biglycan is required for adaptive remodeling after ... 8.pdf · biglycan is required for adaptive...
TRANSCRIPT

ISSN: 1524-4539 Copyright © 2008 American Heart Association. All rights reserved. Print ISSN: 0009-7322. Online
72514Circulation is published by the American Heart Association. 7272 Greenville Avenue, Dallas, TX
DOI: 10.1161/CIRCULATIONAHA.107.714147 2008;117;1269-1276; originally published online Feb 25, 2008; Circulation
Tschöpe and J.W. Fischer Young, H.P. Schultheiss, B. Levkau, H.A. Baba, T. Unger, K. Zacharowski, C.
R. Lüllmann-Rauch, O. Lettau, C. Jacoby, J. Schrader, S.-M. Brand-Herrmann, M.F. D. Westermann, J. Mersmann, A. Melchior, T. Freudenberger, C. Petrik, L. Schaefer,
Biglycan Is Required for Adaptive Remodeling After Myocardial Infarction
http://circ.ahajournals.org/cgi/content/full/117/10/1269located on the World Wide Web at:
The online version of this article, along with updated information and services, is
http://www.lww.com/reprintsReprints: Information about reprints can be found online at
[email protected]. E-mail:
Fax:Kluwer Health, 351 West Camden Street, Baltimore, MD 21202-2436. Phone: 410-528-4050. Permissions: Permissions & Rights Desk, Lippincott Williams & Wilkins, a division of Wolters
http://circ.ahajournals.org/subscriptions/Subscriptions: Information about subscribing to Circulation is online at
at SWETS SUBSCR SVC/25338706 on September 15, 2008 circ.ahajournals.orgDownloaded from

Biglycan Is Required for Adaptive Remodeling AfterMyocardial Infarction
D. Westermann, MD*; J. Mersmann, MD*; A. Melchior, MS*; T. Freudenberger, MS*; C. Petrik;L. Schaefer, PhD; R. Lüllmann-Rauch, MD; O. Lettau, MS; C. Jacoby, PhD; J. Schrader, MD;S.-M. Brand-Herrmann, MD, PhD; M.F. Young, PhD; H.P. Schultheiss, MD; B. Levkau, MD;H.A. Baba, MD; T. Unger, MD; K. Zacharowski, MD; C. Tschöpe, MD; J.W. Fischer, PhD
Background—After myocardial infarction (MI), extensive remodeling of extracellular matrix contributes to scar formationand preservation of hemodynamic function. On the other hand, adverse and excessive extracellular matrix remodelingleads to fibrosis and impaired function. The present study investigates the role of the small leucine-rich proteoglycanbiglycan during cardiac extracellular matrix remodeling and cardiac hemodynamics after MI.
Methods and Results—Experimental MI was induced in wild-type (WT) and bgn�/0 mice by permanent ligation of the leftanterior descending coronary artery. Biglycan expression was strongly increased at 3, 7, and 14 days after MI in WTmice. bgn�/0 mice showed increased mortality rates after MI as a result of frequent left ventricular (LV) ruptures.Furthermore, tensile strength of the LV derived from bgn�/0 mice 21 days after MI was reduced as measured ex vivo.Collagen matrix organization was severely impaired in bgn�/0 mice, as shown by birefringence analysis of Sirius redstaining and electron microscopy of collagen fibrils. At 21 days after MI, LV hemodynamic parameters were assessedby pressure-volume measurements in vivo to obtain LV end-diastolic pressure, end-diastolic volume, and end-systolicvolume. bgn�/0 mice were characterized by aggravated LV dilation evidenced by increased LV end-diastolic volume (bgn�/0,111�4.2 �L versus WT, 96�4.4 �L; P�0.05) and LV end-diastolic pressure (bgn�/0, 24�2.7 versus WT, 18�1.8 mm Hg;P�0.05) and severely impaired LV function (EF, bgn�/0, 12�2% versus WT, 21�4%; P�0.05) 21 days after MI.
Conclusion—Biglycan is required for stable collagen matrix formation of infarct scars and for preservation of cardiachemodynamic function. (Circulation. 2008;117:1269-1276.)
Key Words: collagen � myocardial infarction � extracellular matrix � fibrosis � proteoglycans
Extensive remodeling of the extracellular matrix (ECM)occurs during all phases of infarct healing, including
initial degradation of ECM by matrix metalloproteinases(MMPs), the formation of provisional ECM by de novo synthe-sis, and finally the deposition of collagen-forming scar tissue.1
During scar maturation, inflammatory cells and fibroblastsdisappear as a result of apoptosis, and collagen becomes increas-ingly crosslinked to stabilize the scar and to provide tensilestrength until a hypocellular, collagen-rich scar is formed.2 Thecomposition of the ECM during the individual phases of infarct
healing not only determines the mechanical stability but alsomodulates the cellular phenotype.3 In the long term, adverse leftventricular (LV) remodeling, including fibrosis and ventriculardilatation, often causes progressive heart failure associated withconsiderable morbidity and mortality after acute myocardialinfarction (MI).
Clinical Perspective p 1276The role of MMPs during postinfarct remodeling has been
studied extensively. After MI, an imbalance between MMPs
Received May 8, 2007; accepted December 24, 2007.From the Abteilung für Kardiologie und Pneumologie, Charite-Universitätsklinikum Berlin, Campus Benjamin Franklin, Berlin, Germany (D.W., O.L.,
H.P.S., C.T.); Molecular Cardioprotection and Inflammation Group, Klinik für Anästhesiologie (J.M.) and Molekulare Pharmakologie, Institut fürPharmakologie und Klinische Pharmakologie (A.M., T.F., J.W.F.), Universitätsklinikum Düsseldorf, Düsseldorf, Germany; Center for CardiovascularResearch, Institut für Pharmakologie und Toxikologie, Campus Charite-Mitte, Charite-Universitatsmedizin Berlin, Berlin, Germany (C.P., T.U.);Nephropharmakologie, Allgemeine Pharmakologie und Toxikologie, Klinikum der Johann Wolfgang Goethe Universität Frankfurt, Frankfurt, Germany(L.S.); Institut für Anatomie, Christian-Albrechts-Universität Kiel, Kiel, Germany (R.L.-R.); Institut für Herzkreislauf-Physiologie, Heinrich-Heine-Universität Düsseldorf, Düsseldorf, Germany (C.J., J.S.); Leibniz-Institute for Arteriosclerosis Research, Department of Molecular Genetics ofCardiovascular Disease, University of Muenster, Muenster, Germany (S.-M.B.-H.); Craniofacial and Skeletal Diseases Branch, National Institute ofDental and Craniofacial Research, National Institutes of Health, Bethesda, Md (M.F.Y.); Institut für Pathophysiologie (B.L.) and Institut für Pathologie(H.A.B.), Universitätsklinikum Essen, Essen, Germany; and Molecular Cardioprotection and Inflammation Group, Department of Anaesthesia, UniversityHospital Bristol, Bristol Royal Infirmary, Bristol, UK (K.Z.).
*The first 4 authors contributed equally to this work.The online Data Supplement, which contains an expanded Methods section and figures, can be found with this article at http://circ.ahajournals.org/
cgi/content/full/CIRCULATIONAHA.107.714147/DC1.Correspondence to Jens W. Fischer, Molecular Pharmacology, Institut für Pharmakologie and Klinische Pharmakologie, Heinrich-Heine-Universität
Düsseldorf, Moorenstrasse 5, D-40225 Düsseldorf, Germany. E-mail [email protected]© 2008 American Heart Association, Inc.
Circulation is available at http://circ.ahajournals.org DOI: 10.1161/CIRCULATIONAHA.107.714147
1269
Heart Failure
at SWETS SUBSCR SVC/25338706 on September 15, 2008 circ.ahajournals.orgDownloaded from

and tissue inhibitors of MMPs (TIMPs) is thought to be a keyfactor in promoting adverse remodeling of the ECM and LVdilatation. However, collagen synthesis and fibrillogenesisalso underlie a complex regulation and might in turn beequally important during infarct healing. Self-assembly ofprocollagen � chains forms triple-helical collagen moleculesthat associate into collagen fibrils. This process is regulatedby accessory ECM molecules such as the family of smallleucine-rich proteoglycans, including biglycan and decorin.4
By controlling lateral assembly of triple helixes into fibrilsand the arrangement of collagen fibers, biglycan and decorinare key regulators of collagen fibrillogenesis and fibrildiameter.4,5 Biglycan and decorin are capable of specificbinding to collagen types I, II, III, and VI6,7 via the coreproteins near the d band of collagen fibrils.8,9 The role ofsmall leucine-rich proteoglycans in LV remodeling is onlypartially understood. It has been reported previously thatbiglycan and decorin are induced after MI in rats and thatbiglycan associates with cardiac fibroblasts in the infarctedmyocardium.10,11 bgn�/0 mice have reduced skeletal growthand develop an osteoporosis-like phenotype resulting from adefect in bone formation.12 Furthermore, the diameter ofcollagen fibrils was reduced in bgn�/0 mice.13 Other functionsof biglycan include regulation of transforming growth factor�-1 activity,14 cell proliferation, apoptosis,15 and activation ofinflammatory responses mediated through activation of Toll-like receptor-2 and -4.16 Taken together, biglycan playsimportant roles in collagen matrix assembly, control ofgrowth factor activity, and cellular phenotype, includinginflammatory cells. However, it is still unknown whetherbiglycan plays a role in cardiac physiology or during infarcthealing. The aim of the present study was to investigate therole of biglycan during remodeling and functional adaptationafter experimental MI. For this purpose, bgn�/0 mice weresubjected to permanent occlusion of the left anterior descend-ing coronary artery, followed by analysis of heart morphol-ogy, ECM, and hemodynamic function.
MethodsA detailed description of the methods and materials is available inthe online Data Supplement.
Animalsbgn�/0 mice (bred at the University of Dusseldorf) with a targeteddeletion of the bgn gene12 and male wild-type (WT; C57BL/6)littermates were compared in this study. All procedures were carriedout in accordance with the Association for Assessment and Accred-itation of Laboratory Animal Care guidelines and the Guide for theCare and Use of Laboratory Animals (Department of Health andHuman Services, National Institutes of Health, publication No.86-23) and were approved by the ethics and research boards of theUniversity of Düsseldorf. Animals (12 to 16 weeks old) wererandomized as indicated.
Myocardial InfarctionAt the age of 12 weeks, WT and bgn�/0 mice were anesthetized byinjection of pentobarbital (100 mg/kg IP). Mice were subjected topermanent left anterior descending coronary artery occlusion, fol-lowed by recovery for 7 or 21 days. All procedures were carried outas recently described in detail.17
Hemodynamic MeasurementsMice were anesthetized by injection of thiopental (125 mg/kg IP),intubated, and ventilated with a respirator (type 7025, Ugo Basile,Comerio, Italy). A 1.4F microconductance pressure catheter (ARIASPR-719, Millar Instruments Inc, Houston, Tex) was positioned inthe LV via the right carotid artery for continuous registration of LVpressure-volume loops in closed-chest animals.18 Systolic functionand myocardial contractility were quantified by LV end-systolicpressure, peak rate of rise in LV pressure (dP/dtmax), ejectionfraction, cardiac output, end-systolic volume, and stroke volume.Diastolic performance was measured by LV end-diastolic pressure,peak dP/dtmin, end-diastolic volume, and time constant of isovolu-mic pressure relaxation.
Analysis of Human MIsTissue sections derived from patients who died of ischemic MIpresenting either as an acute, primary event (�5 days, n�4)) orreoccurring infarction on the basis of old, healed MIs (n�4) wereanalyzed for biglycan by immunostaining (LF 51) and for collagenby Sirius red staining. The samples were collected for diagnosticpurposes during autopsy and were examined by a senior pathologist(H.B.). Informed consent was obtained from patients or relatives.The present study was performed according to the Declaration ofHelsinki, and the study protocol was approved by the EthicsCommittee of the University Hospital of Essen.
Statistical AnalysisFor multiple comparisons, 2-way ANOVA, followed by Bonferroniposttest, was performed. Values of P�0.05 were considered significant.Data are expressed as mean�SEM. Survival curves were computed by theKaplan-Meier method and were compared by use of the log-rank test.
The authors had full access to and take responsibility for theintegrity of the data. All authors have read and agree to themanuscript as written.
ResultsIncreased Biglycan Expression After MIReal-time reverse-transcription polymerase chain reaction(RT-PCR) revealed strongly increased bgn mRNA expressionin the LV after experimental MI, peaking 7 days after MI(Figure 1A). These results were confirmed by immunostain-ing for biglycan, showing that biglycan accumulation occursin the infarcted ischemic area and border zone at 7 days(Figure 1C). Furthermore, at 7 days after MI, fibroblasts andinflammatory cells were detected in areas enriched with
A
B C D
Figure 1. Time course of biglycan expression after MI. A, Real-time RT-PCR revealed significantly increased bgn mRNA expres-sion, peaking 7 days after MI. B, Immunohistochemistry of biglycanin LV of sham-operated WT mice (inset, negative control) and WTmice after left anterior descending coronary artery occlusion at 7(C) and 14 (D) days. n�6. Magnification �100. *P�0.05.
1270 Circulation March 11, 2008
at SWETS SUBSCR SVC/25338706 on September 15, 2008 circ.ahajournals.orgDownloaded from

biglycan (online Data Supplement), in line with previousresults in rats.10,11 At 14 days after MI, bgn mRNA levelswere still 5-fold higher than in sham animals (Figure 1A), andbiglycan immunostaining was still increased in the infarct andperi-infarct zones (Figure 1D).
Cardiac Phenotype of Adult bgn�/0 Micebgn�/0 mice were used as a model to study the effect ofbiglycan on cardiac function. First, the cardiac phenotype wasanalyzed in healthy, noninfarcted animals. At 6 months ofage, morphological and hemodynamic analyses of the heartdid not reveal any differences between WT and bgn�/0 mice.In 15-month-old mice, the body weight (WT, 48.5�4.5 gversus bgn�/0, 41.3�5.2 g [mean�SEM]; P�0.05) and ratioof heart to body weight (Data Supplement) were slightlydecreased. At 15 months, hemodynamic measurements inthese mice revealed no differences in LV end-diastolicpressure, end-diastolic volume, end-systolic pressure, end-systolic volume (not shown), and heart rate (Data Supple-ment). However, ejection fraction showed a trend towardlower values, and end-diastolic pressure-volume ratios weredecreased in bgn�/0 mice (end-diastolic pressure-volume ra-tio: WT, 0.32�0.11 mm Hg/�L; bgn�/0, 0.19�0.07 mm Hg/�L [mean�SEM]; n�6; P�0.05; Data Supplement). Inaddition, 15-month-old bgn�/0 mice showed a trend towarddecreased collagen type I mRNA expression but increasedcollagen type III and decorin expression as determined byreal-time RT-PCR (data not shown). Morphological analysisand collagen staining showed no obvious difference betweenWT and bgn�/0 mice at 15 months (Data Supplement),suggesting that both the morphology and cardiac ECMshowed no major difference in noninfarcted bgn�/0 mice.
Decreased Survival After MI in bgn�/0 MiceKaplan-Meier analysis revealed higher mortality in bgn�/0
mice after experimental MI. This increase in mortality wasobvious after 7 days and continued until 3 weeks at the end ofthe study. At 3 weeks, the survival rates were 60% and 20%in WT and bgn�/0 mice, respectively (Figure 2A). To inves-tigate the cause of death, an additional group of animals(n�8) underwent experimental MI and morphological anal-ysis 7 days after MI. During this 7-day period, the animalswere closely monitored, and mice that died underwent au-topsy immediately. Analysis of this group revealed that thefrequency of LV ruptures was �33% in WT and 66% inbgn�/0 mice, explaining the lower survival (Figure 2B). Figure3 shows that the area at risk and infarct size were not differentbetween the 2 genotypes as determined by Evans blue andp-nitro blue tetrazolium 7 days after MI. These data excludethe possibility that increased area at risk or infarct size isresponsible for the higher frequency of rupture.
Matrix Remodeling 7 Days After MITo investigate the mechanisms that caused LV ruptures, ECMremodeling was characterized in both groups. Collagen con-tent of infarct scars was similar between genotypes asdetermined by Sirius red staining (bright field, Sirius redexpressed as area fraction: WT, 21.9�6.7% versus bgn�/0,37.9�7%; n�4; P�1.6). In addition, immunohistochemistryof collagen type 1 revealed no quantitative differences be-tween groups (data not shown). In polarized light using the
birefringence analysis, Sirius red staining revealed increaseddensity of collagen in WT mice as indicated by the bright red(Figure 4C and 4D; Sirius red birefringence expressed as areafraction: bgn�/0, 3.9�1% versus WT, 10.8�1.7%; n�4;P�0.05). Furthermore, Movat’s pentachrome staining re-vealed a higher proportion of proteoglycans in scars of bgn�/0
mice at 7 days after MI (Figure 4E and 4F), which wascorroborated by Alcian blue staining and subsequent quanti-
A B
DC
Figure 3. Infarct size 7 days after MI. Area at risk (A; %LV) andinfarct size (B; % LV) were not different between the genotypes andcontrols (WT, n�8; bgn�/0, n�8). Representative Evans blue/p-nitroblue tetrazolium–stained sections of WT (C) or bgn�/0 (D) mice.
A
CB
Figure 2. Survival after experimental MI is decreased in bgn�/0
mice. A, Mortality after permanent left anterior descending coro-nary artery occlusion in WT and bgn�/0 mice up to 21 days afterMI. Mortality was significantly increased in bgn�/0 mice (P�0.01,log-rank-test; WT, n�20, 8 nonsurvivors; bgn�/0, n�33, 27 nonsur-vivors). B, bgn�/0 mice died more often as a result of ventricularrupture than their WT littermates within 7 days after MI. C, Heart ofa bgn�/0 mouse. Arrows/thread indicate the LV ruptured area.
Westermann et al Role of Biglycan During Post-MI Remodeling 1271
at SWETS SUBSCR SVC/25338706 on September 15, 2008 circ.ahajournals.orgDownloaded from

fication by image analysis (Alcian blue expressed as areafraction: WT, 73.3�4.4% versus bgn�/0, 87.5�3%; n�4;P�0.05). Decorin was upregulated after MI in WT mice asdescribed previously.19 Notably, post-MI immunostaining ofdecorin was much stronger in bgn�/0 mice (Figure 4G and4H). Taken together, the infarct scars of bgn�/0 mice werecharacterized by loosely packed collagen fibers and a higheramount of proteoglycans, including decorin.
The expression patterns of MMP-2, -8, -9, and -13 andTIMP-1, -2, and -4 were analyzed by immunostaining (Figure5) and by real-time RT-PCR (Data Supplement). MMP-2, -8,-9, and -13 and TIMP-1, -2, and -4 were strongly upregulated
in the infarcted area compared with the noninfarcted area in bothgenotypes (Figure 5). The expression of MMP-2 and -13 wassignificantly higher in the infarcted area of bgn�/0 mice com-pared with WT. However, MMP-9 was decreased and TIMP-1and -4 were significantly upregulated in bgn�/0 mice afterexperimental MI. The differences detected by immunostainingwere largely paralleled by mRNA expression (Data Supple-ment). In situ zymography indicating gelatinolytic activitymainly through MMP-2 and -920 revealed no significant differ-ence between genotypes (Figure 5P). Furthermore, the mRNAexpression of procollagen-prolyl-4-hydroxylase, procollagen-lysyl-hydroxylase 2, procollagen-lysyl-hydroxylase 1, and tissuetransglutaminase, key enzymes in the regulation of collagen andECM crosslinking, was not differentially regulated between thegenotypes after MI (Data Supplement).
Hemodynamic ParametersNo difference in cardiac hemodynamic parameters was de-tected between 16-week-old sham-operated WT and bgn�/0
animals (Table), confirming the analysis of WT versus bgn�/0
mice under normal conditions as mentioned above. In allanimals, as predicted, MI caused a decrease in ejectionfraction (%) and an increase in LV end-diastolic pressure(mm Hg), end-diastolic volume (�L), and end-systolic vol-ume (�L) (Figure 6). Additional hemodynamic parametersare presented in the Table, indicating severe impairment ofhemodynamic functions of WT and bgn�/0 mice 3 weeks afterMI such as a decrease in LV end-systolic pressure (mm Hg),dP/dtmax (mm Hg/s), and dP/dtmin (mm Hg/s) and increased� (ms). The most important finding is that bgn�/0 micedemonstrate significantly increased post-MI LV dilatationwith increased LV end-diastolic volume and end-systolicvolume and decreased ejection fraction compared with WTcontrols 21 days after MI (Figure 6A through 6D). Further-more, bgn�/0 mice demonstrated reduced stroke volume andcardiac output despite significantly increased heart rate (bpm)(Table), suggesting compensatory tachycardia. As an addi-tional sign of cardiac dilatation and failure, LV end-diastolic
WT bgn-/0C D
WT bgn-/0A B
WT bgn-/0E F
WT bgn-/0G H
Figure 4. ECM remodeling 7 days after MI. A, B, Sirius redstaining, bright field. C, D, Sirius red viewed under polarizedlight revealing less densely packed collagen in the infarct scarsof bgn�/0 mice. E, F, Movat’s pentachrome staining revealed aproteoglycan-rich ECM in bgn�/0 mice vs WT. G, H, Decorin im-munostaining (LF-113) was markedly increased in bgn�/0 mice.Representative stainings of WT (n�8) and bgn�/0 (n�8). Magnifi-cation �100.
Figure 5. Immunohistochemistry of MMPsand TIMPs. Immunostaining and quantifica-tion (area fraction [AF]) in the infarct scar 7days after MI of MMP-2 (A, B, and I), MMP-8(C, D, and J), MMP-9 (E, F, and K), andMMP-13 (G, H, and L). For TIMP-1 (M),TIMP-2 (N), and TIMP-4 (O), only the resultsof the image analysis (AF) are given. In thescar (MI), MMP-2, MMP-13, TIMP-1, andTIMP-4 were increased in bgn�/0 mice,whereas MMP-9 was decreased. P, Gelati-nolytic activity as determined by in situzymography (WT, n�5; bgn�/0, n�4).*P�0.05, ***P�0.001 vs WT-MI; #P�0.05 vsrespective noninfarcted regions.
1272 Circulation March 11, 2008
at SWETS SUBSCR SVC/25338706 on September 15, 2008 circ.ahajournals.orgDownloaded from

pressure was significantly increased compared with WTcontrols. This increase in LV end-diastolic pressure is furtherverified by increased ratios of lung wet weight to lung dryweight after MI compared with WT as an additional indicatorfor LV heart failure (Table). These data clearly indicate thatbgn�/0 mice develop impaired hemodynamic function inresponse to ischemic injury. In addition, a subset of animalswas investigated with MRI. The longitudinal MRI images(Figure 6E and 6F) showed stronger LV dilation and areconsistent with the hemodynamic data. Calculation of LVend-diastolic wall stress revealed increased wall stress inbgn�/0 mice after MI compared with WT (14.84�1.2�103
dynes · mm�2 [P�0.05 compared with WT MI] versus9.09�1.8�103 dynes · mm�2).
Matrix Remodeling 21 Days After MIIncreased frequency of cardiac ruptures and decreased hemo-dynamic function of bgn�/0 mice after MI and less tightlypacked collagen at 7 days (Figure 4) point toward perturbed
ECM remodeling in the infarct scar of bgn�/0 mice. Therefore,ECM remodeling also was characterized at 21 days after MI.MI induced a marked upregulation of collagen type 1 and 3mRNA as determined by real-time RT-PCR, which wassignificantly stronger in the bgn�/0 mice (Figure 7A). DecorinmRNA showed a trend toward upregulation in WT versusbgn�/0 mice 21 days after MI (Figure 7B). The overallcollagen content was similar as evidenced by Sirius redstaining (Figure 7C and 7D; total Sirius red expressed as areafraction: WT, 41�3.9% versus bgn�/0, 43.8�11.7%; n�4;P�0.05) and collagen type 1 immunostaining (not shown). Astriking difference appeared with respect to collagen organi-zation (Figure 7C and 7D). Under polarized light, Sirius redstaining revealed that the collagen present in the scar of bgn�/0
mice was characterized by a lower percentage of tightlypacked collagen (Figure 7E and 7F; Sirius red birefringenceexpressed as area fraction: WT, 11.1�0.3% versus bgn�/0,5.9�1.4%; n�4; P�0.05). Immunohistochemistry showedstill increased decorin accumulation (not shown); however,the difference was not as pronounced as detected at 7 days(Figure 4). Analysis of the collagen matrix by electronmicroscopy revealed tight and parallel packing of collagenfibrils in the scars of WT mice. In contrast, collagen fibrilorganization was perturbed in bgn�/0 mice (Figure 7H). Thetensile force to rupture LV rings ex vivo was determined toinvestigate whether the differences in matrix composition andorganization correlate with changes of tensile strength. Undercontrol conditions without MI, no difference was evidentbetween LV rings from WT and bgn�/0 mice that ruptured at84 and 90 mN, respectively (Figure 7I). However, 21 daysafter MI, the force to rupture was dramatically decreased inLV rings derived from bgn�/0 mice (69 mN) compared withWT mice (146 mN).
Biglycan Accumulation in Human MITissue sections derived from patients who died of ischemicMI presenting as either an acute, primary event or reoccurringacute infarction on the basis of old healed MI were examined.Figure 8A through 8D represents an area from an acute MIcharacterized by acute myocyte necrosis with eosinophiliccytoplasm and without visible cardiomyocyte nuclei. Hardlyany immunostaining for biglycan nor Sirius red staining ofcollagen was detected under these conditions. In contrast,Figure 8E through 8H depicts a healed infarction with fibroticregions and some remaining vital cardiomyocytes. In thesehealed infarcts, strong accumulation of both biglycan anddensely packed collagen as shown by birefringence analysis
Table. Hemodynamic Measurements at 21 Days After MI
HeartRate, bpm
End-SystolicPressure, mm Hg
dP/dtmax,mm Hg/s
dP/dtmin,mm Hg/s �, ms
StrokeVolume, �L
Cardiac Output,�L/min
Lung Wet/DryWeight
Sham
WT (n�7) 442�12 105�2 7554�818 �5231�485 13.8�0.7 36.6�3.4 13160�1560 4.48�0.1
bgn�/0 (n�8) 428�8 105�5 6890�345 �5763�429 14.0�0.7 35.4�3.2 13640�1728 4.56�0.1
MI
WT (n�10) 433�11 73�4† 4045�414† �3446�314† 17.4�0.8† 19.6�1.1† 8450�441† 5.5�0.2†
bgn�/0 (n�6) 489�25*† 64�2† 3240�406† �2757�346† 17.0�1.4 14.1�1.2† 6723�716† 6.4�0.2*†
*P�0.05, WT vs bgn�/0; †P�0.05, sham vs MI.
Figure 6. Hemodynamic parameters 21 days after MI. A throughD, Hemodynamic measurements in WT vs bgn�/0 under controlconditions (sham) and 21 days after MI. After MI, bgn�/0 micewere characterized by decreased ejection fraction (EF; A) andincreased LV end-diastolic pressure (Ped; B), LV end-diastolicvolume (Ved; C), and LV end-systolic volume (Ves; D) (shamWT, n�7; sham bgn�/0, n�8; MI WT, n�10; MI bgn�/0; n�6).Additional hemodynamic parameters are given in the Table.E and F, MRI representative longitudinal sections showingenhanced LV dilatation in bgn�/0 mice (F; n�7) vs WT (E; n�8)at 21 days after MI. Arrows point toward an area of markedventricular wall thinning. *P�0.05, ***P�0.001, #P�0.05 vsrespective noninfarcted controls.
Westermann et al Role of Biglycan During Post-MI Remodeling 1273
at SWETS SUBSCR SVC/25338706 on September 15, 2008 circ.ahajournals.orgDownloaded from
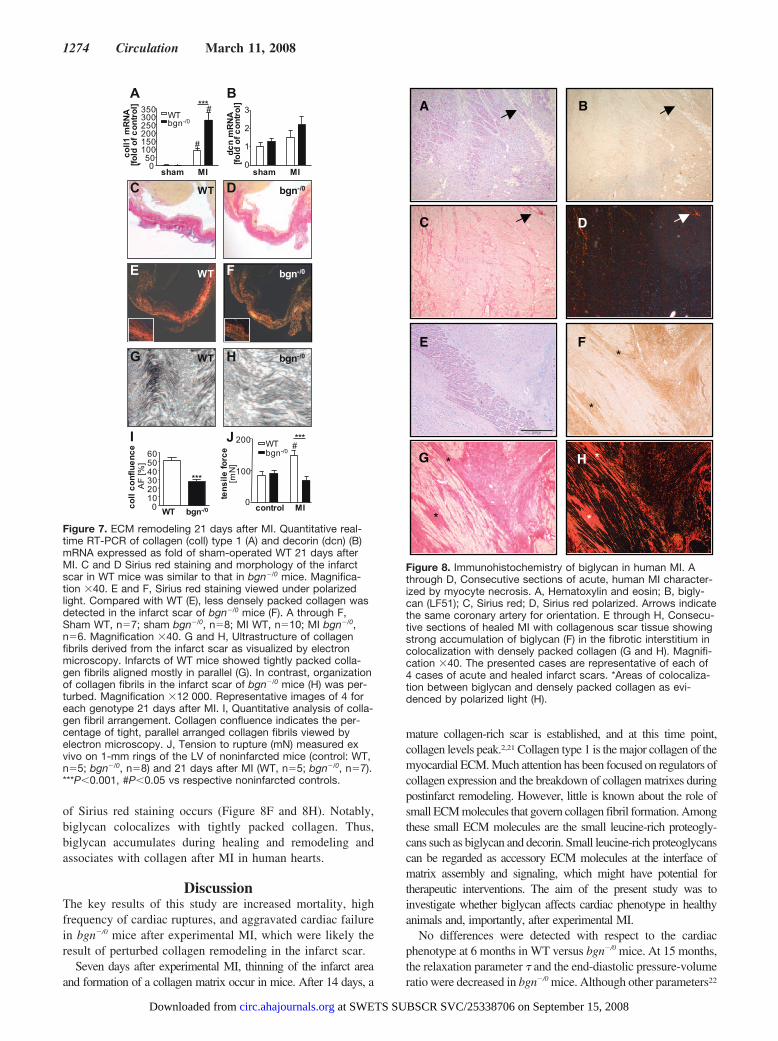
of Sirius red staining occurs (Figure 8F and 8H). Notably,biglycan colocalizes with tightly packed collagen. Thus,biglycan accumulates during healing and remodeling andassociates with collagen after MI in human hearts.
DiscussionThe key results of this study are increased mortality, highfrequency of cardiac ruptures, and aggravated cardiac failurein bgn�/0 mice after experimental MI, which were likely theresult of perturbed collagen remodeling in the infarct scar.
Seven days after experimental MI, thinning of the infarct areaand formation of a collagen matrix occur in mice. After 14 days, a
mature collagen-rich scar is established, and at this time point,collagen levels peak.2,21 Collagen type 1 is the major collagen of themyocardial ECM. Much attention has been focused on regulators ofcollagen expression and the breakdown of collagen matrixes duringpostinfarct remodeling. However, little is known about the role ofsmall ECM molecules that govern collagen fibril formation. Amongthese small ECM molecules are the small leucine-rich proteogly-cans such as biglycan and decorin. Small leucine-rich proteoglycanscan be regarded as accessory ECM molecules at the interface ofmatrix assembly and signaling, which might have potential fortherapeutic interventions. The aim of the present study was toinvestigate whether biglycan affects cardiac phenotype in healthyanimals and, importantly, after experimental MI.
No differences were detected with respect to the cardiacphenotype at 6 months in WT versus bgn�/0 mice. At 15 months,the relaxation parameter � and the end-diastolic pressure-volumeratio were decreased in bgn�/0 mice. Although other parameters22
Figure 7. ECM remodeling 21 days after MI. Quantitative real-time RT-PCR of collagen (coll) type 1 (A) and decorin (dcn) (B)mRNA expressed as fold of sham-operated WT 21 days afterMI. C and D Sirius red staining and morphology of the infarctscar in WT mice was similar to that in bgn�/0 mice. Magnifica-tion �40. E and F, Sirius red staining viewed under polarizedlight. Compared with WT (E), less densely packed collagen wasdetected in the infarct scar of bgn�/0 mice (F). A through F,Sham WT, n�7; sham bgn�/0, n�8; MI WT, n�10; MI bgn�/0,n�6. Magnification �40. G and H, Ultrastructure of collagenfibrils derived from the infarct scar as visualized by electronmicroscopy. Infarcts of WT mice showed tightly packed colla-gen fibrils aligned mostly in parallel (G). In contrast, organizationof collagen fibrils in the infarct scar of bgn�/0 mice (H) was per-turbed. Magnification �12 000. Representative images of 4 foreach genotype 21 days after MI. I, Quantitative analysis of colla-gen fibril arrangement. Collagen confluence indicates the per-centage of tight, parallel arranged collagen fibrils viewed byelectron microscopy. J, Tension to rupture (mN) measured exvivo on 1-mm rings of the LV of noninfarcted mice (control: WT,n�5; bgn�/0, n�8) and 21 days after MI (WT, n�5; bgn�/0, n�7).***P�0.001, #P�0.05 vs respective noninfarcted controls.
A B
C D
E F
G H
Figure 8. Immunohistochemistry of biglycan in human MI. Athrough D, Consecutive sections of acute, human MI character-ized by myocyte necrosis. A, Hematoxylin and eosin; B, bigly-can (LF51); C, Sirius red; D, Sirius red polarized. Arrows indicatethe same coronary artery for orientation. E through H, Consecu-tive sections of healed MI with collagenous scar tissue showingstrong accumulation of biglycan (F) in the fibrotic interstitium incolocalization with densely packed collagen (G and H). Magnifi-cation �40. The presented cases are representative of each of4 cases of acute and healed infarct scars. *Areas of colocaliza-tion between biglycan and densely packed collagen as evi-denced by polarized light (H).
1274 Circulation March 11, 2008
at SWETS SUBSCR SVC/25338706 on September 15, 2008 circ.ahajournals.orgDownloaded from

may influence the end-diastolic pressure-volume ratio, thisfinding suggests that the intrinsic myocardial stiffness is reducedin aged bgn�/0 mice. Taken together, these findings suggest thatduring physiological conditions bgn deficiency has only minoreffects on LV function, which starts to induce hemodynamicchanges only during aging.
After experimental MI, biglycan expression was stronglyupregulated in WT mice, as shown previously in rats.10,11
Interestingly, the upregulation was transient, peaking at 7days, at the same time that collagen content also is highest.2
Using immunohistochemistry, we found biglycan in theinfarct scar and border zone of the infarct at 7 and 14 daysafter MI, in line with a role of biglycan in collagen remod-eling. During the whole experimental period, bgn�/0 miceshowed higher mortality after MI, which was due in large partto LV ruptures. In addition, spontaneous death withoutruptures occurred more often in bgn�/0 mice. This might be aconsequence of aggravated cardiac failure and LV dilatationas indicated by increased LV end-diastolic volume, end-dia-stolic pressure, and ratios of wet lung weight to dry lungweight in bgn�/0 mice after MI.
bgn�/0 mice and WT mice showed no differences in thearea at risk and infarct size, which excludes the possibilitythat decreased survival of bgn�/0 mice after MI is a consequenceof increased ischemia or necrosis. Birefringence analysis ofSirius red–stained sections revealed that the packing of collagenfibrils was disturbed in bgn�/0 mice 7 and 21 days after MI. Thiswas confirmed by analysis of the ultrastructure of collagen inthe infarct scar, showing that the array of collagen washeavily perturbed in bgn�/0 mice. The quantity of collagenaccumulating in the scar was not decreased as analyzed byimmunohistochemistry of collagen type 1 and Sirius redstaining. In contrast, mRNA expression of collagen type 1and collagen type 3 was increased even in bgn�/0 mice, whichmight represent a compensatory mechanism.
Increased MMP activity is known to contribute to ventric-ular dilatation after MI, and expression of MMPs is highlyincreased in the infarcted area.23,24 The upregulation ofMMP-2 and MMP-13 was opposed by upregulation ofTIMP-1 and TIMP-4 and downregulation of MMP-9. MMP-9is a key player in adverse ventricular remodeling after MIbecause MMP-9–dependent collagen degradation is associ-ated with adverse LV myocardial remodeling in animalmodels and patients.25,26 An additional mechanism of how theabsence of biglycan could increase collagen cleavage isconceivable. It was recently demonstrated in vitro that thenoncovalent binding of decorin and biglycan to fibrillarcollagen type 1 inhibits the cleavage of collagen type 1 bycollagenase-1 (MMP-1) and collagenase-3 (MMP-13).27 Theinteraction of biglycan core protein with collagen fibrilsrenders collagen type 1 fibrils less accessible to MMP-mediated cleavage. Of note, biglycan inhibited collagenbreakdown without changing the activation or expression ofMMPs, which would obscure this mechanism even afteranalysis of the MMP/TIMP system as we performed in thepresent study. Taken together, it is conceivable that changes inthe MMP/TIMP levels and/or increased susceptibility to break-down in the absence of biglycan contribute to the cardiacphenotype of bgn�/0 mice described in the present study.
Taken together, in bgn�/0 mice, disturbed collagen matrixassembly might result in decreased mechanical strength ofmyocardial scar tissue after MI. Indeed, ex vivo measurementsrevealed severely reduced tensile strength of LV rings derivedfrom bgn�/0 mice compared with WT mice. The reduced tensilestrength of LV and the increase in LV end-diastolic wall stressmight explain the frequent ruptures in bgn�/0 mice.
Interestingly, the dcn�/� mice also have a strong cardiacphenotype after experimental MI.19 dcn�/� mice developedimpaired LV function after experimental MI that was attributedto abnormal collagen fibril formation. However, ventricularruptures did not occur in dcn�/� mice.19 Therefore, from thepresent study, it appears that the cardiac phenotype is evenstronger in bgn�/0 mice compared with dcn�/� mice. Further-more, upregulation of decorin as observed in bgn�/0 mice 7 daysafter MI was not sufficient to compensate for the loss of biglycanas it does with respect to the osteoporosis phenotype.28
Analysis of scars of MI from humans revealed a strongaccumulation of biglycan in close association with collagenin fibrotic areas of healed scars, whereas biglycan was notdetected after acute human MI. It is therefore possible that thepresent findings are of clinical relevance and that biglycanmight participate in the evolution of a collagenous ECMduring healing of human MI. However, the present data donot allow a conclusion to be drawn as to whether biglycandeposition is characteristic for postinfarct remodeling or isassociated with fibrotic remodeling and hypertrophy in gen-eral. LV wall rupture as observed in bgn�/0 mice also is acomplication of acute MI in patients that is responsible for�20% of infarct-related deaths.29,30 Therefore, it might be ofinterest to investigate the patterns of biglycan expression inhuman MI and the relation to cardiac rupture in future studies.From the present study, it is obvious that the complete absenceof biglycan is deleterious and prevents the establishment of astable infarct scar and hemodynamic adaptation, suggesting thatbiglycan is required for proper infarct healing. However, futurestudies should address how much biglycan is needed for properscar formation and whether too much biglycan might also haveadverse effects such as enhanced fibrosis.
Sources of FundingThis study was in part supported by Deutsche Forschungsgemein-schaft SFB612, SFB TR19, GRK1089, and GRK 754; the Interdis-ciplinary Center for Clinical Research, Münster; and the ExcellenceCluster Cardio-Pulmonary System.
DisclosuresNone.
References1. Welch MP, Odland GF, Clark RA. Temporal relationships of F-actin
bundle formation, collagen and fibronectin matrix assembly, andfibronectin receptor expression to wound contraction. J Cell Biol. 1990;110:133–145.
2. Dobaczewski M, Bujak M, Zymek P, Ren G, Entman ML, FrangogiannisNG. Extracellular matrix remodeling in canine and mouse myocardialinfarcts. Cell Tissue Res. 2006;324:475–488.
3. Gray AJ, Bishop JE, Reeves JT, Laurent GJ. A alpha and B beta chainsof fibrinogen stimulate proliferation of human fibroblasts. J Cell Sci.1993;104(pt 2):409–413.
4. Danielson KG, Baribault H, Holmes DF, Graham H, Kadler KE, IozzoRV. Targeted disruption of decorin leads to abnormal collagen fibrilmorphology and skin fragility. J Cell Biol. 1997;136:729–743.
Westermann et al Role of Biglycan During Post-MI Remodeling 1275
at SWETS SUBSCR SVC/25338706 on September 15, 2008 circ.ahajournals.orgDownloaded from

5. Corsi A, Xu T, Chen XD, Boyde A, Liang J, Mankani M, Sommer B, Iozzo RV,Eichstetter I, Robey PG, Bianco P, Young MF. Phenotypic effects of biglycandeficiency are linked to collagen fibril abnormalities, are synergized by decorindeficiency, and mimic Ehlers-Danlos-like changes in bone and other connectivetissues. J Bone Miner Res. 2002;17:1180–1189.
6. Douglas T, Heinemann S, Bierbaum S, Scharnweber D, Worch H. Fibril-logenesis of collagen types I, II, and III with small leucine-rich proteo-glycans decorin and biglycan. Biomacromolecules. 2006;7:2388–2393.
7. Wiberg C, Hedbom E, Khairullina A, Lamande SR, Oldberg A, Timpl R,Morgelin M, Heinegard D. Biglycan and decorin bind close to the n-terminalregion of the collagen VI triple helix. J Biol Chem. 2001;276:18947–18952.
8. Vogel KG, Paulsson M, Heinegard D. Specific inhibition of type I andtype II collagen fibrillogenesis by the small proteoglycan of tendon.Biochem J. 1984;223:587–597.
9. Schonherr E, Witsch-Prehm P, Harrach B, Robenek H, Rauterberg J,Kresse H. Interaction of biglycan with type I collagen. J Biol Chem.1995;270:2776–2783.
10. Doi M, Kusachi S, Murakami T, Ninomiya Y, Murakami M, Nakahama M,Takeda K, Komatsubara I, Naito I, Tsuji T. Time-dependent changes ofdecorin in the infarct zone after experimentally induced myocardial infarctionin rats: comparison with biglycan. Pathol Res Pract. 2000;196:23–33.
11. Yamamoto K, Kusachi S, Ninomiya Y, Murakami M, Doi M, Takeda K,Shinji T, Higashi T, Koide N, Tsuji T. Increase in the expression ofbiglycan mRNA expression co-localized closely with that of type Icollagen mRNA in the infarct zone after experimentally-induced myo-cardial infarction in rats. J Mol Cell Cardiol. 1998;30:1749–1756.
12. Xu T, Bianco P, Fisher LW, Longenecker G, Smith E, Goldstein S,Bonadio J, Boskey A, Heegaard AM, Sommer B, Satomura K,Dominguez P, Zhao C, Kulkarni AB, Robey PG, Young MF. Targeteddisruption of the biglycan gene leads to an osteoporosis-like phenotype inmice. Nat Genet. 1998;20:78–82.
13. Ameye L, Aria D, Jepsen K, Oldberg A, Xu T, Young MF. Abnormalcollagen fibrils in tendons of biglycan/fibromodulin-deficient mice leadto gait impairment, ectopic ossification, and osteoarthritis. FASEB J.2002;16:673–680.
14. Hildebrand A, Romaris M, Rasmussen LM, Heinegard D, Twardzik DR,Border WA, Ruoslahti E. Interaction of the small interstitial proteo-glycans biglycan, decorin and fibromodulin with transforming growthfactor beta. Biochem J. 1994;302:527–534.
15. Schaefer L, Beck KF, Raslik I, Walpen S, Mihalik D, Micegova M,Macakova K, Schonherr E, Seidler DG, Varga G, Schaefer RM, KresseH, Pfeilschifter J. Biglycan, a nitric oxide-regulated gene, affectsadhesion, growth, and survival of mesangial cells. J Biol Chem. 2003;278:26227–26237.
16. Schaefer L, Babelova A, Kiss E, Hausser HJ, Baliova M, KrzyzankovaM, Marsche G, Young MF, Mihalik D, Gotte M, Malle E, Schaefer RM,Grone HJ. The matrix component biglycan is proinflammatory andsignals through Toll-like receptors 4 and 2 in macrophages. J Clin Invest.2005;115:2223–2233.
17. Petzelbauer P, Zacharowski PA, Miyazaki Y, Friedl P, Wickenhauser G,Castellino FJ, Groger M, Wolff K, Zacharowski K. The fibrin-derivedpeptide Bbeta15-42 protects the myocardium against ischemia-reperfusion injury. Nat Med. 2005;11:298–304.
18. Westermann D, Knollmann BC, Steendijk P, Rutschow S, Riad A,Pauschinger M, Potter JD, Schultheiss HP, Tschope C. Diltiazemtreatment prevents diastolic heart failure in mice with familial hyper-trophic cardiomyopathy. Eur J Heart Fail. 2006;8:115–121.
19. Weis SM, Zimmerman SD, Shah M, Covell JW, Omens JH, Ross J Jr, Dalton N,Jones Y, Reed CC, Iozzo RV, McCulloch AD. A role for decorin in theremodeling of myocardial infarction. Matrix Biol. 2005;24:313–324.
20. Frederiks WM, Mook OR. Metabolic mapping of proteinase activity withemphasis on in situ zymography of gelatinases: review and protocols.J Histochem Cytochem. 2004;52:711–722.
21. Cleutjens JP, Verluyten MJ, Smiths JF, Daemen MJ. Collagenremodeling after myocardial infarction in the rat heart. Am J Pathol.1995;147:325–338.
22. Burkhoff D, Mirsky I, Suga H. Assessment of systolic and diastolicventricular properties via pressure-volume analysis: a guide for clinical,translational, and basic researchers. Am J Physiol Heart Circ Physiol.2005;289:H501–H512.
23. Mukherjee R, Brinsa TA, Dowdy KB, Scott AA, Baskin JM, DeschampsAM, Lowry AS, Escobar GP, Lucas DG, Yarbrough WM, Zile MR,Spinale FG. Myocardial infarct expansion and matrix metalloproteinaseinhibition. Circulation. 2003;107:618–625.
24. Peterson JT, Li H, Dillon L, Bryant JW. Evolution of matrix metallo-protease and tissue inhibitor expression during heart failure progression inthe infarcted rat. Cardiovasc Res. 2000;46:307–315.
25. Ducharme A, Frantz S, Aikawa M, Rabkin E, Lindsey M, Rohde LE,Schoen FJ, Kelly RA, Werb Z, Libby P, Lee RT. Targeted deletion ofmatrix metalloproteinase-9 attenuates left ventricular enlargement andcollagen accumulation after experimental myocardial infarction. J ClinInvest. 2000;106:55–62.
26. Webb CS, Bonnema DD, Ahmed SH, Leonardi AH, McClure CD, ClarkLL, Stroud RE, Corn WC, Finklea L, Zile MR, Spinale FG. Specifictemporal profile of matrix metalloproteinase release occurs in patientsafter myocardial infarction: relation to left ventricular remodeling.Circulation. 2006;114:1020 –1027.
27. Geng Y, McQuillan D, Roughley PJ. SLRP interaction can protect collagenfibrils from cleavage by collagenases. Matrix Biol. 2006;25:484–491.
28. Young MF, Bi Y, Ameye L, Chen XD. Biglycan knockout mice: newmodels for musculoskeletal diseases. Glycoconj J. 2002;19:257–262.
29. Moreno R, Lopez de Sa E, Lopez-Sendon JL, Garcia E, Soriano J,Abeytua M, Elizaga J, Botas J, Rubio R, Moreno M, Garcia-FernandezMA, Delcan JL. Frequency of left ventricular free-wall rupture in patientswith acute myocardial infarction treated with primary angioplasty.Am J Cardiol. 2000;85:757–760.
30. Hutchins KD, Skurnick J, Lavenhar M, Natarajan GA. Cardiac rupture inacute myocardial infarction: a reassessment. Am J Forensic Med Pathol.2002;23:78–82.
CLINICAL PERSPECTIVEKnown mechanisms leading to heart failure after myocardial infarction include subsequent cardiac dilatation and infarctthinning, also called infarct expansion, which may lead to the development of aneurysm and left ventricle rupture.Therefore, adaptive changes in cardiac remodeling are essential so that the ventricle forms a stable scar rather thanrupturing after myocardial infarction. Collagenous networks elaborated by cardiac fibroblasts provide the mechanicalsupport for the scar tissue. It seems important to consider that the stability and 3-dimensional arrangement of cardiaccollagen matrixes are controlled by accessory molecules such as biglycan. Biglycan is a proteoglycan that binds to the dbands of collagen type 1 fibrils and controls fibrillogenesis and proteolysis of collagen by matrix metalloproteinases. Thepresent study reveals that after myocardial infarction, the scar tissue of biglycan-deficient mice was characterized byperturbed collagen fibril arrangement and that ventricular ruptures occur more frequently compared with wild-type mice.Furthermore, the surviving biglycan-deficient mice showed impaired hemodynamic function with subsequent cardiac dilatation,suggesting that in the absence of biglycan, adverse ventricular remodeling is aggravated. Therefore, biglycan appears to play acritical role in the formation of stable infarct scars and the preservation of hemodynamic function after myocardial infarction.Because biglycan accumulates in the scar tissue of human ischemic infarctions, it is possible that the present findings extend tohuman postinfarct remodeling. In the future, it will be of interest to determine whether differences in cardiac biglycan expressionmay be modulated by pharmacological treatment and whether this may change clinical outcome.
1276 Circulation March 11, 2008
at SWETS SUBSCR SVC/25338706 on September 15, 2008 circ.ahajournals.orgDownloaded from