biological mechanisms of phorbol myristate acetate-induced … · phorbol myristate acetate affects...
TRANSCRIPT

[CANCER RESEARCH 43, 88-96, January 1983]0008-5472/83/0043-0000302.00
Biological Mechanisms of Phorbol Myristate Acetate-induced Inhibitionof Proadipocyte Differentiation1
Kankatsu Yun and Robert E. Scott2
Section of Experimental Pathology. Departments of Pathology and Cell Biology, Mayo Clinic and Foundation, Rochester, Minnesota 55905
ABSTRACT
The differentiation of murine proadipocytes can be inducedby two different methods: 7.) by culture of high-density proadipocytes in medium containing high concentrations of serum towhich is added a variety of hormones and 2.) by culture of low-
density proadipocytes in heparinized medium containing onlyhuman plasma. Phorbol myristate acetate influences differentiation in both culture systems, but the biological mechanismsby which phorbol myristate exerts its effect have not beenestablished previously. In this regard, we have recently shownthat five distinct steps mediate the integrated control of proad-
ipocyte proliferation and differentiation (J. Cell Biol., 94: 400,1982). Three steps are involved in the differentiation process:(a) growth arrest at a distinct state in the Gìphase of the cellcycle (Go); (b) nonterminal differentiation; and (c) terminaldifferentiation. Two steps antagonize the differentiation process and cause proliferation. These are (a) induction of loss ofthe differentiated phenotype and (t>) reinitiation of proliferation.In this paper, we have therefore examined the effects of thetumor promoter phorbol myristate acetate on each step ofintegrated control of proliferation and differentiation in high-and low-density proadipocytes. The results show that in thehigh-density system, phorbol myristate acetate can have the
following effects under appropriate experimental conditions. Itcan: (a) block growth arrest at GD; (o) inhibit nonterminaldifferentiation; (c) promote loss of the differentiated phenotype;and (cO promote the proliferation of Go-arrested cells. In thelow-density system by contrast, phorbol myristate acetate (a)
does not block GD arrest and (o) does not block nonterminaldifferentiation but (c) does promote loss of the differentiatedphenotype and (d) does promote the proliferation of Go-arrested cells. These results show that the mechanisms by whichphorbol myristate acetate affects differentiation are significantly influenced by a variety of factors which include changesin the culture medium, cell density, and cell-cell contact. There
fore, attempts to correlate general effects of phorbol myristateacetate on the control of cell proliferation and differentiationwith its action as a tumor promoter may not be justified. Wesuggest that only actions of phorbol myristate acetate whichselectively affect initiated cells should be considered to berelevant to the mechanisms of promotion of carcinogenesis.
INTRODUCTION
The phorbol esters modulate cellular differentiation in avariety of cell types (8, 10, 16, 18,25-27,30,31,38,39,41,45, 46). The specific mechanism(s) which mediates these
biological effects has, however, not been established. Onereason is that phorbol esters have a marked effect on numerousaspects of cellular physiology. For example, PMA3 affects cell
morphology (9); cell proliferation (19, 40); cell cycle arrest(24); nucleic acid and protein synthesis (15); the activity ofornithine decarboxylase (22, 27, 43); fibronectin organization(42); and the intracellular concentration of cyclic nucleotides(3), prostaglandins (21, 43), polyamines (17), and plasmamembrane structure and function (1, 7, 12, 13, 29, 37). Another important reason for the difficulty in determining howphorbol esters modulate differentiation is that differentiation isa complex process which has not been clearly defined in mostcell systems.
We have, however, recently characterized in detail the biological steps which are involved in the integrated control ofmurine proadipocyte proliferation and differentiation (20,33-36, 44). Three stages occur in the process of differentia
tion: (a) growth arrest at a distinct state in the Gìphase of thecell cycle, designated GD; Ob) nonterminal differentiation andarrest at a state designated GD'; and (c) terminal differentiation
Two stages also antagonize the differentiation process: (a)induction of loss of differentiated phenotype; and (b) reinitiationof cell proliferation. A schematic model of the relationship ofthese stages in the integrated control of proadipocyte proliferation and differentiation is presented and described in detailin Chart 1. Based on this model, we have studied the effects ofPMA on proadipocytes in 2 different differentiation-promoting
systems. The results show that changes in cell culture conditions and in cell density have a significant influence on theactions of PMA. These results are discussed with respect tothe action of phorbol myristate as a modulator of cell proliferation and differentiation with its action as a promoter of carcinogenesis (2-6, 11, 15, 19, 23, 32).
MATERIALS AND METHODS
Cell Culture
BALB/3T3 T-proadipocytes, a derivative of clone A31 BALB/3T3
mouse embryo cells (a gift from Dr. Leila Diamond, Wistar Institute,Philadelphia, Pa.) (9,10), were cultured at 37°in a 5% CO2 atmosphere
in antibiotic-free DMEM (Grand Island Biological Co., Grand Island,N. Y.) containing 10% heat-inactivated PCS (KC Biologicals, Lenexa,
Kans.). These cells are similar to Swiss 3T3 L1 proadipocytes described by Green and Kehinde (14). Stock cultures were initiated every6 to 8 weeks from specimens frozen in liquid nitrogen. All stockcultures were maintained at low density by passage at <50% confluence to avoid selection for cells which were not density inhibited. The
1This research was supported in part by NIH Grant CA 28240 and by the
Mayo Foundation to R. E. S.2 To whom requests for reprints should be addressed.
Received May 12, 1982; accepted October 8, 1982.
3 The abbreviations used are: PMA, phorbol myristate acetate; GD. Gìarreststate that precedes differentiation; GD', Gìarrest state for nonterminally differentiated cells; DMEM, Dulbecco's modified Eagle's medium; PCS, fetal calf
serum; MIX, methyl isobutyl xanthine.
88 CANCER RESEARCH VOL. 43
on March 3, 2020. © 1983 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

PMA-induced Inhibition of Proadipocyte Differentiation
Chart 1. Five biological steps are involved in the integrated control of proad-ipocyte proliferation and differentiation (35). Three steps mediate the differentiation process. Cells are first induced to growth arrest at a distinct state in the G,phase of the cell cycle (Arrow 7). Following arrest at this state, designated GD,the cells then undergo a process of nonterminal differentiation (Arrow 2). Cellscan arrest at a state of nonterminal differentiation, designated Go'; or they can
proceed to terminally differentiate (Arrow 3). Two steps antagonize the differentiation process, (a) Go-arrested cells can be induced to reinitiate proliferation(Arrow 5) and (6) GD'-arrested, nonterminally differentiated cells can be induced
to lose the differentiated phenotype (Arrow 4) and then to reinitiate proliferation(Arrow 5). The model shows that the integrated control of cell proliferation anddifferentiation can be mediated at both GD and GD'. This is so because GD cells
can either remain quiescent at GD, they can be induced to differentiate, or theycan be induced to proliferate. Further, GD'-arrested cells can remain quiescentat Go', they can terminally differentiate, or they can be induced to lose the
differentiated phenotype and then reinitiate proliferation. Specific human plasmafractions have been shown to induce G arrest and nonterminal and terminaldifferentiation, and other physiological agents and drugs have been shown tocause loss of the differentiated phenotype and proliferation (34-36).
3T3 T-proadipocyte cell line was also shown to be free of Mycoplasmacontamination as described (33-36).
Differentiation of 3T3 T-Proadipocytes
Two methods were used to induce proadipocyte differentiation. InMethod 1, differentiation was induced in high-density proadipocyte
cultures with high concentrations of serum and insulin (10, 34). InMethod 2, differentiation of low-density proadipocytes was induced by
heparinized medium containing only human plasma (20).High-Density Differentiation System. Differentiation of confluent
3T3 T-proadipocytes was induced by feeding the cells DMEM containing 30% PCS and 50 fig insulin per ml at 4-day intervals. Under these
conditions, growth arrest, determined by autoradiographic analysis ofdecreased [3H]thymidine incorporation into DNA (34), occurred within2 to 4 days. At this time, the cell density was ~1 x 105 cells/sq cm.
Flow microfluorimetry showed that >90% of such cells had a 2ncontent of DNA. In this system, the first evidence of adipocyte differentiation typically occurred ~4 days after growth arrest, and maximum
differentiation was usually observed 12 to 16 days later.Low-Density Differentiation System. The differentiation of low-den
sity proadipocytes was induced by feeding rapidly growing culturesheparinized (30 units/ml) DMEM containing 25% human plasma at 4-
day intervals. G, growth arrest typically occurred within 2 days afterthe initial feeding, and cells reached a final cell density of ~1 x 104
cells/sq cm. In this system, the first evidence of differentiation wasobserved ~6 days after addition of differentiation-promoting medium
(20), and maximum differentiation of 75 to 95% of the cells typicallyoccurred between 5 and 8 days after growth arrest.
Platelet-poor human plasma used in these studies was prepared as
follows. Venous blood was drawn and placed immediately in sodiumcitrate at a final concentration of 0.38%. Citrate anticoagulated blood
was sedimented by centrifugation at 1,000 x g for 30 min at 22°.The
supernatant was again sedimented by centrifugation at 25,000 x g for30 min at 22°to remove platelets. This citrate anticoagulated platelet-poor plasma was frozen at -70° overnight and subsequently thawedat 4°.It was sedimented by centrifugation at 1,000 x g for 10 min at4°and then stored at 4°following Millipore filtration. Human plasma
was then added to heparinized DMEM at the appropriate concentration.Optimum adipocyte differentiation was typically induced by DMEMcontaining 25% human plasma and heparin (30 units/ml).
Quantitation of Adipocyte Differentiation
Adipocyte differentiation was quantitated morphologically by phasemicroscopy or by light microscopic examination of Oil Red "O"-stained
specimens. Cells were designated to be adipocytes when they contained numerous fat droplets within the cytoplasm which displaced thenucleus. Illustrations have been published previously which show suchcells (20, 34). The percentage of adipocytes and the density of adipocytes per sq cm were determined by counting ~500 cells. Previous
data have established that, in these culture systems, there is anexcellent correlation between increased activity of lipoprotein lipaseand glycerol-3-phosphate dehydrogenase, which are differentiationmarker enzymes for adipocytes (20, 34, 35), and morphological differentiation. Therefore, only morphological criteria were used in thestudies described in this paper. The advantage of using morphologicalcriteria for differentiation assessment is that they can be applied toindividual cells so that the actual density and percentage of differentiated cells can be determined. Enzyme assays which have been usedpreviously do not in general afford the opportunity for quantitation onan individual cell basis.
Autoradiography
To determine the extent of DNA synthesis in various cell populations,cultures were pulse labeled for 48 hr with [3H]thymidine (5 jiCi/ml)
(specific activity, 80 Ci/mmol; New England Nuclear, Boston, Mass.).Thereafter, the cells were processed for autoradiography as describedpreviously (20, 34, 35). The mitogenic response of cells to PMA andother compounds was also determined by this method. The percentageof radiolabeled nuclei per 300 to 500 cells was typically assayed ineach determination.
Specific Mitogenic Effect of MIX
We have reported that prior to differentiation, proadipocytes mustfirst growth arrest at a distinct state in the d phase of the cell cycle,GD (34). One important characteristic of Go-arrested cells is that theycan be induced to reinitiate proliferation when exposed to 5 x 10~4 M
MIX, whereas cells Gìgrowth arrested by serum deprivation cannot(34-36). The mitogenic responsiveness of cells to MIX was therefore
used as an assay for cells G, growth arrested at GD.
Flow Microfluorimetric Analysis
Cells were removed from culture flasks with 0.25% tryspin-0.1%EDTA in phosphate-buffered saline (pH 7.4) at room temperature. The
cell suspension was dissociated into single cells by pipetting. To thecell suspension was then added a solution of phosphate-bufferedsaline, pH 7.4, containing 0.75 mw Ca2+, 0.5 mM Mg2*, DNase (0.01
mg/ml), and trypsin inhibitor (0.2 mg/ml). Cells were sedimented bycentrifugation and were fixed with 70% ethanol overnight at 4°.Spec
imens were stained with mithramycin as described (20, 33-36). Flow
microfluorimetric analyses were performed with a computerized FACSIV apparatus (Becton, Dickinson, Mountain View, Calif.).
Materials
Phorbol esters were purchased from Consolidated Midland Corp.,Milwaukee, Wis. They were dissolved in dimethyl sulfoxide; the final
JANUARY 1983 89
on March 3, 2020. © 1983 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
K. Yun and R. E. Scott
concentration of this solvent in the culture medium was <0.1% in allassays. This concentration of dimethyl sulfoxide showed no effect on3T3 T-proadipocyte differentiation, nor was it cytotoxic. MIX, insulin,
and other reagents were purchased in the highest purity available fromSigma Chemical Co., St. Louis, Mo., or other suppliers. The commercialsource of DMEM, serum, and [3H]thymidine was described above.
RESULTS
Effects ofPMA in the High-Density Culture System
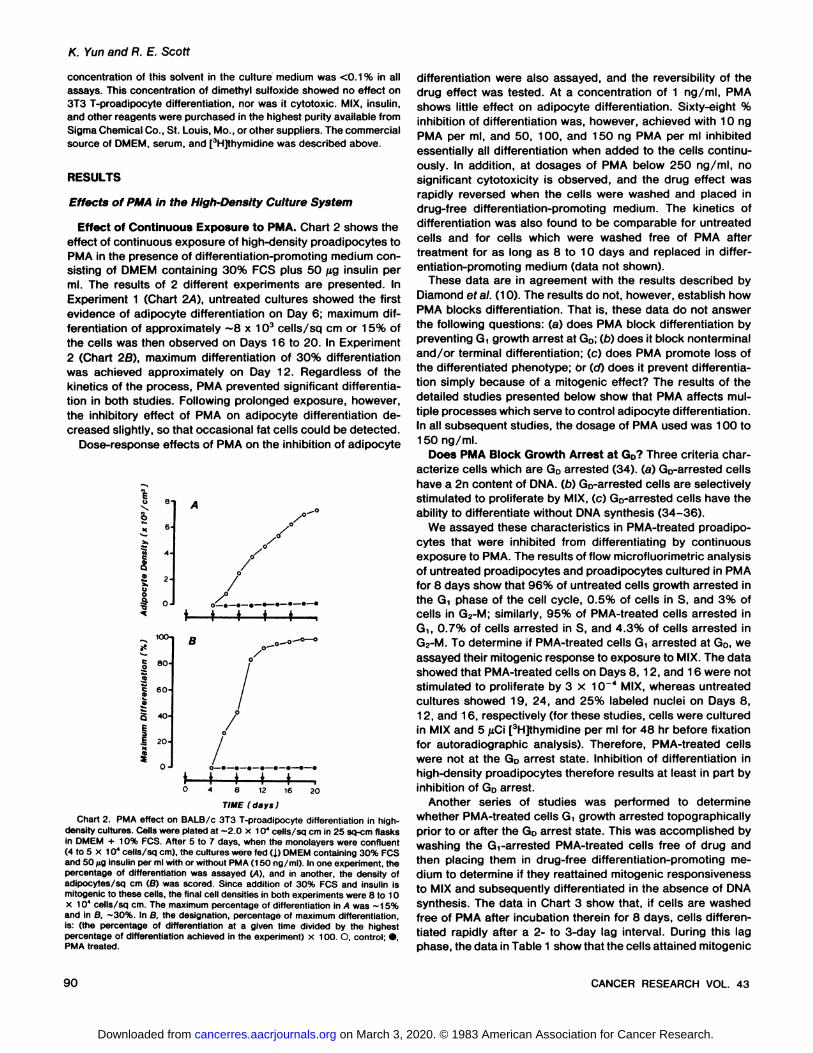
Effect of Continuous Exposure to PMA. Chart 2 shows theeffect of continuous exposure of high-density proadipocytes toPMA in the presence of differentiation-promoting medium consisting of DMEM containing 30% PCS plus 50 jug insulin perml. The results of 2 different experiments are presented. InExperiment 1 (Chart 2/4), untreated cultures showed the firstevidence of adipocyte differentiation on Day 6; maximum differentiation of approximately ~8 x 103 cells/sq cm or 15% of
the cells was then observed on Days 16 to 20. In Experiment2 (Chart 28), maximum differentiation of 30% differentiationwas achieved approximately on Day 12. Regardless of thekinetics of the process, PMA prevented significant differentiation in both studies. Following prolonged exposure, however,the inhibitory effect of PMA on adipocyte differentiation decreased slightly, so that occasional fat cells could be detected.
Dose-response effects of PMA on the inhibition of adipocyte
8 12 16 20
TIME (days)Chart 2. PMA effect on BALB/c 3T3 T-proadipocyte differentiation in high-
density cultures. Cells were plated at -2.0 x 10* cells/sq cm in 25 sq-cm flasks
in DMEM + 10% FCS. After 5 to 7 days, when the monolayers were confluent(4 to 5 x 104 cells/sq cm), the cultures were fed (J.) DMEM containing 30% PCS
and 50 /ig insulin per ml with or without PMA (1 50 ng/ml). In one experiment, thepercentage of differentiation was assayed (A), and in another, the density ofadipocytes/sq cm (ß)was scored. Since addition of 30% PCS and insulin ismitogenic to these cells, the final cell densities in both experiments were 8 to 10x 10* cells/sq cm. The maximum percentage of differentiation in A was -15%and in B, ~30%. In B, the designation, percentage of maximum differentiation,is: (the percentage of differentiation at a given time divided by the highestpercentage of differentiation achieved in the experiment) x 100. O, control; •,PMA treated.
differentiation were also assayed, and the reversibility of thedrug effect was tested. At a concentration of 1 ng/ml, PMAshows little effect on adipocyte differentiation. Sixty-eight %
inhibition of differentiation was, however, achieved with 10 ngPMA per ml, and 50, 100, and 150 ng PMA per ml inhibitedessentially all differentiation when added to the cells continuously. In addition, at dosages of PMA below 250 ng/ml, nosignificant cytotoxicity is observed, and the drug effect wasrapidly reversed when the cells were washed and placed indrug-free differentiation-promoting medium. The kinetics ofdifferentiation was also found to be comparable for untreatedcells and for cells which were washed free of PMA aftertreatment for as long as 8 to 10 days and replaced in differentiation-promoting medium (data not shown).
These data are in agreement with the results described byDiamond et al. (10). The results do not, however, establish howPMA blocks differentiation. That is, these data do not answerthe following questions: (a) does PMA block differentiation bypreventing G! growth arrest at GD;(6) does it block nonterminaland/or terminal differentiation; (c) does PMA promote loss ofthe differentiated phenotype; Or (cOdoes it prevent differentiation simply because of a mitogenic effect? The results of thedetailed studies presented below show that PMA affects multiple processes which serve to control adipocyte differentiation.In all subsequent studies, the dosage of PMA used was 100 to150 ng/ml.
Does PMA Block Growth Arrest at GD? Three criteria characterize cells which are GD arrested (34). (a) GD-arrested cellshave a 2n content of DNA. (ó)Go-arrested cells are selectivelystimulated to proliferate by MIX, (c) GD-arrested cells have theability to differentiate without DNA synthesis (34-36).
We assayed these characteristics in PMA-treated proadipo
cytes that were inhibited from differentiating by continuousexposure to PMA. The results of flow microfluorimetric analysisof untreated proadipocytes and proadipocytes cultured in PMAfor 8 days show that 96% of untreated cells growth arrested inthe Gìphase of the cell cycle, 0.5% of cells in S, and 3% ofcells in G2-M; similarly, 95% of PMA-treated cells arrested in
Gì,0.7% of cells arrested in S, and 4.3% of cells arrested inG2-M. To determine if PMA-treated cells d arrested at GD, we
assayed their mitogenic response to exposure to MIX. The datashowed that PMA-treated cells on Days 8,12, and 16 were notstimulated to proliferate by 3 x 10"4 MIX, whereas untreated
cultures showed 19, 24, and 25% labeled nuclei on Days 8,12, and 16, respectively (for these studies, cells were culturedin MIX and 5 /iCi [3H]thymidine per ml for 48 hr before fixation
for autoradiographic analysis). Therefore, PMA-treated cells
were not at the GD arrest state. Inhibition of differentiation inhigh-density proadipocytes therefore results at least in part by
inhibition of GD arrest.Another series of studies was performed to determine
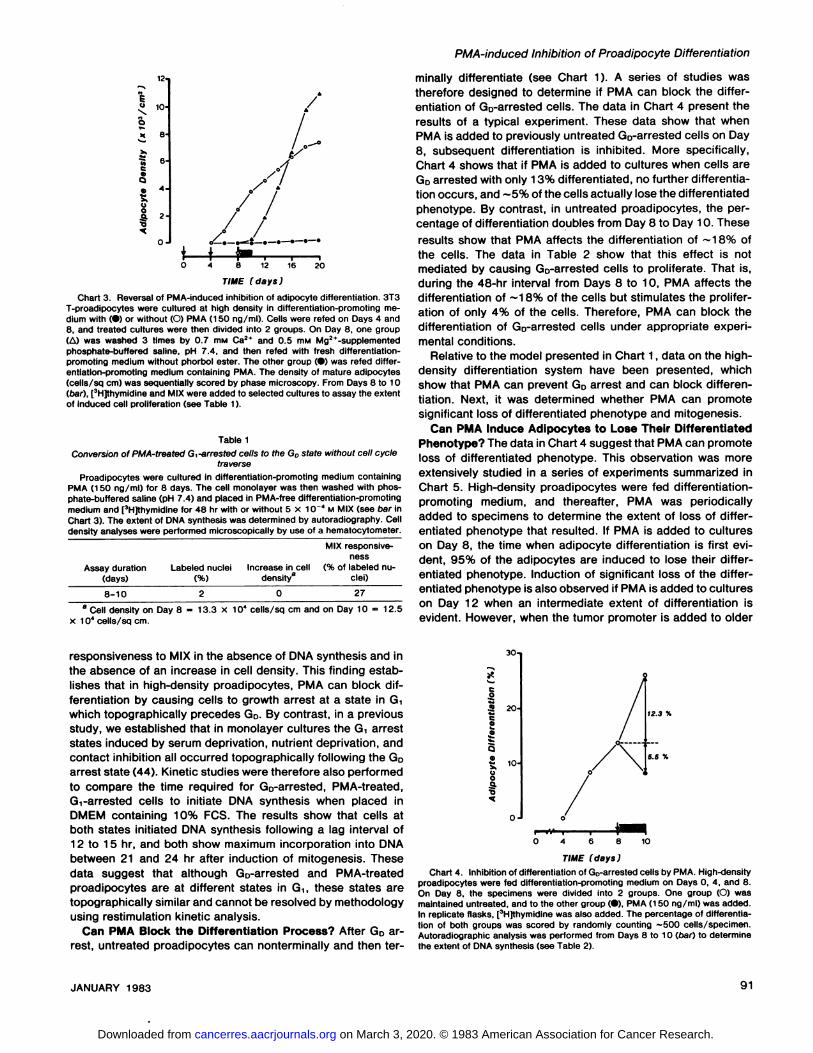
whether PMA-treated cells Gìgrowth arrested topographically
prior to or after the GD arrest state. This was accomplished bywashing the Gi-arrested PMA-treated cells free of drug andthen placing them in drug-free differentiation-promoting me
dium to determine if they reattained mitogenic responsivenessto MIX and subsequently differentiated in the absence of DNAsynthesis. The data in Chart 3 show that, if cells are washedfree of PMA after incubation therein for 8 days, cells differentiated rapidly after a 2- to 3-day lag interval. During this lagphase, the data in Table 1 show that the cells attained mitogenic
90 CANCER RESEARCH VOL. 43
on March 3, 2020. © 1983 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
IP»'
10-
l -
oI -0-/ ///o^._.-eî_._._•—•—•T
t i» . . .8 12 16
T7ME (days;
2O
Chart 3. Reversal of PMA-induced inhibition of adipocyte differentiation. 3T3T-proadipocytes were cultured at high density in differentiation-promoting medium with (•)or without (O) PMA (1 50 ng/ml). Cells were refed on Days 4 and8, and treated cultures were then divided into 2 groups. On Day 8. one group(A) was washed 3 times by 0.7 mw Ca2* and 0.5 mM Mg2*-supplemented
phosphate-buffered saline, pH 7.4, and then refed with fresh differentiation-promoting medium without phorbol ester. The other group (•)was refed differentiation-promoting medium containing PMA. The density of mature adipocytes(cells/sq cm) was sequentially scored by phase microscopy. From Days 8 to 10(bar), [3H]thymidine and MIX were added to selected cultures to assay the extent
of induced cell proliferation (see Table 1).
Table 1Conversion of PMA-treated G,-arrested certs to the GD state without cell cycle
traverseProadipocytes were cultured in differentiation-promoting medium containing
PMA (150 ng/ml) for 8 days. The cell monolayer was then washed with phosphate-buffered saline (pH 7.4) and placed in PMA-free differentiation-promotingmedium and [3H)thymidine for 48 hr with or without 5 x 10~4 M MIX (see bar in
Chart 3). The extent of DNA synthesis was determined by autoradiography. Celldensity analyses were performed microscopically by use of a hematocytometer.
MIX responsiveness
Assay duration Labeled nuclei Increase in cell (% of labeled nu-(days) (%) density8 elei)
8-10 27" Cell density on Day 8
x 10'cells/sq cm.13.3 x 10* cells/sq cm and on Day 10 = 12.5
PMA-induced Inhibition of Proadipocyte Differentiation
minally differentiate (see Chart 1). A series of studies wastherefore designed to determine if PMA can block the differentiation of Go-arrested cells. The data in Chart 4 present theresults of a typical experiment. These data show that whenPMA is added to previously untreated GD-arrested cells on Day
8, subsequent differentiation is inhibited. More specifically,Chart 4 shows that if PMA is added to cultures when cells areGDarrested with only 13% differentiated, no further differentiation occurs, and ~5% of the cells actually lose the differentiatedphenotype. By contrast, in untreated proadipocytes, the percentage of differentiation doubles from Day 8 to Day 10. Theseresults show that PMA affects the differentiation of ~18% of
the cells. The data in Table 2 show that this effect is notmediated by causing Go-arrested cells to proliferate. That is,during the 48-hr interval from Days 8 to 10, PMA affects thedifferentiation of ~18% of the cells but stimulates the prolifer
ation of only 4% of the cells. Therefore, PMA can block thedifferentiation of Go-arrested cells under appropriate experi
mental conditions.Relative to the model presented in Chart 1, data on the high-
density differentiation system have been presented, whichshow that PMA can prevent GD arrest and can block differentiation. Next, it was determined whether PMA can promotesignificant loss of differentiated phenotype and mitogenesis.
Can PMA Induce Adipocytes to Lose Their DifferentiatedPhenotype? The data in Chart 4 suggest that PMA can promoteloss of differentiated phenotype. This observation was moreextensively studied in a series of experiments summarized inChart 5. High-density proadipocytes were fed differentiation-
promoting medium, and thereafter, PMA was periodicallyadded to specimens to determine the extent of loss of differentiated phenotype that resulted. If PMA is added to cultureson Day 8, the time when adipocyte differentiation is first evident, 95% of the adipocytes are induced to lose their differentiated phenotype. Induction of significant loss of the differentiated phenotype is also observed if PMA is added to cultureson Day 12 when an intermediate extent of differentiation isevident. However, when the tumor promoter is added to older
responsiveness to MIX in the absence of DNA synthesis and inthe absence of an increase in cell density. This finding establishes that in high-density proadipocytes, PMA can block dif
ferentiation by causing cells to growth arrest at a state in Gìwhich topographically precedes GD. By contrast, in a previousstudy, we established that in monolayer cultures the d arreststates induced by serum deprivation, nutrient deprivation, andcontact inhibition all occurred topographically following the GDarrest state (44). Kinetic studies were therefore also performedto compare the time required for GD-arrested, PMA-treated,Gt-arrested cells to initiate DNA synthesis when placed in
DMEM containing 10% FCS. The results show that cells atboth states initiated DNA synthesis following a lag interval of12 to 15 hr, and both show maximum incorporation into DNAbetween 21 and 24 hr after induction of mitogenesis. Thesedata suggest that although Go-arrested and PMA-treated
proadipocytes are at different states in G,, these states aretopographically similar and cannot be resolved by methodologyusing restimulation kinetic analysis.
Can PMA Block the Differentiation Process? After Go arrest, untreated proadipocytes can nonterminally and then ter-
30-,
a 20-
I52 10 5.5 %
•468
TIME (days)
1O
Chart 4. Inhibition of differentiation of Go-arrested cells by PMA. High-densityproadipocytes were fed differentiation-promoting medium on Days 0, 4, and 8.On Day 8, the specimens were divided into 2 groups. One group (O) wasmaintained untreated, and to the other group (•),PMA (150 ng/ml) was added.In replicate flasks, [3H]thymidine was also added. The percentage of differentiation of both groups was scored by randomly counting —500 cells/specimen.Autoradiographic analysis was performed from Days 8 to 10 (oar) to determinethe extent of DNA synthesis (see Table 2).
JANUARY 1983 91
on March 3, 2020. © 1983 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
K. Yun and R. E. Scott
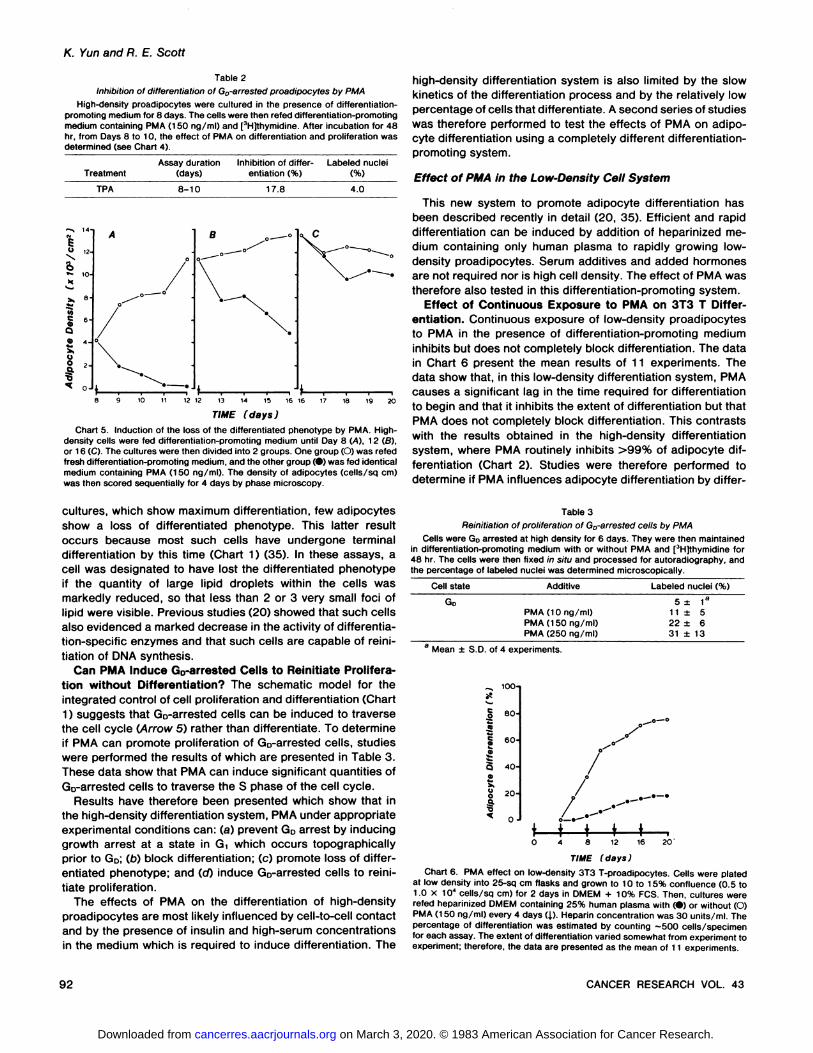
Table 2Inhibition of differentiation of Go-arrested proadipocytes by PMA
High-density proadipocytes were cultured in the presence of differentiation-promoting medium for 8 days. The cells were then refed differentiation-promotingmedium containing PMA (150 ng/ml) and [3H]thymidine. After incubation for 48
hr, from Days 8 to 10, the effect of PMA on differentiation and proliferation wasdetermined (see Chart 4).
TreatmentAssay duration
(days)Inhibition of differ
entiation (%)Labeled nuclei
TPA 8-10 17.8 4.0
I-
v«—
X
OZ H
10
TIME (days)Chart 5. Induction of the loss of the differentiated phenotype by PMA. High-
density cells were fed differentiation-promoting medium until Day 8 (A), 12 (6),or 16 (C). The cultures were then divided into 2 groups. One group (O) was refedfresh differentiation-promoting medium, and the other group (•)was fed identicalmedium containing PMA (150 ng/ml). The density of adipocytes (cells/sq cm)was then scored sequentially for 4 days by phase microscopy.
high-density differentiation system is also limited by the slow
kinetics of the differentiation process and by the relatively lowpercentage of cells that differentiate. A second series of studieswas therefore performed to test the effects of PMA on adipo-cyte differentiation using a completely different differentiation-
promoting system.
Effect of PMA in the Low-Density Cell System
This new system to promote adipocyte differentiation hasbeen described recently in detail (20, 35). Efficient and rapiddifferentiation can be induced by addition of heparinized medium containing only human plasma to rapidly growing low-
density proadipocytes. Serum additives and added hormonesare not required nor is high cell density. The effect of PMA wastherefore also tested in this differentiation-promoting system.
Effect of Continuous Exposure to PMA on 3T3 T Differentiation. Continuous exposure of low-density proadipocytesto PMA in the presence of differentiation-promoting medium
inhibits but does not completely block differentiation. The datain Chart 6 present the mean results of 11 experiments. Thedata show that, in this low-density differentiation system, PMA
causes a significant lag in the time required for differentiationto begin and that it inhibits the extent of differentiation but thatPMA does not completely block differentiation. This contrastswith the results obtained in the high-density differentiation
system, where PMA routinely inhibits >99% of adipocyte differentiation (Chart 2). Studies were therefore performed todetermine if PMA influences adipocyte differentiation by differ-
cultures, which show maximum differentiation, few adipocytesshow a loss of differentiated phenotype. This latter resultoccurs because most such cells have undergone terminaldifferentiation by this time (Chart 1) (35). In these assays, acell was designated to have lost the differentiated phenotypeif the quantity of large lipid droplets within the cells wasmarkedly reduced, so that less than 2 or 3 very small foci oflipid were visible. Previous studies (20) showed that such cellsalso evidenced a marked decrease in the activity of differentiation-specific enzymes and that such cells are capable of reini
tiation of DMA synthesis.Can PMA Induce Go-arrested Cells to Reinitiate Prolifera
tion without Differentiation? The schematic model for theintegrated control of cell proliferation and differentiation (Chart1) suggests that GD-arrested cells can be induced to traversethe cell cycle (Arrow 5) rather than differentiate. To determineif PMA can promote proliferation of Go-arrested cells, studieswere performed the results of which are presented in Table 3.These data show that PMA can induce significant quantities ofGo-arrested cells to traverse the S phase of the cell cycle.
Results have therefore been presented which show that inthe high-density differentiation system, PMA under appropriateexperimental conditions can: (a) prevent GD arrest by inducinggrowth arrest at a state in Gìwhich occurs topographicallyprior to GD; (£>)block differentiation; (c) promote loss of differentiated phenotype; and (d) induce Go-arrested cells to reini
tiate proliferation.The effects of PMA on the differentiation of high-density
proadipocytes are most likely influenced by cell-to-cell contactand by the presence of insulin and high-serum concentrations
in the medium which is required to induce differentiation. The
Table 3Reinitiation of proliferation of Go-arrested cells by PMA
Cells were G0 arrested at high density for 6 days. They were then maintainedin differentiation-promoting medium with or without PMA and [3H]thymidine for
48 hr. The cells were then fixed in situ and processed for autoradiography, andthe percentage of labeled nuclei was determined microscopically.
Cell state Additive Labeled nuclei (%)
PMA (10 ng/ml)PMA (150 ng/ml)PMA (250 ng/ml)
5± 1a
11 ± 522 ± 631 ±13
Mean ±S.D. of 4 experiments.
_, 10OJ*<^,
§ 80
I§ 60-
5 40-
à 20-I
\ t i t l ,04 8 12 16 20'
TIME (days)Chart 6. PMA effect on low-density 3T3 T-proadipocytes. Cells were plated
at low density into 25-sq cm flasks and grown to 10 to 15% confluence (0.5 to1.0 x 104 cells/sq cm) for 2 days in DMEM + 10% FCS. Then, cultures were
refed heparinized DMEM containing 25% human plasma with (•)or without (O)PMA (150 ng/ml) every 4 days (J.). Heparin concentration was 30 units/ml. Thepercentage of differentiation was estimated by counting ~500 cells/specimenfor each assay. The extent of differentiation varied somewhat from experiment toexperiment; therefore, the data are presented as the mean of 11 experiments.
92 CANCER RESEARCH VOL. 43
on March 3, 2020. © 1983 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

PMA-induced Inhibition of Proadipocyte Differentiation
ent mechanisms in high and low cell density differentiationsystems.
Does PMA Block GD Arrest? In the high-density differentiation system, PMA blocks GD arrest by inducing growth arrestof cells at a point in Gt prior to GD-This was found not to be theresult in the low-density differentiation system. If proadipocytesare cultured in heparinized DMEM containing 25% humanplasma and PMA for 8 days, growth arrest in G, occurs asevidenced by markedly decreased [3H]thymidine incorporation
into DMA and by the finding that 93% of such cells are growtharrested in Gì,1% in S, and 6% in G2-M, as determined by
flow microfluorimetric analysis. The data further show thatthese G,-arrested cells show mitogenic responsiveness to MIXand are therefore at GD. Table 4 presents data illustrating thatlow-density G, growth-arrested proadipocytes, induced by culture in differentiation-promoting medium containing PMA for 4,7, 10, or 12 days, all show significant [3H]thymidine incorpo
ration into DNA when treated with MIX (Table 4, Condition A).A comparable result was obtained if proadipocytes werewashed free of PMA after 10 days and refed heparinizedDMEM containing human plasma and MIX (Table 4, ConditionB).
These data establish that, in the low-density differentiation
system, PMA does not induce growth arrest in G, at a stateprior to GD; rather, it allows Gìgrowth arrest to occur at the GDstate. This fact could account for the observation that in thelow-density differentiation system, PMA does not block differ
entiation; it only delays and partially inhibits the differentiationprocess (Chart 6). The inhibition of differentiation in low-density
proadipocytes by PMA therefore does not result from prevention of Go arrest. It could, however, result from a decreasedrate of differentiation of Go-arrested cells, from an increased
proclivity of adipocytes to lose the differentiated phenotype inthe presence of PMA or as a result of PMA-induced mitogen-
esis.Is PMA a Mitogen for Go-arrested Cells? The inhibition of
differentiation induced by PMA in low-density cells illustrated
in Chart 6 could result at least in part from the continuedproliferation of cells. This possibility is supported by the datapresented in Table 5. It shows that low-density cells which
have been GD arrested for various intervals in heparinizedDMEM containing human plasma are highly sensitive to the
Table 4Mix-induced mitogenesis of low-density proadipocytes G, growth arrested in
the presence of PMA and heparinized medium containing human plasmaLow-density proadipocytes were cultured in heparinized DMEM containing
25% human plasma and PMA for the indicated intervals. These cells, which weregrowth arrested in the G phase of the cell cycle, were then tested to determineif they could be induced to incorporate [3H]thymidine into DNA when exposed to5 X 10 ' M MIX. In Condition A, MIX was added directly to the differentiation-
promoting medium containing PMA. In Condition B, the cell monolayer waswashed in DMEM and then refed heparinized DMEM containing 25% humanplasma and MIX. The extent of DNA synthesis was determined during a 48-hrinterval. Thereafter, it was assayed autoradiographically. The data are expressedas the percentage of labeled nuclei in MIX-treated minus untreated cells.
Table 5Mitogenic effect of PMA on low-density Go-arrested cells
Duration of culture(days)
MIX responsiveness(% of labeled nuclei)
Condition A
Condition B
47
1012
10
24432731
21
CellstateGD
arrestedDuration
ofGoarrest(days)468[PMA](ng/ml)10100250150150Mitogenic
response8(%
oflabelednuclei)1353704431
a Determined autoradiographically following culture of Go-arrested cells inPMA and [3H]thymidine for 48 hr.
mitogenic effect of PMA. That is, up to 70% of low-density Go-
arrested cells can be induced to proliferate by PMA. To corroborate this finding, we also established that there is a markeddifference in the cell density of proadipocytes cultured indifferentiation-promoting medium with or without PMA. In cultures in which the initial cell density was 1.4 x 103 cells/sqcm the density increased to 3 x 103 cells/sq cm following
culture in differentiation-promoting medium for 20 days,whereas in cultures containing PMA, the final density was 7-fold higher, i.e., 2.1 x 104 cells/sq cm. The pronounced
mitogenic effect of PMA for low-density proadipocytes there
fore appears to serve a major mechanism for delaying andinhibiting differentiation in this system (Chart 6).
Can PMA Promote Cells to Lose the Differentiated Phenotype? Another mechanism by which PMA could preventdifferentiation of low-density cells is to cause any cells whichdo differentiate following GD arrest to lose the adipocyte phenotype. The data in Chart 7 show that induction of loss of thedifferentiated phenotype can indeed account in part for PMA-
induced differentiation inhibition. These results show that ifPMA is added to low-density cells which have been cultured indifferentiation-promoting medium for 4 (Chart 7A) or 8 (Chart78) days, between 35 and 50% of the cells lose the differentiated phenotype (approximately 50% of the cells which losethe differentiated phenotype progress through the S phase ofthe cell cycle in the presence of PMA as determined by auto-radiographic analysis of [3H]thymidine incorporation into PMA).
At later time intervals (Chart 7C), the ability of PMA to induceloss of differentiated phenotype is decreased, because a significant proportion of the adipocytes have undergone terminaldifferentiation prior to addition of PMA (20, 35). The inhibitionof differentiation in low-density proadipocytes by PMA cantherefore also be induced in part by induction of loss of differentiated phenotype.
Does PMA Block Nonterminal and/or Terminal Differentiation? We first attempted to determine if PMA blocks nonterminal differentiation. Since PMA in this system is both a potentmitogen and a promoter of loss of differentiated phenotype, itwas difficult to unequivocally establish whether PMA-induced
inhibition of differentiation can be mediated solely by inhibitionof nonterminal differentiation. The data in Chart 8, however,suggest that nonterminal differentiation is not significantlyblocked in low-density cells by PMA. If PMA is added tocultures throughout the entire differentiation interval, it causesonly a 2-day lag in differentiation, and the subsequent kineticsof differentiation is similar in untreated and treated cells. Thislag phase is caused by the pronounced mitogenic effect ofPMA for low-density cells. The fact that the total cell density
JANUARY 1983 93
on March 3, 2020. © 1983 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

K. Yun and R. E. Scott
increases during the first 4 days of the experiment also accounts for the observation that, in Chart 8A, the adipocytedensity in the PMA-treated cells exceeds that in control spec
imens. If PMA is added after cells have been cultured indifferentiation-promoting medium for 8 days and are at the Go
state, the results also show a lag before differentiation, butthereafter, differentiation proceeds in a manner similar to thatobserved in untreated cells. These data suggest that PMA doesnot block differentiation.
Experiments were next performed to determine if PMA selectively prevents terminal differentiation. Since adipocyte differentiation does occur to a significant extent in low-densitycells cultured in differentiation-promoting medium containing
PMA (Chart 8), it was possible to assay if the differentiatedcells were in a nonterminal or terminal state. We reportedpreviously that nonterminally differentiated adipocytes can beinduced to lose the differentiated phenotype and proliferate ifthey are refed in DMEM containing 30% serum, whereasterminally differentiated cells cannot (33, 35). We thereforecompared the relative ability of PMA-treated and untreated
differentiated cells to lose their differentiated phenotype. Indata not shown, no differences were detected. That is, afterapproximately 12 to 14 days in the differentiated state, neithertreated nor untreated adipocytes could be induced to lose theirdifferentiated phenotype. Therefore, PMA does not appear toblock the process of terminal differentiation.
DISCUSSION
PMA has two very important biological effects on mammaliancells. It acts as a tumor promoter in vitro (19, 23) and in vivo(4-6), and it modulates the ability of cells to differentiate (8,
10, 18, 26, 30, 38, 39, 41, 45, 46). Based on these observations, the hypothesis has been presented that the effect ofPMA on cell differentiation may account at least in part for itstumor-promoting activity (2). However, since PMA has numerous metabolic effects on cells (3, 11-13, 15, 17, 19, 21, 22,27-29, 31, 37, 40, 43), it has not been possible to establishwhich, if any, actions mediate both tumor promotion and dif-
TIME (days)Chart 7. Induction of loss of the differentiated phenotype by PMA in the low-
density differentiation system. Low-density cells were cultured in heparinizedDMEM containing human plasma for 4, 8, or 12 days and refed on these days.PMA (150 ng/ml) was also added to some specimens when they were refed. Thedensity of adipocyte/sq cm was then determined sequentially by phase microscopy for untreated cells (O) and PMA-treated cells (•).Effect of PMA whenadded on Day 4 (A) Day 8 (8), and Day 12 (C).
ot—
X
14
Q>Q
oot
A-
3-
2-
1 -
•
o—•\—*—*—*4-I
3-
2-
1-
O-l
B
0ÕA8
TIME12
16(days)20
Chart 8. Effect of PMA on the differentiation of low-density proadipocytes.Low-density cells were fed heparinized DMEM containing human plasma with orwithout added PMA (150 ng/ml) on Days 0. 4, 8, 12, and 16 (i) (A) or on Days8, 12. and 16 (i) (B). The effects of these treatments on adipocyte differentiationwere then scored morphologically. A comparison of adipocyte density observedin untreated cells (O) and in cells which had been exposed to PMA continuously(•)is presented in A. A similar comparison is presented in B for untreated cells(O) and cells treated with PMA only from Days 8 through 20 (•).
ferentiation modulation. To accomplish this goal, it is firstnecessary to establish the mechanisms by which phorbol estersaffect specific aspects of both tumor promotion and differentiation.
Our studies have focused on the effects of PMA on adipocytedifferentiation. We reported recently that there are at least 5biological processes involved in the integrated control of cellproliferation and differentiation (33, 35). Three steps are involved in the differentiation process: Go arrest; nonterminaldifferentiation; and terminal differentiation. Two steps antagonize the differentiation process and facilitate cell growth: lossof differentiated phenotype and reinitiation of proliferation (seeChart 1). In the current studies, the effect of PMA on theintegrated control of proadipocyte proliferation and differentiation has therefore been studied in 2 different culture systems.One system uses low-density proadipocytes which are induced
to differentiate by heparinized DMEM containing human plasma
94 CANCER RESEARCH VOL. 43
on March 3, 2020. © 1983 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

PMA-induced Inhibition of Proadipocyte Differentiation
(20), and the second system uses high-density proadipocytes
which are induced to differentiate by DMEM containing highconcentrations of PCS and insulin (10, 34).
PMA is shown to inhibit or block differentiation in bothsystems; however, the data show that different mechanismscause these effects.
In high-density proadipocytes, PMA can prevent GD arrestby causing cells to growth arrest at a G, state topographicallyprior to GD; PMA can inhibit nonterminal differentiation; PMAcan cause loss of the differentiated phenotype; and PMA canreinitiate proliferation of Go-arrested cells.
In low-density cells, PMA does not affect the ability of proad
ipocytes to GD arrest nor their ability to nonterminally or terminally differentiate. Rather, in low-density proadipocytes,
PMA inhibits differentiation because it induces loss of differentiated phenotype and because it is a potent mitogen.
These data show that control of cellular differentiation is acomplex process which can be modulated by numerous factorsincluding cell density, cell-to-cell contact, and the type of
culture medium used. Although it is not possible to establishwhich of these factors is most important, the results show thatPMA can have significant effects on 4 processes involved inthe integrated control of proadipocyte proliferation and differentiation. Since the ability of PMA to modulate differentiationcan be influenced by a variety of microenvironmental factors,we conclude that it is not possible to establish which, if any, ofthe effects of PMA on cell proliferation and differentiationmediate its role as a promoter of carcinogenesis. In view of thefact that we have shown that PMA does not promote tumori-genesis of 3T3 T-proadipocytes (32), the possibility must even
be considered whether any effects of PMA on cell proliferationand differentiation are involved in promotion of carcinogenesis.
Chemical carcinogenesis is a 2-phase process involvinginitiation and promotion processes (2-6), and it has been
shown that promoting agents, such as PMA, only induce thetransformation of cells which have been initiated (2-6). We
therefore suggest that, in order to determine the mechanismsof tumor promotion, specific effects of PMA that act only oninitiated cells must be identified.
ACKNOWLEDGMENTS
We thank John J. Wille, Jr., Ph.D., and Marjorie Wier for their critical discussion; Peter B. Maercklein, Bryan J. Hoerl, and Michael A. Zschunke for technicalassistance; and Patricia Hart for secretarial assistance.
REFERENCES
1. Balmain, A., and Hecker, E. On the biochemical mechanism of tumorigenesisin mouse skin. IV. Early effects of growth-stimulated phorbol esters onphosphate transport and phospholipid synthesis in mouse epidermis.Biochim. Biophys. Acta, 362. 457-468, 1974.
2. Becker, F. F. Recent concepts of initiation and promotion in carcinogenesis.Am. J. Pathol.. ÃŽ05.3-9, 1981.
3. Belman, S., Troll, W., and Garte, S. J. Phorbol-12-myristate-13-acetateeffect on cyclic adenosine 3',5'-monophosphate levels in mouse skin andinhibition of phorbol-myristate-acetate-promoted tumorigenesis by the-ophylline. Cancer Res., 34: 3445-3446, 1974.
4. Berenblum, I. Sequential aspects of chemical carcinogenesis: skin. In: F. F.Becker (ed.), Cancer, a Comprehensive Treatise, Vol. 1. pp. 323-344. NewYork: Pleum Publishing Corp., 1975.
5. Boutwell, R. K. Some biological aspects of skin carcinogenesis. Prog. Exp.Tumor Res., 4: 207-250, 1964.
6. Cairns, J. The origin of human cancer. Nature (Lond.), 289. 353-357, 1981.7. Castagna, M., Rochette-Egly, C., Rosenfeld, C., and Mishal, Z. Altered lipid
microviscosity in lymphoblastoid cells treated with 12-O-tetradecanoylphor-bol-13-acetate, a tumor promoter. FEBS Lett., 100: 62, 1979.
8. Cohen, R., Pacifici, M., Rubinstein, N., Biehl, J., and Holtzer, H. Effect of atumor promoter on myogenesis. Nature (Lond.). 266: 538-540, 1977.
9. Diamond, L., O'Brien, T. G., and Baird, W. M. Tumor promoters and the
mechanism of tumor promotion. Adv. Cancer Res., 32. 1-74, 1980.10. Diamond. L., O'Brien, T. G., and Rovera, G. Inhibition of adipose conversion
of 3T3 fibroblasts by tumor promoters. Nature (Lond.), 269: 247-249,1977.
11. Driedger, P. E., and Blumberg, P. M. Non-phorbol mouse skin tumor promoters do not mimic phorbol myristate acetate in its effects on chickfibroblast. Int. J. Cancer, 22: 63-69, 1978.
12. Driedger, P. E., and Blumberg, P. M. Specific binding of phorbol ester tumorpromoters. Proc. Nati. Acad. Sei. U. S. A., 77: 567-571, 1980.
13. Fisher, P. B., Flamm, M., Schachter, D., and Weinstein, l. B. Tumor promoters induce membrane changes detected by fluorescence polarization.Biochem. Biophys. Res. Commun., 86. 1063-1068, 1979.
14. Green, H., and Kehinde, O. An established preadipose cell line and itsdifferentiation in culture. II. Factors affecting the adipose conversion. Cell,5: 19-27. 1975.
15. Hennings, H., and Boutwell, R. K. Studies on the mechanism of skin tumorpromotion. Cancer Res., 30: 312-320, 1970.
16. Huberman, E., and Callahan, M. F. Induction of terminal differentiation inhuman promyelocytic leukemia cells by tumor-promoting agents. Proc. Nati.Acad. Sei. U. S. A., 76: 1293-1297, 1979.
17. Jane, J., Poso, H., and Raina, A. Polyamines in rapid growth and cancer.Biochim. Biophys. Acta, 473. 241-243, 1978.
18. Koeffler, H. P., Bar-Eli, M., and Territo, M. C. Phorbol ester effect ondifferentiation of human myeloid leukemia cell line, blocked at differentstages of maturation. Cancer Res., 41: 919-926. 1981.
19. Kopelovich, L., Bias, N. E., and Helson, L. Tumor promoter alone inducesneoplastic transformation of fibroblasts from humans genetically predisposed to cancer. Nature (Lond.), 282. 619-621, 1979.
20. Krawisz, B. R., and Scott, R. E. Coupling of proadipocyte growth arrest anddifferentiation. I. Induction by heparinized medium containing human plasma.J. Cell Biol., 94: 394-399, 1982.
21. Levine, L., and Hassid, A. Effects of phorbol-12,13-diesters on prostaglandinproduction and phospholipase activity in canine kidney (MDCK) cells. Biochem. Biophys. Res. Commun., 79: 477-484, 1977.
22. Lichti, U., Patterson, E., Hennings, H., and Yuspa, S. H. Differential retinoicacid inhibition of ornithine decarboxylase induction by 12-O-tetradecanoyl-phorbol-13-acetate and by germicidal ultraviolet light. Cancer Res., 41:49-54, 1981.
23. Mondai, S., and Heidelberger, C. Transformation of C3H/10TVÃŒCL8 mouseembryo fibroblasts by ultraviolet irradiation and/or phorbol ester. Nature(Lond.). 260: 710-711. 1976.
24. Moses, H. L., Proper, J. A., and Volkenant, M. E. Growth arrest of AKR-2Bcells maintained in the presence of epidermal growth factor and! 2-O-tetra-decanoylphorbol-13-acetate: evidence for two separate G, arrest points. J.Cell Physiol., Õ02:367-378, 1980.
25. Nadal-Girard, B. Commitment, fusion, and biochemical differentiation of amyogenic cell line in the absence of DNA synthesis. Cell, Õ5: 855-864,
1978.26. Nagasawa, K., and Mak, T. W. Phorbol esters induce differentiation in
human malignant T lymphoblasts. Proc. Nati. Acad. Sei. U. S. A., 77:2964-2967, 1980.
27. O'Brien, T. G. The induction of ornithine decarboxylase as an early possible
obligatory event in mouse skin carcinogenesis. Cancer Res., 36:2644-2653, 1976.
28. O'Brien, T. G., and Saladik, D. Inhibition of the adipose conversion of BALB/
e 3T3 cells by 12-O-tetradecanoylphorbol-13-acetate: dependence on pHreduction via lactic acid production. J. Cell Physiol., 704: 35-40, 1980.
29. Ojakiam, G. K. Tumor promoter-induced changes in the permeability ofepithelial cell tight junction. Cell, 23:95-103, 1981.
30. Rifkind, R. A., Fibach, E., Reuben, R. C., Gaziti, Y., Yamasaki, H.. Weinstein,l. B., Nudel, U., Sumida, I., Terada, M., and Marks, P. A. Erythroleukemiacells: commitment to differentiate and the role of the cell surface. In: B.Clarkson, P. A. Marks, and J. E. Tills (eds.), The Differentiation of Normaland Neoplastic Hematopoietic Cells, pp. 209-220. New York: Cold SpringHarbor Press, 1978.
31. Rovera, G., Santoli, D., and Damsky, C. Human promyelocytic cells in culturedifferentiate into macrophage-like cells when treated with a phorbol ester.Proc. Nati. Acad. Sei. U. S. A., 76: 2779-2783, 1979.
32. Scott, R. E., and Boone. C. W. Promotion of smooth surface tumorigenicityby phorbol myristate acetate in BALB/3T3 cells and BALB/3T3 T proadipocytes. J. Nati. Cancer Inst., 66: 733-736, 1981.
33. Scott, R. E., and Florine, D. L. Cell cycle models for the aberrant control ofgrowth arrest and differentiation in hyperplasia, metaplasia, and neoplasia.Am. J. Pathol., »07:342-348, 1980.
34. Scott, R. E., Florine, D. L., Wille, J. J., Jr., Yun, K., and Hoerl, B. J. Couplingof growth arrest and differentiation at a distinct state in the G. phase of thecell cycle: GD. Proc. Nati. Acad. Sei. U. S. A., 79. 845-849, 1982.
35. Scott, R. E., Hoerl, B. J., Wille, J. J., Jr., Florine, D. L., Krawisz, B. R., andYun, K. Coupling of proadipocyte growth arrest and differentiation. II. A cellcycle model for the physiological control of cell proliferation. J. Cell Biol.,
JANUARY 1983 95
on March 3, 2020. © 1983 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

K. Yun and R. E. Scott
94:400-405, 1980.36. Scott, R. E., Yun, K., and Florine, D. L. Differential mitogenic effect of methyl
isobutyl xanthine and a tumor growth factor on Gìarrested 3T3 T proadi-pocytes at the predifferentiation GD state and the growth factor deficiencyGs state. Exp. Cell Res., in press, 1982.
37. Shoyab, M., and Todaro, G. J. Specific high affinity cell membrane receptorsfor biologically active phorbol and ingenol esters. Nature (Lond.), 288.451-455, 1980.
38. Sisskin, E. E., and Barrett, J. C. Inhibition of terminal differentiation ofhamster epidermal cells in culture by the phorbol ester, 12-O-tetradeca-noylphorbol-13-acetate. Cancer Res., 41: 593-603, 1981.
39. Steele, V. E., Marchok, A. C., and Nettesheim, P. Establishment of epithelialcell lines following exposure of cultured trachéalepithelium to 12-O-tetra-decanoylphorbol-13-acetate. Cancer Res., 38. 3563-3565, 1978.
40. Tornei, L. D., Chenny. J. C., and Wenner, C. E. The effect of phorbol esterson the proliferation of C3H-10TVÃŒmouse fibroblast: consideration of bothstimulatory and inhibitory effects. J. Cell Physiol., 707: 385-389, 1981.
41. Toyama, Y., West, C. M., and Holtzer, H. Differential response of myofibrils
and 10 nm filaments to a carcinogen. Am. J. Anat., 756: 131-139, 1979.42. Vaheri, A., Ruoslahti, E., and Mosher, D. F. (eds.). Fibroblast surface
proteins. Ann. N. Y. Acad. Sci., 312: 1-456, 1978.43. Verma, A. K.. Ashendel, C. L.. and Boutwell, R. K. Inhibition by prostaglandin
synthesis inhibitors of the induction of epidermal ornithine decarboxylaseactivity, the accumulation of prostaglandins, and tumor promotion causedby 12-O-tetradecanoylphorbol-13-acetate. Cancer Res., 40: 308-315,1980.
44. Wille. J. J., Jr., and Scott, R. E. Topography of predifferentiation GD growtharrest states in the G, phase of the cell cycle. J. Cell. Physiol., 112:115-122, 1982.
45. Yamasaki, H., Fibach, C., Nudel. U., Weinstein, l. B., and Marks, P. Tumorpromoters inhibit spontaneous and induced differentiation of murine eryth-roleukemia cells in culture. Proc. Nati. Acad. Sei. U. S. A., 74: 3451 -3455,1977.
46. Yamasaki, H., Saint-Vincent, L.. and Martel, N. Long-term effect of a tumorpromoter, 12-O-tetradecanoylphorbol-13-acetate, on induced differentiationof Friend leukemia cells. Cancer Res., 40: 3780-3785, 1980.
CANCER RESEARCH VOL. 43
on March 3, 2020. © 1983 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

1983;43:88-96. Cancer Res Kankatsu Yun and Robert E. Scott Inhibition of Proadipocyte DifferentiationBiological Mechanisms of Phorbol Myristate Acetate-induced
Updated version
http://cancerres.aacrjournals.org/content/43/1/88
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/43/1/88To request permission to re-use all or part of this article, use this link
on March 3, 2020. © 1983 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from