bio 127 - section i introduction to developmental biology cell-cell communication in development...
Post on 19-Dec-2015
221 views
TRANSCRIPT

Bio 127 - Section IIntroduction to Developmental Biology
Cell-Cell Communication in DevelopmentGilbert 9e – Chapter 3

• It has to be EXTREMELY well coordinated for the single-celled fertilized ovum to develop into the complex adult
• This coordination requires a systematic way for the cells to know what’s happening around them so that they can change their gene expression correctly
• They must also then change the signals they are sending out to let surrounding cells know what changes they are making

Developmental Activities Coordinated in this Way
1. The formation of tissues from a mix of individual cells
2. The formation of organs from a mix of tissue types
3. The formation of cells, tissues and organs in specific locations
4. The growth and death of cells, tissues and organs
5. The achievement of polarity in cells, tissue and organs

• The plasma membranes of cells are designed to sense what is happening in their environment
• Membrane molecules sense:– other cell membranes– soluble signals sent by other cells– the type of extracellular matrix that surrounds them
• A few signals can get past the plasma membrane

Most cells in the embryo have molecules on their surface that identify who they are
These molecules also instruct themwho they should be in contact with

Sorting out and reconstruction of spatial relationships in aggregates of embryonic amphibian cells
Allcelltypes can do it
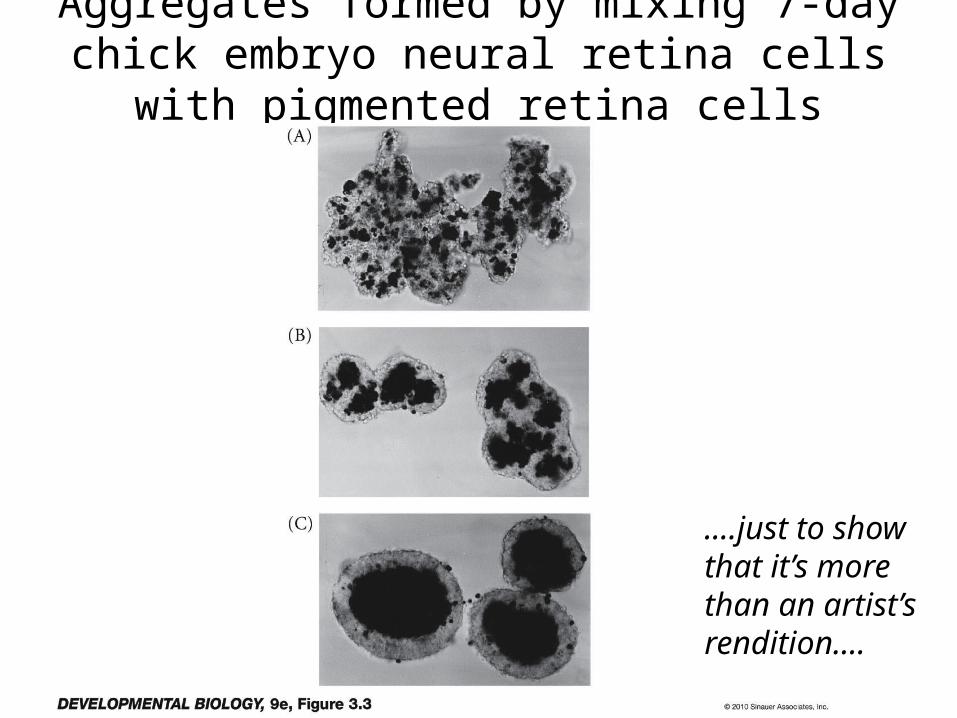
Aggregates formed by mixing 7-day chick embryo neural retina cells with pigmented retina cells
....just to showthat it’s morethan an artist’srendition....

Figure 3.4 Hierarchy of cell sorting in order of decreasing surface tensions
The moreadhesive the cell’s plasma membrane is,the more it migrates to the middle ofa cell mixture.

The molecular biology of cell adhesion: Cadherins
The calcium-dependentadhesion molecules(or cadherins) are the main source of adhesiveactivity on the cell surface
The more you express,the more central youbecome in a mixture

--- A nearly perfect linear relationship
Importance of amount of cadherin for morphogenesis
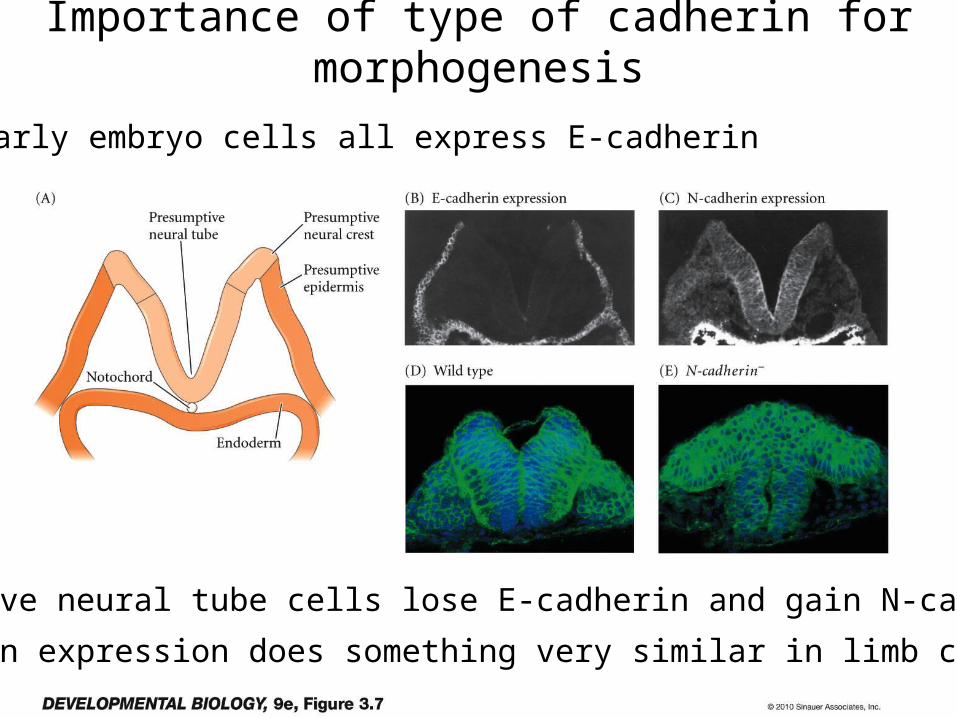
Early embryo cells all express E-cadherin
Presumptive neural tube cells lose E-cadherin and gain N-cadherin.
N-cadherin expression does something very similar in limb cartilage.
Importance of type of cadherin for morphogenesis
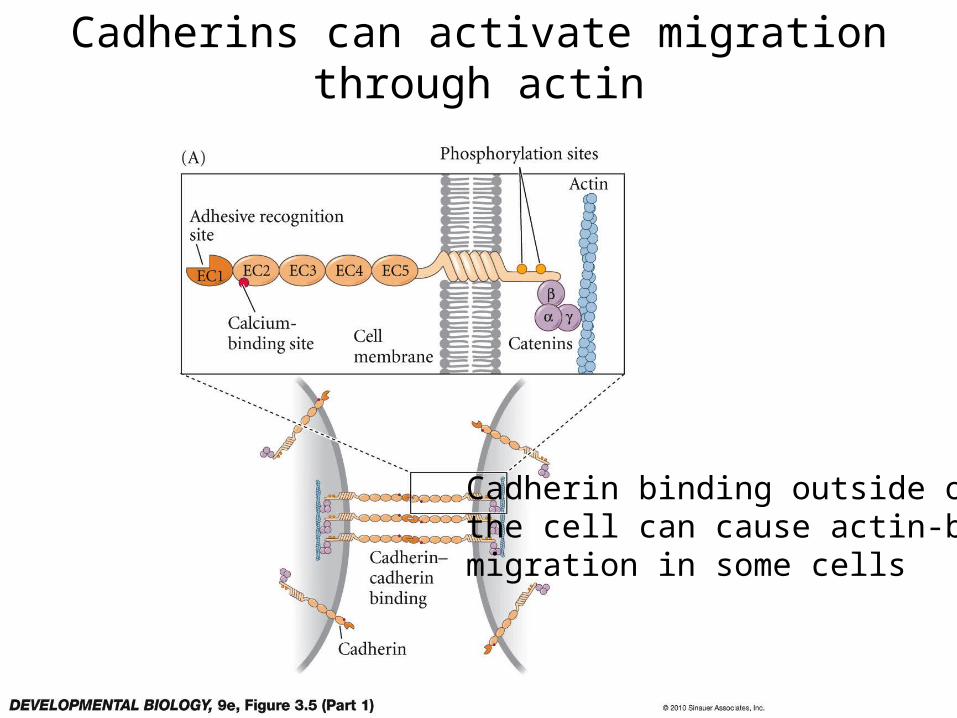
Cadherins can activate migration through actin
Cadherin binding outside of the cell can cause actin-basedmigration in some cells

Disruption of N-Cadherin in Frog Embryos
failed migration failed actin assembly
normalblocked
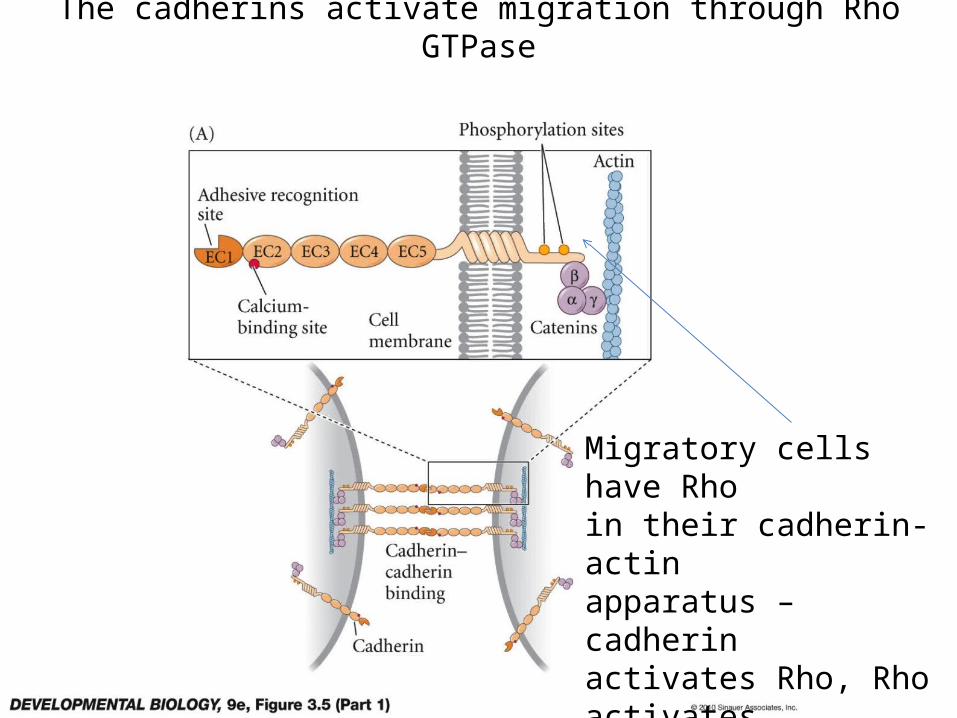
The cadherins activate migration through Rho GTPase
Migratory cells have Rhoin their cadherin-actinapparatus – cadherinactivates Rho, Rho activatesactin-myosin migration.

Drosophila gastrulation
The cells that have Rho activatedmigrate to become the mesoderm.

Rho build-up on E-cad causes actin polymerization and migration
Migration is started by expression of Twist and Snailwhich causes Rho and B-catenin to translocate in cells

Tracheal Development in Drosophila
Rho can also be linked to cellsurface receptors and causechemotactic migration.
The cadherin attachmentsremain strong and the cellsmigrate as a cohesive unit.

Figure 3.12 Cell migration
- not always cadherin-dependent however!
Mesenchymal Cell Migration is also Rho-Dependent

One-Way, Two-Way and Reciprocal Communications Strategies
Ligands
Receptors
2nd Messengers
Target Mechanisms

Cell Signaling Terminology
• Paracrine • Endocrine • Synaptic • Induction • Inducer • Responder • Signal Competence • Signal Transduction • Permissive Signals• Instructive Signals

Ectodermal competence and the ability to respond to the optic vesicle inducer in Xenopus

• Optic vesicle secretes......– BMP 4– Fgf 8
• Head ectoderm expresses......– Sox 2– L-Maf– Pax 6
• Lens genes turned on......– crystallin– others
HOW?

Induced Differentiation

Induction Cascades
• We know that tissues tend to aggregate through cell contact
• It’s common for tissues to play off each other to produce an organ
• Anything from two tissues signaling back and forth to many tissues coordinating each other’s actions
• The common theme is that a change in gene expression internally (TF’s, functional proteins) is often accompanied by a change in secreted proteins (paracrine, endocrine factors)

Eye formation is a classically studied cascade of induction
Simple lens induction...

Reciprocal induction



The reciprocal interaction between an epithelium and a closely associated mesenchyme is a very common means of organ development (organogenesis)

These arebackwards

Different mesenchyme induces different epithelial structures

Also, different epithelium can only become what they are competent to become

A little about some of the actual molecules.....
• Growth Factors carry most of the signals– Hormones and neurotransmitters later on– 4 big families:
• FGF, Hedgehog, Wnt, TGF-b
• Receptors and signaling cascades premade for them make you competent

• Fibroblast growth factor (FGF) family are classic growth factors: FGF 1-8– lots of others: VEGF, HGF, PDGF, etc.– can change transcription of genes 2 ways
• RTK Pathway: receptor tyrosine kinase• JAK-STAT: JAK activates STAT TF’s

• Optic vesicle secretes......– BMP 4– Fgf 8
• Head ectoderm expresses......– Sox 2– L-Maf– Pax 6
• Lens genes turned on......– crystallin– others
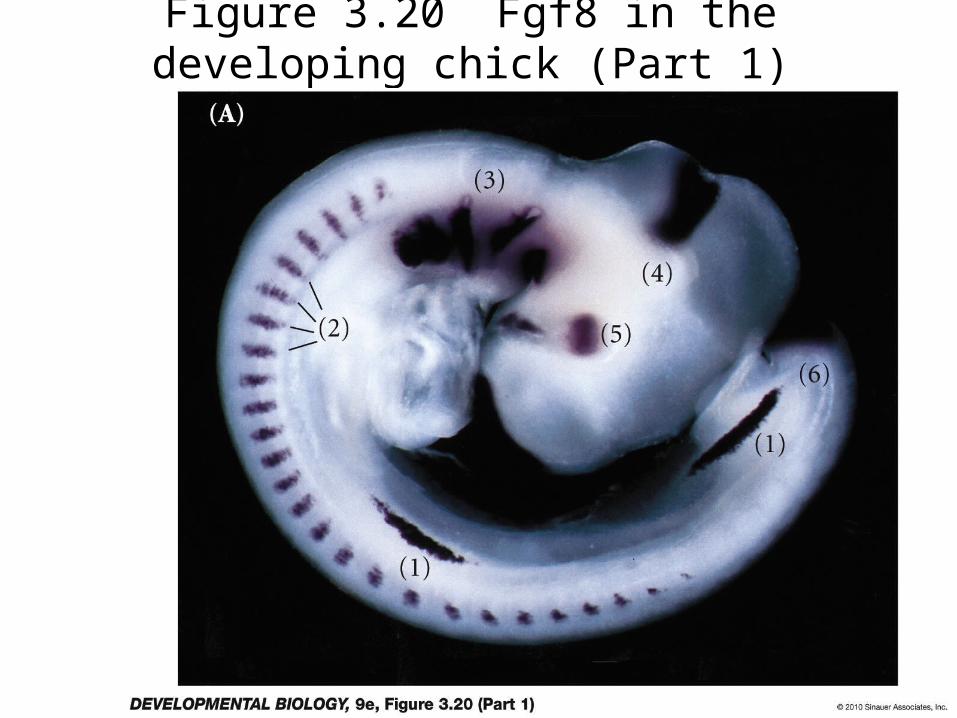
Figure 3.20 Fgf8 in the developing chick (Part 1)

Figure 3.20 Fgf8 in the developing chick (Part 2)
L-Maf expression in ectodermFGF8 in optic vesicle

Figure 3.24 A mutation in the gene for FgfR3 causes the premature constitutive activation of the STAT pathway and
the production of phosphorylated Stat1 protein

• Hedgehog family includes sonic (shh), desert (dhh) and indian (ihh) in vertebrates
• Change transcription through an interesting series of inhibitory activities– the patched receptor inhibits the smoothened
protein until hedgehog binds– smoothened then moves to inhibit proteins that
inhibit the Gli activator protein

Both shh and patched proteins require cholesterol.Blocking its production can cause cyclopism.
Figure 3.26 (A) Sonic hedgehog expression is shown by in situ hybridization in the nervous system, gut, and limb bud of a
chick embryo. (B) Head of a cyclopic lamb

• Wnt family has 15 members in vertebrates– Glycoproteins with lipid tails!– Work through frizzled receptors and disheveled
activators (fly guys!)– Also activate by inhibition of an inhibitor

• Interestingly, Wnt can do much of what cadherins can do
– Send catenins to the nucleus
– Activate rho and change the cytoskeleton
– This is called “crosstalk” and it is very important in cell signaling
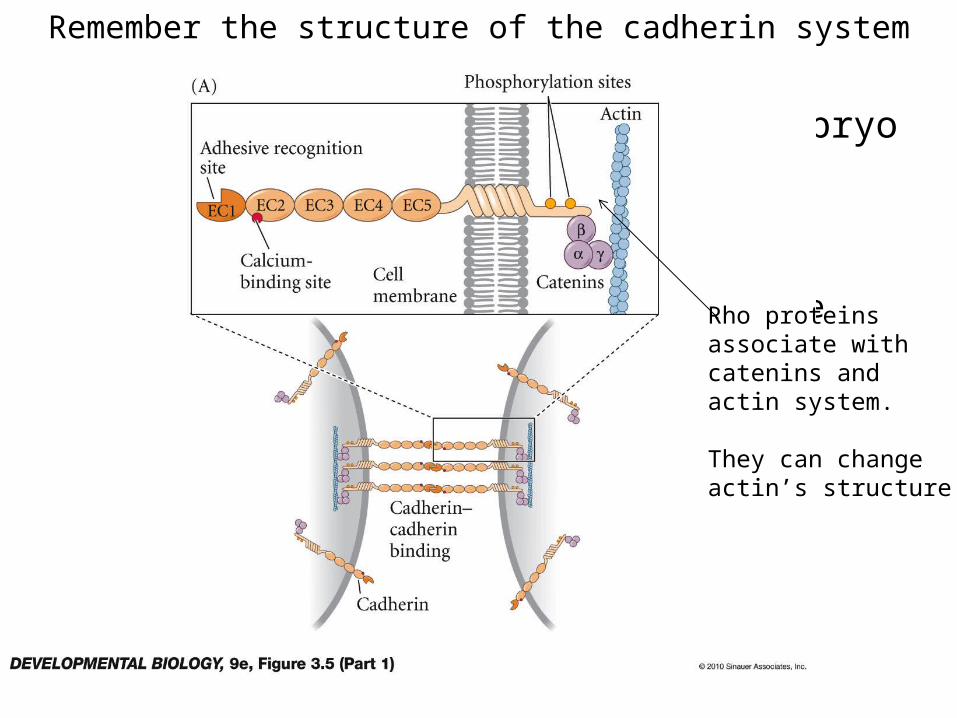
Remember the structure of the cadherin system
• Implantation of the mammalian embryo in adhere to the uterine wall– E- and P-cadherin– Integrin and uterine ECM– Proteins that bind sugars on uterine wall Rho proteins
associate withcatenins andactin system.
They can changeactin’s structure.

• The TGF-b superfamily is a very large family of very active peptide growth factors– involved in the development of most tissues
• The receptors are also a large family of proteins– They are serine-threonine kinases, not RTK
• They work through the activation of SMAD transcription factors
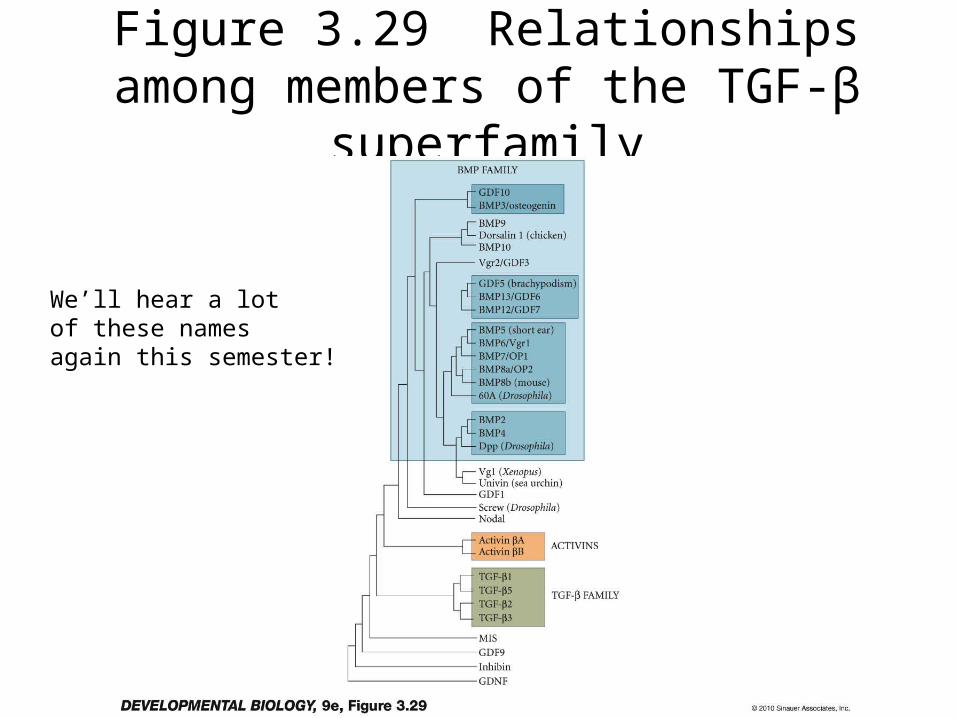
Figure 3.29 Relationships among members of the TGF-β superfamily
We’ll hear a lotof these namesagain this semester!

• Remember:
– The Big 4 are just part of the story
– We’ll talk about others as they come into play

The Delta-Notch family: “Juxtacrine” signals
– Transmembrane proteins on cells in contact– Delta, Jagged or Serrate bind to Notch family– Signals go both ways
– The Notch signal is interesting in that it’s internal domain is cleaved and enters nucleus
– This activates a dormant transcription factor
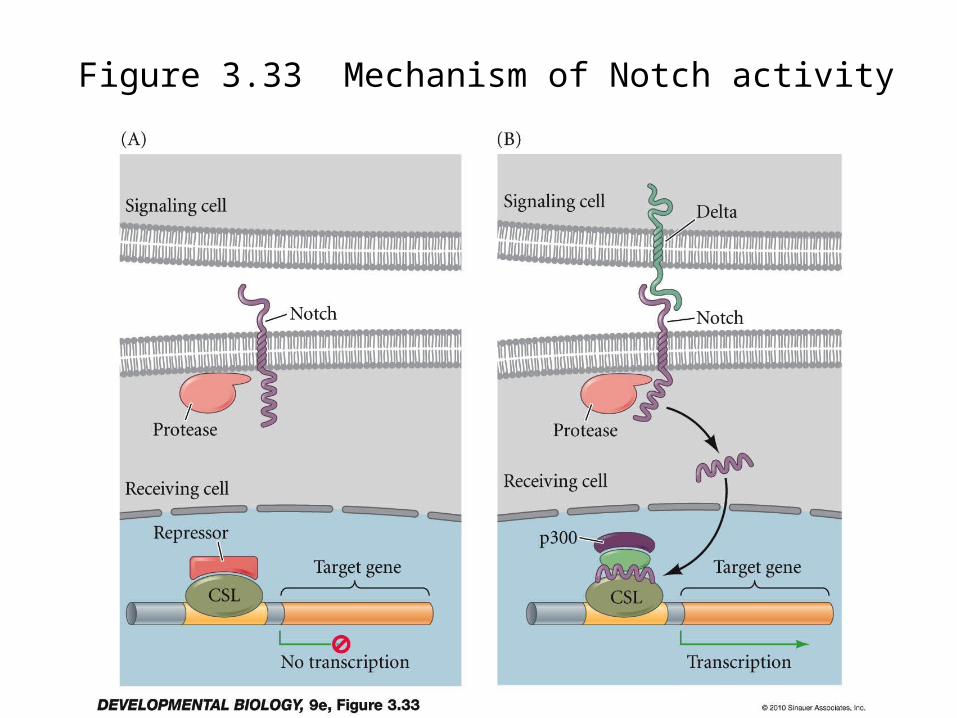
Figure 3.33 Mechanism of Notch activity

Apoptosis: genetically programmed cell death
• Absolutely essential to control cell numbers, cell quality and to create space
– The space between our fingers– 2/3 of all neurons we make– The middle ear– The cerebral ventricles– Frog tails– Male mammary epithelium

Figure 3.32 Disruption of normal brain development by blocking apoptosis

Apoptosis
• Often cells are set to apoptose by default
• They require a signal to keep them alive
• The signal can be soluble or can be attachment, such as cadherins, integrins
• These are guarantees that cells remain where they should be in the body

Figure 3.31 Apoptosis pathways in nematodes and mammals
Early workdone mostlyin worms
Mammalianhomologs
A signal thatturns on CED-9(Bcl2) saves thecell from death

A. Maintaining the differentiated state
B. The extracellular matrix as a source of developmental signals
C. Epithelial-mesenchymal transition
Other Related Strategies of Developmental Biology

• Just changing gene expression is not enough
• Maintaining the new expression pattern is essential for differentiation
• So far, four ways to do this have been described
Maintaining the Differentiated State

Four ways of maintaining differentiation after the initial signal has been given (Part 1)
TF Positive feedback loop Trithorax opens promoter

Four ways of maintaining differentiation after the initial signal has been given (Part 2)
Autocrine loop Paracrine loop

The Role of the Extracellular Matrix (ECM)
• As development proceeds, all cells secrete sugars and proteins to create solid substrate between the cells
• Nearly all cells require adhesion to survive
• Cell migration is also dependent on ECM

Figure 3.37 Extracellular matrices in the developing embryo
A fibronectin tract allows mesoderm migration during gastrulation
The epithelial cells secretefibronectin into basal laminaand then can use it migrate upon.
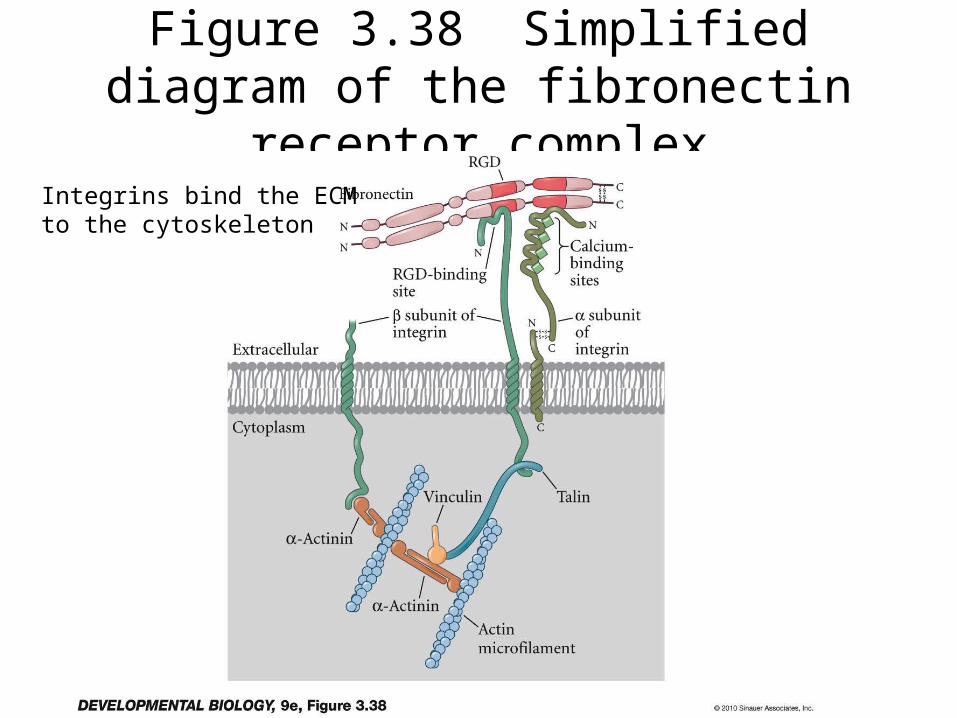
Figure 3.38 Simplified diagram of the fibronectin receptor complex
Integrins bind the ECM to the cytoskeleton

Figure 3.40 Basement membrane-directed gene expression in mammary gland tissue
Plated on plastic(A)
Plated onbasal lamina(B,C,D)

Epithelial to Mesenchymal Transition
• A key type of differentiation in many tissue forming activities in embryos and adults– formation of mesoderm from epiblast– formation of neural crest cells from neural tube– formation of coronary arteries from epicardium– formation of vertebrae from somites– wound healing in skin and vasculature– metastasis of epithelial cancers

Figure 3.41 Epithelial-mesenchymal transition, or EMT (Part 1)
If not accompanied by differentiation,loss of connectionswould lead to death

Figure 3.41 Epithelial-mesenchymal transition, or EMT (Part 2)