biologia molecular reparac
TRANSCRIPT

BIOLOGIA MOLECULAR Blga. Olga Libia Cjuno Huanca
Universidad Nacional de San Antonio Abad del Cusco
Facultad de Ciencias Biologicas
CARRERA PROFESIONAL DE FARMACIA Y BIOQUIMICA
Unidad Académica N° 1: ESTRUCTURA Y FUNCION DEL ADN

FIDELIDAD DE LA REPLICACION
• La replicación del material genético ocurre una sola vez
durante el ciclo de vida de la célula por lo que es imprescindible que sea realizado con una elevada fidelidad de copia, que la secuencia de bases de las moléculas hijas sea exactamente igual a la de las células madre.
• Existen mecanismos que garantizan esta elevada fidelidad de copia entre ellos:
1. La elevada especificidad del apareamiento de bases formando los pares adenina-timina y citosina-guanina.
2. La elevada especificidad de las polimerasas que incorporan los desoxinucleótidos que forman pares complementarios al ADN que se está copiando.
3. Utilización de un ARN iniciador que al ser eliminado y reemplazado por el segmento de ADN correspondiente provoca que estas zonas sean rectificadas siempre.
4. Existencia de un mecanismo de rectificación de errores.
Blga. Olga L. Cjuno H.

Durante la replicación del ADN: • La nucleasa, ADN Polimerasa reparadora y la ligasa son
también una parte de la maquinaria de replicación. • La replicación es mucho más compleja, y aún subsisten
diversas incógnitas: Como, ¿La polimerasa en la cadena adelantada se conecta con la cadena retrasada para permitir que la replicación progreses sincrónicamente en ambas cadenas?. O los proceso de origen de replicación en eucariontes todavía no están bien dilucidadas.
• Dada la necesidad de precisión durante la replicación del DNA y la longitud en la que las células tienen que obtener esta precisión.
Blga. Olga L. Cjuno H.

LESIONES EN LA REPLICACION:
El material genético se halla en constante peligro de ser alterado no sólo por la acción de agentes ambientales sino también espontáneamente, por ejemplo como consecuencias de errores durante la replicación. Cuando las alteraciones del genoma involucran a uno o a unos pocos nucleótidos se denomina mutaciones génicas. Otras veces las alteraciones en de la magnitud que afecta al propio cariotipo motivo por el cual adquieren el nombre de aberraciones cromosómicas. Estas aberraciones pueden ser estructurales o numéricas. • Las estructurales son afectas partes extensas de uno de
los cromosomas, que pueden perderse (delección) o sufrir una inversión, una duplicación o una translocación.
• Las numéricas, en cambio, el cariotipo exhibe un número mayor o menor de cromosomas.
Blga. Olga L. Cjuno H.

TIPOS DE LESIONES GENICAS
Perdida de bases : Producción de sitios apurinicos o apirimidinicos.
Cambio de bases : Desaminaciones (CxU, AxApoxantina, etc).
Modificación Química de bases : Metilación, alquilación, hidroxilación, etc.
Rotura de enlaces fosfodiester : Cortes en las cadenas polinucleótidas.
Entrecruzamientos : Unión covalente en ambas cadenas.
Blga. Olga L. Cjuno H.

CAUSAS DE LAS LESIONES:
Espontáneas o : Desaminaciones por perdida de bases. Error de
Naturales replicación.
Inducidas : Por agentes:
Fisicos : Radiación UV, ionizantes.
Quimicos: Especies reactivas oxigenadas.
Acido nitroso
Agentes alquilantes (metilaciones, alquilaciones).
Agentes bifuncionales (entrecruzamientos.
Carcionógenos.
Blga. Olga L. Cjuno H.

Blga. Olga L. Cjuno H.

MUTACIONES
Las mutaciones se produce por cambios en la estructuras del ADN. Son accidentes de copia de las bases púricas o pirimidicas, que se produce en mayor frecuencia en la replicación del ADN, como consecuencia el ADN sintetizado no es replica exacta del ADN paterno. A pesar de todos los sistemas destinados a prevenir y corregir los posibles errores, estos pueden ser las mutaciones puntuales, que son el cambio de una de las bases de un par en el ADN, un cambio en una base nitrogenada puede alterar la estructura completa de la proteína: Tirosina- arginina- metionina AUG GCU UAC Tirosina- glutamina -metionina AUG GUU UAC Las mutaciones pueden ser: Silenciosas Puntuales
Blga. Olga L. Cjuno H.

SILENCIOSAS: • En un principio, las mutaciones silenciosas fueron irrelevantes para
la salud, ya que se pensaba que no alteraban la composición de las proteínas codificadas por los genes y que cualquier fallo en el proceso de fabricación de una proteína resultaría inocuo para el organismo.
• Esto se desmintió hace unos años ya que se descubrió que la células utilizaban de forma preferente ciertos codones, porque aumenta la velocidad y precisión de la síntesis de proteínas. Una Mutación silenciosa es aquella que provoca una alteración de un nucleotido para crear un codon sinónimo.
• Un codon sinónimo es un codon que se traduce en un mismo aminoacido, es decir, si por ejemplo tenemos el codon GUU el cual forma el aminoacido valina, los codones sinónimos de este serían GUC, GUA, GUG ya que también forman el aminoacido valina. Las mutaciones silenciosas pueden afectar a varios procesos de la síntesis de proteínas en nuestras células.
Blga. Olga L. Cjuno H.

MUTACIONES PUNTUALES: Afectan a un par de bases en la
secuencia del ADN , pueden ser:
Sustitución. Donde debería haber un nucleótido se inserta otro. Por ejemplo, en lugar de la citosina se instala una timina.
Inversión, mediante dos giros de 180° dos segmentos de nucleótidos de hebras complementarias se invierten y se intercambian.
Translocación. Ocurre un traslape de pares de nucleótidos complementarios de una zona del ADN a otra.
Desfasamiento. Al insertarse (inserción) o eliminarse (delección) uno o más nucleótidos se produce un error de lectura durante la traducción que conlleva a la formación de proteínas no funcionales.
Blga. Olga L. Cjuno H.

Blga. Olga L. Cjuno H.

Otras mutaciones incluyen la perdida de algunos pares de bases o incluso secuencias más grandes, multiplicaciones anormales. O traslados de secuencias de unas regiones del cromosoma a otras o entre diferentes cromosomas (translocaciones), causando alteraciones fenotípicas evidentes.
Blga. Olga L. Cjuno H.

Aunque las subidas intensas y rápidas de la temperatura pueden provocar mutaciones, los agentes físicos mutagénicos por excelencia son las radiaciones, que se dividen en ionizantes y no ionizantes. Radiaciones ionizantes. Son radiaciones de longitud de onda muy corta y, por tanto, muy energéticas, que provocan la ionización de los átomos de las sustancias que atraviesan. Entre estas radiaciones se encuentran los rayos X y g, así como las partículas α y b y los neutrones emitidos en procesos radiactivos.Los efectos de las radiaciones ionizantes sobre los seres vivos son de tres tipos: • Fisiológicos. Pueden producir cambios enzimáticos que se traducen en
modificaciones metabólicas. • Citogenéticos. Comportan alteraciones en la estructura de los
cromosomas, como deleciones y translocaciones. • Genéticos. Son producidos por ionizaciones directas del ADN o a través de
otros iones que, a su vez, provocan nuevas ionizaciones y la aparición de radicales libres muy reactivos. Éstos originan cambios químicos en el ADN que se traducen en mutaciones génicas, como la rotura de los enlaces nucleotídicos, la rotura y pérdida de bases nitrogenadas y la aparición de formas tautoméricas.
Blga. Olga L. Cjuno H.
AGENTES MUTAGENICOS FISICOS:


Radiaciones no ionizantes. Son, fundamentalmente, las radiaciones ultravioleta (UV). A diferencia de las anteriores, no producen ionizaciones. Su acción primaria consiste en provocar el paso de electrones a niveles energéticos más altos, lo cual puede dar lugar a formas tautoméricas y dímeros de timina.
• La luz UV une de forma covalente
dos residuos de timina adyacentes en
una hebra cadena de DNA, formando
ciclobutano.
• Los dímeros de timina producen una
distorsión local del DNA.
• Con menos frecuencia se pueden
formar otros dímeros de pirimidinas
T-T > T-C, C-T > C-C
Blga. Olga L. Cjuno H.

Agentes mutagénicos químicos : Numerosas sustancias tienen acción mutagénica (hidrocarburos policíclicos, aminas aromáticas, agentes alquilantes, colorantes industriales, pesticidas, etc.). A diferencia de las radiaciones, sus efectos, entre los que se pueden destacar los siguientes, suelen ser más retardados: • Modificaciones de las bases nitrogenadas. Comprenden las reacciones de
desaminación, alquilación e hidroxilación, que provocan emparejamientos erróneos. Ejm. La desaminación de la adenina da lugar a la hipoxantina, que se aparea con la citosina en lugar de la timina; la guanina se transforma por alquilación en 6-O-metilguanina, que se aparea con timina en lugar de citosina.
• Sustituciones de bases. Están causadas por análogos de bases
nitrogenadas que provocan un emparejamiento erróneo durante la replicación al cambiar una base por otra. Entre ellos se encuentran la 2-aminopurina y el bromouracilo.
• Introducción de ciertas moléculas en la cadena polinucleotídica del
ADN. Estas inserciones provocan la aparición de un exceso de nucleótidos en la hebra de nueva formación durante la replicación. De este modo, a partir de ese punto los tripletes de bases se alteran y el mensaje genético cambia.
Blga. Olga L. Cjuno H.

Agentes mutagénicos biológicos • Algunos agentes biológicos aumentan la frecuencia de la mutación
génica, Destacan entre ellos ciertos virus que pueden producir cambios en la expresión de algunos genes (por ejemplo, los retrovirus, los adenovirus o el virus de la hepatitis B humana, entre otros) y los transposones. Estos últimos son segmentos móviles de ADN que pueden cambiar de posición, trasladándose a otro lugar distinto dentro del mismo cromosoma o incluso a otro cromosoma.
• Estos elementos móviles se han encontrado en todo tipo de organismos (maíz, levaduras, insectos, bacterias, etc.) y pueden originar mutaciones, ya que causan una activación o inactivación génica no deseada al insertarse en los genes estructurales o en los reguladores. Se cree además, que los virus mutagénicos podrían realizar su acción al llevar en su genoma transposones tomados de una célula previamente infectada que incorporarían a la nueva célula parasitada, transposon intercalado en una secuencia génica normal.
Blga. Olga L. Cjuno H.

MUTACIONES GENICAS
• Las mutaciones génicas más comunes consisten en la sustitución de un nucleótido por otro, en la pérdida (delección de uno o varios nucleóticos o en la inserción (intercambio) de uno o varios nucleótidos en la molécula de ADN.
• Cualquiera que sea el tipo de mutación, genera un cambio en la información contenido en el gen y lleva a la producción de una proteína distinta de la esperada o a su falta de producción.
• Como se sabe, el cambio de un nucleótido en un gen da lugar a un codón diferente y en consecuencia, a la presencia en la proteína de un aminoácido que no corresponden (salvo que el nuevo codón sea sinónimo y por lo tanto, codifique al mismo aminoácido).
• Muchas veces el cambio de un solo aminoácido en una molécula proteíca produce alteraciones sustanciales en sus funciones, ya que al modificarse su estructura primaria se alternan también con sus estructuras secundarias y terciarias.
Blga. Olga L. Cjuno H.

CAUSAS DE MUTACIONES GENICAS Considerando las mutaciones génicas como sentido estricto, el estudio de las causas de estas, se puede considerar como el estudio de las causas de las mutaciones. Los otros tipos de mutaciones, se consideran más bien como alteraciones del material genético. Las mutaciones génicas pueden producirse por tres causas: por: • Errores de lectura durante la replicación del ADN, por • Lesiones fortuitas, como, por ejemplo, la rotura del enlace
que une una base nitrogenada a la desoxirribosa, o por • Transposiciones (cambios de posición) de ciertos
segmentos del gen.
Blga. Olga L. Cjuno H.

1.- Errores de lectura. Los errores de lectura que pueden aparecer durante la replicación del ADN pueden deberse a dos causas: a los cambios tautoméricos y a los cambios de fase. • a) Los cambios tautoméricos. Cada base nitrogenada puede
presentarse en dos formas diferentes denominadas formas tautoméricas o tautómeros, una es la normal y la otra la rara. Ambas formas están en equilibrio, y espontáneamente se pasa de una a la otra, lo que se denomina cambio tautomérico. Esto, si sucede durante la replicación, implica mutaciones, ya que cambia la base complementaria en la nueva hebra de ADN. Por ejemplo, la forma normal de la G se complementa con la C, mientras que la forma rara de G, es decir, su forma tautomérica, lo hace con la T.
• b) Los cambios de fase. Son deslizamientos de la hebra que se está formando sobre la hebra molde, de forma que quedan bucles al volverse a emparejar. El crecimiento sigue y la diferencia queda fijada, originándose así la mutación.

En la conformación normal del ADN la guanina y la timina se encuentran en forma ceto (C=O) y pueden cambiar a la forma enol (COH), lo que se conoce como cambio tautomérico. Por su parte, la adenina y la citosina se presentan normalmente en forma amino (NH2) pero pueden tautomerizarse a formas imino (NH)

2. Lesiones fortuitas. Las lesiones fortuitas son alteraciones de la estructura de uno o de varios nucleótidos, que aparecen de forma natural. Las más frecuentes son: • a) Despurinización. Pérdida de purinas por rotura del
enlace entre éstas y las desoxirribosas. Se producen a razón de unas 5.000 a 10.000 por día, en cada célula humana.
• b) Desaminación. Pérdida de grupos amino en las bases nitrogenadas, que entonces se emparejan con una distinta de la normal. Se producen unas 100 por genoma y día.
• c) Dímero de timina. Enlace entre dos timinas contiguas. Generalmente provocado por los rayos ultravioleta de la radiación solar.
Blga. Olga L. Cjuno H.

Despurinación y Desaminación
Sitio apurínico Sitio apurínico 1. Perdida de bases
2. Cambio de bases
DESPIRIMIDIZACIÓN
Blga. Olga L. Cjuno H.

Desaminación de bases
adenina hipoxantina
guanina xantina
citosina uracilo
timina
no hay desaminación
5-metil citosina timina
Blga. Olga L. Cjuno H.
(menos frecuentes)
A Hipoxantina
G Xantina


3. Transposiciones. Son cambios de lugar espontáneos de determinados segmentos de ADN, los denominados elementos genéticos transponibles. Éstos pueden ser menores que un gen (como las llamadas secuencias de inserción), un gen, o un grupo de genes (como los denominados transposones). Las transposiciones pueden producir mutaciones génicas si el elemento genético transpuesto se sitúa dentro de un gen, o mutaciones cromosómicas si pasa a un lugar donde no hay un gen, ya sea dentro del mismo cromosoma o incluso a otro cromosoma.

Rotura de Enlaces Fosfodiester:
Cortes en las cadenas polinucleótidas
Blga. Olga L. Cjuno H.

Entrecruzamientos: Unión covalente de ambas cadenas
Blga. Olga L. Cjuno H.

Modificacion Quimica de Bases:
Metilación
Blga. Olga L. Cjuno H.

Alquilación de bases
Agentes alquilantes
Puede aparearse con C o T,
produciendo transversiones
La alquilación de la posición N7 de un nucleótido de purina, hace que su
enlace glucosídico sea susceptible a la hidrólisis y lleve a la pérdida de la
base: despurinación
Blga. Olga L. Cjuno H.

Oxidación de bases
El metabolismo celular expone el DNA a los efectos perjudiciales de las
especies reactivas de oxígeno (ROS: O2-., HO., H2O2), subproductos naturales
del metabolismo oxidativo.
Se conocen más de 100 modificaciones oxidativas diferentes en el DNA.
p. Ej.: G puede oxidarse a 8-oxoguanina
oxoG puede aparearse con C o con A.
Si oxoG se con A se produce
una TRANSVERSIÓN.
TRANSVERSIÓN: mutación
puntual producida por la
sustitución de una purina por una
pirimidina o al revés.
G C T =A Blga. Olga L. Cjuno H.

EFECTOS SOBRE EL ADN METILACIONES
Blga. Olga L. Cjuno H.

REPARACION DEL ADN Gran N° de lesiones es detectado y
reparado por el sistema de reparación del ADN
Lo que no se puede reparar
Errores en la replicación
Si permanecen estos errores
Cambios de errores permantes de la información genética
“mutación” Blga. Olga L. Cjuno H.

REPARACION DEL ADN
Blga. Olga L. Cjuno H.


SISTEMAS DE REPARACION DEL ADN
• Para cada alteración del ADN existe un mecanismo de reparación particular, dirigido por un conjunto de enzimas específicas.
• En la mayoría de los casos se basan en la información genética complementaria existente entre las dos cadenas de ADN, de modo que si una de ellas sufre alguna alteración (mutación), puede ser reparada a partir de información normal contenida en la otra cadena.
• Asi mismo los mecanismos reparadores de errores también pueden fallar de allí la aparición de una mutación génica.
Blga. Olga L. Cjuno H.

SISTEMAS DE REPARACION DEL ADN
• Por reparación y reversion directa:
• R. de apareamiento incorrectos.
• R. por eliminación de bases alternadas
• R. escisión de nucleótidos.
• R. por recombinación
• R. tendencia al error.
Blga. Olga L. Cjuno H.

- Durante la replicación del ADN, para que un nucleótido pueda ser agregado en el extremo 3´de la cadena hija en crecimiento es imprescindible que el nucleótido incorporado precedentemente sea el adecuado, ya que si la ADN polimerasa inserta en forma accidental el nucleótido incorrecto, percibe el error y no agrega nuevos nucleótidos, la cual detiene transitoriamente el crecimiento de la cadena el error es resuelto por la propia enzima (propiedad de lectura de pruebas).
- Así la polimerasa retrocede e inserta el nucleótido correcto y la sintesis de ADN progresa normalmente. Blga. Olga L. Cjuno H.

Si se da el caso que falle la lectura de pruebas, se pone en marcha un segundo sistema de reparación, que se cumple en tres pasos:
1. El o los nucleótidos erróneos son removidos por una nucleasa
reparadora, la misma que remueve a los cebadores en la síntesis continua y discontínua del ADN, para lo cual la nucleasa corta las uniones fosfodiester donde se ligan los nucleótidos correctos con los incorrectos.
2. El espacio que queda vacio es llenado por nucleótidos adecuados mediante una reacción conducida por el ADN polimerasa ᵦ. Y se completa, cuando
3. El ADN ligasa conecta el extremo 3´del nuevo ADN con el extremo 5´del ADN cortado. Debe existir alguna señal que le permita a la nucleasa reparadora distinguir en cual de las dos cadenas del ADN se encuentran el nucleótido incorrecto.
Blga. Olga L. Cjuno H.

Blga. Olga L. Cjuno H.

• Fuera de la replicación, la aparición del uracilo en lugar de citocina en el ADN, (x desaminaciones espontáneas), desencadena en un mecanismo de reparación distinto, que utiliza una ADN glicosilasa específica, estas reconocen y cortan la conexión química entre la base errónea y la desoxiribosa ligada a ella y deja al nucleótido sin su base.
• Igualmente las ADN glicosilasas específicas remueve la hipoxantina (desaminación de adenina).
Blga. Olga L. Cjuno H.

• La desoxiribosa sin base o sitio AP (x apurinización o apirimidización) es reconocida y por lo tanto removida por las enzimas que actuan en forma sucesiva.
• Los sitios AP apurinicos (x desaminaciones) se producen al perderse alguna purina en forma directa y son reconocidos y reparados por las mismas enzimas correctoras de los sitios AP surgidos las las desaminaciones.
Blga. Olga L. Cjuno H.

• La mayor parte de las mutaciones inducidas por agentes ambientales son reparadas por los mismos mecanismo que utilizan en las correciones de las mutaciones espontáneas.
• Los dímeros de timina generados, por la luz UV son removidos por un sistema de enzimas especiales que hidrolizan simultáneamente dos uniones fosfodiester, una en cada lesión.
Blga. Olga L. Cjuno H.

Blga. Olga L. Cjuno H.

Biología Celular y Molecular 5ª ed., Lodish y col.
Blga. Olga L. Cjuno H.

La replicación del material genético es esencial a la vida. Asegura la continuidad de la información genéticadurante el crecimiento y la reparación de tejidos. Además de la continuidad de la información genética de padres e hijos, a través de las generaciones y hace posible la continuidad de la vida. Los errores en la replicación son el orígenes de las enférmedades hereditarias y también son causas primarias de cáncer.
Durante la Etapa G1, del ciclo celular, la célula “chequea” sus condiciones para dividirse, tales como el tamaño y el estado del ADN. Durante la etapa S, tiene lugar la replicación del ADN. Luego, durante la etapa G2 la “maquinaria se reparación” del ADN repara los errores que podrían haber ocurrido durante la replicación. De esta manera, si durantes la replicación de han acumulado suficientes errores, la etapa G2 se alarga, hasta que el ADN esté en condiciones de continuar el ciclo celular.

CICLO CELULAR Y REGULACION DE LA EXPRESION GENICA
En eucariotas , la sintesis de ADN se realiza en un momento preciso de su cilo de vida, en las bacterias es continua. La proliferación y la progresión del ciclo celular están funcionalmente ligadas a los genes asociados con el control del crecimiento celular e incluyen señales positivas y negativas que regulan el crecimiento como se puede ver en varios casos como: • Ciclo de la mitosis y meiosis para la generación de células somaticas y
gamentos. • Estimulación de células quiescentes para la proliferación de sitios de
cicatrización. • Salida del ciclo celular para que una célula prolifere a diferenciación.
Blga. Olga L. Cjuno H.

El destino de la célula esta programado por su programa genético y por las condiciones ambientales
Blga. Olga L. Cjuno H.
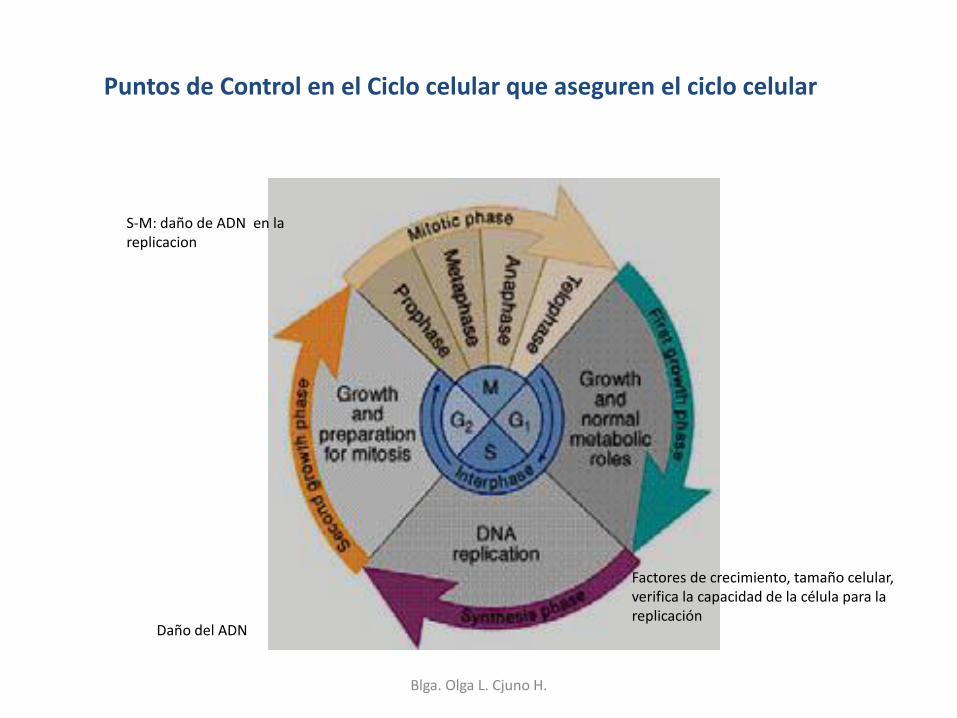
Puntos de Control en el Ciclo celular que aseguren el ciclo celular
S-M: daño de ADN en la replicacion
Daño del ADN
Factores de crecimiento, tamaño celular, verifica la capacidad de la célula para la replicación
Blga. Olga L. Cjuno H.

PROTEINAS IMPLICADAS EN EL CICLO CELULAR: Cdk y Ciclinas. A nivel molecular el ciclo celular está regulado por actividades fluctuantes, parciales o temporales de complejos proteicos como las Ciclinas o las cinasas dependientes de las ciclinas (Cdk).
La división de celulas varian según el tipo celular: - Algunas celulas se dividen durante
toda la vida (cels. Madre que producen sangre).
- Otras se dividen para reparar la lesión (cels. De la piel).
- Otras no se dividen nunca (neuronas, miofibrillas musculares, bloqueado en Go.
Blga. Olga L. Cjuno H.

Cinasas dependientes de la ciclinas (Cdk): constitiyen una nueva clase de cinasas, son activas si se unen a ciclinas, las cinasas fosforilan ciertas Ser/Ther (profase 1) Ciclinas: A y B, primeras descubiertas, se sintetizan constantemente en interfase y despues son degradas bruscamente por enzimas proteolíticas al final de cada mitosis. Existen diferentes ciclinas: A, B, D1, D2, D3, E, G, etc. Se asocian a distintas Cdk durante las sucesivas fases del ciclo para formar complejos enzimáticamente activos.
Blga. Olga L. Cjuno H.

PUNTOS DE CONTROL CRUCIALES EN EL CICLO CELULAR:
Transicion G1-S: Punto sin retorno o punto R de restriccion: Una franqueado la celula entre de manera irreversible en la fase S, aun tendrá dos posibilidades: división o suicidio (apoptosis). Las Cdk2 y Cdk4, estan implicadas en G1.
Blga. Olga L. Cjuno H.

Transicion G2-M: Interviene otro control en punto T de transición. La proteina Cdk1 y la ciclina B se asocian y desempeñan papel esencial para desencadenar la entrada en mitosis. Las enzimas (se fosforilan o desfosforilan) según el caso. La desfosforilación de la CdK1 desencadena la entrada en mitosis, también provoca la fosforilación de una molecula que degradará la ciclina. La Cdk1 se inactiva al asociarse con la ciclina B. La mitosis termina en interfase. Y el ciclo comienza de nuevo.
Blga. Olga L. Cjuno H.

En el curso de la meiosis, se observa la sucesión de dos ciclos de división particulares: en el segundo ciclo no hay Fase S. La célula pasará pues de 2n a n Dos divisiones sucesivas sin fase S sucesivas sin fase M previa, conducirán a un aumento de ploidia (número de copias de cromosomas en una célula) haploide:1, diploide:2.
La fosfatas cdc-25 es inactiva al principio del ciclo se activa alcanzando cierto nivel de ciclina B, la cdc-25 activa la Cdk1
Blga. Olga L. Cjuno H.
