biosynthesis of chondroitin sulfate proteoglycan
TRANSCRIPT

THE JOUHNAL OF BIOLOGICAL CHEMISTRY Vol. 247, No. lZ,Iw~e ofJune25, PP.~W-3847,1972
Printed in U.S.A.
Biosynthesis of Chondroitin Sulfate Proteoglycan
XYLOSYL TRANSPKR TO SMITH-DEGRADED CARTILAGE PROTEOGLYCAN AND OTHER EXOGE- NOUS ACCEPTOR.S*
(Received for publication, November 8, 1971)
JOHN R. BAKER,~ LENNART ROD~N, AND ALLEN C. STOOLMILLER
Prom the Departments of Pediatrics and Biochemistry, La Rabida-University of Chicago In.stitute, the Joseph P. Kelrnedy, J,r., Mental Retardation Center, University of Chicago, Chicago, Illinois 60&V’, and the Department of Biochemistry, l’rinity College, Dublin 2, Ireland
SUMMARY
The chain-initiating reaction in chondroitin sulfate synthe- sis is catalyzed by a xylosyltransferase, which transfers xylose from UDP-xylose to a polypeptide acceptor. The enzyme has previously been detected in several cell-free tissue preparations, which also contain endogenous xylose acceptors. However, reliable quantitation of enzyme levels has not been possible, since such crude systems contain two unknown dependent variables, enzyme concentration and acceptor concentration. The purpose of the present work was (a) to develop exogenous acceptors which would permit accurate determination of enzyme levels and (b) at the same time gain some insight into the substrate specificity of the enzyme.
Many small molecular weight compounds were tested as potential acceptors, including free serine, serine derivatives, and serine-containing peptides. One of the peptides had some acceptor activity, i.e. the tripeptide, Ser-Gly-Gly, al- though it had a rather high K, value of 20.0 mu. Consider- ably higher acceptor activity was observed with peptides derived from the chondroitin sulfate-protein linkage region by Smith degradation of a chondroitin sulfate peptidoglycan preparation, which had been isolated after trypsin digestion of the proteoglycan.
Smith degradation of the entire proteoglycan molecule yielded the best acceptor obtained so far: this material had a K, of 0.064 m&I, expressed in terms of concentration of serine residues. The intact proteoglycan was not an acceptor, nor were the products obtained by digestion with testicular hyaluronidase or bacterial chondroitinase, al- though about half of the serine residues in these three com- pounds are unsubstituted.
Characterization of the [14C]xylosyl-labeled Smith-de- graded proteoglycan showed that (Q) xylose had been in- corporated into the protein in an alkali-labile linkage from which it could be released as xylitol on treatment with alkaline borohydride, and that (b) exhaustive proteolysis
*This work wm supported by United States Public Health Service Grants AM-05996 and HD-04583, and by a grant from the American Heart, Association. Preliminary reports have appeared (1, 2).
$ Recipient of an Intern;tt.ional Postdoctoral Fellowship, ~-FOCI- TW-1401, from the IJnited States Public Health Service.
yielded a compound with electrophoretic and chromatographic properties of xylosyIserine. It is therefore concluded that xylose had been transferred to the serine hydroxyl groups which had been liberated by Smith degradation. Further- more, the failure of the intact proteoglycan and the enzymatic digestion products to serve as acceptors suggests that glycosylation occurs specifically at certain serine sites of the proteoglycan molecule.
In several connective tissue proteoglycans the polysaccharide chains are linked to protein by glycosidic bonds between xylose and the hydroxyl groups of serine residues (3). Transfer of xylose from UDP-xylose to a polypeptide acceptor initiates the biosynthesis of the polysaccharide chains, and this reaction has been shown in cell-free preparations from hen’s oviduct (4), mouse mastocytoma (5), and embryonic chick cartilage (6). These systems contain endogenous acceptors which are probably identical either with the completed protein cores of the pro- teoglycans or with the growing peptide chains.
Exogenous substrates for xylosyltransferase have not been previously available, and it has not been possible to quantitate enzyme activity reliably, since this requires an assay which is independent of Ohe concentration of endogenous acceptors. The present study was undertaken in order to develop such an assay with substrates of well defined nature, and, at the same time, gain some insight into the substrate specificity of the xylosyltransferase reaction. A number of peptides and proteins were t,ested as potential xylose acceptors in the embryonic chick cartilage system described by Robinson et al. (6). Among small peptides of known structure, one acceptor was found, i.e. t.he tripeptide, Ser-Gly-Gly. However, a far better acceptor w&s obtained by Smith degradation of the proteoglycan from bovine nasal cartilage. This paper describes its preparation and use as a xylose acceptor.
EXPERIMENTAL PROCEDURE
Materials
The following materials were purchased from Sigma: bovine serum albumin (Fraction V powder), casein (technical) and
by guest on January 2, 2019http://w
ww
.jbc.org/D
ownloaded from

Issue of June 25, 197” J. R. Balcer, L. Rodt%, and 8. C. Xtoolmiller 3539
casein hydrolysate (enzymatic), histone from calf thymus (type II and type III (lysine-rich)), gelatin from swine skin (type I), lysozyme, grade I, protamine sulfate from salmon, grade I, protamine chloride from salmon, grade V, m-O-phos- phothreonine, glycyl-L-serine, and L-serylglycine. Pronase (B grade), UDP-xylose, MES,’ HEPES, cytochrome c (equine heart, A grade), and 0-phospho-L-serine were obtained from Calbiochem. Ribonuclease B was purchased from Worthing ton, L-serine from Nutritional Biochemicals, poly-L-serine from Miles, bradykinin triacetate hydrate from Seikagaku, ferritin (horse spleen) from Mann, and glycyl-cu-L-aspartyl-L-serine acetate monohydrate from Cycle. The following peptides and serine derivatives were obtained from Fox: L-serylglycylglycine, glycylglycyl-L-serine, L-seryl-L-alanine, cu-L-glutamyl-L-serine, L-alanyl-L-serine, L-leucyl+serine amide hydrochloride, cr- aspartyl-L-serine, L-serine amide hydrochloride, and L-serine methyl ester.
Guanidine hydrochloride (reagent grade) was purchased from Eastman, cetylpyridinium chloride from K and K, potassium borohydride from Rlfa, and TNBS and DNS-chloride from Pierce.
Twice crystallized papain was prepared from crude enzyme (type II; Sigma) by the procedure of Kimmel and Smith (7). 0-/I-D-Xylopyranosyl-L-serine, synthesized by the method of Kum and Roseman (8), was a generous gift from Dr. Alvin Telser.
Pyridine used for chromatography was redistilled after reflux- ing over ninhydrin. DC-l resin was obtained from the Durrum Chemical Co.
UDP-[14C]xylose (174 mCi per mmole) was obtained from New England Nuclear. An increase in blank values in the xylosyl- transferase assays was sometimes observed with storage of the nucleotide sugar, which was then repurified by chromatography on water-washed Whatman No. 3MM paper in Solvent C (see below).
Fresh bovine nasal cartilage was obtained from the Wilson Laboratories, Chicago, and was stored frozen until required. Cartilage proteoglycan which had been degraded with chon- droitinase from Flavobacterium heparinum2 was a generous gift from Dr. V. C. Hascall. In this material, all repeating disac- charide units had been removed by the enzymatic degradation, leaving the oligosaccharide sequence AGlcUA-Gal-Gal-Xyl- of the carbohydrate-protein linkage region. Proteoglycan de- graded by testicular hyaluronidnse was prepared as described previously (9).
!Ileth,ods
Analytical Yethods
Hexosamine was estimated by the method of Boas with omis- sion of the resin treatment (10). IJronic acid was assayed by the carhazole method of Dische (11) and neutral sugars by an an- throne method (12). A Kjeldahl digestion method was used for estilnating total nitrogen.
1 The abbreviations used arc: MES, 2-(iv-morpholino)ethane- sulfonic acid: HEPES. iv-Zhvdroxvethvluiperazine-N’-2-ethane- sulfonic acid; TNBS,’ 2,4,6-&initiobenz&sulfonic acid; DNS, I-dimethylaminonaphthalene-5.sulfo-; PG, proteoglycan; PG-SD, Smith-degraded prot,eoglycan; CS-TP, peptides obtained by Smith degradation of tryptic chondroitin sulfate peptidoglycans; and DFP, diisopropyl flnorophosphate.
2 V. C. Hascall, submitted for publication.
Monosaccharides were separated and quantitated with the Technicon sugar chromatography system (model SC-l). Sam- ples were prepared for chromatography by hydrolysis in 1 M
HCl for 3 hours at loo”, followed by deacidification with Dowex 3 (COs- -) and adsorption of hexosamines and amino acids on Dowex 50 (H+). When this procedure was applied to the determination of xylose in periodate-oxidized proteoglycan, l.O-ml samples of the oxidation mixture (see below) were first reduced by addition of 0.5 ml of 0.1 M Tris, pH 8.4, and 20 mg of KBH+ Reduction was allowed to proceed for 16 hours at room temperature, and the reduced samples were dialyzed overnight against distilled.water prior to hydrolysis.
Amino acid analyses were carried out with a Technicon amino acid analyzer after hydrolysis in 6 M HCl at 100” for 20 hours. A modified gradient was employed which yielded complete separation of glucosamine, galactosamine, and all amino acids (13).
Amino groups were estimated by the TNBS method (14). High Voltage Paper Electrophoreais of Sugars-This was
carried out in 0.2 M borate, pH 10.0 (boric acid, 7.44 g, sodium hydroxide, 4.0 g, in 1 liter of water) on 57.cm sheets of Whatman No. 3MM paper (Savant Flat Plate, model FP-BOA). Other conditions are given in figure legends.
Amino acids and peptides were separated with a Gilson high voltage electrophorator, model D, in 8% formic acid, pH 1.7.
Paper Chromatography-Paper chromatography was per- formed on Whatman No. 3MM paper in: Solvent A, ethyl acetate-pyridine-water (8 :2: 1) ; Solvent B, I-butanol-acetic acid-water (60 : 15 :25) ; Solvent C, 95 y0 ethanol-l M ammonium acetate, pH 5.0 (7:3) ; Solvent D, l-butanol-ethanol-water (10:3:5); Solvent E, I-butanol-acetic acid-water (5:2:1); or Solvent F, I-butanol-pyridine-water (6 : 4: 3).
Thin Layer Chromatography-Thin layer chromatography was carried out on Quanta/Gram* plates (type Q2, microcrystalline cellulose) in phenol-water (100 g of phenol-39 ml of water).
Location Reagents-lx ninhydrin in acetone (containing 1% pyridine) was used for location of amino compounds. Reducing compounds were visualized with silver nitrate-sodium hydrox- ide (15).
Dan&&ion-Dansylation of [14C]xylosylserine was carried out essentially by the method of Gray (16). A 10.~1 sample, con- taining approximately 40 nmoles of xylosylserine and a total radioactivity of about 1000 cpm, was mixed with 0.2 ml of 0.2 M NaHC03, and the solution was evaporated to dryness in uacuo at 60”. After addition of 0.2 ml of water and 0.2 ml of 0.5% DNS-chloride in acetone, the mixture was incubated at 55” for 30 min, concentrated to dryness at 60”, and the residue was extracted vigorously with three 0.1.ml portions of water-sat- urated ethyl acetate. The supernatant fractions obtained after centrifugation were combined and evaporated to dryness. After addition of 20 ~1 of 50% aqueous pyridine, the extract was analyzed by paper electrophoresis in borate buffer, pH 10.
dleasurement of Radioactivity
Aqueous solutions of 0.5 ml were mixed with 10 ml of Perm- afluor solution (Packard Instruments) and 1 ml of Bio-Solv BBS-3 (Beckman). Samples were counted at 4” in a Packard Tri-Carb spectrometer (model 3375). Radioactivity on paper strips was located with a Packard radiochromatogram scanner (model 7201).
by guest on January 2, 2019http://w
ww
.jbc.org/D
ownloaded from

Biosydhesis 0.f Chodroitin Sulfate Proteoglycan
Preparation of Proteoglycan front Bovine Nasal Cartilage Smith Degradation of Proteoglycan
Three hundred grams of cartilage, which had been stripped of perichondrium and cut into 0.5-mm thick slices, were extracted at room temperature for 24 hours in 4.5 liters of 4.0 M guanidine- 0.05 M Tris, pH 7.5, as described by IIascall and Sajdera (17). The extract was poured through a Ruchner funnel to remove coarse particles, and the proteoglycan was precipitated with 2 volumes of ethanol. After 16 hours at 4’, the precipitate was collected by centrifugation, washed with 807, aqueous ethanol, absolute ethanol, and ether. The yield was 36.3 g.
The preparation was dissolved in 0.5 M NaCl and further purified by precipitation with cetylpyridinium chloride. The precipitate was recovered by centrifugation, dissolved in 2.0 M
NaCl, and reprecipitated with 3 volumes of ethanol. ,4fter washing with 800/, ethanol and absolute ethanol, the proteoglycan was redissolved in water and stored frozen. Yield was 31.4 g, based on dry weight determination by lyophilization of an aliquot of this solution. Analyses of this preparation are given in Tables I and II.
TABLE I
Chemical analyses o.f carlilage proteoglycan and PG-SD
PG (l.04 g) was dissolved in 200 ml of 0.05 M sodium meta- periodate-0.25 M sodium perchlorate, pH 5.0 (18), and kept in the da,rk at 25”. The destruction of xylosyl residues was mon- itored‘ every 24 hours as described above. After 5 days, no sylose could be detected, and the mixture was then dialyzed against distilled water for 24 hours. Potassium borohydride was added to a final concentration of 0.15% and reduction was allowed to proceed for 12 hours at 25”. The solution was then acidified to pH 1.0 with HCl and kept at 25” for 6 hours. Sfter neutralization and dialysis against distilled water for 5 hours, the dialyzed material was concentrated to 10 ml and applied to a column (2.1 x 36 cm) of Sephadex G-75 which was eluted with 0.2 M NaCl at a rate of 4 ml per hour (fraction volume, 2 ml). As seen from the elution pattern in Fig. 1, the protein emerged with the void volume and was clearly separated from a large hexosamine-containing peak which consists of the Smith-de- graded chondroitin sulfate chains. Fractions 7 to 15 which con- tained protein were pooled, dialyzed overnight against distilled water, and freeze-dried. Yield was 143 mg. ‘l’he analyses of this material (PG-SD, Preparation ;O are given in Tables I and II. --
component
Uranic acid. ............... Neutral sugars .............
Galactose ................ Xylose ................. Mmnose .............. Fncose. ................
Hexosamine ............... Glucosamine ............. Galact,osamine .........
Nitrogen ..................
Cartilage proteoglycan
7% 25.7
25.4 1.7
23.7 3.5
PC-SD Preparation A
%
0.3 8.4 (5.4 0.2 0.3
Trace 14.2 5.5 8.7 8.0
TABLE II
Amino acid analyses of cartilage proteoglycan and products of Smith deeg?adation
Amino acid
Aspartic acid Threonine.. Serine......... Glutamic acid. Proline. (ilycine. Alanine Valirre. Tsoleucine. Leucine. ._ Tyrosine . I’henylalanine Lysine Histidine. Arginine. Glncosamine Galactosamine.
Cartilage proteo- glyCiIl
.nsoles/m
0.089
O.OG5 0.147 0.148 0.090 0.152 0.088 0.062 0.041 0.094 0.020 0.042 0.040 0.019 0.050 0.095 1.324
-
P
g
PG-SD
repa~tion Prepa;tioI
~mole/nzy
0.2G3 0.466 0.181 0.219 0.304 0.297 0.515 0.614 0.353 0.327 0.350 0.345 0.268 0.286 0.173 0.207 0.160 0.131 0.272 0.335 0.026 0.052 0.120 0.164 0.089 0.158 0.049 0.060 0.116 0.184 0.307 0.300 0.486 0.100
-
-
1
CS-TP
~ld~/lQ!
0.494 0.260 0.5GO 0.700 0.435 0.731 0.400 0.334 0.181 0.398 O.OG7 0.147 0.112 0.082 0.209 0.146 0.155
Vol. 247, Ko. 1%
It is seen from Table II that the Smith-degraded product still contained about 1; times more galactosamine than glucosamine, indicating that the removal of chondroitin sulfate chains was not complete. Slight modification of the conditions of degrada- tion, consisting primarily of an extension of the period of acid cleavage to 3 days, has consistently yielded material in which galactosamine is only about 25OJ, of the total hexosamine. This indicates almost complete removal of chondroitin sulfate chain?, since the major part of the residual galactosamine can be ac- counted for by the galactosamine content of the keratan sulfate component of the product. Amino acid analysis of a tgljical chondroitin sulfate-free product is given in Table II (Prepara- tion 13). This material has been used for some of the experiments described in this paper.
Preparation of Tryptic Chondroitin Lhlfate Peptidoglycans from Cartilage
This material was a generow gift of Dr. Edwin Rosenblum and had been prepared by the following procedure. lzovine nasal septa (535 g) were cleaned of perichondrium and diced
r-
3.0
E g 2.0
,"
10
b *...A ,\
FRACTION NUMBER
FIG. 1. Fractionation of Smith-degraded proteoglycan or1 Sephadex G-75.
by guest on January 2, 2019http://w
ww
.jbc.org/D
ownloaded from

Issue of ,Junc 25, 197” J. R. Raked, Il. Rocldri, and A. C. Stoolmiller 3841
The tissue 1~;~s suspended in 1.5 liters of 0.1 M ammonium bicar- bonate, pH 8.1, and was digested with 1 g of trypsin at 37” with thymol as a preservative. .\fter 1 day, a further addition of 1 g of trypsin and 1 liter of buffer was made, and the illcubat,ion was continued for A t&d of 3 days. The digest was filtered tlrrougb gauze aud the cartilage residue was rinsed with water. The filtrate and washings (approximately 3 liters) were combined and the crude polysncrhnride was precipitated by addition of 5 vol- umes of ethanol and 25 ml of 25% sodium acetate solution. The precipitate was washed twice wit,h absolute ethanol, twice with ether, rind dried irk Z)CMLO over phosphorus pentoxide. Yield was 63 g.
Smith, Degradation, sf Tr!lpfic Chondroitin Sulfate Peptidoglycans
Four grams of tryptic peptidogfycans were dissolved in 400 ml of wat,er, filt,ered t,hrough Wlu~tman No. 1 paper, and passed through a column (4.4 x 18 cm) of AG 50-X2 (H+; 200 to 400 mesh). The column was rinsed with 1 liter of water and the combined effluent and wanhings were concentrated to 200 ml aft.er a.djusting to pH 5.0. The solution was mixed with 200 ml of 1.0 M sodium perchlorate and 400 ml of 0.1 M sodium meta- periodate a11r1 pH was again adjusted to 5.0. After oxidation for 5 days, the continued degradation was carried out as described for the proteoglycan with the exception that the peptide product after acid cleavage was isolated by adsorption to Dowex 50-X2 and elution with 0.25 M ammonium hydroxide. Yield was 44 mg. .inalyses of this nlat,eri:tl ((‘S-TP) are given in Table II.
Preparalion oj S~/losZ/Zlran~~eras~~
Epiphyses from la-day chick embryos were diced and homog- enized in 4 volumes of 0.05 M IIEl’ES-0.05 M KCl, pH 7.5, at’ 0” wit,h a Dual1 homogenizer. The homogenate was centrifuged at 10,000 X g for 10 min, and the supernatant fluid was then centrifuged at 105,000 x q for 1 hour. The resulting pellet was suspended in buffer, dispersed by sonication, and recentrifuged at 105,000 x q for 1 hour. The pellet obtained by this t,reatment ~3s suspended in 0.05 ;\I AIES-0.05 M KCl, pH 6.5, to give a pro- teiu conc*entration of approximately 5 mg per ml, and was stored at -20”. hfter thawing, the particulate enzyme preparation was again dispersed by brief ult,rasonic treatment prior to use.
It should be noted t,hat a large proportion of the total xylosyl- trausferaae activity of a homogenate is present in the 105,000 X g supernatant fluid (6, 19). The purification of the soluble enzyme, with I’G-SD as substrate, and some properties of the purified enzyme are described in a previous paper (19). In the present work, the l)articulat~e enzyme was used except ill one es- periment, where a low level of endogenous acceptor was desirable. Soluble sylosyltransferase for t,his experiment was purified from 285 ml of a 105,000 x q supernatant fraction essentially as de- scribed by Stoolmiller et nl. (19). The material precipitated by ammonium sulfate between 27 aud 68(,>; saturation was dissolved in 0.05 M MES-0.2 M KCl, $1 6.5, and dialyzed against the same buffer for 2 days at 4’. Of the dialyzed solution (total volume, 39 ml), a 2%ml portion was applied to a column (3.3 x 187 cm) of Sephadex G-200, which was eluted with 0.05 M MES-0.2 M KCI, pII 6.5, at a rate of approximately 7 ml per hour. Xylosyltrans- ferase emerged with an effluent volume of 750 to 940 ml. A por- tion (35 ml) of the latter half of the peak was concentrated by ultr~~filtration through a V&4-05 filter (Amicon) t’o about 8 ml.
Assay of Xylosyltransferase Activity
Incubation mixtures contained the followiug components, in a final volume of 77 ~1: 2.5 pmoles of MES buffer, pII 6.5; 2.5 ymoles of KCl; 0.4 pmole of MnCL; 0.75 pmole of KF; 1.5 nmoles of UDP-[14C]xylose (specific activity approximately 30 ,.&i per pmole) ; approximately 0.25 mg of enzyme protein; and acceptor, 0.75 pmole of peptide acceptors or 0.1 to 0.2 mg of PG-SD. ilfter incubation for 30 or 60 min at 37”, the tubes were cooled in an ice bath, and [14C]xylose incorporation was measured by one of the following methods, according to the nature of the accep- tors.
>Vethod a-Incubation mixtures were centrifuged in the cold for 10 min, 50 ~1 of the supernatant fluid were mixed with 0.4 ml of 0.02 M HCl and applied to columns (0.5 X 5 cm) of AG 5OW-X2 (H+, 200 to 400 mesh). After washing with 3 to 4 bed volumes of 0.01 M HCI, adsorbed reaction products were eluted with 3 bed volumes of 2 M ammonium hydroxide. The eluate was evap- orated 60 dryness in a Buchler Evapo-Mix, taken up in 0.5 ml of 0.01 M HCl, and radioactivity was measured. This method was was used to assay acceptor activity of small peptides and other compounds, which even after glycosylation are readily adsorbed to the resin.
Method b-O.05 ml of 1% bovine serum albumin was added, followed by 0.15 ml of 10% trichloroacetic acid-49/ phospho- tungstic acid. Precipitated protein was recovered by centrifuga- tion, washed twice with 5% trichloroacetic acid, and dissolved in 0.1 ml of 1 M NaOH. After dilution to 0.5 ml with water, the radioactivity of the solution wa.s measured. This method was used with substances which are precipitable by the tri- chloroacetic acid-phosphotungstio acid mixture, i.e. proteins, PG-SD, and CS-TP. Occasionally, the incubation mixture was centrifuged and the radioactivity in pellet and supernatant fluid was determined separately.
Method c--In a few experiments, a paper chromatographic procedure was used which is described in detail in a previous report (19).
Enzyme assays were performed under conditions where the incorporation of radioactivity from UDP-[14C]xylose was linear with respect to both time of incubation and enzyme concentra- tion. The concentration of Mn2+ was chosen on the basis of previous studies of Robinson et al. (6). The pH optimum for xylosyl transfer to both endogenous and exogenous acceptor is 6.5 (6, 19). In the interest of economy, UDP-[14C]xylose was used at approximately 60% of t*he caoncentration required for 1’ mnx (19).
Isolation of (~4~Xylosylserine from [14C]Xylosyl-PG-SD
[%]Xylosyl-PG-SD was synthesized by incubating 7 ml of soluble xylosyltransferase (total protein, 4.0 mg) with 28 mg of PG.SD and 0.2 rmole of IyDl’-[14C]xylose (specific activity, 30 PC1 per pmole) in the presence of 0.003 M NhClz and 0.01 M KF (total volume, 10.1 ml). After 2 hours at 37”, the reaction mix- ture was cooled in an ice bath, an aliquot was removed for deter- mination of incorporated radioactivity by Method b, and 7 ml of 1% albumin and 28 ml of 10% trichloroacetic acid were added. The mixture was kept overnight at O”, and the precipitated material was collected by centrifugation and washed three times with IO-ml portions of 10% trichloroacetic scid. On addition of 5 ml of ethanol, the precipitate dissolved and was again pre- cipitated by addition of an equal volume of ether. After an
by guest on January 2, 2019http://w
ww
.jbc.org/D
ownloaded from

3542 Biosynthesis of Chondroitin Sulfate Proteoglycan Vol. 247, n-o. 12
additional wash with ethanol-ether (1: l), the precipitate was washed once with ether and dried in a desiccator over phos- phorus pentoxide.
The product was suspended in 4 ml of 0.12 M sodium acetate, pH 6.5, containing 0.005 M EDTA and 0.005 M cysteine, and was digested with 26 mg of papain at 62” for 43 hours, with a further addition of papain and cysteine after 24 hours. Reaction was stopped by heating the digest at 100’ for 5 min, and the pre- cipitate was sedimented by centrifugation and washed twice with 2-ml portions of 0.2 M Tris-HCI, pH 7.5, containing 0.03 M
CaCh Pronase B (20 mg) was added to the combined supernatant
and washings, and digestion was carried out for 40 hours at 55” with addition of another 20 mg of Pronase after 15 hours. The digest was heated at 100” for 5 min (no precipitate formed) and was diluted to 10 ml. A l-ml portion of the digest was mixed
TABLE III
Assay of xylose acceptor activity Assays were carried out under standard conditions (30-min
incubation; approximately 0.5 mg of enzyme protein per incuba-
tion mixture). Compounds tested were added at a final concen- tration of 10 rnM (0.75 pmole per reaction mixture). Product formation was assayed by Method a. Other compounds which
were tested and found inactive include the following: Ser-Ala, Glu-Ser, Ala-Ser, Asp-Ser, Leu-Ser amide, serine amide, phos- phoserine, phosphothreonine, and bradykinin triacetate.
Compound added -~~-
Radioactivity incorporated
None............. .._...... 322 Serine. ., ._.. .._.............. 322
Serine methyl ester. 368 Ser-Gly. 327 Ser-Gly-Gly . 697 Gly-Ser. ..._..............._........,.. 334 Gly-Gly-Ser............................. 344 Gly-Asp-Ser...... .._............... 315 Polyserine............................... 419
800
l!
1
I I I I
0 20 40 60 80
S,mM
FIG. 2. Substrate concentration curve for Ser-Gly-Gly. Stand- ard incubation mixtures contained up to 0.08 M Ser-Gly-Gly. After incubation for 30 min at 37”, the radioactive products were recovered by Method a. Incorporation into endogenous acceptors was approximately 150 cpm and has been subtracted to give the values shown in the figure.
with carrier 0-/3-n-xylosyl-L-serine (4 pmoles) and applied to a column (0.9 x 49 cm) of DC-l resin, which was eluted with sodium citrate buffer, pH 2.8 (0.2 M with respect to sodium) at a rate of approximately 40 ml per hour (column temperature 55”). Five-minute fractions (3.5 ml) were collected, and 50-~1 aliquots were taken for radioactivity determinations and analysis by the TNBS method. Fractions 9 and 10 (containing xylosyl- serine) were combined, adjusted to pH 1.9 with HCI, and applied to a column (1.7 x 7 cm) of AG 5OW-X2 (H+, 200 to 400 mesh), which was rinsed with 30 ml of 0.01 M HCI and subsequently with 20 ml of water. The adsorbed material was eluted with 0.25 M ammonium hydroxide and, after the alkali breakthrough, a further 10 ml were collected. The eluate was evaporated to dryness twice in a rotary evaporator, taken up in 0.1 ml of water and subjected to further analyses as described under “Results.”
The contribution of endogenous acceptors to the formation of [*4C]xyIosylserine was estimated by carrying an incubation mixture without added PG-SD, at one-tenth the scale, through
the procedure described above.
RESULTS
Xylosyl Transfer to Ser-Gly-Gly-Many small serine-contain- ing peptides of known structure were tested as xylose acceptors. Table III shows that one of these, Ser-Gly-Gly, had significant acceptor activity. The apparent K, for Ser-Gly-Gly n-as 20.0 mM as illustrated in Fig. 2.
The product of xylosyl transfer to Ser-Gly-Gly was presumed to be [*4C]xylosyl-Ser-Gly-Gly, and paper electrophoresis at pH 2 showed that a radioactive compound with a mobility appropriate for Xyl-Ser-Gly-Gly was indeed present in incuba-
tion mixtures containing the acceptor. However, two addi- tional radioactive spots were also observed, and one of these had the same mobility as xylosylserine. This result suggested the presence of peptidases in the xylosyltransferase prepara- tion, and it is shown in Fig. 3 that hydrolysis of the substrate
0 0 S SGG SG
-1 0 CD&
-2
-3 00 0
-4 00 0
-5 00 S SGG G GG
1
FIG. 3. Hydrolysis of Ser-Gly-Gly by the particulate xylosgl- transferase preparation. Standard incubation mixtures con- tained 0.75 pmole of Ser-Gly-Gly and regular or DFP-treated enzyme. After incubation, the mixtures were centrifuged, and aliquots of the supernatant fluids were analyzed by paper elec- trophoresis at 33 volts per cm for 4 hours at pH 2; papers were stained with ninhydrin. S, serine; G, glycine; SG, serylglycine; GG, glycylglycine; SGG, serylglycylglycine. 1, serine, seryl- glycine, and glycylglycine; 2, DFP-treated enzyme plus seryl- glycylglycine; 3, serine, serylglycylglycine, glycylglycine; 4, enzyme plus serylglycylglycine; 5, glycine, glycylglycine.
by guest on January 2, 2019http://w
ww
.jbc.org/D
ownloaded from
Issue of June 25, 1972 J. R. Baker, L. Rod&, and A. C. Stoolmiller 3843
TABLE IV
K,,, values of xylosyl acceptors
Acceptor
Ser-Gly-Gly . .
CS-TP....................... PG.SD....... .._ . . . . .
Km
WC/l mu (serine)
4380 20.0 779 0.44 210 0.064
into serine and glycylglycine actually occurred during the incubation. Treatment of the enzyme preparation with 1% DFP at room temperature overnight effectively inhibited proteolysis without diminishing xylosyltransferase activity.
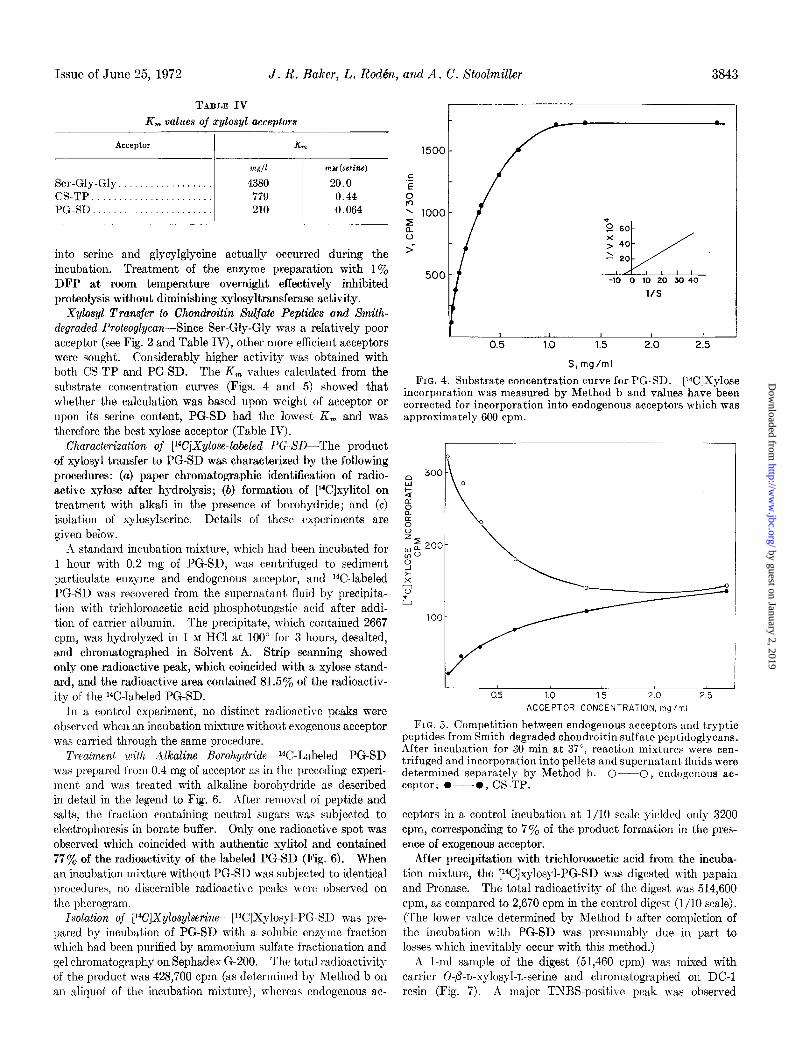
Xylosyl Transfer to Chondroitin Sulfate Peptides and Smith- degraded Proteoglycan-Since Ser-Gly-Gly was a relatively poor acceptor (see Fig. 2 and Table IV), other more efficient acceptors were sought. Considerably higher activity was obtained with both CS-TP and PG-SD. The K, values calculated from the substrate concentration curves (Figs. 4 and 5) showed that whether the calculation was based upon weight of acceptor or upon its serine content, PG-SD had the lowest K, and was therefore the best xylose acceptor (Table IV).
Characterization of [14(JXylose-labeled PG-SD-The product of xylosyl transfer to PG-SD was characterized by the following procedures: (a) paper chromatographic identification of radio- active xylose after hydrolysis; (b) formation of [14C]xylitol on treatment with alkali in the presence of borohydride; and (c) isolation of xylosylserine. Details of these experiments are given below.
A standard incubation mixture, which had been incubated fol 1 hour with 0.2 mg of PG-SD, was centrifuged to sediment particulate enzyme and endogenous acceptor, and 14C-labeled PG-SD was recovered frotn the supernatant fluid by precipita- tion with trichloroacetic acid-phosphotungstic acid after addi- tion of carrier albumin. The precipitate, which contained 2667 cpm, was hydrolyzed in 1 M HCl at 100” for 3 hours, desalt,ed, and chromatographed in Solvent A. St.rip scanning showed only one radioactive peak, which coincided n-it,h a xylose stand- ard, and the radioactive area contained 81.5% of t,he radioactiv- it’y of the 14C-labeled PG-SD.
In a control experiment, no distinct radioactive peaks were observed when an incubation mixture without exogenous acceptor was carried through the same procedure.
Treatment with illkaline BoroI~ydride-14C-Labeled PG-SD was prepared from 0.4 mg of acceptor as in the preceding experi- ment and was treated with alkaline borohydride as described in detail in the legend to Fig;. 6. After removal of peptide and salts, the fraction containing neutral sugars was subjected to electrophoresis in borate buffer. Only one radioactive spot was observed which coincided with authentic xylitol and contained 77% of the radioactivity of the labeled PG-SD (Fig. 6). When an incubation mixture without PG-SD was subjected to identical procedures, no discernible radioactive peaks were observed on the pherogram.
Isolation of [W]Xylosylserine-[14C]Xylosyl-PG-SD was pre- pared by incubation of PG-SD with a soluble enzyme fraction which had been purified by ammonium sulfate fractionation and gel chromatography on Sephadex G-200. The total radioactivity of the product was 428,700 cpm (as determined by Method b on an aliquot of the incubation mixture), whereas endogenous ac-
1500
.c E
8 ' 1000 5 5
Y
500
S, mg/ml
FIG. 4. Substrate concentration curve for PG-SD. [W]Xylose incorporation was measured by Method b and values have been corrected for incorporation into endogenous acceptors which was approximately 600 cpm.
I I 0.5 1.0 1.5 2.0 2.5
ACCEPTOR CONCENTRATION, mg/ml
FIG. 5. Competition between endogenous acceptors and tryptic peptides from Smith-degraded chondroitin sulfate peptidoglycans. After incubation for 30 min at 37”, reaction mixtures were cen- trifuged and incorporation into pellets and supernatant fluids were determined separately by Method b. O-O, endogcnous ac- ceptor; O---O, CS-TP.
ceptors in a control incubation at l/10 scale yielded only 3200 cpm, corresponding to 7% of the product formation in the pres- ence of exogenous acceptor.
After precipitation with trichloroacetic acid from the incuba- tion mixture, the [~4C]xylosyl-PG-SD was digested with papain and Pronase. The total radioactivity of the digest \vas 514,600 cpm, as compared to 2,670 cpm in the control digest (l/10 scale). (The lower value determined by Method b after completion of the incubation with PG-SD was presumably due in part to losses which inevitably occur with this method.)
A l-ml sample of the digest (51,460 cpm) was mixed with carrier 0-P-n-xylosyl-L-serine and chromatographed on DC-1 resin (Fig. 7). A major TNBS-positive peak was observed
by guest on January 2, 2019http://w
ww
.jbc.org/D
ownloaded from

3844
roction
9-10
15 F
350
300
250
200
t 150 o
100
50
XY LITOL
XYLOSE 0 5 10 =p CM
FIG. 6. Formation of [%]xylitol on treat,ment of [“C]PG-SD with alkali and borohydrido. An incubation mixture of twice the standard size, containing 0.4 mg of PG-SD, was incubated at 37” for 1 hour, t.he mixture was centrifuged, and Y-labeled PG-SD was isolated from the supernatant fluid by precipitation with trichloroacctir acid-phosphotungstic acid in the presence of carrier albumin. After drying with ethanol and ether, t,he precipitate (total radioactivity, 5334 cpm) was suspended in 0.2 ml of 0.5 M
NaOH, containing 0.2 M KBH4, and left overnight at 25”. Peptide was precipitated with 0.6 ml of 10°yO trichloroacetic acid, the pre- cipitate was washed with 0.5 ml of 1Oy0 trichloroacetic acid, and the combined supernatant fluids were extracted twice with 5 volumes of ether. The solution was acidified to pH 2 and passed through a column (0.5 X 3 cm) of Dowex 50-X2 (H+, 200 to 400 mesh), which was rinsed with 0.01 IU HCl. The effluent was evap- orated four times with additions of methanol, and the desalted sample (total radioactivity, 4087 cpm) was subjected to paper electrophoresis in 0.2 M borate, pH 10.0. The pherogram was scanned for radioactivity, and sugars were located with AgNOs/ NaOH. A control sample without added PC+-SD was carried through the same procedure.
t I
10 20 Origin
CM
FIG. 8. Paper electrophoresis of Fractions 9 plus 10 at pH 1.7. Electrophoresis was performed on llO-cm strips of Whatman No. 3MM paper at 4000 volts for 4 hours. Xyl-Ser, O-p-D-xylosyl-L- serine. Minor ninhydrin-positive components are indicated by dashed or unfG/led spots.
I )
Fractions
9 -10
j T 10 20
Origin CM
FIG. 9. Paper electrophoresis of Fractions 9 plus 10 at pH 10. Electrophoresis was carried out at 3000 volts for .i hours. Sy2-SW, 0-P-n-xylnsyl-L-serine.
roctions
9- 10
Xyl -Ser
CA 0
4 8 12 16 20
Fraction Number
Fro. 7. Chromatography of a protcolytic digest of [‘~C]xylosyl- PC-SD on DC-1 resin. For details, see “Methods.” a--O, radioactivity; O---O, amino groups (TNBS method wit,h alanine as standard). !
160
120
z
I30 8
40
15 1‘ Origin
5 CM
(Fractions 9 :wi 10) which contained the added sylosylserille and 4574 of the radioactivity applied to the column. Fractions 9 and 10 were pooled (total radioactivity, 23,130 rpm) and de- salted by adsorption to AG 50-X2 and elution with ammonia. The radioactive product was characterized further by several
procedures, including paper electrophoresis at pH 1.7 (Fig. 8) and pH 10 (Fig. 9), thin layer chromatography on cellulose in phenol-water (Fig. lo), and paper chromatography in Solvents II, E, and F for 65 hours. Only one major radioactive prod-
FIG. 10. Thin layer chromatography of Fractions 9 plus 10 on
cellulose in phenol-water. Xyl-Ser, 0./j-n-xylosyl-L-serine. Minor ninhydrin-positive components are indicated by !LnJiZZed
uct was detected by these analyses, which in all cases migrated spoas
by guest on January 2, 2019http://w
ww
.jbc.org/D
ownloaded from

Issue of Juno “5, 197” J. R. Baker, L. Rodth, and A. C. 8oohiller 3s45
I i n 50
40
30 I
20 k
10 IL’ I
T 10 20 30 40 50
Origin CM
FIG. 11. Paper electrophoresis of DNS derivative of Fractions 9 plus 10 at pH 10. Mectrophoresis was carried out at 3000 volts for 5 hours. Also shown are the positions of JINS-serine and DNS-xylosylserine which were prepared as described nnder “Met.hods.” Dasherl spots had a blue fluorescence and were pre- sumably the normal breakdown producm from T)NS chloride (e.g. the sulfonic acid and sulfonamide).
to the same position as the added xylosylserine. Although niu hydrin-positive compounds other thau sylosylserine were also present in Fractions 9 to 10, these substances were presumably nonxylosylated peptides, since little or no radioactivity was found outside the xylosylserine area in the analyses illustrated
in Figs. 8 to 10.
Dausylation of the material in Fractions 9 to 10 yielded 0111) one siguificant radioactive product, which migrated to the same positiou as DNS-xylosylserine on electrophoresis at pH 10 (Fig. 11).
inhibition of Sylosyl Transfer to Ihdogenms hceptors by l?x- ogenous St&rules-Some assurance t,hat q-lose iucorporat,ion
into esogeuous substrate represents the same reaction that occurs with the eudogeuous acceptor has been obtained by competition experimeuts, iii which xylosyl transfer to endogenous acceptor was measured in the presence of increasing concentrations of exogenous substrate. It is seen from Fig. 5 t.hat addition of in- creasing amounts of CS-Tl’ resulted in a progressive decrease in
xylosyl transfer to the endogenous acceptor. However, when the same experiment was carried out with PG-SD as exogenous acceptor, 110 such effect was observed. Reasons for this apparent discrepancy will be discussed later.
Other l’roteins as Xylosyl dcceptors-A uumber of proteins other than PG-SD were assayed for acceptor activity and proved to be inac.tive, with the exception of ribonuclease I3 (Table V). Despite its apparent acceptor activity, this proteiu was not truly a substrate for xylosyl transfer, since no radioactivity could be detected in RNase which had been reisolated by gel chromatography after completed reactiou. The stimulating effect of RNase is due, in part at least, to solubilizstion of the
xylosyltransferase, which occurs concomitantly with digestion of RNA. Details of the RNase effect will be described elsewhere.
It should be uoted that. the intact proteoglycnn as well as the products of degradation by testicular hyaluronidase or bacterial
chondroitiuase were all iuactive as acceptors (Table VI), al- though approximately half of the seriue residues iu these com- pounds are not glycosylated.
The present study was undertaken with a 2-fold purpose: to gain more iuformstiou conceruiug the substrate specificity of the
Proteins tested as xylosyl acceptors
Assays were carried out under standard conditions (30.min incubation) ; proteins were added at a concentration of 13 mg per
ml (1.0 mg per reaction mixture); radioactivity in products was measured by Method b. Other compounds which were test,ed and found inactive include histone, protamine chloride, ferritin, cytochrome c, a casein hydrolysate, and lysozyme. Although lysozyme was not a substrate and did not affect incorporation into
endogenous acceptors, it stimulated xylosyl transfer to PG-SD about a-fold.
Protein added / Radioactivity incorporated
-.
None.
Bovine serum albumin, Gelatin. Ribonuclease B Casein........... _...._............
Protamine sulfate. _.
ce+= l!lO 16X 1x1
761 114 150
Assay of xylose acceptor activity of cartilage proteoglycan
Incubations were carried out for 60 min under standard condi-
tions. In Experiment 1, reaction mixtures were centrifuged aft.er incubation; pellets were assayed by Method b and supernatant, fluids by paper chromatography (19). In Experiment 2, reaction
mixtures were assayed by the paper chromatographic procedure
(19).
Compound added
None............... Proteoglycan (0.2 mg). PG-SD (0.2 mg)
Experiment 1
Radio- Radio- activity
activity in pellet
in super- natant
fluid
cpm cpm
324 183 327 172 727 470
Total Total radio- radio-
activity activity imo;o-
__.
c*?It c@n
507 528 499 474
1197 1243
xylosyltransferase reaction by using exogenous substrat.es of well defined nature, and to develop an assay for the quantitat.ion of enzyme activity independent of the concentration of endoge-
nous acceptors. Such an assay has not been previously available, and earlier studies have relied on the presence of endogenous acceptors in the preparations used (4-6). Consequently, it has not been possible to quantitate enzyme levels satisfactorily, since the reaction systems have contained two unknown dependent variables, i.e. enzyme concentration and acceptor concentratiou. Furthermore, lack of exogenous substrates has so far made it impossible to purify the xylosyltransferase beyond the poiut where endogenous acceptors are removed.
The acceptors tested in this study range from small serine- containing peptides and other serine derivatives to the entire protein core of the chondroitin sulfate proteoglycan. Since in- formation regarding the amino acid sequence of the carbohydrate-
protein linkage region is limited, it was not possible to approach the problem in an entirely rational way. However, it appears likely that the carboxyl group of serine is always bound to a
by guest on January 2, 2019http://w
ww
.jbc.org/D
ownloaded from

3846 Biosynthesis of Chondroitin Sulfate Proteoglycan Vol. 247, No. 12
glycine residue; therefore, it was of particular interest to test peptides containing a Ser-Gly sequence.
One such peptide, Ser-Gly-Gly, had some xylose acceptor ac- tivity and is the only small substrate found so far. However, in view of the rather high K, value, 20 mM, better acceptors were sought, and, initially, peptides from the chondroitin sulfate- protein linkage region which had been obtained by acid hydrolysis of tryptic “doublet” preparations (ZO), were tested. Since such peptides were indeed acceptors, the search for suitable substrates was extended to the isolation of tryptic peptides in more intact form by Smith degradation, and, similarly, the entire proteogly- can was subjected to the same procedure. As indicated by the results shown in Fig. 4 and Table IV, the macromolecular Smith- degraded proteoglycan was a far better acceptor than any of the smaller substrates. This finding is consistent with observations on other transferase reactions in which a polypeptide is glycos- ylated, such as the N-acetylgalactosaminyl transfer in the for- mation of salivary gland mucins (21) and the galactosyl transfer to the protein core of basement membrane collagen (22). In the latter study, galactosyl transfer to tryptic peptides was also obtained, but these peptides were only 23% as active as the en- tire protein core.
In the native proteoglycan, approximately half of the serine residues are bound to polysaccharide chains, and half are not substituted. Nevertheless, the intact proteoglycan is not a xylose acceptor. The failure to obtain xylose transfer to the free serine residues could conceivably be due to the presence of the chondroitin sulfate chains, but the products of hyaluroni- dase or chondroitinase digestion, in which the unsubstituted serines are probably more readily accessible, are also inactive. Although the possibility still exists that the oligosaccharides, which remain attached to the protein, prevent the glycosylation of the unsubstituted residues, the data are more in keeping with the notion that only specific serine residues can serveas acceptors.
In this context, it is also of interest to consider the presence of keratan sulfate in the cartilage proteoglycan. Some of the keratan sulfate chains are bound to serine, whereas others are linked to threonine, in both cases by glycosidic linkages to N- acetylgalactosamine (23). Whereas initiation of chondroitin sulfate and keratan sulfate chains could conceivably occur ran- domly at any serine site in the molecule, this possibility is not supported by the present results nor by current knowledge of proteoglycan structure (for review, see Reference 24). It ap- pears more likely that the proteoglycan has a definite structure and that the xylosyltransferase specifically seeks out certain serine residues in the polypeptide, whereas an N-acetylgalactos- aminyltransferase initiates keratan sulfate synthesis at other serine sites.
Further evidence for a rather specific requirement with respect to the structure of the acceptor was derived from the finding that none of the proteins tested served as a substrate except PG.SD. The stimulation of xylose incorporation into trichloro- acetic acid-precipitable material, which was observed in the presence of ribonuclease, was not due to xylosyl transfer to this substance, as could clearly be shown by reisolation of the ribo- nuclease after completed incubation. As indicated previously, the effect of ribonuclease was due, in part at least, to solubiliea- tion of the particulate portion of the xylosyltransferase. These observations are reminiscent of the findings of Grebner and Neufeld (25) which prompted us to test this substance in the present system. These authors showed that ribonuclease and
lysoayme restored and stimulated activity in aged preparations of xylosyltransferase from mouse mastocytoma. Reisolation of lysozyme after incubation showed that it had not become xylosy- lated, and no ready explanation could be given for the stimula- tory effect.
The finding that a protein like ribonuclease can stimulate xy- lose incorporation greatly without being an acceptor emphasizes the importance of characterizing the reaction products carefully, particularly in view of the complexity of the system used to de- tect the xylosyl transfer. Although the characterization of the [14C]xylose-labeled PG-SD falls short of a definitive identification of the product, the following data would appear sufficient to permit the conclusion that xylose has indeed been transferred to serine hydroxyl groups of the protein core: (1) xylose was incor- porated into the protein in an alkali-labile linkage from which it could be released as xylitol on treatment with alkaline borohy- dride; (9) a radioactive compound with the properties of xylosyl- serine was isolated in 45% yield after proteolysis of [14C]xylose- labeled PG-SD. The identity of the radioactive product with xylosylserine was indicated by (a) its comigration with the au- thentic compound in the course of preparative chromatography on DC-l resin, (b) the electrophoretic mobility at pH 1.7 and pH 10, (c) the RF values observed on paper chromatography and thin layer chromatography in four different solvents, and (d) dansylation, which yielded a radioactive derivative with the same electrophoretic mobility as DNS-xylosylserine.
An apparent discrepancy in the experiments concerned with competition between endogenous and exogenous substrates de- serves further comment. Whereas the small tryptic peptides competed with the endogenous acceptors as substrates for xylosyl transfer, no such effect was observed with the macromolecular PG-SD. This finding does not necessarily invalidate the as- sumption that PG-SD, CS-TP, and the endogenous acceptors are all substrates for the same enzyme, since it seems likely that the macromolecular substrate may easily be excluded from the internal acceptor sites of the particulate enzyme, where much of the transfer to the endogenous acceptors presumably takes place. In this context it should be remembered that the particulate en- zyme preparation consists of healed vesicles of the endoplasmic reticulum (26) and that the dimensions of the membranes are such that only a relatively minor proportion of the transfer sites would be located on the surface of the vesicles. Consequently, competition between the endogenous acceptors and PG-SD would occur only on the surface of the vesicles and might not be expected to have a significant effect on the total incorporation. On the other hand, it seems plausible that the smaller tryptic peptides would be capable of penetrating the particles more readily and thus have access to the entire pool of transfer sites.
Acknowledgments-We are grateful to Dr. Albert Dorfman for stimulating discussions and for reviewing this paper. Skillful technical assistance was given by Miss Gudrun Backstrom, Mrs. Lucila Penas, Mrs. Louise Roth, Miss Mary Lou Spnch, Mrs. Lillian Thompson, and Mrs. Sadako Yamagata.
REFERENCES
1. BAKER, J. R., AND ROD&N, L. (1970) Fed. Proc. 29,338 2. RAKER, J. R., ROD~N, L., AND YAMAGATA, S. (1971) Biochern.
J. 126, 93P 3. LINDAHL, U., AND ROD&N, L. (1971) in A. GOTTSCHALK (Edi-
tor), Glycoproteins, Ed. 2, Elsevier Publishing Co., Amster- dam, in press
by guest on January 2, 2019http://w
ww
.jbc.org/D
ownloaded from

Issue of June 25, 1972 J. R. Baker, L. Rod&, a?zd A. C. Stoolmiller 3847
4. GICEWCCR, E. E:., %LL, C. W., AND NEUFELD, E. F. (1966) Biochem. Biophys. Kes. Commun. 22, 672
5. GILE~XEI~, El. E., HUL, C. W., AXD XIZUFELD, IX. F. (1966) Awh. Biochem. Biophys. 116, 391
6. ROUIN~ON, H. C., TBLSER, A., AND DORFM~N, A. (1966) Proc. Nat. Acad. Sci. U. S. A. 66, 1859
7. KINIMEL, J. R., AND SMITH, YE. L. (1954) J. Biol. Chem. 207,515 8. KUM, K., AND R.OSEMAN, S. (1966) Biochemistry 5,3061X3065 9. GREGORY, J. D., LAURENT, T. C., BND ROD&N, L. (1964) J.
Biol. Chem. 239, 33123320 10. Boas, N. F. (1953) J. Biol. Chem. 204,553 11. DISCHE, Z. (1947) J. Riol. Chum. 167, 189 12. MORRIS, Il. L. (1948) Science 107, 254 13. STFXN, E. L. (1970) Advances in automated analysis, vol. 1,
p. 343, Mediad, Inc., White Plains, N. Y. 14. MOICKXXH, L. C. (1967) Anal. Biochem. 18,64 15. SMITH, I. (Editor) (1960) Chlomatographic and electTophoretic
techniques, Vol. I, p. 252, Interscience Publishers, New York 16. GRAY> w. (1967) illethods ~nzyn/o/. 11, 130
17. H.ISCALL. V. C., AND SAJDER.\, S. W. (1969) J. Biol. Chem. 244.
18. 19.
20.
21.
22.
23.
24.
25.
26,
2384-2i96 ’ SCOTT, J. E. (1968) Biochim. Biophys. Acta 170, 471-473 STOOLMILLER, A. C.. HORWITZ, A. L., AND DORFMAN, A. (1972)
J. Biol. Chem. 24?, 3525-3532 . ANDERSON, B., HOFFMAN, P., AXD MEYER, K. (1965) J. Biol.
Chem. 240, 156-167 MCGUIRE, E. J., AND ROSEMAN, S. (1967) J. Biol. Chem. 242,
3745-3755 SPIRO, M. J., AND SPIRO, R. G. (1971) J. Biol. Chcm. 246, 4910-
4918 ANDERSON BRAY, B., LIEBERMAN, R., AND MEYICR, K. (1967)
J. Biol. Chem. 242, 3373-3380 MUIR, H., in M. A. R. FREEMAN (Editor), Adult articular
cartilnge, Chapter 3, Pitman’s Medical, London, in press GREBNEX, E. E., AND NEUFELD, E. F. (1969) Biochim. Biophys.
A eta 192, 347-349 HORWITZ, A. L., AND DORFMBN, A. (1968) J. Cell Biok 38,
358
by guest on January 2, 2019http://w
ww
.jbc.org/D
ownloaded from

John R. Baker, Lennart Rodén and Allen C. StoolmillerEXOGENOUS ACCEPTORS
SMITH-DEGRADED CARTILAGE PROTEOGLYCAN AND OTHER Biosynthesis of Chondroitin Sulfate Proteoglycan: XYLOSYL TRANSFER TO
1972, 247:3838-3847.J. Biol. Chem.
http://www.jbc.org/content/247/12/3838Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/247/12/3838.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on January 2, 2019http://w
ww
.jbc.org/D
ownloaded from