biosynthetic pathway connects cryptic ribosomally ... · biosynthetic pathway connects cryptic...
TRANSCRIPT

Cell Chemical Biology, Volume 23
Supplemental Information
Biosynthetic Pathway Connects Cryptic Ribosomally
Synthesized Posttranslationally Modified Peptide
Genes with Pyrroloquinoline Alkaloids
Peter A. Jordan and Bradley S. Moore

Figure S1, related to “Time-Course of Ammosamides A-C Production” in the Experimental Procedures section:
LCMS analysis of (A) S. coelicolor M512-pCAP01-amm and (B) Streptomyces sp. CNR-698 culture time-course.
Chromatograms at 254 nm are provided above for each time point. Peaks were verified by retention time in references
to standards (bottom peaks), UV absorbance, and mass spectrometry. Deviations in retention time between (A) S.
coelicolor M512-pCAP01-amm and (B) Streptomyces sp. CNR-698 culture time-courses were attributed to two
different Phenomenex Luna 5 μm C18(2) 100 Å, 100 x 4.6 mm columns used for each analysis. The identity of
ammosamides A-C were verified with standards, UV absorbance and MS analysis.

Figure S2, related to “Chemical Complementation of M512-pCAP01/ammamm3” in the Experimental
Procedures section: LCMS analysis of S. coelicolor M512 pCAP01/ammamm3 mutants chemically complemented
with 4 mM rac-6-chloro-tryptophan, “6-Cl-Trp” (2mM of each enantiomer). Chromatograms at 254 nm are provided
above. Peaks for ammosamides A and B are labeled with 1 and 2, respectively, and 6-chloro-tryptophan is indicated
with an asterisk (*). The top trace corresponds to a media blank and the bottom two traces correspond to cultures
complemented with rac-6-chloro-tryptophan, whereas the remaining samples correspond to cultures treated with L-
tryptophan. The decomposition of 6-chloro-tryptophan is evident in the bottom two traces. The absence of either
ammosamide A or B was verified by retention time with respect to the positive control (pCAP01/amm), UV
absorbance, and mass spectrometry. In particular, the peak corresponding to the same retention time as 1 in the bottom
two traces was not related to ammosamide products based upon UV-Vis profile and MS data.

Figure S3, related to “Secondary Metabolite Profiling of S. coelicolor M512-pCAP01/amm and mutants” in the
Experimental Procedures section: Summary of gene deletions to ‘core’ biosynthetic genes in the amm cluster that
share close homology to lymphostin biosynthesis. The absence of ammosamide A-C was verified by retention time
with respect to a positive control (pCAP01/amm), UV absorbance, and mass spectrometry. Related metabolites were
not observed by UV absorbance or extracting masses.

Figure S4, related to figure 2: A. Alignment of predicted precursor peptides from the from gram-positive and gram-
negative bacteria. MUSCLE alignment performed in Geneious 8.1.5. Abbreviations: LD = lantibiotic dehydratase;
O = oxidoreductase; M = methyl transferase; T = transporter. Entry 2 corresponds to an Amm6 homologue found in
a streptomycete genome carrying the lym biosynthetic gene cluster. Entries 3-5 correspond to Amm6 homologues
found in the lymphostin gen clusters from the genus Salinispora. B. Predicted biosynthetic gene clusters from Bacillus
halodurans C-125 (NC_002570, ORF locus tags ctg1_2032-2062) and Desmospora sp. 8437 (NZ_AFHT00000000,
ORF locus tags HMPREF9374_RS06800-RS06705).
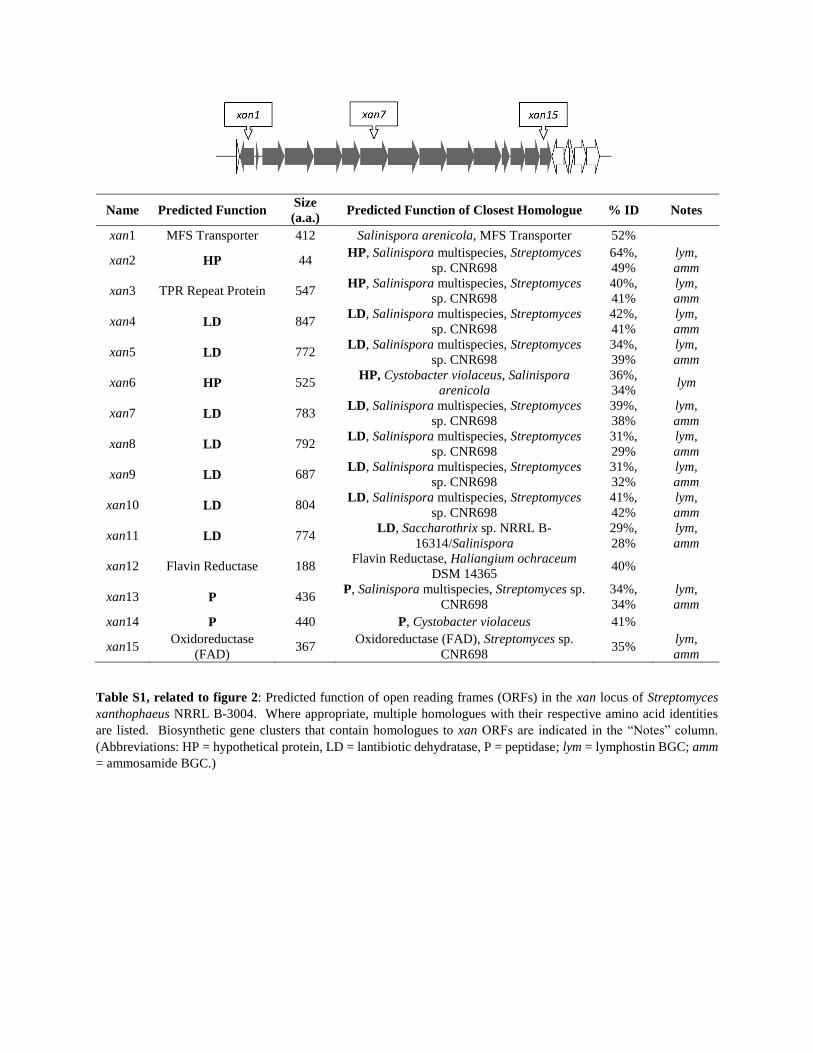
Name Predicted Function Size
(a.a.) Predicted Function of Closest Homologue % ID Notes
xan1 MFS Transporter 412 Salinispora arenicola, MFS Transporter 52%
xan2 HP 44 HP, Salinispora multispecies, Streptomyces
sp. CNR698
64%,
49%
lym,
amm
xan3 TPR Repeat Protein 547 HP, Salinispora multispecies, Streptomyces
sp. CNR698
40%,
41%
lym,
amm
xan4 LD 847 LD, Salinispora multispecies, Streptomyces
sp. CNR698
42%,
41%
lym,
amm
xan5 LD 772 LD, Salinispora multispecies, Streptomyces
sp. CNR698
34%,
39%
lym,
amm
xan6 HP 525 HP, Cystobacter violaceus, Salinispora
arenicola
36%,
34% lym
xan7 LD 783 LD, Salinispora multispecies, Streptomyces
sp. CNR698
39%,
38%
lym,
amm
xan8 LD 792 LD, Salinispora multispecies, Streptomyces
sp. CNR698
31%,
29%
lym,
amm
xan9 LD 687 LD, Salinispora multispecies, Streptomyces
sp. CNR698
31%,
32%
lym,
amm
xan10 LD 804 LD, Salinispora multispecies, Streptomyces
sp. CNR698
41%,
42%
lym,
amm
xan11 LD 774 LD, Saccharothrix sp. NRRL B-
16314/Salinispora
29%,
28%
lym,
amm
xan12 Flavin Reductase 188 Flavin Reductase, Haliangium ochraceum
DSM 14365 40%
xan13 P 436 P, Salinispora multispecies, Streptomyces sp.
CNR698
34%,
34%
lym,
amm
xan14 P 440 P, Cystobacter violaceus 41%
xan15 Oxidoreductase
(FAD) 367
Oxidoreductase (FAD), Streptomyces sp.
CNR698 35%
lym,
amm
Table S1, related to figure 2: Predicted function of open reading frames (ORFs) in the xan locus of Streptomyces
xanthophaeus NRRL B-3004. Where appropriate, multiple homologues with their respective amino acid identities
are listed. Biosynthetic gene clusters that contain homologues to xan ORFs are indicated in the “Notes” column.
(Abbreviations: HP = hypothetical protein, LD = lantibiotic dehydratase, P = peptidase; lym = lymphostin BGC; amm
= ammosamide BGC.)

Figure S5, related to the characterization of compounds 7 and 8 in the Experimental Procedures section:
Scheme for the synthesis of compounds 7 and 8.
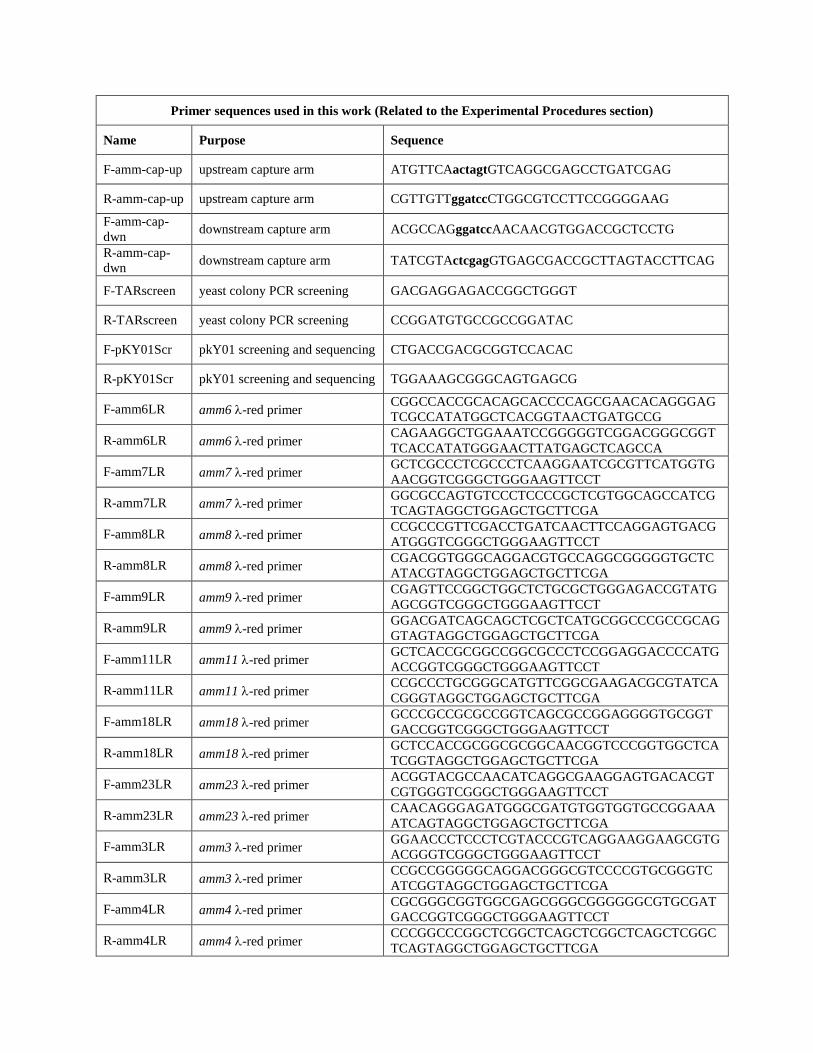
Primer sequences used in this work (Related to the Experimental Procedures section)
Name Purpose Sequence
F-amm-cap-up upstream capture arm ATGTTCAactagtGTCAGGCGAGCCTGATCGAG
R-amm-cap-up upstream capture arm CGTTGTTggatccCTGGCGTCCTTCCGGGGAAG
F-amm-cap-
dwn downstream capture arm ACGCCAGggatccAACAACGTGGACCGCTCCTG
R-amm-cap-
dwn downstream capture arm TATCGTActcgagGTGAGCGACCGCTTAGTACCTTCAG
F-TARscreen yeast colony PCR screening GACGAGGAGACCGGCTGGGT
R-TARscreen yeast colony PCR screening CCGGATGTGCCGCCGGATAC
F-pKY01Scr pkY01 screening and sequencing CTGACCGACGCGGTCCACAC
R-pKY01Scr pkY01 screening and sequencing TGGAAAGCGGGCAGTGAGCG
F-amm6LR amm6 -red primer CGGCCACCGCACAGCACCCCAGCGAACACAGGGAG
TCGCCATATGGCTCACGGTAACTGATGCCG
R-amm6LR amm6 -red primer CAGAAGGCTGGAAATCCGGGGGTCGGACGGGCGGT
TCACCATATGGGAACTTATGAGCTCAGCCA
F-amm7LR amm7 -red primer GCTCGCCCTCGCCCTCAAGGAATCGCGTTCATGGTG
AACGGTCGGGCTGGGAAGTTCCT
R-amm7LR amm7 -red primer GGCGCCAGTGTCCCTCCCCGCTCGTGGCAGCCATCG
TCAGTAGGCTGGAGCTGCTTCGA
F-amm8LR amm8 -red primer CCGCCCGTTCGACCTGATCAACTTCCAGGAGTGACG
ATGGGTCGGGCTGGGAAGTTCCT
R-amm8LR amm8 -red primer CGACGGTGGGCAGGACGTGCCAGGCGGGGGTGCTC
ATACGTAGGCTGGAGCTGCTTCGA
F-amm9LR amm9 -red primer CGAGTTCCGGCTGGCTCTGCGCTGGGAGACCGTATG
AGCGGTCGGGCTGGGAAGTTCCT
R-amm9LR amm9 -red primer GGACGATCAGCAGCTCGCTCATGCGGCCCGCCGCAG
GTAGTAGGCTGGAGCTGCTTCGA
F-amm11LR amm11 -red primer GCTCACCGCGGCCGGCGCCCTCCGGAGGACCCCATG
ACCGGTCGGGCTGGGAAGTTCCT
R-amm11LR amm11 -red primer CCGCCCTGCGGGCATGTTCGGCGAAGACGCGTATCA
CGGGTAGGCTGGAGCTGCTTCGA
F-amm18LR amm18 -red primer GCCCGCCGCGCCGGTCAGCGCCGGAGGGGTGCGGT
GACCGGTCGGGCTGGGAAGTTCCT
R-amm18LR amm18 -red primer GCTCCACCGCGGCGCGGCAACGGTCCCGGTGGCTCA
TCGGTAGGCTGGAGCTGCTTCGA
F-amm23LR amm23 -red primer ACGGTACGCCAACATCAGGCGAAGGAGTGACACGT
CGTGGGTCGGGCTGGGAAGTTCCT
R-amm23LR amm23 -red primer CAACAGGGAGATGGGCGATGTGGTGGTGCCGGAAA
ATCAGTAGGCTGGAGCTGCTTCGA
F-amm3LR amm3 -red primer GGAACCCTCCCTCGTACCCGTCAGGAAGGAAGCGTG
ACGGGTCGGGCTGGGAAGTTCCT
R-amm3LR amm3 -red primer CCGCCGGGGGCAGGACGGGCGTCCCCGTGCGGGTC
ATCGGTAGGCTGGAGCTGCTTCGA
F-amm4LR amm4 -red primer CGCGGGCGGTGGCGAGCGGGCGGGGGGCGTGCGAT
GACCGGTCGGGCTGGGAAGTTCCT
R-amm4LR amm4 -red primer CCCGGCCCGGCTCGGCTCAGCTCGGCTCAGCTCGGC
TCAGTAGGCTGGAGCTGCTTCGA
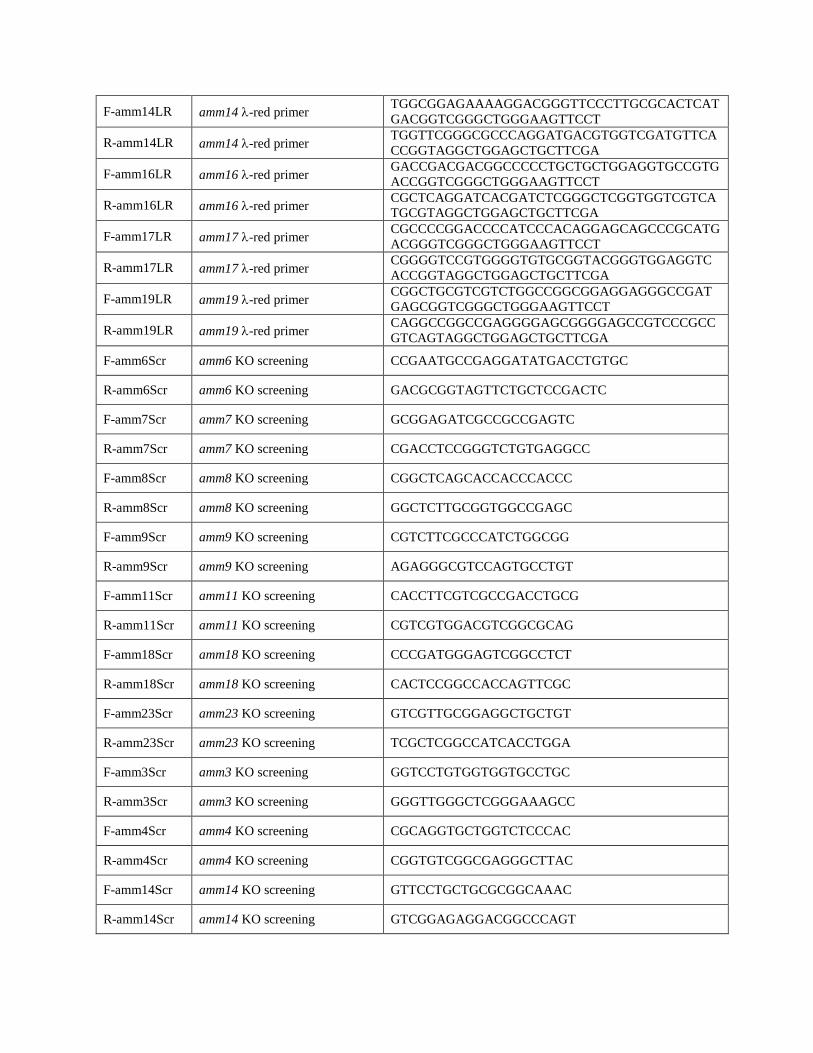
F-amm14LR amm14 -red primer TGGCGGAGAAAAGGACGGGTTCCCTTGCGCACTCAT
GACGGTCGGGCTGGGAAGTTCCT
R-amm14LR amm14 -red primer TGGTTCGGGCGCCCAGGATGACGTGGTCGATGTTCA
CCGGTAGGCTGGAGCTGCTTCGA
F-amm16LR amm16 -red primer GACCGACGACGGCCCCCTGCTGCTGGAGGTGCCGTG
ACCGGTCGGGCTGGGAAGTTCCT
R-amm16LR amm16 -red primer CGCTCAGGATCACGATCTCGGGCTCGGTGGTCGTCA
TGCGTAGGCTGGAGCTGCTTCGA
F-amm17LR amm17 -red primer CGCCCCGGACCCCATCCCACAGGAGCAGCCCGCATG
ACGGGTCGGGCTGGGAAGTTCCT
R-amm17LR amm17 -red primer CGGGGTCCGTGGGGTGTGCGGTACGGGTGGAGGTC
ACCGGTAGGCTGGAGCTGCTTCGA
F-amm19LR amm19 -red primer CGGCTGCGTCGTCTGGCCGGCGGAGGAGGGCCGAT
GAGCGGTCGGGCTGGGAAGTTCCT
R-amm19LR amm19 -red primer CAGGCCGGCCGAGGGGAGCGGGGAGCCGTCCCGCC
GTCAGTAGGCTGGAGCTGCTTCGA
F-amm6Scr amm6 KO screening CCGAATGCCGAGGATATGACCTGTGC
R-amm6Scr amm6 KO screening GACGCGGTAGTTCTGCTCCGACTC
F-amm7Scr amm7 KO screening GCGGAGATCGCCGCCGAGTC
R-amm7Scr amm7 KO screening CGACCTCCGGGTCTGTGAGGCC
F-amm8Scr amm8 KO screening CGGCTCAGCACCACCCACCC
R-amm8Scr amm8 KO screening GGCTCTTGCGGTGGCCGAGC
F-amm9Scr amm9 KO screening CGTCTTCGCCCATCTGGCGG
R-amm9Scr amm9 KO screening AGAGGGCGTCCAGTGCCTGT
F-amm11Scr amm11 KO screening CACCTTCGTCGCCGACCTGCG
R-amm11Scr amm11 KO screening CGTCGTGGACGTCGGCGCAG
F-amm18Scr amm18 KO screening CCCGATGGGAGTCGGCCTCT
R-amm18Scr amm18 KO screening CACTCCGGCCACCAGTTCGC
F-amm23Scr amm23 KO screening GTCGTTGCGGAGGCTGCTGT
R-amm23Scr amm23 KO screening TCGCTCGGCCATCACCTGGA
F-amm3Scr amm3 KO screening GGTCCTGTGGTGGTGCCTGC
R-amm3Scr amm3 KO screening GGGTTGGGCTCGGGAAAGCC
F-amm4Scr amm4 KO screening CGCAGGTGCTGGTCTCCCAC
R-amm4Scr amm4 KO screening CGGTGTCGGCGAGGGCTTAC
F-amm14Scr amm14 KO screening GTTCCTGCTGCGCGGCAAAC
R-amm14Scr amm14 KO screening GTCGGAGAGGACGGCCCAGT
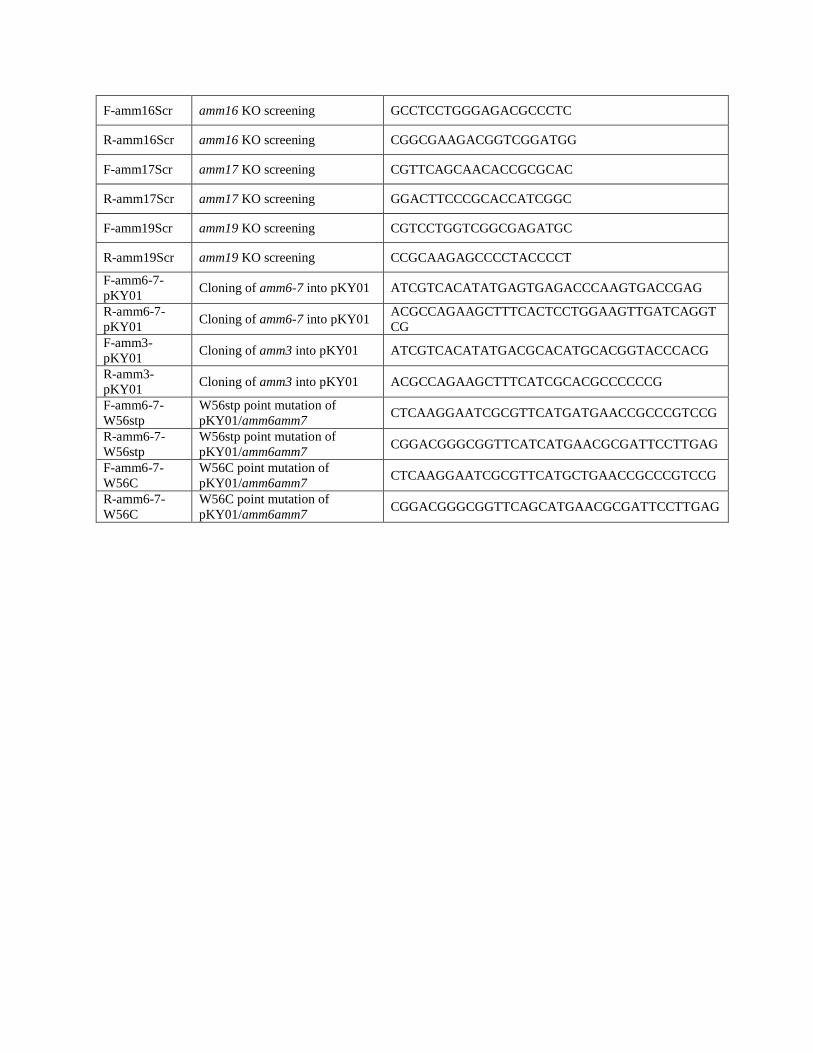
F-amm16Scr amm16 KO screening GCCTCCTGGGAGACGCCCTC
R-amm16Scr amm16 KO screening CGGCGAAGACGGTCGGATGG
F-amm17Scr amm17 KO screening CGTTCAGCAACACCGCGCAC
R-amm17Scr amm17 KO screening GGACTTCCCGCACCATCGGC
F-amm19Scr amm19 KO screening CGTCCTGGTCGGCGAGATGC
R-amm19Scr amm19 KO screening CCGCAAGAGCCCCTACCCCT
F-amm6-7-
pKY01 Cloning of amm6-7 into pKY01 ATCGTCACATATGAGTGAGACCCAAGTGACCGAG
R-amm6-7-
pKY01 Cloning of amm6-7 into pKY01
ACGCCAGAAGCTTTCACTCCTGGAAGTTGATCAGGT
CG
F-amm3-
pKY01 Cloning of amm3 into pKY01 ATCGTCACATATGACGCACATGCACGGTACCCACG
R-amm3-
pKY01 Cloning of amm3 into pKY01 ACGCCAGAAGCTTTCATCGCACGCCCCCCG
F-amm6-7-
W56stp
W56stp point mutation of
pKY01/amm6amm7 CTCAAGGAATCGCGTTCATGATGAACCGCCCGTCCG
R-amm6-7-
W56stp
W56stp point mutation of
pKY01/amm6amm7 CGGACGGGCGGTTCATCATGAACGCGATTCCTTGAG
F-amm6-7-
W56C
W56C point mutation of
pKY01/amm6amm7 CTCAAGGAATCGCGTTCATGCTGAACCGCCCGTCCG
R-amm6-7-
W56C
W56C point mutation of
pKY01/amm6amm7 CGGACGGGCGGTTCAGCATGAACGCGATTCCTTGAG

Supplemental Experimental Procedures
General
All chemicals and reagents were acquired from Fisher Scientific, VWR Scientific, Sigma-Aldrich and used without
further purification. Restrictions enzymes, T4 DNA ligase, Gibson Assembly Master Mix were purchased from New
England Biolabs. PrimeSTAR DNA polymerase from Clontech was used for PCR reactions. Antibiotics for bacterial
selection were used at the following concentrations: Kanamycin (50 µg/mL), Apramycin (50 µg/mL),
Chloramphenicol (25 µg/mL) and Nalidixic Acid (25 µg/mL). All solvents used for LC purposes were LCMS grade
and all solvents for preparative and semi-preparative reverse phase purifications were HPLC grade. LC-MS/MS
analyses were performed on an Agilent 6530 Accurate-Mass Q-TOF MS equipped with a dual electrospray ionization
source and an Agilent 1260 LC system with diode array detector and Phenomenex Kinetex 2.6μ XB-C18 100 A, 150
x 4.6 mm column. MS and UV data were analyzed with Agilent MassHunter Qualitative Analysis version B.05.00.
Preparative HPLC was carried out using an Agilent 218 purification system (ChemStation software, Agilent) equipped
with a ProStar 410 automatic injector, Agilent ProStar UV-Vis Dual Wavelength Detector, a 440-LC fraction collector
and preparative HPLC column indicated below. Semi-preparative HPLC purifications were performed on an Agilent
1260 Series Instrument with a multiple wavelength detector and Phenomenex Luna 5µm C8(2) 250x100 mm semi
preparative column. Unless otherwise specified, all HPLC purifications utilized a 0.1% trifluoracetic acid buffer and
all analytical LCMS methods included a 0.1% formic acid buffer. NMR data were acquired at the UCSD Skaggs
School of Pharmacy, using a 600 MHz Bruker Avance III spectrometer with a 1.7 mm cryoprobe and at the UCSD
Chemistry Department NMR facility on a 500 MHz Varian XSens 5 mm cryoprobe. All signals are reported in ppm
with the internal DMSO-d6 signal at 2.50 ppm or 39.52 ppm. The data is being reported as (s=singlet, d=doublet,
t=triplet, q=quadruplet, m=multiplet or unresolved, br=broad signal, coupling constant(s) in Hz, integration).
Design and Construction of the amm Specific Capture Vector
To directly capture the 37.5 kb region encompassing the ammosamide (amm) biosynthetic gene cluster, we followed
the general procedure described by Yamanaka et al.(Yamanaka et al., 2014) Briefly, 1kb capture arms corresponding
to the upstream and downstream regions of the amm pathway were PCR amplified from Streptomyces sp. CNR-698
genomic DNA using the following primers: F-amm-cap-up, R-amm-cap-up, F-amm-cap-dwn, R-amm-cap-dwn (see
Table S2) with the included SpeI (actagt), XhoI (ctcgag), and BamHI (ggatcc) restriction sites. The two 1 kb fragments
were then PCR assembled into a single 2 kb fragment with F-amm-cap-up and R-amm-cap-dwn, and the assemble
fragment was ligated into pCAP01 using the SpeI and XhoI restriction sites. Prior to TAR capture, the pCAP01/amm
vector was linearized with BamHI.
Direct Capture of the amm gene cluster using transformation associated recombination
Streptomyces sp. CNR-698 genomic DNA was digested with EcoRI, NdeI and NheI-HF prior to its use in the TAR-
capture as described by Yamanaka et al.(Yamanaka et al., 2014) CNR-698 gDNA was also randomly fragmented
using a pipette tip, however capture experiments using this gDNA fragmentation method did not yield any positive
yeast clones Out of 100 colonies screened, where restriction digested gDNA was used, 3 positive hits were identified
by colony PCR using primers for a 1 kb region of amm3 (see Table S3). Candidates from PCR positive clones were
introduced into E. coli Top 10 by electroporation and evaluated by NcoI restriction digestion. The captured construct
was designated pCAP01/amm.
Calculation of RNA secondary structure downstream of amm6
The RNA secondary structure beginning 8 bases downstream of amm6 stop codon consisted of a stem-loop structure
comprising 15 bases, followed by a second stem loop comprising 50 bases. The structure was calculated using the
mFold server (http://unafold.rna.albany.edu/?q=mfold/RNA-Folding-Form) (Zuker, 2003), and RNAstructure
(http://rna.urmc.rochester.edu/RNAstructureWeb/Servers/Predict1/Predict1.html) (Reuter and Mathews, 2010).The
calculated lowest free energy structure, and G of folding were very similar-57.7 kcal/mol (mFold) and -57.4
kcal/mol. (RNAstructure). Minimum free energy calculations were performed at 310.14 K (default) with all other
parameter set at default.
Synthesis of acid 8
The synthesis of compound 9 (Figure S5) was previously reported.(Reddy et al., 2010) Boc protected 9 (29.6 mg,
0.060 mmol) was dissolved in 1.5 mL DCM and 140 µL TFA (1.8 mmol) was added. After stirring for 6 hours at

room temperature, TFA and solvent were removed with a stream of N2 followed by high vacuum. To the resulting
residue was added 6 mL tetrahydrofuran and 1.2 mL 2M LiOH. The reaction was stirred for 1 hour at 40 degrees then
quenched with 0.37 mL TFA (4.8 mmol). The reaction was then poured into a separatory funnel, acidified with 1N
HCl and extracted three times with ethyl acetate. Then organics were dried with acidified brine, followed by
anhydrous sodium sulfate then concentrated. The resulting residue was then purified by preparative HPLC on an
Agilent Pursuit XRS 5 μm C18 100x21.2 mm preparative HPLC column (0-1 min 5% ACN isocratic, 1-10 min 5-
15% ACN at 15 mL/min with a mobile phase buffered with 12.5 mM NH4OH). One third of the purified material was
frozen and lyophilized to yield 2.3 mg of 8 as the ammonium salt. For the other 2/3 of the pure fractions, they were
pooled and the acetonitrile removed by rotovap. The resulting aqueous phase was acidified with HCl and extracted
three times, dried with acidified brine and anhydrous sodium sulfate before. The dried organics were concentrated to
yield 6.3 mg of the free acid. 1H-NMR (500 MHz, DMSO) δ 10.25 (s, 1H), 8.31 (s, 1H), 6.56 (s, 2H), 6.40 (s, 2H). 13C-NMR (126 MHz, dmso) δ 165.7, 164.7, 140.2, 132.5, 131.9, 131.3, 120.1, 117.5, 104.5, 103.9. LCMS analysis
and 1H- and 13C-NMR matched 8 isolated from M512-pCAP01/ammamm3 + pKY01/amm3.
Synthesis of amide 7
The synthesis of compound 10 (Figure S5) was described previously.(Reddy et al., 2010) 2.4 mg of (0.008 mmol)
10 was added 2 mL THF and 200 µL 28% NH4OH. The reaction was stirred at room temperature for 24 hours where
it was determined to be ~40% complete. An additional 200 µL of 28% NH4OH was added and the reaction was then
heated at 40 degrees for 24 hours where it was determined complete. The reaction was diluted with water in separatory
funnel and extracted 3 times with ethyl acetate. The combined organics were dried with brine then sodium sulfate and
concentrated. The resulting residue was then purified by preparative HPLC on an Agilent Pursuit XRS 5 μm C18
100x21.2 mm preparative HPLC column (0-1 min 10% ACN isocratic, 1-15 min 10-58% ACN at 15 mL/min with a
mobile phase buffered with 0.1% trifluoroacetic acid) to yield 0.8 mg. 1H-NMR (500 MHz, DMSO) δ 10.14 (s, 1H),
8.85 (s, 1H), 8.31 (s, 1H), 7.60 (s, 1H), 6.58 (s, 2H), 6.17 (s, 2H). 13C-NMR (126 MHz, DMSO) δ 166.3, 165.1, 144.5,
140.1, 131.7, 131.4, 131.1, 120.0, 115.5, 104.4, 103.7. LCMS analysis and 1H- and 13C-NMR matched 7 isolated
from M512-pCAP01/ammamm23.
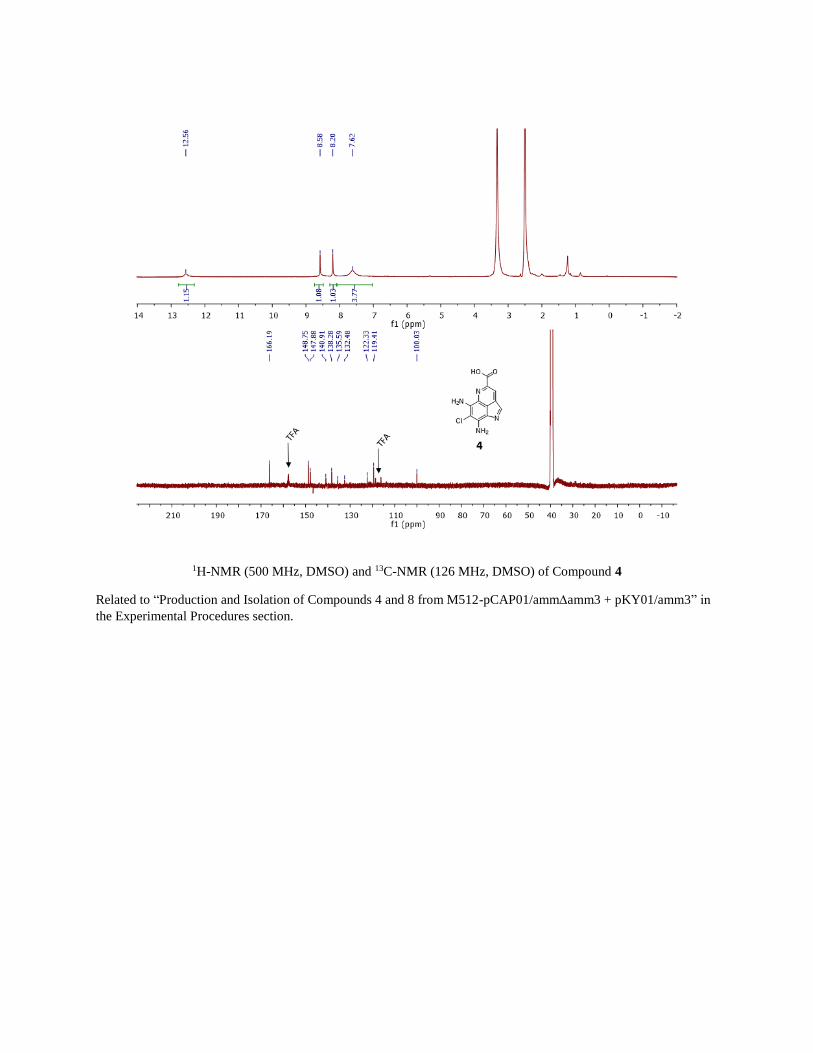
1H-NMR (500 MHz, DMSO) and 13C-NMR (126 MHz, DMSO) of Compound 4
Related to “Production and Isolation of Compounds 4 and 8 from M512-pCAP01/ammamm3 + pKY01/amm3” in
the Experimental Procedures section.
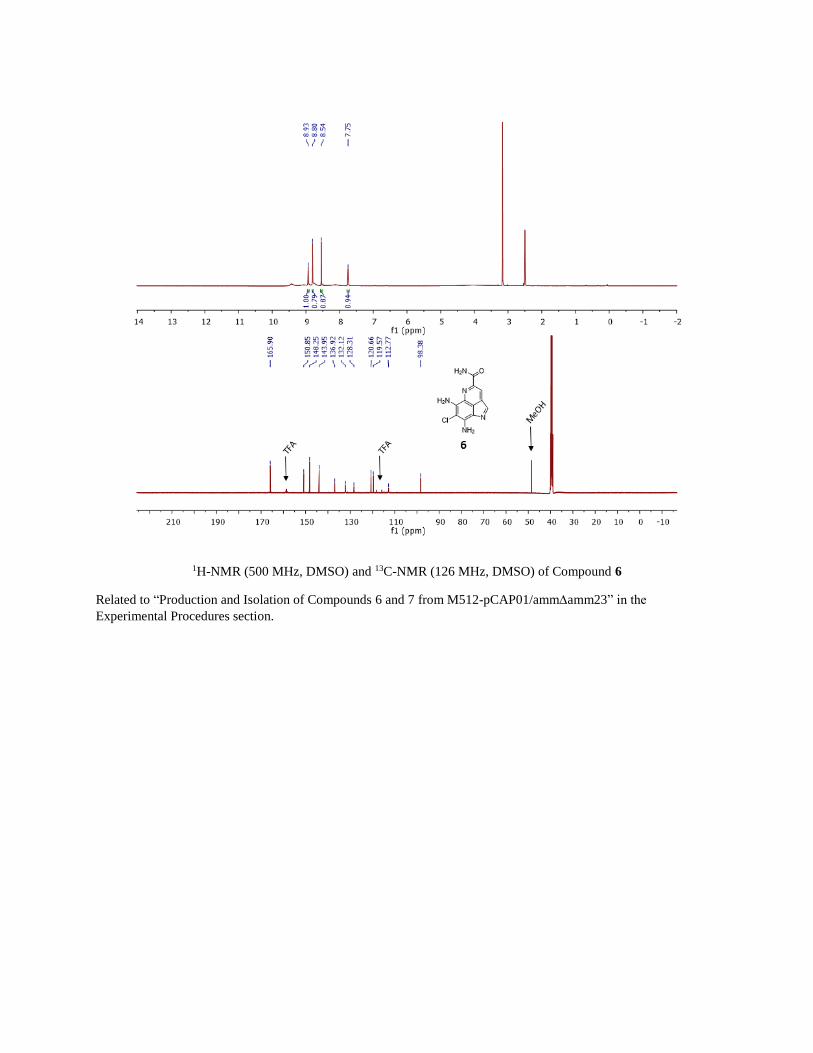
1H-NMR (500 MHz, DMSO) and 13C-NMR (126 MHz, DMSO) of Compound 6
Related to “Production and Isolation of Compounds 6 and 7 from M512-pCAP01/ammamm23” in the
Experimental Procedures section.
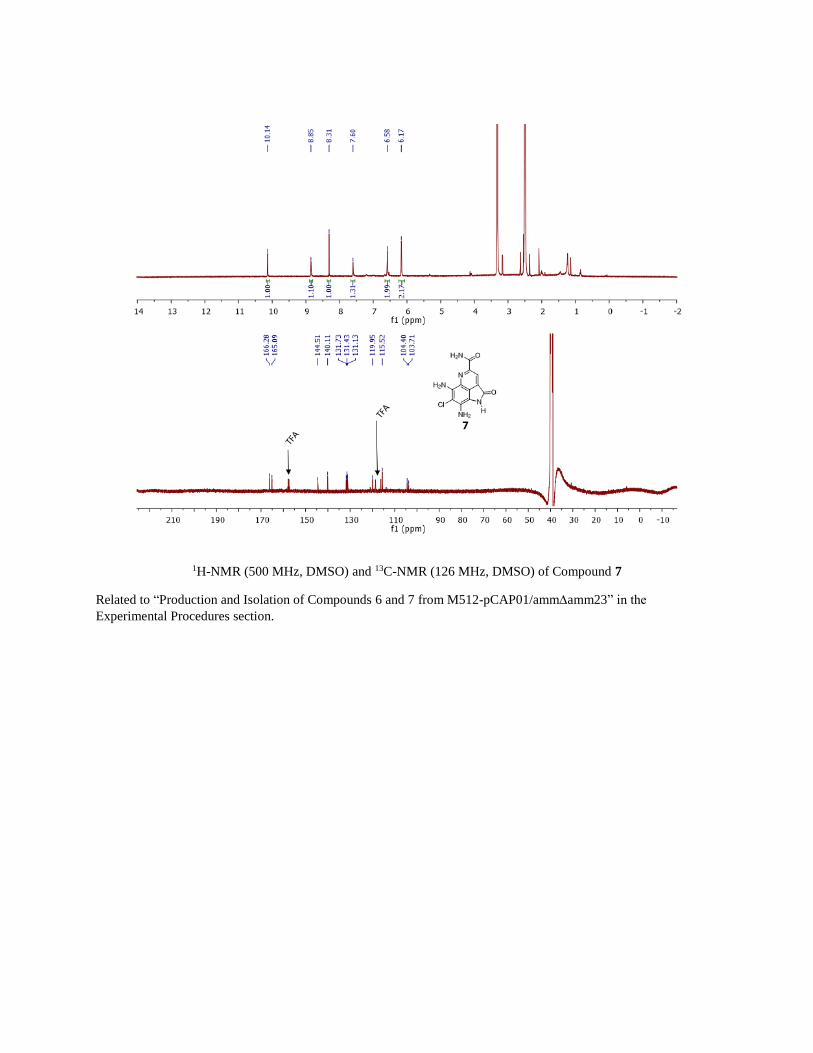
1H-NMR (500 MHz, DMSO) and 13C-NMR (126 MHz, DMSO) of Compound 7
Related to “Production and Isolation of Compounds 6 and 7 from M512-pCAP01/ammamm23” in the
Experimental Procedures section.
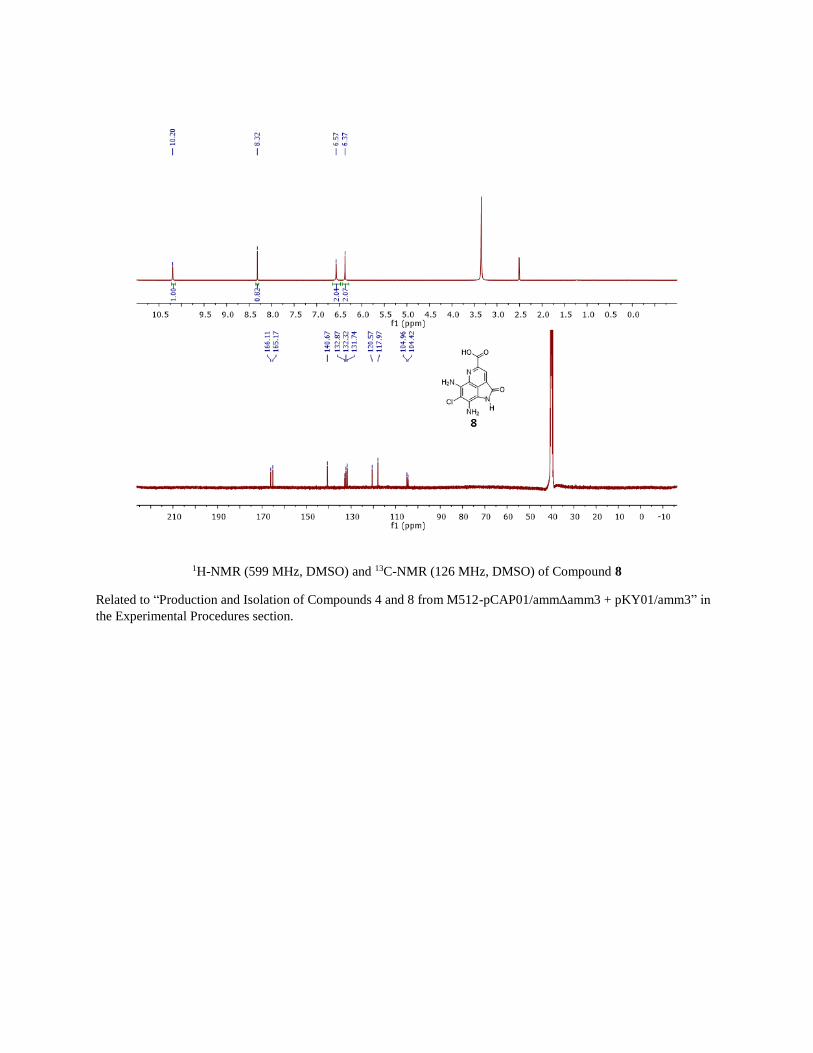
1H-NMR (599 MHz, DMSO) and 13C-NMR (126 MHz, DMSO) of Compound 8
Related to “Production and Isolation of Compounds 4 and 8 from M512-pCAP01/ammamm3 + pKY01/amm3” in
the Experimental Procedures section.
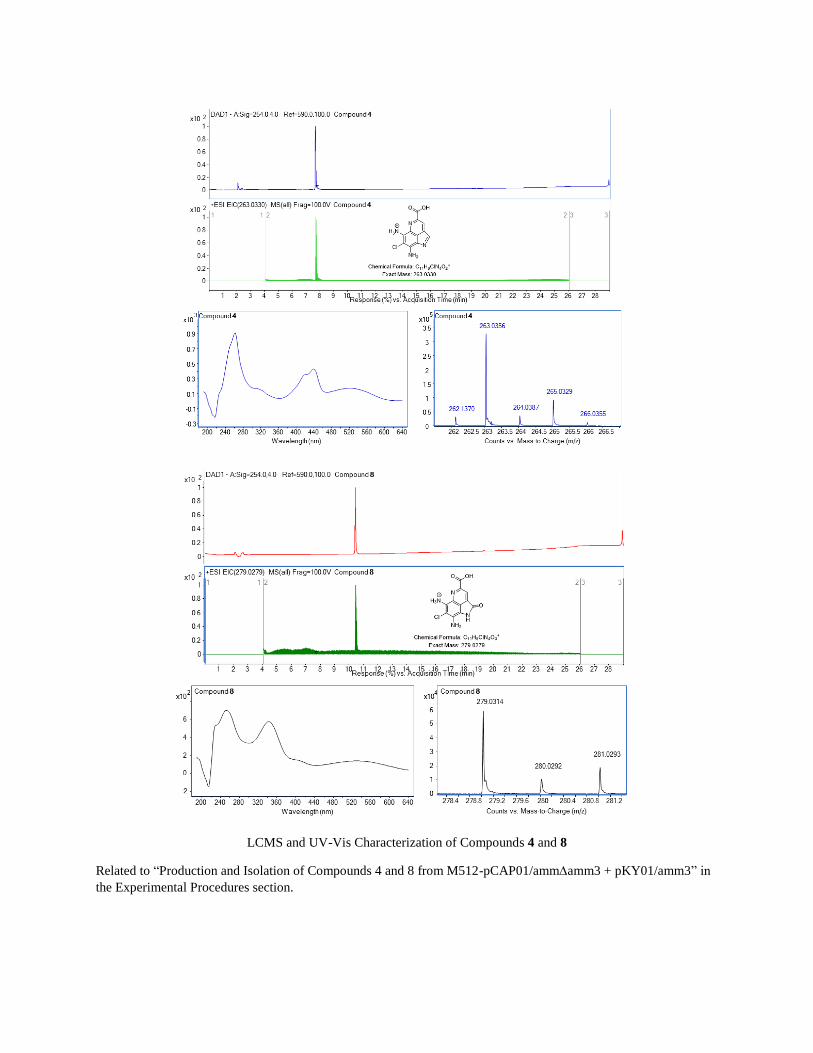
LCMS and UV-Vis Characterization of Compounds 4 and 8
Related to “Production and Isolation of Compounds 4 and 8 from M512-pCAP01/ammamm3 + pKY01/amm3” in
the Experimental Procedures section.
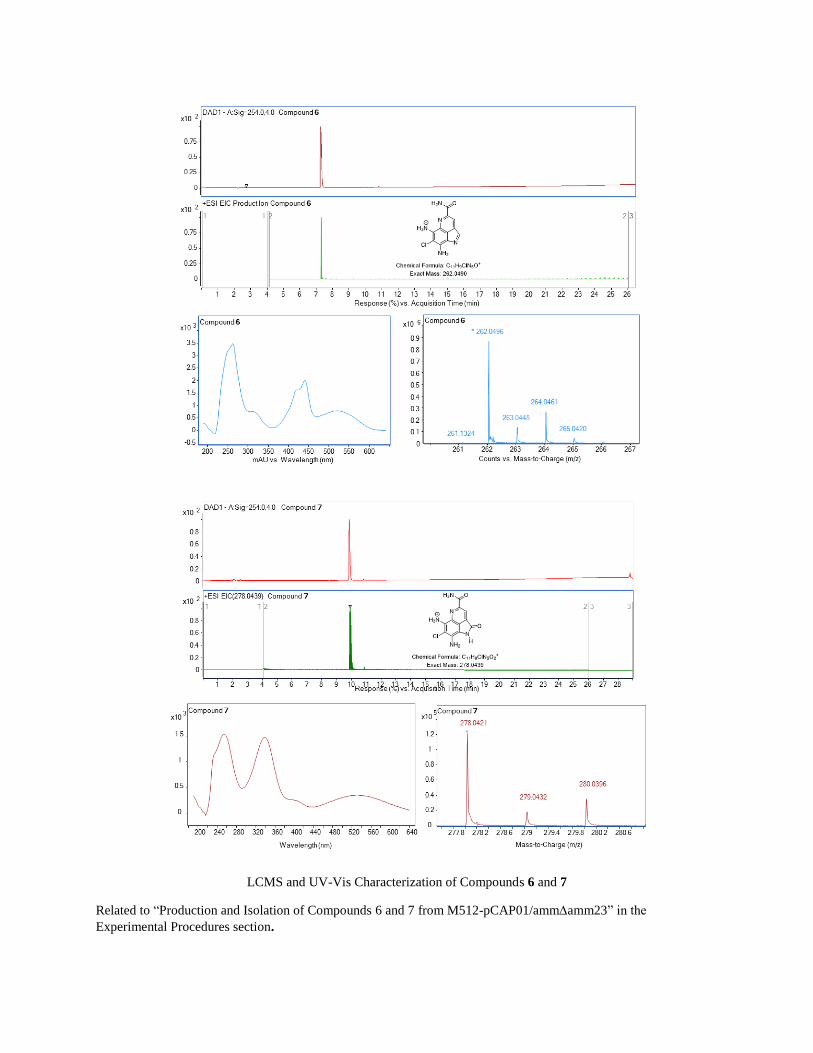
LCMS and UV-Vis Characterization of Compounds 6 and 7
Related to “Production and Isolation of Compounds 6 and 7 from M512-pCAP01/ammamm23” in the
Experimental Procedures section.

Reddy, P.V.N., Banerjee, B., and Cushman, M. (2010). Efficient Total Synthesis of Ammosamide B. Org. Lett. 12, 3112-3114. Reuter, J.S., and Mathews, D.H. (2010). RNAstructure: software for RNA secondary structure prediction and analysis. Bmc Bioinformatics 11, 9. Yamanaka, K., Reynolds, K.A., Kersten, R.D., Ryan, K.S., Gonzalez, D.J., Nizet, V., Dorrestein, P.C., and Moore, B.S. (2014). Direct cloning and refactoring of a silent lipopeptide biosynthetic gene cluster yields the antibiotic taromycin A. Proc. Natl. Acad. Sci. U.S.A. 111, 1957-1962. Zuker, M. (2003). Mfold web server for nucleic acid folding and hybridization prediction. Nucleic Acids Res. 31, 3406-3415.