biochemia stresu oksydacyjnego -...
TRANSCRIPT

Biochemia
stresu oksydacyjnego
Wykład 2
Powstawanie reaktywnych form tlenu w komórkach

Źródła wolnych rodników w komórce
G. Bartosz "Druga twarz tlenu"
- Enzymy generujące H2O2:
np.
* oksydaza aldehydowa
* oksydaza D-aminokwasowa
* okydaza -hydroksykwasowa
* oksydaza ksantynowa
* oksydaza acetylokoenzymu A
* oksydaza glutarylokoenzymu A
* oksydaza galaktozowa
* oksydaza glikolanowa
- Enzymy generujące O2
np.
* oksydaza ksantynowa
* oksydaza aldehydowa
* oksydaza diaminowa
* reduktaza cytochromu P450
* reduktaza glutationowa
* oksydaza galaktozowa
* mieloperoksydaza
* oksydoreduktaza NADPH
* hydroperoksydaza prostaglandynowa
* tyrozynaza
* syntaza tlenku azotu
* reduktaza cytochromu b5
* lipooksygenaza
* dioksygenaza tryptofanowa
.

Źródła wolnych rodników w komórce
G. Bartosz "Druga twarz tlenu"
- Cykle redoks i utleniania ksenobiotyków
- Utlenianie białek oddechowych (hemoglobiny, mioglobiny)
- Samoutlenianie związków niskocząsteczkowych (np. związków tiolowych)
- Łańcuch oddechowy w mitochondriach

Mitochondrium
Stove and Camara. Antioxid Redox Signal 2009.
- Zewnętrzna błona mitochondrialna:
* stosunek wagowy białek do fosfolipidów: ~1:1
* duża zawartość poryn,
- Cząsteczki o masie do ~600 Da mogą swobodnie dyfundować do przestrzeni
międzybłonowej. Większe muszą mieć sekwencję sygnałową na N-końcu,
pozwalającą na wiązanie do translokaz.
- Przestrzeń międzykomórkowa:
* stężenie małych cząsteczek podobne jak w cytozolu
* skład białek jest odmienny niż w cytozolu
- Wewnętrzna błona mitochondrialna:
* stosunek wagowy białek do fosfolipidów: ~3:1
* duża zawartość kardiolipiny (zmniejszenie przepuszczalności błony)
* brak poryn
* transport wszystkich substancji wymaga transporterów
* obecność białek odpowiedzialnych za fosforylację oksydacyjną, syntezę i
hydrolizę ATP, transport białek regulatorowych.
Mitochondrium
L. Sryer. Biochemia; Stove and Camara. Antioxid Redox Signal 2009.
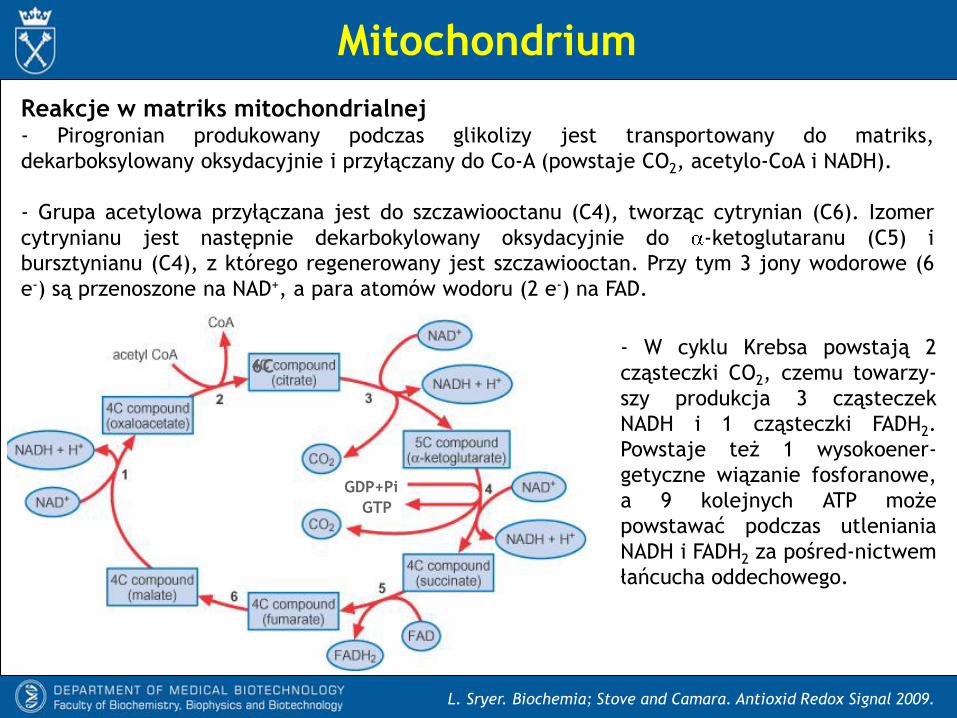
Reakcje w matriks mitochondrialnej- Pirogronian produkowany podczas glikolizy jest transportowany do matriks,
dekarboksylowany oksydacyjnie i przyłączany do Co-A (powstaje CO2, acetylo-CoA i NADH).
- Grupa acetylowa przyłączana jest do szczawiooctanu (C4), tworząc cytrynian (C6). Izomer
cytrynianu jest następnie dekarbokylowany oksydacyjnie do -ketoglutaranu (C5) i
bursztynianu (C4), z którego regenerowany jest szczawiooctan. Przy tym 3 jony wodorowe (6
e-) są przenoszone na NAD+, a para atomów wodoru (2 e-) na FAD.
- W cyklu Krebsa powstają 2
cząsteczki CO2, czemu towarzy-
szy produkcja 3 cząsteczek
NADH i 1 cząsteczki FADH2.
Powstaje też 1 wysokoener-
getyczne wiązanie fosforanowe,
a 9 kolejnych ATP może
powstawać podczas utleniania
NADH i FADH2 za pośred-nictwem
łańcucha oddechowego.
- Elektrony z NADH mogą być
transportowane z cytoplazmy
przez czółenko jabłczanowo-
asparaginowe lub czółenko
glicerolo-3-fosforanowe
6C
GDP+Pi
GTP
Łańcuch oddechowy
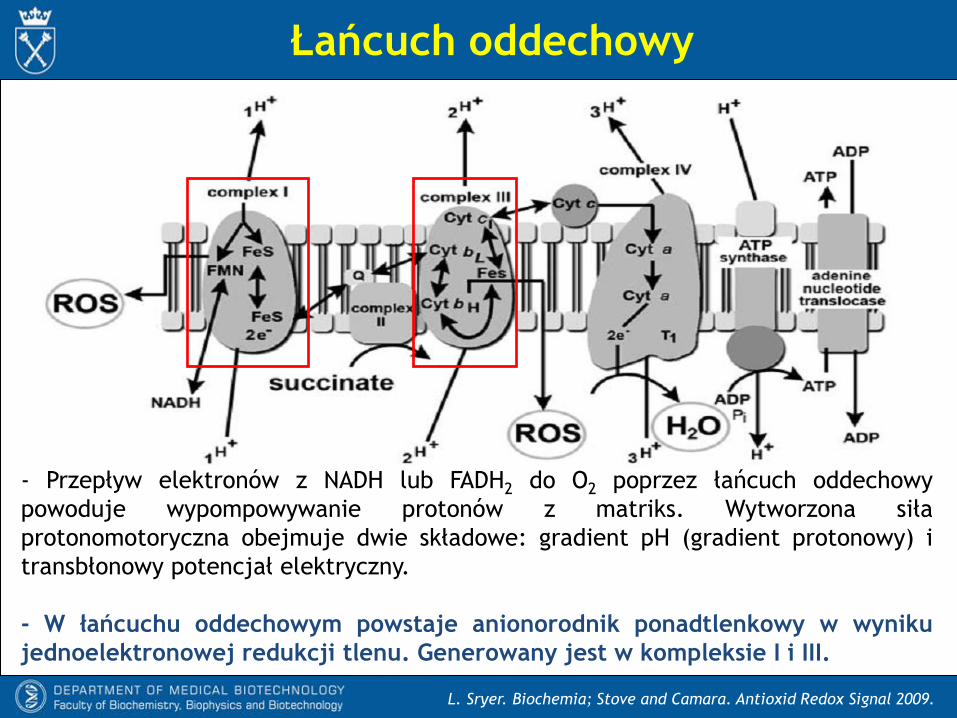
- Przepływ elektronów z NADH lub FADH2 do O2 poprzez łańcuch oddechowy
powoduje wypompowywanie protonów z matriks. Wytworzona siła
protonomotoryczna obejmuje dwie składowe: gradient pH (gradient protonowy) i
transbłonowy potencjał elektryczny.
- W łańcuchu oddechowym powstaje anionorodnik ponadtlenkowy w wyniku
jednoelektronowej redukcji tlenu. Generowany jest w kompleksie I i III.
L. Sryer. Biochemia; Stove and Camara. Antioxid Redox Signal 2009.
Łańcuch oddechowy
Stove and Camara. Antioxid Redox Signal 2009.
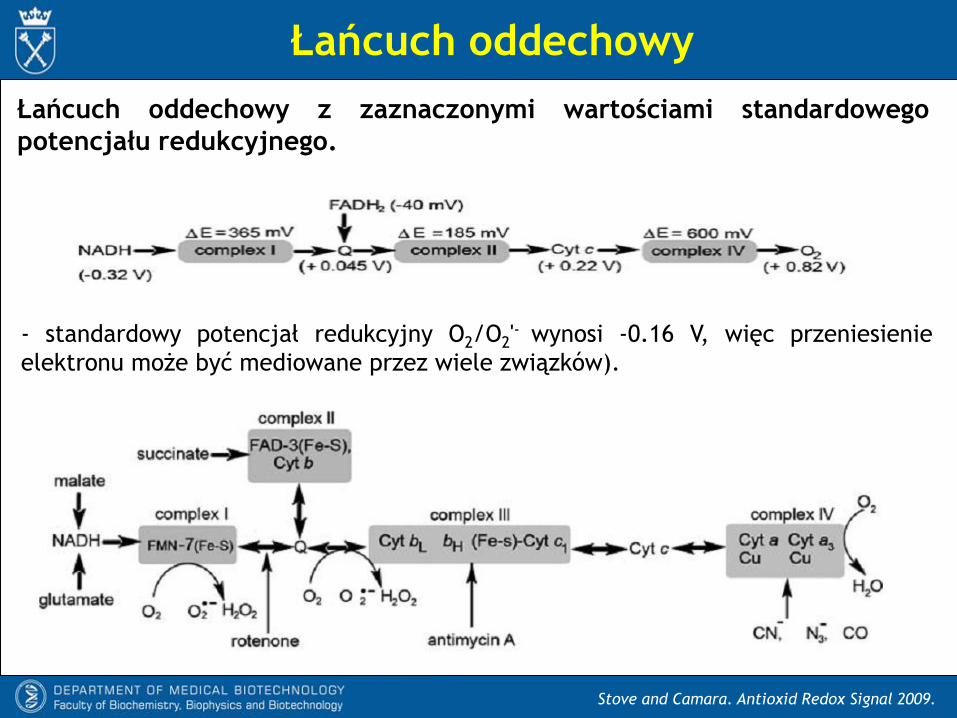
Łańcuch oddechowy z zaznaczonymi wartościami standardowego
potencjału redukcyjnego.
- standardowy potencjał redukcyjny O2/O2'- wynosi -0.16 V, więc przeniesienie
elektronu może być mediowane przez wiele związków).

Związki ułatwiające identyfikację miejsc tworzenia
reaktywnych form tlenu
Stove and Camara. Antioxid Redox Signal 2009

Produkcja O2'- w kompleksie I łańcucha oddechowego
Stove and Camara. Antioxid Redox Signal 2009
Kompleks I (oksydoreduktaza NADH:ubichinon)
- Jest transbłonowym kompleksem enzymatycznym, który:
* utlenia NADH, przekazując elektrony na ubichinon
* jest połączony z pompą protonową, a jego aktywność przyczynia się do powstania
gradientu protonów
* stanowi jedno z dwóch głównych miejsc pobierania równoważników redukcyjnych
(drugie miejsce to kompleks II)
* jest głównym źródłem ROS w komórce w warunkach fizjologicznych
Centra Fe-S
semichinon/
rodnik semichinonowy
Flawina

Stove and Camara. Antioxid Redox Signal 2009
- Kiedy mitochondria utleniają pirogronian, elektrony
są przekazywane z NADH do chinonu (Q) poprzez FMN
i centra Fe-S. Powstający QH' jest redukowany do
chinolu (QH2).
- Kiedy mitochondria utleniają jedynie bursztynian
(przy braku innych substratów) energia gradientu
protonowego wykorzystywana jest do przenoszenia
elektronu wbrew potencjałowi redoks ze
zredukowanego chinonu (chinol, QH2) na NAD+,
zamiast w stronę końcowego akceptora, czyli O2.
utlenianie
pirogronianu
utlenianie samego
bursztynianu
NADO2
Odwrotny transport elektronów
Produkcja O2'- w kompleksie I łańcucha oddechowego
Produkcja O2'- w kompleksie I łańcucha oddechowego
Stove and Camara. Antioxid Redox Signal 2009
-
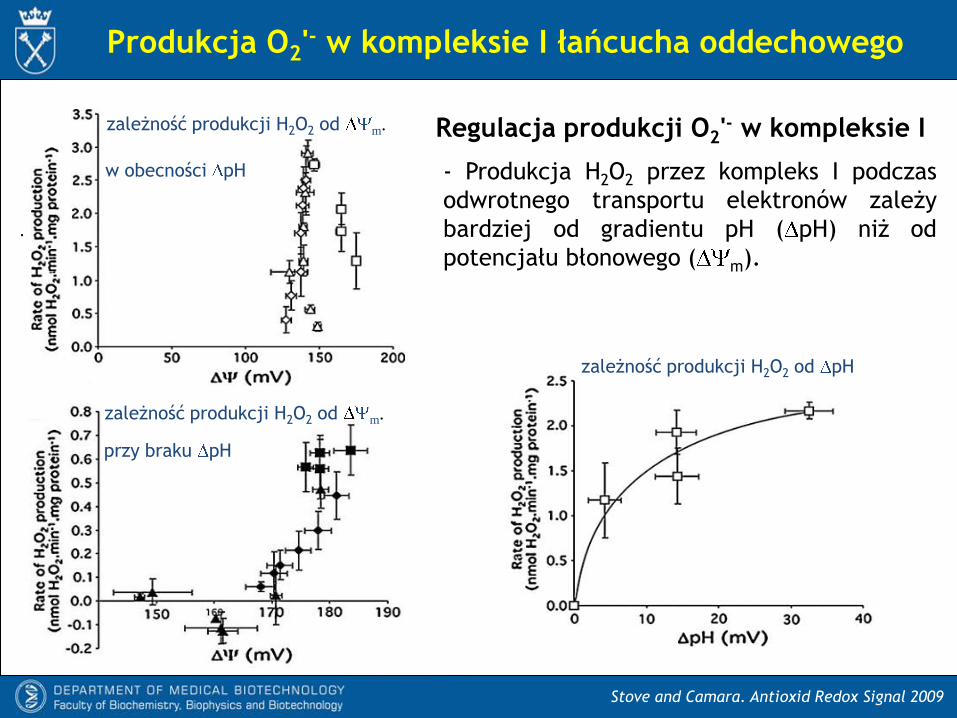
Regulacja produkcji O2'- w kompleksie I
- Produkcja H2O2 przez kompleks I podczas
odwrotnego transportu elektronów zależy
bardziej od gradientu pH ( pH) niż od
potencjału błonowego ( m).
w obecności pH
przy braku pH
zależność produkcji H2O2 od pH
zależność produkcji H2O2 od m.
zależność produkcji H2O2 od m.

Produkcja O2'- w kompleksie III łańcucha oddechowego
Stove and Camara. Antioxid Redox Signal 2009
Kompleks III (oksydoreduktaza
koenzym Q:cytochrom c)
- Budowa kompleksu III:
* zewnętrzną pulę chinonu (Qo)
* wewnętrzną pulę chinonu (Qi)
* cytochrom b566 (cyt b566)
* cytochrom b562 (cyt 562)
* białko Rieske (z kompleksami Fe-S)
* cytochrom c1
* cytochrom c
- Działanie kompleksu III:
* Ubichinon jest redukowany do QH2 po
stronie wewnętrznej (Qi) i migruje do strony
zewnętrznej (Qo) uwalniając 2H+ i przenosząc 1
e- na cyt c1 za pośrednictwem białka Rieske.
Powstaje przy tym QH' i Q.
* Drugi e- redukuje cytochrom b, dzięki czemu
elektrony są przenoszone na wewnętrzną stronę
błony, gdzie redukują chinon do QH2.
* cyt c i cyt c1 przyjmują tylko pojedynczy e-,
dlatego pełna redukcja Q wymaga utlenienia
dwóch cząsteczek QH2 w dwóch kolejnych
cyklach.

Produkcja O2'- w kompleksie III łańcucha oddechowego
Stove and Camara. Antioxid Redox Signal 2009
- Inhibitory kompleksu III:
* Myxothiazol: blokuje miejsce Qo
uniemożliwiając przeniesienie
elektronu z QH2 do centrów Fe-S i
cytochromu b.
* Stigmatellin: blokuje przeniesienie
pierwszego elektronu na centrum Fe-S.
* Antimycin A: wiąże się do miejsca
Qi i blokuje przeniesienie drugiego
elektronu do miejsca Qi. Dzięki temu
hamuje powstawanie QH2 i nasila
tworzenie O2'-.
- Wydaje się, że O2'- tworzony na
kompleksie III jest uwalniany do
przestrzeni międzybłonowej (czyli jest
dysmutowany głównie przez CuZnSOD).
To wciąż jednak nie jest jasne.

Przemiany anionorodnika ponadtlenkowego
Stove and Camara. Antioxid Redox Signal 2009.
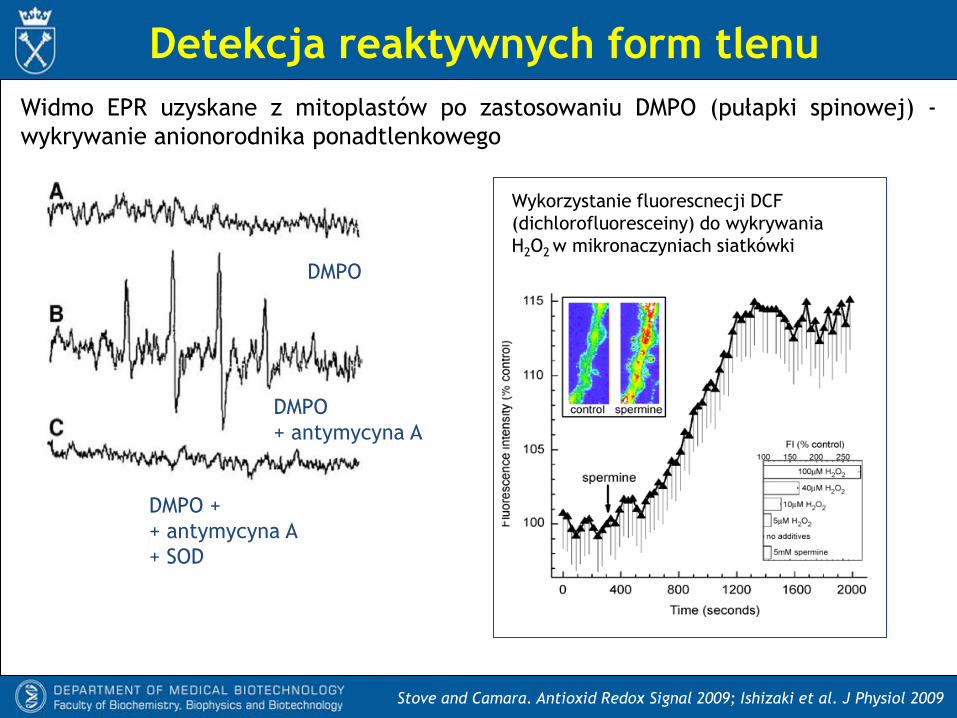
Detekcja reaktywnych form tlenu
Stove and Camara. Antioxid Redox Signal 2009; Ishizaki et al. J Physiol 2009
Widmo EPR uzyskane z mitoplastów po zastosowaniu DMPO (pułapki spinowej) -
wykrywanie anionorodnika ponadtlenkowego
DMPO
DMPO
+ antymycyna A
Wykorzystanie fluorescnecji DCF
(dichlorofluoresceiny) do wykrywania
H2O2 w mikronaczyniach siatkówki
DMPO +
+ antymycyna A
+ SOD

Detekcja anionorodnika ponadtlenkowego
Stove and Camara. Antioxid Redox Signal 2009
- Wykorzystanie Amplex Red do
wykrywania H2O2 w komórkach
hodowanych in vitro.
kontrola stymulacja
- Wykorzystanie Amplex Red do wykrywania H2O2 w
mitochondriach izolowanych z kardiomiocytów świnki
morskiej. Aktywność łańcucha oddechowego stymulowana
bursztynianem.
- CCCP (carbonyl cyanide m-chlorophenyl hydrazone): czynnik rozprzęgający
- rotenon: inhibitor kompleksu I
- AA (antymycyna A): inhibitor kompleksu III
- bursztynian: substrat oddechowy
- pirogronian: substrat oddechowy
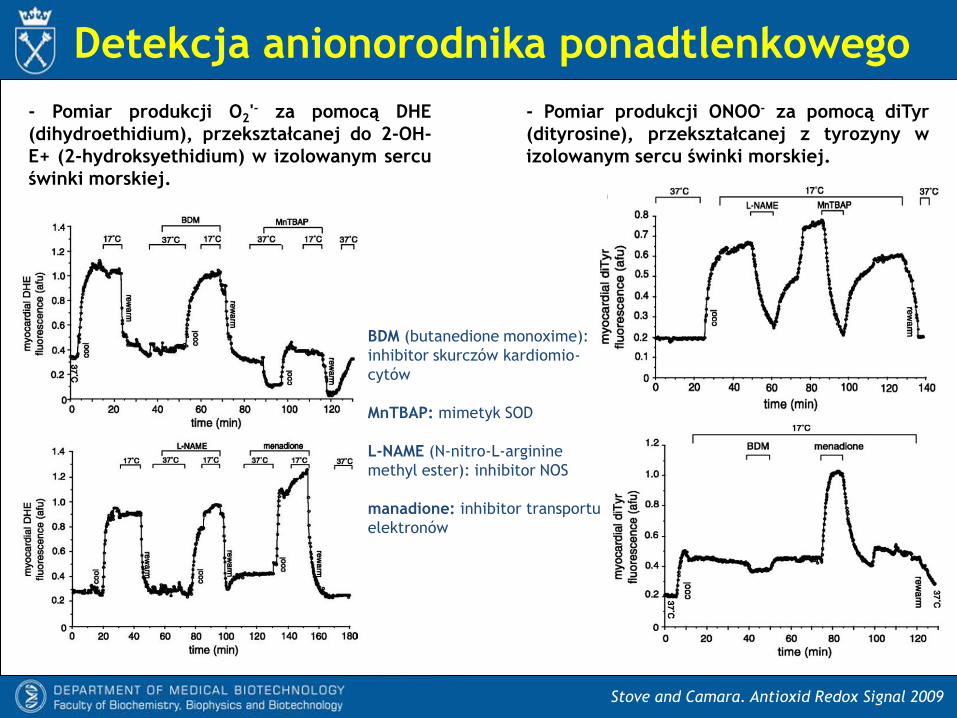
Detekcja anionorodnika ponadtlenkowego
Stove and Camara. Antioxid Redox Signal 2009
- Pomiar produkcji O2'- za pomocą DHE
(dihydroethidium), przekształcanej do 2-OH-
E+ (2-hydroksyethidium) w izolowanym sercu
świnki morskiej.
- Pomiar produkcji ONOO- za pomocą diTyr
(dityrosine), przekształcanej z tyrozyny w
izolowanym sercu świnki morskiej.
BDM (butanedione monoxime):
inhibitor skurczów kardiomio-
cytów
MnTBAP: mimetyk SOD
L-NAME (N-nitro-L-arginine
methyl ester): inhibitor NOS
manadione: inhibitor transportu
elektronów
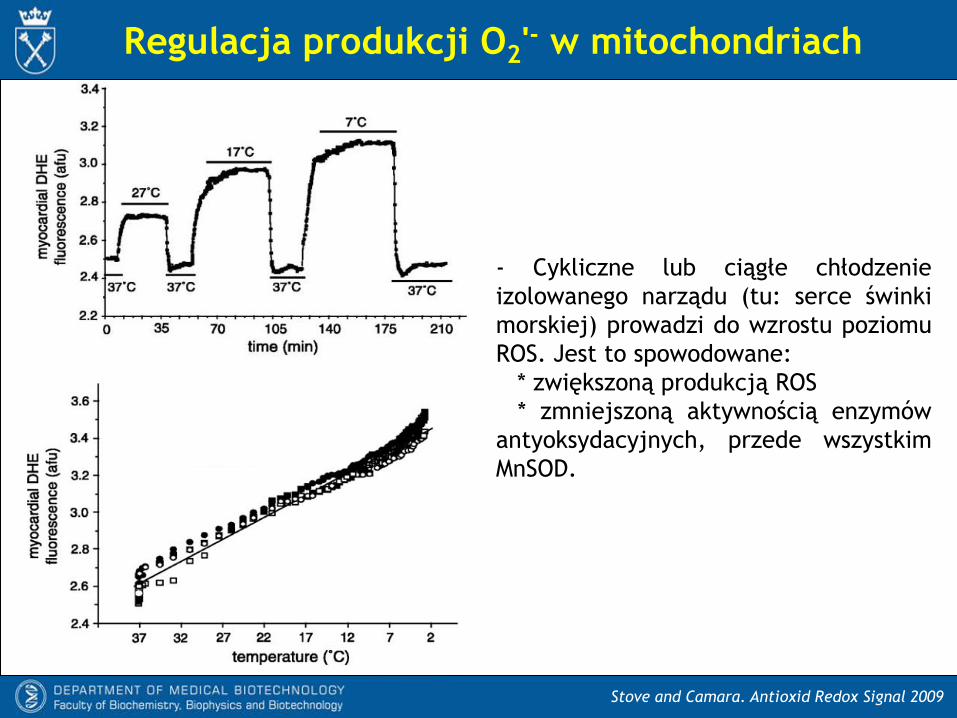
Regulacja produkcji O2'- w mitochondriach
Stove and Camara. Antioxid Redox Signal 2009
- Cykliczne lub ciągłe chłodzenie
izolowanego narządu (tu: serce świnki
morskiej) prowadzi do wzrostu poziomu
ROS. Jest to spowodowane:
* zwiększoną produkcją ROS
* zmniejszoną aktywnością enzymów
antyoksydacyjnych, przede wszystkim
MnSOD.

Ischemia i reperfuzja
- Uszkodzenie tkanek po ischemii i reperfuzji jest
powodowane przez wzrost produkcji ROS
(szczególnie istotne przy transplantacji
narządów):
* Podawanie zmiataczy O2'- i H2O2 (ale nie
jedynie O2'-) zmniejsza uszkodzenia
* Nadekspresja enzymów antyoksydacyjnych
(CuZnSOD, MnSOD, HO-1) zmniejsza uszkodzenia
* Po reperfuzji dochodzi do zwiększonej
produkcji O2'- i ONOO-.
- Większość ROS w ischemii i reperfuzji jest
produkowana w łańcuchu oddechowym, zwłaszcza
przez kompleks III.
- Bardzo istotna jest rola oksydoreduktazy
ksantynowej.
- Schłodzenie narządu przed przeszczepem nasila
produkcję ROS. nabłonek jelitowy
kontrola
I/R
http://www.vivo.colostate.edu/hbooks/pathphys/digestion/stomach/salmonella.jpg
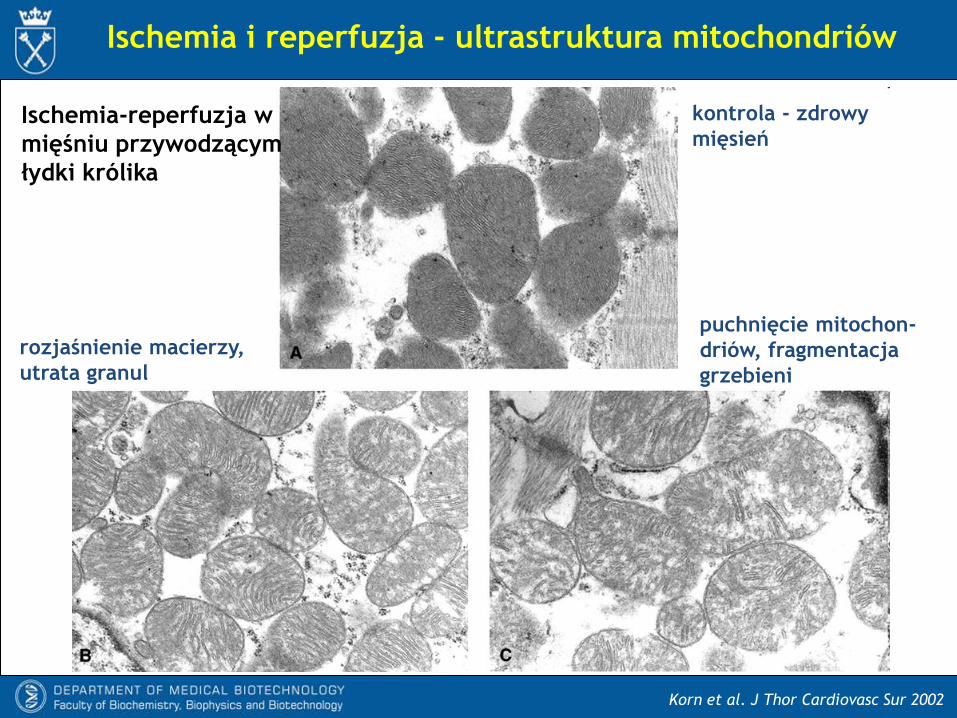
Korn et al. J Thor Cardiovasc Sur 2002
Ischemia i reperfuzja - ultrastruktura mitochondriów
kontrola - zdrowy
mięsieńIschemia-reperfuzja w
mięśniu przywodzącym
łydki królika
rozjaśnienie macierzy,
utrata granul
puchnięcie mitochon-
driów, fragmentacja
grzebieni
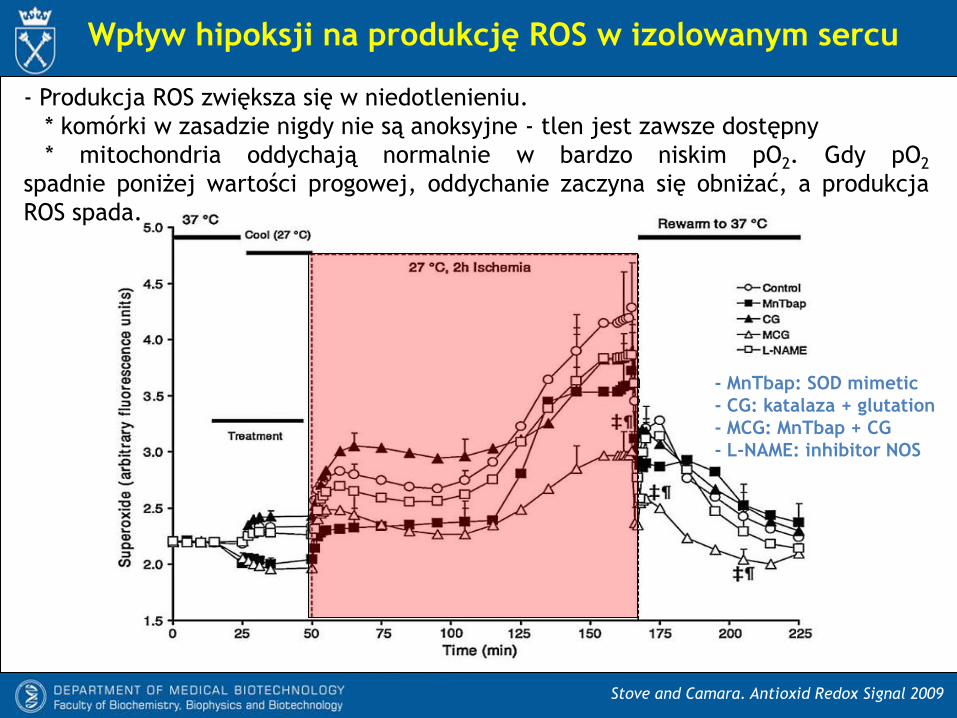
Wpływ hipoksji na produkcję ROS w izolowanym sercu
Stove and Camara. Antioxid Redox Signal 2009
- MnTbap: SOD mimetic
- CG: katalaza + glutation
- MCG: MnTbap + CG
- L-NAME: inhibitor NOS
- Produkcja ROS zwiększa się w niedotlenieniu.
* komórki w zasadzie nigdy nie są anoksyjne - tlen jest zawsze dostępny
* mitochondria oddychają normalnie w bardzo niskim pO2. Gdy pO2
spadnie poniżej wartości progowej, oddychanie zaczyna się obniżać, a produkcja
ROS spada.
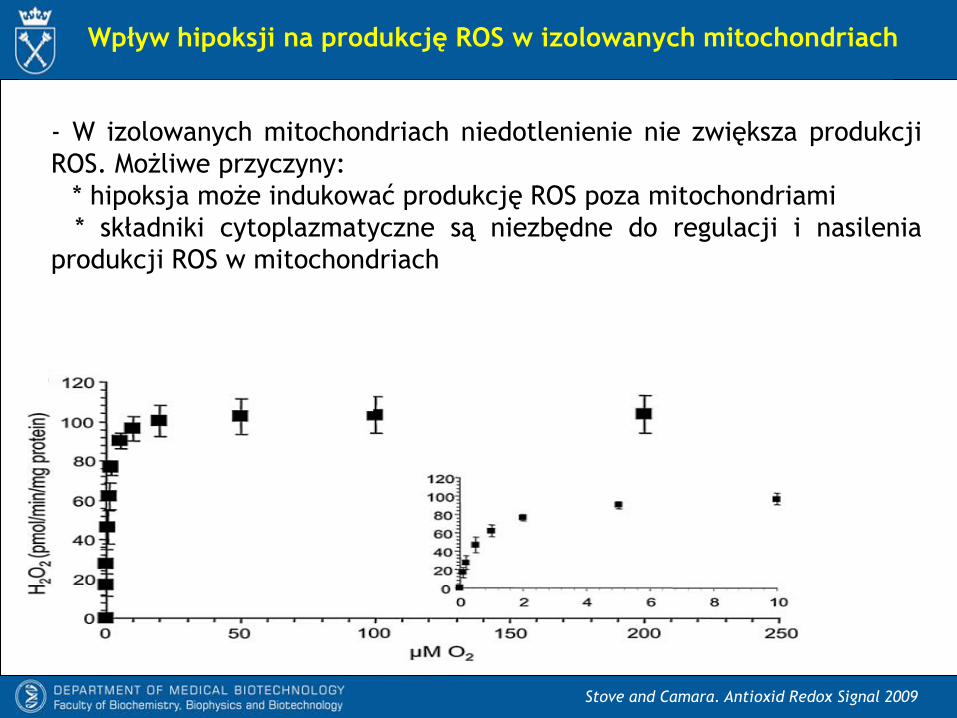
Stove and Camara. Antioxid Redox Signal 2009
Wpływ hipoksji na produkcję ROS w izolowanych mitochondriach
Faza 3
Faza 4
- W izolowanych mitochondriach niedotlenienie nie zwiększa produkcji
ROS. Możliwe przyczyny:
* hipoksja może indukować produkcję ROS poza mitochondriami
* składniki cytoplazmatyczne są niezbędne do regulacji i nasilenia
produkcji ROS w mitochondriach
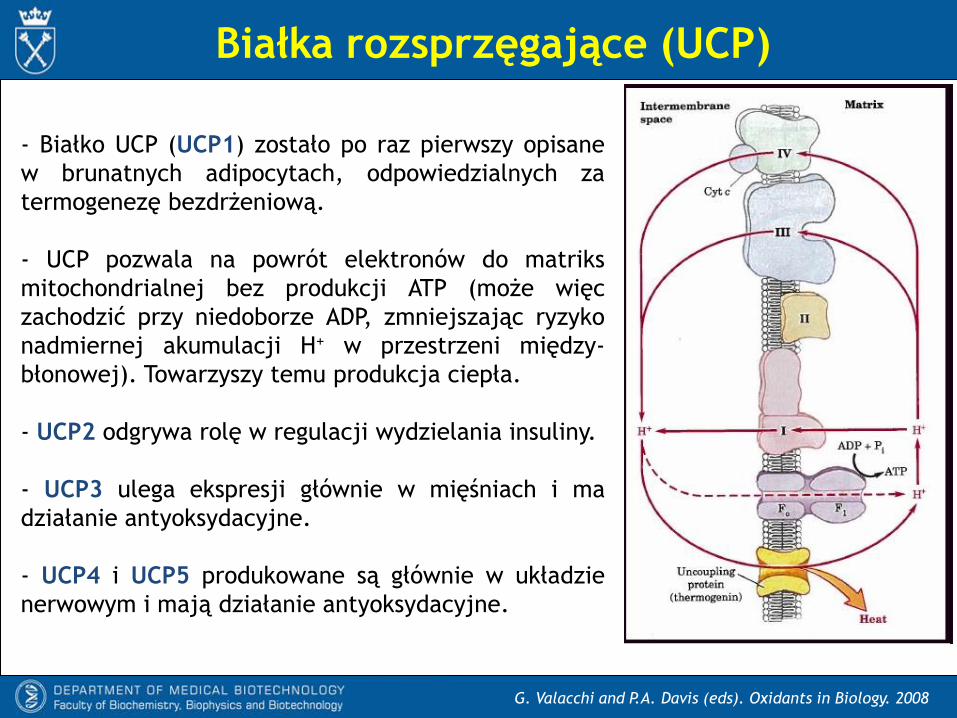
G. Valacchi and P.A. Davis (eds). Oxidants in Biology. 2008
- Białko UCP (UCP1) zostało po raz pierwszy opisane
w brunatnych adipocytach, odpowiedzialnych za
termogenezę bezdrżeniową.
- UCP pozwala na powrót elektronów do matriks
mitochondrialnej bez produkcji ATP (może więc
zachodzić przy niedoborze ADP, zmniejszając ryzyko
nadmiernej akumulacji H+ w przestrzeni między-
błonowej). Towarzyszy temu produkcja ciepła.
- UCP2 odgrywa rolę w regulacji wydzielania insuliny.
- UCP3 ulega ekspresji głównie w mięśniach i ma
działanie antyoksydacyjne.
- UCP4 i UCP5 produkowane są głównie w układzie
nerwowym i mają działanie antyoksydacyjne.
Białka rozsprzęgające (UCP)

UCP w łańcuchu oddechowym
- Myszy pozbawione genu UCP3 wykazują
nasilony stres oksydacyjny
- Nadekspresja UCP2 zwiększa oporność na
stres u Drosophila melanogaster.
Wolkow & Isner. Aging Res Rev. 2006.

Berry and Harre. J Physiol 2004
Oksydoreduktaza ksantynowa (XOR)
NH3
- Odkryta w 1902 w mleku (przez Franza Schardingera), uważana jest za enzym
ułatwiający noworodkom zwalczanie infekcji bakteryjnych dzięki produkcji ROS.
- Jest zaangażowana w hydroksylację puryn, pteryn i aldehydów, ale jej podstawową
rolą jest przekształcanie hypoksantyny do ksantyny, a do kwasu moczowego.

- Należy do hydroksylaz molibdenowych zawierających reszty flawinowe i centra Fe-S.
- Ulega ekspresji w różnych narządach, ale jej najwyższy poziom wykrywany jest w
wątrobie i jelicie.
- Jest obecna w komórkach śródbłonka i może stanowić główne źródło ROS w
śródbłonku.
- Występuje w dwóch formach:
* oksydaza ksantynowa (XO)
* dehydrogenaza ksantynowa (XDH)
- W komórkach ssaczych XOR występuje jako XDR, ale jest łatwo przekształcana do XO
w wyniku utlenienia reszt SH lub proteolizy.
- Zarówno XDH jak i XO mogą produkować ROS.
Oksydoreduktaza ksantynowa (XOR)
Berry and Harre. J Physiol 2004
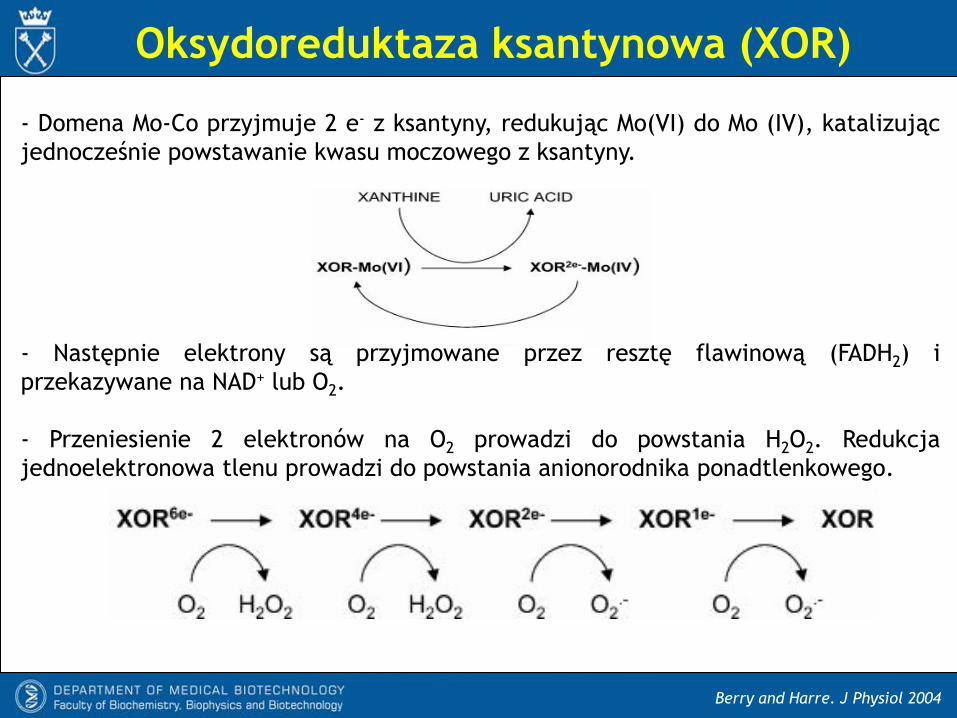
Oksydoreduktaza ksantynowa (XOR)
Berry and Harre. J Physiol 2004
- Domena Mo-Co przyjmuje 2 e- z ksantyny, redukując Mo(VI) do Mo (IV), katalizując
jednocześnie powstawanie kwasu moczowego z ksantyny.
- Następnie elektrony są przyjmowane przez resztę flawinową (FADH2) i
przekazywane na NAD+ lub O2.
- Przeniesienie 2 elektronów na O2 prowadzi do powstania H2O2. Redukcja
jednoelektronowa tlenu prowadzi do powstania anionorodnika ponadtlenkowego.
G. Valacchi and P.A. Davis (eds). Oxidants in Biology. 2008
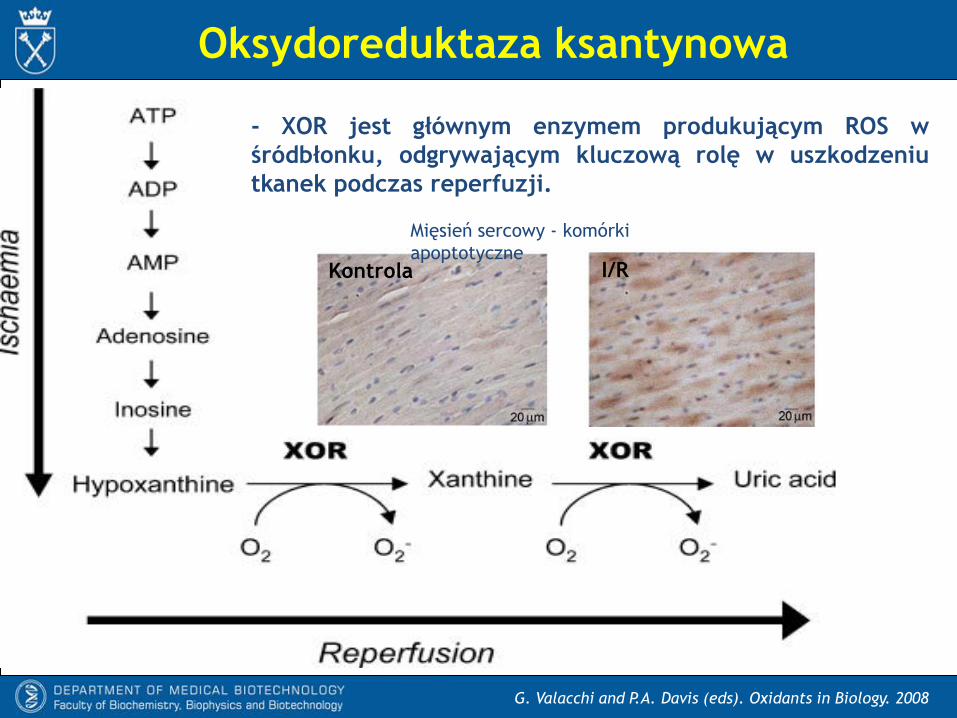
Oksydoreduktaza ksantynowa
- XOR jest głównym enzymem produkującym ROS w
śródbłonku, odgrywającym kluczową rolę w uszkodzeniu
tkanek podczas reperfuzji.
Mięsień sercowy - komórki
apoptotyczneKontrola I/R

Thomas et al. Antioxid Redox Signal 2008
- Każda izoforma oksydazy NADPH zawiera błonowa domenę katalityczną NOX,
posiadającą wszystkie elementy niezbędne do transferu elektronów z NADPH na tlen:
* domenę wiążącą NADPH
* FAD
* dwie grupy hemowe
- Nox1, Nox2 i Nox4 są związane z dodatkowymi czterema dodatkowymi
podjednostkami:
* p22phox (stabilizuje Nox w błonie)
* p47phox (NoxO1, stabilizujące białko cytozolowe)
* p67phox (NoxA1, regulatorowe białko cytozolowe)
* Rac1 (małe białko G)
Podjednostki oksydazy NADPH
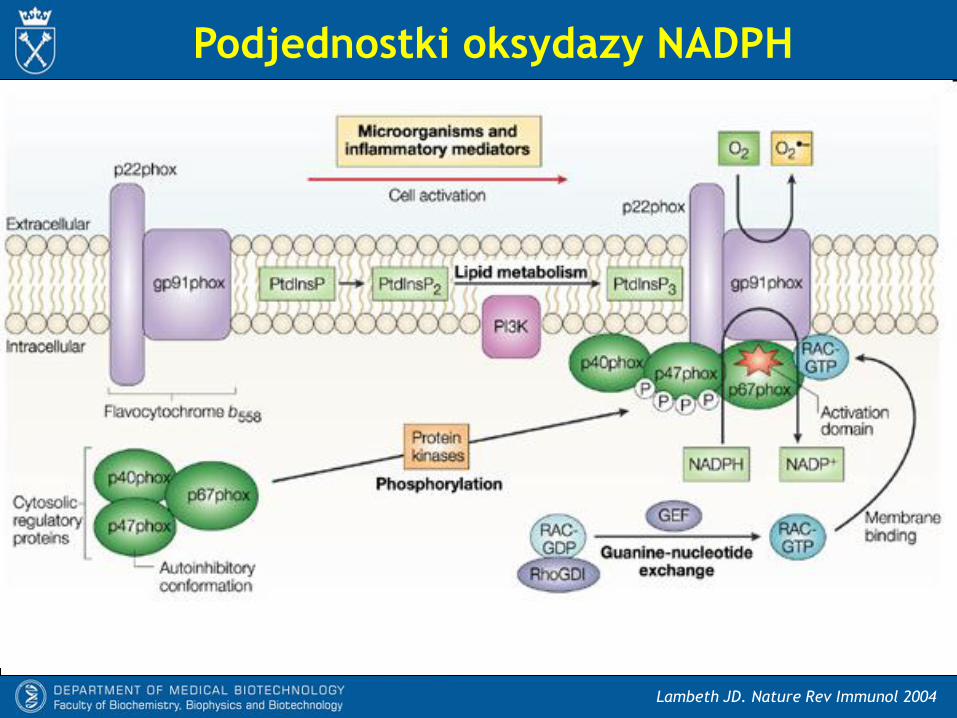
Lambeth JD. Nature Rev Immunol 2004
Podjednostki oksydazy NADPH

Thomas et al. Antioxid Redox Signal 2008
Podjednostki oksydazy NADPH- Struktura NOX1, NOX2, NOX3 i NOX4 jest podobna, a ich domena C-końcowa wiąże
FAD i NADPH.
- Domena C-końcowa NOX5 jest podobna do kalmoduliny i wiąże wapń.
- DUOX1 i DUOX2 są podobne do NOX5, a dodatkowo posiadają domenę transbłonową
przy N-końcu homologiczną do peroksydaz hemowych.
- NOX1, NOX2, NOX3, NOX4 i NOX5 produkują głównie O2-. DUOX1 i DUOX2 produkują
głównie H2O2.
Thomas et al. Antioxid Redox Signal 2008
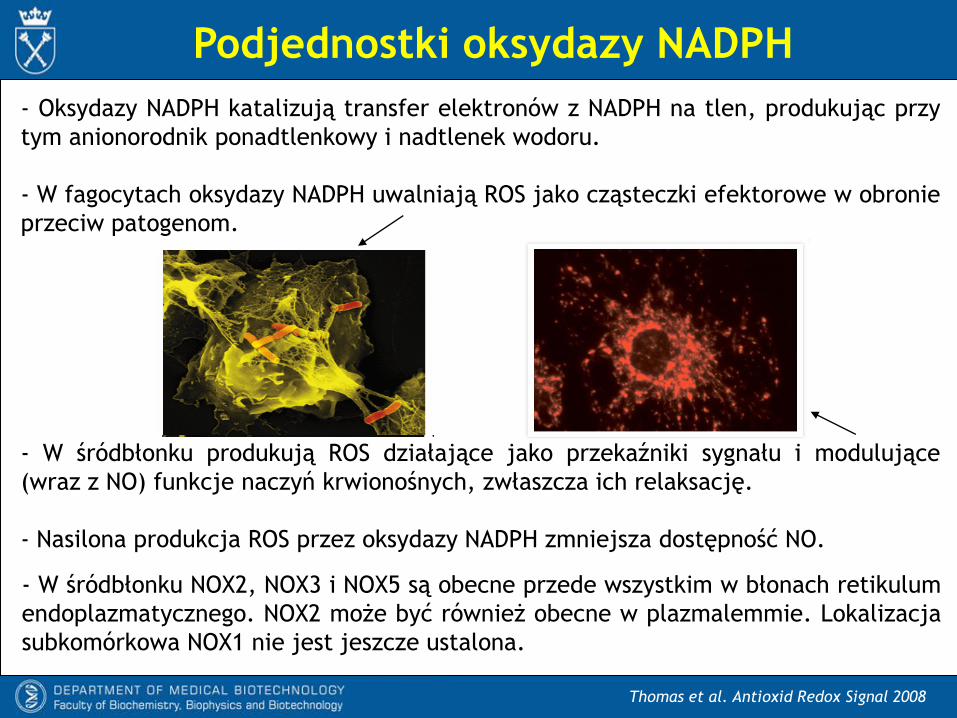
Podjednostki oksydazy NADPH
- Oksydazy NADPH katalizują transfer elektronów z NADPH na tlen, produkując przy
tym anionorodnik ponadtlenkowy i nadtlenek wodoru.
- W fagocytach oksydazy NADPH uwalniają ROS jako cząsteczki efektorowe w obronie
przeciw patogenom.
- W śródbłonku produkują ROS działające jako przekaźniki sygnału i modulujące
(wraz z NO) funkcje naczyń krwionośnych, zwłaszcza ich relaksację.
- Nasilona produkcja ROS przez oksydazy NADPH zmniejsza dostępność NO.
- W śródbłonku NOX2, NOX3 i NOX5 są obecne przede wszystkim w błonach retikulum
endoplazmatycznego. NOX2 może być również obecne w plazmalemmie. Lokalizacja
subkomórkowa NOX1 nie jest jeszcze ustalona.

Oksydazy NADPH
Newsholme et al. J Physiol 2007
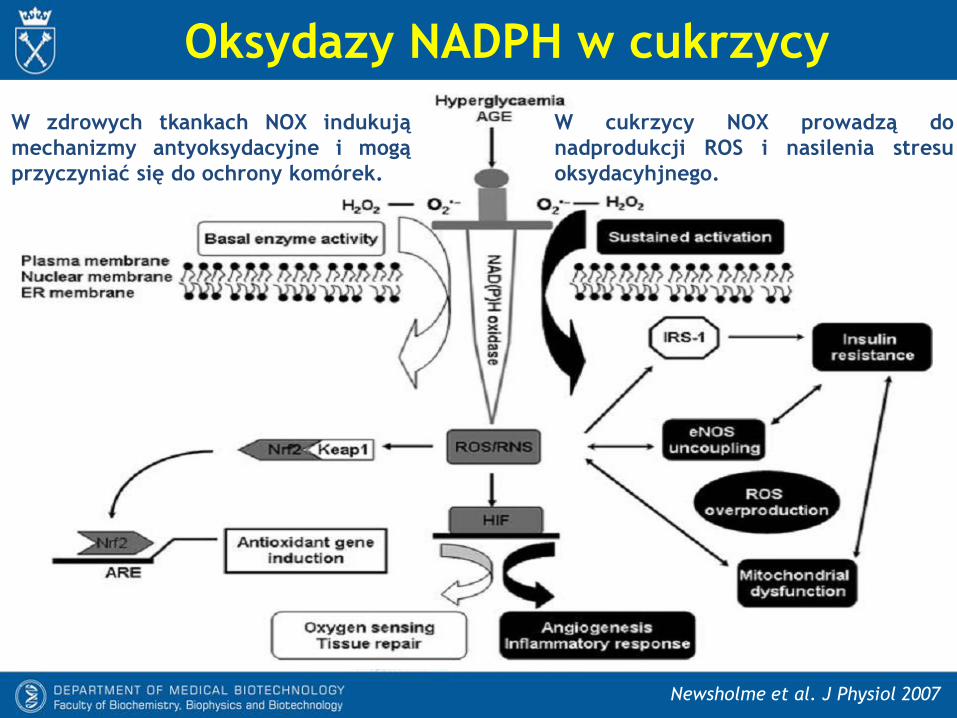
Oksydazy NADPH w cukrzycy
Newsholme et al. J Physiol 2007
W zdrowych tkankach NOX indukują
mechanizmy antyoksydacyjne i mogą
przyczyniać się do ochrony komórek.
W cukrzycy NOX prowadzą do
nadprodukcji ROS i nasilenia stresu
oksydacyhjnego.

Oksydazy NADPH w cukrzycy
Newsholme et al. J Physiol 2007
Wpływ adipokin na ekspresję/aktywność oksydaz NADPH w ścianie naczynia
Aktywacja oksydaz NADPH w różnych modelach cukrzycy
HUVEC, BAEC:
komórki
śródbłonka;
BASM: komórki
mięśni gładkich
aorty

Oksydazy NADPH w cukrzycy
Newsholme et al. J Physiol 2007
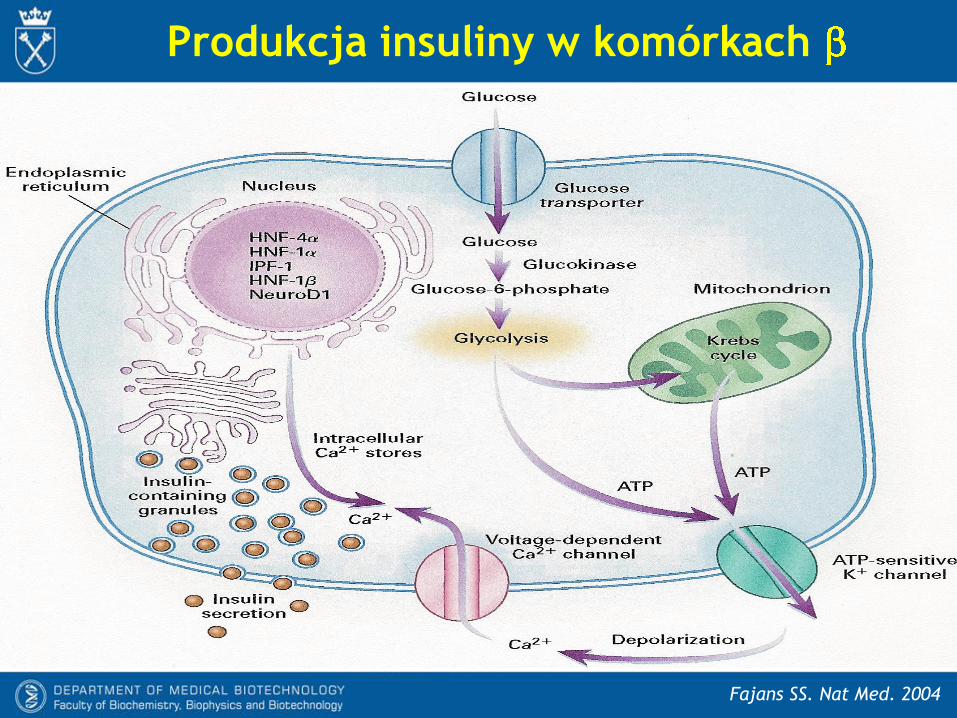
Produkcja insuliny w komórkach
Fajans SS. Nat Med. 2004
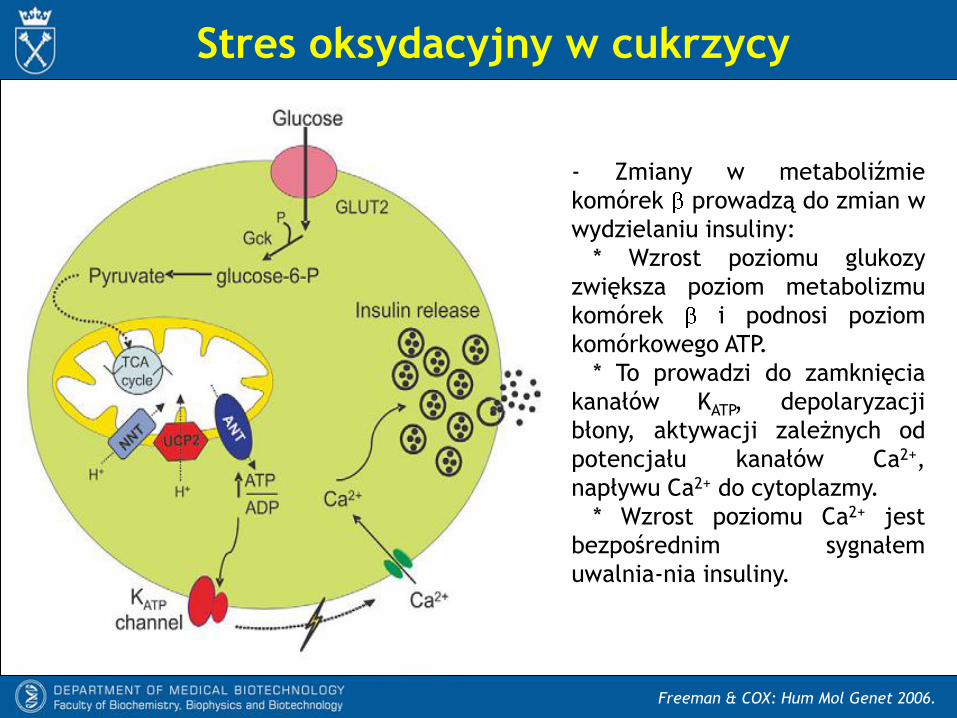
Freeman & COX: Hum Mol Genet 2006.
- Zmiany w metaboliźmie
komórek prowadzą do zmian w
wydzielaniu insuliny:
* Wzrost poziomu glukozy
zwiększa poziom metabolizmu
komórek i podnosi poziom
komórkowego ATP.
* To prowadzi do zamknięcia
kanałów KATP, depolaryzacji
błony, aktywacji zależnych od
potencjału kanałów Ca2+,
napływu Ca2+ do cytoplazmy.
* Wzrost poziomu Ca2+ jest
bezpośrednim sygnałem
uwalnia-nia insuliny.
Stres oksydacyjny w cukrzycy
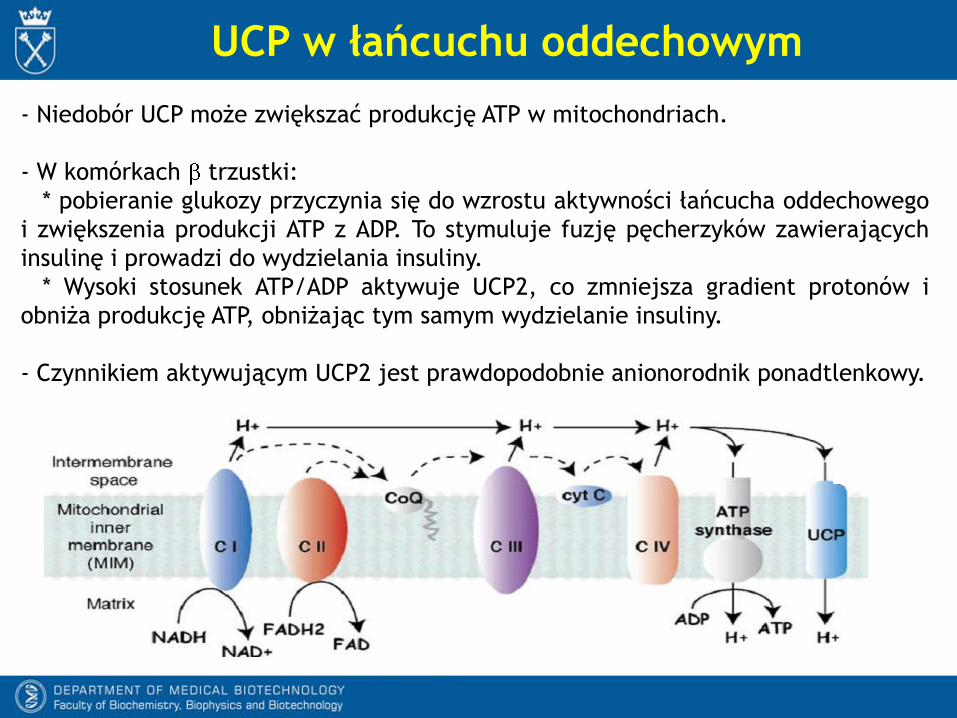
UCP w łańcuchu oddechowym
- Niedobór UCP może zwiększać produkcję ATP w mitochondriach.
- W komórkach trzustki:
* pobieranie glukozy przyczynia się do wzrostu aktywności łańcucha oddechowego
i zwiększenia produkcji ATP z ADP. To stymuluje fuzję pęcherzyków zawierających
insulinę i prowadzi do wydzielania insuliny.
* Wysoki stosunek ATP/ADP aktywuje UCP2, co zmniejsza gradient protonów i
obniża produkcję ATP, obniżając tym samym wydzielanie insuliny.
- Czynnikiem aktywującym UCP2 jest prawdopodobnie anionorodnik ponadtlenkowy.
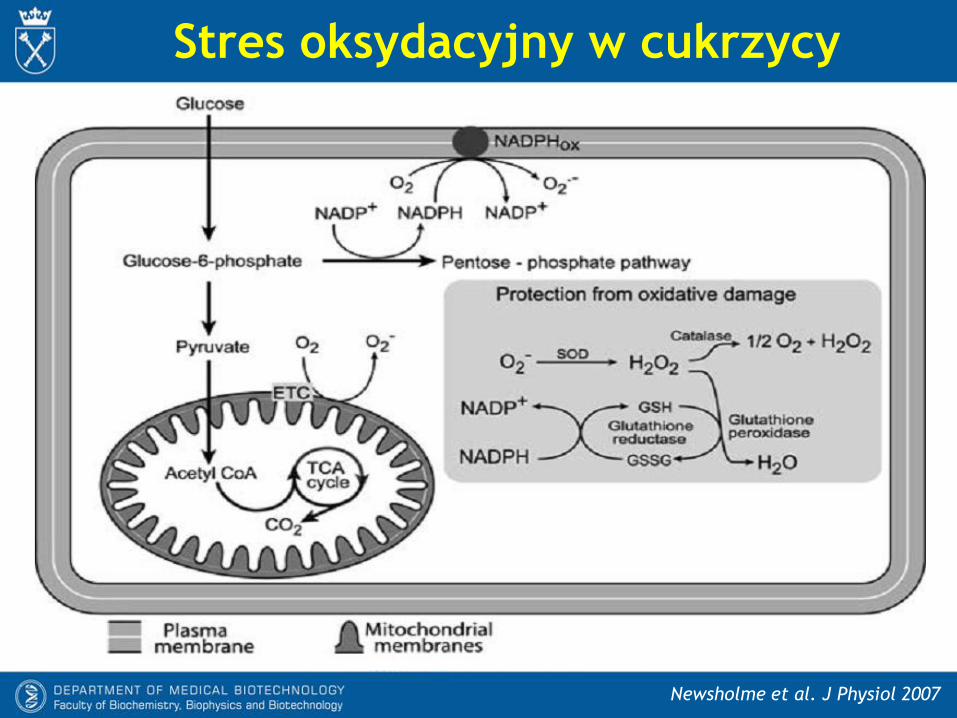
Stres oksydacyjny w cukrzycy
Newsholme et al. J Physiol 2007
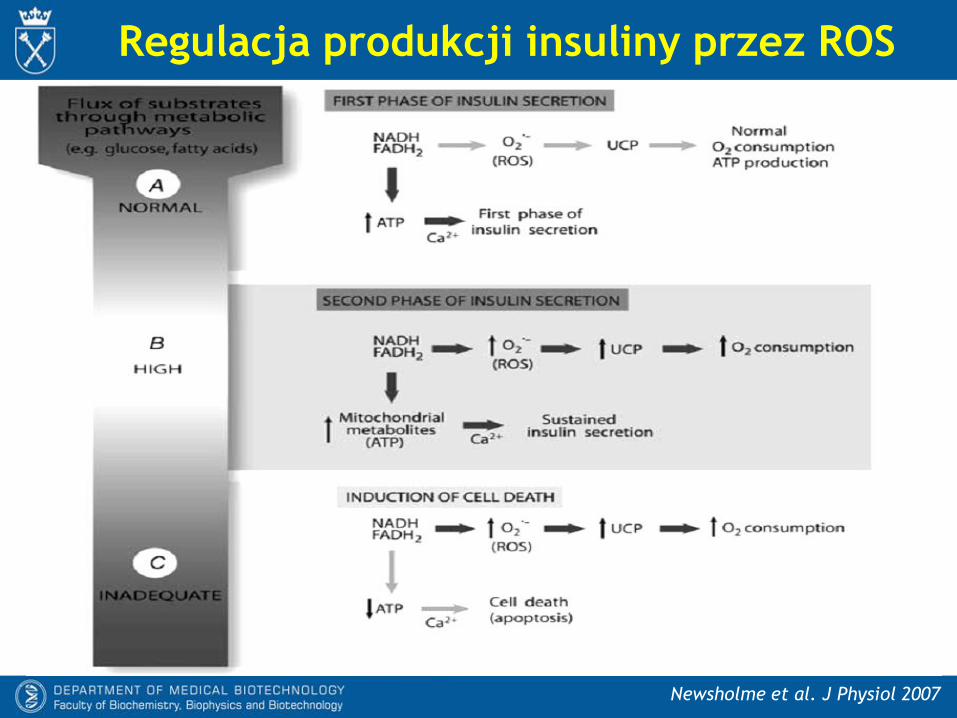
Regulacja produkcji insuliny przez ROS
Newsholme et al. J Physiol 2007

Newsholme et al. J Physiol 2007
Oksydazy NAD(P)H w cukrzycy
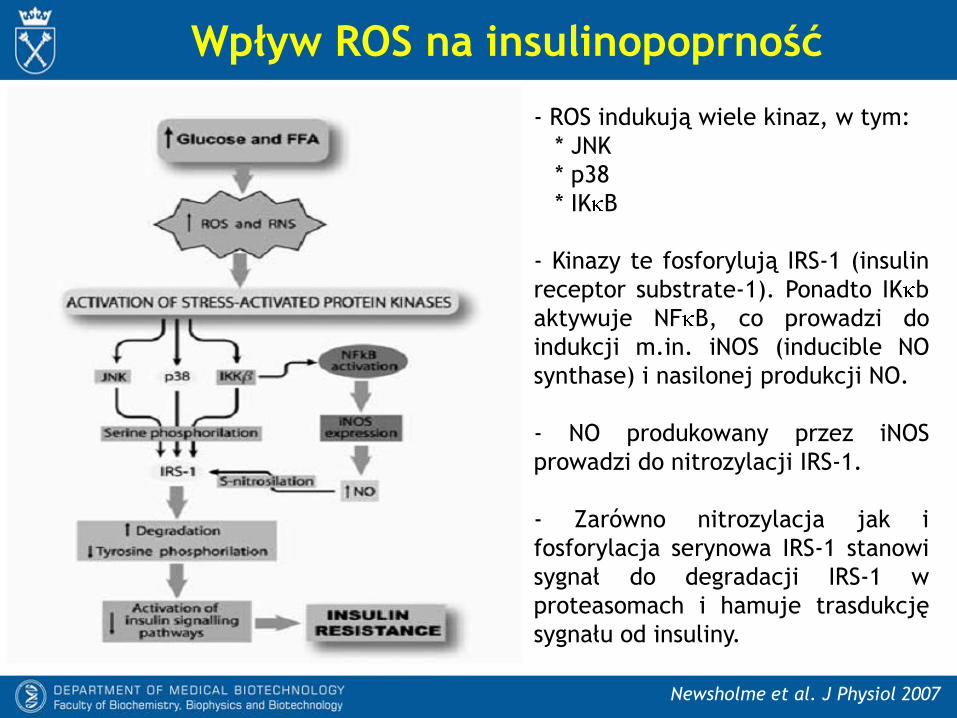
Wpływ ROS na insulinopoprność
Newsholme et al. J Physiol 2007
- ROS indukują wiele kinaz, w tym:
* JNK
* p38
* IK B
- Kinazy te fosforylują IRS-1 (insulin
receptor substrate-1). Ponadto IK b
aktywuje NF B, co prowadzi do
indukcji m.in. iNOS (inducible NO
synthase) i nasilonej produkcji NO.
- NO produkowany przez iNOS
prowadzi do nitrozylacji IRS-1.
- Zarówno nitrozylacja jak i
fosforylacja serynowa IRS-1 stanowi
sygnał do degradacji IRS-1 w
proteasomach i hamuje trasdukcję
sygnału od insuliny.

Dziękuję
Slajdy dostępne na stronie Zakładu Biotechnologii Medycznej
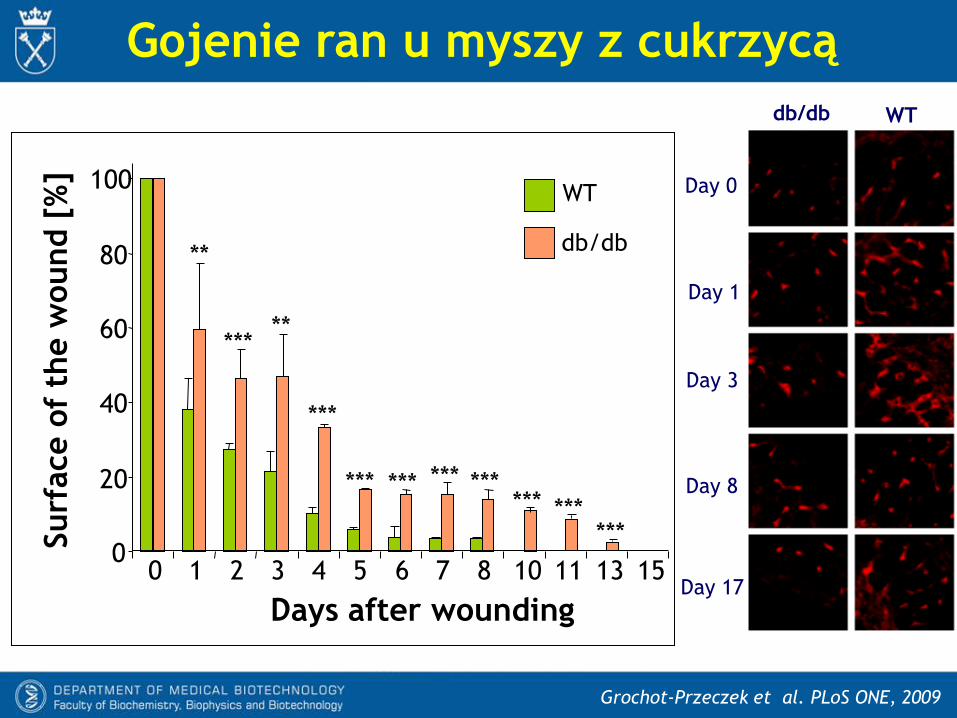
Gojenie ran u myszy z cukrzycą
Days after wounding
0
20
40
60
80
100
0 1 2 3 4 5 6 7 8 10 11 13 15
Surf
ace o
f th
e w
ound [
%]
WT
db/db**
*****
***
*** *** *** ****** ***
***
Grochot-Przeczek et al. PLoS ONE, 2009
db/db WT
Day 1
Day 3
Day 8
Day 17
Day 0
Liu & Velazquez. Antioxid Redox Signal 2008
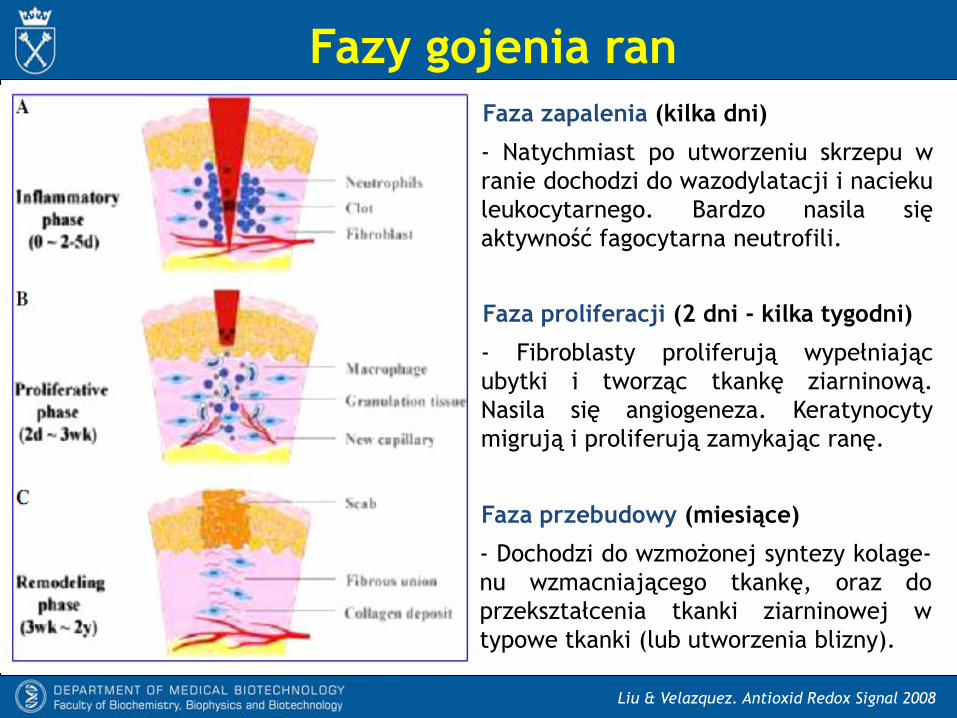
Fazy gojenia ran
Faza zapalenia (kilka dni)
- Natychmiast po utworzeniu skrzepu w
ranie dochodzi do wazodylatacji i nacieku
leukocytarnego. Bardzo nasila się
aktywność fagocytarna neutrofili.
Faza proliferacji (2 dni - kilka tygodni)
- Fibroblasty proliferują wypełniając
ubytki i tworząc tkankę ziarninową.
Nasila się angiogeneza. Keratynocyty
migrują i proliferują zamykając ranę.
Faza przebudowy (miesiące)
- Dochodzi do wzmożonej syntezy kolage-
nu wzmacniającego tkankę, oraz do
przekształcenia tkanki ziarninowej w
typowe tkanki (lub utworzenia blizny).

Jarajapu & Grant. Circ Res 2010.
Upośledzenie funkcji EPC w cukrzycy
EPC: komórki progenitorowe śródbłonka
- U zdrowych osobników czynniki uwalniane
przez ischemiczne lub zranione tkanki
mobilizują komórki progenitorowe (w tym
EPC) ze szpiku, a te po dotarciu do miejsc
zranienia lub niedotlenienia uwalniają
czynniki proangenne (w tym niewielkie ilości
NO i ROS) oraz biorą udział w
neowaskularyzacji i naprawie naczyń.
- W cukrzycy sygnały wysyłane przez
niedotlenione lub zranione tkanki są słabsze,
przez co mobilizacja komórek progenitoro-
wych jest mniejsza. Komórki progenitorowe
które docierają do zranionych tkanek
uwalniają niewiele czynników proangiogen-
nych, natomiast dużo prozapalnych i
antyangiogennych (w tym duże ilości NO i
ROS).
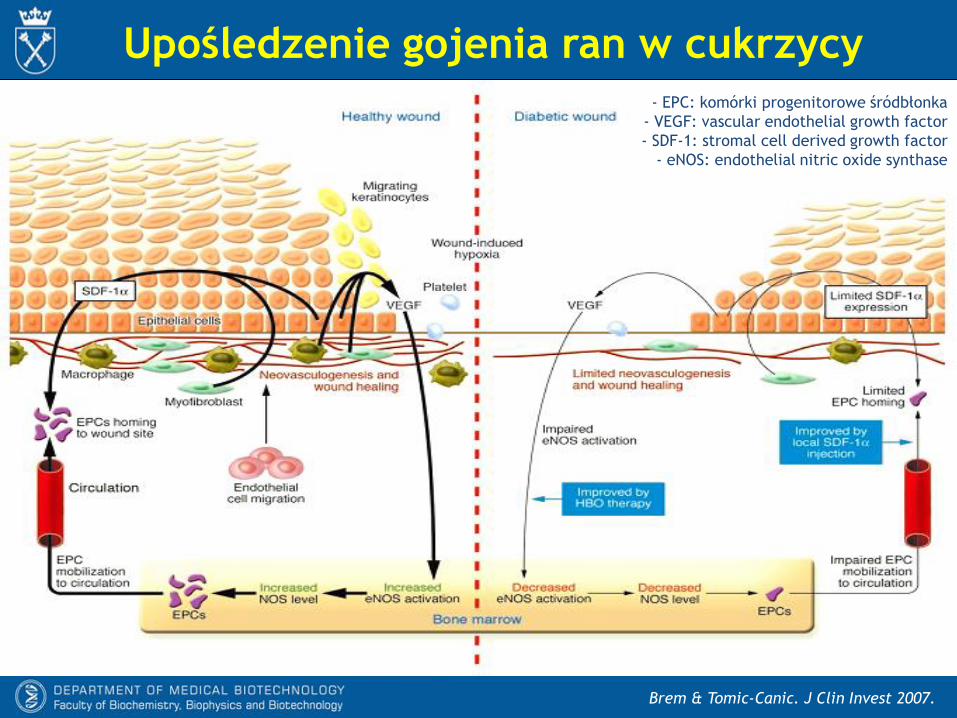
Brem & Tomic-Canic. J Clin Invest 2007.
Upośledzenie gojenia ran w cukrzycy- EPC: komórki progenitorowe śródbłonka
- VEGF: vascular endothelial growth factor
- SDF-1: stromal cell derived growth factor
- eNOS: endothelial nitric oxide synthase

Liu & Velazquez. Antioxid Redox Signal 2008
Mobilizacja EPC
- Zranione tkanki uwalniają między innymi VEGF, który wpływa na komórki
podścieliska szpiku. Prowadzi to do:
* aktywacji enzymatycznej eNOS
* nasilenia produkcji NO.
- To zwiększa aktywność MMP-9, która uwalnia sKitL z błonowego białka mKitL.
- Związanie sKitL do receptora c-Kit prowadzi do uwolnienia EPC z niszy szpikowych
do krążenia.
EPC w szpiku

Mechanisms of blood vessels formation
Formation of blood
vessels de novo
Incorporation to pre-existing
vessels
paracrine stimulation
of endothelial cells
(VEGF, bFGF, IL-8…)
Carmeliet P. Nat Med, 2000

Liu & Velazquez. Antioxid Redox Signal 2008
Regulacja mobilizacji EPC w hiperoksji i hipoksji
- W cukrzycy zmniejszony jest poziom fosforylacji eNOS przez kinazy PI3K/Akt,
przez co zmniejsza się produkcja NO.
- Nasilony stres oksydacyjny i zwiększona produkcja anionorodnika
ponadtlenkowego prowadzi do nasilonej syntezy nadtlenoazotynu (ONOO-) i
zmniejszenia dostępności NO.
- W ranach cukrzycowych osłabiona jest synteza SDF-1, co zmniejsza napływ
komórek progenitorowych do uszkodzonych tkanek. To przyczynia się do
upośledzenia angiogenezy i opóźnia gojenie.
- Cukrzyca powoduje również zmniejszenie liczby EPC w szpiku.
WT db/db
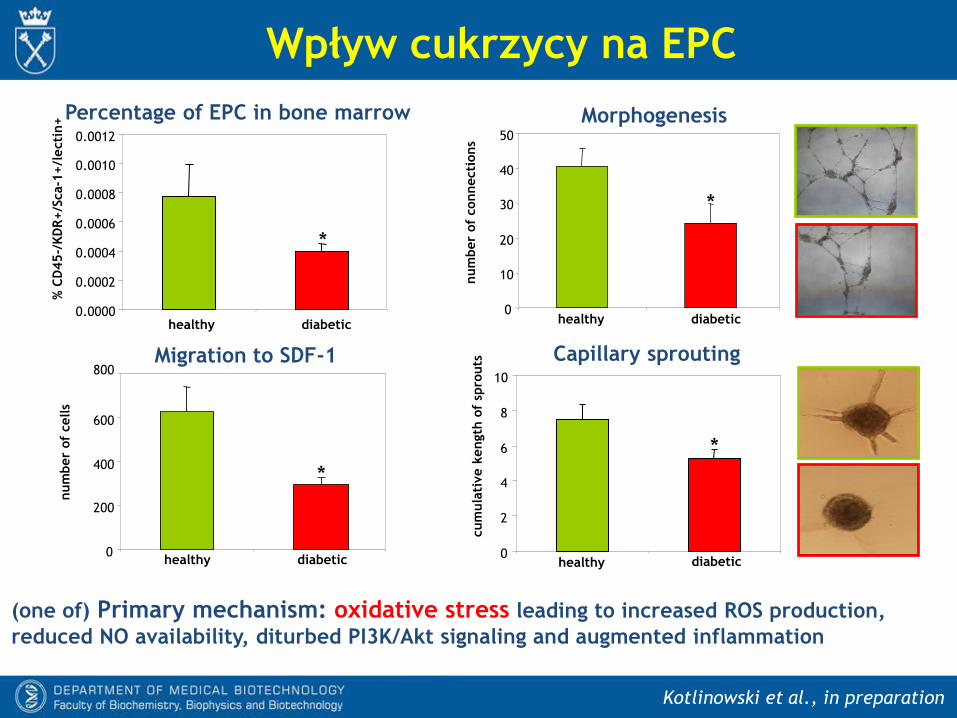
Wpływ cukrzycy na EPC
Kotlinowski et al., in preparation
Percentage of EPC in bone marrow
0.0000
0.0002
0.0004
0.0006
0.0008
0.0010
0.0012
healthy diabetic
% C
D45-/
KD
R+/S
ca-1
+/l
ecti
n+
*
0
200
400
600
800
0
10
20
30
40
50
0
2
4
6
8
10
num
ber
of
cells
*
Migration to SDF-1
healthy diabetic
num
ber
of
connecti
ons
*
healthy diabetic
Morphogenesis
cum
ula
tive k
ength
of
spro
uts
*
healthy diabetic
Capillary sprouting
(one of) Primary mechanism: oxidative stress leading to increased ROS production,
reduced NO availability, diturbed PI3K/Akt signaling and augmented inflammation
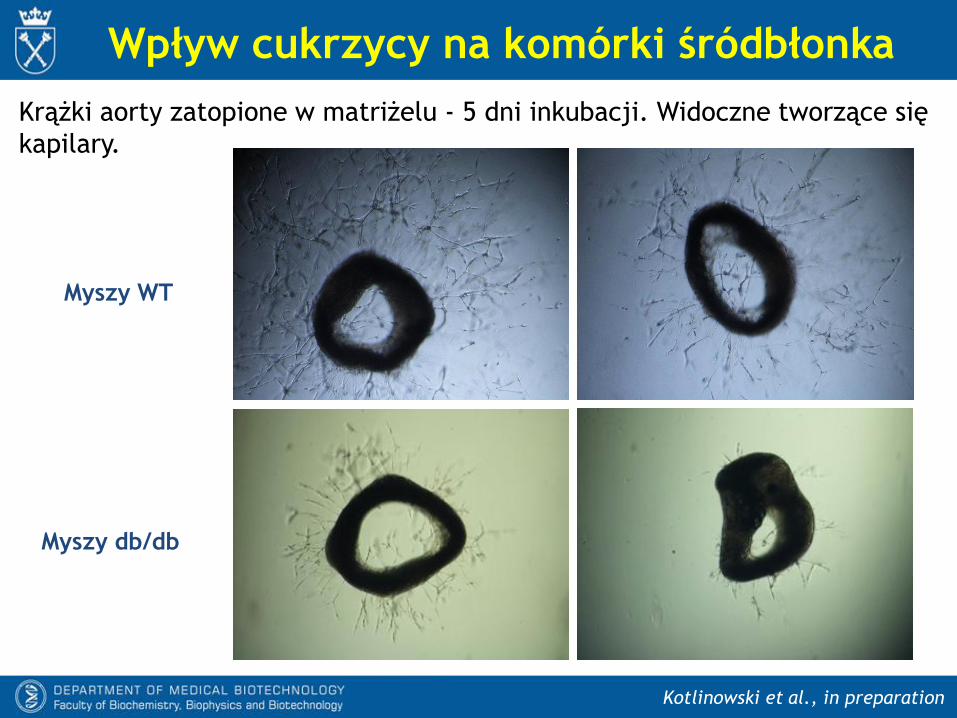
Kotlinowski et al., in preparation
Wpływ cukrzycy na komórki śródbłonka
Krążki aorty zatopione w matriżelu - 5 dni inkubacji. Widoczne tworzące się
kapilary.
Myszy WT
Myszy db/db
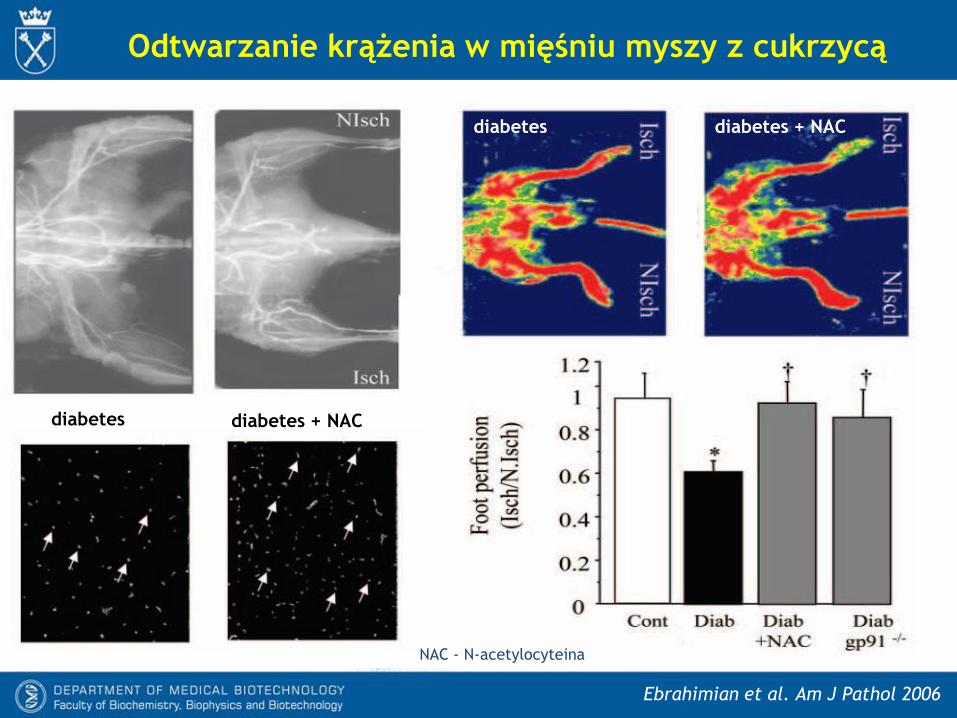
Odtwarzanie krążenia w mięśniu myszy z cukrzycą
Ebrahimian et al. Am J Pathol 2006
diabetes diabetes + NAC
diabetes + NACdiabetes
NAC - N-acetylocyteina
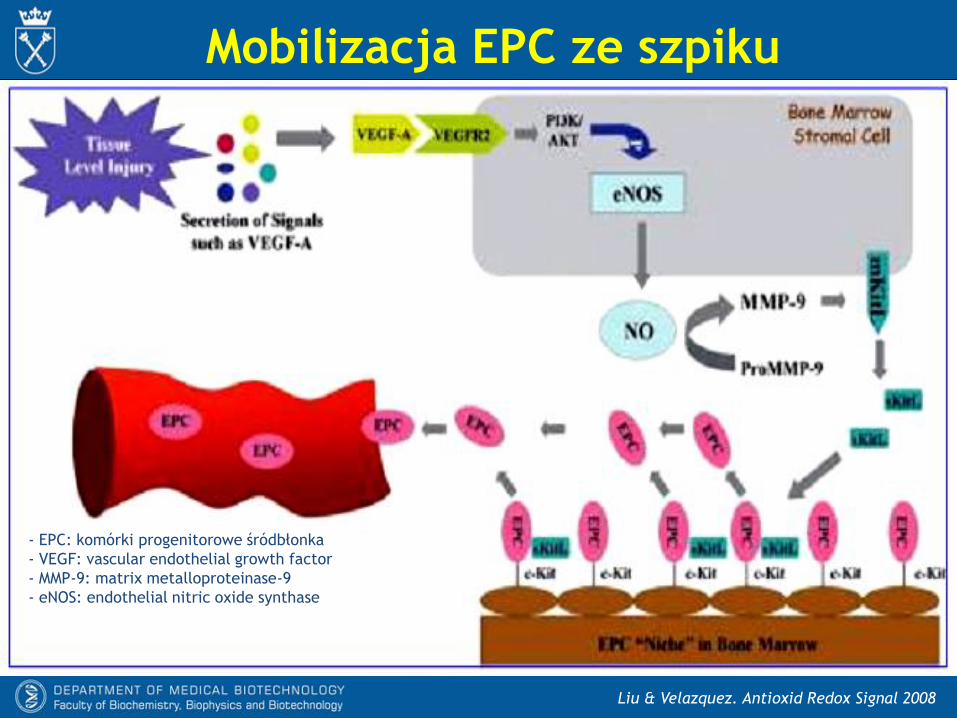
Liu & Velazquez. Antioxid Redox Signal 2008
Mobilizacja EPC ze szpiku
- EPC: komórki progenitorowe śródbłonka
- VEGF: vascular endothelial growth factor
- MMP-9: matrix metalloproteinase-9
- eNOS: endothelial nitric oxide synthase
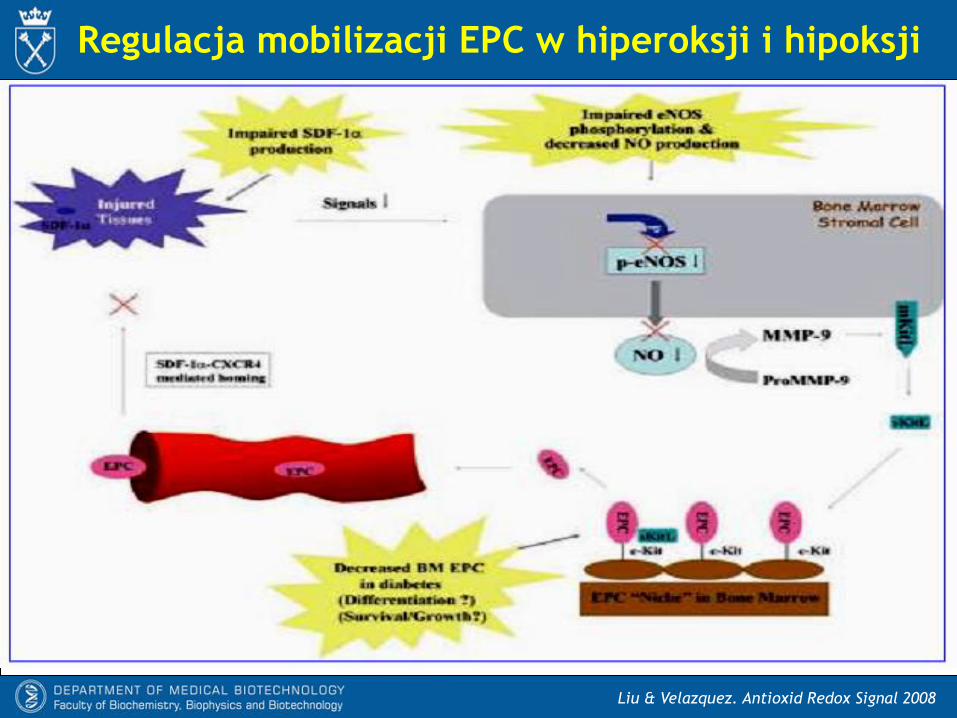
Liu & Velazquez. Antioxid Redox Signal 2008
Regulacja mobilizacji EPC w hiperoksji i hipoksji
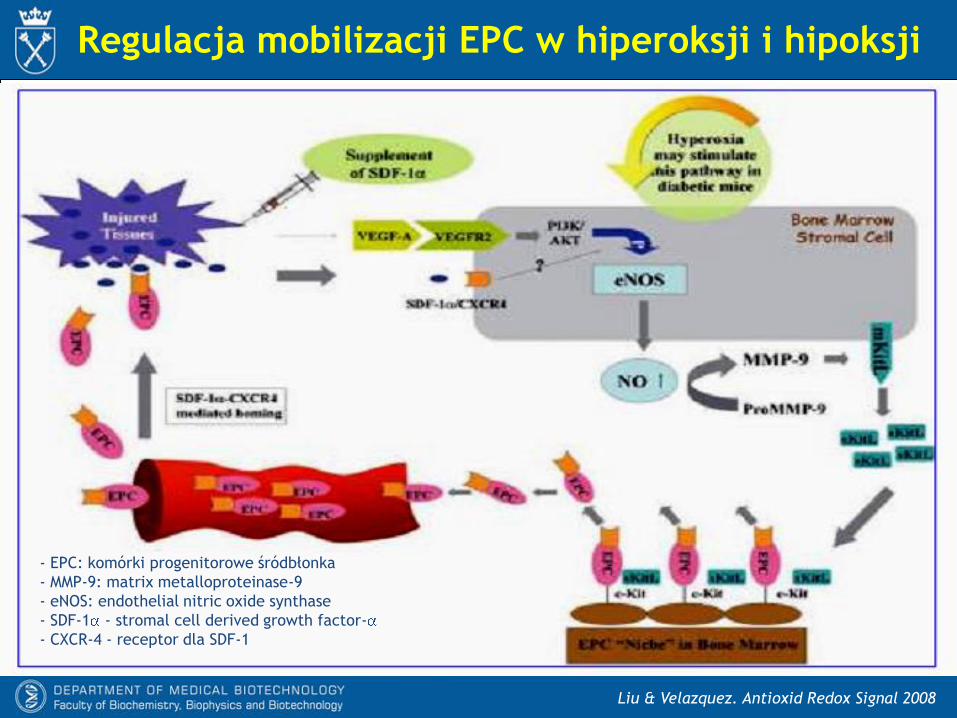
Liu & Velazquez. Antioxid Redox Signal 2008
Regulacja mobilizacji EPC w hiperoksji i hipoksji
- EPC: komórki progenitorowe śródbłonka
- MMP-9: matrix metalloproteinase-9
- eNOS: endothelial nitric oxide synthase
- SDF-1 - stromal cell derived growth factor-
- CXCR-4 - receptor dla SDF-1