bohn chemosphere 2012 - publications...
TRANSCRIPT

Determination of oral uptake and biodistribution of platinum and chromiumby the garden snail (Helix aspersa) employing nano-secondary ion
mass-spectrometry
Tanja Eybe a, Jean-Nicolas Audinot b, Thomas Udelhoven a, Esther Lentzen b, Brahime El Adib b,Johanna Ziebel a, Lucien Hoffmann a, Torsten Bohn a,⇑
aCentre de Recherche Public-Gabriel Lippmann, Environment and Agro-biotechnologies Department, 41 Rue du Brill, L-4422 Belvaux, LuxembourgbCentre de Recherche Public-Gabriel Lippmann, Material Sciences Department, 41 Rue du Brill, L-4422 Belvaux, Luxembourg
h i g h l i g h t s
" Platinum and chromium were detected in midgut gland, mantle, and shell by Nano-SIMS.
" Highest concentrations of heavy metals in midgut gland > mantle > shell.
" Accumulation in midgut gland and mantle was in form of 2–5 lm large agglomerations.
" Platinum and chromium distribution in shell appeared rather diffuse.
a r t i c l e i n f o
Article history:
Received 16 May 2012
Received in revised form 4 August 2012
Accepted 8 September 2012
Available online xxxx
Keywords:
Snails
Chromium
Platinum
Nano-SIMS
ICP-MS
Oral uptake
a b s t r a c t
Environmental heavy metal contamination is a case of concern for both animal and human health. Study-
ing the fate of metals in plant or animal tissues may provide information on pollution. In the present
study, we investigated the possibility to follow the biological fate of chromium and platinum uptake
in common garden snails (Helix aspersa), typically accumulating high concentrations of metals from their
environment. Chromium and platinum were administered orally to snails in 5 groups (n = 25/group):
control, food contaminated by ca. 2.5 lg gÿ1 and 19 lg gÿ1 chromium and 2.5 lg gÿ1 and 25 lg gÿ1
platinum, for 8 weeks. Following exposure, surviving snails were sacrificed, shell and remaining tissue
investigated by ICP-MS, and shell, midgut gland and mantle by nano-secondary ion mass-spectrometry
(Nano-SIMS). 12C14N-normalized platinum and 40Ca-normalized chromium measurements indicated
highest enrichments in cellular vesicles of the midgut gland, and lower concentrations in mantle and
shell, with significantly higher platinum and chromium concentrations in the 2 exposure groups vs.
control (P < 0.05), with somewhat differing distribution patterns for chromium and platinum. Compara-
ble results were obtained by ICP-MS, with both chromium and platinum fed snails showing drastically
elevated concentrations of metals in shell (up to 78 and 122 lg gÿ1 dw platinum and chromium, respec-
tively) and in other tissues (up to 200 and 1125 lg gÿ1 dw platinum and chromium, respectively). Nano-
SIMS allowed for semi-quantitative comparison of metal fate in snail tissues, making this an interesting
technique for future studies in the area of environmental pollution.
Ó 2012 Elsevier Ltd. All rights reserved.
1. Introduction
Many anthropogenic processes have resulted in the release of
‘‘heavy metals’’ into ecosystems, with consequential pollution.
Studying the fate of these metals in environmentally exposed
plants or animals may provide information on the degree of expo-
sure over time and may allow for comparing contamination be-
tween various areas. A potential candidate for monitoring the
fate of metals in the environment is the snail, which is known to
accumulate metals (Dallinger and Wieser, 1984; Notten et al.,
2005). For example, Helix pomatia (Roman snail) does avoid metal
excretion to minimize water losses, thus accumulating metals over
its lifetime (ca. 10 years). However, these snails can reduce the
concentration of undesired free metals such as copper by chelation
with proteins, i.e. metallothionein (MT) isoforms (Menta and Parisi,
2001) and/or intracellular compartmentalization (Dallinger, 1994).
0045-6535/$ - see front matter Ó 2012 Elsevier Ltd. All rights reserved.
http://dx.doi.org/10.1016/j.chemosphere.2012.09.038
Abbreviations: Nano-SIMS, nano-secondary ion mass-spectrometry; ICP-MS,
inductively coupled plasma mass-spectrometry; MT, metallothioneins.⇑ Corresponding author. Tel.: +352 470 261 480; fax: +352 470 264.
E-mail address: [email protected] (T. Bohn).
Chemosphere xxx (2012) xxx–xxx
Contents lists available at SciVerse ScienceDirect
Chemosphere
journal homepage: www.elsevier .com/locate /chemosphere
Please cite this article in press as: Eybe, T., et al. Determination of oral uptake and biodistribution of platinum and chromium by the garden snail (Helix
aspersa) employing nano-secondary ion mass-spectrometry. Chemosphere (2012), http://dx.doi.org/10.1016/j.chemosphere.2012.09.038

Two interesting metals released especially in recent years into
the environment are platinum and chromium. Platinum has been
employed in manifold applications, including its use in medicine
such as dentistry equipment or pharmaceutical industry for anti-
cancer medications, although its predominant capitalization rests
in its use within car catalytic converters (Hoppstock and Sures,
2004), containing approx. 0.08% platinum (Hoffman, 1989). Hot ex-
haust fumes can cause abrasion of platinum and emission into the
environment, via airborne particulate matter. Since its implemen-
tation for car catalytic converters in the 1970s, the platinum con-
centration in the environment has increased worldwide
(Barbante et al., 2001), and its concentrations in soil, dust and
plants in vicinity to heavily frequented streets often exceeds the
natural background level. For example, road dust of European
countries has shown maximum values of 2250 lg kgÿ1 (Gomez
et al., 2001). The toxicity of platinum however is still under discus-
sion. It is known that soluble platinum salts such as PtCl2 possess a
certain genotoxic potential, perhaps due to oxidative stress dam-
age (Migliore et al., 2002). Platinum also has the ability to form
strong complexes and could inhibit a variety of cellular functions
such as respiration (reviewed by Wiseman and Zereini (2009)).
Platinum has been further associated with the emergence of sev-
eral allergies (Burastero et al., 2009). Despite being a noble metal,
a certain bioavailability has been suggested, both via the oral route
(Colombo et al., 2008b) and the lung (Colombo et al., 2008a).
Similar to platinum, chromium has many industrial applica-
tions, due to its anti-corrosive properties, and environmental con-
tamination due to urban and industrial discharges has been
recognized (Losi et al., 1994). For example, manufactured gas plant
sites may have released significant amounts of chromium (Radike
et al., 2002). Relevant environmental concentrations have been
measured in various tissues, such as in rice plants, with up to
500 mg kgÿ1 (Mohanty et al., 2011). The toxicity of chromium de-
pends on its oxidation state. Especially the toxic potential of chro-
mate (VI) is rather high, due to increased solubility in aqueous
systems, and its ability to act as an oxidant. High chromium levels
in humans have been associated with several chronic diseases,
especially with cancer (Holmes et al., 2008), kidney complications
(Barbier et al., 2005), but also with allergic reactions such as by the
skin (Shelnutt et al., 2007).
Due to the potential threat to the environment, monitoring
these elements in plants or animals appears prudent. With respect
to snails, although there is a lack of knowledge concerning the
accumulation of platinum and chromium, the accumulation of
metals in general into snail tissues such as the shell is well known
(Jordaens et al., 2006). For example, metal storage in the midgut
gland (Menta and Parisi, 2001), producing digestive juices, and
mantle, the layer producing the shell, has been described (Chabi-
covsky et al., 2003).
However, following metal distribution in exposed animals is
analytically challenging. Individual metals are typically detected
with inductively coupled plasma mass spectrometry (ICP-MS),
inductively coupled plasma atomic emission spectrometry (ICP-
AES) or atomic absorption spectrometry (AAS) (Ammann, 2007).
However, all of these methods are destructive, and do not allow ex-
act localization of elements in biological tissues. Electron micros-
copy techniques on the other hand do not allow the localization
without ambiguity (Brandenberger et al., 2010), and do not allow
the detection of elements at low concentrations (<1%). Nano-sec-
ondary ion mass-spectrometry (Nano-SIMS) is a relatively new
method to detect pollutants such as xenobiotics in organisms
(Eybe et al., 2009). The investigated tissue or cells remain relatively
intact, and elemental distribution can be observed inside tissues or
even individual cells, at their original location.
Employing the NanoSIMS 50 (Cameca, Genevilliers, France)
with a lateral resolution of approximately 50 nm (Lechene et al.,
2006), and the IMS-6F SIMS (Cameca) with a lateral resolution of
ca. 200 nm (Migeon et al., 2011), allowing for the simultaneous
monitoring of up to five elements in selected tissues, the aim of
this study was to investigate the distribution of platinum and chro-
mium, two elements well detectable by Nano-SIMS, in the midgut
gland, mantle and shell of the common garden snail (Helix aspersa),
following exposure to these elements via the oral route, and to
evaluate the usefulness of studying their biological fate using this
relatively novel technique.
2. Materials and methods
2.1. Chemicals
Unless otherwise stated, all chemicals were obtained from
Sigma–Aldrich (St. Louis, MO, USA) and were of analytical grade
or superior. Platinum standard (1 g Lÿ1 or 10 g Lÿ1 in 2% and 5%
hydrochloric acid, respectively) was purchased from Ultra Scien-
tific (North Kingstown, RI, USA), chromium (1 g Lÿ1 and 10 g Lÿ1
as potassium chromate) was from Fluka Chemicals (Buchs, Swit-
zerland). Nitric acid (Plasma Pure, 67–70%) was from SCP Science
(Baie D’Urfé, Quebec, Canada), hydrogen peroxide (30%, trace anal-
ysis grade) was from Fisher (Schwerte, Germany). All resins used
for embedding were likewise from Sigma–Aldrich. For all experi-
ments, only 18 MX water (Millipore, Brussels, Belgium) was used.
2.2. Exposure experiments
The snail culture (garden snail, H. aspersa) was established at
our institute. One hundred and twenty-five young garden snails
were isolated directly after hatching and lived in plastic boxes
(10 � 10 cm) on 3 cm high, sterilized soil taken from the nature.
Calcium carbonate was offered in form of sepia chalk (Pitti, Rec-
klinghausen, Germany) in approximately 2 � 3 cm pieces ad libi-
tum. The snails were fed zucchini without peel ad libitum until
the age of 2 months. For the exposure experiments, they were sep-
arated into larger glass boxes (50 � 25 cm) with each 25 animals
per exposure group. These boxes likewise contained 3 cm high
sterilized soil. Each exposure group was weighed directly before
the experiments. For this purpose, all 25 animals were sprayed
with water for soil removal. The snails were dried carefully with
a tissue and weighed together.
The snails were then fed with fresh peeled zucchini (ca. 30 g
fresh material) containing the injected metal standards (Table 1),
three times a week (Monday, Wednesday and Friday), for a total
period of 2 months. For this purpose, approx. 100 microlitres of
the metal standards (1 and 10 g Lÿ1) were injected slowly with a
fine needle (0.4 � 20 mm, Terumo, Summerset, NJ) and syringe at
3–4 places in a ca. 30 g zucchini cube and left to equilibrate for
1 h. The exact amount of metal injected was determined by weigh-
ing the syringe and needle prior and after the injection. The zuc-
chini was offered to the snails on a watch-glass dish made of
quartz, accessible to all snails ad libitum. Pieces not consumed
by the snails were collected on the next feeding occasion, re-
weighed, and stored at ÿ20 °C until determination of the amount
of non-consumed metal by ICP-MS. After 2 months, all surviving
snails (Table 1) were weighed again and the average weight of
the remaining snails was determined. Afterwards, the snails were
sacrificed by chloroform vapour (0.5 mL of chloroform on a tissue
in a closed 25 mL plastic centrifuge tube (Sarstedt, Nümbrecht,
Germany)). The shell was then carefully removed with a scalpel
and washed with distilled water; the mantle edge and midgut
gland were either resected for embedding in an epoxy resin for
Nano-SIMS analysis, or the remaining pooled tissue without shell
kept frozen for ICP-MS analyses.
2 T. Eybe et al. / Chemosphere xxx (2012) xxx–xxx
Please cite this article in press as: Eybe, T., et al. Determination of oral uptake and biodistribution of platinum and chromium by the garden snail (Helix
aspersa) employing nano-secondary ion mass-spectrometry. Chemosphere (2012), http://dx.doi.org/10.1016/j.chemosphere.2012.09.038

2.3. Embedding of snail organs
From each exposure group, 3 animals were selected to be ana-
lysed by Nano-SIMS, one of small, one of medium, and one of larger
size, relatively to the total surviving snails. Samples were prepared
similar as described earlier (Eybe et al., 2008, 2009). In short,
freshly prepared organs were placed in 4% glutaraldehyde in
0.1 M phosphate buffered saline (PBS) over night. Following a sin-
gle washing step with PBS, the samples were fixed with 1% osmium
tetroxide for 1 h. After an additional washing step with PBS, the
samples were dehydrated stepwise with increasing concentrations
of acetone in water (30, 50, 70, 90, and 100% acetone). After allow-
ing the resin to infiltrate the tissue with two increasing concentra-
tions (50 and 66%) of epoxy resin (Epon 812 substitute) in acetone
for 1 h each, the samples were soaked in 100% resin over night and
then hardened for 24 h at 60 °C. The snail organs were cut to
500 nm semi-thin sections with an ultramicrotome (Leica Ultracut,
UCT, Le Pecq, France) and placed on silicon supporters (Siltronix,
Archamps, France) that were stored at room temperature in plastic
boxes, until Nano-SIMS analysis.
2.4. Embedding of shells
Pieces from the youngest part of the shells were prepared with a
protocol adapted fromMarkichet al. (2002). The shellwaswashed in
0.05 M Na-ethylenediaminetetraacetic acid (EDTA) to remove
adherent metals. After rinsing in deionized water, the pieces were
ovendried at 60 °C to constantweight. The shellwas thenembedded
with acrylic resin (Unicryl, Polysciences Inc. Eppelheim, Germany),
polished in decreasing steps (40, 15, 6, 3 up to 1 lm) using a carbide
paper with a lubricant (Mercaprex, Presi, France). After cleaning
with distilled water and drying at room temperature, the samples
were coated with a gold layer using amodular high vacuum coating
system (BAL-TEC, Halbergmoos, Germany) to allow for sufficient
conductivity of samples for Nano-SIMS analysis. During the ionic
bombardment, an electron gun was used to allow the evacuation
of the charges during the analysis (Wacey et al., 2010).
2.5. Microwave digestion (tissues and shell) for ICP-MS
Samples of snails were dried in an oven at 105 °C for several
hours, then shells and tissues were mineralized in a microwave
oven (Anton Paar Multiwave 3000, Hertford, UK). For this purpose,
up to 250 mg of a dry homogenized sample aliquot were weighted,
then 7 mL of nitric acid (67–70%) and 3 mL of hydrogen peroxide
(30%) were added. The microwave was programmed to alter power
from 0 to 1400W within 10 min, to rest at 1400W for 15 min and
then to 0 W for 20 min. The samples were finally diluted to 25 mL
with water, centrifuged, and diluted between 20 and 1000 times
according to the analysed element directly before ICP-MS analysis.
Zucchini samples were likewise oven-dried and aliquots homoge-
nized and mineralized and analysed under the same conditions
as the snail samples.
2.6. ICP-MS analyses of snail and zucchini
ICP-MS analyses were done on an ELAN DRC-e (Perkin–Elmer,
Waltham, MA, USA) equipped with a cross-flow nebulizer associ-
ated with a Scott spray chamber, ceramic injector and an AS-93
Plus autosampler. ICP-MS instrumental operating parameters and
settings for the ELAN DRC-e instrument were as follows: RF power:
1250W; plasma gas flow: 15 L minÿ1; nebulizer gas flow: ca.
0.9 L minÿ1; auxiliar gas flow: 1.2 L minÿ1; nebulizer: cross flow;
spray chamber: Scott; number of replicates: three; reaction gas:
5% H2 in Ar; reaction gas flow: 1 mL minÿ1. Standards were pre-
pared in solutions of 1% nitric acid. Standard solutions used were
ICP Multi Element Standard Solution VI CertiPUR (Merck, Darms-
tadt, Germany) for chromium, and platinum standard was from
Ultra Scientific. Each sample was measured in triplicate.
2.7. NanoSIMS 50 investigation for platinum
Measurements were performed with the NanoSIMS 50 in ras-
ter-imaging mode. A Cs+ primary ion beam (acceleration energy
+8 keV, current intensity 1.5 pA) was used for rasterizing over a
typically 40 � 40 lm area on the sample surface (polarized,
ÿ8 keV) with an expected spatial resolution of 150 nm (spot size).
The images were recorded with 256 � 256 pixels, with an acquisi-
tion time of 40 ms pixelÿ1 and with a parallel detection of five dif-
ferent negative secondary ions. For snail platinum measurements,12C3,
12C14N, 16O, 31P and 195Pt (33.8% natural abundance) signals
were simultaneously collected with five electron multipliers. The
instrument was tuned to obtain a mass resolution of M/DM =
4500. Mass calibration of Pt, carried out on a daily base, was
achieved using a Pt foil (Goodfellow, Cambridge, UK).
2.8. IMS-6F investigation for chromium
Analyses were performed with a modified IMS-6F SIMS instru-
ment (Cameca). This instrument was chosen as it allowed SIMS
Table 1
Study design and amount of metals consumed by the snails during the 2 months feeding period with zucchini. Metal standards were injected carefully into zucchini pieces
designated for consumption by snails (Helix aspersa).a
Platinum Chromium
Control Exposure 1 Exposure 2 Exposure 1 Exposure 2
n (snails) 25 25 25 25 25
Surviving snails after experiment (n) 16e 11 12 16 21
Weight of individual snail, average, beginning (g) 0.196 0.194 0.184 0.170 0.174
Weight of surviving snails, average, end (g) 0.930 0.936 0.645 0.667 0.688
% Weight gain 474 482 350 392 395
Metal concentration added (g Lÿ1) – 1.0b 10.0b 1.0c 10.0c
Concentration metal in food (lg gÿ1) <0.1 2.5 25 2.5 19
Number of snails investigated by Nano-SIMS 3 3 3 3 3
Number of snails investigated by ICP-MS 6 4 5 6 9
Total amount of metal given (lg) 0 1839 19950 2044 15403
Total amount of metal consumed (lg)d 0 1770 16404 1850 12722
a Native content of zucchini: <0.1 lg gÿ1 platinum and <0.1 lg gÿ1 chromium.b Applied dissolved in diluted HCl (2% and 5% for 1 and 10 g Lÿ1, respectively).c Applied as K2CrO4.d Estimated based on re-weighed amount of non-consumed zucchini.e From the remaining snails, 3 per group were selected for Nano-SIMS analyses, of which 1 was of small, 1 of middle, 1 of large size.
T. Eybe et al. / Chemosphere xxx (2012) xxx–xxx 3
Please cite this article in press as: Eybe, T., et al. Determination of oral uptake and biodistribution of platinum and chromium by the garden snail (Helix
aspersa) employing nano-secondary ion mass-spectrometry. Chemosphere (2012), http://dx.doi.org/10.1016/j.chemosphere.2012.09.038

measurements in the positive mode, employing a Ga+-source for
the primary ion beam (Migeon et al., 2011). The beam could be
focalized down to 100 nm. For this study, a Ga+ primary beam of
200 pA and an impact energy of 25.5 kV were employed. The
images were recorded with 256 � 256 pixels, a raster size of typi-
cally 40 � 40 lm and a counting time of 4 ms pixelÿ1. Under these
conditions, a lateral resolution of ca. 200 nm is expected. The 23Na,39K, 40Ca, 52Cr and 53Cr signals were collected sequentially by elec-
tron multiplier. Both isotopes of chromium (53Cr, 9.5% natural
abundance, 52Cr, 83.8% natural abundance) were recorded. To limit
mass interferences between the elemental ions and the organic
ions, the mass spectrometer was adjusted for individual species
so that interference-free ions could be sampled. For this purpose,
a conventional energy filtering (75 eV) was applied (MacRae
et al., 1993). Mass calibration of chromium, carried out on a daily
base, was achieved using reference standards (Sigma–Aldrich).
2.9. Normalization of platinum and chromium signals and statistical
interpretation
Of the three animals investigated by the NanoSIMS 50, one rep-
resentative image per animal was selected for 12C14N-normaliza-
tion of 195Pt intensities. Likewise, 3 pictures were chosen from
the IMS-6F measurements of 52Cr, and normalized by 40Ca. These
ions were used for normalization as they generally represented
the strongest signal detected by the NanoSIMS 50 and the IMS-
6F, respectively. Thus, three control images and each three images
of the two exposure levels were selected for the analysis, from the
midgut gland, the shell, and the mantle, respectively. For the statis-
tical analyses, the mean intensity of all positive signals within one
image was determined. The analyses were carried out using Image
J vs. 1.43 (National Institutes of Health, Bethesda, MD). Data were
then logarithmized for statistical comparisons in order to achieve a
normal distribution with equal variance of data, which was verified
by normality plots and box-plots. To compare mean intensity lev-
els between the exposure groups, a linear mixed model was devel-
oped with the logarithmized, normalized intensity values as
observed variable, and the metal (chromium and platinum), tissue
(midgut gland, mantle and shell) and the group (control, exposure
1 and exposure 2) as fixed factors. Upon finding significant interac-
tions, metal and tissue were kept constant and the effect of expo-
sure was investigated for each metal and tissue individually.
Following statistical Fisher-F tests, Bonferroni PostHoc tests were
conducted. The overall significance level was fixed to a = 0.05 (2-
sided).
3. Results
3.1. Snails
A large fraction of the snails survived the metal exposure, albeit
the survival rate between the groups differed. While survival rate
was 64% for the control group not receiving chromium or platinum
(Table 1), the survival rate was lower for the platinum fed groups
(44 and 48% for the low and the highly exposed group, respec-
tively) but similar to the chromium fed groups (64 and 84% for
the low and the highly exposed group, respectively). The develop-
ment of the average weight gain was highest for the control and
the lower platinum exposed group (close to 400% compared to ini-
tial weight), but similar to the other groups (350–400%).
3.2. ICP-MS determined metal content of exposed snails
Exposure of snails to both platinum and chromium resulted in
drastic increases of metals in all snail tissues, including shell and
other tissues (Table 2). Of the total amount platinum applied to
the 2 exposure groups (1840 and 19950 lg, respectively), 96.2%and 82.2% were consumed by the snails, respectively, and 15%
and 1.4% of the original doses were recovered in the snails, respec-
tively. Of the recovered platinum, about 19% were recovered in the
shell, vs. 81% in the remaining tissues in the first exposure group,
and 59% vs. 41% in the second exposure group, similar as for the
control group (58% of platinum recovered in the shell). While the
concentration of platinum in shells increased in the higher exposed
group, this was not the case for other tissues and the total uptake
(Table 2).
Of the total amount of chromium applied to the 2 exposure
groups (2044 and 15403 lg), 91% and 83% were consumed by
the snails, respectively, and 44% and 0.8% were recovered in the
snails, respectively. Of the recovered chromium, a somewhat lower
fraction compared to platinum was recovered in the shell, about
14% vs. 86% in the remaining tissues in the first exposure group,
and 22% vs. 78% in the second exposure group. The control shells
contained higher proportions of chromium, about 44%. For both
shell and other tissues, higher chromium levels were found in
the group exposed to the lower chromium concentration.
3.3. Nano-SIMS measurements
In general, the Nano-SIMS evaluation was in accordance with
the ICP-MS measurements, showing increased concentrations of
both platinum and chromium in the shell and in tissues following
the feeding experiments (Figs. 1–4), with the highest number of
positive signals in the midgut gland followed by the mantle fol-
lowed by the shell. However, a 12C14N normalization and statistical
comparison were not deemed meaningful between the different
tissues due to different concentrations and distribution of carbon
and calcium.
Measuring additional ions including those generated by CN, P, O
(together with Pt), and Na, K and Ca (together with Cr) allowed to
differentiate structures similar as by light microscopy, including
cell membranes and nuclei (Eybe et al., 2008). Structures of the
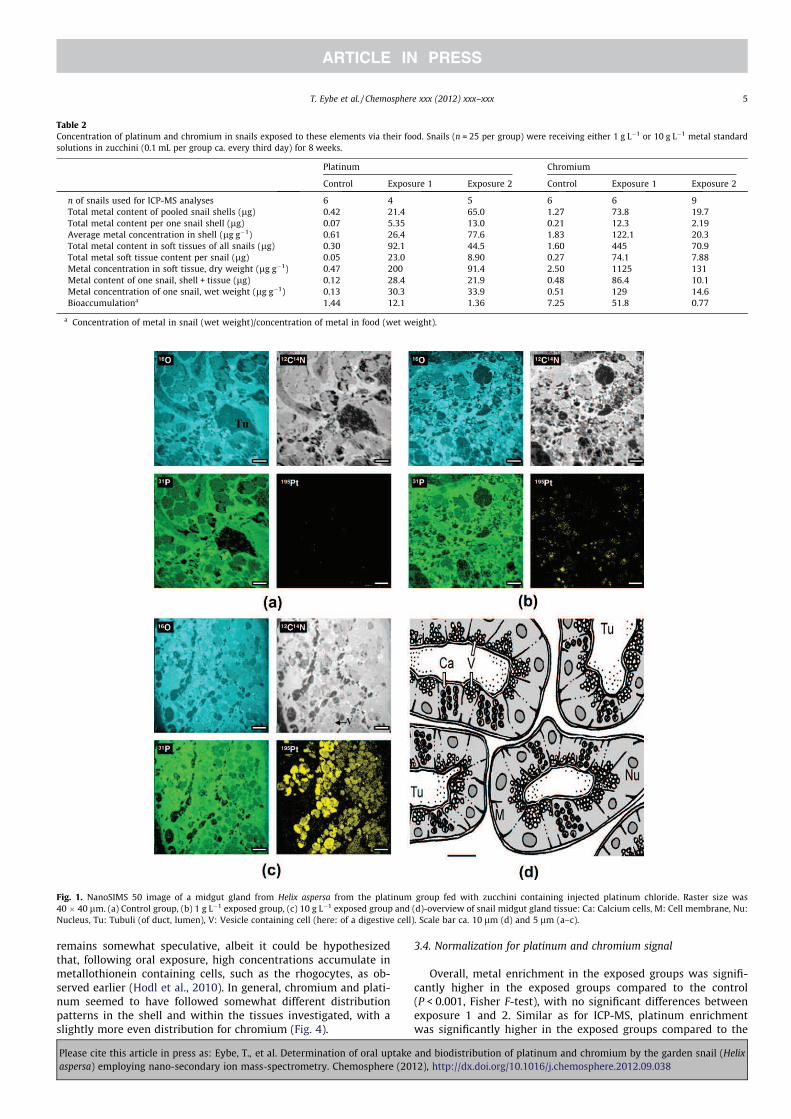
midgut gland further included tubuli (black spots neither rich in31P, 195Pt, 16O, and 12C14N, Fig. 1). Vesicles rich in platinum (ca.
2 lm diameter) were typically associated with higher phosphate
concentrations (Fig. 1). Additional visible cell structure compo-
nents included structures containing several vesicles rich in oxy-
gen and cluster carbon–nitrogen and platinum, which could be
basophilic cells (Fig. 1c).
Concerning chromium, 1–4 lm vesicle spots rich in 40Ca, 39K,23Na, 195Pt and 52Cr were clearly visible, albeit Ca, Na and K rich
structures, possibly granules within calcium cells, were generally
not the same locations that accumulated high amounts of Cr
(Fig. 4).
It appears that compared to controls, certain cellular structures
including tubuli were somewhat shrunken in the metal exposed
animals (Fig. 1), perhaps due to increased turnover of cells associ-
ated with detoxification, a stage that has also been termed as
‘‘shrinkage necrosis’’ (Cobb et al., 1996).
Concerning the mantle, the tissue producing the shell, certain
spots with concentrated amounts of platinum and chromium are
visible; however, their affiliation to a certain cellular tissue was
less clear, and spots were fewer and generally smaller (not above
1 lm) compared to the midgut gland agglomerations, at least for
platinum, while this was less apparent for chromium. In Fig. 2a
and c, nuclei of ca. 5 lm diameter were visible. For the shell, it
was difficult to spot any structures. Due to the absence of cells,
the distribution was more uniform, with much smaller and sparse
agglomeration spots.
Thus, the exact location of platinum and chromium enrichment
in the tissue and cells, especially for the mantle and the shell,
4 T. Eybe et al. / Chemosphere xxx (2012) xxx–xxx
Please cite this article in press as: Eybe, T., et al. Determination of oral uptake and biodistribution of platinum and chromium by the garden snail (Helix
aspersa) employing nano-secondary ion mass-spectrometry. Chemosphere (2012), http://dx.doi.org/10.1016/j.chemosphere.2012.09.038
remains somewhat speculative, albeit it could be hypothesized
that, following oral exposure, high concentrations accumulate in
metallothionein containing cells, such as the rhogocytes, as ob-
served earlier (Hodl et al., 2010). In general, chromium and plati-
num seemed to have followed somewhat different distribution
patterns in the shell and within the tissues investigated, with a
slightly more even distribution for chromium (Fig. 4).
3.4. Normalization for platinum and chromium signal
Overall, metal enrichment in the exposed groups was signifi-
cantly higher in the exposed groups compared to the control
(P < 0.001, Fisher F-test), with no significant differences between
exposure 1 and 2. Similar as for ICP-MS, platinum enrichment
was significantly higher in the exposed groups compared to the
Fig. 1. NanoSIMS 50 image of a midgut gland from Helix aspersa from the platinum group fed with zucchini containing injected platinum chloride. Raster size was
40 � 40 lm. (a) Control group, (b) 1 g Lÿ1 exposed group, (c) 10 g Lÿ1 exposed group and (d)-overview of snail midgut gland tissue: Ca: Calcium cells, M: Cell membrane, Nu:
Nucleus, Tu: Tubuli (of duct, lumen), V: Vesicle containing cell (here: of a digestive cell). Scale bar ca. 10 lm (d) and 5 lm (a–c).
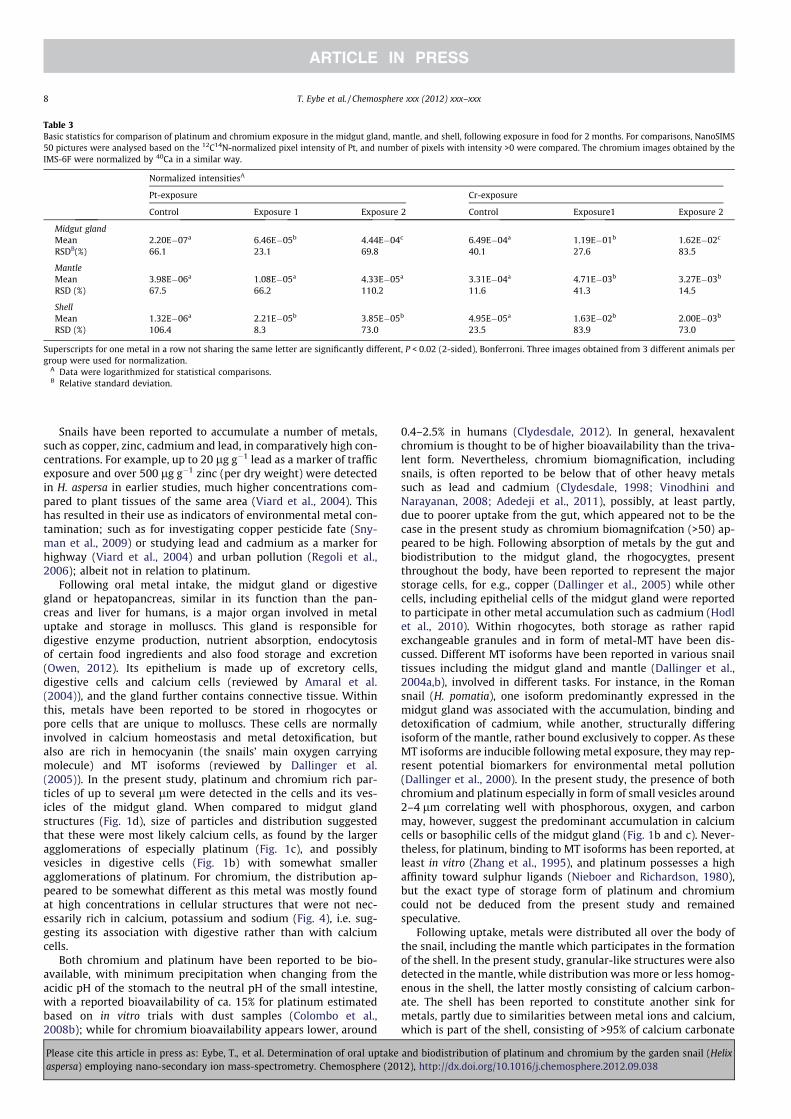
Table 2
Concentration of platinum and chromium in snails exposed to these elements via their food. Snails (n = 25 per group) were receiving either 1 g Lÿ1 or 10 g Lÿ1 metal standard
solutions in zucchini (0.1 mL per group ca. every third day) for 8 weeks.
Platinum Chromium
Control Exposure 1 Exposure 2 Control Exposure 1 Exposure 2
n of snails used for ICP-MS analyses 6 4 5 6 6 9
Total metal content of pooled snail shells (lg) 0.42 21.4 65.0 1.27 73.8 19.7
Total metal content per one snail shell (lg) 0.07 5.35 13.0 0.21 12.3 2.19
Average metal concentration in shell (lg gÿ1) 0.61 26.4 77.6 1.83 122.1 20.3
Total metal content in soft tissues of all snails (lg) 0.30 92.1 44.5 1.60 445 70.9
Total metal soft tissue content per snail (lg) 0.05 23.0 8.90 0.27 74.1 7.88
Metal concentration in soft tissue, dry weight (lg gÿ1) 0.47 200 91.4 2.50 1125 131
Metal content of one snail, shell + tissue (lg) 0.12 28.4 21.9 0.48 86.4 10.1
Metal concentration of one snail, wet weight (lg gÿ1) 0.13 30.3 33.9 0.51 129 14.6
Bioaccumulationa 1.44 12.1 1.36 7.25 51.8 0.77
a Concentration of metal in snail (wet weight)/concentration of metal in food (wet weight).
T. Eybe et al. / Chemosphere xxx (2012) xxx–xxx 5
Please cite this article in press as: Eybe, T., et al. Determination of oral uptake and biodistribution of platinum and chromium by the garden snail (Helix
aspersa) employing nano-secondary ion mass-spectrometry. Chemosphere (2012), http://dx.doi.org/10.1016/j.chemosphere.2012.09.038

non-exposed groups (P < 0.001, Fisher F-test) when all 3 tissues
were combined. When investigated per tissue, only the differences
for the midgut gland and the shell were significantly different be-
tween exposure groups (P < 0.001 and P = 0.005, Fisher-F test,
respectively), while there was a trend (P = 0.057) for higher plati-
num concentrations in the mantle at higher exposure concentra-
tions. However, albeit not exactly encompassing the same
tissues, different to ICP-MS, an increasingly high concentration of
platinum of exposure group 2 > 1 > control was found by the Nano-
SIMS50 (Table 3), at least when combining all tissues (P < 0.031,
Fisher F-test) and for the midgut gland alone (P < 0.020, Bonferron-
i), while ICP-MS did not suggest increased tissue concentrations of
combined digested tissues when comparing exposure group 2 vs. 1
(Table 2).
Chromium enrichment was significantly higher in the exposed
groups compared to the non-exposed groups (P < 0.001, Fisher F-
test) when all 3 tissues were combined, with highest intensities
for exposure group 1 > 2 > control (P < 0.001, Bonferroni, Table 3),
being in line with general results obtained by ICP-MS. When inves-
tigated per tissue, differences between exposure groups for the
midgut gland, mantle and the shell were significantly different
(P < 0.001, Fisher-F test). An increasingly high concentration of
chromium in exposure groups 2 > 1 > control was found for the
midgut gland alone (P = 0.009, Bonferroni).
4. Discussion
In the present study, we have shown that the Nano-SIMS tech-
nique was a useful tool to follow the fate of platinum and chro-
mium, metals correlated with automobile and general industrial
activity, after oral uptake by the garden snail H. aspersa. Both met-
als were detected in several snail tissues by this technique, includ-
ing the shell, mantle, and midgut gland, and results obtained by
Nano-SIMS were generally in agreement with uptakes determined
by ICP-MS. This confirmation was further based on semi-quantita-
tive uptake comparisons following 12C14N-normalization of 195Pt
and 40Ca-normalization of 52Cr. Employing Nano-SIMS, it was fur-
ther possible to highlight that agglomeration of metals did not fol-
low an even distribution, but that chromium and platinum were
found in certain vesicles, especially in the midgut gland. To our
knowledge, this is the first report on studying chromium and plat-
inum fate and distribution in snails by Nano-SIMS.
Fig. 2. NanoSIMS 50 image of the mantle from Helix aspersa from the platinum group fed with zucchini containing injected platinum chloride. Raster size was 40 � 40 lm (a)
or 15 � 15 lm (b) or 20 � 20 lm (c). (a) Control group, (b) 1 g Lÿ1 exposed group, (c) 10 g Lÿ1 exposed group. Nu: Nucleus. Scale bar equals 5 lm.
6 T. Eybe et al. / Chemosphere xxx (2012) xxx–xxx
Please cite this article in press as: Eybe, T., et al. Determination of oral uptake and biodistribution of platinum and chromium by the garden snail (Helix
aspersa) employing nano-secondary ion mass-spectrometry. Chemosphere (2012), http://dx.doi.org/10.1016/j.chemosphere.2012.09.038

Fig. 4. IMS-6F image of the midgut gland from Helix aspersa from the chromium group fed with zucchini containing injected chromium (1 g Lÿ1). Ca = calcium cells, calcium
granules, Tu = tubule. Raster size was 40 � 40 lm. Scale bar equals 5 lm.
Fig. 3. IMS-6F image of midgut gland (MDD), mantle, and shell from Helix aspersa from the chromium group fed with zucchini containing injected potassium dichromate.
Raster size was 40 � 40 lm. (a) Control group, (b) 1 g Lÿ1 exposed group and (c) 10 g Lÿ1 exposed group. Scale bar equals 5 lm.
T. Eybe et al. / Chemosphere xxx (2012) xxx–xxx 7
Please cite this article in press as: Eybe, T., et al. Determination of oral uptake and biodistribution of platinum and chromium by the garden snail (Helix
aspersa) employing nano-secondary ion mass-spectrometry. Chemosphere (2012), http://dx.doi.org/10.1016/j.chemosphere.2012.09.038
Snails have been reported to accumulate a number of metals,
such as copper, zinc, cadmium and lead, in comparatively high con-
centrations. For example, up to 20 lg gÿ1 lead as a marker of traffic
exposure and over 500 lg gÿ1 zinc (per dry weight) were detected
in H. aspersa in earlier studies, much higher concentrations com-
pared to plant tissues of the same area (Viard et al., 2004). This
has resulted in their use as indicators of environmental metal con-
tamination; such as for investigating copper pesticide fate (Sny-
man et al., 2009) or studying lead and cadmium as a marker for
highway (Viard et al., 2004) and urban pollution (Regoli et al.,
2006); albeit not in relation to platinum.
Following oral metal intake, the midgut gland or digestive
gland or hepatopancreas, similar in its function than the pan-
creas and liver for humans, is a major organ involved in metal
uptake and storage in molluscs. This gland is responsible for
digestive enzyme production, nutrient absorption, endocytosis
of certain food ingredients and also food storage and excretion
(Owen, 2012). Its epithelium is made up of excretory cells,
digestive cells and calcium cells (reviewed by Amaral et al.
(2004)), and the gland further contains connective tissue. Within
this, metals have been reported to be stored in rhogocytes or
pore cells that are unique to molluscs. These cells are normally
involved in calcium homeostasis and metal detoxification, but
also are rich in hemocyanin (the snails’ main oxygen carrying
molecule) and MT isoforms (reviewed by Dallinger et al.
(2005)). In the present study, platinum and chromium rich par-
ticles of up to several lm were detected in the cells and its ves-
icles of the midgut gland. When compared to midgut gland
structures (Fig. 1d), size of particles and distribution suggested
that these were most likely calcium cells, as found by the larger
agglomerations of especially platinum (Fig. 1c), and possibly
vesicles in digestive cells (Fig. 1b) with somewhat smaller
agglomerations of platinum. For chromium, the distribution ap-
peared to be somewhat different as this metal was mostly found
at high concentrations in cellular structures that were not nec-
essarily rich in calcium, potassium and sodium (Fig. 4), i.e. sug-
gesting its association with digestive rather than with calcium
cells.
Both chromium and platinum have been reported to be bio-
available, with minimum precipitation when changing from the
acidic pH of the stomach to the neutral pH of the small intestine,
with a reported bioavailability of ca. 15% for platinum estimated
based on in vitro trials with dust samples (Colombo et al.,
2008b); while for chromium bioavailability appears lower, around
0.4–2.5% in humans (Clydesdale, 2012). In general, hexavalent
chromium is thought to be of higher bioavailability than the triva-
lent form. Nevertheless, chromium biomagnification, including
snails, is often reported to be below that of other heavy metals
such as lead and cadmium (Clydesdale, 1998; Vinodhini and
Narayanan, 2008; Adedeji et al., 2011), possibly, at least partly,
due to poorer uptake from the gut, which appeared not to be the
case in the present study as chromium biomagnifcation (>50) ap-
peared to be high. Following absorption of metals by the gut and
biodistribution to the midgut gland, the rhogocygtes, present
throughout the body, have been reported to represent the major
storage cells, for e.g., copper (Dallinger et al., 2005) while other
cells, including epithelial cells of the midgut gland were reported
to participate in other metal accumulation such as cadmium (Hodl
et al., 2010). Within rhogocytes, both storage as rather rapid
exchangeable granules and in form of metal-MT have been dis-
cussed. Different MT isoforms have been reported in various snail
tissues including the midgut gland and mantle (Dallinger et al.,
2004a,b), involved in different tasks. For instance, in the Roman
snail (H. pomatia), one isoform predominantly expressed in the
midgut gland was associated with the accumulation, binding and
detoxification of cadmium, while another, structurally differing
isoform of the mantle, rather bound exclusively to copper. As these
MT isoforms are inducible following metal exposure, they may rep-
resent potential biomarkers for environmental metal pollution
(Dallinger et al., 2000). In the present study, the presence of both
chromium and platinum especially in form of small vesicles around
2–4 lm correlating well with phosphorous, oxygen, and carbon
may, however, suggest the predominant accumulation in calcium
cells or basophilic cells of the midgut gland (Fig. 1b and c). Never-
theless, for platinum, binding to MT isoforms has been reported, at
least in vitro (Zhang et al., 1995), and platinum possesses a high
affinity toward sulphur ligands (Nieboer and Richardson, 1980),
but the exact type of storage form of platinum and chromium
could not be deduced from the present study and remained
speculative.
Following uptake, metals were distributed all over the body of
the snail, including the mantle which participates in the formation
of the shell. In the present study, granular-like structures were also
detected in the mantle, while distribution was more or less homog-
enous in the shell, the latter mostly consisting of calcium carbon-
ate. The shell has been reported to constitute another sink for
metals, partly due to similarities between metal ions and calcium,
which is part of the shell, consisting of >95% of calcium carbonate
Table 3
Basic statistics for comparison of platinum and chromium exposure in the midgut gland, mantle, and shell, following exposure in food for 2 months. For comparisons, NanoSIMS
50 pictures were analysed based on the 12C14N-normalized pixel intensity of Pt, and number of pixels with intensity >0 were compared. The chromium images obtained by the
IMS-6F were normalized by 40Ca in a similar way.
Normalized intensitiesA
Pt-exposure Cr-exposure
Control Exposure 1 Exposure 2 Control Exposure1 Exposure 2
Midgut gland
Mean 2.20Eÿ07a 6.46Eÿ05b 4.44Eÿ04c 6.49Eÿ04a 1.19Eÿ01b 1.62Eÿ02c
RSDB(%) 66.1 23.1 69.8 40.1 27.6 83.5
Mantle
Mean 3.98Eÿ06a 1.08Eÿ05a 4.33Eÿ05a 3.31Eÿ04a 4.71Eÿ03b 3.27Eÿ03b
RSD (%) 67.5 66.2 110.2 11.6 41.3 14.5
Shell
Mean 1.32Eÿ06a 2.21Eÿ05b 3.85Eÿ05b 4.95Eÿ05a 1.63Eÿ02b 2.00Eÿ03b
RSD (%) 106.4 8.3 73.0 23.5 83.9 73.0
Superscripts for one metal in a row not sharing the same letter are significantly different, P < 0.02 (2-sided), Bonferroni. Three images obtained from 3 different animals per
group were used for normalization.A Data were logarithmized for statistical comparisons.B Relative standard deviation.
8 T. Eybe et al. / Chemosphere xxx (2012) xxx–xxx
Please cite this article in press as: Eybe, T., et al. Determination of oral uptake and biodistribution of platinum and chromium by the garden snail (Helix
aspersa) employing nano-secondary ion mass-spectrometry. Chemosphere (2012), http://dx.doi.org/10.1016/j.chemosphere.2012.09.038

(reviewed by White et al. (2007)). Indeed, the present results
showed that up to 60% of the total metal content was detected in
the snail shell, with a higher percentage for platinum (up to 60%)
than for chromium (up to 22%). It is possible that platinum fol-
lowed a more similar distribution than calcium due to its ionisa-
tion state in the +2 form, but this remains speculative. The fact
that this high concentration found by ICP-MS was not very well re-
flected by the NanoSIMS 50 pictures could be due to technical dif-
ficulties in analysing this non-conductive material via layering it
with a conductive gold-surface, a technique which is common
practice for NanoSIMS 50 non-conducting samples (Wacey et al.,
2010), but which might, to some extent, compromise signal inten-
sities, i.e. the charge compensation could not be optimized over the
entire sample. Imaging with the oxygen source as primary beam
(Oÿ) has been envisaged but has not yet been performed. Further-
more, it cannot be completely excluded that the way of chemical
embedding caused certain losses of metals due to wash-out effects.
In the future, it would be interesting to employ alternative prepa-
ration techniques such as cryo-preparation (Smart et al., 2010).
Nevertheless, the 12C14N normalized results (Table 3) showed the
drastic metal increase in this compartment compared to control
samples.
It is interesting to note that the uptake of platinum and chro-
mium in the 10 times higher exposed groups did not result in dras-
tically increased tissue concentrations (Table 2). Except for
platinum in the shell, the concentrations detected by ICP-MS were
even lower in the tissues of snails receiving higher vs. lower metal
concentrations in their food. We can only speculate for the reasons.
The similar weight distribution (Table 1) of snail groups including
weight gain (except for the Pt 2 group) does not suggest that snails
exposed to higher concentrations of metals significantly limited
their food intake. Avoidance of food parts that were higher con-
taminated can, to a large degree, also be excluded, as only a limited
fraction of the metals was recovered in the non-consumed food (4%
and 18% for the low and high platinum group and 10% and 17% for
the low and high chromium group, respectively). A somewhat
higher avoidance of metals in the higher exposure groups however
is suggested by these figures, and it cannot be excluded that the
surviving snails were to some extent able to avoid the metal con-
taminated food. However, earlier studies have reported an EC50
for oral Cr6+ uptake by H. aspersa of around 300 lg gÿ1 (Coeurdas-
sier et al., 2000), thus our concentrations of up to 25 lg gÿ1 could
only be defined as being of intermediate toxicity, which is also re-
flected by a similar survival rate between controls and metal-fed
snails, with survival rates perhaps generally impacted by non-opti-
mized rearing conditions, rather than metal intake alone. Snails
may have avoided metal intake due to altered taste of the zucchini
by its metal content or due to increased acidity resulting from the
metal solution in diluted HCl. Another explanation would be that
snails were able, above a certain threshold, to excrete metals. It
has been reported that such mechanisms are present in snails that
were exposed to very high metal concentrations, e.g. via excretion
of entire cells by diapedesis (Dallinger andWieser, 1984). As higher
ratios of metal concentration in soft tissues:shells for both plati-
num and chromium were found in the lower exposed animals, it
also could be speculated that the body transported surplus concen-
trations of metals into the shell.
Based on the Nano-SIMS images, no huge differences between
platinum and chromium distribution could be suggested, only a
somewhat more evenly dispersed chromium signal. It has to be
noted though that the detection of chromium with the IMS-6F
instrument resulted, due to its lower lateral resolution, in the
depicturing of less details. However, chromium retention in the
snails appeared to be somewhat higher compared to that of plati-
num (Table 2), perhaps due to the higher solubility and bioavail-
ability of chromium VI compared to platinum chloride, despite
the higher presumed toxicity of chromium VI to act as an oxidising
agent with potential carcinogenic characteristics (Nickens et al.,
2010).12C14N normalization was carried out in the present study to
cancel out between-run intensity differences of the Nano-SIMS
analysis, similar as done previously by 12C normalization (Eybe
et al., 2009). 12C14N was used as the limited spread of the magnetic
field did not allow detecting simultaneously 195Pt and 12C. Further-
more, normalization was carried out for Cr with 40Ca, as this ele-
ment was found to have the strongest signal intensity in the
IMS-6F, representing a comparatively abundant element, assuming
to allow for a reasonably accurate normalization.
It also has to be mentioned that not all metals could equally
well be followed by Nano-SIMS techniques in living tissues, due
to differences in detection sensitivity. Metals that could be esti-
mated to be equally well detected include some transition metals
such as vanadium, titanium, and alkaline and earth alkaline ele-
ments and lanthanides, while other interesting ones, in terms of
toxicity and environmental fate, including mercury, cadmium or
lead, can only be detected with lower sensitivity, both in positive
or negative mode.
In summary, the uptake of platinum and chromium was fol-
lowed in snails employing the Nano-SIMS technique, to allow
detection within various tissues and metal distribution in cells. It
is postulated that snails may, due to their high accumulation of
metals over their lifetime (in case of H. aspersa being ca. 2–5 years
(Thompson and Cheney (1996))), serve as indicators of environ-
mental pollution, and that Nano-SIMS could constitute a promis-
ing, emerging technique to study metal fate.
Acknowledgments
We thank Prof. Dallinger for his kind aid in snail cell structure
interpretation. We thank Boris Untereiner for his help in preparing
the snail samples. This work was realized in the framework of the
project ‘‘Nano-Environment’’, supported by the Fonds National de
la Recherche (FNR, Luxembourg) within the ‘‘INNOVATIVE MATE-
RIALS AND NANOTECHNOLOGY’’ program.
Appendix A. Supplementary material
Supplementary data associated with this article can be found, in
the online version, at http://dx.doi.org/10.1016/j.chemosphere.
2012.09.038.
References
Adedeji, O.B., Adeyemo, O.K., Oyedele, M.O., 2011. Heavy metals in snail and watersamples from Alaro river in Oluyole industrial area of Ibadan southwesternNigeria. J. Appl. Sci. Environ. Sanit. 6, 115–121.
Amaral, A.F., Anselmo, H., da Cunha, R.M., Rodrigues, A.S., 2004. The connectivetissue index of Helix aspersa as a metal biomarker. Biometals 17, 625–629.
Ammann, A.A., 2007. Inductively coupled plasma mass spectrometry (ICP MS): aversatile tool. J. Mass Spectrom. 42, 419–427.
Barbante, C., Veysseyre, A., Ferrari, C., Van de Velde, K., Morel, C., Capodaglio, G.,Cescon, P., Scarponi, G., Boutron, C.F., 2001. Greenland snow evidence of largescale atmospheric contamination for platinum, palladium and rhodium.Environ. Sci. Technol. 35, 835–839.
Barbier, O., Jacquillet, G., Tauc, M., Cougnon, M., Poujeol, P., 2005. Effect of heavymetals on, and handling by, the kidney. Nephron Physiol. 99, 105–110.
Brandenberger, C., Clift, M.J., Vanhecke, D., Muhlfeld, C., Stone, V., Gehr, P., Rothen-Rutishauser, B., 2010. Intracellular imaging of nanoparticles: is it an elementalmistake to believe what you see? Part Fibre Toxicol. 7, 15.
Burastero, S.E., Paolucci, C., Fabbri, M., 2009. Ambient pollutants as adjuvant forallergic sensitization: the emerging role of platinum group elements. J. Biol.Regul. Homeost. Agents 23, 207–215.
Chabicovsky, M., Niederstatter, H., Thaler, R., Hodl, E., Parson, W., Rossmanith, W.,Dallinger, R., 2003. Localization and quantification of Cdÿ and Cuÿ specificmetallothionein isoform mRNA in cells and organs of the terrestrial gastropodHelix pomatia. Toxicol. Appl. Pharmacol. 190, 25–36.
T. Eybe et al. / Chemosphere xxx (2012) xxx–xxx 9
Please cite this article in press as: Eybe, T., et al. Determination of oral uptake and biodistribution of platinum and chromium by the garden snail (Helix
aspersa) employing nano-secondary ion mass-spectrometry. Chemosphere (2012), http://dx.doi.org/10.1016/j.chemosphere.2012.09.038

Clydesdale, F.M., 1998. Mineral interactions in foods. In: Bodwell, C.E., Erdman, J.W.(Eds.), Nutrient Interactions. Marcel Decker, New York, pp. 73–113.
Clydesdale, F.M., 2012. Mineral interactions in foods. In: Bodwell, C.E., Erdman, J.W.(Eds.), Nutrient Interactions. Marcel Decker, New York, pp. 73–113.
Cobb, J.P., Hotchkiss, R.S., Karl, I.E., Buchman, T.G., 1996. Mechanisms of cell injuryand death. Br. J. Anaesth. 77, 3–10.
Coeurdassier, M., Vaufleury, A.G., Badot, P.M., 2000. Dose-dependent growthinhibition and bioaccumulation of hexavalent chromium in land snail Helix
aspersa. Environ. Toxicol. Chem. 19, 2571–2578.Colombo, C., Monhemius, A.J., Plant, J.A., 2008a. Platinum, palladium and rhodium
release from vehicle exhaust catalysts and road dust exposed to simulated lungfluids. Ecotoxicol. Environ. Saf. 71, 722–730.
Colombo, C., Monhemius, A.J., Plant, J.A., 2008b. The estimation of thebioavailabilities of platinum, palladium and rhodium in vehicle exhaustcatalysts and road dusts using a physiologically based extraction test. Sci.Total Environ. 389, 46–51.
Dallinger, R., 1994. Invertebrate organisms as biological indicators of heavy metalpollution. Appl. Biochem. Biotechnol. 48, 27–31.
Dallinger, R., Wieser, W., 1984. Patterns of accumulation, distribution and liberationof Zn, Cu, Cd and Pb in different organs of the land snail Helix pomatia L. Comp.Biochem. Physiol. C. 79 (1), 117–124.
Dallinger, R., Berger, B., Gruber, C., Hunziker, P., Stürzenbaum, S., 2000.Metallothioneins in terrestrial invertebrates: structural aspects, biologicalsignificance and implications for their use as biomarkers. Cell. Mol. Biol. 46(2), 331–346.
Dallinger, R., Chabicovsky, M., Berger, B., 2004a. Isoform-specific quantification ofmetallothionein in the terrestrial gastropod Helix pomatia. I. Molecular,biochemical, and methodical background. Environ. Toxicol. Chem. 23 (4),890–901.
Dallinger, R., Chabicovsky, M., Lagg, B., Schipflinger, R., Weirich, H.G., Berger, B.,2004b. Isoform-specific quantification of metallothionein in the terrestrialgastropod Helix pomatia. II. A differential biomarker approach under laboratoryand field conditions. Environ. Toxicol. Chem. 23 (4), 902–910.
Dallinger, R., Chabicovsky, M., Hodl, E., Prem, C., Hunziker, P., Manzl, C., 2005.Copper in Helix pomatia (Gastropoda) is regulated by one single cell type:differently responsive metal pools in rhogocytes. Am. J. Physiol. Regul. Integr.Comp. Physiol. 289, R1185–R1195.
Eybe, T., Audinot, J.N., Guignard, C., Migeon, H.N., Hoffmann, L., 2008. NanoSIMS 50elucidation of the natural element composition in structures of cyanobacteriaand their exposure to halogen compounds. J. Appl. Microbiol. 105 (5), 1502–1511.
Eybe, T., Bohn, T., Audinot, J.N., Udelhoven, T., Cauchie, H.M., Migeon, H.N.,Hoffmann, L., 2009. Uptake visualization of deltamethrin by NanoSIMSand acute toxicity to the water flea Daphnia magna. Chemosphere 76,134–140.
Gomez, B., Gomez, M., Sanchez, J.L., Fernandez, R., Palacios, M.A., 2001. Platinumand rhodium distribution in airborne particulate matter and road dust. Sci.Total Environ. 269, 131–144.
Hodl, E., Felder, E., Chabicovsky, M., Dallinger, R., 2010. Cadmium stress stimulatestissue turnover in Helix pomatia: increasing cell proliferation from metaltolerance to exhaustion in molluscan midgut gland. Cell Tissue Res. 341, 159–171.
Hoffman, J., 1989. Precious metal technologies. In: Thermae, A.E., Gondolier, I.H.(Eds.), Process Metallurgy, vol. 5. Elsevier, Amsterdam.
Holmes, A.L., Wise, S.S., Wise Sr., J.P., 2008. Carcinogenicity of hexavalentchromium. Indian J. Med. Res. 128, 353–372.
Hoppstock, K., Sures, B., 2004. Platinum-group metals. In: Meiran, E., Anke, M.,Ihnat, M., Stoeppler, M. (Eds.), Elements and their Compounds in theEnvironment. Wiley-VCH, Weinheim, pp. 1047–1086.
Jordaens, K., De Wolf, H., Vandecasteele, B., Blust, R., Backeljau, T., 2006.Associations between shell strength, shell morphology and heavy metals inthe land snail Cepaea nemoralis (Gastropoda, Helicidae). Sci. Total Environ. 363(1–3), 285–293.
Lechene, C., Hillion, F., McMahon, G., Benson, D., Kleinfeld, A.M., Kampf, J.P., Distel,D., Luyten, Y., Bonventre, J., Hentschel, D., Park, K.M., Ito, S., Schwartz, M.,Benichou, G., Slodzian, G., 2006. High-resolution quantitative imaging ofmammalian and bacterial cells using stable isotope mass spectrometry. J. Biol.5, 20.
Losi, M.E., Amrhein, C., Frankenberger, W.T., 1994. Environmental biochemistry ofchromium. Rev. Environ. Contam. Toxicol. 36, 91–121.
MacRae, N.D., Bottazzi, P., Ottolini, L., Vannucci, R., 1993. Quantitative REE analysisof silicates by SIMS: conventional energy filtering vs. specimen isolation mode.Chem. Geol. 103, 45–54.
Markich, S.J., Jeffree, R.A., Burke, P.T., 2002. Freshwater bivalve shells as archivalindicators of metal pollution from a copper–uranium mine in tropical northernAustralia. Environ. Sci. Technol. 36, 821–832.
Menta, C., Parisi, V., 2001. Metal concentrations in Helix pomatia, Helix aspersa andArion rufus: a comparative study. Environ. Pollut. 115 (2), 205–208.
Migeon, A., Audinot, J.N., Eybe, T., Richaud, P., Damien, B., Migeon, H.N., Chalot, M.,2011. Cadmium and zinc localization by SIMS in leaves of Populus deltoides (cv.Lena) grown in a metal polluted soil. Surf. Interface Anal. 43, 367–369.
Migliore, L., Frenzilli, G., Nesti, C., Fortaner, S., Sabbioni, E., 2002. Cytogenetic andoxidative damage induced in human lymphocytes by platinum, rhodium andpalladium compounds. Mutagenesis 17, 411–417.
Mohanty, M., Pattnaik, M.M., Mishra, A.K., Patra, H.K., 2011. Chromiumbioaccumulation in rice grown in contaminated soil and irrigated minewastewater – a case study at south Kaliapani chromite mine area, Orissa,India. Int. J. Phytoremediat. 13, 397–409.
Nickens, K.P., Patierno, S.R., Ceryak, S., 2010. Chromium genotoxicity: a double-edged sword. Chem. Biol. Interact. 188, 276–288.
Nieboer, E., Richardson, D.H.S., 1980. The replacement of the nondescript term‘‘heavy metals’’ by a biologically and chemically significant classification ofmetal ions. Environ. Poll. Series B Chem. Phys. 1, 3–26.
Notten, M.J., Oosthoek, A.J., Rozema, J., Aerts, R., 2005. Heavy metal concentrationsin a soil-plant-snail food chain along a terrestrial soil pollution gradient.Environ. Pollut. 138 (1), 178–190.
Owen, G., 2012. Digestion. In: Wilbur, K.M., Yonge, C.M. (Eds.), Physiology ofMollusca. Academic Press, New York, pp. 53–96.
Radike, M., Warshawsky, D., Caruso, J., Goth-Goldstein, R., Reilman, R., Collins, T.,Yaeger, M., Wang, J., Vela, N., Olsen, L., Schneider, J., 2002. Distribution andaccumulation of a mixture of arsenic, cadmium, chromium, nickel, andvanadium in mouse small intestine, kidneys, pancreas, and femur followingoral administration in water or feed. J. Toxicol. Environ. Health A 65 (23), 2029–2052.
Regoli, F., Gorbi, S., Fattorini, D., Tedesco, S., Notti, A., Machella, N., Bocchetti, R.,Benedetti, M., Piva, F., 2006. Use of the land snail Helix aspersa as sentinelorganism for monitoring ecotoxicologic effects of urban pollution: an integratedapproach. Environ. Health Perspect. 114, 63–69.
Shelnutt, S.R., Goad, P., Belsito, D.V., 2007. Dermatological toxicity of hexavalentchromium. Crit. Rev. Toxicol. 37, 375–387.
Smart, K.E., Smith, J.A., Kilburn, M.R., Martin, B.G., Hawes, C., Grovenor, C.R., 2010.High-resolution elemental localization in vacuolate plant cells by nanoscalesecondary ion mass spectrometry. Plant J. 63, 870–879.
Snyman, R.G., Reinecke, A.J., Reinecke, S.A., 2009. Quantitative changes in digestivegland cells and oocytes of Helix aspersa, as biomarkers of copper oxychlorideexposure under field conditions. Bull. Environ. Contam. Toxicol. 83, 19–22.
Thompson, R., Cheney, S., 1996. Raising Snails. The Altnerative Farming SystemsInformation Center, National Agricultural Library. <http://www.nal.usda.gov/afsic/AFSIC_pubs/srb96-05.htm>.
Viard, B., Pihan, F., Promeyrat, S., Pihan, J.C., 2004. Integrated assessment of heavymetal (Pb, Zn, Cd) highway pollution: bioaccumulation in soil, Graminaceae andland snails. Chemosphere 55, 1349–1359.
Vinodhini, R., Narayanan, M., 2008. Bioaccumulation of heavy metals in organs offresh water fish Cyprinus carpio (Common carp). Int. J. Environ. Sci. Tech. 5,1979–1982.
Wacey, D., Gleeson, D., Kilburn, M.R., 2010. Microbialite taphonomy andbiogenicity: new insights from NanoSIMS. Geobiology 8, 403–416.
White, M.M., Chejlava, M., Fried, B., Sherma, J., 2007. The concentration of calciumcarbonate in shells of freshwater snails. Am. Malacol. Bull. 22, 139–142.
Wiseman, C.L., Zereini, F., 2009. Airborne particulate matter, platinum groupelements and human health: a review of recent evidence. Sci. Total Environ.407, 2493–2500.
Zhang, B., Tang, W., Gao, S., Zhou, Y., 1995. Platinum binding to metallothionein.Analysis of circular dichroism spectra of complexes formed betweenmetallothionein and platinum from cis- and trans-diamminedichloroplatinum. J. Inorg. Biochem. 58 (1), 9–19.
10 T. Eybe et al. / Chemosphere xxx (2012) xxx–xxx
Please cite this article in press as: Eybe, T., et al. Determination of oral uptake and biodistribution of platinum and chromium by the garden snail (Helix
aspersa) employing nano-secondary ion mass-spectrometry. Chemosphere (2012), http://dx.doi.org/10.1016/j.chemosphere.2012.09.038