bausteine aus dem Endoplasmatischen Retikulum hervorgehen,
insbesondere die peroxisomalen Membranteile, die bei der de-novo Synthese von
Peroxisomen benötigt werden (HOEPFNER et al. 2005; VAN DER ZAND et al.
2006).
Ausgehend vom Stadium des reifen Peroxisoms werden für eine Verdopplung
folgende Stadien durchlaufen (Abbildung 2-4) :
1) Verlängerung und Abschnürung
2) Teilung und Gruppenbildung
3) Trennung der Peroxisomen

2 Literaturübersicht 33
4) Wachstum der neu entstandenen Peroxisomen,
Wachstum und Reifung durch Matrixproteinimport
Verlängerung/ Abschnürung
Teilung/Gruppenbildung
Trennung
Wachstum/Reifung durch Matrixproteinimport
Reifes Peroxisom
1
2
3
4
Abbildung 2-4 Modell für die Teilung und Proliferation von Peroxisomen (modifiziert nach PLATTA u. ERDMANN 2007)
Wachstum und Reifung der Peroxisomen
Das Verfahren zur Proteinherstellung gilt sowohl für Matrixproteine als auch für
peroxisomale Membranproteine. Die im Endoplasmatischen Retikulum produzierten
peroxisomalen Membranproteine werden in das neu entstehende Peroxisom
eingebaut. Dadurch findet ein Wachstums- und Reifungsprozess statt, bis es
aufgrund der maximalen Größe erneut zu einer Teilung kommt. Damit die neu
synthetisierten Proteine die Peroxisomen erreichen, verfügen diese über
„peroxisomale targeting Signale“ (PTS). Zwei dieser Zielsignale sind bekannt unter
den Abkürzungen PTS1 und PTS2 (GOULD u. VALLE 2000; PLATTA u. ERDMANN
2007).

34 2 Literaturübersicht
Zum besseren Verständnis des peroxisomalen Wachstums kann der gesamte
Prozess in wenige wichtige Schritte unterteilt werden (Abbildung 2-5).
Der erste Schritt umfasst die Bindung eines neu synthetisierten, gefalteten Proteins
an den PTS1/Pex5-Rezeptor oder den löslichen Pex7-Rezeptor (1). Der
überwiegende Anteil der gefalteten Proteine wird über PTS1/Pex5-Rezeptor in das
peroxisomale Lumen transportiert, indem der Rezeptor zunächst das Protein zur
peroxisomalen Membran leitet (2). Anschließend erfolgt der Übertritt des Rezeptor-
Proteinkomplexes auf die luminale Seite der Peroxisomenmembran (3). Daraufhin
trennen sich Rezeptor und Protein (4). Das Protein verbleibt im Peroxisom, während
der Rezeptor in das Zytosol zurückkehrt, um für weitere peroxisomale
Matrixproteinimporte zur Verfügung zu stehen (GOULD u. VALLE 2000; PLATTA u.
ERDMANN 2007). Der Export des Rezeptors aus dem peroxisomalen Lumen zurück
ins Zytosol ist ein energieverbrauchender Prozess (OLIVEIRA et al. 2003) und auch
das Einschleusen der Proteine ist ATP abhängig. Zuerst werden die
Membranproteine synthetisiert, anschließend erfolgt die Synthese der Matrixproteine
(MASTERS u. CRANE 1995).

2 Literaturübersicht 35
Neues Protein
Neues Protein
Neues ProteinNeues
Protein
PTS1/Pex5 Rezeptor
1 2
PTS1/Pex5 Rezeptor
PTS1/Pex5 oder Lösl. Pex7 Rezeptor
3
Neues Protein
4
oder Zytosol
peroxisomaleMatrix
Löslicher Pex7
Rezeptor
PTS1/Pex5 Rezeptor
peroxisomaleMembran
Abbildung 2-5 Translokation von Proteinen zum Wachstum der Peroxisomen (modifiziert nach PLATTA u. ERDMANN 2007)
Auslösende Faktoren der Peroxisomenproliferation
Es gibt extrinsische und intrinsische Faktoren, die eine Peroxisomenproliferation
induzieren können (YAMAMOTO u. FAHIMI 1987). Eine Regeneration der Leber
nach einer partiellen Hepatektomie führt zu einer Peroxisomenproliferation in den
Hepatozyten (YAMAMOTO u. FAHIMI 1987). Auch zahlreiche Chemikalien,
Medikamente und Weichmacher, allgemein als Peroxisomenproliferatoren (PPO)
bezeichnet, können eine Vermehrung der Zellorganelle bewirken (LAZAROW u. DE
DUVE 1976; FAHIMI et al. 1982). Weichmacher, die sich z.B. in Plastikverpackungen
von Lebensmitteln befinden, haben in tierexperimentellen Studien gezeigt, dass sie
eine Peroxisomenproliferation auslösen können (MASTERS u. CRANE 1995).
Langzeitfütterungsversuche mit Ratten führten darüberhinaus zu der Entwicklung von
hepatozellulären Karzinomen (MASTERS u. CRANE 1995). Auch Medikamente, wie
Azetylsalizylsäure, anabole Steroide, Clofibrate, Trimethoprim Sulfonamide,
Spironolacton und orale Kontrazeptiva führen zu einer Erhöhung der

36 2 Literaturübersicht
Peroxisomenzahl (DE CRAEMER 1995). Die Verabreichung von
Schilddrüsenhormonen geht beispielsweise mit einer verminderten Katalaseaktivität
bei gleichzeitiger Erhöhung der Peroxisomenzahl von verminderter Größe einher
(JUST u. HARTL 1983; KERCKAERT et al. 1989). Die Stimulation von kultivierten
humanen Hepatozyten mit mehrfach ungesättigten Fettsäuren oder
Sauerstoffradikalen kann ebenfalls eine Peroxisomenproliferation auslösen
(SCHRADER et al. 1998a; SCHRADER et al. 1999). Zahlreiche Untersuchungen
haben gezeigt, dass es etwa 60 Xenobiotika gibt, die zu einer
Peroxisomenproliferation führen (MOODY et al. 1991). Darüber hinaus können
fettreiche Diäten, Hungerperioden, Vitamin E Mangel, Hyperthyreoidismus und
Anpassung an Kälte zu einer Proliferation der Peroxisomen führen (MASTERS u.
CRANE 1995). Die Peroxisomenproliferation, herbeigeführt durch PPO, fällt je nach
Lokalisation im Leberläppchen sehr variabel aus. In periportalen Hepatozyten finden
sich sehr viele elongierte Peroxisomen, während im perizentralen Bereich eher
rundliche Peroxisomen vorkommen (FAHIMI et al. 1996). Elongierte Peroxisomen
sind ein Zeichen für eine peroxisomale Proliferation und geben somit einen wichtigen
Hinweis auf eine stattgefundene Teilungsaktivität der Peroxisomen (SCHRADER et
al. 1996; SCHRADER et al. 1998b; KOCH et al. 2004). Vergleicht man diese
Beobachtung mit der Proliferation von hepatischen Stammzellen von periportal nach
perizentral (ARBER et al. 1988), kann daraus gefolgert werden, dass die
Peroxisomen dieser Proliferationsrichtung folgen (SCHRADER u. FAHIMI 2006). Bei
Proliferationsvorgängen wird auch eine perinukleäre Verteilung der Peroxisomen
beobachtet (ROELS et al. 1983; DE CRAEMER et al. 1993).
Regulation der Peroxisomenproliferation
Die Regulation der Peroxisomenproliferation im Säugetierorganismus erfolgt über
nukleäre Transkriptionsfaktoren. Diese Transkriptionsfaktoren gehören zur großen
Familie der Steroid-, Thyroid- und Retinoidrezeptoren und werden als „Peroxisome
proliferator activated receptors“ (PPARs) bezeichnet (ISSEMANN u. GREEN 1990).
Es werden drei Typen unterschieden: PPAR α, PPAR β und PPAR γ
(SCHOONJANS et al. 1996).

2 Literaturübersicht 37
PPAR α ist in den Lipidstoffwechsel, die Lipoproteinsynthese und den Ablauf von
Entzündungsreaktionen involviert (FEIGE et al. 2006). PPAR α wird stark in der
Leber exprimiert (KLAUNIG et al. 2003) und wird besonders bei Nagern durch
Lipidsenker sowie mehrfach ungesättigte Fettsäuren aktiviert (BEIER et al. 1992,
1997). PPAR α fungiert als Sensor für den Lipidgehalt der Hepatozyten und reguliert
die Genexpression für die mitochondriale und peroxisomale ß-Oxidation
(HASHIMOTO et al. 2000). Ein entscheidender Punkt in der Karzinogenese ist die
Aktivierung von PPAR α (GONZALEZ et al. 1998). PPAR β bestimmt das Maß des
Fettmetabolismus in Skelettmuskeln und Fettgewebe (LUQUET et al. 2005). Die
Bedeutung von PPAR γ liegt in der Regulation der Proliferation und
Ausdifferenzierung von Fettzellen (LUQUET et al. 2005).
2.2.2 Abnahme der Peroxisomenzahl durch Autophagie
Im Folgenden wird auf die Abnahme der Peroxisomenzahl in der Zelle eingegangen,
was insbesondere über Autophagie reguliert wird.
Autophagische Prozesse sind katabole intrazelluläre Abläufe, die letztendlich in
einen lysosomalen Abbau von Zellorganellen münden. Die Autophagie nimmt eine
wichtige Stellung ein, zum Beispiel in Hepatozyten, indem die nicht mehr benötigten
Zellorganellen abgebaut werden und die Abbauprodukte der Zelle als Nährstoffe
wieder zur Verfügung stehen (CUERVO 2004a; MIZUSHIMA 2005).
Es gibt drei verschiedene Formen der Autophagie, die alle ähnliche Funktionen
erfüllen: die Makroautophagie, die Mikroautophagie und die „Chaperon-mediierte
Autophagie“ (CUERVO 2004a). Diese drei Mechanismen kommen alle in
Säugetierzellen vor (CUERVO 2004a).
Bei der Makroautophagie werden die zu entsorgenden Komponenten in eine
Doppellamelle, dem Autophagosom, verpackt, welches dann mit dem Lysosom
verschmilzt und durch Hydrolasen abgebaut wird (SAKAI et al. 2006). Die
Membranen des lysosomalen Systems sowie die lysosomalen Proteine werden im
glatten Endoplasmatischen Retikulum (sER) synthetisiert (LÖFFLER et al. 2007).

38 2 Literaturübersicht
Im Rahmen der Mikroautophagie wird das zu entsorgende Material von der
lysosomalen Membran direkt umschlossen, als sogenanntes „Microautophagic body“,
und mittels Hydrolasen abgebaut (SAKAI et al. 1998; LÖFFLER et al. 2007).
Der Prozess der „Chaperon mediierten Autophagie“ beinhaltet die Bindung eines
Rezeptors an das zu eliminierende Substrat im Zytosol. Dieser Komplex bindet nun
an einen an der lysosomalen Membran befindlichen Rezeptor. Daraufhin erfolgt der
Übertritt des Substrates in das lysosomale Lumen, in dem die Degradierung durch
Hydrolasen stattfindet (CUERVO 2004b).
Funktion
Der Vorgang der Makroautophagie ist sowohl bei pathologischen, als auch
physiologischen Abläufen, wie der Beseitigung von apoptotischen Zellen, während
der Embryogenese, der Wiederverwertung von Nährstoffen und der Beseitigung von
fehlgefalteten Proteinen, intrazellulären Bakterien und geschädigten Zellorganellen
beteiligt (CUERVO 2004a; LEVINE u. KLIONSKY 2004). Es wird angenommen, dass
es sich in erster Linie um einen protektiven Mechanismus handelt (LEMASTERS et
al. 2002; CUERVO 2004a).
Auch zur Vermeidung von Neoplasien besitzt die Autophagie eine wichtige
Schutzfunktion. Durch die Beseitigung von geschädigten Mitochondrien und der
Reduktion des oxidativen und metabolischen Stresses wird auch die DNA vor
Angriffen durch Radikale geschützt, was Mutationen verhindert und die Stabilität des
Genoms wahrt (NELSON et al. 2004; KARANTZA-WADSWORTH et al. 2007;
MATHEW et al. 2007). Eine gestört ablaufende Autophagie führt jedoch zu einer
Genamplifikation, die letztlich ein Hauptmechanismus in der Onkogenaktivierung
darstellt (LITTLE u. CHARTRAND 2004).
Regulation
Der bedeutendste Faktor, der den Prozess der Autophagie in der Leber einleitet ist
Hungern. Hierbei wird in den ersten 48h bei Ratten und Mäusen 25-45 % des
zelleigenen Proteins abgebaut (ADDIS et al. 1936). Autophagie scheint dabei einen
wichtigen Mechanismus zur alternativen Energieversorgung darzustellen, um

2 Literaturübersicht 39
während Hungerperioden durch Recyling von Zellbestandteilen einen normalen
Metabolismus zu gewährleisten (JIN u. WHITE 2007). Autophagie trägt dadurch zur
Energieversorgung von Zellen in Hungerperioden bei und ist somit an der
Aufrechterhaltung der zellulären Homöostase beteiligt (NELSON et al. 2004).
Autophagie ist ein energieverbrauchender Prozess, der bei ATP Mangel zum
Erliegen kommt (MEIJER u. CODOGNO 2004). Auch Insulin übt einen
inhibitorischen Effekt aus (PFEIFER 1977), während Glukagon Autophagieprozesse
fördert (DETER et al. 1967). Die alimentäre Aufnahme von langen und sehr langen
Fettsäuren, die den Peroxisomen als Substrat dienen, führt ebenfalls zu einem
Abbruch der Pexophagie (LUIKEN et al. 1992).
2.3 Hepatopathien des Hundes
In der Humanmedizin existieren 17 erblich bedingte peroxisomale Erkrankungen, die
vor allem auf einer Biogenese- oder Funktionsstörung der Peroxisomen beruhen
(WANDERS 2004a). Für den Hund sind weder angeborene noch erworbene
peroxisomale Erkrankungen beschrieben. Um jedoch herauszufinden, ob
Peroxisomen bei bestimmten degenerativen bzw. neoplastischen Lebererkrankungen
eine Rolle spielen, wurden in der vorliegenden Arbeit Hunde mit Hepatopathien
untersucht, die im Folgenden hinsichtlich ihres makroskopischen und
mikroskopischen Erscheinungsbildes näher erläutert werden sollen.
Die Leber befindet sich zentral im kranialen Abdomen, dem Zwerchfell direkt
anliegend. Die Oberfläche ist glatt, da sie von einer serösen Kapsel überzogen ist.
Das Leberparenchym erscheint rotbraun, ist von fragiler Konsistenz und beim Hund
in zahlreiche Leberlappen unterteilt, deren Enden im gesunden Zustand scharf
konturiert sind (CULLEN 2007a).
Es können zwei Einteilungen hinsichtlich der architektonischen Bauweise der Leber
unterschieden werden: Die klassische Einteilung der Leber mit dem Leberläppchen
als kleinste anatomische Einheit. Das Leberläppchen bildet eine mehr oder weniger
sechseckige Struktur mit der in der Mitte befindlichen Zentralvene, die das Blut der

40 2 Literaturübersicht
Lebervene zuführt. An den Ecken des Sechseckes befinden sich die Portalfelder , die
auch als Glisson Trias bezeichnet bezeichnet werden. Sie schließen Gallengänge,
Zweige der Portalvene, der Leberarterie, Nervenfasern und Lymphgefäße ein,
gestützt von einem feinen retikulären Bindegewebe (Abbildung 2-6 A). Das Blut wird
aus der Leberarterie und Portalvene in die Sinusoide geleitet, wo es sich vermischt
und eng an den Hepatozyten vorbeifließt, bevor es über die Zentralvene in die V.
hepatica abgeführt wird (CULLEN 2007a).
Bei der funktionalen Einteilung der Leber nach RAPPAPORT (1980) stellt das
Portalgebiet den zentralen Punkt des sogenannten Leberazinus dar, da dort das
sauerstoffreiche Blut über die Arterie ankommt und zu den in der Peripherie
gelegenen Lebervenen abgeführt wird. Dabei nimmt der Sauerstoffgehalt von Zone 1
(periportal oder zentroazinär) über Zone 2 (mittzonal) nach Zone 3 (periazinär)
kontinuierlich ab (Abbildung 2-6 B) (CULLEN 2007a).
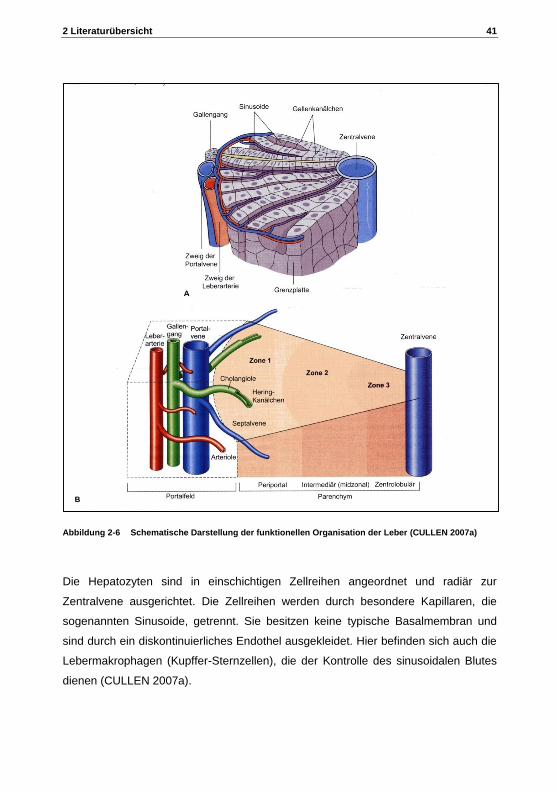
2 Literaturübersicht 41
Abbildung 2-6 Schematische Darstellung der funktionellen Organisation der Leber (CULLEN 2007a)
Die Hepatozyten sind in einschichtigen Zellreihen angeordnet und radiär zur
Zentralvene ausgerichtet. Die Zellreihen werden durch besondere Kapillaren, die
sogenannten Sinusoide, getrennt. Sie besitzen keine typische Basalmembran und
sind durch ein diskontinuierliches Endothel ausgekleidet. Hier befinden sich auch die
Lebermakrophagen (Kupffer-Sternzellen), die der Kontrolle des sinusoidalen Blutes
dienen (CULLEN 2007a).

42 2 Literaturübersicht
Die Hepatozyten besitzen zu der das Lumen der Sinusoide begrenzenden Seite
zahlreiche Mikrovilli, die der Oberflächenvergrößerung, der Aufnahme von
Substanzen aus dem Plasma und der Sekretion von Produkten dienen. An den
basolateralen Seiten der Hepatozyten befinden sich Gallekanälchen, die sich aus
den modifizierten Zellmembranen zweier benachbarter Hepatozyten ergeben. Der
Disse´ Raum ist eine besondere anatomische Struktur zwischen den Hepatozyten
und den Endothelzellen der Lebersinusoide. Hier findet ein Austausch von Stoffen
zwischen Plasma und den mit Mikrovilli besetzten Hepatozyten statt. Eine
Schädigung dieser Zone beeinträchtigt die Leberfunktion erheblich. In diesem Raum
befinden sich auch fettspeichernde Zellen, sogenannte Ito-Zellen, die unter anderem
der Vitamin A-Speicherung dienen. Bei Leberschädigungen sinkt ihr Vitamin A-
Gehalt und sie bilden Kollagen, das die Entstehung einer Leberfibrose fördert
(CULLEN 2007a).
2.3.1 Hepatozelluläre Degeneration
Die hepatozelluläre Degeneration, früher als Hepatose bezeichnet, umfasst nicht
primär entzündlich bedingte Veränderungen der Hepatozyten, beispielsweise durch
Sauerstoffmangel oder Stoffwechselstörungen. Die Veränderungen können folgenlos
bleiben, in ein von Entzündungsprozessen begleitetes chronisches Stadium
übergehen oder in einer chronischen Zerstörung von Lebergewebe mit vergeblichen
bindegewebigen Umbau- und Regenerationsprozessen (Zirrhose) enden (KÄUFER-
WEISS 2007).
Das Verteilungsmuster der auftretenden degenerativen Veränderungen gibt
Aufschluss über die möglichen Ursachen. So wird unterschieden, ob Leberzellen im
gesamten Parenchym (diffus), zufällig verteilt (multifokal) oder in bestimmten Zonen
(1-3) des Leberazinus geschädigt sind (ROTH u. MEYER 1995).
Hydropische Degeneration
Hepatozyten reagieren bei Störung ihrer Homöostase immer in ähnlicher Weise.
Dabei werden im allgemeinen die Elektrolytkonzentration und der Flüssigkeits-
haushalt derart gestört, dass es zu einem Wassereinstrom kommt und die Zelle

2 Literaturübersicht 43
anschwillt (CULLEN et al. 2006). Diesen Vorgang nennt man daher auch
hydropische Degeneration (CULLEN 2007b).
Mikroskopisch kann sich das zunächst in Form einer trüben Schwellung des
Hepatozyten darstellen. Dabei kommt es zu einer Schädigung der Mitochondrien
z. B. durch exogene Gifte, Hypoxie und/oder Stoffwechselentgleisungen, was zu
einer unspezifischen Leistungsschwäche der Hepatozyten führt (KÄUFER-WEISS
2007). Die ebenfalls durch Wassereinstrom vergrößerten Mitochondrien können
lichtmikroskopisch als gleichmäßig in der Leberzelle verteilte Körnchen
wahrgenommen werden (KÄUFER-WEISS 2007).
Abbildung 2-7 Hydropische Degeneration von Hepatozyten im Bereich der Zentralvene (Z=Zentralvene; Rind, Paraffinschnitt, H.E.) (CULLEN 2007a)
Bei fortschreitender hydropischer Schwellung zeigen die Zellen eine sogenannte
ballonierende Degeneration. Dabei nimmt der Einstrom von Wasser und Natrium
stetig zu. Die Konsequenz ist eine massive Schwellung der gesamten Zelle und
deren Organellen, insbesondere der Mitochondrien, der Lysosomen und des
Endoplasmatischen Retikulums (KÄUFER-WEISS 2007; STALKER u. HAYES 2007).

44 2 Literaturübersicht
Eine Zellschwellung kann reversibel sein, wenn das Ausmaß und die Dauer der
schädigenden Noxe begrenzt sind (Abbildung 2-8). Wird dabei jedoch ein kritischer
Punkt überschritten, so kommt es in der Folge zum irreversiblen Zelluntergang
(CULLEN 2007b)
Abbildung 2-8 Physiologische Zelle und Veränderungen bei reversibler und irreversibler Zellschädigung (CULLEN 2007b)
Auslöser einer hydropischen Degeneration ist im Prinzip immer eine gestörte
Zellatmung aufgrund hypoxischer Zustände, die entweder den gesamten
Organismus betreffen können oder nur lokal wirken. Eine Vergiftung oder
Blockierung mitochondrialer Enzyme führt ebenfalls zu einer gestörten Zellatmung
(KÄUFER-WEISS 2007). Daher ist die hydropische Degeneration eine relativ
unspezifische Zellveränderung, der viele Ursachen zugrunde liegen können
(CULLEN 2009). So können bei Jungtieren, die vom Wachstumsverhalten der
Wurfgeschwister abweichen, angeborene Erkrankungen, wie z. B. primäre

2 Literaturübersicht 45
Speicherkrankheiten der Leber eine ätiologische Rolle spielen (CULLEN 2009),
während bei älteren Tieren zahlreiche erworbene Erkrankungen (z. B. chronische
Vergiftungen, hohe Blutkortisolspiegel, Herzinsuffizienz, Adipositas) eine
hepatozelluläre Degeneration auslösen können. Im Rahmen einer hepatozellulären
Degeneration kommt es häufig zu Einlagerungen bestimmter Substanzen, da der
Hepatozyt nicht mehr in der Lage ist, diese angemessen zu verstoffwechseln. Dabei
handelt es sich am häufigsten um Fett oder Glykogen (STALKER u. HAYES 2007;
CULLEN 2009).
Glykogenose
Eine übermäßige hepatozelluläre Glykogeneinlagerung (Glykogenose) tritt im
Rahmen einer steroidinduzierten Hepatopathie auf und ist induziert durch endogene
oder exogene Glukokortikoide bzw. in seltenen Fällen durch Steroidhormone
(Progesteron, Aldosteron) (CULLEN et al. 2006; SEPESY et al. 2006). SEPESY et
al. (2006) gehen davon aus, dass auch eine stressinduzierte Hyperkortisolämie bei
akuten oder chronischen Erkrankungen an der Entstehung einer Glykogenose
beteiligt ist. Weiterhin beobachten sie außerdem Glykogeneinlagerungen im
Zusammenhang mit hepatischen und nicht hepatischen Neoplasien, auch wenn die
Hunde keine exogen zugeführten Glukokortikoide erhielten. Glukokortikoide führen
zu einer Anordnung der Peroxisomen um Glykogentropfen, wobei insgesamt die
Anzahl der Peroxisomen vermindert ist (PHILLIPS et al. 1987).
Im Falle einer steroidinduzierten Hepatopathie ist die Leber vergrößert und weist eine
blassbraune Farbe auf (CULLEN 2007a). Das Bild einer hepatozellulären
Glykogeneinlagerung zeigt sich als intrazytoplasmatische Vakuolisierung von
Hepatozyten mit überwiegend kleinen verwaschenen Vakuolen. Der Nukleus befindet
sich dabei zunächst noch in zentraler Position (CULLEN et al. 2006). Bei
zunehmender Glykogeneinlagerung kann es zu einer zwei bis zehnfachen
Vergrößerung der Hepatozyten mit Verdrängung des Kernes in die Peripherie
kommen. Die Veränderungen beginnen am häufigsten in der Zone 2 (intermediäre
Zone) des Azinus, können sich jedoch im weiteren Krankheitsverlauf zunehmend
ausbreiten. Im fortgeschrittenen Stadium gehören Zelluntergänge (Nekrosen), sowie

46 2 Literaturübersicht
neutrophile Entzündungszellinfiltrate als Reaktion auf den Gewebsuntergang zum
histologischen Bild. Die PAS-Färbung (periodic acid Schiff reaction) gibt Aufschluss
über den intrahepatozellulären Gehalt an Glykogen. Klinisch sind deutlich erhöhte
enzymatische Aktivitäten der ALT und der ALKP zu erwarten (STALKER u. HAYES
2007). Das Erkrankungsbild findet sich häufig bei Hunden. Auch angeborene
Glykogenspeicherkrankheiten führen zu ähnlichen Veränderungen der Hepatozyten
(STALKER u. HAYES 2007).
Lipidose
Der Lipidmetabolismus in der Leber wird sehr eng reguliert. Fettsäuren werden
oxidiert, um dem Energiebedarf zu decken; sie werden zu Triglyzeriden verestert, die
an VLDL (very long density lipoprotein) gebunden in die Fettdepots transportiert oder
in den Hepatozyten gespeichert werden (RAO u. REDDY 2004).
Bei der Anreicherung von Fetttropfen in Hepatozyten spricht man von einer
hepatischen Lipidose oder hepatozellulären Steatose, die im Extremfall zu einer
Leberverfettung führen kann (DE CRAEMER 1995; KÄUFER-WEISS 2007;
STALKER u. HAYES 2007). Dies beinhaltet eine absolute Erhöhung des
Fettgehaltes der Leberzellen (CULLEN et al. 2006; SCHRADER u. FAHIMI 2008).
Akute Lipidosen zeigen sich makroskopisch als vergrößerte blasse Lebern mit
brüchiger bis fettiger Konsistenz ohne Veränderung der Leberarchitektur.
Makroskopisch erscheinen die Ränder abgerundet (STALKER u. HAYES 2007).
Vorangeschrittene toxische Schädigungen von Hepatozyten führen zur
Verschmelzung von kleinen Lipidtropfen zu einem großen Tropfen, der den Kern aus
seiner zentralen Stellung verdrängt und zur Konturveränderung der Zelle führt. Die
Sinusoide werden komprimiert und vermindert durchblutet (STALKER u. HAYES
2007).
Eine Ansammlung von Triglyzeriden in Form von Fetttropfen kann sich histologisch
mikrovesikulär oder makrovesikulär darstellen (RAO u. REDDY 2004; STALKER u.
HAYES 2007). Die mikrovesikuläre Steatose beschreibt intrazelluläre, überwiegend
perinukleär angeordnete Lipidtropfen, die kleiner sind als der Kern, während bei einer

2 Literaturübersicht 47
makrovesikulären Steatose, die Lipidtropfen mindestens so groß wie der Kern oder
größer sind und den Nukleus nach peripher verdrängen (CULLEN et al. 2006).
Eine mikrovesikuläre Lipidose beginnt meistens zentrolobulär (Zone 3) im
Leberläppchen und kommt vor allem bei erhöhter Mobilisation von Fetten z. B. bei
Diabetes mellitus und juveniler Hypoglykämie von Toy Rassen (CULLEN et al. 2006)
und nicht selten bei toxisch bedingten Hepatopathien vor (STALKER u. HAYES
2007). Die mikrovesikuläre Lipidose tritt häufig im Zusammenhang mit Schädigungen
der Mitochondrien auf und ist hinsichtlich des Zellschadens als schwerwiegender
einzuordnen als die makrovesikuläre Lipidose (CULLEN 2009).
Bereits degenerierte Hepatozyten zeigen in besonderem Maße eine hepatische
Lipidose, da der Durchsatz von Fettsäuren und Triglyzeriden durch die Blockade an
entscheidenden Punkten im Lipidstoffwechsel und der Sekretion von VLDL
beträchtlich eingeschränkt ist. Das Fett, das als Substrat der ß-Oxidation dient,
sammelt sich somit im Hepatozyten (DE CRAEMER et al. 1995). Eine krankhafte
Verfettung der Leber kann demnach durch vielfältige alimentäre, metabolische,
toxische und hypoxische Ursachen eintreten (STALKER u. HAYES 2007). Die
hepatische Steatose kann, wie die hydropische Degeneration reversibel verlaufen.
Bei persistierender oder sehr heftiger Noxe im Zusammenhang mit ROS,
Endotoxinen, Tumornekrosefaktor alpha und unter dem Einfluss von bestimmten
Zytokinen kann sie sich zu einer Steatohepatitis ausweiten (DAY u. JAMES 1998).
Zu den verschiedenen Ursachen und Mechanismen, die eine Lipidansammlung in
Hepatozyten auslösen können, gehören:
prähepatische Ursachen: vermehrte Anfuhr und damit Aufnahme von
Fettsäuren (z. B. Mobilisation bei Laktation, Hunger, endokrine Störungen,
Trächtigkeit) (CULLEN 2007a; STALKER u. HAYES 2007)
intrahepatische Ursachen: abnorme Hepatozytenfunktion und Energie-
mangelzustände mit herabgesetztem Lipidstoffwechsel (CULLEN 2007a),
toxische und hypoxische Ursachen (CULLEN 2007a), Reduktion der Enzyme
zur ß-Oxidation in den Peroxisomen (FAN et al. 1998).

48 2 Literaturübersicht
posthepatische Ursachen: verminderte Apoproteinsynthese mit reduzierter
Bildung und Export von Lipoproteinen aus den Hepatozyten (CULLEN 2007a)
Längerfristig bestehende Lipidosen ziehen meist weitere Schädigungen nach sich.
So kommt es bei chronischen Zuständen zu Fibrose, Pigmentablagerungen und
nodulären Hyperplasien. Diese Veränderungen werden durch fortschreitende
peroxidative Schädigung und Aktivierung von Ito-Zellen hervorgerufen. Das erhöhte
Auftreten von Lipidtropfen erhöht auch den oxidativen Stress der Zelle, der zur
Verstärkung der Lipidperoxidation von Membranen beiträgt (STALKER u. HAYES
2007). In der Humanmedizin wird histologisch die Steatose auch mit Auftreten von
inflammatorischen Veränderungen, ballonierender Degeneration, Apoptose und
Entzündung beobachtet (RAO u. REDDY 2004).
2.3.2 Verhalten der Peroxisomen bei degenerativen
Veränderungen der Hepatozyten
Die ultrastrukturellen Veränderungen bei der hydropischen Leberzelldegeneration
umfassen Plasmamembranveränderungen wie „Blebbing“, Zerreißung von Mikrovilli,
Bildung von Myelinfiguren und der Verlust von Zellkontakten. Veränderungen der
Mitochondrien in Form von Schwellung und das Auftreten von amorphen
Einlagerungen sind häufig sowie die Dilatation des Endoplasmatischen Retikulums
und die Ablösung der Ribosomen (CULLEN et al. 2006).
Beim Menschen zeigen Peroxisomen der Hepatozyten im Rahmen einer Steatose
eine variable Katalaseaktivität, werden kleiner und proliferieren, wobei sie
unterschiedliche Erscheinungsformen wie anguläre, gebogene oder irreguläre
Konturen mit Protrusionen und Membranschleifen (Gastruloid cisternae) annehmen
können. Das peroxisomale Nukleoid verschwindet dabei (DE CRAEMER et al. 1995).
Die Mitochondrien der steatotisch veränderten humanen Leber nehmen beachtlich
an Größe zu und werden als Megamitochondrien bezeichnet. Dabei zeigen sie
parakristalline Einschlüsse und parallel arrangierte Cristae (DE CRAEMER et al.

2 Literaturübersicht 49
1995). Das Endoplasmatische Retikulum einer Fettleber ist dilatiert und zeigt
Vesikelbildung (DE CRAEMER et al. 1995).
In nahezu allen geschädigten Zellen, besonders in Hepatozyten, die zu den
metabolisch aktiven Zellen zählen, kommt es im Rahmen von degenerativen
Veränderungen zu einem Anstieg der Peroxisomenzahl. Die sehr pleomorphen
Peroxisomen lagern sich dabei im Bereich der Schädigung, z.B. in der Nähe von
Mitochondrien, gruppenweise an. Insgesamt erscheint die Peroxisomenproliferation
zentrolobulär ausgeprägter als perilobulär (CHEVILLE 1994c).
2.3.3 Hepatitiden
Entzündliche Leberveränderungen können in akute und chronische sowie in
spezifische infektiös bedingte und unspezifische reaktive Formen eingeteilt werden.
Das Verteilungsmuster ist dabei abhängig von der Entzündungsursache, der
Eintrittspforte, dem Infektionserreger bzw. dessen Zelltropismus innerhalb der Leber
(CULLEN 2007a; STALKER u. HAYES 2007).
Hämatogene Infektionen verursachen z. B. ein eher zufällig verteiltes, multifokales
Verteilungsmuster, während über die Gallenwege aszendierende Infektionen vor
allem in Portalbereichen ihre Manifestation finden, wobei schwerwiegende
Infektionen das gesamte Portalfeld und über dessen Grenzen hinaus
Entzündungsprozesse mit einbeziehen können (CULLEN 2007a).
Akute Hepatitis
Die akute Hepatitis ist durch das Auftreten von Entzündungserscheinungen mit
Infiltration von neutrophilen Granulozyten, Aktivierung von Kupffer-Zellen, Nekrosen
und Apoptosevorgängen von Hepatozyten charakterisiert. Die Zellpopulationen, die
an der Entzündungsreaktion beteiligt sind, variieren stark in Abhängigkeit von der
Ursache, der Wirtsantwort und dem Stadium der Läsion. Meist dominieren
neutrophile Granulozyten bei bakteriell bedingten Infektionen und
Protozoeninfektionen. Virale Infektionen führen in erster Linie zum Auftreten von
Nekrosen, Apoptosen einzelner Zellen und häufig nur minimalen

50 2 Literaturübersicht
Entzündungszellinfiltraten, meist dominiert von Lymphozyten (CULLEN 2007a).
Kupffer-Zellen spielen außerdem eine wichtige Rolle bei akuten
Entzündungsprozessen, da sie durch ihre Phagozytosefähigkeit zur Beseitigung von
pathogenen Noxen, wie z. B. Bakterien, beitragen. Im Zuge einer Aktivierung zu
sekretorisch aktiven Histiozyten schütten sie neben Zytokinen weitere Mediatoren
aus, die an hypertrophen und proliferativen Prozessen der Hepatozyten, der Ito-
Zellen und Endothelien beteiligt sind (STALKER u. HAYES 2007). Bakterielle
Leberentzündungen können beim Hund durch Leptospiren verschiedener Serovare
(L. grippothyphosa, L. pomona, L. bratislava, L. canicola, L. icterohaemorrhagiae),
bei Jungtieren durch Clostridium piliforme oder auch nach disseminierter Ausbreitung
im Organismus durch Nocardia asteroides verursacht werden. Ursache viraler
Hepatitiden sind das canine Adenovirus 1 und das canine Herpesvirus 1 (KÄUFER-
WEISS 2007).
Chronische Hepatitis
Bei immer wiederkehrenden Noxen oder unvollständiger Ausheilung akuter
Hepatitiden kommt es zur Entwicklung einer chronischen Hepatitis. Sie ist in erster
Linie gekennzeichnet durch eine Fibrose neben Infiltrationen von mononukleären
Entzündungszellen, wie Lymphozyten, Makrophagen und Plasmazellen.
Regenerationsprozesse in Form von proliferierenden Knötchen sind ebenfalls häufig
zu finden (CULLEN 2007a). Die Diagnose sollte die Dauer und den Grad der
Erkrankung einbeziehen. Die Aktivität einer Hepatitis wird durch den Grad der
Entzündungsreaktion und das Ausmaß von Apoptose und Nekrose bestimmt
(CULLEN et al. 2006). Beim Hund finden sich variable Mengen neutrophiler
Granulozyten. Dies weist daraufhin, dass ein akuter Entzündungsreiz bestehen bleibt
bzw. wiederholt auftritt (CULLEN 2007a).
Es können verschiedene Formen von chronischen Hepatitiden unterschieden werden
(CULLEN 2007a):
granulomatöse Hepatitis (Bildung einzelner Granulome; Auftreten fokal oder
disseminiert, Entzündungsgeschehen wird dominiert durch Makrophagen und

2 Literaturübersicht 51
außerdem mehrkernigen Riesenzellen, wenigen Lymphozyten und
Plasmazellen)
chronisch-eitrige Hepatitis (Abszesse: einzeln oder multifokal auftretend, gut
bindegewebig abgegrenzt/eingekapselte Prozesse)
idiopathische chronische Hepatitis (frühere Bezeichnung: chronisch aktive
Hepatitis)
o persistierende Entzündung der Leber mit gemischtzelligen
Entzündungszellinfiltraten
o Fibrose der Portalfelder
o Noduläre Parenchymregeneration
o Portale und periportale mononukleäre Zellinfiltration
o Cholestase
Weiterhin sind zwei Subtypen der chronischen Hepatitis für Hunde beschrieben:
progressive chronische Hepatitis (rassebedingte Kupfervergiftung bei
Dalmatiner, Bedlington und West Highland White Terrier, Pudel)
familiäre chronische Hepatitis (weibliche Hunde der Rasse Dobermann,
Skye Terrier, Cocker Spaniel und Labrador Retriever)
2.3.4 Lebertumoren
Lebertumoren können primär oder sekundär auftreten. Primäre Tumore des Leber-
und des Gallengangsystems haben ihren Ursprung in epithelialen Zellen, wie den
Hepatozyten, dem Gallengangsepithel sowie dem Epithel der Gallenblase oder
entstehen aus mesenchymalem Gewebe, wie Bindegewebe oder Zellen der
Blutgefäße. Sekundäre Tumore der Leber sind sehr häufig und stellen den größten
Anteil der Lebertumore aufgrund ausgeprägter Metastasierung in dieses Organ dar.
Am häufigsten metastasieren in die Leber maligne Lymphome, darüber hinaus
Melanome und Hämangiosarkome (CULLEN 2007a).

52 2 Literaturübersicht
Virale Erreger, Chemikalien, Mykotoxine und Medikamente spielen bei der
Entstehung von Tumoren der Haussäugetiere entgegen den Erkenntnissen der
Humanmedizin eine eher untergeordnete Rolle. Tumoren der Leber und der
Gallengänge stehen, soweit bekannt, in der Veterinärmedizin nicht in direktem
Zusammenhang mit vorangegangenen Lebererkrankungen (STALKER u. HAYES
2007).
Tabelle 2-2 Übersicht der Lebertumoren (CULLEN 2007a)
Die epithelialen Tumoren der Leber gehen entweder von Hepatozyten, von
Gallengangsepithelien oder deren gemeinsamen undifferenzierten Vorläuferzellen
(oval cells) hervor. Hepatozelluläre Adenome kommen vor allem bei älteren Hunden
sowohl in gesundem Gewebe als auch in zirrhotisch umgebautem Parenchym vor
(KÄUFER-WEISS 2007). Sie unterscheiden sich makroskopisch kaum von nodulären
Hyperplasien oder von Leberzellkarzinomen und treten normalerweise als einzelne
Tumoren auf (STALKER u. HAYES 2007).
Hepatozelluläre Karzinome kommen selten aber mit einer gewissen Regelmäßigkeit
bei Hunden vor (CULLEN 2007a). Sie treten als einzelne oder multinoduläre
Umfangsvermehrungen auf (STALKER u. HAYES 2007), zeigen ein infiltratives
Wachstum und neigen nach Einbruch in das Pfortadersystem zur weiteren
Ausbreitung in der Leber (KÄUFER-WEISS 2007). Hämatogene Metastasen über die
Lebertumoren epithelial mesenchymal
Neuroendokriner
Ursprung
Sonstige
Benigne
Hepatozelluläres
Adenom
Adenom der
Gallenwege
Hämangiom
Maligne
Hepatozelluläres
Karzinom
Gallengangskarzinom
Hepatocholangio-
karzinom
Hepatoblastom
Hämangiosarkom
Hepatisches
Karzinoid
Lymphom
Myeloide Neoplasie
Mastzelltumor

2 Literaturübersicht 53
Vena hepatica finden sich meist zuerst in der Lunge (HEAD et al. 2003; STALKER u.
HAYES 2007). Histologisch werden trabekuläre, adenoide und solide Formen der
Karzinome unterschieden (STALKER u. HAYES 2007), wobei mehr als eine
histologische Variante in einem Tumor auftreten kann (HEAD et al. 2003). Mitosen
kommen häufig vor (STALKER u. HAYES 2007). Die zellulären Eigenschaften der
Hepatozyten in hepatozellulären Karzinomen variieren je nach Differenzierungsgrad.
Gut differenzierte hepatozelluläre Karzinome zeigen Leberzellen, die normalen
Hepatozyten gleichen, mit rundem zentralen Kern und eosinophilem Zytoplasma.
Das Zytoplasma kann auch blass sein, vakuolisiert und gefüllt mit Glykogen oder
Lipiden. Schlecht differenzierte Karzinome weisen dagegen Hepatozyten auf, die
sehr pleomorph sind. Diese Tumorzellen zeigen eine Anisokaryose und nur wenig
basophiles Zytoplasma. Die Nukleoli sind deutlich vergrößert. Eine Sonderform des
hepatozellulären Karzinoms stellt das Klarzellkarzinom dar, das vollständig aus
ballonierten Zellen zusammengesetzt ist (HEAD et al. 2003).
Bei den Tumoren, die vom Gallengangsepithel ausgehen, werden das Cholangiom
und das Cholangiokarzinom unterschieden. Das Cholangiom findet sich vor allem in
Lebern älterer Hunde. Cholangiome treten intrahepatisch auf, selten extrahepatisch
und erscheinen als solide oder zystische Tumore (STALKER u. HAYES 2007). Das
Wachstumsverhalten der Gallengangskarzinome ist solide oder multinodulär
(STALKER u. HAYES 2007) und besitzt häufig ein ausgeprägtes fibröses Stroma
(CULLEN 2007a). Hämatogene Metastasen sind selten, Metastasen in regionale
Lymphknoten dagegen häufig. Das Wachstumsverhalten ist sehr invasiv. Die
Abgrenzung zu anderen metastatischen Adenokarzinomen ist mitunter sehr
schwierig. (STALKER u. HAYES 2007).
Das Hepatoblastom ist ein Tumor ausgehend von pluripotenten Vorläuferzellen,
welcher sowohl bei alten als auch bei jungen Hunden vorkommt (STALKER u.
HAYES 2007).
Tumoren, die aus neuroendokrinen Zellen der Leber oder der Gallengänge
hervorgehen, nennt man hepatische Karzinoide. Diese sind gekennzeichnet durch
ein besonders aggressives Wachstumsverhalten in Form von stark infiltrativem

54 2 Literaturübersicht
Wachstum und Metastasenbildung in lokale Lymphknoten und das Peritoneum.
Typischerweise sind die Tumorzellen nesterartig arrangiert, durchzogen von
Bindegewebssepten (HEAD et al. 2003; STALKER u. HAYES 2007). Hepatische
Karzinoide müssen von metastatischen Tumoren des Nebennierenmarkes
abgegrenzt werden (HEAD et al. 2003).
Primäre mesenchymale Tumoren der Leber können von Bindegewebe der glatten
Muskulatur oder den Endothelzellen ausgehen. Sowohl gutartige Leiomyome als
auch maligne Leimyosarkome und Fibrosarkome treten auf. Hämangiome und
Hämangiosarkome finden sich ebenfalls häufig in der Leber, wobei
Hämangiosarkome dort häufiger als Metastasen vorkommen (STALKER u. HAYES
2007).
Sonstige Neoplasien
Die Leber ist aufgrund ihrer starken Vaskularisation ein bevorzugtes Organ für die
Ansiedlung metastatischer Tumoren (CHEVILLE 1994d; STALKER u. HAYES 2007).
Metastatische Neoplasien sind bei Hunden die am häufigsten vorkommenden
Lebertumoren und entstammen zumeist dem Pankreas, dem Gastrointestinaltrakt,
der Milz, der Milchleiste, den Nebennieren, Knochen und der Schilddrüse (HOSKINS
2005). Karzinome des Pankreas und Gastrointestinaltraktes metastasieren über die
Portalvene in die Leber. Einige Tumore, wie Mammakarzinome, Schildrüsen-
karzinome, Melanome und Sarkome, metastasieren über die Lunge und die
Leberarterie (STALKER u. HAYES 2007). Die periportalen und perisinusoidalen
Bereiche sind sehr empfänglich für hämatopoetische neoplastische Zellen. Aufgrund
dieser Tatsache sind Lymphome, maligne Histiozytose, myeloide und erythroide
Leukämien sowie Mastzelltumoren in der Leber nicht selten (STALKER u. HAYES
2007). Das multizentrische maligne Lymphom ist ein häufiger Tumor der Leber, auch
unter Einbeziehung von Milz (HEAD et al. 2003; STALKER u. HAYES 2007),
Lymphknoten, Thymus, Knochenmark und anderen Organen (KÄUFER-WEISS
2007; STALKER u. HAYES 2007).

2 Literaturübersicht 55
Wie bereits erwähnt sind Hämangiosarkome der Leber häufig als Metastasen
anzutreffen. Der Tumor besteht aus Endothelzellen, die unterschiedlich große
vaskuläre Spalten und Kavernen ausformen. Auch solide Formen treten auf
(STALKER u. HAYES 2007), wobei die neoplastischen Endothelzellen anhand ihrer
„schuhzweckenförmigen“ Gestalt erkannt werden können (KÄUFER-WEISS 2007).
Die bevorzugten Lokalisationen der Primärtumoren sind Milz, Knochenmark und
Leber. Die Metastasierung erfolgt in besonderem Maße in die Lunge und das rechte
Herzohr. Neben der Bösartigkeit dieser Tumore ist, im Gegensatz zu den
Hämangiomen, ihre Tendenz zur spontanen Ruptur auffällig. Häufiger als andere
Rassen erkranken der Deutsche Schäferhund und der Boxer am Hämangiosarkom
(KÄUFER-WEISS 2007).
2.3.4.1 Verhalten der Peroxisomen und Mitochondrien bei
Tumoren
Ultrastrukturelle Untersuchungen von hepatozellulären Tumoren zeigen allgemein
eine große Variabilität der Tumorzellen in der Frühphase der Karzinogenese.
Während der Entstehung eines hepatozellulären Karzinoms zeigen Tumorzellen eine
strukturelle Vereinfachung des Zytoplasmas, der Organellen (simplification).
Vereinfachungen der Organellen treten besonders beim rER und den Mitochondrien
auf. Ein Verlust von Organellen betrifft vor allem das sER, die Peroxisomen und das
Substrat Glykogen.
Bei schnell wachsenden Hepatomen konnte beispielsweise eine Abnahme der
Peroxisomenzahl und deren Enzymausstattung beobachtet werden (MOCHIZUKI et
al. 1971). Darüber hinaus findet während des Tumorwachstums ein Wechsel von
peroxidativem zu mitochondrialem Metabolismus statt, um Energie für Zellwachstum
und Mitose bereitzustellen (TOLBERT u. ESSNER 1981). Bei hepatozellulären
Karzinomen sowie bei Lebermetastasen anderer Tumore konnten Veränderungen
des peroxisomalen Erscheinungsbildes in Form von Invaginationen, Protrusionen
und Membranschleifen (Gastruloid cisternae) beobachtet werden (DE CRAEMER et
al. 1993). Diese Veränderungen sind jedoch nicht spezifisch für Tumorerkrankungen,

56 2 Literaturübersicht
da sie ebenfalls bei Hepatitis und Steatose vorkommen können (DE CRAEMER et al.
1991; DE CRAEMER et al. 1995).
Im Gegensatz zu den zahlreichen Informationen in der Humanmedizin liegen in der
Veterinärmedizin keine Erkenntnisse in Bezug auf das Verhalten der Peroxisomen
und der Mitochondrien bei tumorösen Prozessen, insbesondere des Hundes vor. Ziel
dieser Arbeit ist es, Informationen hinsichtlich des Verhaltens der caninen
Peroxisomen und auch in untergeordneter Stellung, der Mitochondrien, in tumorösen
und degenerativen Erkrankungen der Leber zu erarbeiten.

3 Eigene Untersuchungen 57
3 Eigene Untersuchungen
3.1 Material und Methoden
3.1.1 Einleitung
In der vorliegenden klinisch-prospektiven Studie erfolgte eine qualitative und
quantitative elektronenmikroskopische Untersuchung der hepatozellulären
Peroxisomen von Hunden, die an degenerativen Erkrankungen oder tumorösen
Erkrankungen der Leber litten. Als Kontrollgruppe dienten Hunde ohne
Leberveränderungen.
Alle Gruppen wurden hinsichtlich der Anzahl an Peroxisomen pro Fläche/Hepatozyt
(µm²) miteinander verglichen. Neben der Untersuchung der Peroxisomen hinsichtlich
Morphologie und Anzahl wurden außerdem die übrigen Zellorganellen der
Hepatozyten (rER, sER, Mitochondrien, Lysosomen, Glykogen) auf morphologische
Veränderungen untersucht und mit denen von lebergesunden Hunden verglichen.
Die Patienten wurden einer Anamnese, einer Allgemeinuntersuchung und einer
Ultraschalluntersuchung unterzogen. Ergänzende Untersuchungen, wie die
Bestimmung klinisch-chemischer und hämatologischer Parameter, wurden
standardmäßig eingeleitet. Vor jeder Biopsieentnahme wurden
Gerinnungsparameter, wie aktivierte partielle Thromboplastinzeit und
Prothrombinzeit bestimmt. Zur weiteren Abklärung der Hepatopathie wurde
transabdominal oder laparaskopisch eine Biopsie des pathologisch veränderten
Lebergewebes entnommen und einer histologischen Untersuchung unterzogen.
3.1.2 Patienten
Im Zeitraum Oktober 2006 - Juni 2009 wurden insgesamt 45 Patienten mit
Hepatopathien in die Studie aufgenommen. Davon konnten jedoch insgesamt 26
Patienten aufgrund unterschiedlicher Ausschlusskriterien (Tabelle 3-1) nicht

58 3 Eigene Untersuchungen
verwendet werden. Überwiegend handelte es sich dabei um die Tatsache, dass der
pathohistologische Befund am Paraffin eingebetteten Gewebe nicht in
Übereinstimmung mit der für ultrastrukturelle Untersuchungen in Epon eingebetteten
sehr kleinen Gewebeproben stand, so dass schließlich die Proben von 19 Patienten
für die vorliegende Studie zur Verfügung standen und untersucht wurden.
Auswahlkriterien/Verteilung
Patienten mit degenerativen Leberveränderungen wurden auf der Basis veränderter
klinisch-chemischer Blutparameter und der histologischen Diagnose der
Leberbiopsie in die Studie aufgenommen. Auch die Patienten mit tumorösen
Lebererkrankungen wurden anhand histologischer Befunde der Leberbiopsie und
gegebenfalls veränderter Blutwerte in die Studie integriert.
Tabelle 3-1 Anzahl der Patienten mit Hepatopathien, die nicht mit in die vorliegende Studie einbezogen werden konnten
Letztlich wurden 19 Patienten in die vorliegende Studie aufgenommen, von denen 11
in die Gruppe der degenerativen Lebererkrankungen und 8 Patienten in die Gruppe
der neoplastischen Lebererkrankungen fielen.
Ursachen
Anzahl der Patienten
keine histologischen Krankheitsanzeichen 3
Lokalisation für ultrastrukturelle Untersuchung nicht repräsentativ 23

3 Eigene Untersuchungen 59
Gruppe 1
Diese Patientengruppe umfasst 11 Hunde, die an einer degenerativen
Lebererkrankung litten. In Tabelle 3-2 findet sich eine Übersicht der Patienten.
Tabelle 3-2 Übersicht Patienten der Gruppe 1 mit degenerativen Lebererkrankungen (w=weiblich, wk=weiblich kastriert, m=männlich, M.=Morbus, WHWT=Westhighland White Terrier, JRT=Jack Russel Terrier, RHT=Rauhaarteckel)
K-Nr. Rasse Alter in Jahren
Geschlecht Gewichtsklasse Grunderkrankung Vorbehandlung
2029 Fox Terrier 12 w 10-20 kg Hämangiosarkom
(Milz), hgr.Splenitis -
1972
Welsh Terrier 10
w 0-10 kg
Hypothyreose, IBD, Futtermittelallergie,
Mammamisch-tumor, Anteile
eines Karzinoms
Kortison Trimethoprim Sulfonamide
1979
Mischling 13
wk 20-40 kg
Mitralinsuffizienz ohne
hämodynamische Bedeutung, M.
Cushing
Trilostanum
2015 Mischling 11 m 20-40 kg V. a. Pankreatitis -
2017 Cocker Spaniel
9 m 10-20 kg
Anaplastisches Sarkom (Milz) -
1963 Pudel
6
m 10-20 kg
Kryptorchide, Mitralis-
degeneration mit leichter Insuffizienz
Pimobendan
2062 RHT 12 m 0- 10 kg Hypothyreose
-
2063 RHT 12
w 0- 10 kg maligner
Granulosazelltumor (Ovar)
-
1969 WHWT 11 m 0- 10 kg V. a. Lungenfibrose Benazepril
1974 Golden R. 15 w 20-40 kg Leiomyom Benazepril
Carprofen
2025 JRT 11
w 0- 10 kg Nierentumor mit Einbruch in die V.
cava -
Gruppe 2
Die zweite Gruppe der Patienten umfasste 8 Patienten, die eine neoplastische
Veränderung in der Leber aufwiesen. Dabei handelte es sich entweder um einen
Primärtumor der Leber oder um eine sekundäre Lebermetastase. In Tabelle 3-3 ist
eine Übersicht der Patienten mit histopathologischer Tumordiagnose der
Grunderkrankung, sowie der Auflistung verabreichter Medikamente zu sehen.

60 3 Eigene Untersuchungen
Tabelle 3-3 Übersicht Patienten der Gruppe die an einem Primär- oder Sekundärtumor erkrankten (w=weiblich, m=männlich, JRT=Jack Russel Terrier, RHT=Rauhaarteckel)
K-Nr.
Rasse Alter
(Jahre) Ge-
schlecht Gewichts-
klasse
Histo-pathologische
Diagnose (Tumor)
Grunderkrankung/ Verdachtsdiagnose
Vorbehandlung
Seku
nd
ärtu
mo
r
1965 Rottweiler 11 m 20-40 kg Hämangio-
sarkom Hämangiosarkom der
Milz -
2006 Mischling 7 m 20-40 kg Hämangio-
sarkom
Hämangiosarkom der Milz
Pimobendan Benazepril
Spironolacton
2009 Hovawart 7 m 20-40 kg Hämangio-
sarkom
Gekrösetumor in Verbindung zu
Lebertumor L-Thyroxin
2030 Staffordshire
Terrier 10 w 20-40 kg anaplastisches
malignes Lymphom
multizentrisches Lymphom
2027 RHT 13 w 0- 10 kg Metastase eines Plattenepithel-
karzinoms -
Benazepril Propentophyllin
Furosemid Diazepam
Theophyllin
2021
JRT 12 m
0- 10 kg
Metastase eines
Phäochromo- blastoms
Phäochromoblastom der
Nebenniere Ramipiril
Pri
mär
-
tum
or 1968 Mischling 12 m 10-20 kg
Gallengangsadenom, hepatozelluläres
Karzinom - -
2011 Mischling 9 m
10-20 kg Klarzellkarzinom - -
Gruppe 3
Außerdem wurden als Kontrolltiere zwei Hunde der Rasse Beagle herangezogen, die
zuvor im Rahmen einer pharmakologischen Studie bereits als Kontrolltiere dienten.
Die Tiere waren 365 Tage alt und wurden mit einem Standardfutter gefüttert. Zur
Probenentnahme wurden die Tiere über eine überdosierte intravenöse Gabe von
Pentobarbital (0,5 ml/kg KGW Release®) eingeschläfert. Im Anschluss folgte die
Probenentnahme mittels Biopsie. Dabei wurden bei jedem Tier an drei zuverlässigen
Lokalisationen der Leber Bioptate entnommen, sodass insgesamt sechs Proben als
Kontrollmaterial zur Verfügung standen und elektronenmikroskopisch untersucht
werden konnten.

3 Eigene Untersuchungen 61
Tabelle 3-4 Kontrolltiere (m=männlich, d=Tage)
K-Nr.
Rasse
Alter (d)
Geschlecht
Gewicht
2103 Beagle 365 m
10-20 kg
(mittelgroße Rasse)
2104 Beagle 365 m
10-20 kg
(mittelgroße Rasse)
Laboruntersuchungen
Die im Labor der Kleintierklinik des Tierärztlichen Institutes der Georg-August-
Universität Göttingen durchgeführten Blutuntersuchungen erfolgten mit dem
Hämatologiegerät „Abbott Cell-Dyn® 3500“ (Fa. Abbott GmbH & Co. KG,
Wiesbaden). Die Messung der klinisch-chemischen Parameter erfolgte anfangs
mittels des Gerätes „RX Daytona“ (Fa. Randox Laboratories Ltd., United Kingdom),
ab März 2007 stand zur täglichen Routinediagnostik das Analysegerät „Konelab 20i“
(Fa. Thermo Fisher Scientific Inc., Dreieich) zur Verfügung. Die Untersuchung der
Gerinnungsparameter erfolgte mit dem Gerät „SCA 2000 Veterinary Coagulation
Analyzer“ (Fa. Synbiotics, San Diego, California). Die Ergebnisse der durchgeführten
Blutuntersuchungen finden sich in tabellarischer Form im Anhang (Anhangstabelle
10-1 bis Anhangstabelle 10-8).
Gewinnung der Leberprobe
Die Leberbiopsie wurde am sedierten Patienten transabdominal unter
Ultraschallkontrolle durchgeführt. In einigen Fällen machte der sonographische
Befund der Patienten eine sofortige Probelaparotomie in Allgemeinanästhesie
notwendig, sodass bei diesen Tieren unter Sichtkontrolle ein Bioptat entnommen
werden konnte. In beiden Fällen war eine 12-stündige Nahrungskarenz notwendig.
Notfallsituationen bildeten eine Ausnahme.

62 3 Eigene Untersuchungen
Alle Patienten erhielten einen intravenösen Zugang über die Vena cephalica
antebrachii oder Vena saphena lateralis mit einem sterilen Venenkatheter
„Vasocan® “ der Größe: 1,1 x 33 mm der 0,9 x 25 mm (Fa. Braun Melsungen AG,
Melsungen).
Die Sedation zur transabdominalen Biopsieentnahme wurde über die intravenöse
Gabe von Propofol (Narcofol®, Fa. Cp-pharma, Burgdorf) erreicht. Die intravenöse
Gabe erfolgte nach individueller Wirkung. Die maximale Dosis an Propofol belief sich
auf bis zu 6 mg/kg KGW.
Zur intravenösen Einleitung der Allgemeinanästhesie für die Durchführung der
Probelaparatomie wurden folgende Narkotika in Kombination angewendet:
Levomethadon (1 mg/kg KGW L-Polamivet®, Fa. Intervet, Unterschleißheim)
mit Acepromacin (0,4 mg/kg KGW Vetranquil®, Fa. Albrecht, Aulendorf)
Levomethadon (1 mg/kg KGW L-Polamivet®, Fa. Intervet, Unterschleißheim)
und Diazepam (maximal 0,5 mg/kg KGW Diazepam®, Roche Holding GmbH,
Grenzach-Whyhlen) getrennt voneinander verabreicht.
Das Inhalationsnarkosegerät „Sulla 808 V“ der Fa. Dräger, Lübeck, mit Sauerstoff-
Isofluran Gemisch, stand zur Aufrechterhaltung der Injektionsnarkose zur Verfügung.
Die Überprüfung der Vitalparameter erfolgte mit dem Pulsoximeter (Fa. Smiths
Medical PM, Inc. Veterinary Division, Norwell, USA).
Die Leberbiopsie wurde mittels einer sterilen Vet Core Biopsy needle, Surgi VetTM der
Größe: 20 mm, 14 GA, 15 cm, (Fa. Smiths Medical PM, Inc. Veterinary Division,
Norwell, USA) genommen. Die im Rahmen der Biopsie entnommenen
Gewebeproben wurden geteilt. Der Biopsieanteil für die histologische Untersuchung
wurde direkt nach der Entnahme in 10 %igem neutral gepuffertem Formaldehyd für
mindestens 24 Stunden fixiert. Der Anteil der Leberbiopsie für die TEM-
Untersuchung wurde in 2,5 %igem Glutaraldehyd fixiert und unverzüglich in 1mm x
1mm große Blöcke geschnitten.

3 Eigene Untersuchungen 63
3.1.3 Lichtmikroskopische Untersuchung
Präparation
Die Leberbioptate wurden direkt nach der Entnahme in 10 %igem neutral
gepuffertem Formaldehyd für mindestens 24 Stunden fixiert. Im Anschluss wurden
die Gewebeproben zugeschnitten und in Einbettkassetten gelegt. Die
Paraffineinbettung erfolgte automatisch durch das Gerät „Excelsior ES“ (Fa. Thermo
Scientific, Dreieich) nach einem laborüblichen Standardprotokoll (siehe Anhang). Von
den Paraffinblöcken wurden mit dem Schlittenmikrotom (Mikrotom HM 400R, Fa.
Thermo Fisher Scientific, Dreieich) 3-4 µm dicke Schnitte angefertigt. Zum
Abnehmen der Schnitte vom Paraffinblock diente ein im Eisbad angefeuchteter
Durchschlagpapierstreifen. Zum Strecken der Schnitte wurden diese in ein 40°C
warmes Wasserbad überführt. Die Paraffinschnitte wurden dann auf Objektträger
aufgezogen und bei 37°C im Wärmeschrank über Nacht getrocknet. Bis zur weiteren
Verwendung wurden sie bei Raumtemperatur aufbewahrt. Für die histologische
Untersuchung wurden die Paraffinschnitte mit der Standard-Färbung Hämatoxilin-
Eosin (H.&E.) gefärbt. Weitere Spezialfärbungen wurden durchgeführt, wie die
Berliner Blau Reaktion zum Nachweis von Eisen in Hepatozyten und die PAS
Reaktion zum Nachweis von Glykogen in Hepatozyten. Die Protokolle zu den
einzelnen Färbungen sind im Anhang aufgeführt.
Histologische Kriterien zur Diagnosestellung
Die histologische Untersuchung der Leberproben erfolgte in Zusammenarbeit mit
anerkannten Fachtierärzten für Pathologie der Tiermedizinischen Praxis für
Histopathologie Göttingen.
3.1.4 Präparation für die Transmissionselektronenmikroskopie
Für die ultrastrukturelle Untersuchung der hepatozellulären Peroxisomen wurden die
Leberproben in 2,5 %igem Glutaraldehyd fixiert und anschließend in Epon
eingebettet. Die Einbettung erfolgte in einem Epongemisch nach (LUFT 1961) (siehe
Anhang) im Lynx-Einbettautomaten (Fa. Leica, Bensheim, Deutschland). Die genaue

64 3 Eigene Untersuchungen
Anleitung zum Einbetten findet sich ebenfalls im Anhang. Die Bioptate wurden in
bereits mit vorpolymerisiertem Epon zu einem Drittel gefüllten Einbettungsformen
aus Silikongummi seitlich positioniert, so dass beim späteren Anschneiden der
Gewebeblöcke ein günstiger Anschnitt getroffen werden konnte. Die Blockformen
wurden mit Epon aufgefüllt. Es schloss sich eine 24-stündige Polymerisation des
Epoxidharzes bei 60°C im Wärmeschrank an. Die auspolymerisierten Epon-Blöcke
wurden im Anschluss mit einer Fräse (Reichert Ultratrim, Fa.Leica, Bensheim)
getrimmt. Ziel war es, die Oberfläche des Gewebeblockes so zu formen, dass das
Gewebe nur noch von einem schmalen Eponsaum umgeben war. Im weiteren
Verlauf wurden mit Hilfe des Ultramikrotoms (Reichert, Ultracut S, Fa. Leica,
Bensheim) unter Verwendung von Histo-Diamantmessern (Fa. Diatome; Bienne,
Schweiz) 0,5 µm dicke Semidünnschnitte mit einer Kantenlänge von 1-2 mm erstellt.
Die durch den Schneidevorgang anfallenden Schnitte sammelten sich auf dem
Flüssigkeitstrog und wurden auf einen Objektträger mit einem Tropfen Aqua dest.
mittels eines Glasstäbchens übertragen. Anschließend wurden die Schnitte auf einer
Wärmebank bei 60°C für mindestens eine Stunde getrocknet. Die Semidünnschnitte
wurden einer Methylenblau/Azur III Färbung nach RICHARDSON (ausführliches
Protokoll: siehe Anhang) unterzogen und mit Eukitt (Fa. Kindler, Freiburg)
eingedeckt.
Die hergestellten Semidünnschnitte dienten der lichtmikroskopischen Vororientierung
und morphologischen Beurteilung sowie der gezielten Erstellung von
Ultradünnschnitten für die elektronenmikroskopische Untersuchung.
Für die Herstellung der Ultradünnschnitte wurde der lichtmikroskopisch interessante
Bereich des Semidünnschnittes mit den relevanten und repräsentativen
pathologischen Veränderungen in einer Skizze dargestellt, sodass von dieser Region
anschließend der Ultradünnschnitt erstellt werden konnte. Für die Anfertigung der
Ultradünnschnitte wurde der Bereich des Semidünnschnittes ausgewählt, der die
histologisch diagnostizierte Lebererkrankung am besten repräsentierte und die
Hepatozyten im Anschnitt mit Nukleus auch morphologisch ein gut erhaltenes
Erscheinungsbild zeigten. Bei Patienten mit einem Lebertumor wurden die zu
untersuchenden Hepatozyten aus dem an den Tumor angrenzenden Lebergewebe

3 Eigene Untersuchungen 65
entnommen, da nur dort die Zelle in einem Zustand war, der die morphologische
Beurteilung ermöglichte. Die Auswahl der zu untersuchenden Hepatozyten erfolgte
unabhängig von ihrer periportalen, mittzonalen oder zentralen Ausrichtung im
Leberläppchen. Mit Hilfe eines Diamantmessers (Firma Diatome; Bienne, Schweiz)
wurden die 60-80 nm dünnen Schnitte in Form von Schnittketten mit einer
Kantenlänge von 0,1- 0,3 mm erstellt. Die so entstandenen Ultradünnschnitte wurden
auf Kupferobjektträgernetze, sogenannte Grids (single slot 1 x 2, Fa. Plano, Marburg)
aufgezogen. Die Kontrastierung der Schnitte wurde per Hand mit Uranylacetat und
Bleicitrat (REYNOLDS 1963; STEMPAK u. WARD 1964) durchgeführt. Die
Kontrastierungsprotokolle finden sich im Anhang.
3.1.5 Auswertung und Dokumentation
Die lichtmikroskopische Untersuchung (Mikroskop: Axioplan 2-Imaging; Fa. Zeiss,
Göttingen) erfolgte in Zusammenarbeit mit der tierärztlichen Praxis für
Histopathologie Göttingen. Die fotografische Dokumentation und Bearbeitung der
Bilder erfolgte mit der Digitalkamera Axio Cam (Fa. Carl Zeiss, Göttingen) sowie dem
Computerprogramm Axio Vision (Fa. Carl Zeiss, Göttingen).
Für die elektronenmikroskopische Untersuchung stand das ZEISS-
Elektronenmikroskop (EM 10 C, Fa. Zeiss, Oberkochen) sowie das
Bildanalyseprogramm Analysis (Soft Imaging System, Münster) zur Verfügung.
Bei insgesamt 21 Patienten wurde elektronenmikroskopisch die Anzahl der
Peroxisomen in Bezug zur Fläche (µm²) von 6 Hepatozyten bestimmt. Die Auswahl
der Hepatozyten erfolgte in erster Linie nach deren Erscheinungsbild und
Erhaltungszustand, wobei insbesondere die Morphologie der Peroxisomen gut
erhalten sein sollte.
Darüber hinaus wurden die Hepatozyten der Tiere aus Gruppe 1 so ausgewählt,
dass alle eine vergleichbare Menge von Fetttropfen enthielten. Dabei wurde die
Fläche der Fetttropfen nicht von der Gesamtfläche der Hepatozyten abgezogen (DE
CRAEMER et al. 1995).

66 3 Eigene Untersuchungen
Bei der Auswahl der Hepatozyten von Tieren der Gruppe 2 wurden nur gut erhaltene
Hepatozyten aus dem Randbereich der Neoplasie ausgewählt. Von der Auswahl von
Hepatozyten direkt aus dem Zentrum eines Tumors wurde abgesehen, da eine
angemessene Beurteilung der Organellen dieser Zellen aufgrund von nekrotischen
Veränderungen in Folge des tumorösen Geschehens nicht möglich war.
Bei 2500 facher Vergrößerung wurde der Ultradünnschnitt nach gut erhaltenen
Hepatozyten mit Nukleus durchsucht und der ausgewählte Hepatozyt fotografiert
(siehe Abbildung 3-1). Zur Identifikation und Auszählung der Peroxisomen wurde der
Hepatozyt anschließend bei 6300 facher Vergrößerung fotografiert. In dieser Ver-
größerung waren die Peroxisomen gut erkennbar. Die Einzelaufnahmen wurden
schließlich zu einem Bild zusammengefügt. Eine digitale Erfassung von sehr großen
Hepatozyten, erfolgte mit der Multiple Image Alignment (MIA)-Funktion des
Bildanalyseprogrammes. Die Fläche (µm²) eines Hepatozyten wurde ebenfalls mit
Hilfe der Bildanalysesoftware morphometrisch am Computerbildschirm bei 2500
facher Vergrößerung ermittelt. Als Bezugsgröße für die quantitative Bestimmung der
Peroxisomen diente die gesamte zytoplasmatische Fläche des jeweiligen
Hepatozyten (RYOO u. BUSCHMANN 1989).

3 Eigene Untersuchungen 67
Fläche Hepatozyt 173,17 µm²
Abbildung 3-1 Übersicht eines Hepatozyten mit Bestimmung der Fläche in µm², (K2011, Ultradünnschnitt, MIA, Transmissionselektronenmikroskopie Gerätevergr. 2500x)
Auf den ausgedruckten Übersichtsdarstellungen der Hepatozyten wurden die
einzelnen Peroxisomen/Fläche µm² des Hepatozyten ausgezählt. Die Identität der
Peroxisomen musste teilweise in höheren Vergrößerungen auf dem
Computerbildschirm überprüft werden.
Neben der quantitativen Auswertung wurde auch das morphologische
Erscheinungsbild der Peroxisomen sowie das der übrigen Zellorganellen untersucht
und qualitativ erfasst.
Die gesamte elektronenmikroskopische Auswertung erfolgte anhand eines
selbsterstellten Befundbogens (Anhangstabelle 10-9).

68 3 Eigene Untersuchungen
3.1.6 Statistik
Die statistische Auswertung erfolgte mit dem Programm Statistica (Fa. StatSoft,
Tulsa, Oklahoma, USA).
Die statistische Auswertung der Peroxisomen/Fläche µm² sowie die Gesamt-
betrachtung der drei Gruppen hinsichtlich Alter und Rasse erfolgte unter Verwendung
eines einfaktoriellen ANOVA Testes. Ein Signifikanzwert von p <0.05 ( 5 %) wurde
der Untersuchung zu Grunde gelegt. Die vergleichende Betrachtung der Gruppen
untereinander erfolgte unter Verwendung des t-Tests. Eine Signifikanz lag vor, wenn
p <0,017 war (Bonforoni Anpassung).
Zu den Untersuchungskriterien gehörten: Peroxisomen (Px) klein, Peroxisomen (Px)
groß, Erhaltungszustand, Verteilung der Peroxisomen (Px) in der Zelle, Protrusion,
Marginalplatte, Nukleoid, Mitochondrien, Schwellung der Mitochondrien,
parakristalline Einschlüsse der Mitochondrien, Riesenmitochondrien, Lipidtropfen,
Myelinfiguren, Glykogen. Die Auswertung der Untersuchungskriterien erfolgte bei
globaler Betrachtung mit dem Kruskal- Wallis Test (p <0,05). Bei Betrachtung der
Einzelgruppen untereinander wurde der Mann-Withney U Test verwendet (p <0,017).
3.2 Ergebnisse
Ziel der vorliegenden Arbeit ist die elektronenmikroskopische, quantitative und
qualitative Untersuchung hepatozellulärer Peroxisomen von Hunden, die entweder
neoplastische oder degenerative Leberveränderungen aufweisen, im Vergleich zu
einer gesunden Kontrollgruppe.
Zunächst werden die Patientengruppen, die in die vorliegende Untersuchung
einbezogen wurden, hinsichtlich der Alters-, Rasse- und Gewichtsklassenverteilung
beschrieben.
Im Anschluss werden die wichtigsten klinisch-chemischen Laborergebnisse
vorgestellt. Schließlich erfolgt eine vergleichende Darstellung der licht- und
elektronenmikroskopischen Untersuchungsergebnisse der einzelnen Gruppen sowie

3 Eigene Untersuchungen 69
in Abhängigkeit vom Alter und den Gewichtsklassen der Hunde mit entsprechenden
statistischen Auswertverfahren.
3.2.1 Zusammensetzung der Patientengruppen
Rasseverteilung
Die Gruppe der Hunde mit degenerativen Leberveränderungen (Gruppe 1) besteht,
wie in Tabelle 3-5 ersichtlich, aus zwei Mischlingen, zwei Rauhaarteckeln, und je
einem Fox Terrier, Welsh Terrier, Cocker Spaniel, Pudel, Westhighland White
Terrier, Golden Retriever und Jack Russel Terrier.
Die Gruppe der Hunde mit neoplastischen Leberveränderungen (Gruppe 2) setzt sich
aus drei Mischlingen und jeweils einem Hund der Rasse Rottweiler, Hovawart,
Staffordshire Terrier, Rauhaarteckel und Jack Russel Terrier zusammen.
Die Gruppe der gesunden Kontrolltiere (Gruppe 3) besteht aus zwei Beagle mit
einem Alter von 365 Tagen.

70 3 Eigene Untersuchungen
Tabelle 3-5 Rasseverteilung der Gruppen 1-3
Altersverteilung
Gruppe 1 besteht aus vier Patienten im Alter zwischen sechs und zehn Jahren
(36 %) und sieben Patienten im Alter von elf bis 15 Jahren (64 %). Keiner der Hunde
aus Gruppe 1 war jünger als sechs Jahre.
Gruppe 2 bestand zur Hälfte aus Hunden zwischen sechs und zehn Jahren und zur
anderen Hälfte aus Hunden im Alter von elf bis 15 Jahren.
Zwei gesunde Kontrolltiere im Alter von 365 Tagen bildeten die Gruppe 3.
Eine vergleichende Darstellung der Altersverteilung findet sich in Tabelle 3-6.
Rasse
Anteil an Gesamtpatientenzahl
Absolute Häufigkeit (n)
Gruppe 1
(Degeneration)
Gruppe 2
(Tumor)
Gruppe 3
(Kontrolle)
Mischling 5 (23 %) 2 3 -
Rauhaarteckel 3 (14 %) 2 1 -
Jack Russel Terrier 2 ( 9 %) 1 1 -
Fox Terrier 1 ( 5 %) 1 - -
Welsh Terrier 1 ( 5 %) 1 - -
Cocker Spaniel 1 ( 5 %) 1 - -
Pudel 1 ( 5 %) 1 - -
Westhighland White Terrier
1 (5 %) 1 - -
Golden Retriever 1 (5 %) 1 - -
Rottweiler 1 (5 %) - 1 -
Hovawart 1 (5 %) - 1 -
Staffordshire Terrier 1 (5 %) - 1 -
Beagle 2 (9 %) - - 2

3 Eigene Untersuchungen 71
Tabelle 3-6 Altersverteilung: absolute und relative Häufigkeit der Patienten der Gruppen 1-3
Gewichtsverteilung
Die Hunde der Gruppe 1 verteilen sich anhand ihres Körpergewichtes in folgende
Gewichtsklassen. Fünf Hunde kleiner Rassen mit einem Körpergewicht (KGW) bis zu
10 kg, drei Hunde mittelgroßer Rasse mit einem KGW von 10-20 kg sowie drei
Hunden großer Rassen mit einem KGW von 20-40 kg.
Tabelle 3-7 Gewichtsverteilung der Patienten der Gruppen 1-3
Gewichtsklasse Anteil an
Gesamtpatientenzahl
Absolute Häufigkeit (n)
Gruppe 1
(Degeneration)
Gruppe 2
(Tumor)
Gruppe 3
(Kontrolle)
kleine Rasse: 0-10 kg 7 (33 %) 5 2 0
mittelgroße Rasse: 10-20 kg 7 (33 %) 3 2 2
große Rasse:
20-40 kg 7 (33 %) 3 4 0
Die Gruppe 2 besteht aus jeweils zwei Hunden (25 %), die kleinen bzw. mittelgroßen
Rassen angehören. Vier Hunde (50 %) gehören großen Rassen an.
Zwei Hunde mittelgroßer Rasse (Beagle) bilden die Gruppe 3 (Kontrolle).
Geschlechtsverteilung
Sechs (54,5 %) von elf Patienten der Gruppe 1 sind weiblichen Geschlechts. Eine
dieser Hündinnen (K 1979) ist kastriert. Diese Hündin wird aufgrund der geringen
Anzahl bei der statistischen Auswertung den weiblichen Tieren zugeordnet. Die
übrigen 5 (45,5 %) Hunde sind männlichen Geschlechts und nicht kastriert.
Altersgruppe
Anteil an Gesamtpatientenzahl
Absolute Häufigkeit (n)
Gruppe 1
(Degeneration)
Gruppe 2
(Tumor)
Gruppe 3
(Kontrolle)
1-5 Jahre 2 (10 %) 0 0 2
6-10 Jahre 8 (38 %) 4 4 -
11-15 Jahre 11 (52 %) 7 4 -

72 3 Eigene Untersuchungen
Tabelle 3-8 Geschlechtsverteilung der Hunde der Gruppen 1-3
Geschlecht Anteil an Gesamtpatientenzahl
Absolute Häufigkeit (n)
Gruppe 1
(Degeneration)
Absolute Häufigkeit (n)
Gruppe 2
(Tumor)
Absolute Häufigkeit (n)
Gruppe 3
(Kontrolle)
weiblich total 8 (38 %) 6 2 0
weiblich intakt 7 (33 %) 5 2 0
weiblich kastriert 1 (5 %) 1 0 0
männlich total 13 (62 %) 5 6 2
männlich intakt 13 (62 %) 5 6 2
männlich kastriert 0 0 0 0
Von den acht Patienten der Gruppe 2, sind zwei (25 %) weiblichen und sechs (75 %)
männlichen Geschlechts, wobei keiner der Hunde kastriert ist.
Gruppe 3 besteht aus zwei männlichen Tieren, die ebenfalls nicht kastriert sind.
3.2.2 Veränderungen der labordiagnostischen Leberwerte
Es folgt ein Überblick über einige wichtige labordiagnostische Leberwerte (ALT,
GLDH, ALKP, Gallensäure), die in der vorliegenden Arbeit bei den Hunden aller
Gruppen untersucht wurden. Die Einteilung der Laborwertveränderungen erfolgt
nach CENTER (2007) in folgende Grade: geringgradig (<5fache Erhöhung des
Referenzwertes), mittelgradig (5-10fache Erhöhung des Referenzwertes), hochgradig
(>10fache Erhöhung des Referenzwertes) sowie die Kategorie „nicht gemessen“ und
„normal“ (Werte im Referenzbereich). Die Laboruntersuchungen wurden für die
Gruppen 1 (Degeneration) und Gruppe 2 (Tumor) durchgeführt. Labordiagnostische
Untersuchungsergebnisse der Kontrolltiere lagen nicht vor.
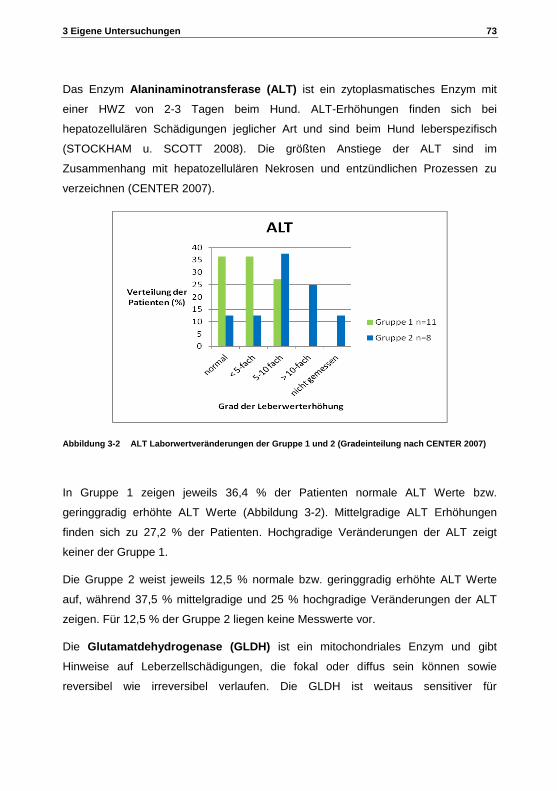
3 Eigene Untersuchungen 73
Das Enzym Alaninaminotransferase (ALT) ist ein zytoplasmatisches Enzym mit
einer HWZ von 2-3 Tagen beim Hund. ALT-Erhöhungen finden sich bei
hepatozellulären Schädigungen jeglicher Art und sind beim Hund leberspezifisch
(STOCKHAM u. SCOTT 2008). Die größten Anstiege der ALT sind im
Zusammenhang mit hepatozellulären Nekrosen und entzündlichen Prozessen zu
verzeichnen (CENTER 2007).
Abbildung 3-2 ALT Laborwertveränderungen der Gruppe 1 und 2 (Gradeinteilung nach CENTER 2007)
In Gruppe 1 zeigen jeweils 36,4 % der Patienten normale ALT Werte bzw.
geringgradig erhöhte ALT Werte (Abbildung 3-2). Mittelgradige ALT Erhöhungen
finden sich zu 27,2 % der Patienten. Hochgradige Veränderungen der ALT zeigt
keiner der Gruppe 1.
Die Gruppe 2 weist jeweils 12,5 % normale bzw. geringgradig erhöhte ALT Werte
auf, während 37,5 % mittelgradige und 25 % hochgradige Veränderungen der ALT
zeigen. Für 12,5 % der Gruppe 2 liegen keine Messwerte vor.
Die Glutamatdehydrogenase (GLDH) ist ein mitochondriales Enzym und gibt
Hinweise auf Leberzellschädigungen, die fokal oder diffus sein können sowie
reversibel wie irreversibel verlaufen. Die GLDH ist weitaus sensitiver für

74 3 Eigene Untersuchungen
hepatozelluläre Schädigungen beim Hund als die ALT (STOCKHAM u. SCOTT
2008).
Abbildung 3-3 GLDH Laborwertveränderungen der Gruppe 1 und 2 (Gradeinteilung nach CENTER 2007)
Bei Gruppe 1 ist die GLDH insgesamt etwas seltener und nicht so stark erhöht wie
bei Gruppe 2. Hier zeigen 36,4 % der Patienten keine Abweichungen vom
Normalwert, 27,2 % geringgradige Erhöhungen und jeweils 18,2 % mittelgradige
bzw. hochgradige Zunahmen der GLDH. In Gruppe 2 ist bei 37,5 % der Patienten
eine hochgradige Zunahme der GLDH zu verzeichnen, während jeweils 25 %
mittelgradige bzw. gar keine Abweichungen vom Normalwert aufweisen. Für 37,5 %
der Gruppe 2 liegen keine Messwerte vor.
Die Alkalische Phosphatase (ALKP) gehört zur Familie der Phosphatasen. In
nahezu allen Geweben lassen sich Enzymaktivitäten nachweisen. Die ALKP ist nur
bedingt als ein leberspezifisches Enzym zu betrachten (KRAFT u. DÜRR 2005).
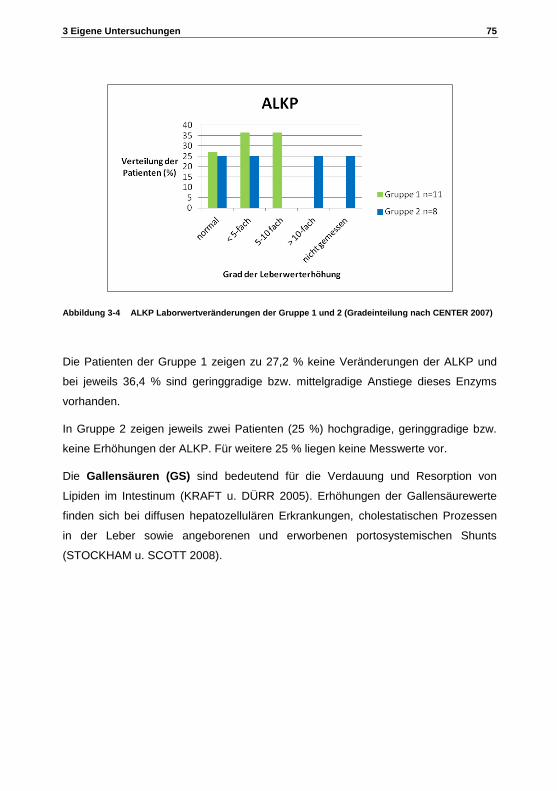
3 Eigene Untersuchungen 75
Abbildung 3-4 ALKP Laborwertveränderungen der Gruppe 1 und 2 (Gradeinteilung nach CENTER 2007)
Die Patienten der Gruppe 1 zeigen zu 27,2 % keine Veränderungen der ALKP und
bei jeweils 36,4 % sind geringgradige bzw. mittelgradige Anstiege dieses Enzyms
vorhanden.
In Gruppe 2 zeigen jeweils zwei Patienten (25 %) hochgradige, geringgradige bzw.
keine Erhöhungen der ALKP. Für weitere 25 % liegen keine Messwerte vor.
Die Gallensäuren (GS) sind bedeutend für die Verdauung und Resorption von
Lipiden im Intestinum (KRAFT u. DÜRR 2005). Erhöhungen der Gallensäurewerte
finden sich bei diffusen hepatozellulären Erkrankungen, cholestatischen Prozessen
in der Leber sowie angeborenen und erworbenen portosystemischen Shunts
(STOCKHAM u. SCOTT 2008).

76 3 Eigene Untersuchungen
Abbildung 3-5 Gallensäure Laborwertveränderungen der Gruppe 1 und 2 (Gradeinteilung nach CENTER 2007)
Die Gallensäurewerte der Gruppen 1 und 2 sind überwiegend „normal“. Bei 18,2 %
der Tiere liegen geringgradige Zunahmen vor (Abbildung 3-5). Bei der Hälfte der
Gruppe 2 (50 %) wurden keine Messungen durchgeführt. Bei den übrigen Patienten
zeigen 25 % keine Erhöhungen und jeweils 12,5 % geringgradige bzw. hochgradige
Erhöhungen der Gallensäurewerte. Ausführliche Informationen zu den übrigen
gemessenen klinisch-chemischen, wie auch hämatologischen labordiagnostischen
Parametern der Gruppe 1 und 2 sind im Anhang (Anhangstabelle 10-1 bis
Anhangstabelle 10-8) zu finden.
3.2.3 Lichtmikroskopische Untersuchungsergebnisse
Die Leberbioptate aller in die Untersuchung mit einbezogenen Patienten wurden
einer umfangreichen histopathologischen Untersuchung unterzogen. Bei der Gruppe
1 wurden die degenerativen Veränderungen qualitativ hinsichtlich ihres Charakters
beschrieben sowie semiquantitativ hinsichtlich ihres Ausprägungsgrades in gering-,
mittel- oder hochgradige Veränderungen eingeteilt. Dabei wurden insbesondere auf
die Art der hepatozellulären Degeneration (vakuolig, hydropisch, ballonier-

3 Eigene Untersuchungen 77
end/Pflanzenzelltyp), den Charakter des begleitenden Entzündungszellinfiltrates, den
Lipidgehalt der Hepatozyten, Anzahl und Morphologie der Itozellen, intrazellulär und
extrazelluläres Vorkommen von Pigmenten sowie entsprechende Begleit-
erscheinungen, wie Cholestase, stauungsbedingte Hyperämien oder Proliferation
des Gallengangsepithels geachtet. Eine Zusammenfassung der histopathologischen
Befunde von Gruppe 1 ist in Tabelle 3-9 dargestellt.
In Gruppe 1 zeigten 5 Hunde (45,5 %) insgesamt geringgradige degenerative
Leberveränderungen, während 4 Hunde (36,3 %) mittelgradige Degenerationen
aufwiesen. 2 Tiere (18,2 %) zeigten eine hochgradige Degeneration der Hepatozyten
(Abbildung 3-6). Über 50 % der degenerativen Veränderungen wurden von
Entzündungszellinfiltraten begleitet, die häufig von neutrophilen Granulozyten
dominiert wurden, aber auch gemischtzellige Infiltrate, die fokal, multifokal
herdförmig, zentrolobulär und perivaskulär auftraten. Sie kamen in minimaler,
geringgradiger und mittelgradiger Ausprägung vor. In 27 % der Fälle traten
Lipidansammlungen mit minimaler bis mittelgradiger Ausprägung auf.
Bei Gruppe 2 wurde ebenfalls das nicht-neoplastisch veränderte Leberparenchym
hinsichtlich degenerativer Veränderungen untersucht und beurteilt. Vier (50 %)
Hunde zeigten eine geringgradige Degeneration, eine mittelgradige Degeneration
konnte bei drei Hunden (37,5 %) diagnostiziert werden, während ein Patient (12,5 %)
eine hochgradige Degeneration zeigte. Außerdem wurde der vorhandene Tumor
charakterisiert und entweder als primärer Lebertumor oder als Metastase anderen
Ursprungs eingeteilt.
Eine Zusammenfassung der histopathologischen Befunde von Gruppe 2 ist in
Tabelle 3-10 und Tabelle 3-11 dargestellt.
Bei Gruppe 3 standen zwei Patienten als Kontrolltiere zur Verfügung. Das
Lebergewebe dieser Tiere wurde ebenfalls hinsichtlich degenerativer Veränderungen
untersucht und beurteilt. Beide Patienten wiesen minimale degenerative
Veränderungen auf.
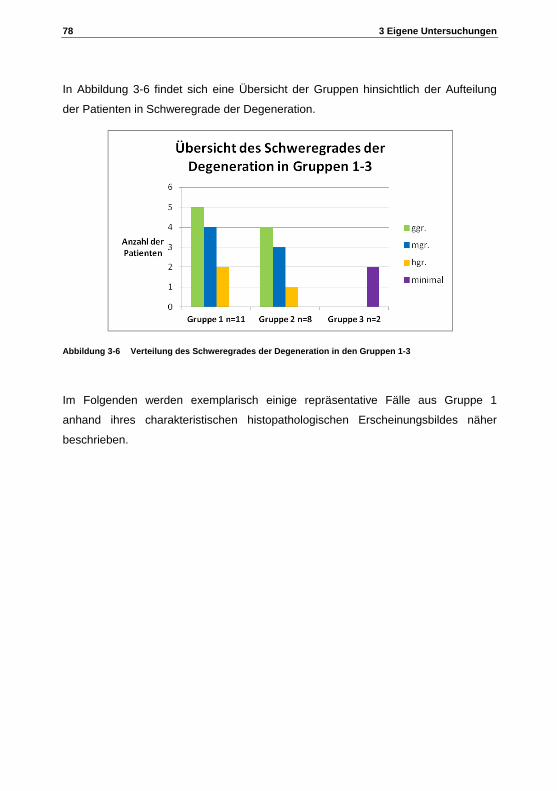
78 3 Eigene Untersuchungen
In Abbildung 3-6 findet sich eine Übersicht der Gruppen hinsichtlich der Aufteilung
der Patienten in Schweregrade der Degeneration.
Abbildung 3-6 Verteilung des Schweregrades der Degeneration in den Gruppen 1-3
Im Folgenden werden exemplarisch einige repräsentative Fälle aus Gruppe 1
anhand ihres charakteristischen histopathologischen Erscheinungsbildes näher
beschrieben.
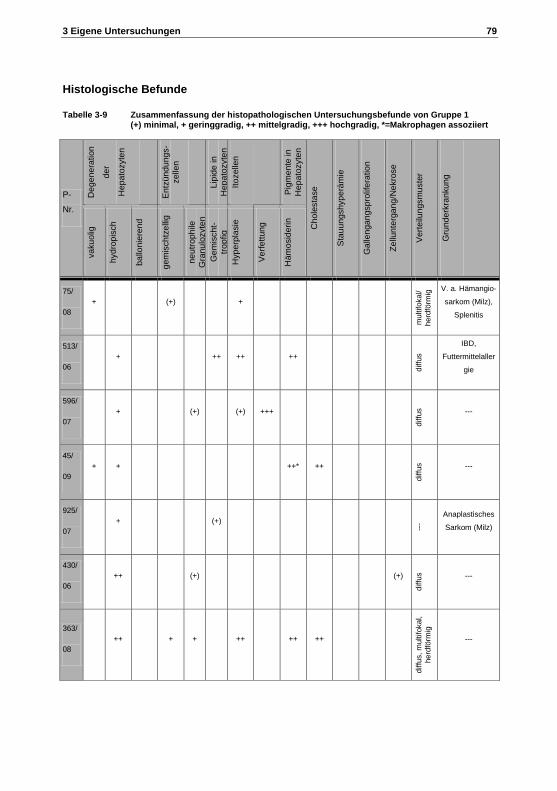
3 Eigene Untersuchungen 79
Histologische Befunde
Tabelle 3-9 Zusammenfassung der histopathologischen Untersuchungsbefunde von Gruppe 1 (+) minimal, + geringgradig, ++ mittelgradig, +++ hochgradig, *=Makrophagen assoziiert
P-
Nr.
De
ge
ne
ration
de
r
Hep
ato
zyte
n
En
tzü
nd
ung
s-
ze
llen
Lip
ide
in
Hep
ato
zyte
n
Ito
zelle
n
Pig
men
te in
Hep
ato
zyte
n
Cho
lesta
se
Sta
uu
ng
shyp
erä
mie
Ga
llen
ga
ngsp
rolif
era
tio
n
Ze
llun
terg
an
g/N
ekro
se
Ve
rte
ilun
gsm
uste
r
Gru
nd
erk
ran
ku
ng
va
ku
olig
hyd
rop
isch
ba
llon
iere
nd
ge
mis
ch
tze
llig
ne
utr
op
hile
Gra
nu
lozyte
n
Ge
mis
ch
t-
tro
pfig
Hyp
erp
lasie
Ve
rfe
ttun
g
Häm
osid
erin
75/
08
+ (+) +
multifokal/
herd
förm
ig V. a. Hämangio-
sarkom (Milz),
Splenitis
513/
06
+ ++ ++ ++
diffu
s
IBD,
Futtermittelaller
gie
596/
07
+ (+) (+) +++ diffu
s
---
45/
09
+ + ++* ++
diffu
s
---
925/
07
+ (+)
---
Anaplastisches
Sarkom (Milz)
430/
06
++ (+) (+)
diffu
s
---
363/
08
++ + + ++ ++ ++
diffu
s,
multifokal,
herd
förm
ig
---

80 3 Eigene Untersuchungen
Fortsetzung von Tabelle 3-9
P-
Nr.
Deg
en
era
tion
de
r
Hep
ato
zyte
n
En
tzü
nd
ung
s-
ze
llen
Lip
ide
in
Hep
ato
zyte
n
Ito
zelle
n
Pig
men
te in
Hep
ato
zyte
n
Cho
lesta
se
Sta
uu
ng
shyp
erä
mie
Ga
llen
ga
ngsp
rolif
era
tio
n
Ze
llun
terg
an
g/N
ekro
se
Ve
rte
ilun
gsm
uste
r
Gru
nd
erk
ran
ku
ng
va
ku
olig
hyd
rop
isch
ba
llon
iere
nd
ge
mis
ch
tze
llig
ne
utr
op
hile
Gra
nu
lozyte
n
Ge
mis
ch
tt-
rop
fig
Hyp
erp
lasie
Ve
rfe
ttun
g
Häm
osid
erin
371/
08
++ ++ (+) + + ++
diffu
s maligner
Granulosa-zelltumor
476/
06
++/++
+ + (+) +/++
---
Kortison-dauer-
medikation (Hautpatient)
539/
07
++/++
+ ++ ++ + +
diffu
s Leiomyom
(kaudal der Blase)
31/
08
+++ ++ ++ m
ultifokal,
zentr
o-lobulä
r Niere: ana-plastischer
Tumor
Geringgradige hepatozelluläre Degeneration
Geringgradige Degenerationen der Hepatozyten waren histologisch durch vakuolige,
hydropische oder auch hydropisch-vakuolige Schwellungen der Hepatozyten
charakterisiert. Die Verteilungsmuster der histologischen Veränderungen reichten
von multifokal herdförmig und herdförmig bis diffus. Die entzündlichen
Begleiterscheinungen waren gemischtzelligen Charakters, rein granulozytärer Natur
oder eine Kombination beider Merkmale. Ein Patient zeigte eine gemischttropfige
Verfettung der Hepatozyten, weitere Patienten zeigten hochgradige Verfettungen
oder auch nur vereinzelt Einlagerungen von Fett in Hepatozyten. Itozellhyperplasien
traten in geringgradigem bis mittelgradigem Umfang auf. Eine Hämosiderose

3 Eigene Untersuchungen 81
assoziiert mit Makrophagen oder Hepatozyten trat bei nahezu allen Patienten auf.
Eine repräsentative Darstellung einer geringgradigen Degeneration findet sich in
Abbildung 3-7.
Abbildung 3-7 Geringgradige Leberzelldegeneration mit ggr. Itozellhyperplasie (Pfeil) und minimalen gemischtzelligen perivaskulären Entzündungszellinfiltraten (Pfeilkopf),
(P75/08, Paraffinschnitt, H.E.).
Mittelgradige hepatozelluläre Degeneration
Die Veränderungen der Hepatozyten in der als mittelgradig eingestuften
Degeneration reichten von rein hydropischen, rein vakuoligen bis zu Mischformen
von vakuolären-hydropischen Veränderungen der Leberzellen. Das
Verteilungsmuster gestaltete sich diffus und multifokal herdförmig in den
untersuchten Lebergeweben. Entzündliche Infiltrate traten in Form von multifokal
geringgradig perivaskulär befindlichen neutrophilen Granulozyten sowie
gemischtzelligen perivaskulären Entzündungszellinfiltraten unter Beteiligung von
neutrophilen Granulozyten auf. Der überwiegende Anteil der Patienten zeigte eine
Hämosiderose und ein Patient zeigte eine Cholestase. Eine repräsentative
Darstellung einer mittelgradigen Degeneration findet sich in Abbildung 3-8.
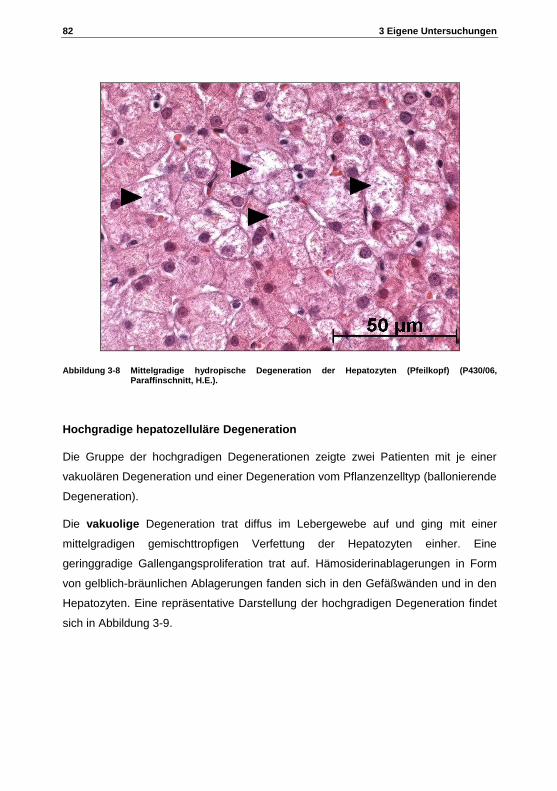
82 3 Eigene Untersuchungen
Abbildung 3-8 Mittelgradige hydropische Degeneration der Hepatozyten (Pfeilkopf) (P430/06, Paraffinschnitt, H.E.).
Hochgradige hepatozelluläre Degeneration
Die Gruppe der hochgradigen Degenerationen zeigte zwei Patienten mit je einer
vakuolären Degeneration und einer Degeneration vom Pflanzenzelltyp (ballonierende
Degeneration).
Die vakuolige Degeneration trat diffus im Lebergewebe auf und ging mit einer
mittelgradigen gemischttropfigen Verfettung der Hepatozyten einher. Eine
geringgradige Gallengangsproliferation trat auf. Hämosiderinablagerungen in Form
von gelblich-bräunlichen Ablagerungen fanden sich in den Gefäßwänden und in den
Hepatozyten. Eine repräsentative Darstellung der hochgradigen Degeneration findet
sich in Abbildung 3-9.

3 Eigene Untersuchungen 83
Abbildung 3-9 Hochgradige Degeneration der Hepatozyten. Inset zeigt eine ausgeprägte gemischttropfige Verfettung (539/07, Paraffinschnitt, H.E.).
Leberzelldegeneration vom Pflanzenzelltyp (ballonierende Degeneration)
Die Veränderungen der Hepatozyten in Form von hochgradigen
Leberzelldegenerationen vom Pflanzenzelltyp (ballonierend) fanden sich multifokal
zentrolobulär. Vor allem periportal traten multifokal mittelgradige gemischtzellige
Entzündungszellinfiltrate mit hoher Beteiligung von neutrophilen Granulozyten auf.
Eine repräsentative Darstellung einer ballonierenden Degeneration der Hepatozyten
findet sich in Abbildung 3-10.

84 3 Eigene Untersuchungen
Abbildung 3-10 Hochgradige zentrolobuläre Leberzelldegeneration vom Pflanzenzelltyp (P31/08, Paraffinschnitt, H.E.).
Sekundäre Begleiterscheinungen
Die meisten Pigmente, die im Rahmen der degenerativen Veränderungen in Gruppe
1 gefunden werden konnten, waren intrazellulär in Hepatozyten, gelblich-braun und
fein granulär. Eine positive Berliner Blau Färbung sprach für das Vorliegen von
Hämosiderin (Abbildung 3-11).
Einige Patienten zeigten eine Cholestase (Abbildung 3-12). Zahlreiche Patienten
zeigen Itozellhyperplasien (Abbildung 3-13).
Zwei Patienten zeigten Stauungshyperämien. Nekroseherde/Zelluntergänge fanden
sich nur in wenigen Fällen.

3 Eigene Untersuchungen 85
Abbildung 3-11 Geringgradige hydropische Leberzelldegeneration mit Hämosiderinspeicherung in Kupffer Sternzellen (schwarze Pfeile) (P596/07, Paraffinschnitt, Berliner Blau Färbung).
Abbildung 3-12 Geringgradige hydropische bis vakuoläre hepatozelluläre Degeneration mit Cholestase (schwarze Pfeile), (P45/09, Paraffinschnitt, H.E.).

86 3 Eigene Untersuchungen
Gruppe 2
Zwei Patienten der Gruppe 2 hatten primäre Lebertumoren, die hepatozellulären
Ursprungs waren (Tabelle 3-10). Die übrigen Tiere der Gruppe 2 wiesen Metastasen
in der Leber auf, die von Primärtumoren anderen Ursprungs entstammten. So hatten
drei Hunde ein metastatisches Hämangiosarkom. Zwei dieser Patienten zeigten
einen Primärtumor in der Milz. Der Sitz des Primärtumors des dritten Patienten war
unklar (multizentrisches Auftreten Darm/Mesenterium). Jeweils ein Hund wies eine
Lebermetastase eines Plattenepithelkarzinoms bzw. eines Phäochromoblastoms auf
und ein weiterer Hund wies hepatische Anteile eines multizentrischen Lymphoms
auf.
Tabelle 3-10 Tumorverteilung in Gruppe 2
Entstehung des Tumors
Tumorart
Anzahl
primär (n=2) Klarzellkarzinom 1
hepatozelluläres Karzinom 1
sekundär (n=6)
Hämangiosarkom 3
multizentrisches Lymphom 1
Plattenepithelkarzinom 1
Phäochromoblastom 1
Für die Untersuchung hepatozellulärer Veränderungen in der Gruppe 2 wurden nicht
neoplastisch veränderte Hepatozyten im Randbereich der Tumoren ausgewählt und
hinsichtlich Degeneration, Auftreten von Entzündungszellen, Lipidgehalt, Itozellen,
Pigmentgehalt sowie sekundären Begleiterscheinungen, wie Cholestase, Hyperämie
und Gallengangsproliferationen histologisch untersucht. Die Ergebnisse sind in
Tabelle 3-11 zusammengefasst.

3 Eigene Untersuchungen 87
Tabelle 3-11 Histopathologische Untersuchungsbefunde der Gruppe 2 (+) minimal, + geringgradig, ++ mittelgradig, +++ hochgradig
P-
Nr.
Deg
en
era
tion
En
tzü
nd
ung
s-
ze
llen
Lip
ide
in
Hep
ato
zyte
n
Ito
zelle
n
Pig
men
t in
Hep
ato
zyte
n
Cho
lesta
se
Sta
uu
ng
shyp
erä
mie
Ga
llen
ga
ngsp
rolif
era
tio
n
Ze
llun
terg
an
g/N
ekro
se
Ve
rte
ilun
gsm
uste
r
vakuolig
hydro
pis
ch
ballo
nie
rend
gem
ischtz
elli
g
mononukle
är
neutr
ophile
Gra
nulo
zyte
n
kle
invakuolig
gro
ßvakuolig
gem
ischttro
pfig
Hyperp
lasie
Verf
ettung
Häm
osid
erin
Bili
rubin
444/
06
+ + + ++
+
mulif
okal
771/
07
+ + + + ++ +
+
multifokal
803/
07
+++ +++ ++ ++ ++
467/
06
+ + ++ +/
++
++ 77/
08
++ + +
zentr
o-
lobulä
r
67/
08
+ ++ ++ ++ + + (+)
multifokal
853/
07
++ (+) +
periport
al
992/
08
++ + ++ ++
+
diffu
s

88 3 Eigene Untersuchungen
Vier Hunde (50 %) zeigten eine geringgradige Degeneration, eine mittelgradige
Degeneration konnte bei 3 Hunden (37,5 %) diagnostiziert werden, während ein
Patient (12,5 %) eine hochgradige Degeneration in der Umgebung der Neoplasie
zeigte (Abbildung 3-6).
Im Folgenden werden exemplarisch einige repräsentative Fälle aus Gruppe 2
anhand des histopathologischen Erscheinungsbildes der nicht neoplastischen
hepatozellulären Veränderungen/Degeneration näher beschrieben.
Geringgradige hepatozelluläre Degeneration im tumorangrenzenden
Lebergewebe
Geringgradige Degenerationen (Abbildung 3-13) des an den Tumor angrenzenden
Lebergewebes zeigten sich in vakuolären, hydropischen Veränderungen oder einer
Kombination beider Charakteristika sowie als ballonierende Degeneration. Es lag ein
multifokal bis diffuses Verteilungsmuster der degenerativen Veränderungen vor. Es
traten gemischtzellige und neutrophile entzündliche Infiltrate auf. Die Patienten
zeigten eine geringgradige bis mittelgradige Itozellhyperplasie und geringgradige bis
mittelgradige Hämosiderinansammlungen. Zwei Hunde zeigten eine mittelgradige
Cholestase. Teils fanden sich gering- bis mittelgradige Gallengangsproliferationen,
geringgradige Stauungshyperämien sowie minimale bis mittelgradige Nekrosen. Eine
repräsentative Darstellung einer geringgradigen vakuolären Degeneration in der
Nähe eines Tumorareals findet sich in Abbildung 3-13.

3 Eigene Untersuchungen 89
a
a a
a
B
A
A
Abbildung 3-13 Itozellhyperplasie mit Verfettung (a), geringgradige vakuoläre Leberzelldegeneration (A), Inset (B): Metastase eines Plattenepithelkarzinoms in der Leber, (P67/08, Paraffinschnitt, H.E.).
Mittelgradige hepatozelluläre Degeneration im tumorangrenzenden
Lebergewebe
Mittelgradige Leberzelldegenerationen (Abbildung 3-14) der an den Tumor
angrenzenden Leberzellen hatten einen hydropischen oder vakuolären Charakter.
Das Verteilungsmuster der degenerativen Veränderungen der Hepatozyten zeigte
sich diffus, periportal oder zentrolobulär. Die Veränderungen wurden von minimalen
bis geringgradigen gemischtzelligen und mononukleären oder neutrophilen
Entzündungsinfiltraten begleitet. Zwei der Patienten zeigten eine gering- bis
mittelgradige Itozellhyperplasie. Geringgradige Ansammlungen von Hämosiderin in
den Hepatozyten sowie Anzeichen einer Cholestase fanden sich bei je einem
Patient. In geringgradigem Umfang fanden sich bei einem Patienten Nekrosen. Eine
repräsentative Darstellung einer mittelgradigen hydropischen Degeneration im
tumorfreien Lebergewebe findet sich in Abbildung 3-14.

90 3 Eigene Untersuchungen
A
B C
D
Abbildung 3-14 Mittelgradige hydropische Leberzelldegeneration (A), intrazelluläre Pigmentablagerung (dünne Pfeile), minimale Infiltrate neutrophiler Granulozyten (dicke Pfeile); Inset (B): Metastase eines Phäochromoblastoms (C) in der Leber, tumorfreies Lebergewebe (D), (P992/08, Paraffinschnitt, H.E.).
Hochgradige hepatozelluläre Degeneration im tumorangrenzenden
Lebergewebe
Die hochgradige Degeneration (Abbildung 3-15) wurde als hydropisch bis
ballonierend charakterisiert. In mittelgradigem Umfang fand sich eine Cholestase und
Stauungshyperämie im Gewebe und in mittelgradigem Umfang traten
Gallengansproliferationen auf. Eine repräsentative Darstellung einer hochgradigen
hydropischen Degeneration im Zusammenhang mit einem Tumor findet sich in
Abbildung 3-15.

3 Eigene Untersuchungen 91
B
A
Abbildung 3-15 Hochgradige hydropische bis ballonierende Degeneration der Hepatozyten (A) im an den Tumor (Hämangiosarkom) angrenzenden Lebergewebe (großes Bild); Inset: Hämangiosarkom, (P803/07, Paraffinschnitt, H.E.).
Sonstige Veränderungen
Hyperplasien der Itozellen traten in geringgradigem bis mittelgradigem Umfang auf
(Abbildung 3-13).
Hämosiderose
Hämosiderin trat in geringgradigem bis mittelgradigem Umfang in den Makrophagen
und Hepatozyten auf (Abbildung 3-16).
Gallengangsproliferation und Stauungshyperämie
Gallengangsproliferationen fanden sich in gering- bis mittelgradigem Umfang.
Stauungshyperämien fanden sich in geringgradigem und mittelgradigem Umfang
(Abbildung 3-17).

92 3 Eigene Untersuchungen
Abbildung 3-16 Geringgradige hepatozelluläre Degeneration bei hepatozellulärem Karzinom, Hämosiderinablagerung in Kupffer Sternzellen (schwarze Pfeile) und intrahepatozellulär (rote Pfeile) (P467/06-1, Paraffinschnitt, Berliner Blau Färbung).

3 Eigene Untersuchungen 93
A
Abbildung 3-17 Ausgeprägte Gallengangsproliferation (dünne Pfeile), Stauungshyperämie (A) und hochgradige hydropische Leberzelldegeneration (Paraffinschnitt, H.E.) bei einem Tier (P803/07) mit einem Hämangiosarkom.
Gruppe 3
Für die vorliegende Arbeit wurden zwei Kontrolltiere herangezogen, die klinisch keine
Hinweise hinsichtlich einer Lebererkrankung aufwiesen. Bei der histologischen
Untersuchung der Lebern zeigten sich jedoch bei beiden Hunden geringgradige
hepatozelluläre Veränderungen, die mit minimaler hydropischer Schwellung
einhergingen (Abbildung 3-18).

94 3 Eigene Untersuchungen
Abbildung 3-18 Geringgradige hydropische Degeneration, (P2103/3, Paraffinschnitt, H.E.).

3 Eigene Untersuchungen 95
3.2.4 Elektronenmikroskopische Untersuchungsergebnisse
Neben der histologischen Untersuchung von Leberbiopsien wurden die
hepatozellulären Veränderungen der Tiere aus den drei Untersuchungsgruppen
ultrastrukturell analysiert und im Hinblick auf morphologische und quantitative
Veränderungen der Peroxisomen miteinander verglichen. Für die ultrastrukturelle
Untersuchung mittels Elektronenmikroskopie wurden zunächst Semidünnschnitte
hergestellt und auf repräsentative Veränderungen der Leberzellen für die spätere
Anfertigung von Ultradünnschnitten abgesucht. Dabei zeigte sich, dass im
Semidünnschnitt die degenerativen Veränderungen teilweise deutlicher zum
Vorschein kamen als in den zuvor untersuchten, mit H.E.-gefärbten histologischen
Schnitten, so dass im Folgenden ergänzend einige repräsentative Semidünnschnitte
mit charakteristischen hepatozellulären degenerativen Veränderungen dargestellt
wurden.
3.2.4.1 Semidünnschnitte
Geringgradige Degeneration
Abbildung 3-19 zeigt, die schon im H.E. Schnitt deutlich erkennbare diffuse
hydropische Schwellung der Hepatozyten. Sehr eindrücklich stellt sich im
Semidünnschnitt die zusätzliche multifokale gemischttropfige Verfettung dar.
Während es sich bei den weißen Punkten in anderen Schnitten um geschwollene
Mitochondrien handelt (Abbildung 3-22), sind es hier eindeutig Fetttropfen. Der
Semidünnschnitt lässt deutlich die Zellgrenzen im Gegensatz zum H.E. Schnitt
erkennen.

96 3 Eigene Untersuchungen
Abbildung 3-19 Gegenüberstellung von Semidünnschnitt und Paraffinschnitt bei einem Fall (K1972) mit geringgradiger hydropischer Leberzelldegeneration mit assoziierter gemischttropfiger Verfettung (Semidünnschnitt, Methylenblau; Paraffinschnitt, H.E.).
Der Patient K1968/1 (Abbildung 3-20) erkrankte an einem hepatozellulären Karzinom
mit adenomatöser Gallengangsproliferation. In Abbildung 3-20 ist eine geringgradige
hydropische Degeneration im an den Tumor angrenzenden Gewebe dargestellt. Der
Semidünnschnitt zeigt sehr deutlich die sich verändernde Anordnung der
Leberzellen, die auch schon im H.E. Schnitt (Inset) nicht mehr gegeben ist. Die
Hepatozyten weisen eine inhomogene Anfärbung des Zytoplasmas auf.

3 Eigene Untersuchungen 97
Abbildung 3-20 Geringgradige hepatozelluläre Degeneration (K1968/1). Der Semidünnschnitt zeigt inhomogene zytoplasmatische Färbungen mit eingelagerten grünlichen Fettvakuolen in den Hepatozyten. Daneben sieht man einen Fetttropfen (Pfeil), der im H.E. Schnitt (Inset) als leere Vakuole erkennbar ist. Im H.E. Schnitt sind zusätzlich Herde mit Hämosiderinablagerungen (Kreis). (Semidünnschnitt, Methylenblau; Paraffinschnitt, H.E. Färbung).
Mittelgradige hepatozelluläre Degeneration
Im Semidünnschnitt sind im Vergleich zum Paraffinschnitt verschiedene Einzelheiten
der jeweiligen hepatozellulären Degeneration besonders sichtbar. Während der
Paraffinschnitt eher nur diffuse Veränderungen einer Degeneration mit mittelgradiger
Ausprägung aufweist, zeigen die Semidünnschnitte deutlich abgegrenzte
Hepatozyten, bei denen die Zellkerne überwiegend noch gut erhalten sind. Im
Zytoplasma sind eindeutige Charakteristika der Degeneration sichtbar, z. B.
ausgeprägte Glykogenfelder, die den Hepatozyten im Paraffinschnitt hydropisch
geschwollen erscheinen lassen (Abbildung 3-21).

98 3 Eigene Untersuchungen
Abbildung 3-21 Mittelgradige hepatozelluläre Degeneration bei einem Hund mit Klarzellkarzinom (K2011/20). Die im Inset sichtbare mittelgradige hydropische Schwellung der Hepatozyten ist im Semidünnschnitt auf eine massive Glykogenablagerung mit einzelnen feinen Fettvakuolen zurückzuführen. (Semidünnschnitt, Methylenblau; Paraffinschnitt, H.E. Färbung).
Hochgradige hepatozelluläre Degeneration vom Pflanzenzelltyp
Auch bei den Fällen mit hochgradigen hepatozellulären Degenerationen lassen sich
in Semidünnschnitten von Biopsien der Tiere mit und ohne tumoröse Veränderungen
deutlich mehr Einzelheiten als im H.E. gefärbten Paraffinschnitt erkennen (Abbildung
3-22). Da sich die individuellen Zellen deutlich darstellen, ist die ausgeprägte
Schwellung sichtbar. Die Zellkerne werden an die Peripherie des Zytoplasmas
gedrängt, oft fehlen Anschnitte der Kerne in der Schnittebene oder es fallen
Kernuntergangsformen auf. Durch die unterschiedliche Schwellung der Zellen wird
die Balkenstruktur der Lobuli sehr unregelmäßig und teilweise aufgelöst. Bei
Schwellung der Zellen mit Akzentuierung der Zellwände wird bei einigen Fällen die
charakteristische pflanzenzellähnliche Erscheinung der Hepatozyten deutlich, die die
Degeneration vom Pflanzenzelltyp lichtmikroskopisch charakterisiert.

3 Eigene Untersuchungen 99
Im Vergleich mit Kontrolltieren (Abbildung 3-23) färbt sich das Zytoplasma der
Hepatozyten unterschiedlich und inhomogen an. Im Zytoplasma finden sich je nach
Art der Degeneration Einschlüsse, die von Fetttropfen über Pigmentablagerungen
und ausgeprägten Vakuolisierungen reichen. Grundsätzlich zeigen die mehr oder
weniger gut erhaltenen Hepatozyten eine Vielzahl kleiner Vakuolen, bei denen es
sich um geschwollene Mitochondrien handelt.
Bei den Tieren mit diffusen hepatozellulären Degenerationen zeigen sich auf
lichtmikroskopischer Ebene am Semidünnschnitt keine Unterschiede zu den Tieren
mit Tumorerkrankungen, bei denen degenerative Hepatozytenareale in Tumornähe
ausgewählt wurden.
F
F
F
F
G
G
Abbildung 3-22 Ausgeprägte hepatozelluläre Degenerationserscheinungen bei einem Tier (K2025/3) aus der Gruppe 1. Neben Fetttropfen (F) zeigen sich Degenerationserscheinungen durch starke deutliche Schwellung der Hepatozyten (Pfeilspitzen). Andere Hepatozyten zeigen intrazytoplasmatische Pigmentablagerungen (dünne Pfeile) oder sind durch unregelmäßige Glykogenablagerungen (G) charakterisiert. In fast allen Zellen sind kleine Vakuolen zu erkennen, bei denen es sich um geschwollene Mitochondrien handelt (Semidünnschnitt, Methylenblau).

100 3 Eigene Untersuchungen
Kontrolltiere
Wie bei der lichtmikroskopischen Auswertung an H.E. gefärbten Paraffinschnitten
zeigen auch die Semidünnschnitte der beiden Kontrolltiere an einzelnen Hepatozyten
vereinzelte Anzeichen einer hepatozellulären Degeneration. Diese äußern sich in
Form von leichten Vakuolisierungen im Zytoplasma oder hydropischen Schwellungen
einzelner Hepatozyten. Überwiegend zeigt die Leber der Kontrolltiere einen
regulären Aufbau der Leberbälkchen mit Hepatozyten, die ein homogen angefärbtes
Zytoplasma aufweisen (Abbildung 3-23).
Abbildung 3-23 Kontrolltier (K2103) mit einzelnen Vakuolen in den ansonsten regelmäßig angeordneten Hepatozyten, die ein homogen gefärbtes Zytoplasma aufweisen (Semidünnschnitt, Methylenblau).

3 Eigene Untersuchungen 101
3.2.4.2 Ultradünnschnitte
3.2.4.2.1 Quantitative Unterschiede der Peroxisomenzahl
Nachdem anhand der Semidünnschnitte repräsentative Ultradünnschnitte aus den
Leberproben angefertigt wurden, konnten diese im Hinblick auf morphologische
sowie quantitative Unterschiede der Peroxisomen zwischen den Gruppen
ultrastrukturell untersucht werden.
Für die quantitative Untersuchung der hepatozellulären Peroxisomen und deren
Vergleich zwischen den Untersuchungsgruppen sowie in Abhängigkeit von Alter und
Rasse/Gewichtsklasse der untersuchten Tiere wurden die Peroxisomen in jeweils
sechs Hepatozyten pro Patient ausgezählt und auf die Gesamtfläche der
Hepatozyten (µm²) bezogen.
3.2.4.2.2 Morphologische Unterschiede der Peroxisomen
Für die morphologische Untersuchung wurden die hepatozellulären Peroxisomen
hinsichtlich ihrer Größe, ihrer Verteilung innerhalb der Zelle sowie ihrer Form
(Protrusion, Marginalplatte, Nukleoid) beurteilt. Darüber hinaus wurden die
hepatozellulären Mitochondrien beschrieben und die Anwesenheit von Lipidtropfen,
Myelinfiguren und der Glykogengehalt semiquantitativ erfasst.
Die Ergebnisse der morphologischen Untersuchungen der hepatozellulären
Peroxisomen und Mitochondrien sowie der semiquantitativen Auswertung von
hepatozellulären Lipidtropfen, Myelinfiguren und Glykogengehalt sind für Gruppe
1(Tiere ohne Tumor) in Tabelle 3-12, für Gruppe 2 (Tiere mit Tumor) in Tabelle 3-13
und für Gruppe 3 (Kontrolltiere) in Tabelle 3-14 zusammengefasst.

102 3 Eigene Untersuchungen
Gruppe 1
Tabelle 3-12 Semiquantitative Beurteilung der Morphologie der Peroxisomen und der Hepatozyten bei Tieren ohne Lebertumoren (+) vereinzelt (<10 %), + geringgradig (10-30 %), ++ mittelgradig (30-50 %), +++ hochgradig (50-100 %), x vorhanden, - nicht vorhanden, n.b. nicht beurteilbar
Merkmal P75/
08
K 2029
P513/
06
K 1972
P596/
07
K 1979
P45/
09
K 2015
P925/
07
K 2017
P430/
06
K 1963
P363/
08
K 2062
P371/
08
K 2063
P476/
06
K 1969
P539/
07
K 1974
P31/
08
K 2025
Px klein
Durchmesser:
ca.555nm
+ + ++ ++ ++ + (+) + (+) ++ ++
Px groß
Durchmesser:
ca. 918 nm
+ + + + + + + - ++ (+) (+)
Verteilung in der
Zelle
Nähe
Fett-
tropfen
Cluster Pan-
zytär
Pan-
zytär
Cluster Pan-
zytär
Cluster
Pan-
zytär
Pan-
zytär
Cluster
Pan-
zytär
Peri-
nukleär
Pan-
zytär
Protrusion x x x - - x x - - - -
Marginalplatte x x x - x x x x x x x
Nukleoid x x x x - x x x x - -
Mitochondrien
Schwellung x - x - x - x x - x x
Lyse - - x - x - - x - x x
Parakristalline
Einschlüsse - - x - x x - - x - -
Riesen-
mitochondrien x x x x x x x x x x -
Lipidtropfen ++ + (+) ++ (+) + - - + ++ -
Myelinfiguren + - ++ ++ + + + ++ (+) + +
Glykogen - +++ +++ +++ ++ +++ + +++ +++ +++ ++

3 Eigene Untersuchungen 103
Gruppe 2
Tabelle 3-13 Semiquantitative Beurteilung der Morphologie der Peroxisomen und der Hepatozyten bei Lebertumoren (+) vereinzelt (<10 %), + geringgradig (10-30 %), ++ mittelgradig (30-50 %), +++ hochgradig (50-100 %), x vorhanden, - nicht vorhanden, n.b. nicht beurteilbar
Merkmal
P 444/06
K 1965
P 771/07
K 2006
P 803/07
K 2009
P 467/06
K 1968
P 77/08
K 2030
P 67/08
K 2027
P 853/07
K 2011
P992/08
K 2021
Px klein
Durchmesser:
ca. 555nm
+ + + + + + + +
Px groß
Durchmesser:
ca. 918 nm
+ + + + + + + -
Verteilung in
der Zelle
panzytär panzytär panzytär panzytär panzytär
Cluster
panzytär panzytär panzytär
Cluster
Protrusion x - x x x x - x
Marginalplatte x - x x x x - x
Nukleoid x - x - x x - x
Mitochondrien
Schwellung x x x - x x - x
Lyse x x x - x x - x
Parakristalline
Einschlüsse
- - x - - - x x
Riesenmitochondrien - - x x - x x -
Lipidtropfen - - ++ + - + - -
Myelinfiguren +++ +++ ++ (+) (+) ++ - ++
Glykogen ++ + +++ ++ ++ + +++ ++

104 3 Eigene Untersuchungen
Gruppe 3
Tabelle 3-14 Semiquantitative Beurteilung der Morphologie der Peroxisomen und der Hepatozyten bei Kontrolltieren (+) vereinzelt (<10 %), + geringgradig (10-30 %), ++ mittelgradig (30-50 %), +++ hochgradig (50-100 %), x vorhanden, - nicht vorhanden, n.b. nicht beurteilbar
Merkmal K 2104-7 K 2104-8 K 2104-9 K 2103-3 K 2103-12 K 2103-15
Px klein
Durchmesser:
ca. 555nm
+ + + ++ + ++
Px groß
Durchmesser:
ca. 918 nm
+++ +++ +++ + +++ ++
Verteilung in der Zelle
panzytär
Cluster
panzytär
Cluster
panzytär
Cluster
perinukleär
Cluster
perinukleär
Cluster
panzytär
Cluster
Protrusion - x x x x -
Marginalplatte x - x - x -
Nukleoid x x x x - x
Mitochondrien
Schwellung x x x x x x
Lyse - x - x x -
Parakristalline
Einschlüsse
- - x - - -
Riesenmitochondrien x x - x x x
Lipidtropfen (+) + + (+) (+) +
Myelinfiguren + + (+) + ++ +
Glykogen +++ +++ +++ +++ +++ +++

3 Eigene Untersuchungen 105
Unterschiede in der Peroxisomengröße
Bei der ultrastrukturellen Untersuchung der Hepatozyten wurde das Vorkommen von
Peroxisomen unterschiedlicher Größe semiquantitativ erfasst, wobei in der
Auswertung die Anzahl der Peroxisomen mit keine (0%), vereinzelt (<10 %),
geringgradig (10-30 %), mittelgradig (30-50 %) und hochgradig (50-100 %) erfasst
wurde. Zusätzlich wurde dabei zwischen kleinen (Durchmesser ca. 555 nm) und
großen (Durchmesser ca. 918 nm) Peroxisomen unterschieden.
Es zeigte sich, dass in Gruppe 1 45,4 % der Patienten eine mittelgradige Anzahl
kleiner Peroxisomen aufwies, während bei 36,5 % der Patienten geringgradig kleine
und bei 18,1 % nur vereinzelt kleine Peroxisomen gefunden wurden. Das
Vorkommen großer Peroxisomen war dagegen in Gruppe 1 anders verteilt. So
enthielten die Hepatozyten von 63,6 % der Patienten große Peroxisomen in geringer
Anzahl, während zwei Patienten (18,2 %) nur vereinzelt Peroxisomen aufwiesen. Bei
einem Patienten dieser Gruppe konnten keine Peroxisomen festgestellt werden,
während ein Tier große Peroxisomen in mittelgradiger Anzahl besaß.
In Gruppe 2 zeigten sich in den Hepatozyten aller Patienten nahezu gleichermaßen
kleine sowie große Peroxisomen in geringgradigem Ausmaß, wobei ein Patient gar
keine großen Peroxisomen aufwies.
Die Kontrollgruppe (Gruppe 3) zeichnete sich dadurch aus, dass die meisten der
untersuchten Hepatozyten (66,6 %) hochgradig große Peroxisomen enthielten und
nur in einem Schnittpräparat jeweils eine gering- bis mittelgradige Anzahl großer
Peroxisomen gefunden wurde. Kleine Peroxisomen waren in 66,7 % der Fälle in
geringgradiger Anzahl feststellbar.
Eine Übersicht über die Verteilung großer und kleiner Peroxisomen in den drei
Untersuchungsgruppen ist in Abbildung 3-24 und Abbildung 3-25 dargestellt.
Die vergleichende statistische Auswertung aller Gruppen hinsichtlich des Auftretens
von kleinen Peroxisomen in den Hepatozyten ergab keinen signifikanten Unterschied
(Kruskal Wallis Test), während bei den großen Peroxisomen ein signifikanter

106 3 Eigene Untersuchungen
Unterschied festzustellen war (Kruskal Wallis Test p-Wert 0,0288; signifikant ab
p <0,05).
Ein paarweiser Gruppenvergleich ist aufgrund einer zu kleinen Gruppengröße aller
Untersuchungsgruppen nicht durchgeführt worden.
Abbildung 3-24 Verteilung kleiner Peroxisomen in den Gruppen 1-3
Abbildung 3-25 Verteilung großer Peroxisomen in den Gruppen 1-3

3 Eigene Untersuchungen 107
Verteilung der Peroxisomen in der Zelle
Bei der Untersuchung des Verteilungsmusters der Peroxisomen innerhalb der
Hepatozyten wurde zwischen einem panzytären Auftreten, Gruppenbildung (Cluster)
(Abbildung 3-27), Anhäufungen im Bereich des Kernes (perinukleär) (Abbildung
3-28) und sonstigen Verteilungsmustern unterschieden. Wie in Abbildung 3-26
ersichtlich, waren die Peroxisomen in Gruppe 1 und 2 am häufigsten panzytär
verteilt, gefolgt von Clusterbildung. In der Kontrollgruppe hingegen waren die
Peroxisomen am häufigsten in Clustern angeordnet und weniger häufig panzytär
bzw. perinukleär. In Gruppe 1 konnte außerdem bei einem Patienten eine deutlich
mit Fetttropfen und degenerierten Mitochondrien (Abbildung 3-29) assoziierte
Lokalisation der Peroxisomen beobachtet werden. Hinsichtlich der statistischen
Auswertung der Verteilung der Peroxisomen in den vier Merkmalen (Nähe
Fettropfen, Cluster, panzytär, perinukleär) ergab sich lediglich in einem Merkmal
(Cluster) ein signifikantes Ergebnis. Gruppe 1 zeigte das Auftreten der Peroxisomen
in Form eines Clusters zu 36,4 %, während Gruppe 2 das Merkmal zu 25 % aufwies
und Gruppe 3 zu 100 %. Der Unterschied erwies sich zwischen allen drei Gruppen
im Merkmal (Cluster) als signifikant p 0,01233 (p Wert signifikant ab p <0,05).
Abbildung 3-26 Peroxisomales Verteilungsmuster der Gruppen 1-3

108 3 Eigene Untersuchungen
M
Abbildung 3-27 Verteilungsmuster der Peroxisomen (Pfeile) als Cluster. Mitochondrium (M) (K2103/15, Ultradünnschnitt, Transmissionselektronenmikroskopie, Gerätevergr. 16.000x).
N MM
MM
M
Abbildung 3-28 Perinukleäres Verteilungsmuster der Peroxisomen (Pfeile). Nukleus (N), Mitochondrium (M) (K2103/12, Ultradünnschnitt, Transmissionselektronenmikroskopie, Gerätevergr. 6300x).

3 Eigene Untersuchungen 109
Px
Px
Px
Px F
FF
F
M
Abbildung 3-29 Verteilung der Peroxisomen (Px) im Bereich von Fetttropfen (F) und Mitochondrium (M). (K2029, Ultradünnschnitt, Transmissionselektronenmikroskopie, Gerätevergr. 16.000x).
Morphologische Veränderungen der Peroxisomen
Die hepatozellulären Peroxisomen wurden darüber hinaus hinsichtlich ihrer
morphologischen Eigenschaften in den verschiedenen Untersuchungsgruppen
beurteilt (Abbildung 3-30). Dabei zeigte sich, dass in Gruppe 1 Marginalplatten
(Abbildung 3-32) am häufigsten mit 90,1 % zu finden waren, gefolgt vom Auftreten
eines Nukleoids (72,3 %) (Abbildung 3-33). In weniger als 50 % der Fälle wiesen die
Peroxisomen in Gruppe 1 Protrusionen (Abbildung 3-31) auf. In Gruppe 2 dagegen
wurden Protrusionen (75 %) ebenso häufig wie Marginalplatten (75 %) festgestellt.
Das Auftreten eines Nukleoids war geringfügig (62,5 %). In der Kontrollgruppe waren
Nukleoide dagegen am häufigsten nachweisbar (83,3 %), gefolgt von Protrusionen
(66,7 %) und dem Vorhandensein einer Marginalplatte (50 %). Bei der statistischen
Auswertung mit Hilfe des Pearson Chi-Quadrat Testes konnten in keinem Fall
signifikante Unterschiede zwischen den Untersuchungsgruppen ermittelt werden
(Protrusion p-Wert 0,40036; Marginalplatte p-Wert 0,16792; Nukleoid p-Wert 0,68960
signifikant ab p<0,05000).

110 3 Eigene Untersuchungen
Abbildung 3-30 Morphologische Veränderungen der Peroxisomen der Gruppen 1-3
PxM
N
Abbildung 3-31 Protrusion (roter Pfeil) eines Peroxisoms (Px). Mitochondrium (M), Nukleus (N) (K1965, Ultradünnschnitt, Transmissionselektronenmikroskopie, Gerätevergr. 12.000x).

3 Eigene Untersuchungen 111
M
Px
Px
Px
Px
Abbildung 3-32 Marginalplatten (rote Pfeile) der Peroxisomen (Px). Mitochondrium (M), das geschwollen ist. (K2029, Ultradünnschnitt, Transmissionselektronenmikroskopie, Gerätevergr. 16.000x).
Px
Nk
Abbildung 3-33 Peroxisom (Px) mit Nukleoid (Nk) und Marginalplatte (Pfeil). (K1969, Ultradünnschnitt, Transmissionselektronenmikroskopie, Gerätevergr. 25.000x).

112 3 Eigene Untersuchungen
Mitochondriale Eigenschaften
Neben den Peroxisomen wurden außerdem die hepatozellulären Mitochondrien
hinsichtlich Schwellungen, lytischen Prozessen und parakristallinen Einschlüssen
sowie der Anwesenheit von Riesenmitochondrien ultrastrukturell untersucht
(Abbildung 3-34).
Dabei zeigten sich in Gruppe 1 am häufigsten Riesenmitochondrien (90,1 %)
(Abbildung 3-35) gefolgt von Schwellungen (63,6 %) (Abbildung 3-37), Lysen (45,5
%) (Abbildung 3-36) und Einschlüssen (36,4 %) (Abbildung 3-35). In Gruppe 2 waren
die am häufigsten beobachteten mitochondrialen Veränderungen in Form von
Schwellung (75 %) und Lyse (75 %) zu finden. Riesenmitochondrien und
parakristalline Einschlüsse kamen dagegen nur bei der Hälfte bzw. etwas mehr als
einem Drittel der untersuchten Patienten von Gruppe 2 vor. In der Kontrollgruppe
zeigten sämtliche Lokalisationen eine ausgeprägte Schwellung der Mitochondrien
und sehr häufig waren Riesenmitochondrien anzutreffen (83,3 %). Die Hälfte der
untersuchten Lokalisationen wiesen lytisch veränderte Mitochondrien auf und nur zu
16,7 % konnten parakristalline Einschlüsse gefunden werden. Die statistische
Auswertung mit dem Pearson Chi square-Test hinsichtlich der mitochondrialen
Eigenschaften Schwellung, Lyse, parakristallinen Einschlüssen und
Riesenmitochondrien ergab keinen signifikanten Unterschied (Schwellung p-Wert
2,4403; Lyse p-Wert 0,41558; Einschlüsse p-Wert 0,65189; Riesenmitochondrien p-
Wert 0,10634; signifikanter p-Wert <0,0500).

3 Eigene Untersuchungen 113
Abbildung 3-34 Eigenschaften der Mitochondrien der Gruppen 1-3
RM
M
Abbildung 3-35 Riesenmitochondrium (RM) mit parakristallinen Einschlüssen (Pfeile) in Nachbarschaft zu einem normal großen Mitochondrium (M) verdeutlicht den Größenunterschied. (K1979, Ultradünnschnitt, Transmissionselektronenmikroskopie, Gerätevergr. 16.000x).
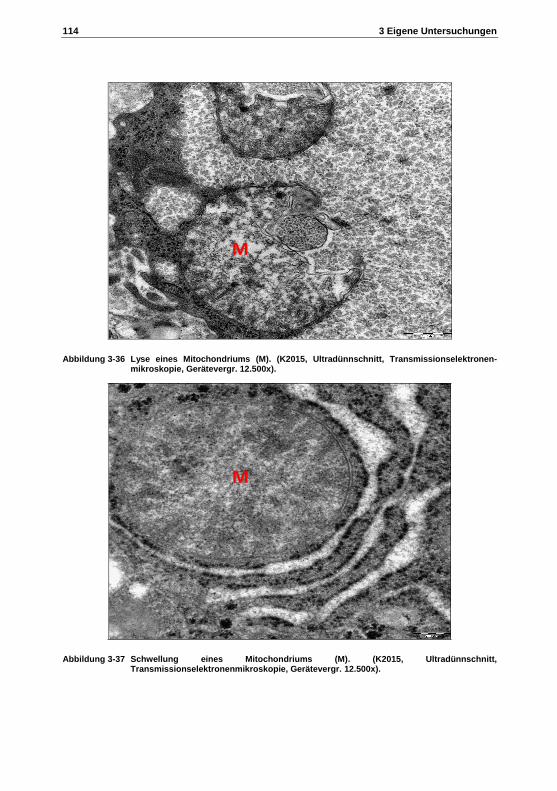
114 3 Eigene Untersuchungen
M
Abbildung 3-36 Lyse eines Mitochondriums (M). (K2015, Ultradünnschnitt, Transmissionselektronen-mikroskopie, Gerätevergr. 12.500x).
M
Abbildung 3-37 Schwellung eines Mitochondriums (M). (K2015, Ultradünnschnitt, Transmissionselektronenmikroskopie, Gerätevergr. 12.500x).

3 Eigene Untersuchungen 115
Lipidgehalt, Myelinfiguren und Glykogengehalt
Auch elektronenmikroskopisch konnten intrahepatozelluläre Lipidtropfen in allen
Untersuchungsgruppen nachgewiesen werden (Abbildung 3-39). Entsprechend
wurden ausgewählte Hepatozyten aus den drei Untersuchungsgruppen auf ihren
Lipid- und Glykogengehalt sowie auf das Vorkommen von Myelinfiguren untersucht
und semiquantitativ ausgewertet (Abbildung 3-38). Dabei zeigte sich, dass in Gruppe
1 tendenziell die meisten Lipidtropfen mit mittelgradigen Mengen bei fast 30 % der
untersuchten Hepatozyten nachgewiesen werden konnten. In Gruppe 2 zeigten
dagegen nur 12,5 % einen mittelgradigen Lipidgehalt und die Mehrheit der Patienten
wiesen überhaupt keine Lipidtropfen auf (62,5 %). In der Kontrollgruppe waren
minimale bis geringgradige Lipidgehalte gleichermaßen in den untersuchten
Schnittebenen verteilt. Zur statistischen Auswertung wurde der Kruskal-Wallis Test
herangezogen. Dabei wurden die drei Untersuchungsgruppen hinsichtlich des
Merkmals Lipidtropfen global betrachtet. Hierbei ergab sich ein signifikanter p-Wert
0,0107 (p-Wert signifikant <0,05000). Im Paarvergleich, der mittels des Mann-
Whitney U-Testes durchgeführt wurde, fand sich zwischen Gruppe 2 und 3 ein
signifikanter Unterschied p-Wert 0,008132 (p-Wert signifikant <0,017).
Abbildung 3-38 Häufigkeit des Vorkommens von Lipidtropfen in den Hepatozyten der Untersuchungsgruppen 1-3

116 3 Eigene Untersuchungen
Hepatozyt
F
F
FF
F
FF
Abbildung 3-39 Gemischttropfige Verfettung (F) eines Hepatozyten. (K1972, Ultradünnschnitt, Transmissionselektronenmikroskopie, Gerätevergr. links 2500x und rechts 3150x).
Myelinfiguren als Einschlüsse in Mitochondrien oder Autophagosomen
repräsentieren möglicherweise pathologische Konformationen von Membran-
proteinen und Lipoproteinschichten und wurden vor allem in Gruppe 2 in gehäufter
Form beobachtet. Hier zeigten 37,5 % der Hepatozyten mittelgradige und 25 %
hochgradige Anhäufungen von Myelinfiguren (Abbildung 3-40 und Abbildung 3-41),
während sie bei 25 % der Hepatozyten nur in minimaler Ausprägung gefunden
wurden. In 12,5 % der Gruppe 2 konnten keine Myelinfiguren nachgewiesen werden.
In Gruppe 1 waren vor allem geringgradige Mengen von Myelinfiguren erkennbar
(54,5 %) und bei 27,3 % traten sie in mittelgradiger Ausprägung auf. Der Rest zeigte
entweder keine (9,1 %) oder nur minimale (9,1 %) Anzeichen von Myelinfiguren. Bei
keinem Patienten der Gruppe 1 wurden hochgradige Anhäufungen von Myelinfiguren
festgestellt. In der Kontrollgruppe wurden zu gleichen Teilen minimale bis
geringgradige Mengen von Myelinfiguren nachgewiesen. Die statistische Auswertung
mit Hilfe des Kruskal-Wallis-Tests ergab keinen signifikanten Unterschied p-Wert

3 Eigene Untersuchungen 117
0,2402 (p-Wert signifikant ab p <0,05000). Ein Paarvergleich wurde nicht
durchgeführt.
Abbildung 3-40 Häufigkeit des Auftretens von Myelinfiguren
M
MF
Abbildung 3-41 Myelinfigur (MF) in unmittelbarer Nachbarschaft zu einem Mitochondrium (M). (K2015, Ultradünnschnitt, Transmissionselektronenmikroskopie, Gerätevergr. 16.000x).

118 3 Eigene Untersuchungen
In Gruppe 1 und 3 war der Glykogengehalt der untersuchten Leberzellen am
höchsten (Abbildung 3-42). Das Glykogen stellt sich dabei elektronenmikroskopisch
als Ansammlung feingranulären Materials dar (Abbildung 3-43). Die Kontrollgruppe
zeigte dabei einen hochgradigen Glykogengehalt in 100 % der untersuchten Zellen.
In Gruppe 1 wurden bei 63,6 % hochgradige Mengen und bei 18,2 % mittelgradige
Mengen an Glykogen festgestellt. Der Rest zeigte entweder nur wenig oder gar kein
Glykogen. In Gruppe 2 zeigte die Hälfte der untersuchten Patienten mittelgradige
Glykogenmengen, während jeweils ein Viertel entweder geringgradige oder
hochgradige Glykogengehalte aufwies. Die statistische Auswertung mittels des
Kruskall-Wallis-Test ergab eine Signifikanz. Der p-Wert lag bei 0,0217 (p-Wert
signifikant ab <0,05000). Der Paarvergleich der Gruppen untereinander mittels des
Mann-Whitney U-Testes ergab keine signifikanten Werte (p-Wert signifikant ab p
<0,017).
Abbildung 3-42 Glykogengehalt der Hepatozyten der Untersuchungsgruppen 1-3

3 Eigene Untersuchungen 119
PxNk
Abbildung 3-43 Ausgeprägte Glykogenansammlungen (rote Pfeile) neben einem Peroxisom (Px) mit Nukleoid (N) und sichtbarer Marginalplatte (schwarzer Pfeil). (K1972, Ultradünnschnitt, Transmissionselektronenmikroskopie, Gerätevergr.16.000x).
3.2.4.2.4 Quantitative Unterschiede
Um herauszufinden, ob sich die Anzahl der Peroxisomen in Abhängigkeit von
unterschiedlichen Krankheitsprozessen in der Leber sowie je nach Alter bzw.
Rasse/Gewichtsklasse der untersuchten Hunde unterscheidet, wurden die
Peroxisomen in sechs repräsentativen Hepatozyten pro Patient bzw. 18 Hepatozyten
pro Kontrolltier (jeweils 6 Hepatozyten aus 3 verschiedenen Lokalisationen)
ausgezählt. Dabei wurde die ausgezählte Peroxisomenmenge auf die Fläche des
untersuchten Hepatozyten bezogen und aus allen untersuchten Hepatozyten der
Mittelwert gebildet. Die Tabellen mit den ausgezählten Peroxisomen pro µm²
Hepatozyt sowie deren Mittelwerte sind im Anhang (Anhangstabelle 10-11)
aufgeführt. Im Anhang (Anhangstabelle 10-12) findet sich eine Gesamtübersicht der
zur statistischen Untersuchung verwendeten Daten.

120 3 Eigene Untersuchungen
Unterschiede zwischen den Gruppen
Die durchschnittliche Peroxisomenzahl/µm² Hepatozyt betrug für die Gruppe 1 0,13
und lag damit etwas höher als in den Gruppen 2 und 3 mit jeweils 0,1
Peroxisomen/µm² Hepatozyt (Abbildung 3-44). Beim Vergleich der untersuchten
Patientengruppen untereinander sowie mit den Kontrolltieren konnten keine
signifikanten Unterschiede zwischen den Hunden der Gruppe 1, Gruppe 2 und den
Kontrolltieren (Gruppe 3) gefunden werden (einfaktorielle ANOVA, p-Wert 0,17515).
Abbildung 3-44 Darstellung der Mittelwerte der Peroxisomenzahl/Hepatozytenfläche (µm²) in den drei Untersuchungsgruppen
Altersabhängige Unterschiede
Da sich zwischen den Untersuchungsgruppen keine statistisch signifikanten
Unterschiede der Peroxisomenzahl/µm² Hepatozyt ergaben und die Patientenzahl
pro Untersuchungsgruppe für eine Einteilung in unterschiedliche Altersstufen
innerhalb der Gruppe zu niedrig war, wurden sämtliche untersuchten Tiere aus allen
Gruppen zusammengefasst und in drei Altersklassen (1-5 Jahre, 6-10 Jahre, 11-15
Jahre) eingeteilt, um mögliche altersabhängige Unterschiede der
0,00
0,02
0,04
0,06
0,08
0,10
0,12
0,14
0,16
0,18
Gruppe 1 Gruppe 2 Gruppe 3
n/µm²
Durchschnittliche Px Zahl pro µm² Hepatozyt (Mittelwerte mit Standardabweichung)

3 Eigene Untersuchungen 121
Peroxisomenanzahl/µm² Hepatozyt zu ermitteln (Anhangstabelle 10-12). Dabei
zeigte sich, dass Tiere im Alter von 11-15 Jahren durchschnittlich etwas mehr
Peroxisomen/µm² Hepatozyt (0,12) aufwiesen, als jüngere Tiere (jeweils ca. 0,1
Peroxisom/µm² Hepatozyt). Die statistische Analyse mittels eines einfaktoriellen
ANOVA Testes ergab jedoch keine signifikanten Unterschiede (p-Wert 0,38906)
zwischen den verschiedenen Altersklassen (Abbildung 3-45).
1-5 Jahre 6-10 Jahre 11-15 Jahre
Altersgruppen
0,06
0,07
0,08
0,09
0,10
0,11
0,12
0,13
0,14
0,15
0,16
Mitte
lwert
e d
er
Pero
xis
om
en/ F
läche µ
m²
Abbildung 3-45 Mittelwerte der Peroxisomen/Hepatozytenfläche(µm²) in Bezug zu den Altersgruppen
Gewichts- bzw. rasseabhängige Unterschiede
Neben altersabhängigen Unterschieden wurden außerdem quantitative Unterschiede
der hepatozellulären Peroxisomen in Abhängigkeit der Rasse bzw. des Gewichtes
der untersuchten Hunde ausgewertet. Hierzu wurden ebenfalls sämtliche Tiere aus
allen Gruppen zusammengefasst und in drei Gewichtsklassen (0-10 kg, 10-20 kg,
20-40 kg) eingeteilt (Anhangstabelle 10-12). Beim Vergleich der durschnittlichen
Peroxisomenanzahl/µm² Hepatozyt fiel auf, dass die Hunde kleiner Rassen bzw. der

122 3 Eigene Untersuchungen
Gewichtsklasse1 (0-10 kg) im Mittel deutlich mehr Peroxisomen aufwiesen als Hunde
mittlerer (10-20 kg) bzw. großer Rassen (20-40 kg) Gewichtsklassen. Die statistische
Auswertung mittels des einfaktoriellen ANOVA Tests der verschiedenen
Gewichtsklassen in Bezug auf die Mittelwerte der Peroxisomenanzahl/µm² Hepatozyt
ergab einen signifikanten Unterschied (p-Wert 0,04023). Der Paarvergleich ergab
unter Verwendung des t-Testes nach Bonforoni-Anpassung (p-Wert signifikant ab
<0,017) einen signifikanten Unterschied (p-Wert 0,016221) zwischen den Gruppen 1
(0-10 kg) und Gruppe 3 (20-40 kg).
0-10kg 10-20kg 20-40kg
Gewichtsklassen
0,05
0,06
0,07
0,08
0,09
0,10
0,11
0,12
0,13
0,14
0,15
0,16
0,17
0,18
Mitte
lwert
e d
er
Pero
xis
om
en/ µ
m²
Flä
che
Abbildung 3-46 Mittelwerte der Peroxisomen/Hepatozytenfläche (µm²) in Bezug zu den Gewichtsklassen

4 Diskussion 123
4 Diskussion
Ziel der vorliegenden Arbeit war die vergleichende lichtmikroskopische und
elektronenmikroskopische Untersuchung von caninen Hepatozyten im Hinblick auf
morphologische und quantitative Unterschiede der Peroxisomen bei Tieren mit
primären degenerativen Lebererkrankungen sowie bei Tieren mit neoplastischen
Veränderungen in der Leber. Zur Diagnostik der Lebererkrankung wurden neben der
pathohistologischen Untersuchung der Leberbiopsien auch leberspezifische klinisch-
chemische Blutparameter untersucht, die in der Diagnostik von Lebererkrankungen
von Bedeutung sind.
4.1 Lichtmikroskopische Untersuchungen
Die geringgradige hepatozelluläre Leberzelldegeneration war die am häufigsten
histologisch gestellte Diagnose, gefolgt von der mittelgradigen hepatozellulären
Degeneration. Die Patienten der Gruppe 1 waren an extrahepatischen Tumoren,
Pankreatitis, IBD, Insuffizienz der Mitralklappe, Hypothyreose und Lungenfibrose
erkrankt. Die Patienten der Gruppe 2 zeigten als Grunderkrankung entweder einen
Primärtumor der Leber oder eine sekundäre Tumorerkrankung infolge einer
Metastasierung in die Leber. Der Charakter der Leberzelldegenerationen reichte in
Gruppe 1 von rein vakuolär (n=2), ballonierend (n=2), vakuolär-hydropisch (n=2) bis
zu rein hydropischer Degeneration (n=5), während Gruppe 2 einen Patienten mit
vakuolär-hydropischer Degeneration, drei Patienten mit rein vakuolärer Degeneration
und vier mit hydropischer Degeneration einschloss. Sicherlich können die einzelnen
Degenerationstypen (vakuolär, hydropisch, ballonierend, Mischformen) aufgrund des
unterschiedlichen Schweregrades nicht gleichgesetzt werden, jedoch lässt sich
feststellen, dass die Untersuchungsergebnisse Parallelen mit den Untersuchungs-
ergebnissen einer Studie von SEPESY et al. (2006) aufweisen, in denen 336
Patienten mit einer vakuolären Degeneration hinsichtlich der Grunderkrankung
untersucht wurden. Sie konnten beweisen, dass die vakuoläre Degeneration von

124 4 Diskussion
Hepatozyten, wie früher angenommen, nicht ausschließlich die Folge übermäßiger
endogener oder exogener Glukokortikoidwirkung ist, sondern zahlreiche
internistische Erkrankungen ursächlich dafür sein können (DE NICOLA et al. 2005;
SEPESY et al. 2006). Das Untersuchungsergebnis wird unterstützt durch die
Tatsache, dass lediglich aus Gruppe 1 je ein Patient (K1972) unter exogenem bzw.
endogenem Glukokortikoideinfluss (Morbus Cushing) (K1979) stand. Sekundär
bedingte Leberveränderungen mit abnormalen Leberenzymaktivitäten sind weitaus
häufiger als primäre Ursachen (DE NICOLA et al. 2005). Darüber hinaus ist der
Anstieg der Leberenzymaktivitäten nicht gleichzusetzen mit dem Schweregrad der
hepatischen Störung. Lediglich 10 % der vorgestellten Patienten mit erhöhten
Leberenzymaktivitäten haben tatsächlich eine primäre Lebererkrankung (DE NICOLA
et al. 2005). Das Leberenzym Alaninaminotransferase (ALT) steigt besonders bei
hepatozellulärer Nekrose, Entzündung (CENTER 2007) und hepatozellulärer
Degeneration (DIAL 1995) an. 37,5 % der Patienten von Gruppe 2 zeigten 5-10fache
ALT Wert Erhöhungen, während sogar 25 % über 10fache Erhöhungen aufwiesen.
Auffallend dabei ist, dass sich die histologischen Veränderungen nicht immer in der
Höhe der ALT Werte widerspiegeln. In Gruppe 1 zeigte Patient K2025 eine
hochgradige Degeneration der Hepatozyten bei gleichzeitig physiologischem ALT
Wert. In Gruppe 2 wies Patient K 2027 histologisch eine geringgradige vakuoläre
Degeneration der Hepatozyten auf, während die ALT um das >10fache erhöht war.
Die geringgradige Degeneration war somit nicht die Grundursache dieser
labordiagnostischen Veränderung, sondern eine Metastase eines
Plattenepithelkarzinoms, das vermutlich zu Nekrose von Leberzellen mit begleitender
Entzündung führte. Erstaunlicherweise zeigte Patient K2030 keine ALT Erhöhung,
ließ jedoch histologisch eine mittelgradige vakuoläre Degeneration in
Zusammenhang mit einem anaplastischen malignen Lymphom in der Leber
erkennen. Diese Beobachtungen decken sich mit der Studie von CENTER et al.
(2007), die ebenfalls feststellten, dass Tiere mit schweren pathologischen
Veränderungen normale ALT Werte aufweisen können. Die Arbeitsgruppe merkt an,
dass normale ALT Werte auch ein Hinweis sein können, dass kaum noch
lebensfähige Hepatozyten vorhanden sind oder aber die ALT schon wieder im

4 Diskussion 125
Absinken begriffen ist (CENTER 2007). Leider können über den Verlauf der
Laborwerte in dieser Studie keine Aussagen getroffen werden, da keine
Untersuchungen der Laborwerte in zeitlichem Verlauf vorgenommen wurden.
Ähnliche Veränderungen finden sich bei dem Enzym Glutamatdehydrogenase
(GLDH), welches ein mitochondriales Enzym darstellt (STOCKHAM u. SCOTT 2008).
Der überwiegende Anteil der Patienten der Gruppe 1 zeigte Werte im
Referenzbereich oder Werte, die unter einer fünffachen Erhöhung lagen. Patient
K2025 wies trotz hochgradiger hepatozellulärer Degeneration einen normalen GLDH
Wert auf. Gruppe 2 hingegen zeigte überwiegend Patienten (37,5 %) mit über
10facher GLDH Erhöhung und 25 % mit fünf bis zehnfacher GLDH Erhöhung. Durch
das Vorhandensein eines Tumors in der Leber als primäre Ursache kam es hier im
Gegensatz zur Gruppe 1 zu stärkeren Schädigungen des Lebergewebes, was sich in
erhöhten enzymatischen Aktivitäten niederschlug. Elektronenmikroskopisch fanden
sich ebenfalls in Gruppe 2 stärkere degenerative Veränderungen der Mitochondrien
in Form von Schwellung und lytischen Veränderungen als in Gruppe 1. Die GLDH
Erhöhungen deuten daraufhin, dass die Mitochondrien in besonderem Maße in ihrer
Funktion beeinträchtigt sind (KÄUFER-WEISS 2007), da das Enzym ansonsten nicht
in dem Ausmaß im Serum messbar wäre. Die Mitochondrien können eine
Schädigung durch Hypoxie, freie Radikale und Toxinbelastung erfahren (VIEIRA u.
KROEMER 1999). Neben den vielseitigen Aufgaben, tragen die Mitochondrien aber
auch selbst zur Radikalbildung bei (ROTH 1997). O2- und H2O2 reagieren häufig mit
Eisen, das sich in den Mitochondrien befindet. Dadurch kann die mitochondriale DNA
geschädigt werden. Die Folge dieser Schädigung ist eine Verstärkung des oxidativen
Stresses aufgrund der Expression veränderter Proteine, die für die Elektronen-
transportkette verantwortlich sind. Es entsteht ein Circulus vitiosus mit verstärkter
Radikalbildung (LEE u. WEI 1997).
Die höchsten ALKP (Alkalische Phosphatase) Erhöhungen fanden sich in der Gruppe
1 zu 35,4 % mit geringgradigen bis mittelgradigen Erhöhungen. Allein 25 % der
Patienten von Gruppe 2 zeigten hochgradige ALKP Wert Erhöhungen. Die höchsten,
also bis zu 100facher ALKP Erhöhung finden sich bei diffusen oder fokalen
cholestatischen Veränderungen, beim hepatozellulären Karzinom, Gallengangs-

126 4 Diskussion
karzinom und Patienten mit einer Glukokortikoidbehandlung oder endogener
Exposition (CENTER 2007). Erstaunlicherweise zeigten in dieser Studie gerade die
Patienten der Gruppe 2 eine normale (K2011) bzw. eine bis zu fünffache (K2009)
Erhöhung des Referenzwertes, die histologisch eine mittelgradige Cholestase
aufwiesen. Eine mögliche Erklärung könnte sein, dass es sich bei dem Pigment doch
um Lipofuscin oder Hämosiderin handelte, da es schwierig ist, dies ohne
Spezialfärbungen nachzuweisen. Patient K2009 erkrankte an einem
Hämangiosarkom der Leber und Patient K2011 an einem Klarzellkarzinom. Der
Patient K1968 zeigte wider Erwarten lediglich eine bis zu fünffache
Referenzwerterhöhung, wobei gerade bei Karzinomen deutlich höhere Werte zu
finden sein können (CENTER 2007).
Die Gruppe 1 zeigte bei drei Patienten histologisch mittelgradige Cholestasen, auch
hier waren die ALKP Werte wider Erwarten normal (K2063) oder nur bis zum
fünffachen Wert erhöht (K2062 und K2015). Lediglich die Patienten, die unter
endogenem (K1979) und exogenem (K1972) Kortisoneinfluss standen, zeigten eine
um das fünf- bis zehnfache Erhöhung des Referenzwertes. Wobei hier natürlich
durchaus höhere ALKP Werte (bis zum 100fachen des Referenzwertes (CENTER
2007) erwartet werden können. Patient K1979 wurde schon mit Trilostan behandelt,
sodass deshalb der ALKP Wert niedriger ausfiel und Patient K1972 erhielt sechs
Monate vor der Biopsieentnahme die letzten Kortisongaben; aufgrund zu starker
Nebenwirkungen wurde die Gabe abgebrochen. Der dennoch relativ hohe ALKP
Wert kann, trotz fehlender Kortisongabe, dadurch erklärt werden, dass dieser Hund
chronisch an einer „Inflammatory bowel disease“ (IBD) und Futtermittelallergie
erkrankt war. Bei derartigen Patienten können erhöhte ALKP Werte durch
endogenen chronischen Stress erzeugt werden (CENTER 2007). Die Gallensäuren
wurden routinemäßig bestimmt. Sie zeigten überwiegend keine Auffälligkeiten,
sodass sie hier nicht weiter diskutiert werden.
Auffallend in dieser Arbeit war das übermäßige Vorkommen von sekundären
Lebertumoren (n=6) im Gegensatz zu primären Lebertumoren (n=2). Die primären
Tumoren zeigten begleitend je eine mittelgradige und eine geringgradige
Leberzelldegeneration, während in der Gruppe der sekundär bedingten Tumoren drei

4 Diskussion 127
Patienten begleitend eine geringgradige, zwei eine mittelgradige zeigten und ein
Patient eine hochgradige Degeneration aufwies. Eine mögliche Erklärung für den
unterschiedlichen Ausprägungsgrad ist sicher die Dauer der Erkrankung, jedoch
auch die Lokalisation der Biopsieentnahme des annähernd gesunden Gewebes der
Leber. Der Umstand, dass in dieser Arbeit mehr Patienten mit einem sekundär
bedingten Tumor untersucht wurden ist darin begründet, dass die Leber neben der
Lunge ein Organ ist, in dem sich bevorzugt Metastasen ansiedeln (CULLEN 2009),
da sie ein sehr stark vaskularisiertes Organ ist (CHEVILLE 1994d; STALKER u.
HAYES 2007). Metastatische Neoplasien sind bei Hunden die am häufigsten
vorkommenden Lebertumoren und entstammen zumeist dem Pankreas, dem
Gastrointestinaltrakt, der Milz, der Milchleiste, den Nebennieren, den Knochen und
der Schilddrüse (HOSKINS 2005).
Letztlich müssen noch die minimalen Veränderungen der klinisch gesunden
Kontrolltiere besprochen werden. Histologisch fanden sich in den histologischen
Proben minimale degenerative Veränderungen. So können die minimalen
geringgradigen Degenerationen der Hepatozyten als Reaktion auf die Hypoxie
betrachtet werden (KÄUFER-WEISS 2007), die im Rahmen einer Allgemein-
anästhesie, wie bei den Kontrolltieren vorliegend eintreten können.
4.2 Elektronenmikroskopische Untersuchung
Für die vorliegende elektronenmikroskopische Untersuchung standen 19
Leberbiopsien von Patienten der Kleintierklinik des Tierärztlichen Institutes der
Georg-August-Universität Göttingen zur Verfügung. Gruppe 1 mit degenerativen
Lebererkrankungen umfasste elf Patienten. Die Gruppe 2 umfasste acht Patienten
mit einem Primär- oder Sekundärtumor der Leber. Als Kontrollgruppe dienten jeweils
3 Leberbiopsien unterschiedlicher Lokalisation von zwei Hunden der Rasse Beagle,
die als klinisch gesund eingestuft wurden. Sämtliche Leberproben wurden an
repräsentativen Lokalisationen hinsichtlich morphologischer Veränderungen der
hepatozellulären Peroxisomen sowie auf deren quantitative Unterschiede untersucht
und zwischen den Gruppen verglichen.

128 4 Diskussion
4.2.1 Quantitative Erfassung der Peroxisomen/Hepatozyt- Fläche
Aufgrund der physiologischen Eigenschaften der Peroxisomen kann angenommen
werden, dass bei primären degenerativen Lebererkrankungen (Gruppe 1) die Anzahl
der Peroxisomen zunimmt. Bei Untersuchungen aus der Humanmedizin an
degenerativ veränderten Hepatozyten, begleitet von steatotischen Veränderungen,
konnte eine Proliferation mit Verringerung der peroxisomalen Größe festgestellt
werden (DE CRAEMER et al. 1995). Auch ein an Lipodystrophie erkrankter Junge
zeigte bei milder Lipidakkumulation eine Steigerung der Peroxisomenanzahl pro
Hepatozyt (DE CRAEMER et al. 1992). Degenerative Veränderungen der
Hepatozyten haben vielfältige Ursachen wie z.B. die Einwirkung verschiedenster
Xenobiotika und Umweltgifte, aber auch hypoxische Zustände mit der Bildung von
freien Radikalen als Folge des oxidativen Stresses. Bei all diesen pathologischen
Veränderungen hat das Peroxisom die Hauptaufgabe die schädlichen Substrate
durch den Peroxidmetabolismus und der damit verbundenen Aktivität der Katalase
zu neutralisieren und weiteren Zellschaden zu verhindern (MANNAERTS u. VAN
VELDHOVEN 1992; SCHRADER u. FAHIMI 2004). In der vorliegenden Arbeit konnte
jedoch keine signifikante Zunahme der Peroxisomenzahl im Vergleich zu den
gesunden Kontrolltieren (Gruppe 3) gefunden werden. Zwischen Gruppe 1 (primäre
hepatozelluläre Degeneration) und Gruppe 2 (Tiere mit Tumoren) waren ebenso
keine signifikanten Unterschiede feststellbar.
Die vorhandene veterinärmedizinische Literatur zu dieser Fragestellung ist sehr
gering. Eine mögliche Erklärung findet sich bei CENTER et al.(1993). Die
Arbeitsgruppe untersuchte Katzen, die an einer Lipidose erkrankt waren. Die Anzahl
der hepatozellulären Peroxisomen der an der Lipidose erkrankten Katzen war wider
Erwarten deutlich verringert, sodass CENTER et al. (1993) vermuteten, dass die
Hepatozyten nicht in der Lage waren neue Peroxisomen zu produzieren und die
Funktion der Organelle aufrechtzuerhalten. Es bestand die Annahme, dass ein
Feedbackmechanismus existiert, der die Biogenese neuer Peroxisomen unterbindet.
Weiterhin bestand die Vermutung, dass die Verminderung der Peroxisomen dem
Krankheitsbild der hepatischen Lipidose zu zuordnen war oder aber die Peroxisomen

4 Diskussion 129
aufgrund peroxisomaler Dysfunktion der Grund der hepatischen Lipidose sein
konnten.
Eine weitere mögliche Erklärung für die ausgebliebene Erhöhung der Peroxisomen in
Gruppe 1 könnte in der Qualität bzw. der Kettenlänge der Fette liegen, die in den
Hepatozyten akkumulieren. Im Rahmen der ß-Oxidation können durch Peroxisomen
lediglich sehr lange oder auch toxische Fettsäuren abgebaut werden (REDDY u.
MANNAERTS 1994; MASTERS u. CRANE 1995). Die Untersuchung der Qualität der
Fette in den Hepatozyten war jedoch nicht Gegenstand dieser Dissertation, wäre
jedoch sicherlich interessant für weiterführende Untersuchungen.
Die ausgebliebene Proliferation von Peroxisomen der Gruppe 1 könnte darüber
hinaus mit dem Prozess der Autophagie in Zusammenhang stehen. Autophagische
Prozesse sind katabole intrazelluläre Abläufe, die letztendlich in einen lysosomalen
Abbau von Zellorganellen münden. Die Autophagie nimmt eine wichtige Stellung ein,
zum Beispiel in Hepatozyten, indem sie die Zelle durch den Abbau von
intrazellulären Materialien mit Nährstoffen versorgt, aber auch nicht mehr benötigte
Zellorganellen abbaut (CUERVO 2004a; MIZUSHIMA 2005). Möglicherweise
unterliegen einige der Peroxisomen aufgrund einer starken Beanspruchung der Zelle
bereits der Autophagie, sodass sie der quantitativen Erfassung nicht mehr zur
Verfügung standen.
Bei der Diskussion um die ausgebliebene Peroxisomenproliferation sollte auch die
Methodik dieser Dissertation beleuchtet werden. Zum einen erfolgte die Auswahl der
Hepatozyten in Anlehnung an eine humanmedizinische Untersuchung, unabhängig
von der periportalen, mittzonalen oder zentralen Ausrichtung, da in dieser
Untersuchung mittels Katalaseanfärbung der Peroxisomen gezeigt werden konnte,
dass es keine bevorzuge periportale oder zentrolobuläre Anhäufung von
Peroxisomen gibt (ROELS et al. 1983). Es ist jedoch nicht auszuschließen, dass der
Hund gänzlich anders veranlagt ist und bei dieser Spezies durchaus eine regionale
hepatozelluläre Häufung von Peroxisomen besteht. Darüber hinaus könnten z. B.
Proliferationsvorgänge der Peroxisomen vor allem zentrolobulär stattgefunden
haben. Andererseits wurden, um ein vergleichbares Verhältnis von Fläche und

130 4 Diskussion
Anzahl der Peroxisomen zu gewährleisten, bei allen Patienten der Gruppe 1-3
ähnliche Hepatozyten ausgewählt. Diese Auswahl sollte eine Konstanz und
Homogenität der untersuchten Hepatozyten ermöglichen.
Fetttropfen innerhalb des Hepatozyten wurden nicht von der Gesamtfläche
abgezogen, da dies laut DE CRAEMER et al. (1995) nicht notwendig sei, weil sich
dabei keine Unterschiede bei der Zählung von Zellorganellen ergeben.
Fünf Patienten der Gruppe 1 erkrankten an einer geringgradigen Degeneration der
Hepatozyten, vier Patienten an einer mittelgradigen Degeneration und zwei an einer
hochgradigen Degeneration. In der Gruppe 2 zeigten vier Patienten eine
geringgradige und drei Patienten eine mittelgradige Degeneration der Hepatozyten,
während ein Patient eine hochgradige Degeneration aufwies. Die Kontrolltiere
(Gruppe 3) zeigten minimale degenerative Veränderungen. Die Patienten mit
hochgradigen degenerativen Veränderungen zeigten keine höheren durch-
schnittlichen Peroxisomenzahlen je µm² Hepatozytenfläche als Fälle mit gering- oder
mittelgradigen Degenerationen, was eigentlich die Erwartung gewesen wäre. Die
breite Variation des Schweregrades der degenerativen Veränderungen könnte eine
weitere Erklärung dafür sein, dass die durchschnittliche Peroxisomenzahl in beiden
Gruppen nicht zugenommen hat. Es wäre möglich, dass bei gering- bis
mittelgradigen Degenerationsvorgängen der Impuls zur Biogenese von Peroxisomen
zu gering ist oder die Anzahl der vorhandenen Peroxisomen mit der darin
befindlichen Katalase und der Kapazität zur ß-Oxidation ausreicht, um den zellulären
Schädigungen entgegenzuwirken. Darüber hinaus müssten in einer Folgestudie
deutlich mehr Patienten integriert werden, die an einer hochgradigen
hepatozellulären Degeneration leiden, um statistisch die Aussagen besser absichern
zu können.
Im Rahmen der Diskussion der Ergebnisse der vorliegenden Arbeit müssen auch
speziesspezifische Unterschiede berücksichtigt werden. Die Gabe von als
Peroxisomenproliferatoren wirkenden hypolipidämisch wirksamen Medikamenten
führt beim Hund und Primaten, eingeschlossen der Mensch zu keiner
Peroxisomenproliferation im Gegensatz zu Ratten (DE LA IGLESIA et al. 1981). Die

4 Diskussion 131
unterschiedliche Reaktion der Säugetiere auf die einzelnen Peroxisomen-
proliferatoren korreliert mit der Expression des PPAR α (KLAUNIG et al. 2003;
PETERS et al. 2005). Die schwache Antwort auf Peroxisomenproliferatoren zur
Indukution des PPAR α Rezeptors bei Nicht-Nagern, wie dem Hund, liegt in einer
unterschiedlichen Aktivierung der PPAR α Transkription begründet (BOSGRA et al.
2005). Die Gabe von PPAR α Agonisten verursacht bei der Ratte eine erhöhte
Aktivität der Enzyme zur ß-Oxidation der Mitochondrien und der Peroxisomen,
während beim Hund nur die mitochondriale ß-Oxidationsaktivität gesteigert wird.
Daher führen beim Hund vermehrt anfallende Fettsäuren bedingt durch Behandlung
mit PPAR α Agonisten zu einer erhöhten Aktivität der Mitochondrien, da eine
peroxisomale Proliferation ausbleibt. Es kommt zu einer Mehrbelastung der
Mitochondrien (GUO et al. 2007). Demnach könnte es sein, dass der Hund auf
Einlagerung von Fetttropfen, wie wir sie besonders bei den Patienten K1972/P513/06
(ggr. Leberzelldegeneration) und K1974/P539/07 (hgr. Leberzelldegeneration) mit
mittelgradigen gemischttropfigen hepatozellulären Verfettungen sehen, gar nicht mit
einer Peroxisomenproliferation reagieren kann.
Humanmedizinische Untersuchungen hinsichtlich der Anzahl der Peroxisomen in
Hepatozyten bei tumorösen Erkrankungen der Leber fallen nicht einheitlich aus. Die
konträren Ergebnisse liegen in der Wahl der zu untersuchenden Zelltypen. Werden
die Peroxisomen in stark veränderten Tumorzellen untersucht, sind aufgrund der
Entdifferenzierung der Zellen infolge neoplastischer Veränderungen auch weniger
Organellen, insbesondere Peroxisomen, vorhanden. Sind gesunde oder
degenerierte, dem Tumor benachbarte Hepatozyten Gegenstand der Untersuchung,
so ist davon auszugehen, dass die Organellen wesentlich besser erhalten sind und
gegebenfalls zunehmen. In dieser Studie wurden ausschließlich gut erhaltene
Hepatozyten untersucht. Die Auswahl von annähernd gesund erscheinenden
Hepatozyten für die elektronenmikroskopische Untersuchung, anstatt der tumorös
veränderten Zellen, stellt eine mögliche Erklärung dar, warum die Zahl der
Peroxisomen nicht signifikant verändert ist.
Aufgrund ihrer Untersuchungen kamen BERNHARD u. ROUILLER (1956) zu der
Erkenntnis, dass eine Färbung der Peroxisomen zur eindeutigen Identifikation nicht

132 4 Diskussion
notwendig sei. Daher erfolgte in dieser Arbeit ebenfalls keine Anfärbung. DE
CRAEMER (1995) vertritt ebenfalls die Ansicht, dass sich die Peroxisomen mittels
Elektronenmikroskopie eindeutig in ihrer Morphologie zuverlässig von anderen
Zellorganellen unterscheiden lassen, obwohl die Gefahr bestand, die Zahl der
Peroxisomen zu überschätzen oder zu unterschätzen. Da jedoch in der vorliegenden
Arbeit einige Leberproben nicht die optimale erwünschte Qualität für eine
elektronenmikroskopische Untersuchung aufwiesen, kann nicht ausgeschlossen
werden, dass einige Peroxisomen nicht als solche erkannt wurden. Aufgrund dieser
Erfahrungen ist entgegen der Ansicht von DE CRAEMER (1995) bzw. BERNHARD
u. ROUILLER (1956) eine Katalasefärbung zur Identifikation von Peroxisomen
gerade in stark beeinträchtigten bzw. nicht gut erhaltenen Gewebeproben
ausdrücklich empfehlenswert.
In den Leberbiopsien der Kontrollpatienten fielen histologisch minimale Anzeichen
einer hydropischen Schwellung der Hepatozyten auf. Die Patienten waren 365 Tage
alt, lebten in stabilen Kleingruppen und bekamen ein standardisiertes
Feuchtfuttermittel. Die Hunde zeigten bei der klinischen Allgemeinuntersuchung
keine Auffälligkeiten, wurden aber vor der Biopsieentnahme narkotisiert. Die Narkose
mit begleitender Hypoperfusion des Organismus und der damit verbundenen
Hypoxie, insbesondere im Lebergewebe, könnte ein Grund für die minimalen
hepatozellulären Degenerationserscheinungen sein. Klinisch chemische
Untersuchungsergebnisse der Leberenzyme lagen für diese Tiere leider nicht vor.
Die Tatsache, dass vergleichsweise junge Hunde bereits derartige Veränderungen
zeigen, lässt die Frage aufkommen, ob es überhaupt möglich ist, vollständig gesunde
Hepatozyten für eine derartige Untersuchung unter Narkose zu gewinnen. Allgemein
bleibt festzustellen, dass die Leber auf alle systemischen Einflüsse des Körpers
reagiert, also auch auf nicht hepatische Störungen, die in der Folge die Funktion der
Hepatozyten beeinflussen (CENTER 2007).

4 Diskussion 133
4.2.2 Quantitative Erfassung der Peroxisomen/Hepatozyt-
Fläche/Alter
Da sich zwischen den Untersuchungsgruppen keine statistisch signifikanten
Unterschiede der Peroxisomenzahl/µm² Hepatozyt ergaben und die Patientenzahl
pro Untersuchungsgruppe für eine Einteilung in unterschiedliche Altersstufen
innerhalb der Gruppe zu niedrig war, wurden sämtliche untersuchten Tiere aus allen
Gruppen zusammengefasst und in drei Altersklassen (1-5 Jahre, 6-10 Jahre, 11-15
Jahre) eingeteilt, um mögliche altersabhängige Unterschiede der
Peroxisomenanzahl/µm² Hepatozyt zu ermitteln (Anhangstabelle 10-12). Die
statistische Analyse ergab jedoch keine signifikanten Unterschiede zwischen den
verschiedenen Altersklassen. Es zeigte sich das Tiere im Alter von 11-15 Jahre
durchschnittlich etwas mehr Peroxisomen/µm² Hepatozyt (0,12) aufwiesen als
jüngere Tiere (0,10 Peroxisomen/µm² Hepatozyt). Die Tendenz der Peroxisomen,
verstärkt in alternden Zellen aufzutreten, deckt sich mit einer humanmedizinischen
Untersuchung. Es wurde festgestellt, dass alternde Fibroblasten, deren
peroxisomaler Enzymgehalt z. B. Katalase absinkt, diesem Zustand mit einer
Peroxisomenproliferation begegnen (LEGAKIS et al. 2002). Es handelte sich bei
dieser Untersuchung zwar um Fibroblasten, jedoch ist nicht auszuschließen, dass
dieses Phänomen auch für canine Hepatozyten zutrifft. Für weiterführende
Untersuchungen sollten in den Gruppen 1-3 deutlich größere Patientenzahlen in den
Altersgruppen gewählt werden, um statistisch aussagekräftigere Untersuchungen
machen zu können.
4.2.3 Quantitative Erfassung der Peroxisomen/Hepatozyt-
Fläche/Rasse
Die Hepatozyten der kleinen Rassen (0-10 kg) zeigen signifikant mehr
Peroxisomen/µm² Hepatozytenfläche als die Tiere, die den Gewichtsklassen 10-20
kg und 20-40 kg angehörten. Für dieses Phänomen liegen in der Literatur keine
Hinweise vor. Wird in die Betrachtung der Gewichtsklasse (0-10 kg) das Alter
einbezogen, gehören zwei Patienten der Altersgruppe 6-10 Jahre an und fünf

134 4 Diskussion
Patienten fallen in die Gruppe 11-15 Jahre. Sowohl die Gewichtsklasse 10-20 kg als
auch die Gewichtsklasse 20-40 kg weisen je drei Patienten mit einem Alter von 6-10
Jahren auf. Zwei Patienten der Gewichtsklasse 10-20 kg sowie vier Patienten der
Gewichtsklasse 20-40 kg haben ein Alter von 11-15 Jahren erreicht.
So hatte die vermehrte Anzahl der Peroxisomen in den Hepatozyten wahrscheinlich
weniger mit der Rasse als mit dem Alter zu tun. Aus Untersuchungen in der
Humanmedizin ist bekannt, dass auch Fibroblasten mit zunehmendem Alter mit einer
Peroxisomenproliferation reagierten (LEGAKIS et al. 2002).
4.2.4 Morphologische Untersuchungen
Bei der morphologischen Untersuchung wurden die Peroxisomen anhand ihrer
Größe (klein/groß), ihres Verteilungsmusters innerhalb der Zelle sowie ihrer
morphologischen Veränderungen (Protrusion, Marginalplatte, Nukleoid) beurteilt.
Peroxisomengröße
In Gruppe 1 und 2 konnte aufgrund der vorhandenen degenerativen sowie der
Lokalisation der Hepatozyten in unmittelbarer Nachbarschaft zu neoplastischen
Prozessen, dem teilweise Vorhandensein von Fetttropfen und dem aus den
degenerativen Veränderungen für die Zelle resultierenden oxidativen Stress, erwartet
werden, dass dies eine Proliferation von Peroxisomen bei gleichzeitiger
Verkleinerung der Peroxisomengröße zur Folge hatte.
Im Rahmen von Proliferationsvorgängen verringern Peroxisomen ihre Größe,
während ihre Gesamtzahl zunimmt (DE CRAEMER 1995).
In der vorliegenden Untersuchung konnten jedoch keine signifikanten Unterschiede
in den einzelnen Gruppen 1-3 im Hinblick auf das Vorkommen kleiner Peroxisomen
festgestellt werden. Da es sich hierbei um eine semiquantitative Auswertung
handelte, konnte im vorliegenden Fall keine Aussage über absolute Zahlen getroffen
werden, aber es liegt die Vermutung nahe, dass in den untersuchten Zellen keine
Proliferation stattgefunden hatte. Diese Feststellung wurde unterstützt durch die
Tatsache, dass bei Betrachtung der Gruppen 1-3 ein signifikanter Unterschied

4 Diskussion 135
bestand, was das Auftreten von großen Peroxisomen betraf. Große Peroxisomen
fanden sich in Zellen, in denen keine Peroxisomenproliferation stattgefunden hatte.
Verteilungsmuster in der Zelle
Hinsichtlich der statistischen Auswertung der Verteilung der Peroxisomen in vier
Merkmalen (Nähe Fettropfen, Cluster, panzytär, perinukleär) ergab sich lediglich in
einem Merkmal (Cluster) ein signifikantes Ergebnis. Gruppe 1 zeigte das Auftreten
der Peroxisomen in Form eines Clusters zu 36,4 %, während Gruppe 2 das Merkmal
zu 25 % aufwies und Gruppe 3 zu 100 %. Der Unterschied erwies sich zwischen
allen drei Gruppen als signifikant p 0,01233 (p-Wert signifikant ab p <0,05).
Keine der Gruppen 1-3 zeigte ein perinukleäres Auftreten von Peroxisomen, sodass
auch diese Untersuchung die Annahme unterstützt, dass keine
Proliferationsvorgänge in den untersuchten Hepatozyten stattgefunden hatten.
Unsere Untersuchungen werden von ROELS et al. (1983) und DE CRAEMER et al.
(1991) bestätigt, die ebenfalls bei Proliferationsvorgängen und in pathologisch
veränderten Lebern eine perinukeäre Verteilung der Peroxisomen fanden. In der
Kontrollgruppe der vorliegenden Studie lagen die Peroxisomen zu 100 % als Cluster
vor, was durch die Untersuchung von ROELS et al. (1983) bestätigt wird, der
ebenfalls bei gesunden Lebern eine homogene Verteilung der Peroxisomen
beobachtet oder die Ansammlung in Form von Gruppen (Cluster). Die degenerativ
veränderten Hepatozyten (Untersuchungsgruppe 1 und 2) zeigten deutlich weniger
Peroxisomen in Form von Clustern. Die Beobachtung, dass Peroxisomen sich
insbesondere in der Nähe von Fetttropfen positionierten (SCHRADER 2001) ließ sich
in der vorliegenden Studie nicht bestätigen, was sicherlich daran lag, das nur zwei
Patienten K1972/P513/06 (ggr. Leberzelldegeneration) und K1974/P539/07 (hgr.
Leberzelldegeneration) hepatozelluläre Verfettungen im Untersuchungsgut
aufwiesen.
Morphologische Veränderungen (Protrusion, Marginalplatte, Nukleoid)
Bei der statistischen Auswertung mit Hilfe des Pearson Chi-Quadrat Testes konnten
in keinem Fall signifikante Unterschiede zwischen den Untersuchungsgruppen

136 4 Diskussion
ermittelt werden (Protrusion p-Wert 0,40036; Marginalplatte p-Wert 0,16792;
Nukleoid p-Wert 0,68960 signifikant ab p<0,05000).
Protrusion:
Das Auftreten von Protrusionen findet man insbesondere bei degenerativen
Veränderungen der Hepatozyten in Zusammenhang mit Steatose, Hepatitiden und
auch Tumorerkrankungen (DE CRAEMER 1995). Die Formveränderungen findet
man ebenfalls bei Proliferationsvorgängen der Peroxisomen. Sie führen zusätzlich zu
einer Oberflächenvergrößerung, die einen erhöhten peroxisomalen Metabolismus
ermöglichen (ROELS et al. 1991). Auffallend ist, dass die Kontrolltiere annähernd so
viele Peroxisomen aufweisen wie die Gruppen 1-2. Eine mögliche Erklärung stellt
neben der Lokalisation der Entnahme der Leberprobe auch der Zeitpunkt im
Krankheitsverlauf dar, sodass zu Beginn einer degenerativen Veränderung der
Impuls zur Peroxisomenproliferation und damit einhergehender Formveränderung
stärker ausfällt als im späteren Verlauf und mit zunehmender Dauer der Erkrankung
abnimmt, gänzlich schwindet oder auf einem Niveau bleibt. Leider liegen über die
Dauer der einzelnen Erkrankungen der Patienten keine Daten vor.
Marginalplatte und Nukleoid:
Die Marginalplatte als elektronendichte Verdickung unter der Peroxisomenmembran
und das Nukleoid als elektronendichter Kern zentral im Peroxisom zeigten sich bei
allen drei Untersuchungsgruppen in nahezu ähnlichem Umfang. Eine Untersuchung
an Katzen, die entweder an einer Lipidose erkrankt waren oder an einer
Gallengangsobstruktion, zeigte, dass bei der erstgenannten Gruppe weder
Marginalplatten noch Nukleoide und bei den Patienten, die an einer
Gallengangsobstruktion litten, beide morphologischen Merkmale vorhanden waren
(CENTER et al. 1993). Die Arbeitsgruppe vermutete, dass das komplette
Verschwinden der morphologischen Merkmale direkt mit dem Krankheitsbild der
Lipidose verbunden ist (CENTER et al. 1993). Bei der quantitativen Erfassung sollte
ebenso berücksichtigt werden, dass das Vorhandensein des Nukleoids abhängig von
der Schnittebene des Präparates ist (DE DUVE u. BAUDHUIN 1966).

4 Diskussion 137
Degenerative Veränderungen, wie sie bei Gruppe 1 und 2 in unterschiedlichem
Schweregrad auftraten, scheinen demnach nicht mit Veränderungen der
Marginalplatte oder dem Nukleoid einherzugehen.
Mitochondriale Eigenschaften
Die Mitochondrien wurden hinsichtlich des Auftretens von Schwellungen, lytischen
Erscheinungen, parakristallinen Einschlüssen und dem Auftreten von
Riesenmitochondrien untersucht. Es bestand kein signifikanter Unterschied in den
vier untersuchten Merkmalen zwischen den Gruppen 1-3. Eine Schwellung zeigten
nahezu 100 % der Gruppe 3 und in etwa gleichen Anteilen Gruppe 1 und 2 (60-70
%). Hinsichtlich der lytischen Veränderungen zeigten beinahe 80 % der Gruppe 2
dieses Merkmal, während dies bei ungefähr 50 % der Gruppe 1 und 3 zutraf.
Parakristalline Einschlüsse fanden sich bei knapp 40 % der Gruppe 1 und 2 sowie
bei knapp 20 % der Gruppe 3. Die Riesenmitochondrien traten zu 80-90 % in Gruppe
1 und 3 auf, während Gruppe 2 zu etwa 50 % das Merkmal aufwies.
Zu den mitochondrialen Veränderungen bei beeinträchtigter Zellhomöostase gehören
Schwellung und Lyse der Mitochondrien, parakristalline Einschlüsse und die Bildung
von Riesenmitochondrien (CHEVILLE 1994c). Mitochondriale Veränderungen
werden dann als bedeutsam betrachtet, wenn sie weitläufig, diffus und immer
wiederkehrend auftreten (CENTER et al. 1993). Allerdings muss einschränkend
gesagt werden, dass mitochondriale Veränderungen in einer Vielzahl von
humanmedizinischen Lebererkrankungen auftreten und diese als unspezifisch
betrachtet werden (PHILLIPS et al. 1987). Veränderungen in Form von
Schwellungen der Mitochondrien fanden sich ebenfalls in tumorös als auch an den
Tumor angrenzenden Lebergeweben (GHADIALLY u. PARRY 1966). Insbesondere
die Gruppen 1 und 2 zeigten überwiegend histologisch geringgradige und
mittelgradige sowie wenige hochgradige Degenerationen der Hepatozyten.
Die Interpretation der elektronenmikroskopischen Bilder muss immer unter dem
Gesichtspunkt der Fixationsmethode betrachtet werden, denn Glutaraldehyd hat
einen negativen Einfluss auf Membranen, insbesondere auf die sehr empfindlichen

138 4 Diskussion
mitochondrialen Membranen, sodass es leicht zu Artefakten kommen kann
(CHEVILLE 1994c).
Die mitochondrialen Größenveränderungen lassen sich nicht in den Hämalaun-Eosin
gefärbten Paraffinschnitten erkennen. Dagegen werden sie in den
Semidünnschnitten, in Form von intrahepatozellulären kleinen weißen Punkten auf
„blauem Hintergrund“ deutlich. CENTER et al. (1993) beobachteten in ihrer
Untersuchung bei den an Lipidose erkrankten Katzen ebenfalls eine starke
Größenzunahme der Mitochondrien, die sie als Kompensationsmechanismus für eine
verminderte Anzahl an Mitochondrien betrachten.
Lipidtropfen, Glykogengehalt und Myelinfiguren in den Hepatozyten
In Gruppe 1 konnten tendenziell die meisten Lipidtropfen nachgewiesen werden. 30
% der untersuchten Hepatozyten zeigten entsprechende Lipidgehalte. In Gruppe 2
zeigte die Mehrheit der Patienten überhaupt keine Lipidtropfen (62,5 %). Die
Kontrollgruppe zeigte minimale Anzeichen einzelner kleiner Lipidtropfen.
Hepatozelluläre Degenerationen sind nicht entzündliche Veränderungen der
Hepatozyten, bedingt durch eine Hypoxie oder allgemeine Stoffwechselstörungen
(KÄUFER-WEISS 2007). Neben der Einlagerung von Wasser (KÄUFER-WEISS
2007) kann es auch zur Anreicherung von Lipiden oder Glykogen kommen
(STALKER u. HAYES 2007; CULLEN 2009). Die Kontrollgruppe zeigte deutlich mehr
Lipidtropfen als die Gruppe 2. Ein mögliche Erklärung liegt in einer prähepatischen
Ursache wie z.B. Hungern (STALKER u. HAYES 2007). Die Tiere insbesondere der
Gruppe 3 wurden vor der Narkose 24h nüchtern belassen. Bei Gruppe 2 konnten die
Tiere nicht in allen Fällen nüchtern gelassen werden, da es sich um Notfälle
handelte, die der direkten Behandlung (Operation) bedurften. Eine weitere mögliche
Erklärung wäre eine Reduktion von Enzymen zur ß-Oxidation in den Peroxisomen
der Kontrollgruppe. Um dies beweisen zu können, müsste man in einer
Folgeuntersuchung die peroxisomalen Enzyme quantitativ bestimmen.
Die Gruppen 1 und 3 zeigten die größten Mengen an Glykogeneinlagerungen in den
untersuchten Hepatozyten. Mögliche Erklärungen finden sich in Gruppe 1 im Hinblick

4 Diskussion 139
auf Grunderkrankungen der untersuchten Patienten. So litt Patient K1972 an einer
Futtermittelunverträglichkeit/Allergie und bekam regelmäßig Glukokortikoide iatrogen
verabreicht. Der Patient mit K1979 war an einem Morbus Cushing erkrankt, sodass
er unter endogenem Glukokortikoideinfluss stand. Beide Patienten zeigten bei der
Bestimmung der klinisch-chemischen Blutparameter hochgradige ALT und ALKP
Erhöhungen (Anhangstabelle 10-1). Die Ursache der starken Glykogeneinlagerung
bei den Kontrolltieren könnte darin begründet liegen, dass es sich um gesunde Tiere
handelte, die keine Anorexie im Vorfeld aufwiesen, während die anderen Patienten
teilweise Anzeichen einer Anorexie als klinisches Begleitsymptom ihrer Erkrankung
zeigten. Die Hepatozyten der Gruppe 2, die an den Tumor angrenzend lagen,
zeigten überwiegend eine gering- bis mittelgradige Einlagerung von Glykogen.
Myelinfiguren als Einschlüsse in Mitochondrien oder Autophagosomen
repräsentieren möglicherweise pathologische Konformationen von Membran-
proteinen und Lipoproteinschichten (CASTEJON u. CASTEJON 2008). Die
statistische Auswertung aller drei Untersuchungsgruppen ergab kein signifikantes
Ergebnis. Die Gruppe 2 zeigte in der elektronenmikroskopischen Untersuchung zu
37,5 % in mittelgradiger und zu 25 % Myelinfiguren in hochgradiger Ausprägung.
Myelinfiguren entstehen durch die Degeneration des Endoplasmatischen Retikulums,
aber auch durch andere Organellen. Die Membranen kollabieren, aggregieren und
formieren sich neu zu „Myelinfiguren“. Diese finden sich frei im Zytosol (CHEVILLE
1994c, b). Die untersuchten Hepatozyten der Gruppe 2 zeigten besonders auffällig
degenerative Veränderungen der Mitochondrien. Damit geht das starke Auftreten der
Myelinfiguren einher, so dass davon auszugehen ist, dass die Hepatozyten der
Gruppe 2 in dem Tumor angrenzenden Gebiet deutlich stärker degeneriert und in
ihrer Funktion beeinträchtigt sind als die Hepatozyten der Gruppe 1 und 3.
Insgesamt lässt sich festhalten, dass die ultrastrukturellen Veränderungen im Bereich
der Peroxisomen und angrenzender Strukturen bei Tumorpatienten stärker zu Tage
treten als bei den Tieren mit reiner Leberdegeneration und den Kontrolltieren.

140 4 Diskussion
4.3 Beeinflussung der Peroxisomenzahl durch Medikamente
Alle in der vorliegenden Arbeit untersuchten Patienten wurden zur Entnahme einer
Leberbiopsie in Allgemeinnarkose versetzt. In den meisten Fällen handelte es sich
um eine Kombination aus Levomethadon (L-Polamivet®, Fa. Intervet,
Unterschleißheim) und Acepromacin (Vetranquil®, Fa. Albrecht, Aulendorf). In einigen
Fällen wurde die Kombination aus Levomethadon (L-Polamivet®, Fa. Intervet,
Unterschleißheim) und Diazepam (Diazepam®, Roche Holding GmbH, Grenzach-
Whyhlen) verwendet, die getrennt voneinander appliziert wurden. Eine dritte Variante
der Allgemeinnarkose bestand aus der Kombination von Levomethadon (L-
Polamivet®, Fa. Intervet, Unterschleißheim) und Xylazin (Xylazin®, Fa. Cp Pharma,
Burgdorf). Eine letzte Variante bestand aus Propofol und Xylazin (Xylazin®, Fa. Cp
Pharma, Burgdorf).
Zwar liegen keine detaillierten Informationen zur Wirkung dieser Medikamente
explizit auf hepatozelluläre Peroxisomen vor, aber allgemein kann angenommen
werden, dass eine Narkose zu einer Hypoperfusion und damit zu einer Hypoxie der
Zellen führt, die wiederum in der Zelle einen erhöhten oxidativen Stress nach sich
zieht. Dies könnte eine Peroxisomenproliferation induzieren.
In der Gruppe 2 wurde dem Tier K1972 mit geringgradiger Leberzelldegeneration
wiederholt Kortison und dauerhaft Sulfonamide verabreicht. Glukokortikoide führen
zu einer Verminderung der Peroxisomen (PHILLIPS et al. 1987). Dies gilt es bei der
Zählung der Peroxisomen zu berücksichtigen, wenngleich es sich hier nur um einen
Patienten handelte, so dass dies sicher nicht den Ausschlag für das Gesamtergebnis
ergab. Dem Hund K1974 mit einer hochgradigen Degeneration der Hepatozyten
wurde Carprofen verabreicht, ein nicht steroidales Antiphlogistikum. Aufgrund der
Tatsache, dass Acetylsalicylsäure der gleichen Gruppe angehört und dies zu einer
Erhöhung der Peroxisomenzahl beim Menschen führt, kann eine solche Wirkung
theoretisch auch für den Hund von Bedeutung sein. Es fehlen jedoch
wissenschaftliche Untersuchungen dazu.

4 Diskussion 141
Bei Patient K1979 mit einer geringgradig eingestuften Degeneration wurde Trilostan,
ein Wirkstoff zur Behandlung des Morbus Cushing verabreicht. Es konnte keine
Literatur gefunden werden, welche Auswirkungen Trilostan auf die Peroxisomenzahl
von Hepatozyten hat. Es sollte jedoch bedacht werden, dass körpereigene
Glukokortikoide, wie auch exogen zugeführte Glukokortikoide, ebenfalls zu einer
Verminderung der Peroxisomen führen können. Der Patient K2006 wurde mit
Spironolacton behandelt. Aus wissenschaftlichen Untersuchungen der
Humanmedizin ist bekannt, dass dies zu einer Erhöhung der Peroxisomenzahl führt.
Aufgrund einer Hypothyreose des Patienten K 2009 wurden diesem L-Thyroxin
verabreicht. Dieser Wirkstoff führt in den Peroxisomen zu einer erhöhten
peroxisomalen ß-Oxidationskapazität bei gleichzeitiger Erhöhung der
Peroxisomenzahl mit verminderter Größe (JUST u. HARTL 1983; KERCKAERT et al.
1989).
Zusammenfassend ergeben die Untersuchungen dieser Studie keine signifikanten
Unterschiede in der Anzahl der Peroxisomen in degenerativen Veränderungen der
Leber sowie in dem Tumor angrenzenden degenerativ veränderten Gewebe. Der
Hund reagiert entgegen den Untersuchungen in der Humanmedizin nicht mit einer
Peroxisomenproliferation bei degenerativen Veränderungen oder bei oxidativem
Stress in der Leberzelle. Es konnten keine signifikanten Unterschiede hinsichtlich der
Peroxisomengröße und des Verteilungsmusters der Peroxisomen in der Zelle
zwischen den drei Untersuchungsgruppen ermittelt werden. Dies spricht zusätzlich
für eine nicht stattgefundene Peroxisomenproliferation. Signifikant war die Anzahl der
Peroxisomen in den unterschiedlichen Altersklassen. So zeigten ältere Hunde
deutlich mehr Peroxisomen als jüngere Tiere. In einer folgenden Studie sollten neben
der quantitativen Zählung auch die peroxisomalen Enzyme bestimmt werden.
Vielleicht ist die Menge an Katalase in den Peroxisomen ausreichend, um dem
oxidativen Stress entgegenzuwirken bzw. sind die Enzyme der ß- Oxidation
ausreichend zum Abbau von angereicherten Fettsäuren. Zusätzlich sollten auch die
Mitochondrien quantitativ bestimmt werden, denn sie sind ebenfalls an der
Homöostase der Zelle und der Bekämpfung von freien Radikalen beteiligt.

142 5 Zusammenfassung
5 Zusammenfassung
Julia Schüttler (2012)
Zur Bedeutung von Peroxisomen im Metabolismus von degenerativen und
neoplastischen Lebererkrankungen beim Hund
Peroxisomen sind Zellorganellen, die besonders in Leberzellen über ihre
enzymatische Ausstattung in eine Vielzahl physiologischer Stoffwechselwege
eingebunden sind. Vor diesem Hintergrund wurden bei einem Patientenkollektiv der
Kleintierklinik des Tierärztlichen Instituts (Georg-August-Universität Göttingen), die
Auswirkungen von Lebererkrankungen auf die Integrität der hepatozellulären
Peroxisomen bei Hunden untersucht.
Für die Untersuchungen standen 19 Hunde mit Hepatopathien und zwei gesunde
Kontrolltiere zur Verfügung. Aufgrund der histologischen Bewertung erfolgte eine
Zuordnung der Tiere in eine Gruppe mit primären Leberdegenerationen (n=11) und in
eine Gruppe mit neoplastischen Veränderungen (n=8). Neben labordiagnostischen
und umfangreichen lichtmikroskopischen Auswertungen wurden umfangreiche
ultrastrukturelle Untersuchungen an Hepatozyten durchgeführt, um quantitative und
qualitative Veränderungen an den hepatozellulären Peroxisomen festzustellen. Die
dabei ermittelten Ergebnisse lassen sich wie folgt zusammenfassen.
1. Bei den labordiagnostischen Werten (ALT, GLDH, ALKP, Gallensäuren)
zeigen sich im Vergleich besonders in der Gruppe der Tumorpatienten
erhöhte Leberwerte.
2. Die histologischen Leberveränderungen wurden in die drei Gruppen gering-,
mittel- und hochgradige Leberdegenerationen eingeteilt, wobei die de-
generativen Veränderungen durch vakuolisierende, hydropische und
ballonierende Muster mit und ohne Verfettung der Hepatozyten charakterisiert
waren und von verschiedenen reaktiven Prozessen begleitet wurden. Diese
Veränderungen lagen sowohl in der Gruppe der primär degenerativ

5 Zusammenfassung 143
veränderten Hepatozyten vor als auch in der Tiergruppe, die neben
degenerierten Leberzellen Tumorveränderungen aufwiesen.
3. Zu den Tumoren gehörten ein Klarzellkarzinom, ein hepatozelluläres
Karzinom, drei Fälle mit Metastasen eines Hämangiosarkoms, je eine
Metastase eines Plattenepithelkarzinoms, eines Phäochromoblastoms sowie
ein multizentrisches Lymphom. In der Umgebung der neoplastischen
Strukturen zeigten die Hepatozyten graduell unterschiedliche degenerative
Prozesse.
4. Ultrastrukturell sind die Peroxisomen des Hundes durch eine Marginalplatte
und ein zentrales Nukleoid gezeichnet. Weiterhin finden sich vereinzelt
Protrusionen. Die Peroxisomen liegen diffus einzeln im Zytoplasma (panzytär)
und im perinukleären Bereich. Fokal finden sich intrazytoplasmatische Cluster.
5. Beim Gruppenvergleich zeigt sich, dass große Peroxisomen (Durchmesser
>918 nm) besonders bei den Kontrolltieren vorhanden sind, bei denen die
Peroxisomen regelmäßig als Cluster festzustellen sind. Bei den übrigen
Merkmalen treten keine signifikanten Unterschiede auf. Auch hinsichtlich der
Peroxisomenzahl pro µm2 Hepatozytenfläche liegen keine signifikanten
Unterschiede zwischen den Gruppen vor.
6. In allen drei Gruppen können unterschiedliche Veränderungen an den
Mitochondrien der Hepatozyten festgestellt werden, die von lytischen
Prozessen über Schwellungen von Mitochondrien bis zur Bildung von so-
genannten Riesenmitochondrien mit und ohne parakristalline Einschlüsse
reichen, ohne dass statistisch signifikante Unterschiede vorliegen. Dies trifft im
Wesentlichen auch für Veränderungen wie Lipidtropfen, Myelinfiguren und
Glykogengehalte der Hepatozyten zu.
Zusammenfassend lässt sich sagen, dass auch bei Hunden die Peroxisomen ein
wesentlicher struktureller Bestandteil der Hepatozyten sind. Bei degenerativen
Prozessen an Hepatozyten zeigen sich unabhängig von der zugrunde liegenden

144 5 Zusammenfassung
Kausalität Veränderungen, die Ausdruck eines entsprechenden Zellmetabolismus
sein dürften. Bei der Heterogenität der dystrophischen Prozesse und der
unterschiedlichen Ausgangssituation (primäre Degeneration, Degenerationen im
Rahmen einer Neoplasie) sowie den vielseitigen Wechselwirkungen mit reaktiven
Vorgängen wie entzündlichen Zellinfiltrationen ist die Untersuchung von Biopsien mit
Hilfe der Transmissionselektronenmikroskopie ein weniger geeignetes Verfahren, um
anhand punktueller Ergebnisse generelle Aussagen zum Ausmaß der
Leberschädigung zu ermöglichen. Dennoch zeigen die Ergebnisse, dass die
Peroxisomen neben anderen Zellorganellen Rückschlüsse auf das Ausmaß von
hepatozellulären Zellschädigungen bei Hepatopathien des Hundes zulassen.
Möglicherweise lassen sich an den Peroxisomen durch eine stringente Selektion der
Patienten anhand festgelegter klinischer Befunde, labordiagnostischer Parameter
und histologischer Eingrenzung auf ein einheitliches Veränderungsmuster
krankheitstypische ultrastrukturelle Veränderungen nachweisen.

6 Summary 145
6 Summary
Julia Schüttler (2012)
The role of peroxisomes in the metabolism in degenerative and neoplastic
diseases of the canine liver
Peroxisomes are characterized by their content of enzymes. Especially in
hepatocytes the peroxisomes are involved in a variety of metabolic functions. The
purpose of this study was to investigate the effect of liver diseases on the integrity of
canine hepatocellular peroxisomes. The study was performed with patients from the
clinic of small animals (Veterinary institute, Georg-August-university; Göttingen).
The population of dogs included in the study consisted of 19 dogs with hepatopathies
and two healthy dogs as control animals. Dogs were grouped on the basis of results
of histologic examination of biopsy specimens. Disease groups included patients with
primary hepatocellular degeneration (n=11) and patients with neoplastic diseases
(n=8). To determine quantitative and qualitative changes of hepatocellular
peroxisomes, hematologic and biochemical blood profiles, extensive light microscopy
examinations and detailed transmission electron microscopy studies were performed.
The results can be summarized as follows.
1. In comparison to the group of patients with primary degenerative liver disease
the group of patients with neoplastic disease showed elevated serum
concentrations of ALT, GLDH, ALKP and bile acids.
2. The results of histologic examinations of liver tissue were divided into mild,
moderate and severe degenerative hepatocellular changes. Degenerative
alterations of hepatocytes were characterized as vacuolar, hydropic or
ballooning with or without steatosis of hepatocytes. Degenerative hepato-
cellular changes were associated with variable reactive processes. These
pathohistological changes were associated with patients showing primary
degenerative disease as well as with patients suffering from degenerative
hepatocellular disease and neoplasia of liver tissue.

146 5 Summary
3. The group of patients with neoplastic liver disease included one clear cell
carcinoma, a hepatocellular carcinoma, three cases with liver metastasis of a
hemangiosarcoma, metastasis of a squamous cell carcinoma and a
metastasis of a pheochromoblastoma as well as one multicentric lymphoma.
Hepatocytes next to neoplastic structures showed gradually different
degenerative processes.
4. Ultrastructurally canine hepatocellular peroxisomes contained a marginal plate
and a central nucleoid. Protrusions of peroxisomes appeared also. The
peroxisomal distribution pattern in the hepatocytes was arranged diffuse
solitary (pancytoplasmic) and perinuclear. Focal intracytoplasmatic clusters of
peroxisomes existed.
5. Comparing the groups, big peroxisomes (diameter >918nm) were especially
observed in patients of the control group. The peroxisomes were arranged
regulary in clusters. Comparison of the other characteristics revealed no
significant differences. Regarding the feature “peroxisomal number per
hepatocellular surface (µm²)” we determined no significant results among the
groups.
6. Hepatocellular mitochondria of all three groups showed alterations. Lytic
processes, swelling of mitochondria and giant mitochondria with and without
paracrystal inclusions were observed. However there was no significant
difference between the three groups. The mitochondrial aberrations, the
occurrence of lipid droplets, myelin figures and glycogen content of
hepatocytes did not differ significantly.
Results suggest that peroxisomes are common organelles in canine hepatocytes.
Alterations associated with degenerative processes occur apart from the underlying
cause and accordingly reflect cell metabolism. The examination of biopsy specimens
with transmission electron microscopy is not a very suitable method to obtain general
information about hepatocellular damage based on punctual results. An explanation
might be the variability of dystrophic processes, the quite different start points of
disease (primary degeneration, degeneration combined with neoplasia) and the
pathophysiological interactions for example characterized by inflammatory cell

6 Summary 147
infiltrates. The results in the present study reveal that peroxisomes beyond other cell
organelles are a suitable indicator for hepatocellular damage. If patients are selected
stringently based on defined clinical results, laboratory parameters and defined
histologic diagnosis referring to a uniform pattern of changes the investigator might
be able to determine disease related typical ultrastructural changes in peroxisomes.

148 7 Literaturverzeichnis
7 Literaturverzeichnis
ADDIS, T., L. POO u. W. LEW (1936): The quantities of protein lost by the various organs and tissues of the body during a fast. J Biol Chem 115, 111-118 ANGERMULLER, S., M. ISLINGER u. A. VOLKL (2009): Peroxisomes and reactive oxygen species, a lasting challenge. Histochem Cell Biol 131, 459-463 ARBER, N., G. ZAJICEK u. I. ARIEL (1988): The streaming liver. II. Hepatocyte life history. Liver 8, 80-87 BAUDHUIN, P., H. BEAUFAY u. C. DE DUVE (1965): Combined biochemical and morphological study of particulate fractions from rat liver. Analysis of preparations enriched in lysosomes or in particles containing urate oxidase, D-amino acid oxidase, and catalase. J Cell Biol 26, 219-243 BEIER, K., A. VOLKL u. H. D. FAHIMI (1992): Suppression of peroxisomal lipid beta-oxidation enzymes of TNF-alpha. FEBS Lett 310, 273-276 BEIER, K., A. VOLKL u. H. D. FAHIMI (1997): TNF-alpha downregulates the peroxisome proliferator activated receptor-alpha and the mRNAs encoding peroxisomal proteins in rat liver. FEBS Lett 412, 385-387 BERNHARD, W. u. C. ROUILLER (1956): Microbodies and the problem of mitochondrial regeneration in liver cells. J Biophys Biochem Cytol 2, 355-360 BOSGRA, S., W. MENNES u. W. SEINEN (2005): Proceedings in uncovering the mechanism behind peroxisome proliferator-induced hepatocarcinogenesis. Toxicology 206, 309-323 CAMOES, F., N. A. BONEKAMP, H. K. DELILLE u. M. SCHRADER (2009): Organelle dynamics and dysfunction: A closer link between peroxisomes and mitochondria. J Inherit Metab Dis 32, 163-180

7 Literaturverzeichnis 149
CASTEJON, O. J. u. H. D. CASTEJON (2008): Electron Microscopy of myelin figures in normal and pathological tissue. Acta Microscopia 17, 13-19 CENTER, S. A. (2007): Interpretation of liver enzymes. Vet Clin North Am Small Anim Pract 37, 297-333, vii CENTER, S. A., L. GUIDA, M. J. ZANELLI, E. DOUGHERTY, J. CUMMINGS u. J. KING (1993): Ultrastructural hepatocellular features associated with severe hepatic lipidosis in cats. Am J Vet Res 54, 724-731 CHEVILLE, N. F. (1994a): Blockade of Metabolic Pathways. In: N. F. CHEVILLE (Hrsg.): Ultrastructural Pathology- An Introduction to Interpretation 1stedition, Iowa State University Press, Ames, Iowa, Ames, S. 126-191 CHEVILLE, N. F. (1994b): Consequences of Acute cell injury: Necrosis, Recovery and Hypertrophy. In: N. F. CHEVILLE (Hrsg.): Ultrastructural Pathology- An Introduction to Interpretation 1st edition, Iowa State University Press, Ames, Iowa, Ames, S. 80-123 CHEVILLE, N. F. (1994c): Interpretation of Acute cell injury: Degeneration. In: N. F. CHEVILLE (Hrsg.): Ultrastructural Pathology- An Introduction to Interpretation 1st edition, Iowa State University Press, Ames, Iowa, Ames, S. 51-79 CHEVILLE, N. F. (1994d): Neoplasia. In: N. F. CHEVILLE (Hrsg.): Ultrastructural Pathology- An Introduction to Interpretation 1st edition, Iowa State University Press, Ames, Iowa, Ames, S. 229-273 CHEVILLE, N. F. (1994e): Normal Cell Structure: An Overview. In: N. F. CHEVILLE (Hrsg.): Ultrastructural Pathology- An Introduction to Interpretation 1st edition, Iowa State University Press, Ames, Iowa, Ames, S. 2-50 CUERVO, A. M. (2004a): Autophagy: in sickness and in health. Trends Cell Biol 14, 70-77

150 7 Literaturverzeichnis
CUERVO, A. M. (2004b): Autophagy: many paths to the same end. Mol Cell Biochem 263, 55-72 CULLEN, J. M. (2007a): Leber, Gallensystem und exokrines Pankreas. In: M. D. MC GAVIN und J. F. ZACHARY (Hrsg.): Pathologie der Haustiere Elsevier GmbH München, Urban & Fischer Verlag, München, S. 367-425 CULLEN, J. M. (2007b): Schädigung von Zellen und Geweben. In: R. K. MYERS, MC GAVIN, M.D. (Hrsg.): Pathologie der Haustiere Elsevier GmbH München, Urban & Fischer Verlag München, S. CULLEN, J. M. (2009): Summary of the World Small Animal Veterinary Association standardization committee guide to classification of liver disease in dogs and cats. Vet Clin North Am Small Anim Pract 39, 395-418 CULLEN, J. M., T. VAN DEN INGH, T. VAN WINKLE, J. CHARLES u. V. J. DESMET (2006): Morphological classification of parenchymal disorders of the canine and feline liver. In: J. ROTHUIZEN, S. E. BUNCH, J. A. CHARLES, J. M. CULLEN, V. J. DESMET, V. SZATMARI, D. TWEDT, T. VAN DEN INGH, T. VAN WINKLE und R. WASHABAN (Hrsg.): WSAVA Standards for clinical and histological Diagnosis of Canine and Feline Liver Diseases Saunders Elsevier, Philadelphia, S. 77-83 DAY, C. P. u. O. F. JAMES (1998): Steatohepatitis: a tale of two "hits"? Gastroenterology 114, 842-845 DE CRAEMER, D. (1995): Secondary alterations of human hepatocellular peroxisomes. J Inherit Metab Dis 18 Suppl 1, 181-213 DE CRAEMER, D., I. KERCKAERT u. F. ROELS (1991): Hepatocellular peroxisomes in human alcoholic and drug-induced hepatitis: a quantitative study. Hepatology 14, 811-817 DE CRAEMER, D., M. PAUWELS, M. HAUTEKEETE u. F. ROELS (1993): Alterations of hepatocellular peroxisomes in patients with cancer. Catalase cytochemistry and morphometry. Cancer 71, 3851-3858

7 Literaturverzeichnis 151
DE CRAEMER, D., M. PAUWELS u. C. VAN DEN BRANDEN (1995): Alterations of peroxisomes in steatosis of the human liver: a quantitative study. Hepatology 22, 744-752 DE CRAEMER, D., L. VAN MALDERGEM u. F. ROELS (1992): Hepatic ultrastructure in congenital total lipodystrophy with special reference to peroxisomes. Ultrastruct Pathol 16, 307-316 DE DUVE, C. u. P. BAUDHUIN (1966): Peroxisomes (microbodies and related particles). Physiol Rev 46, 323-357 DE LA IGLESIA, F. A., S. M. PINN, J. LUCAS u. E. J. MC GUIRE (1981): Quantitative stereology of peroxisomes in hepatocytes from hyperlipoproteinemic patients receiving Gemfibrozil. Micron 12, 97-98 DE NICOLA, D., S. CENTER, R. HAWTHORNE, B. POTEET, J. M. STEINER, D. TWEDT u. D. A. WILLIAMS (2005): Diagnosing Liver Disease: A roundtable discussion. IDEXXLaboratories, (http://www.idexx.co.uk/animalhealth/analysers/snapreader/lr0905final.pdf) DETER, R. L., P. BAUDHUIN u. C. DE DUVE (1967): Participation of lysosomes in cellular autophagy induced in rat liver by glucagon. J Cell Biol 35, C11-16 DIAL, S. M. (1995): Clinicopathologic evaluation of the liver. Vet Clin North Am Small Anim Pract 25, 257-273 DJORDJEVIC, V. B. (2004): Free radicals in cell biology. Int Rev Cytol 237, 57-89 FAHIMI, H. D., K. BEIER, M. LINDAUER, A. SCHAD, J. ZHAN, J. PILL, W. REBEL, A. VOLKL u. E. BAUMGART (1996): Zonal heterogeneity of peroxisome proliferation in rat liver. Ann N Y Acad Sci 804, 341-361 FAHIMI, H. D., A. REINICKE, M. SUJATTA, S. YOKOTA, M. OZEL, F. HARTIG u. K. STEGMEIER (1982): The short- and long-term effects of bezafibrate in the rat. Ann N Y Acad Sci 386, 111-135

152 7 Literaturverzeichnis
FAN, C. Y., J. PAN, N. USUDA, A. V. YELDANDI, M. S. RAO u. J. K. REDDY (1998): Steatohepatitis, spontaneous peroxisome proliferation and liver tumors in mice lacking peroxisomal fatty acyl-CoA oxidase. Implications for peroxisome proliferator-activated receptor alpha natural ligand metabolism. J Biol Chem 273, 15639-15645 FARRE, J. C. u. S. SUBRAMANI (2004): Peroxisome turnover by micropexophagy: an autophagy-related process. Trends Cell Biol 14, 515-523 FEIGE, J. N., L. GELMAN, L. MICHALIK, B. DESVERGNE u. W. WAHLI (2006): From molecular action to physiological outputs: peroxisome proliferator-activated receptors are nuclear receptors at the crossroads of key cellular functions. Prog Lipid Res 45, 120-159 GEROK, W., BLUM H.E. (1995): Hepatologie. Urban & Schwarzenberg, München, Wien GHADIALLY, F. N. u. E. W. PARRY (1966): Ultrastructure of a human hepatocellular carcinoma and surrounding non-neoplastic liver. Cancer 19, 1989-2004 GIESECKE, D., W. KRAFT u. W. TIEMEYER (1985): [Why Dalmatians excrete uric acid. Causes and consequences of a classical metabolic disorder]. Tierarztl Prax 13, 331-341 GONZALEZ, F. J., J. M. PETERS u. R. C. CATTLEY (1998): Mechanism of action of the nongenotoxic peroxisome proliferators: role of the peroxisome proliferator-activator receptor alpha. J Natl Cancer Inst 90, 1702-1709 GOULD, S. J. u. D. VALLE (2000): Peroxisome biogenesis disorders: genetics and cell biology. Trends Genet 16, 340-345 GRABENBAUER, M., K. SATZLER, E. BAUMGART u. H. D. FAHIMI (2000): Three-dimensional ultrastructural analysis of peroxisomes in HepG2 cells. Absence of peroxisomal reticulum but evidence of close spatial association with the endoplasmic reticulum. Cell Biochem Biophys 32, 37-49

7 Literaturverzeichnis 153
GUO, Y., R. A. JOLLY, B. W. HALSTEAD, T. K. BAKER, J. P. STUTZ, M. HUFFMAN, J. N. CALLEY, A. WEST, H. GAO, G. H. SEARFOSS, S. LI, A. R. IRIZARRY, H. R. QIAN, J. L. STEVENS u. T. P. RYAN (2007): Underlying mechanisms of pharmacology and toxicity of a novel PPAR agonist revealed using rodent and canine hepatocytes. Toxicol Sci 96, 294-309 HARMAN, D. (1956): Aging: a theory based on free radical and radiation chemistry. J Gerontol 11, 298-300 HASHIMOTO, T. (1987): Comparison of enzymes of lipid ß-oxidation in peroxisomes and mitochondria. In: H. D. FAHIMI und H. SIES (Hrsg.): Peroxisomes in Biology and Medicine Springer Verlag, Berlin, Heidelberg, New York, S. 97-104 HASHIMOTO, T. (1999): Peroxisomal beta-oxidation enzymes. Neurochem Res 24, 551-563 HASHIMOTO, T., W. S. COOK, C. QI, A. V. YELDANDI, J. K. REDDY u. M. S. RAO (2000): Defect in peroxisome proliferator-activated receptor alpha-inducible fatty acid oxidation determines the severity of hepatic steatosis in response to fasting. J Biol Chem 275, 28918-28928 HAYASHI, H. u. A. MIWA (1989): The role of peroxisomal fatty acyl-CoA beta-oxidation in bile acid biosynthesis. Arch Biochem Biophys 274, 582-589 HAYASHI, H. u. S. TAKAHATA (1991): Role of peroxisomal fatty acyl-CoA beta-oxidation in phospholipid biosynthesis. Arch Biochem Biophys 284, 326-331 HAYASHI, H., K. TAYA, T. SUGA u. S. NIINOBE (1976): Studies on peroxisomes. VI. Relationship between the peroxisomal core and urate oxidase. J Biochem 79, 1029-1034 HAYASHI, S., S. FUJIWARA u. T. NOGUCHI (2000): Evolution of urate-degrading enzymes in animal peroxisomes. Cell Biochem Biophys 32 Spring, 123-129

154 7 Literaturverzeichnis
HEAD, K. W., J. M. CULLEN, R. R. DUBILZIEG, R. W. ELSE, W. MISDORP, A. K. PATNAIK, S. TATEYAMA u. I. GAAG (2003): Histological Classification of Tumors of the Alimentary System of Domestic Animals. In: F. Y. SCHULMAN (Hrsg.): World Health Organization International Histological Classification of Tumors of Domestic animals 2.Auflage, Armed Forces Institute of Pathology, Washington, DC, Washington, S. 122-124 HESS, F. A., E. R. WEIBEL u. R. PREISIG (1973): Morphometry of dog liver: normal base-line data. Virchows Arch B Cell Pathol 12, 303-317 HOEPFNER, D., D. SCHILDKNEGT, I. BRAAKMAN, P. PHILIPPSEN u. H. F. TABAK (2005): Contribution of the endoplasmic reticulum to peroxisome formation. Cell 122, 85-95 HOSKINS, J. D. (2005): Liver disease in the geriatric patient. Vet Clin North Am Small Anim Pract 35, 617-634 HRUBAN, Z. (1979): Ultrastructure of hepatocellular tumors. J Toxicol Environ Health 5, 403-433 HRUBAN, Z. u. M. RECHCIGL, JR. (1969): Microbodies and related particles. Morphology, biochemistry, and physiology. Int Rev Cytol Suppl 1:1-296 HRUBAN, Z., H. SWIFT u. A. SLESERS (1966): Ultrastructural alterations of hepatic microbodies. Lab Invest 15, 1884-1901 ISSEMANN, I. u. S. GREEN (1990): Activation of a member of the steroid hormone receptor superfamily by peroxisome proliferators. Nature 347, 645-650 JIN, S. u. E. WHITE (2007): Role of autophagy in cancer: management of metabolic stress. Autophagy 3, 28-31 JUST, W. W. u. F. U. HARTL (1983): Rat liver peroxisomes, II. Stimulation of peroxisomal fatty-acid beta-oxidation by thyroid hormones. Hoppe Seylers Z Physiol Chem 364, 1541-1547

7 Literaturverzeichnis 155
KARANTZA-WADSWORTH, V., S. PATEL, O. KRAVCHUK, G. CHEN, R. MATHEW, S. JIN u. E. WHITE (2007): Autophagy mitigates metabolic stress and genome damage in mammary tumorigenesis. Genes Dev 21, 1621-1635 KÄUFER-WEISS, I. (2007): Leber und Gallenwege. In: E. DAHME, WEISS, E. (Hrsg.): Grundriss der speziellen pathologischen Anatomie der Haustiere Verlag Enke, Stuttgart, S. 148-166 KERCKAERT, I., A. CLAEYS, W. JUST, A. CORNELIS u. F. ROELS (1989): Automated image analysis of rat liver peroxisomes after treatment with thyroid hormones: changes in number, size and catalase reaction. Micron Microsc Acta 20, 9-18 KLAUNIG, J. E., M. A. BABICH, K. P. BAETCKE, J. C. COOK, J. C. CORTON, R. M. DAVID, J. G. DELUCA, D. Y. LAI, R. H. MCKEE, J. M. PETERS, R. A. ROBERTS u. P. A. FENNER-CRISP (2003): PPARalpha agonist-induced rodent tumors: modes of action and human relevance. Crit Rev Toxicol 33, 655-780 KLAUNIG, J. E. u. L. M. KAMENDULIS (2004): The role of oxidative stress in carcinogenesis. Annu Rev Pharmacol Toxicol 44, 239-267 KOCH, A., G. SCHNEIDER, G. H. LUERS u. M. SCHRADER (2004): Peroxisome elongation and constriction but not fission can occur independently of dynamin-like protein 1. J Cell Sci 117, 3995-4006 KOCH, A., Y. YOON, N. A. BONEKAMP, M. A. MCNIVEN u. M. SCHRADER (2005): A role for Fis1 in both mitochondrial and peroxisomal fission in mammalian cells. Mol Biol Cell 16, 5077-5086 KOEPKE, J. I., C. S. WOOD, L. J. TERLECKY, P. A. WALTON u. S. R. TERLECKY (2008): Progeric effects of catalase inactivation in human cells. Toxicol Appl Pharmacol 232, 99-108 KRAFT, W. u. U. M. DÜRR (2005): 13 Leber In: W. KRAFT und U. M. DÜRR (Hrsg.): Klinische Labordiagnostik in der Tiermedizin 6.Auflage, Schattauer Verlag, Stuttgart, S. 145-167

156 7 Literaturverzeichnis
KUHN, C. (1968): Particles resembling microbodies in normal and neoplastic perianal glands of dogs. Z Zellforsch Mikrosk Anat 90, 554-562 LAGEWEG, W., J. M. TAGER u. R. J. WANDERS (1991): Topography of very-long-chain-fatty-acid-activating activity in peroxisomes from rat liver. Biochem J 276, 53-56 LAGING, C., A. POSPISCHIL u. D. GIESECKE (1988): [Peroxisomes in the liver and kidney of dogs on a purine-rich and purine-deficient diet. An ultrastructural-morphometric study]. Zentralbl Veterinarmed A 35, 285-290 LAZAROW, P. B. (1978): Rat liver peroxisomes catalyze the beta oxidation of fatty acids. J Biol Chem 253, 1522-1528 LAZAROW, P. B. (1987): The role of peroxisomes in mammalian cellular metabolism. J Inherit Metab Dis 10 Suppl 1, 11-22 LAZAROW, P. B. u. C. DE DUVE (1976): A fatty acyl-CoA oxidizing system in rat liver peroxisomes; enhancement by clofibrate, a hypolipidemic drug. Proc Natl Acad Sci USA 73, 2043-2046 LAZAROW, P. B. u. Y. FUJIKI (1985): Biogenesis of peroxisomes. Annu Rev Cell Biol 1, 489-530 LAZO, O., M. CONTRERAS u. I. SINGH (1990): Topographical localization of peroxisomal acyl-CoA ligases: differential localization of palmitoyl-CoA and lignoceroyl-CoA ligases. Biochemistry 29, 3981-3986 LEE, H. C. u. Y. H. WEI (1997): Mutation and oxidative damage of mitochondrial DNA and defective turnover of mitochondria in human aging. J Formos Med Assoc 96, 770-778 LEGAKIS, J. E., J. I. KOEPKE, C. JEDESZKO, F. BARLASKAR, L. J. TERLECKY, H. J. EDWARDS, P. A. WALTON u. S. R. TERLECKY (2002): Peroxisome senescence in human fibroblasts. Mol Biol Cell 13, 4243-4255

7 Literaturverzeichnis 157
LEMASTERS, J. J., T. QIAN, L. HE, J. S. KIM, S. P. ELMORE, W. E. CASCIO u. D. A. BRENNER (2002): Role of mitochondrial inner membrane permeabilization in necrotic cell death, apoptosis, and autophagy. Antioxid Redox Signal 4, 769-781 LEVINE, B. u. D. J. KLIONSKY (2004): Development by self-digestion: molecular mechanisms and biological functions of autophagy. Dev Cell 6, 463-477 LITTLE, K. C. u. P. CHARTRAND (2004): Genomic DNA is captured and amplified during double-strand break (DSB) repair in human cells. Oncogene 23, 4166-4172 LÖFFLER, G., P. E. PETRIDES u. P. C. HEINRICH (2007): Biochemie und Pathobiochemie. Springer Medizin Verlag, Heidelberg LUFT, J. H. (1961): Improvements in epoxy resin embedding methods. J Biophys Biochem Cytol 9, 409-414 LUIKEN, J. J., M. VAN DEN BERG, J. C. HEIKOOP u. A. J. MEIJER (1992): Autophagic degradation of peroxisomes in isolated rat hepatocytes. FEBS Lett 304, 93-97 LUQUET, S., C. GAUDEL, D. HOLST, J. LOPEZ-SORIANO, C. JEHL-PIETRI, A. FREDENRICH u. P. A. GRIMALDI (2005): Roles of PPAR delta in lipid absorption and metabolism: a new target for the treatment of type 2 diabetes. Biochim Biophys Acta 1740, 313-317 MANDELKER, L. (2008): Introduction to oxidative stress and mitochondrial dysfunction. Vet Clin North Am Small Anim Pract 38, 1-30 MANNAERTS, G. P., L. J. DEBEER, J. THOMAS u. P. J. DE SCHEPPER (1979): Mitochondrial and peroxisomal fatty acid oxidation in liver homogenates and isolated hepatocytes from control and clofibrate-treated rats. J Biol Chem 254, 4585-4595

158 7 Literaturverzeichnis
MANNAERTS, G. P., P. VAN VELDHOVEN, A. VAN BROEKHOVEN, G. VANDEBROEK u. L. J. DEBEER (1982): Evidence that peroxisomal acyl-CoA synthetase is located at the cytoplasmic side of the peroxisomal membrane. Biochem J 204, 17-23 MANNAERTS, G. P. u. P. P. VAN VELDHOVEN (1992): Role of peroxisomes in mammalian metabolism. Cell Biochem Funct 10, 141-151 MARKLUND, S. (1980): Distribution of CuZn superoxide dismutase and Mn superoxide dismutase in human tissues and extracellular fluids. Acta Physiol Scand Suppl 492, 19-23 MARKLUND, S. L. (1992): Regulation by cytokines of extracellular superoxide dismutase and other superoxide dismutase isoenzymes in fibroblasts. J Biol Chem 267, 6696-6701 MASTERS, C. J. u. D. CRANE (1995): The peroxisome: a vital organelle. Cambridge University Press, Cambridge MATHEW, R., V. KARANTZA-WADSWORTH u. E. WHITE (2007): Role of autophagy in cancer. Nat Rev Cancer 7, 961-967 MEIJER, A. J. u. P. CODOGNO (2004): Regulation and role of autophagy in mammalian cells. Int J Biochem Cell Biol 36, 2445-2462 MIZUSHIMA, N. (2005): The pleiotropic role of autophagy: from protein metabolism to bactericide. Cell Death Differ 12 Suppl 2, 1535-1541 MOCHIZUKI, Y., Z. HRUBAN, H. P. MORRIS, A. SLESERS u. E. L. VIGIL (1971): Microbodies of Morris hepatomas. Cancer Res 31, 763-773 MOODY, D. E., J. K. REDDY, B. G. LAKE, J. A. POPP u. D. H. REESE (1991): Peroxisome proliferation and nongenotoxic carcinogenesis: commentary on a symposium. Fundam Appl Toxicol 16, 233-248

7 Literaturverzeichnis 159
NELSON, D. A., T. T. TAN, A. B. RABSON, D. ANDERSON, K. DEGENHARDT u. E. WHITE (2004): Hypoxia and defective apoptosis drive genomic instability and tumorigenesis. Genes Dev 18, 2095-2107 NEUSPIEL, M., A. C. SCHAUSS, E. BRASCHI, R. ZUNINO, P. RIPPSTEIN, R. A. RACHUBINSKI, M. A. ANDRADE-NAVARRO u. H. M. MCBRIDE (2008): Cargo-selected transport from the mitochondria to peroxisomes is mediated by vesicular carriers. Curr Biol 18, 102-108 NOVIKOFF, A. B. u. S. GOLDFISCHER (1969): Visualization of peroxisomes (microbodies) and mitochondria with diaminobenzidine. J Histochem Cytochem 17, 675-680 OLIVEIRA, M. E., A. M. GOUVEIA, R. A. PINTO, C. SA-MIRANDA u. J. E. AZEVEDO (2003): The energetics of Pex5p-mediated peroxisomal protein import. J Biol Chem 278, 39483-39488 OSMUNDSEN, H., J. BREMER u. J. I. PEDERSEN (1991): Metabolic aspects of peroxisomal beta-oxidation. Biochim Biophys Acta 1085, 141-158 PETERS, J. M., C. CHEUNG u. F. J. GONZALEZ (2005): Peroxisome proliferator-activated receptor-alpha and liver cancer: where do we stand? J Mol Med 83, 774-785 PFEIFER, U. (1977): Inhibition by insulin of the physiological autophagic breakdown of cell organelles. Acta Biol Med Ger 36, 1691-1694 PHILLIPS, M. J., S. POUCELL, P. VALENCIA u. J. PATTERSON (1987): The liver. An atlas and text of ultrastructural pathology. Raven Press, New York PLATTA, H. W. u. R. ERDMANN (2007): The peroxisomal protein import machinery. FEBS Lett 581, 2811-2819 POIRIER, Y., V. D. ANTONENKOV, T. GLUMOFF u. J. K. HILTUNEN (2006): Peroxisomal beta-oxidation--a metabolic pathway with multiple functions. Biochim Biophys Acta 1763, 1413-1426

160 7 Literaturverzeichnis
RAO, M. S. u. J. K. REDDY (2004): PPARalpha in the pathogenesis of fatty liver disease. Hepatology 40, 783-786 RAPPAPORT, A. M. (1980): Hepatic blood flow: morphologic aspects and physiologic regulation. Int Rev Physiol 21, 1-63 REDDY, J. K. u. G. P. MANNAERTS (1994): Peroxisomal lipid metabolism. Annu Rev Nutr 14, 343-370 REYNOLDS, E. S. (1963): The use of lead citrate at high pH as an electron-opaque stain in electron microscopy. J Cell Biol 17, 208-212 RHODIN, J. (1954): Correlation of ultrastructural organization and function in normal and experimentally changed proximal convoluted tubule cells of the mouse kidney. Thesis.Stockholm: Aktiebolaget Godvil, Karolinska Institute. ROELS, F., M. ESPEEL u. D. DE CRAEMER (1991): Liver pathology and immunocytochemistry in congenital peroxisomal diseases: a review. J Inherit Metab Dis 14, 853-875 ROELS, F., M. PAUWELS, A. CORNELIS, I. KERCKAERT, P. VAN DER SPEK, G. GOOVAERTS, J. VERSIECK u. S. GOLDFISCHER (1983): Peroxisomes (microbodies) in human liver: cytochemical and quantitative studies of 85 biopsies. J Histochem Cytochem 31, 235-237 ROTH, E. (1997): Oxygen free radicals and their clinical implications. Acta Chir Hung 36, 302-305 ROTH, L. u. D. J. MEYER (1995): Interpretation of liver biopsies. Vet Clin North Am Small Anim Pract 25, 293-303 RYOO, J. W. u. R. J. BUSCHMANN (1989): Morphometry of liver parenchyma in needle biopsy specimens from patients with alcoholic liver disease: preliminary variables for the diagnosis and prognosis of cirrhosis. Mod Pathol 2, 382-389

7 Literaturverzeichnis 161
SACKSTEDER, K. A., J. M. JONES, S. T. SOUTH, X. LI, Y. LIU u. S. J. GOULD (2000): PEX19 binds multiple peroxisomal membrane proteins, is predominantly cytoplasmic, and is required for peroxisome membrane synthesis. J Cell Biol 148, 931-944 SAKAI, Y., A. KOLLER, L. K. RANGELL, G. A. KELLER u. S. SUBRAMANI (1998): Peroxisome degradation by microautophagy in Pichia pastoris: identification of specific steps and morphological intermediates. J Cell Biol 141, 625-636 SAKAI, Y., M. OKU, I. J. VAN DER KLEI u. J. A. KIEL (2006): Pexophagy: autophagic degradation of peroxisomes. Biochim Biophys Acta 1763, 1767-1775 SCHOONJANS, K., B. STAELS u. J. AUWERX (1996): Role of the peroxisome proliferator-activated receptor (PPAR) in mediating the effects of fibrates and fatty acids on gene expression. J Lipid Res 37, 907-925 SCHRADER, M. (2001): Tubulo-reticular clusters of peroxisomes in living COS-7 cells: dynamic behavior and association with lipid droplets. J Histochem Cytochem 49, 1421-1429 SCHRADER, M. (2006): Shared components of mitochondrial and peroxisomal division. Biochim Biophys Acta 1763, 531-541 SCHRADER, M., J. K. BURKHARDT, E. BAUMGART, G. LUERS, H. SPRING, A. VOLKL u. H. D. FAHIMI (1996): Interaction of microtubules with peroxisomes. Tubular and spherical peroxisomes in HepG2 cells and their alterations induced by microtubule-active drugs. Eur J Cell Biol 69, 24-35 SCHRADER, M. u. H. D. FAHIMI (2004): Mammalian peroxisomes and reactive oxygen species. Histochem Cell Biol 122, 383-393 SCHRADER, M. u. H. D. FAHIMI (2006): Growth and division of peroxisomes. Int Rev Cytol 255, 237-290 SCHRADER, M. u. H. D. FAHIMI (2008): The peroxisome: still a mysterious organelle. Histochem Cell Biol 129, 421-440

162 7 Literaturverzeichnis
SCHRADER, M., K. KRIEGLSTEIN u. H. D. FAHIMI (1998a): Tubular peroxisomes in HepG2 cells: selective induction by growth factors and arachidonic acid. Eur J Cell Biol 75, 87-96 SCHRADER, M., B. E. REUBER, J. C. MORRELL, G. JIMENEZ-SANCHEZ, C. OBIE, T. A. STROH, D. VALLE, T. A. SCHROER u. S. J. GOULD (1998b): Expression of PEX11beta mediates peroxisome proliferation in the absence of extracellular stimuli. J Biol Chem 273, 29607-29614 SCHRADER, M., M. THIEMANN u. H. D. FAHIMI (2003): Peroxisomal motility and interaction with microtubules. Microsc Res Tech 61, 171-178 SCHRADER, M., R. WODOPIA u. H. D. FAHIMI (1999): Induction of tubular peroxisomes by UV irradiation and reactive oxygen species in HepG2 cells. J Histochem Cytochem 47, 1141-1148 SCHRADER, M. u. Y. YOON (2007): Mitochondria and peroxisomes: are the 'big brother' and the 'little sister' closer than assumed? Bioessays 29, 1105-1114 SEPESY, L. M., S. A. CENTER, J. F. RANDOLPH, K. L. WARNER u. H. N. ERB (2006): Vacuolar hepatopathy in dogs: 336 cases (1993-2005). J Am Vet Med Assoc 229, 246-252 SHARMA, K. D., L. A. ANDERSSON, T. M. LOEHR, J. TERNER u. H. M. GOFF (1989): Comparative spectral analysis of mammalian, fungal, and bacterial catalases. Resonance Raman evidence for iron-tyrosinate coordination. J Biol Chem 264, 12772-12779 SHNITKA, T. K. (1966): Comparative ultrastructure of hepatic microbodies in some mammals and birds in relation to species differences in uricase activity. J Ultrastruct Res 16, 598-625 SINGH, A. K., G. S. DHAUNSI, M. P. GUPTA, J. K. ORAK, K. ASAYAMA u. I. SINGH (1994): Demonstration of glutathione peroxidase in rat liver peroxisomes and its intraorganellar distribution. Arch Biochem Biophys 315, 331-338

7 Literaturverzeichnis 163
SINGH, H. u. A. POULOS (1988): Distinct long chain and very long chain fatty acyl CoA synthetases in rat liver peroxisomes and microsomes. Arch Biochem Biophys 266, 486-495 STALKER, M. u. M. HAYES (2007): Liver and Biliary System. In: M. G. MAXIE (Hrsg.): Jubb, Kennedy and Palmer´s Pathology of Domestic animals Elsevier Saunders Philadelphia PA, Philadelphia, S. 297-387 STEMPAK, J. G. u. R. T. WARD (1964): An Improved Staining Method for Electron Microscopy. J Cell Biol 22, 697-701 STOCKHAM, S. L. u. M. A. SCOTT (2008): Enzymes Liver function. In: S. L. STOCKHAM und M. A. SCOTT (Hrsg.): Fundamentals of Veterinary Clinical Pathology 2nd edition, Blackwell Publishing Professional Iowa, S. 639-706 TERLECKY, S. R., J. I. KOEPKE u. P. A. WALTON (2006): Peroxisomes and aging. Biochim Biophys Acta 1763, 1749-1754 TOLBERT, N. E. u. E. ESSNER (1981): Microbodies: peroxisomes and glyoxysomes. J Cell Biol 91, 271s-283s USUDA, N., M. K. REDDY, T. HASHIMOTO, M. S. RAO u. J. K. REDDY (1988): Tissue specificity and species differences in the distribution of urate oxidase in peroxisomes. Lab Invest 58, 100-111 VAN DER ZAND, A., I. BRAAKMAN, H. J. GEUZE u. H. F. TABAK (2006): The return of the peroxisome. J Cell Sci 119, 989-994 VEENHUIS, M., J. A. KIEL u. I. J. VAN DER KLEI (2003): Peroxisome assembly in yeast. Microsc Res Tech 61, 139-150

164 7 Literaturverzeichnis
VIEIRA, H. L. u. G. KROEMER (1999): Pathophysiology of mitochondrial cell death control. Cell Mol Life Sci 56, 971-976 WALLACE, D. C. (2001): A mitochondrial paradigm for degenerative diseases and ageing. Novartis Found Symp 235, 247-263; discussion 263-246 WANDERS, R. J. (2000): Peroxisomes, lipid metabolism, and human disease. Cell Biochem Biophys 32, 89-106 WANDERS, R. J. (2004a): Metabolic and molecular basis of peroxisomal disorders: a review. Am J Med Genet A 126A, 355-375 WANDERS, R. J. (2004b): Peroxisomes, lipid metabolism, and peroxisomal disorders. Mol Genet Metab 83, 16-27 WANDERS, R. J. u. H. R. WATERHAM (2006): Biochemistry of mammalian peroxisomes revisited. Annu Rev Biochem 75, 295-332 WEBB, C. u. D. TWEDT (2008): Oxidative stress and liver disease. Vet Clin North Am Small Anim Pract 38, 125-135, v WIEMER, E. A., T. WENZEL, T. J. DEERINCK, M. H. ELLISMAN u. S. SUBRAMANI (1997): Visualization of the peroxisomal compartment in living mammalian cells: dynamic behavior and association with microtubules. J Cell Biol 136, 71-80 YAMAMOTO, K. u. H. D. FAHIMI (1987): Three-dimensional reconstruction of a peroxisomal reticulum in regenerating rat liver: evidence of interconnections between heterogeneous segments. J Cell Biol 105, 713-722 YAN, M., N. RAYAPURAM u. S. SUBRAMANI (2005): The control of peroxisome number and size during division and proliferation. Curr Opin Cell Biol 17, 376-383 YASMINEH, W. G., J. L. PARKIN, J. I. CASPERS u. A. THEOLOGIDES (1991): Tumor necrosis factor/cachectin decreases catalase activity of rat liver. Cancer Res 51, 3990-3995

7 Literaturverzeichnis 165
ZAAR, K. (1992): Structure and function of peroxisomes in the mammalian kidney. Eur J Cell Biol 59, 233-254

166 8 Abbildungsverzeichnis
8 Abbildungsverzeichnis
Abbildung 2-1 Aufbau einer idealisierten tierischen Zelle (LÖFFLER et al. 2007) 15
Abbildung 2-2 Elektronenmikroskopische Darstellung eines Peroxisoms (Pfeil)
mit Marginalplatte und Nukleoid. (P596-07/K1979),
Ultradünnschnitt, Transmissionselektronenmikroskopie,
Gerätevergr. 16.000x). 16
Abbildung 2-3 Abbau geradzahliger Fettsäuren durch β-Oxidation (LÖFFLER et
al. 2007) 26
Abbildung 2-4 Modell für die Teilung und Proliferation von Peroxisomen
(modifiziert nach PLATTA u. ERDMANN 2007) 33
Abbildung 2-5 Translokation von Proteinen zum Wachstum der Peroxisomen
(modifiziert nach PLATTA u. ERDMANN 2007) 35
Abbildung 2-6 Schematische Darstellung der funktionellen Organisation der
Leber (CULLEN 2007a) 41
Abbildung 2-7 Hydropische Degeneration von Hepatozyten im Bereich der
Zentralvene (Z=Zentralvene; Rind, Paraffinschnitt, H.E.)
(CULLEN 2007a) 43
Abbildung 2-8 Physiologische Zelle und Veränderungen bei reversibler und
irreversibler Zellschädigung (CULLEN 2007b) 44
Abbildung 3-1 Übersicht eines Hepatozyten mit Bestimmung der Fläche in µm²,
(K2011, Ultradünnschnitt, MIA, Transmissionselektronen-
mikroskopie Gerätevergr. 2500x) 67
Abbildung 3-2 ALT Laborwertveränderungen der Gruppe 1 und 2
(Gradeinteilung nach CENTER 2007) 73
Abbildung 3-3 GLDH Laborwertveränderungen der Gruppe 1 und 2
(Gradeinteilung nach CENTER 2007) 74

8 Abbildungsverzeichnis 167
Abbildung 3-4 ALKP Laborwertveränderungen der Gruppe 1 und 2
(Gradeinteilung nach CENTER 2007) 75
Abbildung 3-5 Gallensäure Laborwertveränderungen der Gruppe 1 und 2
(Gradeinteilung nach CENTER 2007) 76
Abbildung 3-6 Verteilung des Schweregrades der Degeneration in den Gruppen
1-3 78
Abbildung 3-7 Geringgradige Leberzelldegeneration mit ggr. Itozellhyperplasie
(Pfeil) und minimalen gemischtzelligen perivaskulären
Entzündungszellinfiltraten (Pfeilkopf), 81
Abbildung 3-8 Mittelgradige hydropische Degeneration der Hepatozyten
(Pfeilkopf) (P430/06, Paraffinschnitt, H.E.). 82
Abbildung 3-9 Hochgradige Degeneration der Hepatozyten. Inset zeigt eine
ausgeprägte gemischttropfige Verfettung (539/07, Paraffinschnitt,
H.E.). 83
Abbildung 3-10 Hochgradige zentrolobuläre Leberzelldegeneration vom
Pflanzenzelltyp (P31/08, Paraffinschnitt, H.E.). 84
Abbildung 3-11 Geringgradige hydropische Leberzelldegeneration mit
Hämosiderinspeicherung in Kupffer Sternzellen (schwarze Pfeile)
(P596/07, Paraffinschnitt, Berliner Blau Färbung). 85
Abbildung 3-12 Geringgradige hydropische bis vakuoläre hepatozelluläre
Degeneration mit Cholestase (schwarze Pfeile), (P45/09,
Paraffinschnitt, H.E.). 85
Abbildung 3-13 Itozellhyperplasie mit Verfettung (a), geringgradige vakuoläre
Leberzelldegeneration (A), Inset (B): Metastase eines
Plattenepithelkarzinoms in der Leber, (P67/08, Paraffinschnitt,
H.E.). 89
Abbildung 3-14 Mittelgradige hydropische Leberzelldegeneration (A),
intrazelluläre Pigmentablagerung (dünne Pfeile), minimale

168 8 Abbildungsverzeichnis
Infiltrate neutrophiler Granulozyten (dicke Pfeile); Inset (B):
Metastase eines Phäochromoblastoms (C) in der Leber,
tumorfreies Lebergewebe (D), (P992/08, Paraffinschnitt, H.E.). 90
Abbildung 3-15 Hochgradige hydropische bis ballonierende Degeneration der
Hepatozyten (A) im an den Tumor (Hämangiosarkom)
angrenzenden Lebergewebe (großes Bild); Inset: Hämangio-
sarkom, (P803/07, Paraffinschnitt, H.E.). 91
Abbildung 3-16 Geringgradige hepatozelluläre Degeneration bei
hepatozellulärem Karzinom, Hämosiderinablagerung in Kupffer
Sternzellen (schwarze Pfeile) und intrahepatozellulär (rote Pfeile)
(P467/06-1, Paraffinschnitt, Berliner Blau Färbung). 92
Abbildung 3-17 Ausgeprägte Gallengangsproliferation (dünne Pfeile),
Stauungshyperämie (A) und hochgradige hydropische
Leberzelldegeneration (Paraffinschnitt, H.E.) bei einem Tier
(P803/07) mit einem Hämangiosarkom. 93
Abbildung 3-18 Geringgradige hydropische Degeneration, (P2103/3, Paraffin-
schnitt, H.E.). 94
Abbildung 3-19 Gegenüberstellung von Semidünnschnitt und Paraffinschnitt bei
einem Fall (K1972) mit geringgradiger hydropischer Leberzell-
degeneration mit assoziierter gemischttropfiger Verfettung
(Semidünnschnitt, Methylenblau; Paraffinschnitt, H.E.). 96
Abbildung 3-20 Geringgradige hepatozelluläre Degeneration (K1968/1). Der
Semidünnschnitt zeigt inhomogene zytoplasmatische Färbungen
mit eingelagerten grünlichen Fettvakuolen in den Hepatozyten.
Daneben sieht man einen Fetttropfen (Pfeil), der im H.E. Schnitt
(Inset) als leere Vakuole erkennbar ist. Im H.E. Schnitt sind
zusätzlich Herde mit Hämosiderinablagerungen (Kreis).
(Semidünnschnitt, Methylenblau; Paraffinschnitt, H.E. Färbung). 97

8 Abbildungsverzeichnis 169
Abbildung 3-21 Mittelgradige hepatozelluläre Degeneration bei einem Hund mit
Klarzellkarzinom (K2011/20). Die im Inset sichtbare mittelgradige
hydropische Schwellung der Hepatozyten ist im Semidünnschnitt
auf eine massive Glykogenablagerung mit einzelnen feinen
Fettvakuolen zurückzuführen. (Semidünnschnitt, Methylenblau;
Paraffinschnitt, H.E. Färbung). 98
Abbildung 3-22 Ausgeprägte hepatozelluläre Degenerationserscheinungen bei
einem Tier (K2025/3) aus der Gruppe 1. Neben Fetttropfen (F)
zeigen sich Degenerationserscheinungen durch starke deutliche
Schwellung der Hepatozyten (Pfeilspitzen). Andere Hepatozyten
zeigen intrazytoplasmatische Pigmentablagerungen (dünne
Pfeile) oder sind durch unregelmäßige Glykogenablagerungen
(G) charakterisiert. In fast allen Zellen sind kleine Vakuolen zu
erkennen, bei denen es sich um geschwollene Mitochondrien
handelt (Semidünnschnitt, Methylenblau). 99
Abbildung 3-23 Kontrolltier (K2103) mit einzelnen Vakuolen in den ansonsten
regelmäßig angeordneten Hepatozyten, die ein homogen
gefärbtes Zytoplasma aufweisen (Semidünnschnitt,
Methylenblau). 100
Abbildung 3-24 Verteilung kleiner Peroxisomen in den Gruppen 1-3 106
Abbildung 3-25 Verteilung großer Peroxisomen in den Gruppen 1-3 106
Abbildung 3-26 Peroxisomales Verteilungsmuster der Gruppen 1-3 107
Abbildung 3-27 Verteilungsmuster der Peroxisomen (Pfeile) als Cluster.
Mitochondrium (M) (K2103/15, Ultradünnschnitt, Transmissions-
elektronenmikroskopie, Gerätevergr. 16.000x). 108
Abbildung 3-28 Perinukleäres Verteilungsmuster der Peroxisomen (Pfeile).
Nukleus (N), Mitochondrium (M) (K2103/12, Ultradünnschnitt,
Transmissionselektronenmikroskopie, Gerätevergr. 6300x). 108

170 8 Abbildungsverzeichnis
Abbildung 3-29 Verteilung der Peroxisomen (Px) im Bereich von Fetttropfen (F)
und Mitochondrium (M). (K2029, Ultradünnschnitt,
Transmissionselektronenmikroskopie, Gerätevergr. 16.000x). 109
Abbildung 3-30 Morphologische Veränderungen der Peroxisomen der Gruppen
1-3 110
Abbildung 3-31 Protrusion (roter Pfeil) eines Peroxisoms (Px). Mitochondrium
(M), Nukleus (N) (K1965, Ultradünnschnitt, Transmissions-
elektronenmikroskopie, Gerätevergr. 12.000x). 110
Abbildung 3-32 Marginalplatten (rote Pfeile) der Peroxisomen (Px).
Mitochondrium (M), das geschwollen ist. (K2029,
Ultradünnschnitt, Transmissionselektronenmikroskopie,
Gerätevergr. 16.000x). 111
Abbildung 3-33 Peroxisom (Px) mit Nukleoid (Nk) und Marginalplatte (Pfeil).
(K1969, Ultradünnschnitt, Transmissionselektronenmikroskopie,
Gerätevergr. 25.000x). 111
Abbildung 3-34 Eigenschaften der Mitochondrien der Gruppen 1-3 113
Abbildung 3-35 Riesenmitochondrium (RM) mit parakristallinen Einschlüssen
(Pfeile) in Nachbarschaft zu einem normal großen Mitochondrium
(M) verdeutlicht den Größenunterschied. (K1979,
Ultradünnschnitt, Transmissionselektronenmikroskopie,
Gerätevergr. 16.000x). 113
Abbildung 3-36 Lyse eines Mitochondriums (M). (K2015, Ultradünnschnitt,
Transmissionselektronenmikroskopie, Gerätevergr. 12.500x). 114
Abbildung 3-37 Schwellung eines Mitochondriums (M). (K2015, Ultradünnschnitt,
Transmissionselektronenmikroskopie, Gerätevergr. 12.500x). 114
Abbildung 3-38 Häufigkeit des Vorkommens von Lipidtropfen in den Hepatozyten
der Untersuchungsgruppen 1-3 115

8 Abbildungsverzeichnis 171
Abbildung 3-39 Gemischttropfige Verfettung (F) eines Hepatozyten. (K1972,
Ultradünnschnitt, Transmissionselektronenmikroskopie, Geräte-
vergr. links 2500x und rechts 3150x). 116
Abbildung 3-40 Häufigkeit des Auftretens von Myelinfiguren 117
Abbildung 3-41 Myelinfigur (MF) in unmittelbarer Nachbarschaft zu einem
Mitochondrium (M). (K2015, Ultradünnschnitt, Transmissions-
elektronenmikroskopie, Gerätevergr. 16.000x). 117
Abbildung 3-42 Glykogengehalt der Hepatozyten der Untersuchungsgruppen 1-3 118
Abbildung 3-43 Ausgeprägte Glykogenansammlungen (rote Pfeile) neben einem
Peroxisom (Px) mit Nukleoid (N) und sichtbarer Marginalplatte
(schwarzer Pfeil). (K1972, Ultradünnschnitt, Transmissions-
elektronenmikroskopie, Gerätevergr. 16.000x). 119
Abbildung 3-44 Darstellung der Mittelwerte der Peroxisomenzahl/
Hepatozytenfläche (µm²) in den drei Untersuchungsgruppen 120
Abbildung 3-45 Mittelwerte der Peroxisomen/Hepatozytenfläche( µm²) in Bezug
zu den Altersgruppen 121
Abbildung 3-46 Mittelwerte der Peroxisomen/Hepatozytenfläche (µm²) in Bezug
zu den Gewichtsklassen 122

172 9 Tabellenverzeichnis
9 Tabellenverzeichnis
Tabelle 2-1 Unterschiede peroxisomaler und mitochondrialer β-Oxidation 27
Tabelle 2-2 Übersicht der Lebertumoren (CULLEN 2007a) 52
Tabelle 3-1 Anzahl der Patienten mit Hepatopathien, die nicht mit in die
vorliegende Studie einbezogen werden konnten 58
Tabelle 3-2 Übersicht Patienten der Gruppe 1 mit degenerativen
Lebererkrankungen (w=weiblich, wk=weiblich kastriert,
m=männlich, M.=Morbus, WHWT=Westhighland White Terrier,
JRT=Jack Russel Terrier, RHT=Rauhaarteckel) 59
Tabelle 3-3 Übersicht Patienten der Gruppe die an einem Primär- oder
Sekundärtumor erkrankten (w=weiblich, m=männlich,
JRT=Jack Russel Terrier, RHT=Rauhaarteckel) 60
Tabelle 3-4 Kontrolltiere (m=männlich, d=Tage) 61
Tabelle 3-5 Rasseverteilung der Gruppen 1-3 70
Tabelle 3-6 Altersverteilung: absolute und relative Häufigkeit der Patienten
der Gruppen 1-3 71
Tabelle 3-7 Gewichtsverteilung der Patienten der Gruppen 1-3 71
Tabelle 3-8 Geschlechtsverteilung der Hunde der Gruppen 1-3 72
Tabelle 3-9 Zusammenfassung der histopathologischen Untersuchungs-
befunde von Gruppe 1 79
Tabelle 3-10 Tumorverteilung in Gruppe 2 86

1 173
Tabelle 3-11 Histopathologische Untersuchungsbefunde der Gruppe 2 (+)
minimal, + geringgradig, 87
Tabelle 3-12 Semiquantitative Beurteilung der Morphologie der
Peroxisomen und der Hepatozyten bei Tieren ohne
Lebertumoren 102
Tabelle 3-13 Semiquantitative Beurteilung der Morphologie der
Peroxisomen und der Hepatozyten bei Lebertumoren 103
Tabelle 3-14 Semiquantitative Beurteilung der Morphologie der
Peroxisomen und der Hepatozyten bei Kontrolltieren 104

174 10 Anhang
10 Anhang
10.1 Protokolle für die Histologie
10.1.1 Fixierlösungen
10 %iges neutral gepuffertes Formalin
35 %ige Formaldehydlösung 285 ml
Aqua dest. 215 ml
Stammlösung A Phosphatpuffer (s.u.) 100 ml
Stammlösung B Phosphatpuffer (s.u.) 400 ml
2,5 %ige gepufferte Glutaraldehydlösung
25 %ige Glutaraldehydlösung 100 ml
Aqua dest. 400 ml
Lösung A Phosphatpuffer (s.u.) 100 ml
Lösung B Phosphatpuffer (s.u.) 400 ml
10.1.1.1 Phosphatpuffer
Stammlösung A (0,2 M)
Natriumdihydrogenphosphat-Monohydrat (Fa. Merck, Darmstadt, Nr.6346)
27,6g
Aqua bidest. ad 1000 ml
Stammlösung B (0,2 M)
Di-Natriumhydrogenphosphat-Dihydrat (Fa. Merck, Darmstadt, Nr. 6580)
35,6 g
Aqua bidest. ad 1000 ml

10 Anhang 175
Gebrauchslösung (0,1M, pH 7,4-7,6)
Stammlösung A 10 ml
Stammlösung B 40 ml
Aqua bidest. 50 ml
10.1.1.2 Epongemisch nach LUFT (1961)
Mischung A:
Glycidether 100
(Fa. Serva, Heidelberg, BRD, Nr.21045) 71,3 g
DDSA (2-Dodecenylsuccinic acid anhydrid)
(Fa. Serva, Heidelberg, BRD, Nr.20755) 115,0g
auf einem Magnetrührer 1 h rühren
Mischung B:
Glycidether 100
(Fa. Serva, Heidelberg, BRD, Nr.21045) 100,0 g
MNA (Methylnadic anhydride)
(Fa. Serva, Heidelberg, BRD, Nr.29452) 89,0 g
auf einem Magnetrührer 1 h rühren
Mischung A und B im Verhältnis 1:1 mischen und 30 min rühren
1,8 % Beschleuniger 2,4,6-Tris-dimethylaminomethylphenol
(Fa. Serva, Heidelberg, Nr. 36975)
tropfenweise dazugeben und 15 min rühren

176 10 Anhang
10.1.2 Färbungen
10.1.2.1 Hämalaun & Eosin Färbung
Entparaffinisierung und Rehydratisierung:
Xylol: 5 min
Xylol: 2 min
Xylol: 2 min
100 %iger Alkohol: 2 min
96 %iger Alkohol: 2 min
80 %iger Alkohol: 1 min
70 %iger Alkohol: 1 min
Aqua dest. 1 min
Zur Kernfärbung: Hämalaun nach Mayer (Fa. Merck, Darmstadt) für 3 min
Zum Bläuen 10 min fließend wässern
Zur Gegenfärbung: wässriges Eosin (Fa. Thermo Fisher Scientific, Dreieich)
für 5 min
Fließend wässern für 5 s
Entwässerung in der aufsteigenden Alkoholreihe:
70 %iger Alkohol: 1 min
80 %iger Alkohol: 1 min
96 %iger Alkohol: 1 min
100 %iger Alkohol: 1 min
100 %iger Alkohol: 2 min
Xylol: 1 min
Xylol: 3 min
Xylol: 3 min
Eindecken mit Eukitt (Fa. Kindler, Freiburg)

10 Anhang 177
10.1.2.2 Berliner Blau Reaktion (Eisennachweis nach Perls)
Entparaffinisierung und Rehydratisierung:
Xylol: 5 min
Xylol: 2 min
Xylol: 2 min
100 %iger Alkohol: 2 min
96 %iger Alkohol: 2 min
80 %iger Alkohol: 1 min
70 %iger Alkohol: 1 min
Aqua dest.: 1 min
Kaliumferrozyanid 10 %ig: 5 min
1:1 HCl 20 %ig und Kaliumferrozyanid 20 %ig: 30 min
3 x in Aqua dest. spülen
Kernechtrot (Darstellung der Kerne): 5 min
3 x in Aqua dest. spülen
Entwässerung in der aufsteigenden Alkoholreihe:
70 %iger Alkohol: 1 min
80 %iger Alkohol: 1 min
96 %iger Alkohol: 1 min
100 %iger Alkohol: 1 min
100 %iger Alkohol: 2 min
Xylol: 1 min
Xylol: 3 min
Xylol: 3 min
Eindecken mit Eukitt (Fa. Kindler, Freiburg)

178 10 Anhang
10.1.2.3 Periodic-Acid-Schiff-Reaktion (PAS)
Entparaffinisierung und Rehydratisierung:
Xylol: 5 min
Xylol: 2 min
Xylol: 2 min
100 %iger Alkohol: 2 min
96 %iger Alkohol: 2 min
80 %iger Alkohol: 1 min
70 %iger Alkohol: 1 min
Aqua dest.: 1 min
0,5 % Perjodsäure 10 min
Aqua dest. fließend 10 min
Schiff´s Reagenz (Fa. Merck, Darmstadt): 15 min
Leitungswasser fließend: 10 min
Gegenfärbung mit Hämalaun nach Mayer: 30 s
Bläuen in Leitungswasser: 10 min
Entwässerung in der aufsteigenden Alkoholreihe:
70 %iger Alkohol: 1 min
80 %iger Alkohol: 1 min
96 %iger Alkohol: 1 min
100 %iger Alkohol: 1 min
100 %iger Alkohol: 2 min
Xylol: 1 min
Xylol: 3 min
Xylol: 3 min
Eindecken mit Eukitt (Fa. Kindler, Freiburg)

10 Anhang 179
10.1.2.4 Methylenblaufärbung nach RICHARDSON (1960)
Lösung A:
Azur II für die Lichtmikroskopie (Fa. Merck, Darmstadt, Nr. 9211)
1,0 g
Aqua dest. ad 100 ml
Lösung B:
di-Natriumtetraborat p.a. (Fa. Merck, Darmstadt, Nr. 6306)
1,0 g
Aqua dest. ad 100 ml
Methylenblau für die Lichtmikroskopie (Fa. Merck, Darmstadt, Nr. 15943)
1,0 g darin lösen
Lösungen A und B im Verhältnis 1:1 mischen und filtrieren
Schnitte auf der Wärmebank (60°C) 1 min mit dieser Mischung färben

180 10 Anhang
10.1.2.5 Doppelkontrastierung mit Uranylacetat und Bleicitrat nach
Reynolds
1 Tropfen des 5 %igen Uranylacetates in eine mit Paraplast gefüllte Petri-
schale pipettieren
Kupferobjektträgernetz für 30 min. in diesen Tropfen legen
Kupferobjektträgernetz gründlich in Aqua dest. abspülen
von der gebrauchsfertigen, zentrifugierten Lösung 1 Tropfen in eine mit
Paraplast ausgegossene Petrischale
Kupferobjektträgernetz für 7 Minuten in diesen Tropfen legen
Petrischale verschließen
nach dem Kontrastieren gründlich mit Aqua bidest. spülen
Grids trocknen lassen
Ansetzen des Bleicitrates :
Bleinitrat (Fa. Merck, Darmstadt, Nr. 7398)
1,33 g
Natriumcitrat (Fa. Merck, Darmstadt, Nr. 6448)
1,67 g
Aqua dest. ad 30 ml
Reagenzien stark schütteln
nach 30 min erfolgt Umwandlung von Bleinitrat zu Bleicitrat
8 ml 1N NaOH (carbonatfrei) zusetzen
auf 50 ml Aqua dest. auffüllen und schwenken
Nach Auflösen des Bleicitrates ist die Lösung fertig (pH 12)
Uranylacetatlösung
5 %ige wässrige Uranylacetat-Lösung (Fa. Merck, Darmstadt, Nr. 8473
lichtgeschützt aufbewahren!)- vorher zentrifugieren

10 Anhang 181
10.1.3 Laborprotokolle für Einbettungen
Paraffineinbettung- Excelsior ES Protokoll
Wasser (entmineralisiert) für 2 h bei Zimmertemperatur
aufsteigende Alkoholreihe (50 %ig-70 %ig-80 %ig-96 %ig-100 %ig-100 %ig)
für jeweils 45 min bei 35°C und Vakuum
2x Isopropanol für je1 h bei Zimmertemperatur und Vakuum
3x Paraffin für jeweils 1,0 h bei 60°C und Vakuum
Eponeinbettung nach Luft (1961)
1. Spülen in 0,1 M Phosphatpuffer (pH 7,4) 1,4 h/4°C
(3x wechseln)
2. Nachfixieren in 1 %iger Osmiumtetroxidlösung
0,1 M Phosphatpuffer (pH 7,4-7,6) 2,0 h/4°C
3. Spülen in 0,1 M Phophatpuffer (pH 7,4) 1,5 h/4°C
(3x wechseln)
4. Dehydrieren in einer aufsteigenden Ethanolreihe
30 %iger Alkohol 30 min/4°C
50 %iger Alkohol 30 min/4°C
70 %iger Alkohol 30 min/4°C
96 %iger Alkohol 30 min/4°C
100 %iger Alkohol 30 min/4°C
100 %iger Alkohol 30 min/4°C
Isopropanol 30 min/4°C
5. Infiltration mit Propylenoxid 30 min/4°C
(2x wechseln)
6. Infiltration mit Propylenoxid-Epon 2:1 1,0 h/10°C
7. Infiltration mit Propylenoxid-Epon 1:1 2,0 h/10°C
8. Infiltration mit Propylenoxid-Epon 1:2 4,0 h/10°C
9. Infiltration mit reinem Epon über Nacht/20°C
10. Infiltration mit reinem Epon 3,0h/20°C

182 10 Anhang
10.2 Anhangstabellen
Anhangstabelle 10-1 Klinische Chemie der Patienten mit einer hepatozelulären Degeneration (Gruppe 1) mit klinikeigenen Referenzwerten
Parameter Ref.wert
K-Nummer
2029 1972 1979 2015 2017 1963 2062 2063 1969 1974 2025
Glukose (mg/dl) 70-120 81 95 135
sie
he A
nhangsta
belle
10-4
86 94 107 112 124 96
sie
he A
nhangsta
belle
10-6
Fructosamin(µmol/l) 0-370 283 187 108 60 90 199 323 88 42
Gallensäure(µmol/l) 0-20 3 17 22 6 12 53 2 13 3
GLDH (U/l) 0-6 39 28 >73 4 >73 38 21 6 2
ALT (U/l) 0-55 260 237 363 36 417 307 79 88 30
AST (U/l) 0-30 398 47 42 47 72 73 37 21 19
T-Bil (mg/dl) 0-0,4 0,15 0,2 * 0,1 0,4 0,2 0,2 0,3 0,2
AP (U/l) 0-150 380 >800 >800 102 270 385 93 743 800
Ges.-Eiweiß (g/dl) 5,4-7,5 6,2 8,0 8,0 5,9 8,1 6,0 5,8 6,6 5,5
Albumin (g/dl) 2,5-4,4 2,7 3,4 4,3 2,8 4,0 2,6 3,1 3,2 2,4
Globulin (g/dl) 2,51-4,51
3,4 4,6 3,7 3,1 4,1 3,4 2,6 3,4 3,1
Kreatinin (mg/dl) 0,4-1,4 1,2 0,84 0,94 0,8 0,6 0,9 0,8 0,8 0,9
Harnstoff (mg/dl) 20-50 70 20 29 28 23 35 25 17 25
Calcium (mmol/l) 2,3-3,0 2,8 2,7 2,9 2,2 2,5 3,0 2,7 2,7 2,2
Phosphor (mmol/l) 0,7-1,6 1,7 1,0 1,3 1,5 1,7 0,9 1,2 1,1 1,2
α- Amylase (U/l) 0-1650 1285 355 661 1200 571 532 577 435 430
Cholesterin (mg/dl) 120-390 258 590 253 152 281 245 308 288 371
Lipase (U/l) 0-300 161 26 57 109 14 77 71 22 19
Kreatininkinase (U/l) 0-90 641 93 96 139 172 104 179 100 61
Natrium (mmol/l) 140-155 145 * * * * 148 144 * *
Kalium (mmol/l) 3,5-5,1 4,9 * * * * 5,2 4,4 * *
CRP (mg/l) 0-15 7 * * * * 3 4 * *
PT (sec.) 12-17 14 12 16 * 12 12 * * 14
PTT (sec.) 71-102 80 70 97 * 70 ** * * 83
* nicht gemessen ** nicht messbar (out of range)

10.2 Anhangstabellen 183
Anhangstabelle 10-2 Klinische Chemie der Patienten mit einem primären oder sekundären Tumor der Leber (Gruppe 2) mit klinikeigenen Referenzwerten
Parameter Ref.wert
K-Nummer
1965 2006 2009 1968 2030 2027 2011 2021
Glukose (mg/dl) 70-120 v * 84 117 91 95
sie
he A
nhangsta
belle
10-4
I
Fructosamin(µmol/l) 0-370 e * 156 52 347 315 D
Gallensäure(µmol/l) 0-20 t * 9 26 7 986 E
GLDH (U/l) 0-6 t * 69 73 5 sie
he
Anhangsta
belle
10-4
X
ALT (U/l) 0-55 e * 317 399 56 X
AST (U/l) 0-30 s * 169 71 80
T-Bil (mg/dl) 0-0,4 t * 0,2 0,4 0,3
AP (U/l) 0-150 * 261 236 33
Ges.-Eiweiß (g/dl) 5,4-7,5 * 6,8 6,0 7,0 6,3
Albumin (g/dl) 2,5-4,4 * 3,5 2,5 2,9 3,4
Globulin (g/dl) 2,51-4,51
* 3,3 3,5 2,9
Kreatinin (mg/dl) 0,4-1,4 * 1,4 1,0 1,2 1,2
Harnstoff (mg/dl) 20-50 * 84 31 28 33
Calcium (mmol/l) 2,3-3,0 * 2,4 2,4 3,6 2,8
Phosphor (mmol/l) 0,7-1,6 * 1,0 1,5 1,0 1,2
α- Amylase (U/l) 0-1650 * 367 735 805 708
Cholesterin (mg/dl) 120-390 * 285 250 246 769
Lipase (U/l) 0-300 * 22 36 60 294
Kreatininkinase (U/l) 0-90 * 73 315 65 328
Natrium * * * 148 144
Kalium * * * 4,17 3,8
CRP * * * 11 10
PT (sec.) 12-17 * 14 * * 43 12 *
PTT (sec.) 71-102 * 74 * * 114 87 *
* nicht gemessen ** nicht messbar (out of range)

184 10 Anhang
Anhangstabelle 10-3 Differentialblutbilder der Patienten Gruppe 1 und Gruppe 2
K-Nr.
Leuk.
(K/µl)
Erythr.
(M/µl)
Neutr.
(%)
Lymph.
. (%)
Mono.
(%)
Eos.
(%)
Baso.
(%)
Hb
(g/dl)
HKT
(%)
MCV
(fL)
MCH
(pg)
MCHC
(g/dl)
PLT
(K/µl)
Gru
pp
e 1
2029 24 6,8 87 5 7 0 1 15 43 64 22 35 365
1972 8,0 8,3 74 15 5 5 1 18 62 75 22 30 556
1979 12 9,3 74 18 7 1 1 20 60 64 21 33 436
2015 siehe Anhangstabelle 10-5
2017 26 4,7 91 6 3 0 0 11 33 69 24 34 136
1963 12 7,5 81 13 5 0 1 18 54 72 24 34 16
2062 16 5,4 94 5 1 0 0 12 34 64 22 34 391
2063 14 7,0 76 10 12 0 2 16 47 67 23 34 1079
1969 8 7,0 71 19 8 1 1 18 54 74 24 32 536
1974 10 7,5 78 6 15 0 1 17 51 68 22 32 796
2025 107 2,7 98 1 1 0 0 6 19 72 23 32 553
Gru
pp
e 2
1965 siehe Anhangstabelle 10-8
2006 * * * * * * * * * * * * *
2009 6 6,4 51 47 0 1 1 14 42 65 22 34 128
1968 29 6,1 74 6 15 0 4 14 38 63 22 35 549
2030 5 7,0 71 12 14 2 1 15 44 65 22 34 330
2027 14 8,0 92 7 1 0 0 19 58 70 22 32 270
2011 siehe Anhangstabelle 10-5
2021 15 6,0 89 6 1 4 0 13 41 68 22 32 381
Ref.-Wert 6-12
5,5-
8,5
55-
75 13-33 0-5 0-4 0-1
15-
19
44-
52
60-
77
17-
23 31-34 200-400
* nicht gemessen ** nicht messbar (out of range)

10.2 Anhangstabellen 185
Anhangstabelle 10-4 Klinische Chemie Patienden der Gruppen 1 und 2 (Laboklin)
* nicht gemessen ** nicht messbar (out of range)
Parameter Ref.Wert
Gruppe 1 Gruppe 2
2015 2027 2011
Fructosamin (µmol/l) 0-374 218 * 329
GLDH (U/l) 0-6 9 263 53
GGT (U/l) 0-5 9 * 5,8
ALT (U/l) 0-55 54 1268 1071
AST (U/l) 0-25 8 182 32
T-Bil (µmol/l) 0-3,4 1,5 39 1,2
AP (U/l) 0-108 115 4787 109
Ges.-Eiweiß (g/l) 54-75 63 * 73
Albumin (g/l) 25-44 39 * 40
Globulin (g/l) 25-45 24 * 33
Kreatinin (µmol/l) 35-106 52 * 71
Harnstoff (mmol/l) 3,3-8,3 3,8 * 3,9
Calcium (mmol/l) 2,3-3,0 2,7 * 2,7
Phosphor (mmol/l) 0,7-1,6 1,4 * 1,7
α- Amylase (U/l) 0-1650 1016 * 965
Cholesterin (mmol/l) 3,1-10,1 5,2 * 7,3
Lipase (U/l) 0-300 593 * 45
Kreatininkinase (U/l) 0-90 68 * 34
Natrium (mmol/l) 140-155 147 * 146
Kalium (mmol/l) 3,5-5,1 4,4 * 4,5
CRP (mg/l) 0-15 * * *
PT (sec.) 12-17 13 43 12
PTT (sec.) 71-102 69 114 87

186 10 Anhang
Anhangstabelle 10-5 Differentialblutbilder der Patienten Gruppe 1 und Gruppe 2 (Laboklin)
K-Nr.
Leuko.
(G/l)
Erythr.
(T/l)
Neutr.
(%)
Lymph
. (%)
Mono.
(%)
Eos.
(%)
Baso.
(%)
Hb
(g/l)
HKT
(l/l)
PLT
(G/l)
Gru
pp
e 1
2015 7,4 8,0 71 18 7 4 0 186 0,57 440
Gru
pp
e 2
2011 9,7 7,5 69 25 6 0 0 190 0,48 315
Ref.-Wert 6-12 5,5-8,5 55-75 13-30 0-4 0-6 0-1
150-
190
0,44-
0,52
150-
500
Anhangstabelle 10-6 Klinische Chemie (Synlab) Gruppe 1
Parameter Ref.Wert
Gruppe 1
2025
GLDH (U/l) 0-9,6 4,9
ALT (U/l) 0-80 17
AST (U/l) 0-76 35
T-Bil (µmol/l) 0-8,55 3,42
AP (U/l) <141 20
Ges.-Eiweiß (g/l) 55-75 54
Albumin (g/l) 28-46 24,0
Kreatinin (µmol/l) <141 99,9
Harnstoff (mmol/l) 3,3-8,3 12,8
Calcium (mmol/l) 2,0-3,0 2,2
Phosphor (mmol/l) 0,68-1,74 1,14
Cholesterin (mmol/l) 3,49-6,99 3,44
Natrium (mmol/l) 140-155 153
Kalium (mmol/l) 3,5-5,1 3,9
Glukose (mmol/l) 3,05-5,55 5,27
Eisen (µmol/l) 11,3-34,0 20,4

10.2 Anhangstabellen 187
Anhangstabelle 10-7 Klinische Chemie Vettest Gruppe 2
Anhangstabelle 10-8 Hämatologie (Gruppe 2) Vet Abc
K-Nr.
WBC
(10³/µl)
Erythr.
(10 6 /µl)
Neutr.
(%)
Lymph.
(%)
Mono.
(%)
Hb
(gdl)
HKT
(%)
PLT
(10³/ µl)
Gru
pp
e 2
1965 13,3 3,2 88 7 5 7,04 21 49
Ref.-
Wert 6-12 5,5-8,5 0-100 0-100 0-100 15-20
44,0-
57,0 150-500
Parameter Ref.Wert
Gruppe 2
1965 2021
ALT (U/l) 10-100 59 776
AST (U/l) 0-50 55 *
AP (U/l) 23-212 - 2000
Urea (mmol/l) 2,5-9,6 17,1 -
Kreatinin (µmol/l) 44-159 241 -
Kreatinin (mg/dl) 0,5-1,8 - 0,6
Glukose (mmol/l) 3,89-7,94 6,59 -
Glukose (mg/dl) 70-143 - 74
T-Bil (µmol/l) 0-15 2 -
- T-Bil (mg/dl) 0,0-0,9 - 5,0
BUN (mg/dl) 7-27 - 13
Albumin ( g/dl) 2,2-3,9 - 2,8
Total Protein (g/dl) 5,2-8,2 - 6,3
Globulin ( g/dl) 2,5-4,5 - 3,4
Amylase ( U/l) 500-1000 - 827
Cholesterin (mg/dl) 110-320 - 261
Calcium (mg/dl) 7,9-12,0 - 10,5
Phosphor (mg/dl) 2,5-6,8 - 2,6

188 10 Anhang
Anhangstabelle 10-9 Auswertungsbogen der elektronenmikroskopischen Untersuchung
Elmi
K-Nummer
Schnittqualität
Fotos
Morphologie Peroxisomen
PX Zahl
PX Größe
Verteilung innerhalb der Zelle
Invagination
Protrusion
Nukleoid in PX
Marginalplatte
Besonderheiten
Mitochondrien
Endoplasmat. Retikulum
Glykogen
Nukleus
Lysosomen
Cytoplasma
Endothel
Mikrovilli
Anzahl Px pro Hepatozyt
Anzahl Px und Fläche des
Hepatozyten
Hep A Px
Fläche µm² Hep B Px
Fläche µm²
Hep C Px
Fläche µm² Hep D Px
Fläche µm²
Hep E Px
Fläche µm² Hep F Px
Fläche µm²
Px/Fläche (Zytosol)

10.2
An
ha
ng
sta
be
lle
n
189
An
ha
ng
sta
be
lle
10
-10
Sta
mm
da
ten
de
r s
tati
sti
sc
he
n A
us
we
rtu
ng
de
r m
orp
ho
log
isc
he
n U
nte
rsu
ch
un
ge
n d
er
Pe
rox
iso
me
n u
nd
Mit
och
on
dri
en
so
wie
in
traze
llu
läre
r B
es
tan
dte
ile
Gru
pp
e
Tie
r K
-
Nr.
Px k
lein
P
x g
roß
V
er-
teilu
ng
in d
er
Zelle
(1)
Ver-
teilu
ng
in d
er
Zelle
(2)
Pro
-
trusio
n
Mar-
gin
al-
pla
tte
Nukle
o-
id
Mito-
chon-
drien
Schw
el-
lung
Mito-
chon-
drien
Lyse
Mito-
chon.
para
kris
talli
ne
Ein
schl.
Rie
sen-
mito-
chon-
drien
Lip
id-
tropfe
n
Myelin
-
figure
n
Gly
ko-
gen
1
202
9
2
2
1
1
1
1
1
2
2
1
3
2
5
1
197
2
2
2
2
1
1
1
2
2
2
1
2
5
4
1
197
9
3
2
3
1
1
1
1
1
1
1
1
3
4
1
201
5
3
2
3
2
2
1
2
2
2
1
3
3
4
1
201
7
3
2
2
2
1
2
1
1
1
1
1
2
3
1
196
3
2
2
2
3
1
1
1
2
2
1
1
2
2
4
1
206
2
1
2
3
1
1
1
1
2
2
1
5
2
2
1
206
3
2
5
2
3
2
1
1
1
1
2
1
5
3
4
1
196
9
1
3
3
2
1
1
2
2
1
1
2
1
4
1
197
4
3
1
4
2
1
2
1
1
2
1
3
2
4
1
202
5
3
1
4
2
1
2
1
1
2
1
3
2
4
2
196
5
2
2
3
1
1
1
1
1
2
2
5
4
3
2
200
6
2
2
3
2
2
2
1
1
2
2
5
4
2
2
200
9
2
2
3
1
1
1
1
1
1
1
3
3
4
2
196
8
2
2
3
1
1
2
2
2
2
1
2
1
3
2
203
0
2
2
2
3
1
1
1
1
1
2
2
5
1
3
2
202
7
2
2
3
1
1
1
1
1
2
1
2
3
2
2
201
1
2
2
3
2
2
2
2
2
1
1
5
5
4
2
202
1
2
5
2
3
1
1
1
1
1
1
2
5
3
3
3
210
4-7
2
4
2
3
2
1
1
1
2
2
1
1
2
4
3
210
4-8
2
4
2
3
1
2
1
1
1
2
1
2
2
4
3
210
4-9
2
4
2
3
1
1
1
1
2
1
2
2
1
4
3
210
3-3
3
2
2
4
1
2
1
1
1
2
1
1
2
4
3
210
3-1
2
2
4
2
4
1
1
2
1
1
2
1
1
3
4
3
210
3-1
5
3
3
2
3
2
2
1
1
2
2
1
2
2
4
Lege
nd
e
Px
Grö
ße
Ve
r-te
ilu
ng
in
Z
ell
e
P
ro-
tru
sio
n
Ma
r-g
ina
lpl.
Nu
kle
oid
M
ito
ch
. S
ch
we
ll.
Mit
oc
h.
Lys
e
Mit
oc
h.
Pa
rak
ris
t.
Ein
sc
hl.
R
ies
en
mi.
L
ipid
-tr
op
fen
M
ye
lin
fig
. G
lyk
og
en
(+
) ve
rein
zelt
1
<1
0 %
N
äh
e
Fe
tttr
op
f 1
Ja
=1
Ja
= 1
Ja
=1
Ja
=1
Ja
=1
Ja
=1
Ja
=1
(+
)min
ima
l=
1
(+)m
inim
al
=1
(+
)min
ima
l=
1
+
gg
r.
2
10
-30
%
Clu
ste
r 2
N
ein
=2
Nein
=2
Nein
=2
Nein
=2
Nein
=2
Nein
=2
Nein
=2
+ g
gr.
=2
+ g
gr.
=2
+ g
gr.
=2
+
+m
gr.
3
3
0-5
0 %
P
an
zytä
r 3
++
mg
r.=
3
++
mg
r.=
3
++
mg
r.=
3
+
++
hg
r.
4
50
-10
0 %
P
eri
-n
ukle
är
4
+
++
hg
r. =
4
++
+ h
gr.
=4
+
++
hg
r=4
.
ke
ine
5
Ke
ine
=5
Ke
ine
=5
Ke
ine
=5

190 10 Anhang
Anhangstabelle 10-11 Ergebnisse der Peroxisomenzählung pro Hepatozyt mit Mittelwert
Tier K Nummer Gruppe Hep1 Px/µm²Hep2 Px/µm²Hep3 Px/µm²Hep4 Px/µm²Hep5 Px/µm²Hep6 Px/µm²Mittelwerte
1 2029 kein Tumor 0,08 0,05 0,08 0,09 0,07 0,12 0,08
2 1972 kein Tumor 0,16 0,14 0,17 0,19 0,10 0,14 0,15
3 1979 kein Tumor 0,09 0,18 0,15 0,17 0,19 0,14 0,15
4 2015 kein Tumor 0,07 0,12 0,11 0,07 0,08 0,16 0,10
5 2017 kein Tumor 0,11 0,13 0,14 0,15 0,13 0,14 0,13
6 1963 kein Tumor 0,18 0,16 0,17 0,17 0,13 0,12 0,16
7 2062 kein Tumor 0,12 0,15 0,15 0,09 0,11 0,10 0,12
8 2063 kein Tumor 0,13 0,24 0,15 0,16 0,15 0,11 0,16
9 1969 kein Tumor 0,11 0,19 0,19 0,12 0,12 0,14 0,15
10 1974 kein Tumor 0,05 0,07 0,07 0,09 0,11 0,09 0,08
11 2025 kein Tumor 0,09 0,08 0,08 0,09 0,11 0,12 0,10
12 1965 Tumor 0,07 0,06 0,10 0,09 0,07 0,10 0,08
13 2006 Tumor 0,07 0,07 0,04 0,04 0,04 0,08 0,06
14 2009 Tumor 0,09 0,08 0,11 0,06 0,09 0,10 0,09
15 1968 Tumor 0,11 0,08 0,18 0,09 0,08 0,10 0,11
16 2030 Tumor 0,08 0,05 0,07 0,08 0,09 0,09 0,08
17 2027 Tumor 0,17 0,13 0,11 0,10 0,08 0,10 0,11
18 2011 Tumor 0,05 0,02 0,06 0,02 0,06 0,05 0,04
19 2021 Tumor 0,17 0,16 0,15 0,21 0,27 0,18 0,19
20 2104-7 Kontrolle 0,04 0,12 0,06 0,04 0,06 0,05 0,06
21 2104-8 Kontrolle 0,08 0,10 0,09 0,06 0,06 0,08 0,08
22 2104-9 Kontrolle 0,10 0,13 0,09 0,09 0,12 0,13 0,11
23 2103-3 Kontrolle 0,11 0,11 0,12 0,11 0,16 0,14 0,12
24 2103-12 Kontrolle 0,11 0,22 0,11 0,20 0,19 0,07 0,15
25 2103-15 Kontrolle 0,08 0,07 0,05 0,10 0,06 0,04 0,07

10.2 Anhangstabellen 191
Anhangstabelle 10-12 Stammdaten der statistischen Untersuchung (Mittelwert der Peroxisomen/µm², Alter, Gewichtsklasse)
K Nummer Gruppe Hep1 Px/µm²Hep2 Px/µm²Hep3 Px/µm²Hep4 Px/µm²Hep5 Px/µm²Hep6 Px/µm²Mittelwerte Alter in Jahren Gewichtsklasse
2029 kein Tumor 0,08 0,05 0,08 0,09 0,07 0,12 0,08 11-15 10-20 kg
1972 kein Tumor 0,16 0,14 0,17 0,19 0,10 0,14 0,15 6-10 0-10 kg
1979 kein Tumor 0,09 0,18 0,15 0,17 0,19 0,14 0,15 11-15 20-40 kg
2015 kein Tumor 0,07 0,12 0,11 0,07 0,08 0,16 0,10 11-15 20-40 kg
2017 kein Tumor 0,11 0,13 0,14 0,15 0,13 0,14 0,13 6-10 10-20 kg
1963 kein Tumor 0,18 0,16 0,17 0,17 0,13 0,12 0,16 6-10 10-20 kg
2062 kein Tumor 0,12 0,15 0,15 0,09 0,11 0,10 0,12 11-15 0-10 kg
2063 kein Tumor 0,13 0,24 0,15 0,16 0,15 0,11 0,16 11-15 0-10 kg
1969 kein Tumor 0,11 0,19 0,19 0,12 0,12 0,14 0,15 11-15 0-10 kg
1974 kein Tumor 0,05 0,07 0,07 0,09 0,11 0,09 0,08 11-15 20-40 kg
2025 kein Tumor 0,09 0,08 0,08 0,09 0,11 0,12 0,10 6-10 0-10 kg
1965 Tumor 0,07 0,06 0,10 0,09 0,07 0,10 0,08 11-15 20-40 kg
2006 Tumor 0,07 0,07 0,04 0,04 0,04 0,08 0,06 6-10 20-40 kg
2009 Tumor 0,09 0,08 0,11 0,06 0,09 0,10 0,09 6-10 20-40 kg
1968 Tumor 0,11 0,08 0,18 0,09 0,08 0,10 0,11 11-15 10-20 kg
2030 Tumor 0,08 0,05 0,07 0,08 0,09 0,09 0,08 6-10 20-40 kg
2027 Tumor 0,17 0,13 0,11 0,10 0,08 0,10 0,11 11-15 0-10 kg
2011 Tumor 0,05 0,02 0,06 0,02 0,06 0,05 0,04 6-10 10-20 kg
2021 Tumor 0,17 0,16 0,15 0,21 0,27 0,18 0,19 11-15 0-10 kg
2104-7 Kontrolle 0,04 0,12 0,06 0,04 0,06 0,05 0,06 1-5 10-20 kg
2104-8 Kontrolle 0,08 0,10 0,09 0,06 0,06 0,08 0,08 1-5 10-20 kg
2104-9 Kontrolle 0,10 0,13 0,09 0,09 0,12 0,13 0,11 1-5 10-20 kg
2103-3 Kontrolle 0,11 0,11 0,12 0,11 0,16 0,14 0,12 1-5 10-20 kg
2103-12 Kontrolle 0,11 0,22 0,11 0,20 0,19 0,07 0,15 1-5 10-20 kg
2103-15 Kontrolle 0,08 0,07 0,05 0,10 0,06 0,04 0,07 1-5 10-20 kg

192 Danksagung
Danksagung
An erster Stelle danke ich meinem Doktorvater, Herrn Univ. Prof. Dr. F.-J. Kaup, für
die Überlassung des Dissertationsthemas, die uneingeschränkte Unterstützung des
Vorhabens, in Form von Diskussionen, fachlichem Rat und freundlichen Zuspruch.
Ich danke für die Begeisterung für die Pathologie, insbesondere für die
Elektronenmikroskopie, die ich, als Klinikerin, stets durch ihn erfahren habe.
Ich bedanke mich sehr herzlich bei Herrn Prof. Dr. Stephan Neumann für die
Hilfestellung bei der Umsetzung dieser Dissertation, sowohl in fachlichen Fragen und
wissenschaftlichen Diskussionen als auch für die freundschaftliche Unterstützung.
Danke an Frau Prof. Dr. Marion Hewicker-Trautwein für die Bereitschaft, das
Zweitgutachten zu erstellen.
Darüberhinaus danke ich den Mitarbeitern der Abteilung Medizinische Statistik der
Universität Göttingen, insbesondere Frau Dr. Katharina Lange, Herrn Simon
Schneider und Herrn Thomas Asendorf, für die statistische Auswertung meiner
Daten.
Ich danke allen Mitarbeitern der Kleintierklinik des Tierärztlichen Institutes bei der
Durchführung und Umsetzung dieses Vorhabens. Insbesondere danke ich Uta,
Wendy und Sonja für Ihren Zuspruch, ihre Hilfsbereitschaft und Freundschaft. Ganz
besonders möchte ich Almuth für ihre Ideen und die Unterstützung im Hinblick auf
die Dissertation danken, insbesondere für die Freundschaft, die Diskussionen und
wertvollen Gespräche in allen Lebenslagen über all die Jahre. Danke.

Danksagung 193
Die Abteilung Infektionspathologie des Deutschen Primatenzentrums mit all ihren
Mitarbeitern hat mich immer herzlich aufgenommen - ich danke Euch. Ich danke Dr.
Martina Zöller für die wertvollen Hinweise zur Lichtmikroskopie und der
fotografischen Erfasssung der histologischen Befunde. Frau PD Dr. Martina
Schlumbohm für Ratschläge und Ideen hinsichtlich der Statistik dieser Arbeit und
Frau Stephanie Heiduck für die Hilfe bei der Beschaffung sehr ausgefallener
Literaturquellen. Besonders möchte ich Frau Nadine Schminke meinen Dank
aussprechen, die mit fachkundigem Rat und großem Engagement an der Umsetzung
dieses Projektes beteiligt war. Ebenso geht ein großes Dankeschön an Frau Kaiser-
Jarry für ihre Hilfe bei der Umsetzung der Arbeit. Ich werde sie in wertvoller
Erinnerung behalten. Ebenfalls danke ich sehr herzlich Anne Gereke für die
abschließende sorgfältige Durchsicht der Dissertation.
Ganz besonders danke ich Frau Dr. Eva Gruber-Dujardin für die überaus
sorgfältigen, wertvollen und kritischen Korrekturen meiner Dissertation. Ich danke für
die zahlreichen Ratschläge und fachlichen Diskussionen, die von unschätzbarem
Wert für das Gelingen dieser Arbeit waren. Danke, Eva.
Ich empfinde große Dankbarkeit gegenüber meiner Familie. Dieses Projekt nahm viel
Zeit in Anspruch, die ich nicht mit der Familie verbringen konnte. Ich danke Euch für
euren Rückhalt, das stetige Verständnis und die Liebe.
Mein größter Dank geht an meine Eltern Karl-Heinz und Reinhild, die mir über all die
Jahre, die Zeit des Veterinärstudiums inbegriffen, Rückhalt, Hilfe zu jeder Zeit, ein
geborgenes Zuhause, ein offenes Ohr und letztlich auch finanzielle Unterstützung
gegeben haben. Danke.

194 Danksagung
Zum Abschluss möchte ich Raimund für sein unendliches Vertrauen, seine
Zuversicht und seine unermessliche Geduld danken. Ich bin sehr glücklich, dass Du
mich in dieser Zeit begleitet hast.

Göttingen, den 27.Februar 2012
Erklärung
Hiermit erkläre ich, dass ich die Dissertation mit dem Titel „Zur Bedeutung von
Peroxisomen im Metabolismus von degenerativen und neoplastischen
Lebererkrankungen beim Hund“ selbstständig verfasst habe. Bei der Anfertigung
wurden folgende Hilfen Dritter in Anspruch genommen:
Abteilung Medizinische Statistik der Universität Göttingen
Ich habe keine entgeltliche Hilfe von Vermittlungs- bzw. Beratungsdiensten
(Promotionsberater oder anderer Personen) in Anspruch genommen. Niemand hat
von mir unmittelbar oder mittelbar entgeltliche Leistungen für Arbeiten erhalten, die
im Zusammenhang mit dem Inhalt der vorgelegten Dissertation stehen.
Ich habe die Dissertation an folgenden Institutionen angefertigt:
Deutsches Primatenzentrum, Abt. Infektionspathologie; Göttingen
Kleintierklinik, Tierärztliches Institut, Georg-August-Universität; Göttingen
Die Dissertation wurde bisher nicht für eine Prüfung oder Promotion oder für einen
ähnlichen Zweck zur Beurteilung eingereicht.
Ich versichere, dass ich die vorstehenden Angaben nach bestem Wissen vollständig
und der Wahrheit entsprechend gemacht habe.
Julia Schüttler


