bioproducts from sulfite pulping: lisa x. lai a thesis
TRANSCRIPT

Bioproducts from sulfite pulping:
Bioconversion of sugar streams from pulp, sludge, and spent sulfite liquor
Lisa X. Lai
A thesis
submitted in partial fulfillment of the
requirements for the degree of
Master of Science
University of Washington
2010
Program Authorized to Offer Degree:
School of Forest Resources

University of Washington
Graduate School
This is to certify that I have examined this copy of a master’s thesis by
Lisa X. Lai
and have found that it is complete and satisfactory in all respects,
and that any and all revisions required by the final
examining committee have been made.
Committee Members:
_____________________________________________________
Renata Bura
_____________________________________________________
Rick Gustafson
_____________________________________________________
William McKean
Date:__________________________________

i
TABLE OF CONTENTS Page
List of figures ............................................................................................ ii
List of tables ............................................................................................ iii
Chapter 1: Introduction ............................................................................ 1
1.1 Lignocellulosic biomass................................................................. 3
1.2 Pretreatment ................................................................................... 6
1.3 Sulfite pulping ................................................................................ 8
1.4 Enzymatic hydrolysis ..................................................................... 8
1.5 Fermentation ................................................................................ 10
1.6 SHF vs. SSF ................................................................................. 12
1.7 Products from a sugar platform .................................................... 13
Chapter 2: Draft paper ........................................................................... 14
Introduction ..................................................................................... 15
Methods .......................................................................................... 16
Results and discussion .................................................................... 20
Conclusion ...................................................................................... 26
Chapter 3: Additional results and discussion ......................................... 28
3.1 Fermentation of SSL using S. cerevisiae ..................................... 28
3.2 Effect of solids loading on enzymatic hydrolysis ........................ 29
3.3 Effect of enzyme loading on hydrolysis ...................................... 30
Chapter 4: Additional conclusions and future work .............................. 32
Acknowledgements ................................................................................. 33
References ............................................................................................... 34

ii
LIST OF FIGURES
Page
1. Bioconversion process schematic………………………………2
2. Experimental design…………………………………………...17
3. Spent sulfite liquor C. guilliermondii fermentation……………22
4. Pulp and sludge hydrolysis….…………………………………23
5. Fermentation of pulp hydrolysates…………………………….24
6. Fermentation of sludge hydrolysates…………………………..25
7. Combined feedstock SSF………………………………………26
8. Spent sulfite liquor S. cerevisiae fermentation………………...28
9. Variable consistency hydrolysis……………………………….29
10. Variable enzyme loading hydrolysis…………………………..31

iii
LIST OF TABLES
Page
1. Pulp and sludge composition…………………………………..20
2. Spent sulfite liquor composition……………………………….21
3. Solid stream result summary…………………………………..27

1
Chapter 1: Introduction
The global impact of human energy consumption has been made increasingly apparent in recent years
by growing rates of atmospheric CO2 accumulation, likely caused by a rise in anthropogenic burning
of fossil fuels (Canadell, Le Quéré et al. 2007). It has been shown that mitigation of these effects
may be achieved through displacement of petroleum-based transportation fuels with a biofuel
producing less harmful emissions, such as ethanol (Tyson 1993; von Sivers and Zacchi 1995;
Bergeron 1996; Galbe and Zacchi 2002). In addition to environmental benefits, ethanol addresses
concerns about the exhaustion of available fossil fuels, energy security, and ease of adaptation to the
current fuel infrastructure in the US.
Though ethanol is overwhelmingly produced from sugar cane in Brazil, and corn in North America,
lignocellulosic biomass, such as agricultural and wood residue, is a widely available and largely
untapped feedstock (Wiselogel, Tyson et al. 1996). In addition, ethanol production from
lignocellulosics, commonly called bioethanol, has been shown to have a more favorable energy ratio
than that originating from starch or sugar sources (Keeney and DeLuca 1992; Lorenz, Morris et al.
1995; Gnansounou and Dauriat 2005). Major process steps for the traditional bioconversion scheme
of lignocellulosic biomass to bioethanol are well known and illustrated in Figure 1. First,
lignocellulosic biomass is pretreated to break up its rigid structure and prepare it for the next step,
hydrolysis. During enzymatic hydrolysis, sugars are released from the pretreated biomass.
Fermentation utilizes microorganisms to convert these sugars to bioproducts, for example ethanol and
xylitol.
Though laboratory-scale bioethanol production has been increasingly studied in recent years, no full-
scale commercial plants exist in the US today. One reason is that challenges exist in each step of the
bioconversion scheme, which will be discussed in greater detail in subsequent chapters. First, an
ideal biomass is difficult to find, as it must be inexpensive, readily-available in large quantities, and
consistent in chemical composition. Pretreatment requires optimization of severity, as the ideal
degree of biomass fractionation is often limited by the formation of fermentation inhibitors.
Hydrolysis necessitates consideration for the cost of enzymes, potentially requiring sacrifices in sugar
yield. Finally, fermentation requires selection of an ideal microorganism for the specific operation at
hand, making considerations for inhibitor tolerance and possible fermentation of co-products
alongside of bioethanol.

2
Sugar streams from sulfite pulping, described in greater detail later, were studied in this thesis as a
means of mitigating the problems associated with the biomass acquisition and pretreatment steps
described above. Spent sulfite liquor (SSL), pulp, and sludge, the streams studied in this thesis, are
concurrently produced with bleached sulfite pulp at a rapid rate, thus meeting the criteria of being
inexpensive and available in large quantities. In addition, due to the majority of fractionation and
delignification already being accomplished by the sulfite pulping process, the streams studied here do
not require pretreatment.
Figure 1. Simplified process schematic for the conversion of lignocellulosic
biomass to bioproducts. This process flow illustrates the separate hydrolysis and
fermentation (SHF) process, however hydrolysis and fermentation can alternatively
be combined into one step, or a process known as simultaneous saccharification and
fermentation (SSF).
Objective and Outline
The main objective of the work presented in this thesis is to examine the potential of producing single
and mixed sugars from sulfite pulping streams to use for the production of bioethanol and other
valuable bioproducts. This was accomplished first through the fermentation of SSL with Candida
guilliermondii, a microorganism capable of fermentation mixed 5-and 6-carbon sugars. Next, single
sugars streams were produced from the separate hydrolysis and fermentation (SHF) of pulp and
sludge in water. Finally, mixed sugars streams were produced from SHF and simultaneous
saccharification and fermentation (SSF) of pulp and sludge mixed with SSL. Success of this process
would create the potential to exploit a new feedstock for bioethanol conversion, as well as address
some of the challenges associated with bioconversion described above.
Several background sections are presented in order to put this research into context. Section 1.1
provides a background about biomass chemistry. Section 1.2 discusses pretreatment methods and
Lignocellulosic biomass
Pretreatment Hydrolysis Fermentation
Bioproducts (ethanol, xylitol, etc.)

3
limitations. Section 1.3 discusses sulfite pulping as a means of pretreatment. Section 1.4 describes
enzymatic hydrolysis. Section 1.5 provides an overview of fermentation in terms of the
microorganisms used in this study and their associated products. Section 1.6 compares separate
hydrolysis and fermentation (SHF) and simultaneous saccharification and fermentation (SSF)
bioconversion processes. Finally, Section 1.7 contains an overview of the most promising
biochemical building blocks that can be made from biomass. Chapter 2 contains a draft paper that
will be submitted for publication regarding the use of sulfite pulping streams as a sugar platform from
which to produce higher-value bioproducts. Additional results and discussion not presented in Paper
I are shown in Chapter 3, followed by additional conclusions and future work in Chapter 4.
1.1 Lignocellulosic biomass
Feedstocks for ethanol production can be categorized into sucrose-, starch-, and lignocellulosic-based
materials. Sucrose-based materials are readily fermentable to ethanol and other products, while
starch-based feedstocks can easily be converted to sugars via enzymatic hydrolysis. Lignocellulosic
feedstocks represent great production value, but due to their chemistry, are the most challenging to
utilize for bioconversion. Lignocellulosics encompass a broad range of materials, including woody
biomass, herbaceous plants (switchgrass and giant reed), and agricultural residues (corn stover and
wheat straw). Common among all lignocellulosic biomass types are their three major components of
cellulose, hemicellulose, and lignin, and minor components of extractives and ash. These constituent
compounds are present in varying quantities among biomass types, depending on their species of
origin.
Cellulose
Lignocellulosic feedstocks are derived from plant biomass, whose defining characteristic at the
cellular level is the presence of cell walls comprised primarily of cellulose. The most abundant
natural polysaccharide, cellulose is the target of bioethanol conversion processes. Cellulose is
comprised of D-glucose units linked by β-1,4 glycosidic bonds. A dimer of two linked glucose units
is referred to as cellobiose. It is because of this linkage that each individual cellulose molecule forms
a linear polymer. Bioconversion necessitates the release of these individual glucose units for
microbial fermentation to bioproducts. The degree of polymerization (DP) of cellulose, or number of
monomeric units in an oligomeric molecule, varies by source. Sulfite pulp, one of the feedstocks
used in this study, averages a DP of 1255, while softwood tree species, the primary source of the
sulfite pulp used in this study, average a DP of 8000 (Fengel and Wegener 1989). Cellulose

4
molecules with a DP less than 8 are considered water-soluble, while at higher DPs, they have a
greater affinity for one another than for water (Brown 2004). In native cellulose, individual
molecules form rigid microfibrils in which they are aligned with reducing ends oriented in the same
direction. This structure is stabilized by rigidly arranged intra- and intermolecular hydrogen bonds,
thus giving native cellulose a highly crystalline conformation that is difficult to degrade (Klemm,
Heublein et al. 2005). Amorphous (non-crystalline) regions have also been found to comprise 10-
50% of native cellulose (Fan, Lee et al. 1982), but the influence of crystallinity on cellulose
degradability is still being investigated (Atalla and VanderHart 1999; Jarvis 2003; Ding and Himmel
2006). The rigid structure of cellulose is resistant to chemical and mechanical alteration, thus posing
a challenge for the bioconversion process (Ding and Himmel 2008).
Hemicellulose
Similar to cellulose, hemicellulose is also a polysaccharide, but its constituent sugars are both hexoses
(6-C sugars; D-glucose, D-mannose, and D-galactose) and pentoses (5-C sugars; D-xylose and L-
arabinose). With a few exceptions, pentoses are generally not readily fermentable by naturally
occurring yeasts. Hemicellulose has a much lower DP, and therefore lower molecular weight, than
cellulose, and is often a branched, rather than linear polymer. Hemicellulose composition and DP
varies widely across species. Softwood hemicellulose is majorly comprised of galactoglucomannans,
composed of a backbone of β-1,4 linked mannose and glucose units in roughly a 3:1 ratio. Acetyl
groups occur on the C2 or C3 carbon of roughly every third backbone unit. The glucomannan
backbone is branched with α-1,6 linked galactose side chains occurring on glucose backbone units.
Softwood galactoglucomannans constitute about 20% of dry wood by mass, and ranges from 100-150
in DP (Fengel and Wegener 1989). Softwoods also contain 10-15% xylans, composed of a β-1,2
linked xylose backbone. One in 10 xylose units has a branched α-1,4 linked arabinose unit, and one
in five has a α-1,2 linked 4-O-methylglucuronic acid unit (Fengel and Wegener 1989). In contrast,
hardwood hemicellulose is mainly comprised of xylans (15-30%) characterized by a β-1,4 linked
xylose backbone, where many of its xylose units are acetylated at their C2 or C3 carbons. Side chains
of α -1,2 linked 4-O-methylglucuronic acid units occur at every 6-11 xylose units. Hardwood xylan
has a DP of 100-200. Mannans occur in only 3-5% of hardwood by mass, and are characterized by β
-1,4 linked backbone of mannose and glucose units in roughly a 2:1 ratio with no side chains. Native
hemicellulose is embedded within the cell wall, binding to cellulose and components, such as
structural proteins. The heterogeneity of hemicellulose makes it non-crystalline, and easier to
hydrolyze.

5
Lignin
The second most abundant organic substance within plant biomass is lignin, a highly complex
polymer consisting of phenolic compounds. Lignin composition again varies greatly between
species, but its three constituent phenolic ring structures are ρ-hydroxyphenyl, guaiacyl (containing
one methoxyl group), and syringyl (containing two methoxyl groups), which are bound to one another
through a complex network of carbon-carbon and ether bonds. Softwood lignin is comprised
overwhelmingly by guaiacyl units. Common lignin linkages include β-O-4, α-O-4, 4-O-5, 5-5, β-β,
β-5, and β-1, all of which, with the exception of α-O-4, are formed through free radical coupling of
precursor ring structures (Fengel and Wegener 1989). Roughly 50% of softwood lignin is β-O-4
linked, a reactive linkage under alkaline pulping conditions. The next most common linkages for
softwood lignin are β-5, comprising 9-12%, and 5-5, comprising 10-11%. All other linkages each
comprise less than 8% of softwood lignin (Fengel and Wegener 1989). Like glue, lignin binds the
cell wall components together, giving lignocellulosic biomass its structural integrity, while also
contributing to its flexibility (Pan 2008). The presence of lignin poses one of the biggest challenges
to bioconversion, as its removal is necessary in order to allow hydrolysis enzymes access to cellulose
and its constituent sugars (Akin 2007; Pan 2008).
Extractives and ash
Though present in very small quantities, typically below 5%, organic extractives can still greatly
affect certain properties of biomass, including its color, odor, and density (Kai 1991). In nature,
extractives perform the functions of protection from diseases and parasites, attraction of pollinators
and seed dispersers, and food storage. Softwood extractives consist mainly of terpenes, fats, waxes,
and phenolics (Fengel and Wegener 1989).
Ash is composed of the inorganic compounds remaining after complete combustion of biomass.
Though comprising only about 1% of wood biomass, ash is composed of Ca, K, and Na oxides that
are essential for plant growth (Ohlsson 2004).
Ideal feedstock
Though lignocellulosic bioconversion has been increasingly studied in recent years, an ideal
feedstock has yet to be found. Pretreatment and other steps in the bioconversion process must be
optimized in a manner that is dependent on feedstock composition, so the wide compositional
variability in biomass is problematic. Also, a desirable feedstock should be high in cellulose content,
while low in lignin content to maximize the ease and efficiency of bioconversion. Ideal feedstocks

6
must also be free of non-native contaminants that may interfere with hydrolysis or fermentation,
rendering many industrial residues (e.g. wood and paper waste) undesirable.
There are also economic and social considerations. Biomass can account for 25- 40% of the cost of
bioethanol production (Nguyen and Saddler 1991; Gregg and Saddler 1996; Galbe and Zacchi 2002;
Wingren, Galbe et al. 2003). An inexpensive feedstock with year-round availability is ideal, making,
for instance, agricultural grasses less desirable due to their high cultivation costs and seasonally-
dependent growth rates. Also, the acquisition of an ideal biomass should not be in competition with
other industries, such as agriculture or wood products manufacturing. This topic has become more
relevant with the advent of recent studies regarding the downfalls of corn ethanol (Pimentel 2003).
An ideal feedstock exhibiting all of these characteristics has proven to be difficult to find.
1.2 Pretreatment
As previously described, the crystallinity of cellulose, its particle size, available surface area, and the
presence of lignin make raw lignocellulosic biomass resistant to direct saccharification by hydrolysis
enzymes. Therefore, a pretreatment step is needed in order to alter biomass’ rigid structure (Figure
1). During pretreatment, the surface area of biomass is increased, particle size decreased, and, to
some degree, the crystallinity of cellulose is decreased (Mosier, Wyman et al. 2005). The cell wall
matrix is disrupted, accompanied by partial removal of lignin and hemicellulose. Mechanical,
chemical, and biological methods, or any combination of these, are utilized in pretreatment.
Mechanical methods involve milling or grinding, often in combination with a thermal treatment such
a steaming. Chemical pretreatment involves the addition of acid or alkali solvents that result in
partial degradation of lignin or hemicellulose. Biological pretreatment subjects the biomass to lignin-
degrading microorganisms.
Steam explosion is a method that has been found to be very efficient for woody feedstocks. Biomass
is subjected to high temperature (185-240°C) and pressure (87-500 psi) for a period of 10 s to 10 m,
followed by a rapid release of pressure, which is said to cause an “explosion” of biomass into its
fractionated form (Mason 1933; DeLong 1977). Optimal conditions for temperature, pressure, and
time are dependent on feedstock composition. The addition of an acid catalyst such as H2SO4 or SO2
to raw biomass before steam explosion has been found to improve pretreatment efficiency (Ramos,
Breuil et al. 1992; Eklund, Galbe et al. 1995).

7
Ammonia fiber expansion (AFEX) pretreatment is a similar process to steam explosion, in which
biomass is combined with ammonia at 100-400 psi and 70-200C before a rapid release of pressure.
Yields of up to 90% of the theoretical ethanol yield have been observed using AFEX pretreatment in
switchgrass (Alizadeh, Teymouri et al. 2005), however, while effective on agricultural residues, this
method has not been found to work with softwoods (Hsu 1996).
Organosolv pulping has emerged as a pretreatment method for ethanol production. In this process,
biomass is combined with ethanol, water, and H2SO4, and cooked under high temperature and
pressure. The resultant slurry is filtered, and solids are hydrolyzed, while lignin is precipitated from
the filtrate. The residual liquid fraction, containing hydrolyzed sugars, can be concentrated and also
fermented (Lora and Aziz 1985).
Biological pretreatment is carried out through the delignification of biomass by white rot fungi,
mostly belonging to the phylum Basidiomycotina (Blanchette 1991). Lignin degradation by these
microorganisms occurs via the enzyme activity of lignin peroxidase, manganese peroxidase, and
laccase, though some cellulose degradation is frequently observed due to non-specific enzyme
binding (Pointing, Pelling et al. 2005). The degree of preferential lignin consumption can vary
widely between organisms. For example, cellulose loss of 65% has been observed for Trametes
versicolor, while for Perenniporia medulla-panis, 73% delignification has been achieved without any
loss of cellulose (Blanchette 1991). Due to a slow reaction rate, the largest drawback of solely using
biological pretreatment is the long residence time that is required, up to 60 days, relative to that of
steam explosion (Taniguchi, Suzuki et al. 2005; Yu, Zhang et al. 2009). Microbial delignification has
more prominently been studied as a pretreatment method to use in conjunction with organosolv
pulping, where it has been found to decrease energy consumption by up to 68% (Blanchette 1991).
The severity factor, or the degree to which biomass is fractionated during pretreatment, is dependent
on conditions such as temperature and residence time, and can be represented by the following
formula:
Log RO = t * e(Tr-Tb)/14.75
Where t = residence time (m), Tr = reaction temperature, and Tb = reference temperature (100°C)
(Abatzoglou, Chornet et al. 1992). Too low of a severity can lead to incomplete fractionation of
biomass, causing low hydrolysis yields in the next step. Higher severity pretreatments result in more
complete delignification and a higher degree of polysaccharide hydrolysis, which are desirable

8
reactions. However, unfavorable reactions also occur during pretreatment. Hexose sugars are
degraded to hydroxymethylfurfural (HMF), and pentoses to furfural. Further degradation of HMF
leads to the formation of formic and levulinic acids (Fengel and Wegener 1989). Acetic acid is
produced through the cleavage of acetyl groups in hemicellulose. These compounds are inhibitory to
fermentation, and, at high enough concentrations, toxic to yeast (Delgenes, Moletta et al. 1996).
Therefore, pretreatment optimization is essential to achieve efficient bioconversion.
In addition, pretreatment can account for a large portion of the energy and/or chemical input needed
in any bioconversion process. For this reason, and the need for optimization described above,
pretreatment is often seen as the limiting step of bioconversion.
1.3 Sulfite pulping
The feedstocks utilized here for bioconversion were sugar streams produced from ammonia-based
acid sulfite pulping. This process involves the mixing of raw, mainly softwood chips with SO2 at
high temperature and acidic pH. Pulping occurs through the interaction of lignin with SO2, whereby
lignin is solubilized through the addition of hydrophillic sulfonate groups. Sulfonation mainly
occurs on the α, and sometime γ carbon of lignin groups (Gellerstedt 1973). In addition,
carbohydrates are cleaved from lignin-carbohydrate-lignin linkages, reducing lignin’s molecular
weight. In hemicellulose, acetyl groups are cleaved, as well as α-1, 6 galactosidic linkages (Glennie
1971). Therefore, the resulting spent sulfite liquor (SSL), as it is commonly referred, is high in
monomeric sugar and sulfonated lignin (lignosulfonate) content. The resulting pulp is mostly
delignified, and high in glucose content. Sludge, which is made up of pulping fines and rejects, is
similar to pulp in sugar composition, but contains a high amount of residual lignin.
These three streams were obtained for this study from collaborators at Kimberly-Clark in Everett,
WA. Utilization of these as feedstocks for bioconversion makes pretreatment obsolete, as a high
degree of solids delignification can be expected from the sulfite cooking process, and sugars in the
liquid stream are expected to be mostly in monomeric form. Also, sulfite pulping provides the benefit
of a central location where sugar streams are already being generated at no additional cost.
1.4 Enzymatic hydrolysis
Saccharification of cellulose into its monomeric glucose components is achieved by the action of a
group of enzymes collectively referred to as cellulases. These enzymes are synthesized by various
naturally-occurring bacteria and fungi, the most prominent of these being Trichoderma reesei, an

9
aerobic mesophilic fungus. Its cellulase enzymes are classified into three groups: endoglucanases
(EG), cellobiohydrolases (CBH), and β-glucosidase (βG), each associated with a specific action
(Ghose 1987). EG attacks internal bonds within cellulose, exposing free ends and disrupting its
crystalline structure. Starting from these exposed free ends, CBH travels along the length of the
cellulose chain, cleaving cellobiose units, which are subsequently hydrolyzed into two glucose units
by βG. One explanation for this system of synergistic enzymatic activity is limitation of end-product
inhibition (Eriksson, Karlsson et al. 2002; Väljamäe, Kipper et al. 2003). T. reesei is cultured
industrially and its enzymes are separated and purified for use in large-scale hydrolysis.
Two user-controlled factors affecting hydrolysis rate are enzyme loading and solids consistency.
Since enzyme quantity cannot be directly measured, enzyme activity for EG and CBH is measured in
terms of filter paper units (FPU) per volume. The quantity of enzyme activity required to produce 2.0
mg of cellobiose from 50 mg of filter paper (essentially pure cellulose), or a 4% conversion, after 1 hr
at 50°C and 4.8 pH (the ideal conditions for cellulase activity) is defined as 0.1875 FPU (Decker,
Adney et al. 2003). The effect of enzyme loading on glucose yield is not linear. For example, a 50%
increase in enzyme loading would result in less than a 50% increase in glucose yield (Ghose 1987).
Enzyme loading mainly affects the initial rate of hydrolysis, and therefore, the residence time
required to reach maximum glucose yield. Therefore, a change in enzyme loading may not have a
proportionate effect on final glucose yield itself. Even at very high enzyme loading, the complete
hydrolysis of cellulose is often difficult to achieve due to the presence of residual lignin, and end-
product inhibition (Tengborg, Galbe et al. 2001). Both cellobiose and glucose act as noncompetitive
inhibitors of T. reesei cellulases (Holtzapple, Cognata et al. 1990; Xiao, Zhang et al. 2004). Ethanol
is also inhibitory, but to a far lesser extent (Holtzapple, Cognata et al. 1990). Accumulation of these
compounds during hydrolysis is disadvantageous. Glucose has been found to be less inhibitory than
cellobiose, so a proportionately higher loading of βG is typically used during enzymatic hydrolysis to
limit inhibition, compared to EG or CBH. Enzyme cost occupies a significant portion of the total cost
for bioconversion, so minimizing enzyme loading to the extent that it is not detrimental to glucose
yield is favorable (Shen and Agblevor 2008).
Solids consistency is described as dry weight of biomass (substrate) divided by the volume of total
liquid in a hydrolysis reaction, expressed as a percentage. Enzyme kinetic curves for cellulose
hydrolysis typically exhibit a biphasal shape, with an initial logarithmic phase followed by an
asymptotic phase as maximum glucose conversion is approached (Ramos, Breuil et al. 1993). When
expressed in terms of percent glucose conversion from cellulose, increasing solids consistency is

10
typically found to reduce the initial conversion rate. Glucose percent conversion is typically found to
decrease, but final glucose concentration may improve due to greater substrate availability (Cara,
Moya et al. 2007). High consistency hydrolysis represents a promising source of cost reduction. A
previous assessment has shown that total cost can be reduced by nearly 20% when solids consistency
is increased from 5% to 8% for softwood (Wingren, Galbe et al. 2003). However, it has been
supposed that bioconversion using solids consistencies of greater than 8% encounters three key
barriers. First, there is a reduction in the amount of free water available for enzyme transport due to
hydrogen bonding with the released sugars (Felby, Thygesen et al. 2008). Secondly, higher
consistencies can introduce a high concentration of lignin, a fermentation inhibitor (Delgenes,
Moletta et al. 1996). Finally, increased sugar production can cause severe end-product inhibition
(Holtzapple, Cognata et al. 1990). Solids loading of up to 40% has been reported in wheat straw,
resulting in a decrease in percent glucose conversion to one third that of 2% consistency hydrolysis
(Jørgensen, Vibe-Pedersen et al. 2007). However, a more promising result has been published for
hardwoods, where 20% consistency hydrolysis resulted in 158 g/L glucose (Zhang, Qin et al. 2009).
Low enzyme loading and high consistency hydrolysis represent great potential in terms of cost
reduction for bioconversion technologies.
1.5 Fermentation
Microbial fermenters include a variety of bacteria and fungi. The well-studied Saccharomyces
cerevisiae, known commonly as baker’s or brewer’s yeast, is one such organism utilized in this
report. Originally isolated from the skin of grapes, it has become a model eukaryotic organism in
biological research due to its size, ease of genetic manipulation, and high economic value. Like many
organisms, S. cerevisiae metabolizes glucose, and other 6-carbon sugars, via the Embden-Meyerhof-
Parnas glycolytic pathway, mainly producing ethanol. Genetically modified S. cerevisiae such as
424A (LNH-ST) can also be made to metabolize 5-carbon sugars (Sedlak and Ho 2004). The overall
reaction of hexose fermentation to ethanol is:
C6H12O6 2 C2H5OH + 2 CO2
The mass balance of this process indicates that the maximum possible yield of ethanol is 51% of the
mass of starting glucose. The strain of S. cerevisiae utilized in this study (ATCC 96581) was isolated
from spent sulfite liquor, and had previously demonstrated tolerance of 8 g/L of acetic acid with
superior galactose fermentation to a commercially available S. cerevisiae (Linden, Peetre et al. 1992).
Yields of about 75% of the theoretical maximum ethanol were achieved for SSL at pH 6, but other S.

11
cerevisiae strains have shown near 100% yield from synthetic sugars (Keating, Panganiban et al.
2006). Similar to other ethanologenic yeasts, the strain exhibits preferential consumption of glucose
first, followed by mannose, then galactose (Linden, Peetre et al. 1992).
A second organism, Candida guilliermondii, (ATCC 201935) was also utilized in this study to
demonstrate cofermentation of xylitol alongside of ethanol. Xylitol is primarily used as a sweetener,
and due to being sugar alcohol, does not impact insulin levels when ingested, and contains 36% fewer
calories than sucrose (Hassinger, Sauer et al. 1981). C. guilliermondii converts xylose to xylitol via
the Xylose Reductase-Xylitol Dehydrogenase (XR-XDH) pathway, represented by the net equation:
60 C5H10O5 + 12 ADP + 12 Pi + 12 H2O + 3 O2 54 C5H12O5 + 12 ATP + 30 CO2
The theoretical yield of xylitol is 91% of the starting xylose mass (Barbosa, de Medeiros et al. 1988).
C. guilliermondii also concurrently ferments hexoses to ethanol. Because of this unique ability, the
composition of media used to propagate the yeast prior to fermentation has an effect on its
productivity, an phenomenon that has been explained by enzyme induction (Lee, Sopher et al. 1996).
C. guilliermondii pre-grown in xylose media has previously been found to produce a significantly
higher xylitol yield than yeast grown in glucose alone or in mixed media. By pre-growing yeast on
xylose, yields of approximately 60% of the theoretical maximum for xylitol were found during
fermentation of hydrolysate containing both glucose and xylose (da Silva and de Almeida Felipe
2006). This yield has been found to further improve when xylose is the only sugar in the
fermentation media, to about 80% (Barbosa, de Medeiros et al. 1988).
Fermentation inhibitors released from biomass during pretreatment include of a variety of compounds
that are categorized into three groups. Weak acids (acetic acid) and furan derivatives (furfural and
HMF) are sugar degradation products, while phenolic compounds result from the degradation of
lignin during pretreatment. Inhibitors have been topic of great interest, but their mechanisms of
inhibition are still a matter of investigation (Palmqvist and Hahn-Hägerdal 2000). Though seemingly
contradictory, it has been reported by several studies that small quantities of inhibitors can enhance
fermentation yields (Banerjee, Bhatnagar et al. 1981; Sanchez and Bautista 1988; Pampulha and
Loureiro-Dias 1990; Taherzadeh, Niklasson et al. 1997). In particular, ethanologenic yeasts have
been reported to tolerate up to 10 g/L acetic acid, 3 g/L HMF, and 1.6 g/L furfural with no significant
effect on ethanol yield, and even improved yields in some cases (Keating, Panganiban et al. 2006).

12
Fermentation products, such as ethanol (Brown, Oliver et al. 1981), xylitol (Meyrial, Delgenes et al.
1991), lactic acid (Ohara, Hiyama et al. 1992), and glycerol (Zeng, Ross et al. 1994), can furthermore
act as inhibitors by limiting yeast growth and product formation. Metals released from reactors and
equipment during industrial bioconversion processes have also been found to be inhibitory
(Oleszkiewicz and Sharma 1990). Due to the variation in their fermentation products and inhibitor
tolerance, different microorganisms can yield dissimilar results under the same conditions. Therefore,
the appropriate microorganisms must be selected in each individual case of fermentation, with regard
to the sugars and inhibitors that are present, as well as desired product(s).
1.6 SHF vs. SSF
In SHF, hydrolysis and fermentation are carried out as completely separate steps, whereby enzymes
are added and saccharification is executed to completion, only after which, yeasts are added. The
main advantage of this process is it allows for each step to be performed at its optimum conditions for
temperature and pH. As previously mentioned, cellulase enzymes are maximally active at 50C and a
pH of 4.8, whereas most microbes ferment better at temperatures near 30C and pH near 6.0.
However, a major disadvantage of SHF is that the accumulation of glucose and cellobiose during
hydrolysis can lead to end-product inhibition, as described earlier. To maintain acceptable ethanol
yields, the hydrolysis step in SHF must often be carried out at low solid loadings, resulting in a
relatively dilute ethanol stream. Economically, this is not ideal, as it can increase costs for
downstream processing, namely for distillation (Wingren, Galbe et al. 2003).
In SSF, hydrolysis and fermentation take place concurrently, thereby reducing the possibility of end-
product inhibition. As described previously, ethanol is far less inhibitory to cellulases than are the
sugars released during hydrolysis, 1/16 as inhibitory as cellobiose according to one study (Holtzapple,
Cognata et al. 1990). However, the main disadvantage of SSF is that it takes place under
compromised conditions of temperature (around 37C) and pH (around 5.5), which can have an effect
on total yield.
Generally, studies comparing SSF and SHF have illustrated that determination of the better method is
dependent on a number of factors. Several studies have confirmed that SHF produces higher overall
yields, while SSF requires less time (Alfani, Gallifuoco et al. 2000; Cantarella, Cantarella et al. 2004).
An ideal operating process would be flexible to allow for either method to be used in accordance to
feedstock availability and the desired product(s).

13
1.7 Products from a sugar platform
A number of building blocks for high value bioproducts can be made using the platform of microbial
conversion from sugars. Of the 12 most promising of these reported by the U.S. Department of
Energy, seven can currently be produced using known biological pathways with microorganisms.
The remaining five are presently produced using chemical pathways (Werpy, Petersen et al. 2004).
Four carbon 1,4-diacids, including succinic fumaric, and malic acids, are synthesized through
microbial fermentation via overexpression of Krebs Cycle pathways associated with C4
diacid formation, primarily with Escherichia coli (Millard, Chao et al.). Chemical reduction
of these building blocks produces derivatives that can be used to make solvents, water soluble
polymers, and fibers such as lycra.
3-Hydroxypropionic acid (3-HPA) can be produced via microbial fermentation, though the
pathway is not known, nor is its chemical conversion pathway. Sonora fiber can be derived
from 1,3-propane diol, the product of 3-HPA reduction. Acrylates are formed by 3-HPA
dehydration, which are used to make super absorbent polymers.
Aspartic acid is produced microbially through fermentation or enzymatic action on
oxaloacetate in the Krebs cycle, or chemically from the animation of fumaric acid. Amine
butanediol can be produced through its chemical reduction, aspartic anhydride through its
dehydration, or polyaspartic from its polymerization.
Glutamic acid is made through microbial fermentation of glucose, chiefly by
Corynebacterium glutamicum (Georgi, Rittmann et al. 2005). Reduction of glutamic acid
produces diol, diacid, and aminodiol derivatives, which can be made into monomers for
polyesters and polyamides.
Itaconic acid can be produced through fermentation by aerobic fungi, and is primarily used as
a copolymer and polymer precursor, namely for acrylic or methacrylic acid (Willke and
Vorlop 2001).
Glycerol is produced through the transesterification of oils or from anaerobic microbial
fermentation of sugars (Yazdani and Gonzalez 2007). It can be oxidized to generate glyceric
acid, or directly polymerized to produce polyesters and polyols, utilized in polyurethane
resins. Propylene glycol and 1,3-propanediol can also be produced from glycerol through its
hydrogenolysis.
Sugar alcohols, including xylitol and arabinitol, are made through microbial fermentation of
xylose and arabinose, respectively, though its current primary production through the

14
chemical pathway of hydrogenation of those sugars. These building blocks may be used
directly as sweetners, or oxidized to produce sugar acids (Prakasham, Rao et al. 2009).
Though not produced microbially, levulinic acid can be generated from the degradation of HMF
during pretreatment. It is currently produced through the chemical pathway of acid catalyzed
dehydration of hexose sugars. Levulinic acid can be reduced to make derivatives for fuel oxygenates
and solvents, oxidized to acetyl acrylates or acetic-acrylic succinic acids, used as copolymers. In
addition, condensation of levilinic acid produced diphenolic acid, used in polycarbonate synthesis.
This thesis presents xylitol and ethanol fermentation as examples of biochemical production, but a
wide variety of bioproducts can potentially be made from the sugars available in sulfite pulping
streams.
Chapter 2: Draft paper
Biochemical production from sulfite pulping sugar streams
Abstract
The production of single- and mixed-sugar streams and their conversion to bioproducts were studied
using sulfite pulping streams as feedstocks. Sulfite pulp, sludge, and spent sulfite liquor (SSL) were
utilized because they are concurrently generated alongside of bleached pulp, and because the pulping
process renders pretreatment unnecessary. SSL, comprised of mostly monomeric hexose and pentose
sugars, was directly fermented to ethanol and xylitol with Candida guilliermondii. Single-sugar
streams were generated through hydrolysis of pulp and sludge in buffered water, followed by
fermentation to ethanol with Saccharomyces cerevisiae. Mixed-sugar streams were generated
through both SHF and SSF of pulp and sludge in SSL using S. cerevisiae. Direct fermentation of SSL
to ethanol and xylitol produced yields consistent with that of a synthetic sugar control (89.5%, 40.3%,
respectively). The best utilization of pulp was determined to be as a single-sugar stream, derived from
SHF in water, which yielded a cellulose to ethanol conversion of 62.2% of the theoretical maximum
(28.3 g/L). Sludge produced the highest yield when mixed with SSL during SSF (50.0%, 23.7 g/L).
Sulfite pulp, sludge, and SSL currently represent untapped industrial resources for the production of
single- and mixed-sugar streams from which high-value bioproducts can be made.

15
Introduction
The biochemical industry currently lacks an abundant sugar source from which to make higher value
products. An ideal sugar source must be inexpensive, readily available year-round, and relatively
pure in composition. Using biological conversion, these sugars can be fermented to valuable products
such as ethanol, xylitol, arabitol, succinic acid, and lactic acid (Clark and Deswarte 2008).
Lignocellulosic biomass is a widely abundant sugar source, but requires expensive and energy-
intensive pretreatment, and rarely is consistent in composition. Large scale biomass-to-bioproduct
plants do not currently exist in the US. Coupling biochemical production with an existing industry,
such as sulfite pulping, creates the potential to use sugar streams already being generated by the
pulping process, and simultaneously eliminates the need for pretreatment, as the resultant streams are
mostly delignified.
We explored the potential of converting sulfite pulping streams to sugar sources from which higher
value bioproducts can be made. The three sulfite pulping streams examined in this study were pulp,
primary clarifier sludge, and spent sulfite liquor (SSL). The hydrolysability and fermentability to
ethanol of Kraft mill sludges has already been demonstrated by Sjöde et al. 2007 and Kang et al.
2010. The fermentability of SSL to ethanol using Rhizopus oryzae and Saccharomyces cerevisiae,
respectively, has been exemplified by Taherzadeh et al. 1997, and Helle et al. 2008. However, aside
from ethanol, these materials can alternatively be converted to sugar streams, from which a host of
biochemical can be produced using the model of a sugar platform (Clark and Deswarte 2008).
Without bioconversion, sludge and SSL can require costly treatment techniques prior to disposal or
burning, so their conversion to higher value products is both desirable and economically beneficial.
Pulp is itself a highly valued product, so its conversion to biochemicals is less economically desirable.
However, conversion to a very high value bioproduct may be desirable in the future if there is an
excess of sulfite pulp on the market.
In addition, reconfiguration of the traditional bioconversion scheme using these three pulping streams
has not been well studied. The objective of this paper is to examine the potential of generating single
and mixed sugars streams from sulfite pulp, sludge, and SSL for bioproduct conversion via SHF and
SSF. We explored high consistency, low enzyme loading SHF of each stream separately, but we also
analyzed SHF and SSF of combined feedstocks, SSL fortified with either of the two solid streams.
This offers the potential to produce highly concentrated sugar streams without using large quantities
of enzyme, as would be the case in traditional, single-feedstock enzymatic hydrolysis. Ethanol and

16
xylitol were produced to demonstrate examples of biological conversion, but many other bioproducts
are possible.
Methods
Raw materials
Ammonia-based sulfite pulping at Kimberly-Clark in Everett, WA produces woodpulp for direct sale
or conversion to tissue products. Byproducts include primary clarifier sludge, and spent sulfite
liquor. Pulp is produced at a nominal rate of rate of 500 air dry metric tons per day (admt/d). Under
the right conditions, up to 90 admt/d could be provided for hydrolysis (Sande 2010). Current practice
at Kimberly-Clark is to dry, press, and store pulp when it is not being used for paper production.
Sludge is produced at a rate of 45 dry short tons/day, and SSL at 500 short tons dry solids/admt pulp
at 14% solids (Sande 2010). Sludge is dewatered and burned as hog fuel onsite, and SSL is
evaporated and burned to recover SO2 and heat. Collaborators at Kimberly-Clark provided us with
the pulp, sludge and SSL used in this study.
All materials were derived from primarily softwoods. The mill also produces a hardwood grade, so
pulp and sludge contained a small, unknown amount of hardwood fiber. Pulp was collected from the
mill’s pre-bleach washers and had not been treated with ClO2. Sludge was collected from primary
clarifiers before introduction to aerobic bacteria, and contained a mixture of pulping fines and rejects,
tissue mill sludge, and boiler house effluent. SSL was taken directly from brown stock washers at
14% solids and had not been evaporated. Solids were washed with ten times their mass in water, and
stored at -20°C until use. Moisture content was 77.3% for pulp, and 75% for sludge. SSL was stored
at 4°C.
Compositional analysis
Insoluble carbohydrates and lignin
TAPPI method T-222 om-98 (TAPPI 1998) was used to gravimetrically analyze insoluble lignin, and
photometrically analyze soluble lignin. Carbohydrate content was measured using HPLC. Dried
samples of 0.2 g were ground to 40-mesh size and combined with 3.0 mL of 72% (w/w) H2SO4 for 2
h. Samples were then diluted to 4% (w/w) H2SO4, autocalved at 121°C for 1 h, and filtered through
glass fritted crucibles. Filtrate was collected, carbohydrate content analyzed by HPLC, and acid-

17
insoluble lignin content calculated by measuring UV at 205 nm. Oven-dried crucibles were weighed
to determine acid insoluble lignin content.
Soluble carbohydrates
Soluble monomeric and oligomeric carbohydrate content was determined using NREL LAP TP-510-
42623 (Sluiter, Hames et al. 2004). Five mL SSL was added to 0.697 mL of 72% (w/w) H2SO4 and
filled to a 20 mL total volume with water. Samples were autoclaved at 121°C for 1 h and analyzed by
HPLC to determine total sugar content. Monomeric sugars were analyzed on the raw SSL, and
oligomeric sugar was calculated as the difference between total and monomeric sugar content.
Oligomeric standards containing a range of arabinose, galactose, glucose, xylose, and mannose
concentrations were treated in the same manner as samples, and a sugar degradation factor was
applied to oligomeric sugar calculations.
Figure 2. Experimental design of converting pulp, sludge, and SSL to bioproducts
via fermentation, SHF, and SSF. In SHF, solid streams are combined with either
water or SSL. In SSF, solid streams are combined with SSL and converted in one
step.
Hydro
lysis
SS
F
Fe
rme
nta
tio
n
Fe
rme
nta
tio
n
Pulp or Sludge SSL
Bioproducts (e.g. ethanol, xylitol)
add to
SSL H2O or SSL
Sugars

18
Enzymatic hydrolysis and fermentation
We explored process designs by which pulp, sludge, and SSL could be converted to a sugar platform
from which biochemicals could be produced. The resulting schematic of converting pulp and sludge
to ethanol, and SSL to ethanol and xylitol, is shown in Figure 2. Separate hydrolysis and
fermentation (SHF) was used to convert the three streams separately, and simultaneous
saccharification and fermentation (SSF) was used to convert SSL fortified with each of the two solid
streams in one step. SSL was fermented to ethanol and xylitol.
Saccharification
Hydrolysis was carried out in 125 mL (50 mL reaction volume) Erlenmeyer flasks in triplicate on
washed solid materials. Solids were enzymatically hydrolyzed at 10% (w/v) consistency in both
water and SSL with pH adjusted to 4.8. Flasks were incubated at 50°C and 150 rpm in an orbital
shaker (New Brunswick). Enzymes added were cellulase at 5 FPU/g cellulose (Spezyme, Genencor,
Palo Alto, CA) and β-glucosidase at 10 CBU/g cellulose (Novozymes 188, Bagsverd, Denmark). For
controls, the same amount of enzyme added to pulp and sludge flasks was respectively added to flasks
containing plain SSL. Samples of 1 mL volume were taken periodically over 48 h, boiled at 100°C
for 5 min to denature enzymes, and stored at -20°C until HPLC analysis.
Fermentation (Saccharomyces cerevisiae)
Prior to fermentation, Saccharomyces cerevisiae (ATCC 96581) isolated from spent sulfite liquor
(Linden, Peetre et al. 1992) was streaked onto YPD agar plates and allowed to grow for 48 h. Sterile
liquid media containing 10 g/L each of glucose, yeast extract, and peptone was inoculated with one
colony from the plate. Cells were grown for a total of 48 h at 30°C and 150 rpm in an orbital shaker,
with fresh media replaced at 24 h. Cells were then spun down, washed twice in water, and
resuspended in a small volume of 0.5% NaCl. Cell concentration was determined by comparing
optical density of the cell suspension at 600 nm to a calibration curve.
After completion of hydrolysis, the remaining liquid hydrolysate was boiled at 100°C to denature
enzymes and vacuum filtered through filter paper. The resulting filtrate was collected and nutrients
were added in the form of (NH4)2HPO4 at 2 g/L, Na2SO4 at 0.2 g/L, and NaNO3 at 2 g/L. Hydrolyzed
SSL, and a solution of 10 g/L each of glucose, galactose, and mannose in water were also treated in
this manner to use as controls. The pH was adjusted to 6.0 with 50% w/v NaOH and S. cerevisiae

19
was added at a concentration of 5 g/L. Fermentation was done in 50 mL volume in 125 mL
Erlenmeyer flasks, incubated at 30°C and 150 rpm for 72 h in an orbital shaker. Samples of 1 mL
volume were taken periodically and centrifuged at 10,000 rpm for 5 min. Supernatant was collected
through 0.22 μm syringe filters and stored at -20°C, while pellets were washed and optical density
measured at 600 nm to determine cell concentration.
Fermentation (Candida guilliermondii)
Fermentation was also performed on hydrolyzed SSL spiked with synthetic xylose up to a 30 g/L total
concentration using C. guilliermondii (ATCC 201935) to demonstrate xylitol production. This was
done in an identical manner as fermentation with S. cerevisiae except that the nutrients added in this
case were 5 g/L urea, 1.7 g/L yeast nitrogen base, and 1 g/L yeast extract. Two controls containing
30 g/L each of glucose and xylose were handled in the same manner.
SSF
SSL containing the same nutrient concentrations as in SHF was adjusted to pH 5.5 with 50% w/v
NaOH. The solution was fortified with pulp or sludge at 10% consistency, and enzyme and S.
cerevisiae were added at 5 FPU/g cellulose (10 CBU/g β-glucosidase) and 5 g/L, respectively.
Samples were run in triplicate in 100 mL volume at 37°C in an orbital shaker for 72 h. A control
containing 10 g/L each of galactose, glucose, and mannose was prepared and analyzed identically.
Analysis of sugars and ethanol
Monosaccharides were quantified using a Dionex (Sunnyvale, CA) HPLC (ICS-3000) system
equipped with an AS50 autosampler, ED50 electrochemical detector, GP50 gradient pump, and anion
exchange column (Dionex, CarboPac PA1). Deionized water at 1 mL/min was used as an eluent,
with a postcolumn addition of 0.2 M NaOH. The injection volume was 5 μL. Standards were
prepared containing arabinose, galactose, glucose, xylose, and mannose in an appropriate range of
concentrations that fully encompassed the range found respectively in the samples. Samples were
filtered through 0.22 µm syringe filters and an internal standard, fucose, was added at 0.2 g/L.
Ethanol, xylitol, acetic acid, furfural, and hydroxymethylfurfural (HMF) were measured using a
Shimadzu (Columbia, MD) HPLC system equipped with a RID-10A differential refractometric
detector, LC-20AD solvent delivery module, SIL-20AHT autosampler, and a Rezex RHM
monosaccharide H+ anion exchange column (Phenomenex, Torrance, CA). An isocratic mobile
20
phase of 5 μM H2SO4 was used at a flow rate of 0.6 mL/min, and a 20 μL injection volume was used
for each sample. Standards of ethanol, xylitol, acetic acid, furfural, and HMF were prepared in
appropriate concentrations. Samples were analyzed by relating to standard curves generated by the
prepared standards of known concentration. Calculation of standard curves and standard deviation
was done in Microsoft Excel.
Results and discussion
Chemical composition of pulp, sludge, and SSL
The chemical composition of pulp and sludge in terms of polysaccharides, lignin, and ash is shown in
Table 1. Both solid streams were mainly comprised of glucan (pulp 88.6%, 70.2%), contained a
small amount of acid insoluble lignin (pulp 3.3%, sludge 12.3%), and very minimal acid soluble
lignin (pulp 1.6%, sludge 0.7% (Table 1). Pulp contained more glucan and less total lignin than
sludge, an expected outcome of the pulping process. Mannan was the next most abundant
polysaccharide (pulp 6.6%, sludge 4.4%) for both streams due to their derivation from mostly
softwoods. As expected, neither stream contained a detectable amount of arabinan or galactan. At
9.9%, sludge contained more ash than pulp, at 0.2%, a result of being partially sourced from boiler
house effluent. It has been observed that similar fiber sludges, derived from Kraft pulping, and
containing as little as 32.3% glucan and as much as 24% total lignin, can still be hydrolyzed into
monomeric sugars for ethanol production (Sjöde, Alriksson et al. 2007).
Table 1. Percent composition of solid streams based on TAPPI method T-222 om-98. Acid insoluble
lignin is shown as AIL, and acid soluble lignin is ASL.
Glucan (%) Xylan (%) Mannan (%) AIL (%) ASL (%) Ash (%)
Pulp 88.6 2.8 6.6 3.3 1.6 0.2
Sludge 70.2 3.1 4.4 12.3 0.7 9.9
The monomeric and oligomeric sugar composition of SSL is shown in Table 2. Due to softwood
hemicellulose being mainly comprised of galactoglucomannans and Arabino-4-O-
methylglucuronoxylans (Fengel and Wegener 1989), the most abundant sugar was mannose (14.0 g/L
monomers), followed by xylose (7.6 g/L monomers). Glucose (5.1 g/L) was the next most abundant
sugar in SSL, followed by minimal amounts of galactose (3.4 g/L) and arabinose (1.2 g/L). All five
sugars were mostly in monomeric form (83%), making SSL a readily-available monosaccharide
21
source. Acetic acid, hydroxymethylfurfural (HMF) and furfurals were also present in SSL at
concentrations of 5.0, 0.11 and 0.16 g/L, respectively (Table 2). An unevaporated SSL stream
containing 31.3 g/L total sugar was used for this study, but this can be concentrated onsite up to 97
g/L total sugar if a higher sugar concentration is desired (Sande 2010).
Table 2. Chemical composition of SSL and oligomer % of total based on NREL LAP TP-510-42623
(Sluiter, Hames et al. 2004). *Acetic acid 5 g/L, HMF 0.11 g/L, furfural 0.16 g/L.
Ara (g/L) Gal (g/L) Glu (g/L) Xyl (g/L) Man (g/L) Total 5-C
sugars (g/L)
Total 6-C
sugars (g/L)
Monomer 1.2 3.4 5.1 7.6 14.0 8.8 22.5
Oligomer 0.11 0.40 0.92 0.04 3.86 0.15 5.18
Monomer
% of total
91.5 98.6 84.7 99.5 78.4 98.4 81.3
Due to its high glucan content and minimal lignin and ash, sulfite pulp, if successfully hydrolyzed,
can be expected to produce a clean, single-sugar stream (glucose). Sludge may produce similar
results, though its higher lignin content is expected to reduce hydrolysis yields as compared to pulp.
SSL, having a higher concentration of both hexoses and pentoses, is already an excellent source of
mixed sugars, most of which are in monomeric form.
Fermentation of SSL
As the high concentration of monomeric sugars in SSL has been demonstrated through its
compositional analysis, we next utilized this mixed sugar stream for fermentation to test its potential
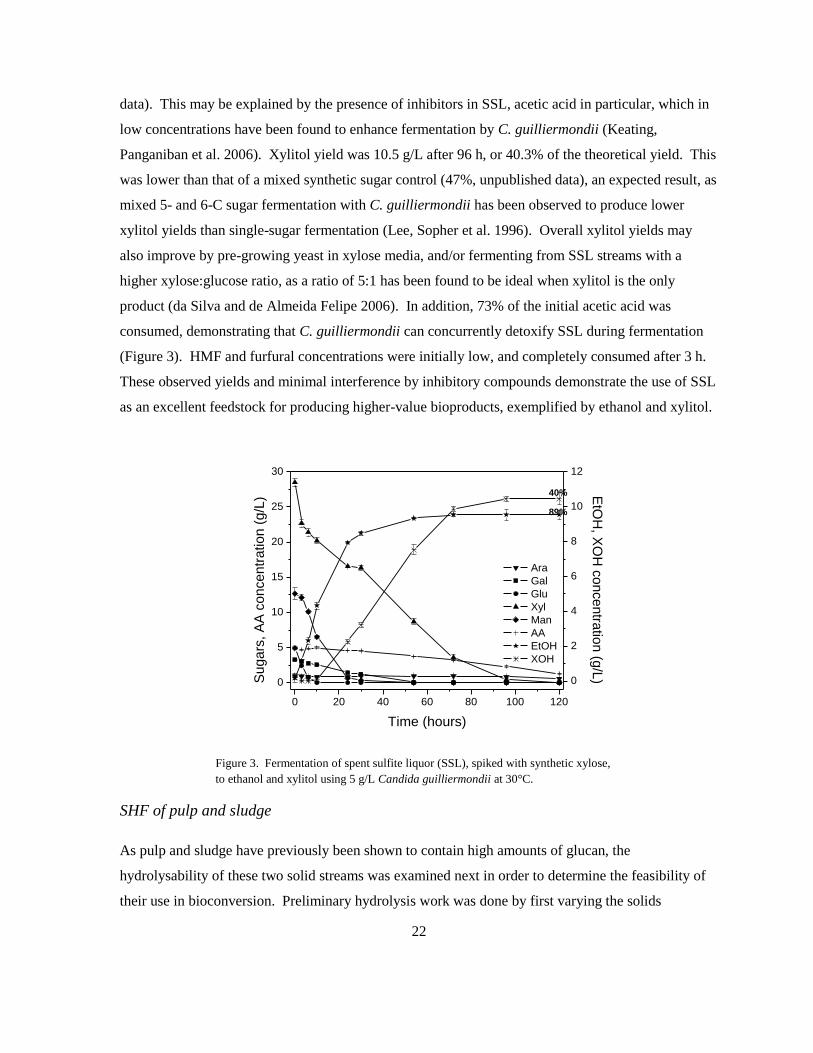
conversion to bioproducts. Figure 3 shows fermentation of SSL utilizing C. guilliermondii, which
ferments hexoses to ethanol and xylose to xylitol. Due to the low concentration of native xylose (7.6
g/L), SSL was spiked with synthetic sugar to demonstrate the full potential of xylose to xylitol
fermentation. Glucose was consumed after 21 h, followed mannose and galactose at 33 h. This
pattern of consumption was consistent with that of a Tembec yeast strain and S. cerevisiae during
mixed synthetic sugar fermentation (Keating, Robinson et al. 2004). Total consumption of
approximately 30 g/L xylose required 120 h. Ethanol and xylitol were produced concurrently.
Ethanol concentration reached 9.5 g/L after 72h, corresponding to 89.5% of the theoretical yield.
This was higher than the ethanol yield achieved with a mixture of synthetic sugars (77%, unpublished
22
data). This may be explained by the presence of inhibitors in SSL, acetic acid in particular, which in
low concentrations have been found to enhance fermentation by C. guilliermondii (Keating,
Panganiban et al. 2006). Xylitol yield was 10.5 g/L after 96 h, or 40.3% of the theoretical yield. This
was lower than that of a mixed synthetic sugar control (47%, unpublished data), an expected result, as
mixed 5- and 6-C sugar fermentation with C. guilliermondii has been observed to produce lower
xylitol yields than single-sugar fermentation (Lee, Sopher et al. 1996). Overall xylitol yields may
also improve by pre-growing yeast in xylose media, and/or fermenting from SSL streams with a
higher xylose:glucose ratio, as a ratio of 5:1 has been found to be ideal when xylitol is the only
product (da Silva and de Almeida Felipe 2006). In addition, 73% of the initial acetic acid was
consumed, demonstrating that C. guilliermondii can concurrently detoxify SSL during fermentation
(Figure 3). HMF and furfural concentrations were initially low, and completely consumed after 3 h.
These observed yields and minimal interference by inhibitory compounds demonstrate the use of SSL
as an excellent feedstock for producing higher-value bioproducts, exemplified by ethanol and xylitol.
0 20 40 60 80 100 120
0
5
10
15
20
25
30
Ara
Gal
Glu
Xyl
Man
AA
EtOH
XOH
Time (hours)
Sugars
, A
A c
oncentr
ation (
g/L
)
0
2
4
6
8
10
12
EtO
H, X
OH
concentra
tion (g
/L)
Figure 3. Fermentation of spent sulfite liquor (SSL), spiked with synthetic xylose,
to ethanol and xylitol using 5 g/L Candida guilliermondii at 30°C.
SHF of pulp and sludge
As pulp and sludge have previously been shown to contain high amounts of glucan, the
hydrolysability of these two solid streams was examined next in order to determine the feasibility of
their use in bioconversion. Preliminary hydrolysis work was done by first varying the solids
89%
40%
23
consistency, then the enzyme loading, to determine the most desirable conditions for SHF, based on
yield. Solids were enzymatically hydrolyzed at 2%, 10%, and 15% (w/v) consistency and 10 FPU/g
cellulose enzyme loading. In both cases, hydrolysis at 10% consistency yielded high glucose
concentrations (87 g/L for pulp, 62 g/L for sludge, not shown) after 24 h. Next, enzymes loadings of
1, 2, 5, 10, and 15 FPU/g cellulose were used to hydrolyze 2% (w/v) consistency slurries of each
solid. In this case, 5 FPU/g cellulose resulted in reasonably high glucose yields (18 g/L for pulp, 14
g/L for sludge) after 24 h without sacrificing enzyme cost. Therefore, SHF experiments were all
performed at 10% (w/v) solids consistency and 5 FPU/g cellulose enzyme loading.
0 20 40 60 80 100
0
10
20
30
40
50
60
70
80
Cellu
lose to g
lucose c
onvers
ion (
%)
Time (hours)
Pulp / H2O
Pulp / SSL
Sludge / H2O
Sludge / SSL
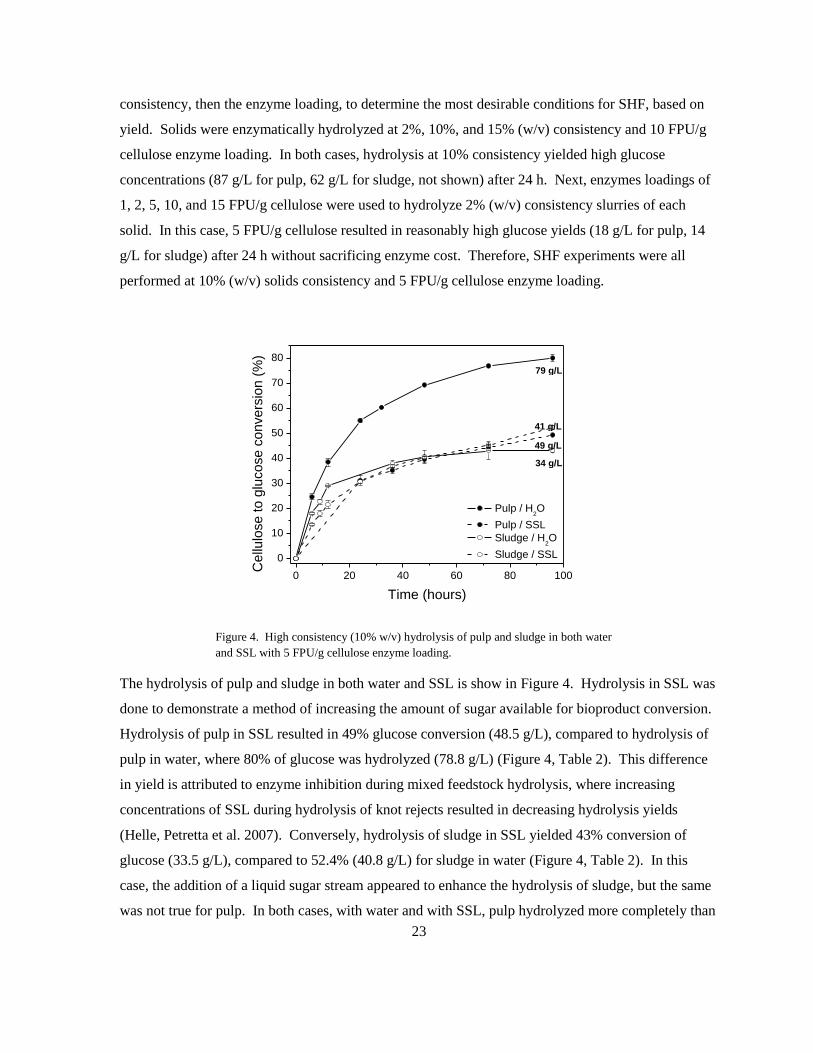
Figure 4. High consistency (10% w/v) hydrolysis of pulp and sludge in both water
and SSL with 5 FPU/g cellulose enzyme loading.
The hydrolysis of pulp and sludge in both water and SSL is show in Figure 4. Hydrolysis in SSL was
done to demonstrate a method of increasing the amount of sugar available for bioproduct conversion.
Hydrolysis of pulp in SSL resulted in 49% glucose conversion (48.5 g/L), compared to hydrolysis of
pulp in water, where 80% of glucose was hydrolyzed (78.8 g/L) (Figure 4, Table 2). This difference
in yield is attributed to enzyme inhibition during mixed feedstock hydrolysis, where increasing
concentrations of SSL during hydrolysis of knot rejects resulted in decreasing hydrolysis yields
(Helle, Petretta et al. 2007). Conversely, hydrolysis of sludge in SSL yielded 43% conversion of
glucose (33.5 g/L), compared to 52.4% (40.8 g/L) for sludge in water (Figure 4, Table 2). In this
case, the addition of a liquid sugar stream appeared to enhance the hydrolysis of sludge, but the same
was not true for pulp. In both cases, with water and with SSL, pulp hydrolyzed more completely than
79 g/L
34 g/L 49 g/L
41 g/L
24
did sludge, an expected result due to the higher lignin and ash content of sludge, which has been
found to inhibit hydrolysis enzymes (Kang, Wang et al. 2010). Sludge hydrolysis was enhanced by
the addition of SSL, however, pulp hydrolyzed more completely in water.
0 5 10 15 20
0
10
20
30
40
50
60
70
Pulp / H2O Glucose
Pulp / SSL Hexose
Pulp / H2O EtOH
Pulp / SSL EtOH
Time (hours)
Sugar
concentr
ation (
g/L
)
0
10
20
30
EtO
H c
oncentra
tion (g
/L)
Figure 5. Fermentation of pulp hydrolysates using 5 g/L Saccharomyces cerevisiae
at 30°C.
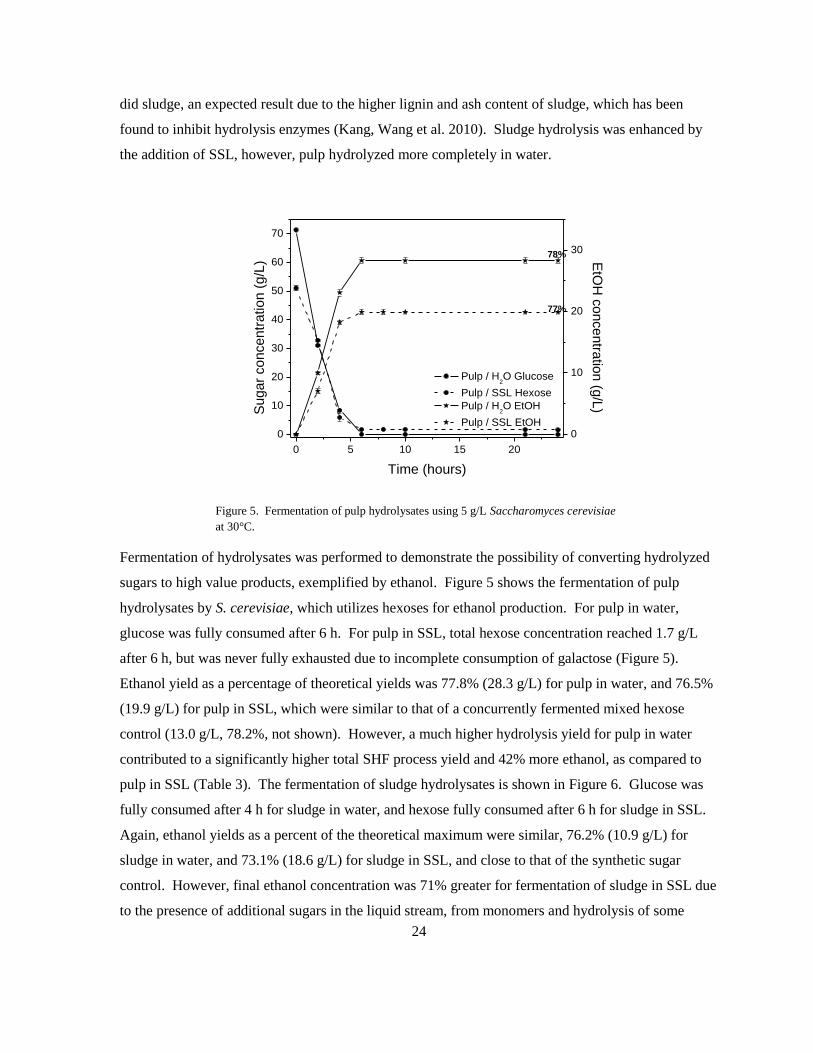
Fermentation of hydrolysates was performed to demonstrate the possibility of converting hydrolyzed
sugars to high value products, exemplified by ethanol. Figure 5 shows the fermentation of pulp
hydrolysates by S. cerevisiae, which utilizes hexoses for ethanol production. For pulp in water,
glucose was fully consumed after 6 h. For pulp in SSL, total hexose concentration reached 1.7 g/L
after 6 h, but was never fully exhausted due to incomplete consumption of galactose (Figure 5).
Ethanol yield as a percentage of theoretical yields was 77.8% (28.3 g/L) for pulp in water, and 76.5%
(19.9 g/L) for pulp in SSL, which were similar to that of a concurrently fermented mixed hexose
control (13.0 g/L, 78.2%, not shown). However, a much higher hydrolysis yield for pulp in water
contributed to a significantly higher total SHF process yield and 42% more ethanol, as compared to
pulp in SSL (Table 3). The fermentation of sludge hydrolysates is shown in Figure 6. Glucose was
fully consumed after 4 h for sludge in water, and hexose fully consumed after 6 h for sludge in SSL.
Again, ethanol yields as a percent of the theoretical maximum were similar, 76.2% (10.9 g/L) for
sludge in water, and 73.1% (18.6 g/L) for sludge in SSL, and close to that of the synthetic sugar
control. However, final ethanol concentration was 71% greater for fermentation of sludge in SSL due
to the presence of additional sugars in the liquid stream, from monomers and hydrolysis of some
78%
77%
25
oligomers present in SSL. Fermentation yields were consistent between all four hydrolysates and the
hexose control, demonstrating that fermentation did not account for differences in final ethanol
concentration. Instead, ethanol yield is more likely to be improved by (1) improving cellulose to
glucose conversion during hydrolysis, and (2) increasing the initial availability of sugars by, in some
cases, mixing solid and liquid sugar streams.
0 5 10 15 20
0
10
20
30
40
50
Sludge / H2O Glucose
Sludge / SSL Hexose
Sludge / H2O EtOH
Sludge / SSL EtOH
Time (hours)
Sugar
concentr
ation (
g/L
)
0
5
10
15
20
EtO
H c
oncentra
tion (g
/L)
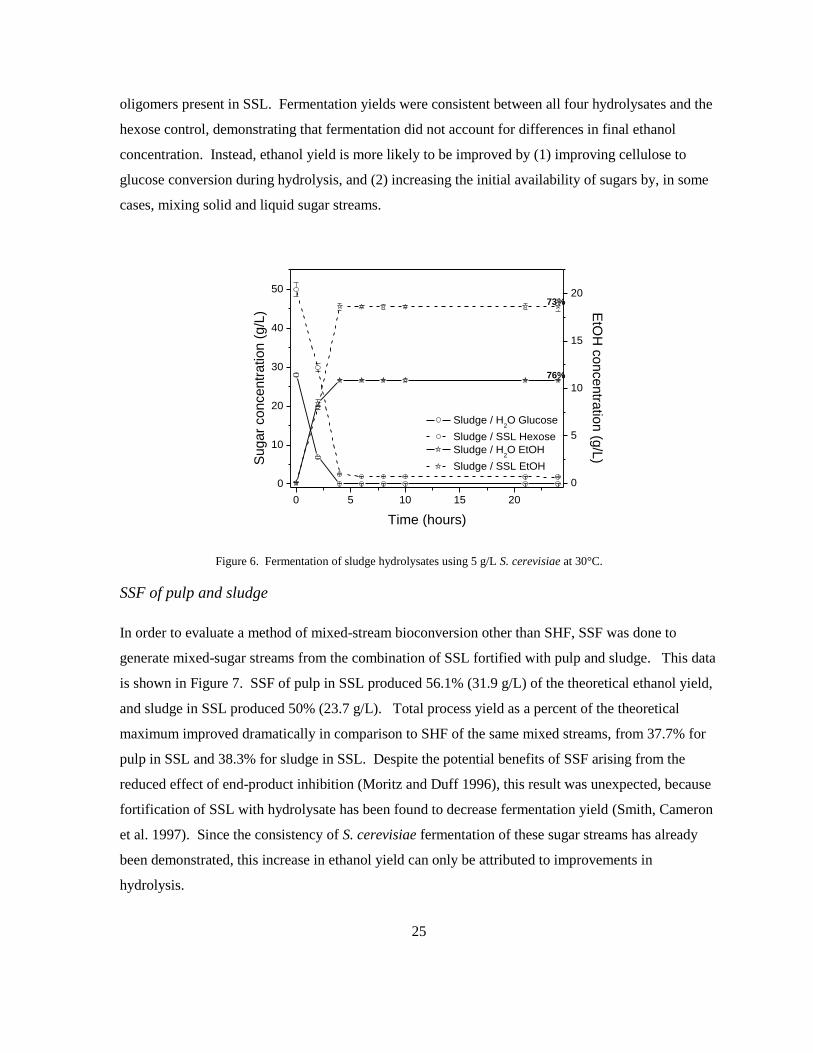
Figure 6. Fermentation of sludge hydrolysates using 5 g/L S. cerevisiae at 30°C.
SSF of pulp and sludge
In order to evaluate a method of mixed-stream bioconversion other than SHF, SSF was done to
generate mixed-sugar streams from the combination of SSL fortified with pulp and sludge. This data
is shown in Figure 7. SSF of pulp in SSL produced 56.1% (31.9 g/L) of the theoretical ethanol yield,
and sludge in SSL produced 50% (23.7 g/L). Total process yield as a percent of the theoretical
maximum improved dramatically in comparison to SHF of the same mixed streams, from 37.7% for
pulp in SSL and 38.3% for sludge in SSL. Despite the potential benefits of SSF arising from the
reduced effect of end-product inhibition (Moritz and Duff 1996), this result was unexpected, because
fortification of SSL with hydrolysate has been found to decrease fermentation yield (Smith, Cameron
et al. 1997). Since the consistency of S. cerevisiae fermentation of these sugar streams has already
been demonstrated, this increase in ethanol yield can only be attributed to improvements in
hydrolysis.
73%
76%
26
0 10 20 30 40 50 60 70
0
2
4
6
8
10
12
14
Pulp / SSL Hexoses
Sludge / SSL Hexoses
Pulp / SSL EtOH
Sludge / SSL EtOH
Time (hours)
Sugar
concentr
ation (
g/L
)
0
5
10
15
20
25
30
35
EtO
H c
oncentra
tion (g
/L)
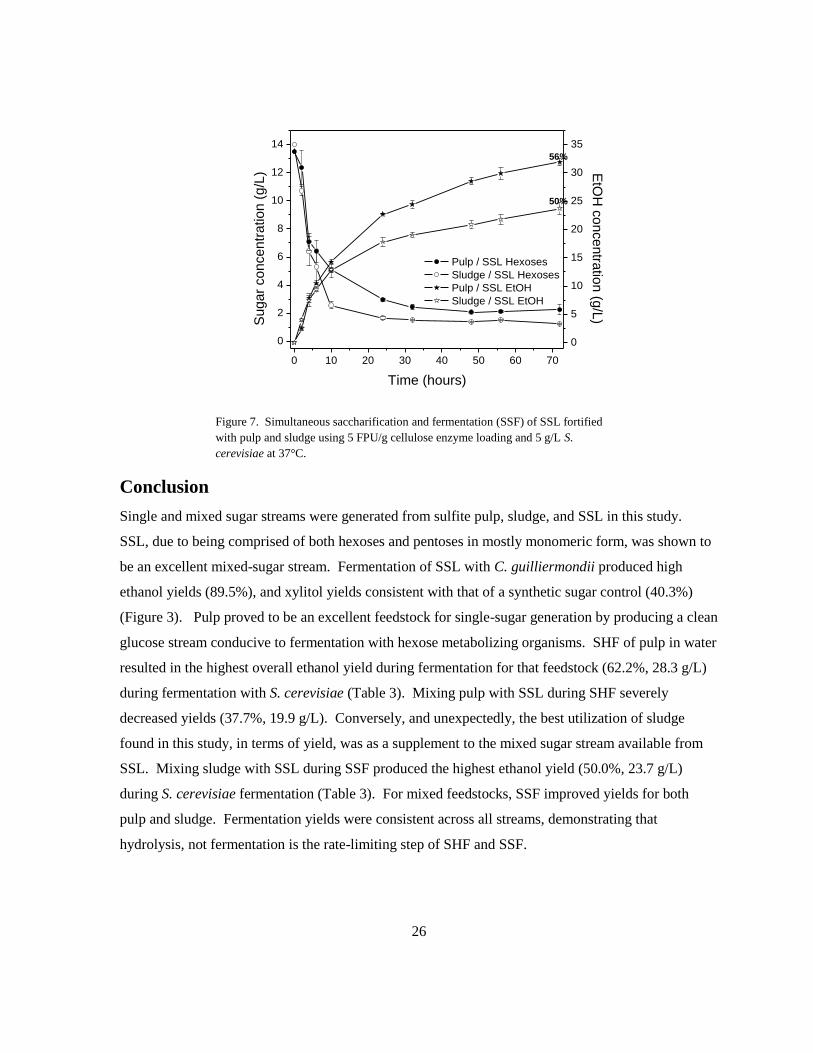
Figure 7. Simultaneous saccharification and fermentation (SSF) of SSL fortified
with pulp and sludge using 5 FPU/g cellulose enzyme loading and 5 g/L S.
cerevisiae at 37°C.
Conclusion
Single and mixed sugar streams were generated from sulfite pulp, sludge, and SSL in this study.
SSL, due to being comprised of both hexoses and pentoses in mostly monomeric form, was shown to
be an excellent mixed-sugar stream. Fermentation of SSL with C. guilliermondii produced high
ethanol yields (89.5%), and xylitol yields consistent with that of a synthetic sugar control (40.3%)
(Figure 3). Pulp proved to be an excellent feedstock for single-sugar generation by producing a clean
glucose stream conducive to fermentation with hexose metabolizing organisms. SHF of pulp in water
resulted in the highest overall ethanol yield during fermentation for that feedstock (62.2%, 28.3 g/L)
during fermentation with S. cerevisiae (Table 3). Mixing pulp with SSL during SHF severely
decreased yields (37.7%, 19.9 g/L). Conversely, and unexpectedly, the best utilization of sludge
found in this study, in terms of yield, was as a supplement to the mixed sugar stream available from
SSL. Mixing sludge with SSL during SSF produced the highest ethanol yield (50.0%, 23.7 g/L)
during S. cerevisiae fermentation (Table 3). For mixed feedstocks, SSF improved yields for both
pulp and sludge. Fermentation yields were consistent across all streams, demonstrating that
hydrolysis, not fermentation is the rate-limiting step of SHF and SSF.
56%
50%
27
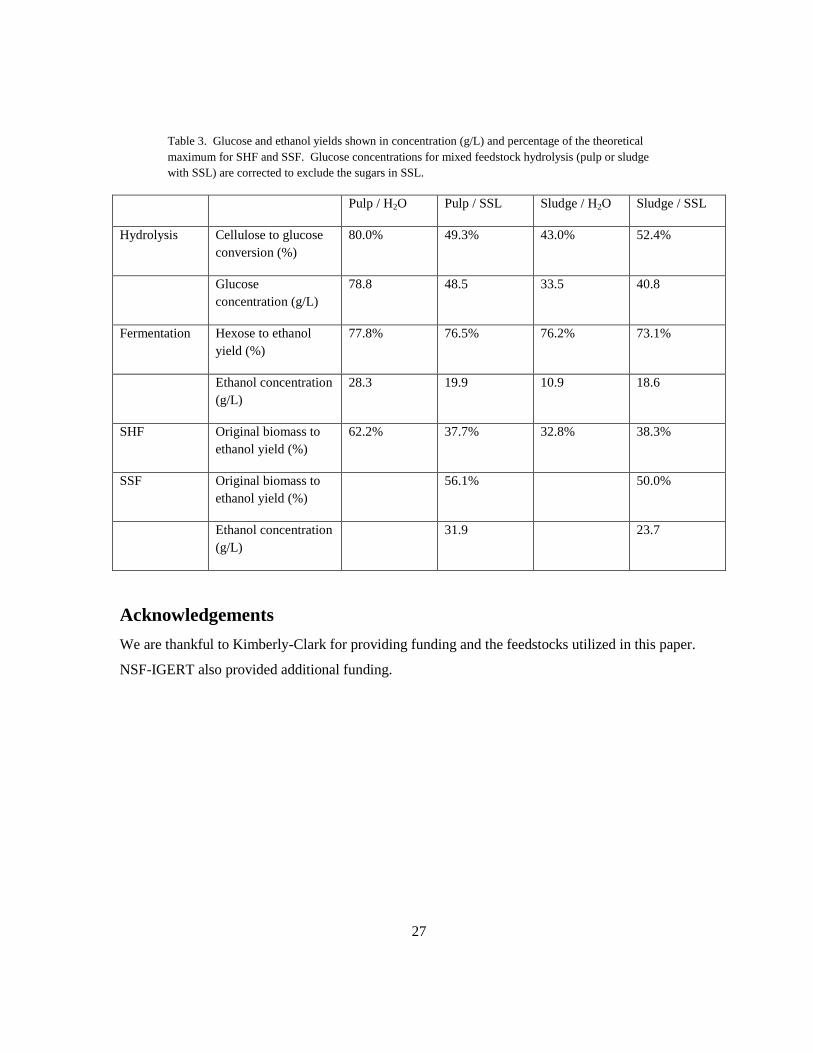
Table 3. Glucose and ethanol yields shown in concentration (g/L) and percentage of the theoretical
maximum for SHF and SSF. Glucose concentrations for mixed feedstock hydrolysis (pulp or sludge
with SSL) are corrected to exclude the sugars in SSL.
Pulp / H2O Pulp / SSL Sludge / H2O Sludge / SSL
Hydrolysis Cellulose to glucose
conversion (%)
80.0% 49.3% 43.0% 52.4%
Glucose
concentration (g/L)
78.8 48.5 33.5 40.8
Fermentation Hexose to ethanol
yield (%)
77.8% 76.5% 76.2% 73.1%
Ethanol concentration
(g/L)
28.3 19.9 10.9 18.6
SHF Original biomass to
ethanol yield (%)
62.2% 37.7% 32.8% 38.3%
SSF Original biomass to
ethanol yield (%)
56.1% 50.0%
Ethanol concentration
(g/L)
31.9 23.7
Acknowledgements
We are thankful to Kimberly-Clark for providing funding and the feedstocks utilized in this paper.
NSF-IGERT also provided additional funding.
28
Chapter 3: Additional results and discussion
3.1 Fermentation of SSL using S. cerevisiae
0 5 10 15 20 25 30
0
2
4
6
8
10
12
Time (hours)
Concentr
ation (
g/L
)
Gal
Glu
Man
Yeast
Gly
AA
EtOH
0
2
4
6
8
EtO
H c
oncentra
tion (g
/L)
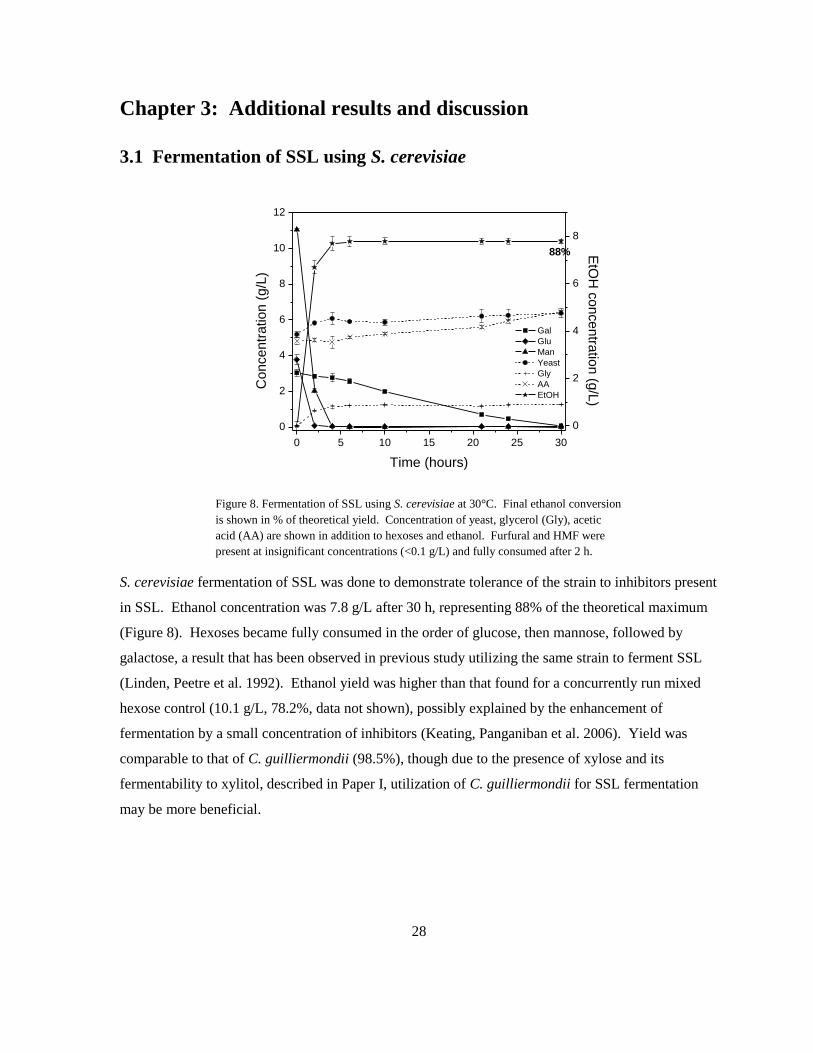
Figure 8. Fermentation of SSL using S. cerevisiae at 30°C. Final ethanol conversion
is shown in % of theoretical yield. Concentration of yeast, glycerol (Gly), acetic
acid (AA) are shown in addition to hexoses and ethanol. Furfural and HMF were
present at insignificant concentrations (<0.1 g/L) and fully consumed after 2 h.
S. cerevisiae fermentation of SSL was done to demonstrate tolerance of the strain to inhibitors present
in SSL. Ethanol concentration was 7.8 g/L after 30 h, representing 88% of the theoretical maximum
(Figure 8). Hexoses became fully consumed in the order of glucose, then mannose, followed by
galactose, a result that has been observed in previous study utilizing the same strain to ferment SSL
(Linden, Peetre et al. 1992). Ethanol yield was higher than that found for a concurrently run mixed
hexose control (10.1 g/L, 78.2%, data not shown), possibly explained by the enhancement of
fermentation by a small concentration of inhibitors (Keating, Panganiban et al. 2006). Yield was
comparable to that of C. guilliermondii (98.5%), though due to the presence of xylose and its
fermentability to xylitol, described in Paper I, utilization of C. guilliermondii for SSL fermentation
may be more beneficial.
88%
29
3.2 Effect of solids loading on enzymatic hydrolysis
(a)
0 10 20 30 40 50
0
20
40
60
80
100
Cellu
lose to g
lucose c
onvers
ion (
%)
Time (hours)
Pulp 2%
Pulp 10%
Pulp 15%
(b)
0 10 20 30 40 50
0
20
40
60
80
100
Cellu
lose to g
lucose c
onvers
ion (
%)
Time (hours)
Sludge 2%
Sludge 10%
Sludge 15%
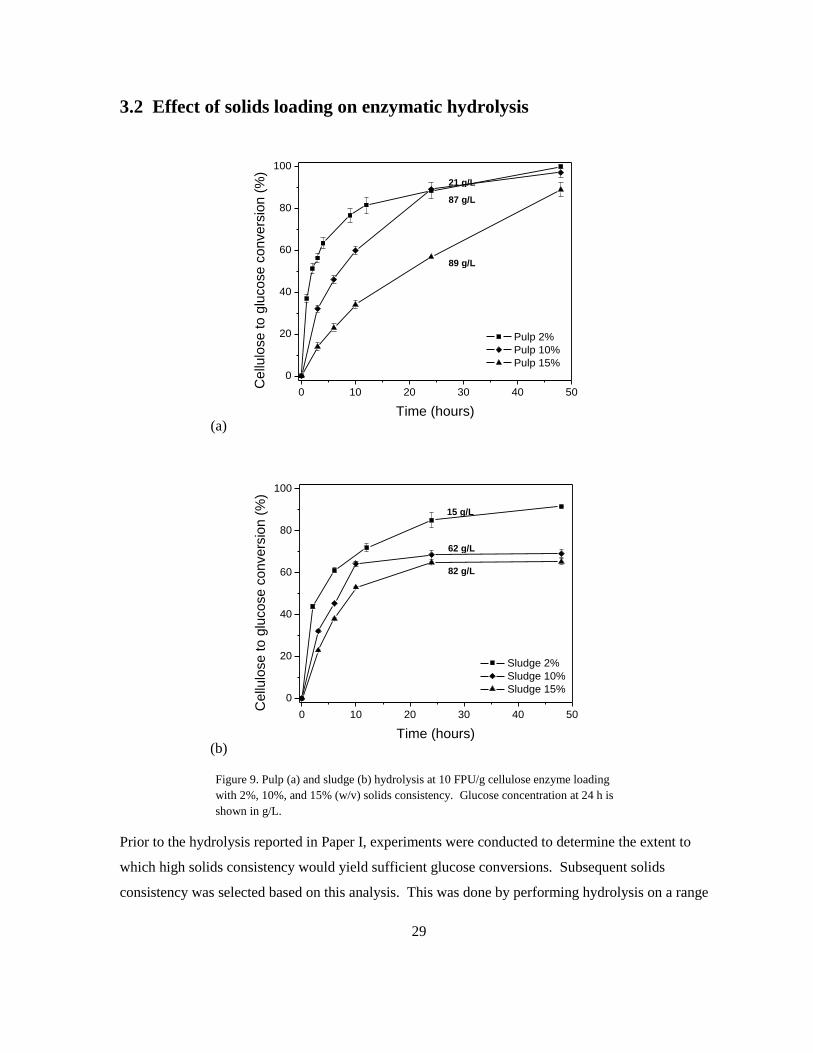
Figure 9. Pulp (a) and sludge (b) hydrolysis at 10 FPU/g cellulose enzyme loading
with 2%, 10%, and 15% (w/v) solids consistency. Glucose concentration at 24 h is
shown in g/L.
Prior to the hydrolysis reported in Paper I, experiments were conducted to determine the extent to
which high solids consistency would yield sufficient glucose conversions. Subsequent solids
consistency was selected based on this analysis. This was done by performing hydrolysis on a range
21 g/L
87 g/L
89 g/L
15 g/L
62 g/L
82 g/L

30
of solids loadings (2-15% w/v) while keeping enzyme loading at 10 FPU/g cellulose (Figure 9).
Though carried out for 48 h, a short turnover time is favorable in an industrial setting, so conversion
at 24 h was examined to determine the desired solids loading for all subsequent SHF and SSF. As
expected, percent glucose conversion decreased with increased solids loading for both pulp and
sludge. For pulp, percent glucose conversion did not differ between 2% and 10% consistency (both
89%), but dropped significantly to 57% when consistency was increased to 15%. For sludge, percent
conversion between 2% and 10% consistency at 24 h differed by a greater margin (83% and 68%,
respectively), however this also corresponded to a desirable increase in glucose concentration from
14.4 g/L to 61.4 g/L. High consistency hydrolysis has previously been described as a desirable way
of reducing cost, because it produces a highly concentrated sugar streams, and therefore results in a
more concentrated product stream. Taking into account both % glucose conversion and sugar
concentration in the final hydrolysate, a solid consistency of 10% was selected for subsequent the
analyses presented in Paper I.
3.3 Effect of enzyme loading on hydrolysis
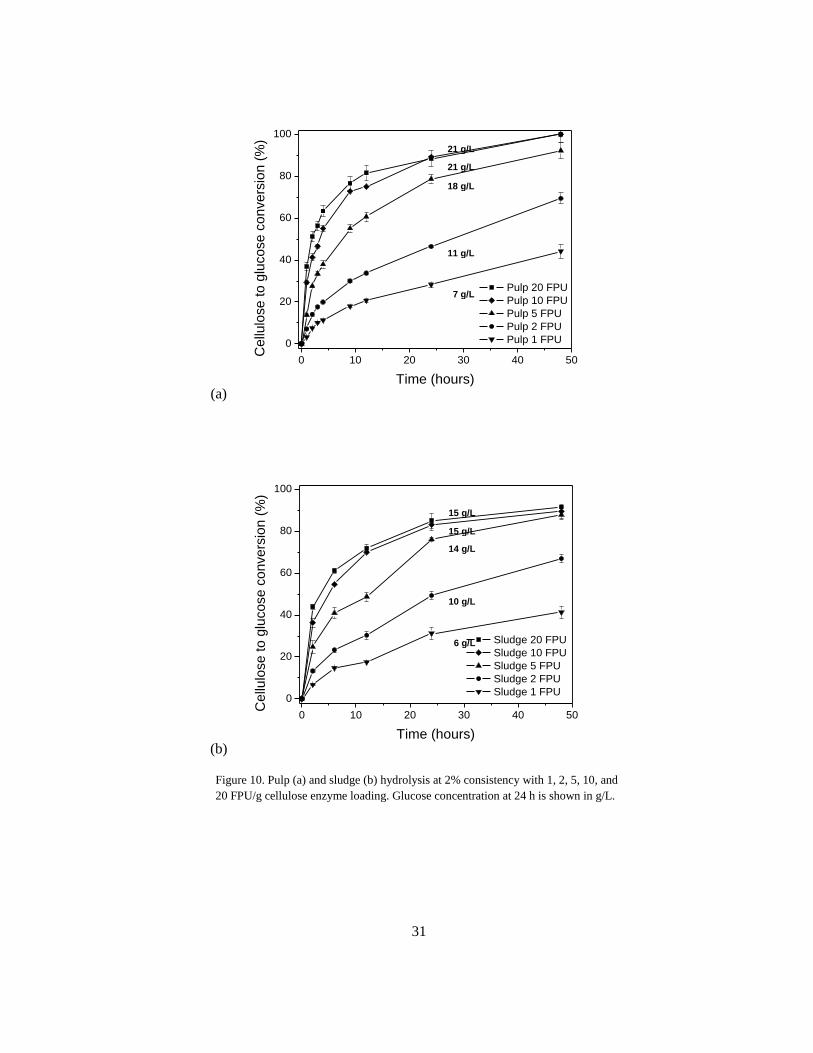
Enzyme loading was also selected based on analyses that took place prior to those presented in Paper
I. The effect of enzyme loading on hydrolysis was explored by hydrolyzing pulp and sludge at 2%
consistency using a gradient of enzyme loading of 1-20 FPU/g cellulose (Figure 10). As predicted,
increased enzyme loading had a positive, nonlinear effect on percent glucose conversion. For pulp,
doubling the amount of enzyme from 5 to 10 FPU/g cellulose resulted in only a 10% gain in glucose
conversion (79% to 89%). Comparatively, reducing to 2 FPU/g cellulose enzyme loading resulted in
a significant drop in percent conversion (46%). Similar results were found for sludge, where glucose
conversion at 2, 5, and 10 FPU/g cellulose for sludge was 49%, 76%, and 83%, respectively after 24 h
(Fig. 3b). As previously described, reducing enzyme loading to the extent that glucose conversion is
not significantly effected is a favorable industrial practice that reduces cost. Based on this analysis,
an enzyme loading of 5 FPU/g cellulose was selected for subsequent SHF and SSF.
31
(a)
0 10 20 30 40 50
0
20
40
60
80
100
Cellu
lose to g
lucose c
onvers
ion (
%)
Time (hours)
Pulp 20 FPU
Pulp 10 FPU
Pulp 5 FPU
Pulp 2 FPU
Pulp 1 FPU
(b)
0 10 20 30 40 50
0
20
40
60
80
100
Cellu
lose to g
lucose c
onvers
ion (
%)
Time (hours)
Sludge 20 FPU
Sludge 10 FPU
Sludge 5 FPU
Sludge 2 FPU
Sludge 1 FPU
Figure 10. Pulp (a) and sludge (b) hydrolysis at 2% consistency with 1, 2, 5, 10, and
20 FPU/g cellulose enzyme loading. Glucose concentration at 24 h is shown in g/L.
21 g/L
21 g/L
18 g/L
11 g/L
7 g/L
15 g/L
15 g/L
14 g/L
10 g/L
6 g/L

32
Chapter 4: Additional conclusions and future work
The production of single and mixed sugar streams from pulp, sludge, and SSL has been demonstrated
in this thesis and the enclosed draft paper. Fermentation of SSL with S. cerevisiae produced ethanol
yields equivalent to that of C. guilliermondii, presented in the draft paper (88%, and 89.5%,
respectively). However, the prospect of exploiting xylose, present in SSL at 5.1 g/L, makes the use of
C. guilliermondii or a similar pentose-metabolizing microorganism a more appealing option due to
the potential of making two products from SSL. From the hydrolysis results presented in Chapter 3,
10% (w/v) consistency and 5 FPU/g cellulose enzyme loading was determined to be the most
desirable parameters for the high consistency, low enzyme loading hydrolysis results presented in the
draft paper. However, this determination was based on laboratory-scale yield alone, and the
economic viability of these conditions for a full-scale industrial operation must be evaluated prior to
implementation. For the purposes of this thesis and draft paper, these hydrolysis parameters were
sufficient.
From the draft paper presented in Chapter 2, it was concluded that SSL is a readily-available source
of mixed 5- and 6-C sugars, which are mostly in monomeric form. Pulp proved to a be desirable
feedstock for a single sugar stream (glucose) due to its 88.6% glucan content, 80% of which was
hydrolyzed in water. Mixing of pulp with SSL yielded low cellulose to ethanol conversions for both
SHF (37.7%), and SSF (56.1%). Sludge hydrolyzed more completely when combined with SSL
during SSF, yielding 50% total cellulose to ethanol conversion.
Future work
The objective of generating single- and mixed-sugar streams from sulfite pulp, sludge, and SSL has
been achieved in this thesis. However, the full potential of bioproduct conversion during the
fermentation step has not been entirely explored. First, growth of yeast on SSL prior to fermentation
can be explored as a means of better adapting the yeast to inhibitory compounds within SSL. In
addition to the potential of boosting yields, this reduces the cost spent on synthetic sugars for the
propagation of yeast. Secondly, production of other biochemicals via fermentation with a wider
variety of sugar-metabolizing microorganisms can be explored. On an industrial scale, flexibility in
the choice of microorganism would allow the bioconversion process to adjust according to the market
value of the variety of bioproducts that can be made. Finally, techno-economic and life-cycle
analyses of the proposed bioconversion processes of pulping streams to bioproducts would benefit
decisions regarding industrial-scale operation. In particular, comparison of the processes utilized here

33
with dedicated lignocellulose-to-ethanol facilities would be valuable. Continued research along these
lines would allow the biochemicals industry to tap into a currently unused source of sugars for
bioproduct conversion.
Acknowledgements
This thesis is the result of 2 ½ years of work in the School of Forest Resources at the University of
Washington. I give my sincere thanks to my advisor and committee chair, Renata Bura, and
committee members Rick Gustafson and Bill McKean for their guidance through the duration of my
research and thesis writing. I would also like to thank Doug Asbe, Wally Sande, and Jeff Ross at
Kimberly-Clark for their collaboration on this project and for providing the feedstocks. I’m also
grateful to NSF-IGERT for providing funding through the duration of my graduate work.

34
References
Abatzoglou, N., E. Chornet, et al. (1992). "Phenomenological kinetics of complex systems: the
development of a generalized severity parameter and its application to lignocellulosics
fractionation." Chemical Engineering Science 47(5): 1109-1122.
Akin, D. E. (2007). Grass Lignocellulose. Applied Biochemistry and Biotecnology. J. R. Mielenz, K.
T. Klasson, W. S. Adney and J. D. McMillan, Humana Press: 3-15.
Alfani, F., A. Gallifuoco, et al. (2000). "Comparison of SHF and SSF processes for the bioconversion
of steam-exploded wheat straw." Journal of Industrial Microbiology & Biotechnology
25(4): 184-192.
Alizadeh, H., F. Teymouri, et al. (2005). "Pretreatment of switchgrass by ammonia fiber explosion
(AFEX)." Applied Biochemistry and Biotechnology 124(1): 1133-1141.
Atalla, R. H. and D. L. VanderHart (1999). "The role of solid state 13C NMR spectroscopy in studies
of the nature of native celluloses." Solid State Nuclear Magnetic Resonance 15(1): 1-19.
Banerjee, N., R. Bhatnagar, et al. (1981). "Development of resistance in Saccharomyces cerevisiae
against inhibitory effects of Browning reaction products." Enzyme and Microbial Technology
3(1): 24-28.
Barbosa, M. F. S., M. B. de Medeiros, et al. (1988). "Screening of yeasts for production of xylitol
fromd-xylose and some factors which affect xylitol yield in Candida guilliermondii." Journal
of Industrial Microbiology & Biotechnology 3(4): 241-251.
Bergeron, P. (1996). Environmental impacts of bioethanol. Handbook on ethanol: Production and
utilization. C. E. Wyman, Taylor & Francis: 90-93.
Blanchette, R. A. (1991). "Delignification by Wood-Decay Fungi." Annual Review of
Phytopathology 29(1): 381-403.
Brown, R. M. (2004). "Cellulose structure and biosynthesis: What is in store for the 21st century?"
Journal of Polymer Science Part A: Polymer Chemistry 42(3): 487-495.
Brown, S. W., S. G. Oliver, et al. (1981). "Ethanol inhibition of yeast growth and fermentation:
Differences in the magnitude and complexity of the effect." Applied Microbiology and
Biotechnology 11(3): 151-155.
Canadell, J. G., C. Le Quéré, et al. (2007). "Contributions to accelerating atmospheric CO2 growth
from economic activity, carbon intensity, and efficiency of natural sinks." Proceedings of the
National Academy of Sciences 104(47): 18866-18870.

35
Cantarella, M., L. Cantarella, et al. (2004). "Comparison of different detoxification methods for
steam-exploded poplar wood as a substrate for the bioproduction of ethanol in SHF and SSF."
Process Biochemistry 39(11): 1533-1542.
Cara, C., M. Moya, et al. (2007). "Influence of solid loading on enzymatic hydrolysis of steam
exploded or liquid hot water pretreated olive tree biomass." Process Biochemistry 42(6):
1003-1009.
Clark, J. and F. Deswarte, Eds. (2008). Introduction to chemicals from biomass. Renewable
resources. West Sussex, Wiley.
da Silva, D. D. V. and M. d. G. de Almeida Felipe (2006). "Effect of glucose:xylose ratio on xylose
reductase and xylitol dehydrogenase activities from Candida guilliermondii in sugarcane
bagasse hydrolysate." Journal of Chemical Technology & Biotechnology 81(7): 1294-1300.
Decker, S., W. Adney, et al. (2003). "Automated filter paper assay for determination of cellulase
activity." Applied Biochemistry and Biotechnology 107(1): 689-703.
Delgenes, J. P., R. Moletta, et al. (1996). "Effects of lignocellulose degradation products on ethanol
fermentations of glucose and xylose by Saccharomyces cerevisiae, Zymomonas mobilis,
Pichia stipitis, and Candida shehatae." Enzyme and Microbial Technology 19(3): 220-225.
DeLong, E. A. (1977). Method of rendering lignin separable from cellulose and hemicellulose in
lignocellulosic material and the product so produced. C. P. a. D. Limited. Canada.
Ding, S.-Y. and M. E. Himmel (2006). "The Maize Primary Cell Wall Microfibril: A New Model
Derived from Direct Visualization." Journal of Agricultural and Food Chemistry 54(3): 597-
606.
Ding, S.-Y. and M. E. Himmel (2008). Anatomy and ultrastructure of maize cell walls: An example
of energy plants. Biomass recalcitrance: Deconstructing the plant cell wall for bioenergy. M.
E. Himmel, Wiley-Balckwell: 38-60.
Eklund, R., M. Galbe, et al. (1995). "The influence of SO2 and H2SO4 impregnation of willow prior
to steam pretreatment." Bioresource Technology 52(3): 225-229.
Eriksson, T., J. Karlsson, et al. (2002). "A model explaining declining rate in hydrolysis of
lignocellulose substrates with cellobiohydrolase I (Cel7A) and endoglucanase I (Cel7B) of
Trichoderma reesei." Applied Biochemistry and Biotechnology 101(1): 41-60.
Fan, L., Y.-H. Lee, et al. (1982). The nature of lignocellulosics and their pretreatments for enzymatic
hydrolysis. Microbial Reactions, Springer Berlin / Heidelberg. 23: 157-187.
Felby, C., L. Thygesen, et al. (2008). "Cellulose–water interactions during enzymatic hydrolysis as
studied by time domain NMR." Cellulose 15(5): 703-710.

36
Fengel, D. and G. Wegener (1989). Wood: Chemistry, ultrastructure, reactions. Berlin, New York,
Walter de Gruyter.
Galbe, M. and G. Zacchi (2002). "A review of the production of ethanol from softwood." Applied
Microbiology and Biotechnology 59(6): 618-628.
Gellerstedt, G. (1973). The reactions of lignin during sulphite pulping. Stockholm.
Georgi, T., D. Rittmann, et al. (2005). "Lysine and glutamate production by Corynebacterium
glutamicum on glucose, fructose and sucrose: Roles of malic enzyme and fructose-1,6-
bisphosphatase." Metabolic Engineering 7(4): 291-301.
Ghose, T. K. (1987). "Measurement of cellulase ativities." Pure & Applied Chemistry 59(2): 257-268.
Glennie, D. W. (1971). Reactions in sulfite pulping. Lignins: Occurrence, formation, structure, and
reactions. K. V. Sarkanen and C. H. Ludwig. New York, Wiley: 597-637.
Gnansounou, E. and A. Dauriat (2005). Energy balance of bioethanol: a synthesis.
Gregg, D. J. and J. N. Saddler (1996). "Factors affecting cellulose hydrolysis and the potential of
enzyme recycle to enhance the efficiency of an integrated wood to ethanol process."
Biotechnology and Bioengineering 51(4): 375-383.
Hassinger, W., G. Sauer, et al. (1981). "The effects of equal caloric amounts of xylitol, sucrose and
starch on insulin requirements and blood glucose levels in insulin-dependent diabetics."
Diabetologia 21(1): 37-40.
Helle, S. S., R. A. Petretta, et al. (2007). "Fortifying spent sulfite pulping liquor with hydrolyzed
reject knots." Enzyme and Microbial Technology 41(1-2): 44-50.
Holtzapple, M., M. Cognata, et al. (1990). "Inhibition of Trichoderma reesei cellulase by sugars and
solvents." Biotechnology and Bioengineering 36(3): 275-287.
Hsu, T.-A. (1996). Pretreatment of biomass. Handbook on bioethanol: Production and utilization. C.
E. Wyman. Washington, Taylor & Francis: 179-212.
Jarvis, M. (2003). "Chemistry: Cellulose stacks up." Nature 426: 611-612.
Jørgensen, H., J. Vibe-Pedersen, et al. (2007). "Liquefaction of lignocellulose at high-solids
concentrations." Biotechnology and Bioengineering 96(5): 862-870.
Kai, Y. (1991). Chemistry of extractives. Wood and cellulosic chemistry. D. N.-S. Hon and N.
Shiraishi, CRC Press.
Kang, L., W. Wang, et al. (2010). "Bioconversion of Kraft Paper Mill Sludges to Ethanol by SSF and
SSCF." Applied Biochemistry and Biotechnology 161(1): 53-66.
Keating, J. D., C. Panganiban, et al. (2006). "Tolerance and adaptation of ethanologenic yeasts to
lignocellulosic inhibitory compounds." Biotechnology and Bioengineering 93(6): 1196-1206.

37
Keating, J. D., J. Robinson, et al. (2004). "An ethanologenic yeast exhibiting unusual metabolism in
the fermentation of lignocellulosic hexose sugars." Journal of Industrial Microbiology &
Biotechnology 31(5): 235-244.
Keeney, D. R. and T. H. DeLuca (1992). "Biomass as an energy source for the midwestern U.S."
American Journal of Alternative Agriculture 7(03): 137-144.
Klemm, D., B. Heublein, et al. (2005). "Cellulose: Fascinating Biopolymer and Sustainable Raw
Material." Angewandte Chemie International Edition 44(22): 3358-3393.
Lee, H., C. R. Sopher, et al. (1996). "Induction of xylose reductase and xylitol dehydrogenase
activities on mixed sugars in Candida guilliermondii." Journal of Chemical Technology &
Biotechnology 65(4): 375-379.
Linden, T., J. Peetre, et al. (1992). "Isolation and characterization of acetic acid-tolerant galactose-
fermenting strains of Saccharomyces cerevisiae from a spent sulfite liquor fermentation
plant." Appl. Environ. Microbiol. 58(5): 1661-1669.
Lora, J. H. and S. Aziz (1985). "Organosolv pulping: a versatile approach to wood refining." Journal
Name: Tappi; (United States); Journal Volume: 68:8: Medium: X; Size: Pages: 94-97.
Lorenz, D., D. Morris, et al. (1995). How much energy does it take to make a gallon of ethanol?
Minneapolis, MN, Institute for Local Self-Reliance.
Mason, W. H. (1933). Process and apparatus for disintegration of material. United States Patent,
MASONITE CORP.
Meyrial, V., J. P. Delgenes, et al. (1991). "Xylitol production from D-xylose by<i>Candida
guillermondii</i>: Fermentation behaviour." Biotechnology Letters 13(4): 281-286.
Millard, C. S., Y. P. Chao, et al. Enhanced production of succinic acid by overexpression of
phosphoenolpyruvate carboxylase in Escherichia coli.
Moritz, J. and S. Duff (1996). "Ethanol production from spent sulfite liquor fortified by hydrolysis of
pulp mill primary clarifier sludge." Applied Biochemistry and Biotechnology 57-58(1): 689-
698.
Mosier, N., C. Wyman, et al. (2005). "Features of promising technologies for pretreatment of
lignocellulosic biomass." Bioresource Technology 96(6): 673-686.
Nguyen, Q. A. and J. N. Saddler (1991). "An integrated model for the technical and economic
evaluation of an enzymatic biomass conversion process." Bioresource Technology 35(3):
275-282.
Ohara, H., K. Hiyama, et al. (1992). "Non-competitive product inhibition in lactic acid fermentation
from glucose." Applied Microbiology and Biotechnology 36(6): 773-776.

38
Ohlsson, K. A. (2004). "Carbonation of wood ash recycled to a forest soil as measured by isotope
ratio mass spectrometry." Soil Science Society of America Journal 64(6): 2155-2161.
Oleszkiewicz, J. A. and V. K. Sharma (1990). "Stimulation and inhibition of anaerobic processes by
heavy metals--A review." Biological Wastes 31(1): 45-67.
Palmqvist, E. and B. Hahn-Hägerdal (2000). "Fermentation of lignocellulosic hydrolysates. II:
inhibitors and mechanisms of inhibition." Bioresource Technology 74(1): 25-33.
Pampulha, M. E. and M. C. Loureiro-Dias (1990). "Activity of glycolytic enzymes of Saccharomyces
cerevisiae in the presence of acetic acid." Applied Microbiology and Biotechnology 34(3):
375-380.
Pan, X. (2008). "Role of Functional Groups in Lignin Inhibition of Enzymatic Hydrolysis of
Cellulose to Glucose." Journal of Biobased Materials and Bioenergy 2: 25-32.
Pimentel, D. (2003). "Ethanol Fuels: Energy Balance, Economics, and Environmental Impacts Are
Negative." Natural Resources Research 12(2): 127-134.
Pointing, S. B., A. L. Pelling, et al. (2005). "Screening of basidiomycetes and xylariaceous fungi for
lignin peroxidase and laccase gene-specific sequences." Mycological Research 109(1): 115-
124.
Prakasham, R. S., R. S. Rao, et al. (2009). "Current trends in biotechnological production of xylitol
and future prospects." Current Trends in Biotechnology and Pharmacy 3(1): 8-36.
Ramos, L., C. Breuil, et al. (1992). "Comparison of steam pretreatment of eucalyptus, aspen, and
spruce wood chips and their enzymatic hydrolysis." Applied Biochemistry and Biotechnology
34-35(1): 37-48.
Ramos, L. P., C. Breuil, et al. (1993). "The use of enzyme recycling and the influence of sugar
accumulation on cellulose hydrolysis by Trichoderma cellulases." Enzyme and microbial
technology. 15(1): 19.
Sanchez, B. and J. Bautista (1988). "Effects of furfural and 5-hydroxymethylfurfural on the
fermentation of Saccharomyces cerevisiae and biomass production from Candida
guilliermondii." Enzyme and Microbial Technology 10(5): 315-318.
Sande, W. (2010). Personal communication. L. Lai. Everett, WA.
Sedlak, M. and N. Ho (2004). "Production of ethanol from cellulosic biomass hydrolysates using
genetically engineered <i>saccharomyces</i> yeast capable of cofermenting
glucose and xylose." Applied Biochemistry and Biotechnology 114(1): 403-416.

39
Shen, J. and F. A. Agblevor (2008). "Optimization of enzyme loading and hydrolytic time in the
hydrolysis of mixtures of cotton gin waste and recycled paper sludge for the maximum profit
rate." Biochemical Engineering Journal 41(3): 241-250.
Sjöde, A., B. Alriksson, et al. (2007). The Potential in Bioethanol Production From Waste Fiber
Sludges in Pulp Mill-Based Biorefineries. Applied Biochemistry and Biotecnology. J. R.
Mielenz, K. T. Klasson, W. S. Adney and J. D. McMillan, Humana Press: 327-337.
Sluiter, A., B. Hames, et al. (2004). Determination of sugars, byproducts, and degradation products in
liquid fraction process samples. Golden, CO, NREL. NREL/TP-510-42623.
Smith, M. T., D. R. Cameron, et al. (1997). "Comparison of industrial yeast strains for fermentation
of spent sulphite pulping liquor fortified with wood hydrolysate." Journal of Industrial
Microbiology & Biotechnology 18(1): 18-21.
Taherzadeh, M. J., C. Niklasson, et al. (1997). "Acetic acid--friend or foe in anaerobic batch
conversion of glucose to ethanol by Saccharomyces cerevisiae?" Chemical Engineering
Science 52(15): 2653-2659.
Taniguchi, M., H. Suzuki, et al. (2005). "Evaluation of pretreatment with Pleurotus ostreatus for
enzymatic hydrolysis of rice straw." Journal of Bioscience and Bioengineering 100(6): 637-
643.
TAPPI (1998). TAPPI standard methods, T222 om-98. TAPPI Journal, TAPPI Press.
Tengborg, C., M. Galbe, et al. (2001). "Influence of enzyme loading and physical parameteres on the
enzymatic hydrolysis of steam-pretreated softwood." Biotechnology Progress 17(1): 110-117.
Tyson, K. S. (1993). Fuel cycle evaluations of biomass-ethanol and reformulated gasoline. Volume 1.
Other Information: PBD: Nov 1993: Medium: ED; Size: 130 p.
Väljamäe, P., K. Kipper, et al. (2003). "Synergistic cellulose hydrolysis can be described in terms of
fractal-like kinetics." Biotechnology and Bioengineering 84(2): 254-257.
von Sivers, M. and G. Zacchi (1995). "A techno-economical comparison of three processes for the
production of ethanol from pine." Bioresource Technology 51(1): 43-52.
Werpy, T., G. Petersen, et al. (2004). Top value added chemicals from biomass, Volume I: Results
from screening for potential candidates from sugars and synthesis gas. T. Werpy and G.
Petersen, NREL, Pacific Northwest National Laboratory.
Willke, T. and K. D. Vorlop (2001). "Biotechnological production of itaconic acid." Applied
Microbiology and Biotechnology 56(3): 289-295.

40
Wingren, A., M. Galbe, et al. (2003). "Techno-Economic Evaluation of Producing Ethanol from
Softwood: Comparison of SSF and SHF and Identification of Bottlenecks." Biotechnology
Progress 19(4): 1109-1117.
Wiselogel, A., S. Tyson, et al. (1996). Biomass feedstock resources and composition. Handbook on
bioethanol: Production and utilization. C. E. Wyman, Taylor & Francis: 106-117.
Xiao, Z., X. Zhang, et al. (2004). "Effects of Sugar Inhibition on Cellulases and β-Glucosidase
During Enzymatic Hydrolysis of Softwood Substrates." Applied Biochemistry and
Biotechnology 115(1/3): 1115-1126.
Yazdani, S. S. and R. Gonzalez (2007). "Anaerobic fermentation of glycerol: a path to economic
viability for the biofuels industry." Current Opinion in Biotechnology 18(3): 213-219.
Yu, J., J. Zhang, et al. (2009). "Combinations of mild physical or chemical pretreatment with
biological pretreatment for enzymatic hydrolysis of rice hull." Bioresource Technology
100(2): 903-908.
Zeng, A. P., A. Ross, et al. (1994). "Multiple product inhibition and growth modeling of clostridium
butyricum and klebsiella pneumoniae in glycerol fermentation." Biotechnology and
Bioengineering 44(8): 902-911.
Zhang, X., W. Qin, et al. (2009). "High consistency enzymatic hydrolysis of hardwood substrates."
Bioresource Technology 100(23): 5890-5897.