bordetella bronchisepticadermonecrotizing toxin, which activates a small gtp-binding protein rho,...
TRANSCRIPT

EXPERIMENTAL CELL RESEARCH 230, 163–168 (1997)ARTICLE NO. EX963414
SHORT NOTE
Bordetella bronchiseptica Dermonecrotizing Toxin, Which Activates aSmall GTP-Binding Protein Rho, Induces Membrane Organelle
Proliferation and Caveolae Formation
TAKAO SENDA,*,1 YASUHIKO HORIGUCHI,† MASANORI UMEMOTO,‡NAKABA SUGIMOTO,† AND MORIHIRO MATSUDA†
*Department of Anatomy and ‡Department of Otorhinolaryngology, Osaka University Medical School, 2-2 Yamadaoka,Suita, Osaka 565, Japan; and †Department of Bacterial Toxinology, Research Institute for Microbial Diseases,
Osaka University, 3-1 Yamadaoka, Suita, Osaka 565, Japan
been identified [3]. Although some of them were shownWe recently demonstrated that Bordetella bronchi- to be functionally regulated by rho protein, it is not
septica dermonecrotizing toxin (DNT) modifies a small known how these proteins elicit a variety of cell re-GTP-binding protein rho and causes assembly of actin sponses. A great variety of rho-associated proteinsstress fibers and focal adhesions in MC3T3-E1 cells, raises the possibility that rho protein may conduct cel-indicating that DNT activates the function of rho pro- lular functions other than those so far reported.tein (Horiguchi et al., 1995, J. Cell Sci. 108, 3243–3251). Dermonecrotizing toxin (DNT) produced by Borde-In this study, we examined by electron microscopy ul- tella species causes ischemic dermonecrosis when ittrastructural changes in DNT-treated MC3T3-E1 cells is injected intradermally into guinea pigs, mice, rab-under conditions in which DNT elicited the above mor- bits, and other animals [7, 8]. Using purified DNT [9,phological changes. We found that DNT induced prolif- 10] and an osteoblastic cell line, MC3T3-E1 [11], weeration of cytoplasmic membrane organelles, such as have shown that DNT changes cell morphology [12],Golgi apparatus, smooth and rough endoplasmic retic- inhibits expression of cell differentiation markers,ulum, and mitochondria, and formation of plasmalem- such as alkaline phosphatase and type I collagen inmal caveolae. Therefore we examined also the effect
osteoblasts [12], stimulates protein and DNA synthe-of Clostridium botulinum C3 exoenzyme (C3), whichses [13], and blocks cytokinesis [13]. Recently, wehas been known to ADP-ribosylate and inactivate rhofound that the direct target of DNT is a small GTP-protein, and found that C3 exoenzyme inhibited thebinding protein rho and that DNT activates the rho-development of membrane organelles in the MC3T3-dependent signaling pathway, which regulates as-E1 cells. These findings show that proliferation ofsembly of actin stress fibers and focal adhesions, re-membrane organelles and caveolae formation are con-sulting in changes in cell morphology [14].trolled by the function of rho, which is the target of
In this study we investigated by electron microscopyDNT action. q 1997 Academic Press
changes in the ultrastructure of DNT-treated MC3T3-E1 cells. Here we report that Bordetella bronchisepticaDNT, which activates a small GTP-binding protein rho,INTRODUCTIONinduces membrane organelle proliferation and plas-malemmal caveolae formation.Rho protein is considered to trigger and regulate a
variety of cell responses including phospholipid turn-over [1–3], protein phosphorylation [3], rearrange- MATERIALS AND METHODSments of the actin cytoskeleton in mammalian cells [3,4], and growth and remodeling of the cell wall in yeasts DNT and C3 exoenzyme. DNT was purified from the cell extracts
of B. bronchiseptica strain S798 by the method of Horiguchi et al.[5, 6]. Recently, a number of proteins that associate[10]. Clostridium botulinum C3 exoenzyme (C3) was provided by S.with the rho protein in a GTP-dependent manner haveKozaki, University of Osaka Prefecture (Osaka, Japan).
Cell culture. MC3T3-E1 cells [11] were cultivated at 377C under5% CO2 in air as described previously [14].1 To whom correspondence and reprint requests should be ad-
dressed at present address: Department of Anatomy I, Nagoya Uni- Phase-contrast microscopic examination. Cells seeded at an ini-tial density of 2.5 1 104 cells/cm2 into wells of 24-well plates wereversity School of Medicine, 65 Tsurumai-cho, Showa-ku, Nagoya 466,
Japan. cultivated for 1 day. Then the cells were preincubated in serum-free
163 0014-4827/97 $25.00Copyright q 1997 by Academic Press
All rights of reproduction in any form reserved.
AID ECR 3414 / 6i16$$$$21 12-12-96 21:41:42 ecal
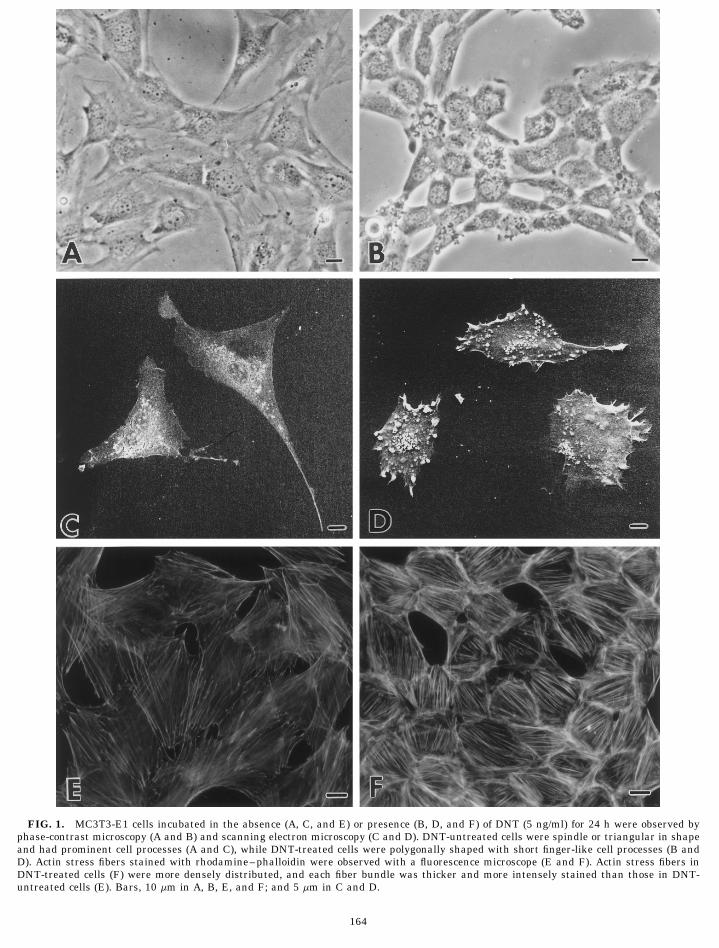
FIG. 1. MC3T3-E1 cells incubated in the absence (A, C, and E) or presence (B, D, and F) of DNT (5 ng/ml) for 24 h were observed byphase-contrast microscopy (A and B) and scanning electron microscopy (C and D). DNT-untreated cells were spindle or triangular in shapeand had prominent cell processes (A and C), while DNT-treated cells were polygonally shaped with short finger-like cell processes (B andD). Actin stress fibers stained with rhodamine–phalloidin were observed with a fluorescence microscope (E and F). Actin stress fibers inDNT-treated cells (F) were more densely distributed, and each fiber bundle was thicker and more intensely stained than those in DNT-untreated cells (E). Bars, 10 mm in A, B, E, and F; and 5 mm in C and D.
164
12-12-96 21:41:42 ecal

165SHORT NOTE
medium overnight before adding DNT at a final concentration of 5 1C), DNT-treated cells were polygonally shaped withng/ml. Then the cells were incubated at 377C for 24 h. The cells with short finger-like cell processes (Figs. 1B and 1D). Theseor without DNT treatment were examined under a phase-contrast morphological changes were considered to be due tomicroscope.
rearrangement of the cytoskeletal system. Figures 1ERhodamine–phalloidin staining. Actin stress fibers in DNT-and 1F show that actin stress fibers in DNT-treatedtreated and -untreated cells were stained with rhodamine–phalloi-
din as described previously [14]. cells were more densely distributed and that each fiberElectron microscopic examinations. Cells seeded at an initial den- bundle was thicker and more intensely stained than in
sity of 1.0 1 104 cells/cm2 in 6-cm plastic dishes were cultivated for DNT-untreated cells.1 day. After 24 h incubation in serum-free medium with or without We then examined changes in the ultrastructure inDNT, the cells were fixed with 2.5% glutaraldehyde in PBS for 30
the cells treated with DNT under the above conditionsmin and subsequently in 1% osmium tetroxide in PBS for 10 min.Then the cells were washed with 10% sucrose in distilled water, using a transmission electron microscope. Figure 2incubated with 3% uranyl acetate in distilled water, and dehydrated shows that the cytoplasm of untreated cells was filledin graded concentrations of ethanol. (1) For scanning electron micros- with abundant free ribosomes and a small amount ofcopy, they were immersed in t-butyl alcohol, freeze-dried, coated with poorly developed membrane organelles, such as Golgiplatinum, and subjected to examination. (2) For transmission elec-
apparatus, rough endoplasmic reticulum, and mito-tron microscopy, they were embedded in epoxy resin. Ultrathin sec-tions cut with an ultramicrotome were stained with uranyl acetate chondria (Figs. 2A–2D). Golgi apparatuses, whichand lead citrate and subjected to examination. For transmission elec- were composed of two or three short Golgi lamellae andtron microscopic analyses on the effects of C3, cells were preincu- a few surrounding vesicles and vacuoles, were alwaysbated in 10% fetal calf serum (FCS)-containing medium at 377C for localized in the perinuclear region (Fig. 2A). A few24 h and consequently incubated in the FCS-containing medium with
rough endoplasmic reticulum were seen in the perinu-or without 10 mg/ml C3 at 377C for 40 h. Then the cells were fixedand processed for transmission electron microscopic examination. clear cytoplasm (Figs. 2A and 2B). Mitochondria with
tubular and/or lamellar cristae were scattered both inthe perinuclear cytoplasm and in the cell process (Figs.RESULTS AND DISCUSSION2A–2D). Actin stress fibers were running along both
Figure 1 shows morphological changes in DNT- upper (apical) and basal plasma membranes and weretreated MC3T3-E1 cells detected by phase-contrast mi- hardly seen in the inner cytoplasm (Figs. 2A–2D). Thecroscopy (Figs. 1A and 1B) and scanning electron mi- basal plasma membrane was frequently in contact withcroscopy (Figs. 1C and 1D). In contrast to the cells incu- the dish surface along the cell processes (Figs. 2C andbated without DNT, which have spindle or triangular 2D). Most of these cell-to-dish attachment sites, how-
ever, were not associated with the stress fibers. In con-cell shapes and prominent cell processes (Figs. 1A and
FIG. 2. Transmission electron micrographs of DNT-untreated MC3T3-E1 cells. Poorly developed membrane organelles, such as Golgiapparatus (G), rough endoplasmic reticulum, and mitochondria were seen (A–D). Cell-to-dish attachment sites (arrows) were not associatedwith the stress fibers (C and D). Bars, 500 nm.
AID ECR 3414 / 6i16$$$$22 12-12-96 21:41:42 ecal

FIG. 3. Transmission electron micrographs of DNT-treated MC3T3-E1 cells. Membrane organelles, such as Golgi apparatus (A), smooth(B) and rough (C) endoplasmic reticulum, and mitochondria (C) were markedly developed. Thick stress fiber bundles were running not onlyalong the plasma membrane but also in the inner cytoplasm (D). Many omega-shaped caveolae (arrowheads) were formed along the basalplasma membrane (E and F). Cell-to-dish focal adhesions were undercoated with stress fibers (G and H). Bars, 200 nm in A, B, C, D, G,and H; and 100 nm in E and F.
166
12-12-96 21:41:42 ecal
167SHORT NOTE
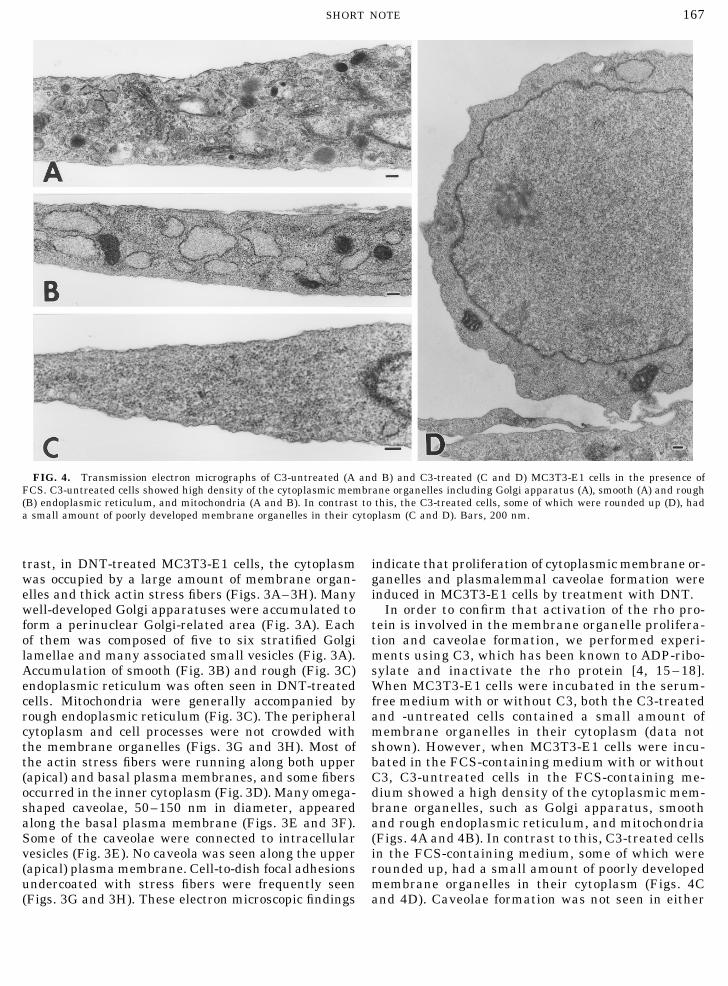
FIG. 4. Transmission electron micrographs of C3-untreated (A and B) and C3-treated (C and D) MC3T3-E1 cells in the presence ofFCS. C3-untreated cells showed high density of the cytoplasmic membrane organelles including Golgi apparatus (A), smooth (A) and rough(B) endoplasmic reticulum, and mitochondria (A and B). In contrast to this, the C3-treated cells, some of which were rounded up (D), hada small amount of poorly developed membrane organelles in their cytoplasm (C and D). Bars, 200 nm.
trast, in DNT-treated MC3T3-E1 cells, the cytoplasm indicate that proliferation of cytoplasmic membrane or-ganelles and plasmalemmal caveolae formation werewas occupied by a large amount of membrane organ-
elles and thick actin stress fibers (Figs. 3A–3H). Many induced in MC3T3-E1 cells by treatment with DNT.In order to confirm that activation of the rho pro-well-developed Golgi apparatuses were accumulated to
form a perinuclear Golgi-related area (Fig. 3A). Each tein is involved in the membrane organelle prolifera-tion and caveolae formation, we performed experi-of them was composed of five to six stratified Golgi
lamellae and many associated small vesicles (Fig. 3A). ments using C3, which has been known to ADP-ribo-sylate and inactivate the rho protein [4, 15–18].Accumulation of smooth (Fig. 3B) and rough (Fig. 3C)
endoplasmic reticulum was often seen in DNT-treated When MC3T3-E1 cells were incubated in the serum-free medium with or without C3, both the C3-treatedcells. Mitochondria were generally accompanied by
rough endoplasmic reticulum (Fig. 3C). The peripheral and -untreated cells contained a small amount ofmembrane organelles in their cytoplasm (data notcytoplasm and cell processes were not crowded with
the membrane organelles (Figs. 3G and 3H). Most of shown). However, when MC3T3-E1 cells were incu-bated in the FCS-containing medium with or withoutthe actin stress fibers were running along both upper
(apical) and basal plasma membranes, and some fibers C3, C3-untreated cells in the FCS-containing me-dium showed a high density of the cytoplasmic mem-occurred in the inner cytoplasm (Fig. 3D). Many omega-
shaped caveolae, 50–150 nm in diameter, appeared brane organelles, such as Golgi apparatus, smoothand rough endoplasmic reticulum, and mitochondriaalong the basal plasma membrane (Figs. 3E and 3F).
Some of the caveolae were connected to intracellular (Figs. 4A and 4B). In contrast to this, C3-treated cellsin the FCS-containing medium, some of which werevesicles (Fig. 3E). No caveola was seen along the upper
(apical) plasma membrane. Cell-to-dish focal adhesions rounded up, had a small amount of poorly developedmembrane organelles in their cytoplasm (Figs. 4Cundercoated with stress fibers were frequently seen
(Figs. 3G and 3H). These electron microscopic findings and 4D). Caveolae formation was not seen in either
AID ECR 3414 / 6i16$$$$22 12-12-96 21:41:42 ecal

168 SHORT NOTE
C3-treated or C3-untreated cells. We have previously REFERENCESshown that DNT action is mediated through the rhoprotein, and the present data lead to assignment of 1. Zhang, J., King, W. G., Killon, S., Hall, A., Feig, L., and Ritten-
house, S. E. (1993) J. Biol. Chem. 268, 22251–22254.new functions to rho apart from the previous docu-2. Chong, L. D., Traynor-Kaplan, A., Bokoch, G. M., and Schwartz,mentation on rho’s role in actin stress fiber assembly.
M. A. (1994) Cell 79, 507–513.Activation of formation of membrane organelle and3. Machesky, L. M., and Hall, A. (1996) Trends Cell Biol. 6, 304–caveolae requires synthesis of membrane proteins. A
310.remarkable increase in number and development of4. Ridley, A. J., and Hall, A. (1992) Cell 70, 389–399.rough endoplasmic reticulum and Golgi apparatus in5. Drgonova, J., Drgon, T., Tanaka, K., Kollar, R., Chen, G., Ford,DNT-treated cells indicates acceleration of protein syn-
R. A., Chan, C. S. M., Takai, Y., and Cabib, E. (1996) Sciencethesis, in good agreement with the results of our previ- 272, 277–279.ous biochemical studies [13].
6. Qadota, H., Python, C. P., Inoue, S. B., Arisawa, M., Anraku,Various cellular functions, including the organiza- Y., Zheng, Y., Watanabe, T., Levin, D. E., and Ohya, Y. (1996)tion of the cytoskeletal system, are suggested to be reg- Science 272, 279–281.ulated by signal transduction pathways in which a 7. Bruckner, I. E., and Evans, D. G. (1939) J. Pathol. Bacteriol. 48,small GTP-binding protein rho plays important roles. 67–78.Our previous studies showed that DNT caused assem- 8. Evans, D. G. (1940) J. Pathol. Bacteriol. 51, 49–58.bly of actin stress fibers and focal adhesions, resulting 9. Horiguchi, Y., Nakai, T., and Kume, K. (1989) Microb. Pathog.in changes in the cell shape of MC3T3-E1 cells, and 6, 361–368.that rho protein is the direct target of DNT [14]. In the 10. Horiguchi, Y., Nakai, T., and Kume, K. (1990) FEMS Microbiol.
Lett. 66, 39–43.present study we demonstrated for the first time that11. Sudo, H., Kodama, H., Amagai, Y., Yamamoto, S., and Kasai, S.DNT induces the proliferation of membrane organelles
(1983) J. Cell Biol. 96, 191–198.and caveolae formation in addition to assembly of actin12. Horiguchi, Y., Nakai, T., and Kume, K. (1991) Infect. Immun.stress fibers and transformation of cell shape. C3 exo-
59, 1112–1116.enzyme, which ADP-ribosylates and inactivates the rho13. Horiguchi, Y., Sugimoto, N., and Matsuda, M. (1993) Infect. Im-protein, inhibited the development of membrane organ-
mun. 61, 3611–3615.elles. This indicates that rho controlled positively and14. Horiguchi, Y., Senda, T., Sugimoto, N., Katahira, J., and Ma-negatively the membrane organelle proliferation and
tsuda, M. (1995) J. Cell Sci. 108, 3243–3251.caveolae formation. Little has been known on the mech-15. Morii, N., Sekine, A., Ohashi, Y., Nakao, K., Imura, H., Fujiwara,anism of membrane organelle formation and its regula-
M., and Narumiya, S. (1988) J. Biol. Chem. 263, 12420–12426.tion. Here we present a new approach to elucidate the16. Narumiya, S., Sekine, A., and Fujiwara, M. (1988) J. Biol. Chem.role of a small GTP-binding protein rho in the regula- 263, 17255–17257.
tion of formation of membrane organelles.17. Rubin, E. J., Gill, D. M., Boquet, P., and Popoff, M. R. (1988)One of the new findings in the present study, that Mol. Cell. Biol. 8, 418–426.
DNT induces the formation of plasmalemmmal caveo- 18. Chardin, P., Boquet, P., Madaule, P., Popoff, M. R., Rubin, E. J.,lae, is very interesting, since recently evidence has and Gill, D. M. (1989) EMBO J. 8, 1087–1092.been accumulated to indicate the important function 19. Fujimoto, T., Nakade, S., Miyawaki, A., Mikoshiba, K., andof caveolae in signal transduction. The inositol triphos- Ogawa, K. (1992) J. Cell Biol. 119, 1507–1513.phate receptor-like protein and the Ca2/ pump that 20. Fujimoto, T. (1993) J. Cell Biol. 120, 1147–1157.regulate transcellular Ca2/ mobilization [19, 20] and 21. Rothberg, K. G., Heuser, J. E., Donzell, W. C., Ying, Y.-S., Glen-several signal transduction-related proteins [21–23] ney, J. R., and Anderson, R. G. W. (1992) Cell 68, 673–682.have been shown to be associated with caveolae. DNT 22. Schnitzer, J. E., Liu, J., and Oh, P. (1995) J. Biol. Chem. 270,will be an useful tool to elucidate the exact relationship 14399–14404.between activation of rho protein and proliferation of 23. Schnitzer, J. E., McIntosh, D. P., Dvorak, A. M., Liu, J., and Oh,
P. (1995) Science 269, 1435–1439.membrane organelles and formation of caveolae.
Received May 17, 1996Revised version received October 18, 1996
AID ECR 3414 / 6i16$$$$22 12-12-96 21:41:42 ecal