biologie notion de rÉflexe -...
TRANSCRIPT

Bio
logi
eNOTION DE RÉFLEXE
Juin 2010
CUEEP - UNIVERSITÉ LILLE 1 SCIENCES ET TECHNOLOGIES
DÉPARTEMENT SCIENCES
JACQUES COGET


Table des matières
Table des matières 3
I - Introduction 5
II - Les éléments anatomiques du réflexe 7
A. Les récepteurs.................................................................................7
B. Les centres......................................................................................8
C. Les afférences et les efférences..........................................................9
D. Les effecteurs................................................................................10
III - Étude expérimentale 11
IV - Le réflexe myotatique 15
V - Exercices d'application 19
A. Exercice d'application N°1...............................................................19
B. Exercice d'application N°2...............................................................20
C. Exercice d'application N°3...............................................................20
D. Exercice d'application N°4...............................................................21
Solution des exercices de TD 23
CUEEP-Université Lille 1 Sciences et Technologies3


I - Introduction I
C'est au cours des XVIIe et XVIIIe siècles que le terme réflexe, emprunté au langage des physiciens et au latin reflexus (du verbe reflectere = ramener en arrière), s'est peu à peu imposé en physiologie nerveuse pour décrire les mouvements animaux initiés par des stimulations sensorielles.
Il faudra toutefois attendre les travaux du neurophysiologiste anglais Sir Charles Scott Sherrington (1857-1952), prix Nobel de physiologie et de médecine en 1932 avec son compatriote Edgar Adrian (1899-1977) pour l'ensemble de leurs travaux sur la physiologie du système nerveux, pour que les principaux mécanismes qui régissent le réflexe soient élucidés, que le rôle de chaque élément soit précisé et qu'on en arrive à la notion d'arc réflexe pouvant être schématisé de la manière suivante.
Comme on le voit, le réflexe présente cinq composantes anatomiques qui sont indispensables à son accomplissement :
un récepteur qui détecte la stimulation sensorielle, physique ou chimique, et qui la convertit en potentiels d'action ;une afférence qui conduit le message sensoriel produit par le récepteur au centre nerveux chargé de l'analyser ; un centre qui met en relation afférence et efférence et qui élabore une réponse appropriée à la stimulation ;une efférence qui véhicule le nouveau message jusqu'à l'effecteur chargé de produire la réponse souhaitée ; un effecteur qui, comme son nom l'indique, effectue la réponse et qui, selon les cas, peut être un muscle ou une glande.
La réalité est toutefois plus complexe dans la mesure où plus la stimulation est importante, plus elle met en jeu de récepteurs, ce qui a pour conséquence de mobiliser de nombreuses afférences et de nombreuses efférences et, au final, plusieurs effecteurs. Pensez, par exemple, au réflexe de toux qui permet l'expulsion d'un corps étranger des voies respiratoires ou au réflexe de retrait après avoir posé
CUEEP-Université Lille 1 Sciences et Technologies5

une main sur un objet brûlant. Quoi qu'il en soit, le réflexe présente toujours quatre caractéristiques :
il est adapté car il permet à l'organisme de produire une réponse physiologique en rapport avec la stimulation ;il est involontaire car il se déroule sans intervention consciente du sujet et sans contrôle possible de sa part ;il est prévisible car il se répète à l'identique après chaque stimulation du même ordre et de même intensité ;il est inné car il ne nécessite aucun apprentissage et apparaît spontanément une fois le système nerveux mature.
Cette dernière caractéristique est toutefois absente d'un type particulier de réflexe, dénommé réflexe conditionné, qui nécessite un apprentissage et qui fut découvert en 1902 par le russe Ivan Petrovitch Pavlov (1849-1936), prix Nobel de physiologie et de médecine en 1904 pour ses travaux de physiologie cardiovasculaire et digestive. Ayant observé que la présentation d'un repas à un chien suffit pour déclencher une sécrétion salivaire, il associe la présentation du mets à un signal sonore et constate qu'au bout d'un certain temps, le signal sonore seul provoque la salivation du chien. Le réflexe a donc été appris. Par contre, une fois ce type de comportement acquis, si le signal sonore n'est plus jamais suivi d'une présentation de nourriture, le réflexe conditionné finit par disparaître ce qui ne s'observe jamais pour les réflexes classiques qui seuls seront étudiés ici.
CUEEP-Université Lille 1 Sciences et Technologies6
Introduction

II - Les élémentsanatomiques duréflexe
II
Les récepteurs 7
Les centres 8
Les afférences et les efférences 9
Les effecteurs 10
A. Les récepteurs
D'une manière générale, les récepteurs qui participent aux réflexes sont ceux de la sensibilité générale ou somesthésie – par opposition à ceux de la sensibilité spéciale qui sont ici peu sollicités et qui regroupent les récepteurs de l'audition, de la gustation, de l'olfaction et de la vision.
Depuis les travaux de Sherrington, l'habitude est prise de les classer en trois catégories correspondant à trois territoires anatomiques distincts.
Les extérocepteurs assurent la sensibilité superficielle de l'organisme au tact, au chaud, au froid et à la douleur. Situés dans la peau et dans les tissus sous-cutanés, ils se présentent sous forme de terminaisons nerveuses qui peuvent être nues, associées à des cellules particulières ou encapsulées dans des petits corpuscules.Les propriocepteurs assurent la sensibilité profonde de l'organisme à sa position, à ses déplacements et à ses mouvements. Localisés dans les muscles, les tendons et les articulations, ils se présentent également sous forme de terminaisons nerveuses qui peuvent être nues, intégrées dans des structures spécifiques aux muscles (fuseaux neuromusculaires) ou encapsulées dans des petits corpuscules. On y ajoute généralement les récepteurs labyrinthiques de l'oreille interne qui se présentent sous forme de cellules ciliées et qui sont responsables de l'équilibration.Les intérocepteurs assurent la sensibilité viscérale de l'organisme à différents stimuli physiques et chimiques. Moins connus que les précédents, ils sont disposés dans les parois des viscères et des vaisseaux sanguins et impliqués dans la régulation des grandes fonctions végétatives telles que la digestion, la circulation ou la respiration.
CUEEP-Université Lille 1 Sciences et Technologies7

Toutefois, cette classification des récepteurs basée sur la répartition anatomique n'apportant aucune indication sur leur fonction, on y ajoute un certain nombre de catégories transversales de manière à préciser leur spécificité. On parle ainsi, pour les plus répandus :
de barorécepteurs sensibles à la pression,de chémorécepteurs sensibles à des substances chimiques,de mécanorécepteurs sensibles à la déformation,de nocicepteurs sensibles à la douleur,d'osmorécepteurs sensibles à la pression osmotique,de photorécepteurs sensibles à la lumière,de thermorécepteurs sensibles au chaud ou au froid.
Enfin, précisons que si leur fonctionnement interne varie avec le mode d'excitation, tous répondent à la stimulation par une modification de leur potentiel de membrane, ce qui a pour effet de convertir le stimulus en signal électrique et de le coder sous forme de potentiels d'action.
B. Les centres
Lieu de mise en relation des afférences et des efférences, les centres réflexes sont aussi le lieu où s'élabore la réponse physiologique à la stimulation.
Bien qu'il en existe en différents points du névraxe (les réflexes faisant intervenir les récepteurs de la sensibilité spéciale, comme l'accommodation visuelle, et de nombreux réflexes viscéraux, comme la déglutition, sont par exemple traités au niveau du tronc cérébral), la moelle épinière apparaît comme le principal centre réflexe de l'organisme.
Une coupe transversale révèle une parfaite symétrie droite et gauche ainsi qu'une dualité de structure entre :
la substance blanche périphérique organisée en trois cordons,la substance grise centrale formée de trois cornes.
C'est dans cette dernière que sont localisés les corps cellulaires :
des motoneurones à l'origine des efférences,des interneurones reliant afférences et efférences,des neurones d'association qui permettent aux différents étages médullaires d'être reliés entre eux.
CUEEP-Université Lille 1 Sciences et Technologies8
Les éléments anatomiques du réflexe
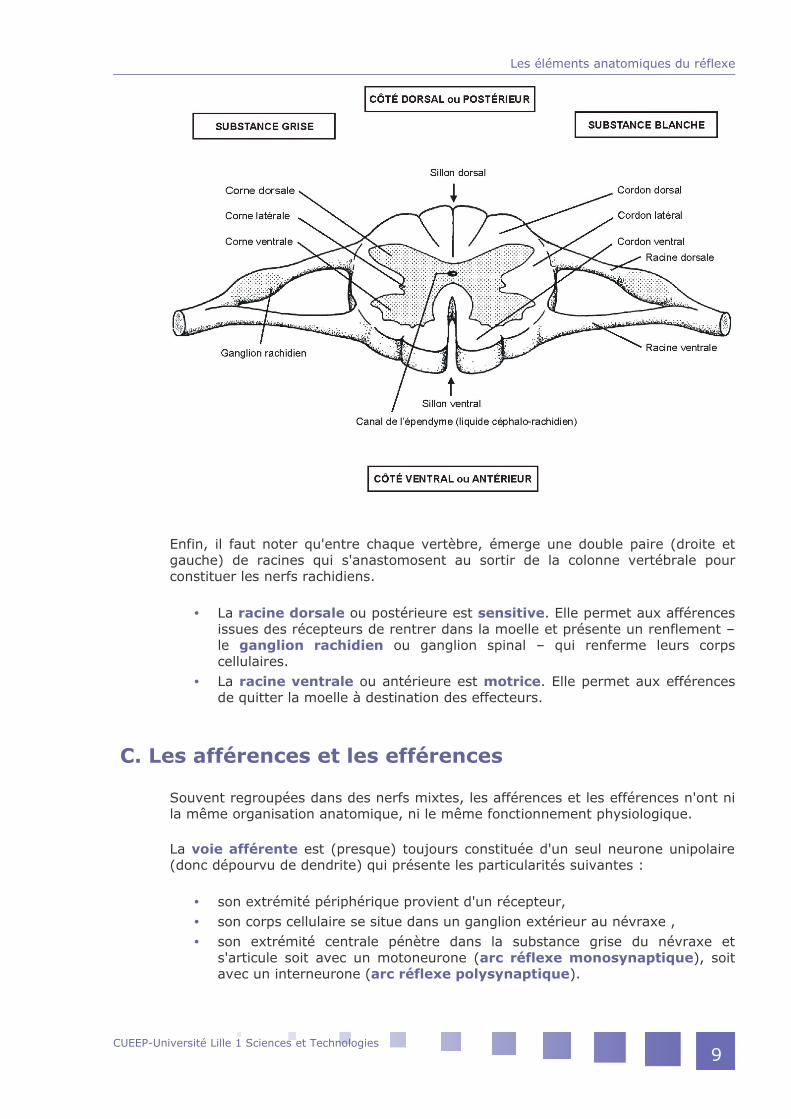
Enfin, il faut noter qu'entre chaque vertèbre, émerge une double paire (droite et gauche) de racines qui s'anastomosent au sortir de la colonne vertébrale pour constituer les nerfs rachidiens.
La racine dorsale ou postérieure est sensitive. Elle permet aux afférences issues des récepteurs de rentrer dans la moelle et présente un renflement – le ganglion rachidien ou ganglion spinal – qui renferme leurs corps cellulaires.La racine ventrale ou antérieure est motrice. Elle permet aux efférences de quitter la moelle à destination des effecteurs.
C. Les afférences et les efférences
Souvent regroupées dans des nerfs mixtes, les afférences et les efférences n'ont ni la même organisation anatomique, ni le même fonctionnement physiologique.
La voie afférente est (presque) toujours constituée d'un seul neurone unipolaire (donc dépourvu de dendrite) qui présente les particularités suivantes :
son extrémité périphérique provient d'un récepteur,son corps cellulaire se situe dans un ganglion extérieur au névraxe ,son extrémité centrale pénètre dans la substance grise du névraxe et s'articule soit avec un motoneurone (arc réflexe monosynaptique), soit avec un interneurone (arc réflexe polysynaptique).
Les éléments anatomiques du réflexe
CUEEP-Université Lille 1 Sciences et Technologies9

La voie efférente renferme quant à elle un ou deux neurones selon qu'elle appartient :
au système nerveux central (efférences à destination des muscles striés squelettiques),au système nerveux végétatif (efférences à destination des muscles viscéraux et des cellules glandulaires).
Dans le premier cas – le seul qui sera envisagé ici –, il s'agit toujours d'un neurone multipolaire à l'axone myélinisé.
D. Les effecteurs
Destination finale du réflexe, les effecteurs ont en charge la réalisation concrète de la réponse à la stimulation.
Comme on vient de le voir, il en existe trois catégories.
Les muscles striés squelettiques (muscles rouges) permettent les mouvements des segments osseux. Ils sont tous attachés au squelette par l'intermédiaire des tendons, à l'exception des orbiculaires et des sphincters qui ont une forme en anneau.Les muscles lisses (muscles blancs) permettent les mouvements internes des viscères et la modification du calibre des vaisseaux sanguins en réponse aux besoins de l'organisme. Tous sont localisés dans la paroi des organes.Certaines glandes exocrines et endocrines peuvent également être impliquées dans des activités réflexes. Elles sont alors innervées par le système nerveux végétatif et libèrent leur contenu après réception d'un influx moteur qualifié de viscéro-sécrétoire.
CUEEP-Université Lille 1 Sciences et Technologies10
Les éléments anatomiques du réflexe

III - Étudeexpérimentale
III
Une grenouille spinale (dont l'encéphale a été détruit mais la moelle épinière laissée intacte) est suspendue par la mâchoire inférieure à une potence. On porte à l'extrémité du pied droit une stimulation électrique de plus en plus intense et on observe les réactions de l'animal.
Tant que la stimulation est infraliminaire, aucune réponse n'est observée. Par contre, dès que la stimulation atteint un certain seuil, on constate une légère flexion du pied droit, à l'endroit même où a été portée la stimulation. C'est la loi de la localisation.
Puis, au fur et à mesure que la stimulation augmente, on note :
le fléchissement complet de la patte excitée tendant à soustraire l'animal à la stimulation douloureuse (loi d'unilatéralité) ;le fléchissement des deux pattes postérieures montrant que le réflexe de retrait s'est étendu au membre contralétéral (loi de symétrie) ;l'extension du mouvement au quatre membres indiquant qu'un nombre
CUEEP-Université Lille 1 Sciences et Technologies11

croissant de circuits neuronaux a été activé (loi d'irradiation) ;enfin des contorsions du corps entier de l'animal, toujours dans le but de se soustraire à la stimulation de plus en plus intense (loi de généralisation).
Cet ensemble de lois, désormais connu sous le nom de lois de Pflüger – du nom du neurophysiologiste allemand Eduard Pflüger (1829-1910) qui, le premier, les a observées en trempant l'extrémité d'une patte postérieure de grenouille spinale dans des solutions d'acide de plus en plus concentrées –, montre que la complexité de la réponse augmente avec l'intensité de la stimulation.
Le réflexe est ici à point de départ cutané. Lorsque la stimulation est supraliminaire, les récepteurs situés dans la peau de l'animal sont excités et élaborent un message sensoriel dont les caractéristiques (nombre de potentiels d'action, fréquence, durée de la salve) sont en rapport avec l'intensité de stimulation. Le message est ensuite véhiculé par des neurones en T qui constituent la voie afférente du réflexe et dont les fibres périphériques empruntent le nerf sciatique avant de rejoindre la moelle épinière par la racine dorsale. Le message sensoriel pénètre alors dans la corne dorsale ipsilatérale (du même côté) où il est transmis à des interneurones qui le relaient jusqu'aux motoneurones commandant la musculature du pied situés dans la corne ventrale ipsilatérale. À ce niveau l'information est traitée et une réponse motrice permettant la flexion du pied est élaborée. Le nouveau message, efférent et véhiculé par les axones des motoneurones du fléchisseur du pied, quitte alors la moelle par la racine ventrale et descend dans le nerf sciatique jusqu'au muscle fléchisseur du pied.
Puis, au fur et à mesure que l'intensité de la stimulation augmente, la réponse se complexifie grâce à l'intervention d'un nombre croissant de neurones d'association qui relaient le message sensoriel de manière à mobiliser de plus en plus de motoneurones et, donc, de plus en plus de masses musculaires.
Toutefois, pour qu'il y ait flexion d'un membre, il faut que les muscles fléchisseurs se contractent mais il faut aussi que leurs antagonistes, c'est-à-dire les extenseurs, soient relâchés. Ceci implique que le message sensoriel à l'origine du réflexe doit tout à la fois :
exciter les motoneurones qui commandent les fléchisseurs,inhiber les motoneurones qui commandent les extenseurs.
Cette double innervation, découverte par Sherrington et désormais connue sous le
CUEEP-Université Lille 1 Sciences et Technologies12
Étude expérimentale
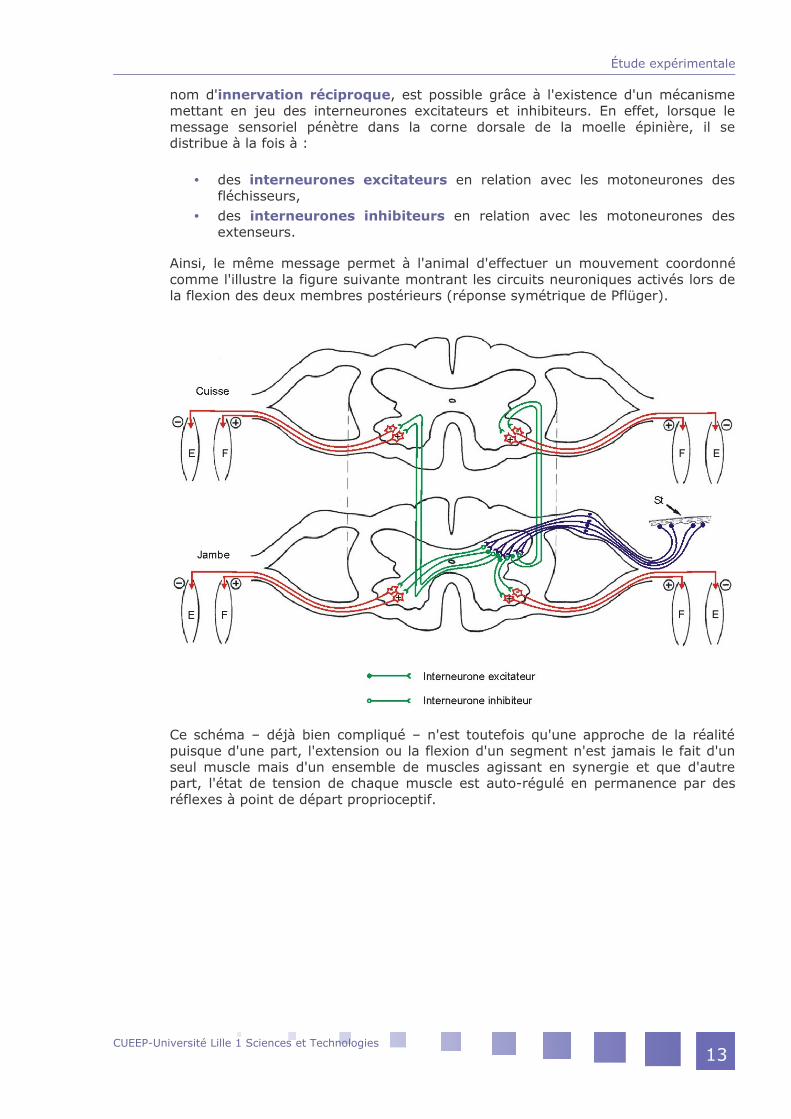
nom d'innervation réciproque, est possible grâce à l'existence d'un mécanisme mettant en jeu des interneurones excitateurs et inhibiteurs. En effet, lorsque le message sensoriel pénètre dans la corne dorsale de la moelle épinière, il se distribue à la fois à :
des interneurones excitateurs en relation avec les motoneurones des fléchisseurs,des interneurones inhibiteurs en relation avec les motoneurones des extenseurs.
Ainsi, le même message permet à l'animal d'effectuer un mouvement coordonné comme l'illustre la figure suivante montrant les circuits neuroniques activés lors de la flexion des deux membres postérieurs (réponse symétrique de Pflüger).
Ce schéma – déjà bien compliqué – n'est toutefois qu'une approche de la réalité puisque d'une part, l'extension ou la flexion d'un segment n'est jamais le fait d'un seul muscle mais d'un ensemble de muscles agissant en synergie et que d'autre part, l'état de tension de chaque muscle est auto-régulé en permanence par des réflexes à point de départ proprioceptif.
Étude expérimentale
CUEEP-Université Lille 1 Sciences et Technologies13


IV - Le réflexemyotatique
IV
Le réflexe myotatique est un réflexe d'étirement qui permet à un muscle donné de s'opposer, en se contractant, à son propre étirement et donc de conserver un certain tonus.
À l'origine du réflexe se trouvent des petites structures dispersées dans le muscle – les fuseaux neuromusculaires – qui possèdent une double innervation motrice et sensitive. Elles renferment en effet des fibres musculaires d'un type particulier (commandées par des motoneurones spécifiques) autour desquelles s'enroule l'extrémité périphérique d'un neurone en T dont le corps cellulaire se trouve dans un ganglion rachidien. Ainsi, lorsque le muscle est étiré, les fibres musculaires intrafusoriales le sont également, ce qui provoque la décharge des fibres nerveuses sensitives qui en sont issues. Le message est alors transmis directement aux motoneurones du muscle concerné ce qui a pour effet de les exciter et de provoquer la contraction du muscle en réponse à son étirement initial.
Ici encore, le principe d'innervation réciproque s'applique de manière à ce que la contraction du muscle concerné s'accompagne du relâchement de son antagoniste. Le mécanisme est automatique car lorsque les fibres sensitives à l'origine du réflexe myotatique pénètrent dans la moelle épinière, elles s'articulent directement avec les motoneurones du muscle concerné (l'agoniste) mais également avec ceux du muscle antagoniste par l'intermédiaire d'interneurones inhibiteurs.
L'étirement initial provoque donc simultanément :
la contraction du muscle agoniste (par un arc réflexe monosynaptique),le relâchement du muscle antagoniste (par un arc réflexe polysynaptique).
CUEEP-Université Lille 1 Sciences et Technologies15
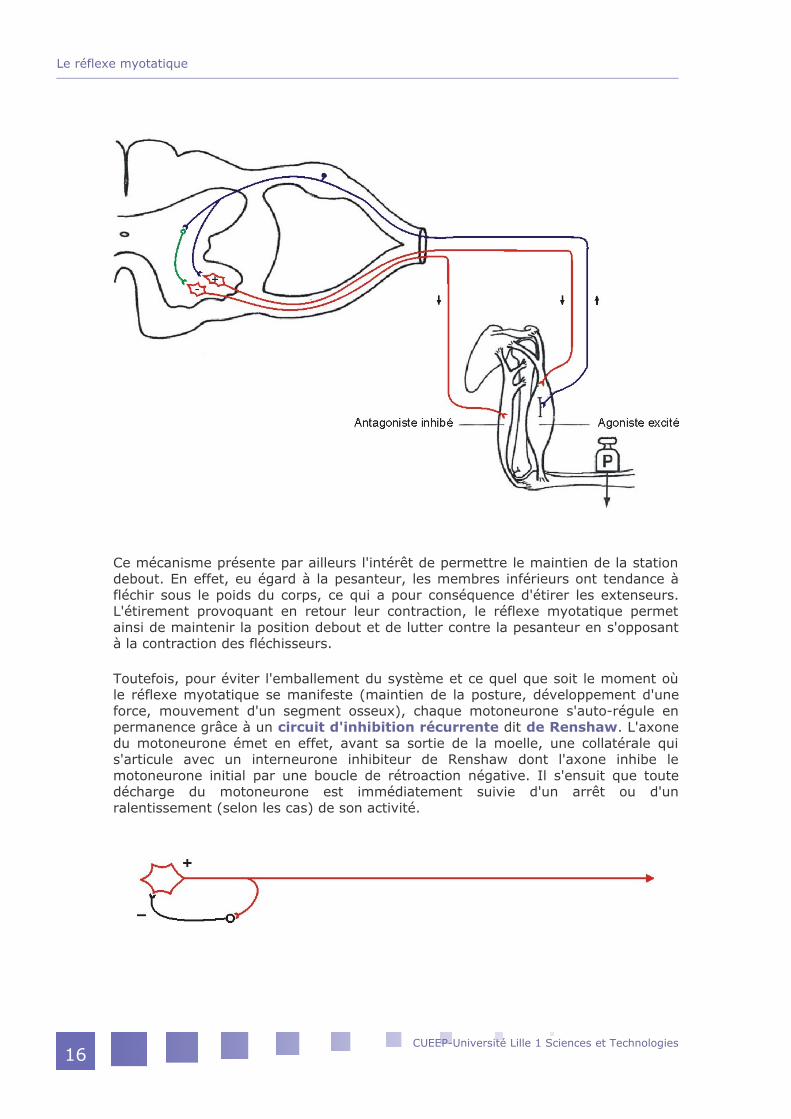
Ce mécanisme présente par ailleurs l'intérêt de permettre le maintien de la station debout. En effet, eu égard à la pesanteur, les membres inférieurs ont tendance à fléchir sous le poids du corps, ce qui a pour conséquence d'étirer les extenseurs. L'étirement provoquant en retour leur contraction, le réflexe myotatique permet ainsi de maintenir la position debout et de lutter contre la pesanteur en s'opposant à la contraction des fléchisseurs.
Toutefois, pour éviter l'emballement du système et ce quel que soit le moment où le réflexe myotatique se manifeste (maintien de la posture, développement d'une force, mouvement d'un segment osseux), chaque motoneurone s'auto-régule en permanence grâce à un circuit d'inhibition récurrente dit de Renshaw. L'axone du motoneurone émet en effet, avant sa sortie de la moelle, une collatérale qui s'articule avec un interneurone inhibiteur de Renshaw dont l'axone inhibe le motoneurone initial par une boucle de rétroaction négative. Il s'ensuit que toute décharge du motoneurone est immédiatement suivie d'un arrêt ou d'un ralentissement (selon les cas) de son activité.
CUEEP-Université Lille 1 Sciences et Technologies16
Le réflexe myotatique

Par ailleurs, comme la cellule de Renshaw innerve aussi l'interneurone inhibiteur de l'innervation réciproque, il s'ensuit que la décharge initiale du motoneurone commandant l'agoniste provoque également en retour une inhibition de l'inhibiteur du motoneurone de l'antagoniste, c'est-à-dire une levée de son inhibition.
Enfin, précisons pour terminer que :
le réflexe myotatique se double d'un réflexe myotatique inverse, à point de départ tendineux, déclenché par une contraction excessive du muscle ;des messages nerveux issus des centres supérieurs se projettent sur les motoneurones et modulent l'activité réflexe de la moelle épinière en fonction de la vigilance du sujet et de sa position dans l'espace.
De sorte que les circuits mis en jeu lors d'un réflexe, aussi simple soit-il, sont toujours en réalité beaucoup plus complexes que la représentation que nous en avons faite.
Le réflexe myotatique
CUEEP-Université Lille 1 Sciences et Technologies17


V - Exercicesd'application
V
Exercice d'application N°1 19
Exercice d'application N°2 20
Exercice d'application N°3 20
Exercice d'application N°4 21
A. Exercice d'application N°1
On effectue plusieurs expériences chez une pieuvre, un animal où le système nerveux comprend deux parties : une partie sus-œsophagienne constituée d'un ganglion céphalique et une partie sous-œsophagienne faite d'une chaîne ganglionnaire reliée aux organes par des nerfs mixtes.
Dans un premier temps, le ganglion céphalique est détruit. L'animal semble indifférent à ce qui l'entoure. Toutefois, la piqûre d'un tentacule provoque son retrait et si l'on renouvelle plusieurs fois l'expérience, le tentacule se retire toujours de la même manière.
Question 1[Solution n°1 p 23]
Quel type de comportement a-t-on mis en évidence et que pouvez-vous en déduire quant au rôle de la chaîne ganglionnaire ?
On réalise alors les observations suivantes sur plusieurs tentacules.
a) La piqûre d'un tentacule anesthésié n'entraîne plus de comportement de retrait.
b) La piqûre d'un tentacule dont le nerf a été sectionné n'est pas suivie d'effet.
c) La stimulation du nerf d'un tentacule anesthésié provoque son retrait.
d) La même expérience répétée sur un tentacule curarisé n'est pas suivie d'effet (rappelons que le curare est un agent bloquant la transmission neuromusculaire).
e) La destruction de la chaîne ganglionnaire sous-œsophagienne abolit tout comportement de retrait suite à une piqûre.
CUEEP-Université Lille 1 Sciences et Technologies19

Question 2[Solution n°2 p 23]
Interprétez ces données expérimentales et précisez quels sont les éléments anatomiques mis en jeu dans le comportement de retrait.
B. Exercice d'application N°2
Afin d'étudier le rôle des racines rachidiennes, on réalise plusieurs types de sections chez un animal spinal.
Question 1[Solution n°3 p 24]
Quelles seront les conséquences immédiates de chaque section ?
Question 2[Solution n°4 p 24]
Sachant que les axones qui sont rattachés au corps cellulaire régénèrent et qu'ils sont capables d'établir de nouvelles synapses fonctionnelles, quelles séquelles résulteraient de chaque section si l'animal était gardé en vie et en supposant que la régénération ait été totale ?
C. Exercice d'application N°3
François Magendie (1784-1855), l'un des pionniers de la physiologie expérimentale en France, fut le premier à mettre en évidence le rôle des racines rachidiennes. Un extrait de son journal, publié en 1822, relate ses premières expérimentations chez le Chien.
« Depuis longtemps, je désirais faire une expérience dans laquelle je couperais sur un animal les racines postérieures des nerfs qui naissent de la moelle épinière (...) J'eus alors sous les yeux les racines postérieures des paires lombaires et sacrées et, en les soulevant successivement avec les lames de petits ciseaux, je pus les couper d'un côté, la moelle restant intacte. J'ignorais quel serait le résultat de cette tentative (...) et j'observais l'animal ; je crus d'abord le membre correspondant aux
CUEEP-Université Lille 1 Sciences et Technologies20
Exercices d'application

nerfs coupés entièrement paralysé ; il était insensible aux piqûres et aux pressions les plus fortes ; il me paraissait immobile, mais bientôt, à ma grande surprise, je le vis se mouvoir d'une manière très apparente, bien que la sensibilité y fut toujours tout à fait éteinte. Une seconde, une troisième expérience me donnèrent exactement le même résultat (...) Il se présentait naturellement à l'esprit de couper les racines antérieures en laissant intactes les postérieures (...) Comme dans les expériences précédentes, je ne fis la section que d'un seul côté, afin d'avoir un terme de comparaison. On conçoit avec quelle surprise je suivis les effets de cette section. Ils ne furent point douteux : le membre était complètement immobile et flasque tandis qu'il conservait une sensibilité sans équivoque. Enfin, pour ne rien négliger, j'ai coupé à la fois les racines antérieures et postérieures : il y eut perte absolue de sentiment et de mouvement. »
Question [Solution n°5 p 24]
Interprétez ces données expérimentales.
D. Exercice d'application N°4
Chez un animal spinal, la stimulation supraliminaire d'une patte postérieure déclenche un réflexe ipsilatéral de flexion. On enregistre alors les variations de tension mécanique consécutives à la stimulation de deux muscles de la cuisse : le quadriceps fémoral (extenseur de la jambe) et le biceps crural (fléchisseur de la jambe). Les résultats sont les suivants.
Question 1[Solution n°6 p 24]
Que met-on en évidence ?
Exercices d'application
CUEEP-Université Lille 1 Sciences et Technologies21

Question 2[Solution n°7 p 25]
En conditions normales, on observe que le réflexe ipsilatéral de flexion d'un membre postérieur s'accompagne généralement d'un réflexe contralatéral d'extension. Quel peut en être l'intérêt pour l'animal ?
Question 3[Solution n°8 p 25]
Décrivez les circuits neuroniques mobilisés dans ce dernier cas.
CUEEP-Université Lille 1 Sciences et Technologies22
Exercices d'application

Solution des exercices de TD
> Solution n°1 (exercice p. 19)
Le ganglion céphalique ayant été détruit, seule la chaîne ganglionnaire sous-œsophagienne peut être impliquée dans le comportement de retrait de l'animal. Par ailleurs, le fait que le tentacule se retire toujours de la même manière, à la suite d'une stimulation identique, évoque un comportement réflexe. La chaîne ganglionnaire sous-œsophagienne est ici l'équivalent de la moelle épinière chez les Mammifères.
> Solution n°2 (exercice p. 20)
Chaque expérience apporte une information sur les différents éléments anatomiques mis en jeu dans le comportement de retrait.
a) L'anesthésie empêche les récepteurs de réagir à la stimulation. Comme aucun message sensoriel ne parvient à la chaîne ganglionnaire, il est donc normal que le réflexe n'apparaisse pas.
b) Cette fois les récepteurs ont réagi à la piqûre, un message sensoriel a été élaboré mais, l'afférence étant sectionnée, il ne peut parvenir à la chaîne ganglionnaire et donc déclencher le réflexe de retrait.
c) Comme précédemment, les récepteurs anesthésiés n'ont pu élaborer de message sensoriel. Toutefois la stimulation du nerf qui relie le tentacule à la chaîne ganglionnaire revient à déclencher artificiellement le réflexe et donc à permettre le retrait. En revanche, le nerf étant mixte, il est impossible avec cette seule expérience de savoir si on a stimulé les afférences, de manière à simuler un message sensoriel en provenance des récepteurs, ou les efférences, de manière à simuler un message moteur à destination des muscles permettant le retrait du tentacule.
d) Le tentacule étant curarisé, les messages nerveux qui parviennent aux muscles ne peuvent être efficaces. Cela revient à paralyser les effecteurs qui ne peuvent donc accomplir la phase finale du réflexe.
e) Cette fois le centre réflexe étant détruit, le message sensoriel élaboré par les récepteurs ne peut plus arriver à destination. Il ne peut donc plus y avoir de réflexe puisque l'arc est interrompu entre les voies afférente et efférente.
En conclusion, cette série d'expériences illustre la nécessité de respecter l'intégrité
CUEEP-Université Lille 1 Sciences et Technologies23

des cinq éléments indispensables au bon accomplissement du réflexe. À savoir : les récepteurs, les afférences, le centre, les efférences et les effecteurs.
> Solution n°3 (exercice p. 20)
La section de la racine dorsale (A ou B) ne permettant plus aux messages afférents de pénétrer dans la moelle épinière, on observera une anesthésie du territoire concerné. En revanche, la section de la racine ventrale (C) se traduira par une paralysie du territoire concerné. En effet, les axones issus des motoneurones étant interrompus, les messages efférents ne pourront arriver à destination. Enfin, la section du nerf rachidien (D) affectant à la fois la voie afférente et la voie efférente, on observera une anesthésie et une paralysie du territoire concerné.
> Solution n°4 (exercice p. 20)
À la suite de la section A, les axones centraux des neurones en T du ganglion rachidien repousseront et établiront de nouvelles synapses dans la moelle, de sorte que si la régénération est totale, il n'y aura aucune séquelle pour l'animal. En revanche, à la suite de la section B, les axones périphériques des neurones en T ayant été séparés de leurs corps cellulaires, la partie comprise entre les récepteurs et le ganglion rachidien ne régénèrera pas, ce qui se traduira par une perte de la sensibilité du territoire concerné. La section C n'engendrera par contre aucune séquelle. En effet, les axones des motoneurones n'ayant jamais été séparés de leurs corps cellulaires, ils repousseront dans la racine ventrale et, une fois la régénération achevée, l'animal retrouvera la motricité du territoire concerné. Enfin, la section D revenant à cumuler les sections B et C, l'animal perdra sa sensibilité mais conservera sa motricité.
> Solution n°5 (exercice p. 21)
En coupant les racines postérieures (dorsales) d'un seul côté, Magendie empêche les messages sensoriels en provenance du membre ipsilatéral de pénétrer dans la moelle épinière. Il obtient donc une insensibilité du territoire concerné sans que la motricité du membre soit en apparence altérée (elle l'est en réalité car la section des racines postérieures ne permet plus aux muscles de réguler leur tonus par voie réflexe). En revanche, lorsqu'il sectionne les racines antérieures (ventrales), il interrompt la voie efférente et empêche donc les axones des motoneurones d'atteindre leurs cibles. Il obtient donc une paralysie du membre concerné qui conserve ici sa sensibilité. Enfin, la section des racines antérieures et postérieures provoquant l'interruption des voies afférentes (sensitives) et efférentes (motrices), il est normal que Magendie observe une « perte absolue de sentiment et de mouvement ».
> Solution n°6 (exercice p. 21)
On constate que suite à la stimulation, la tension mécanique du fléchisseur (le biceps crural) augmente alors que celle de l'extenseur (le quadriceps fémoral) diminue. C'est donc que les motoneurones du premier ont été activés et que les motoneurones du second ont été inhibés, conformément au principe d'inhibition réciproque décrit par Sherrington, de manière à permettre la flexion de la jambe.
CUEEP-Université Lille 1 Sciences et Technologies24
Annexes

> Solution n°7 (exercice p. 22)
Le fait que le membre contralatéral réagisse par une extension permet à l'animal de mieux conserver sa position debout alors que l'autre membre est fléchi et qu'il ne participe donc plus au maintien du corps.
> Solution n°8 (exercice p. 22)
Les circuits neuroniques mobilisés sont les suivants : à leur arrivée dans la moelle, les messages afférents issus des récepteurs stimulés se distribuent à la fois aux motoneurones de la corne ventrale ipsilatérale par l'intermédiaire d'interneurones excitateurs pour ceux qui commandent les fléchisseurs et inhibiteurs pour ceux qui commandent les extenseurs et aux motoneurones de la corne ventrale contralatérale par l'intermédiaire d'interneurones excitateurs pour ceux qui commandent les extenseurs et inhibiteurs pour ceux qui commandent les fléchisseurs.
Annexes
CUEEP-Université Lille 1 Sciences et Technologies25