br. j. cancer (1972) 26, - celldeath-apoptosis.org · br. j. cancer (1972) 26, 239 apoptosis:...
TRANSCRIPT

Br. J. Cancer (1972) 26, 239
APOPTOSIS: A BASIC BIOLOGICAL PHENOMENON WITH WIDE-RANGING IMPLICATIONS IN TISSUE KINETICS
J. F. R. KERR*, A. H. WYLLIE AND A. R. CURRIEt
From the Department of Pathology, University of Aberdeen
Received for publication April 1972
Summary.-The term apoptosis is proposed for a hitherto little recognized mechan-ism of controlled cell deletion, which appears to play a complementary but oppositerole to mitosis in the regulation of animal cell populations. Its morphologicalfeatures suggest that it is an active, inherently programmed phenomenon, and ithas been shown that it can be initiated or inhibited by a variety of environmentalstimuli, both physiological and pathological.
The structural changes take place in two discrete stages. The first comprisesnuclear and cytoplasmic condensation and breaking up of the cell into a number ofmembrane-bound, ultrastructurally well-preserved fragments. In the secondstage these apoptotic bodies are shed from epithelial-lined surfaces or are takenup by other cells, where they undergo a series of changes resembling in vitro autolysiswithin phagosomes, and are rapidly degraded by lysosomal enzymes derived fromthe ingesting cells.
Apoptosis seems to be involved in cell turnover in many healthy adult tissuesand is responsible for focal elimination of cells during normal embryonic develop-ment. It occurs spontaneously in untreated malignant neoplasms, and participatesin at least some types of therapeutically induced tumour regression. It is implicatedin both physiological involution and atrophy of various tissues and organs. It canalso be triggered by noxious agents, both in the embryo and adult animal.
IN recent years it has become widelyrecognized that spontaneous loss of cellsis an important parameter in neoplasticgrowth (Iversen, 1967; Refsum and Berdal,1967; Steel, 1967; Frindel, Malaise andTubiana, 1968; Laird, 1969; Clifton andYatvin, 1970; Weinstein and Frost, 1970;Lala, 1971, 1972). However, although itis agreed that cell death probably accountsfor most of this loss, little appears to beknown about the mechanisms involved(Lala, 1972).
It has long been tacitly assumed thatcells must be lost continuously frommany normal tissues to balance the celldivision that is readily demonstrable, andthere seems little doubt that loss of cells
often accompanies atrophy and physio-logical involution of tissues and organs.The term necrobiosis is sometimes usedfor this " physiological cell death ", butits morphological features have not beenclearly defined.
The morphological type of cellulardeath described in almost all standard textsis coagulative necrosis, and there iscertainly nothing to suggest that it isinvolved in the control of cell populations.It appears to be invariably caused bynoxious stimuli, and is probably the resultof an irreversible disturbance of cellularhomoeostatic mechanisms (Judah, Ahmedand McLean, 1965; Trump and Ginn,1969), electron microscopy revealing signs
* On study leave from the University of Queensland. Present address: Department of Pathology,University of Queensland Medical School, Herston, Brisbane, Australia, 4006.
t Requests for reprints should be sent to Professor A. R. Currie at his present address: Department ofPathology, University of Edinburgh Medical School, Teviot Place, Edinburgh EH8 9AG, Scotland.
18

J. F. R. KERR, A. H. WYLLIE AND A. R. CURRIE
(1
(2)
(3a
(4)(3b)
FIG. 1.-Normal human secretory-phase endometrium. Apoptotic bodies in the epithelial lining of agland are indicated by arrows: others have been shed into the gland lumen. H. and E. x 400.
FIG. 2.-Prostate of a rat killed 4 days after orchidectomy. There are many apoptotic bodiesamongst the rather shrunken glandular epithelial cells. H. and E. x 750.
FIG. 3.-(a) Scattered apoptotic bodies (arrows) in the adrenal cortex of a normal 4-day old rat.H. and E. x 750. (b) By contrast, coagulative necrosis is almost always confluent, and the " ghosts "of dead cells retain their size and shape. This example illustrates coagulative necrosis induced inthe inner adrenal cortex of an adult rat by 7,12-dimethylbenz(a)anthracene (DMBA). Note thatthe light-microscope appearance3 of nuclear pyknosis occur in both types of cell death despitetheir differences. H. and E. x 200.
FIG. 4.-Apoptotic body (arrow) in a human rectal adenocarcinoma. H. and E. x 1200.
240
p.r. -,. .Bw:- ." i.IW 'VW. W .71-mlopmT" -:;

A BASIC BIOLOGICAL PHENOMENON WITH WIDE-RANGING IMPLICATIONS 241
of degeneration like those found in invitro autolysis (Trump, Goldblatt andStowell, 1965) at an early stage of itsdevelopment (Trump and Ginn, 1969).The recent discovery of a distinctlydifferent mode of cellular death withultrastructural features that are consistentwith an active, inherently controlledphenomenon (Klion and Schaffner, 1966;Farbman, 1968; Kerr, 1969, 1971) istherefore of interest, and we have nowshown that it plays an important role inthe regulation of cell numbers in a varietyof tissues under both physiological andpathological conditions. It can alwaysbe detected in untreated malignant neo-plasms (Kerr and Searle, 1972a), and itparticipates in the regression that followsat least some forms of therapy (Currieet al., 1972; Kerr and Searle, 1972b). Itis also found in many of the tissues ofhealthy animals (Kerr, 1965, 1971, 1972a;Wyllie, Kerr and Currie, 1972a), and itsfocal appearance at specific times duringnormal ontogenesis indicates that it isimplicated in the fashioning of developingorgans and digits, and in the involutionof phylogenetic vestiges in the embryo(Glucksmann, 1951; Saunders, 1966;Farbman, 1968; Webster and Gross, 1970).It is prominent in the adrenal glandfollowing withdrawal of adrenocortico-trophic hormone (ACTH) (Wyllie et al.,1972b), and it is involved in other types ofatrophy (Kerr, 1971).
Thus, although the development ofthis distinctive type of necrosis, which haspreviously been called shrinkage necrosison morphological grounds (Kerr, 1965,1971), can, in fact, be triggered by noxiousagents (Kerr, 1971), it often appearsspontaneously or in response to knownphysiological stimuli, and it is clear thatits implications in tissue kinetics are ofwidely ranging importance. It is notconfined to vertebrates (Goldsmith, 1966),
and we suspect that further work willconfirm it as a general mechanism ofcontrolled cell deletion, which is comple-mentary to mitosis in the regulation ofanimal cell populations. Because of itsimportant kinetic significance we suggestthat it be called " Apoptosis ".*
In this paper we review the morpho-logical changes that take place duringthe evolution of apoptosis, we considersome of its biological and pathologicalimplications with particular reference totumour growth and, on the basis of these,propose our concept of apoptosis as avital biological phenomenon.
THE MORPHOLOGY OF APOPTOSISApoptosis characteristically affects
scattered single cells, and is manifestedhistologically by the formation of small,roughly spherical or ovoid cytoplasmicfragments, some of which contain pyknoticremnants of nuclei (Fig. 1-4). In theliver, these have sometimes been referredto as Councilman or Councilman-likebodies, but since there is uncertaintyabout the nature of the structures des-cribed by Councilman in yellow fever(Klion and Schaffner, 1966), we shall callthem apoptotic bodies.
Electron microscopy shows that thestructural changes in apoptosis takeplace in two discrete stages (Fig. 5): thefirst comprises the formation of apoptoticbodies, the second their phagocytosis anddegradation by other cells.
We have so far studied the evolutionof the process with the electron micro-scope in the normal neonatal rat adrenal(Wyllie et al., 1972a), in embryonic mesen-chyme (Crawford, Kerr and Currie, 1972),in both human (Kerr and Searle, 1972aand b) and animal (Currie et al., 1972)neoplasms, in the adrenal cortex followingACTH withdrawal (Wyllie et al., 1972b),and in various types of liver and adrenal
* We are most grateful to Professor James Cormack of the Department of Greek, University ofAberdeen, for suggesting this term. The word " apoptosis " (:T6:cToaIa) is used in Greek to describe the" dropping off " or " falling off " of petals from flowers, or leaves from trees. To show the derivationclearly, we propose that the stress should be on the penultimate syllable, the second half of the wordbeing pronounced like " ptosis " (with the " p " silent), which comes from the same root " to fall "
and is already used to describe drooping of the upper eyelid.

J. F. R. KERR, A. H. WYLLIE AND A. R. CURRIE
PARENCHYMAL CELLS
residual bod
FIG 5 -Diagram to illustrate the morphological features of apoptosis.
f
I(
ragmentation
o
242
K&

A BASIC BIOLOGICAL PHENOMENON WITH WIDE-RANGING IMPLICATIONS 243
FIG 6.-24 Electron micrographs of sections of Epon-embedded tissues stained withuranyl acetate and lead citrate.
FIG. 6 and 7. Cluster of extracellular apoptotic bodies in atrophying rat liver lobe 3 days afterobstruction of its portal blood supply. Organelles are crowded together but appear essentiallyintact: the proportion of different cytoplasmic constituents varies from body to body. Densenuclear remnants (arrows) are present in some bodies but not in others. " Histiocytes " (H) containpartly degraded residues of phagocytosed bodies. Fig. 6: x 2500; Fig. 7: x 9000.

J. F. R. KERR, A. H. WYLLIE AND A. R. CURRIE
injury (Kerr, 1969, 1970, 1971, 1972aand b); in every case the ultrastructuralfeatures are essentially the same.
The formation of apoptotic bodiesinvolves marked condensation of bothnucleus and cytoplasm, nuclear fragmen-tation, and separation of protuberancesthat form on the cell surface (Fig. 5;Kerr, 1971) to produce many membrane-bounded, compact, but otherwise well-preserved cell remnants of greatly varyingsize (Fig. 5, 6, 7; see also Fig. 11, 12, 15).The initial morphological events havenot been identified: cells that are recog-nizable with certainty as undergoingapoptosis have already condensed andseparated from their neighbours, and thenuclear chromatin is aggregated in densemasses beneath the nuclear envelope(Kerr, 1971). Fully developed apoptoticbodies show closely packed organelles,which may themselves be condensed(Kerr, 1972b; Wyllie et al., 1972a), butwhich are apparently intact, both chemi-cally (Kerr, 1965, 1967; Ballard and Holt,1968) and structurally (Fig. 7 and 11;Klion and Schaffner, 1966; Farbman, 1968;Kerr, 1969, 1971, 1972a and b; Kerr andSearle, 1972a; Wyllie et al., 1972a;Wyllie et al., 1972b). Lucent cyto-plasmic vacuoles and dense masses ofnuclear material are seen in some bodies(Fig. 6). The content of an apoptoticbody depends on the cellular constituentsthat happened to be present in the cyto-plasmic protuberance that gave rise to it(Fig. 5); small bodies thus occasionallyconsist almost entirely of condensednuclear chromatin (Fig. 12), whereasothers are composed only of cytoplasmicelements (Fig. 7 and 11).
Apoptotic bodies frequently occur inclusters in the intercellular space (Fig. 6),and it is only the larger members ofsuch clusters that can be discerned withthe light microscope. The smaller bodiestend to disperse from their site of originand, in organs such as the liver andadrenal cortex, are often seen in thespaces between parenchymal and sinusoid-lining cells. Those arising from glandular
and mucosal epithelium and from renaltubules are frequently shed into thelumen (Fig. 1; Kerr, 1972a). A few mayenter blood vessels. It should be empha-sized that free extracellular bodies nevershow ultrastructural evidence of degenera-tion, and it is probable that at this stagethey are still capable of metabolic activity(Kerr, 1971) though irreversibly com-mitted to destruction.
The condensation is presumably aconsequence of the extrusion of water,but its mechanism is still unknown.Rough estimates of the degree of conden-sation suggest that the small membrane-bounded cell fragments might be formedwithout new synthesis of plasma mem-brane; detailed quantitative electron-microscope studies of serial sections ofclusters of apoptotic bodies would berequired to verify this impression.
In all the tissues so far studied, themajority of the apoptotic bodies havebeen found within the cytoplasm of intactcells. This suggests that they are rapidlyphagocytosed, possibly because of changesin the properties of their surface mem-branes. Those that develop under physio-logical conditions in post-natal life have,as yet, been seen only within connectivetissue cells (sinusoid-lining cells, " histio-cytes ") (Fig. 13 and 19; Wyllie et al.,1972a), but during embryonic developmentand in various pathological states in theadult, the bodies are also avidly ingestedby epithelial cells (Fig. 8; Farbman, 1968;Kerr, 1971, 1972a and b). Moreover, incarcinomata, apoptotic bodies appear tobe phagocytosed more frequently byneoplastic epithelial cells than by" histiocytes " (Fig. 20 and 23; Currieet al., 1972; Kerr and Searle, 1972a andb). The conventional functional distinc-tion between " histiocytes " and paren-chymal cells is clearly not an absolute one.Epithelial cells in adult animals can, whensuitably stimulated, display marked phago-cytic activity (Kerr, 1972a), and it isperhaps not surprising that in the embryo,where cellular functions are probably lesssharply demarcated, epithelial cells should
244

Fig. 10FIG. 8-10.-Atrophying rat liver tissue 3 days after obstruction of its portal blood supply. The twoapoptotic bodies indicated by arrows in Fig. 8 lie within phagosomes in the cytoplasm of a hepato-
cyte; their ergastoplasm is degenerate and their mitochondria are swollen and show focal matrixdensities (autolytic changes). Note that normal numbers of secondary lysosomes (L) are stillpresent in the cytoplasm bordering the bile canaliculus (BC). In Fig. 9, residues of degradedapoptotic bodies (arrows) are seen in the paracanalicular cytoplasm, and secondary lysosomes of thetype found in normal hepatocytes have disappeared, suggesting that they have previously fusedwith phagosomes. FIG. 10 shows a large apoptotic body without a nuclear remnant in the cyto-plasm of a "histiocyte " (H): note the autolytic changes. Fig. 8: x 4600; Fig. 9: X 9200; Fig.10: x 6600.

246 J. F. R. KERR, A. H. WYLLIE AND A. R. CURRIE
^
:::a i . s: ..*|: .:
*} v
.e..$. i,., .i.
a,
..:Oj.:,.* . .. : . ........ 2... X
w.
,r..; ..
;s} - e
:s... ffi
::'.S4>
...... .i.
.<,..
:.?:.
.t,.
X
,.l_.
.Iig. 1 1oigr. 12
FiG. 11-14.-Adrenal cortex of a healthy 5-day old rat. The extracellular apoptotic body illustratedin Fig. 11 shows closely aggregated but apparently intact mitochondria of epithelial cell type (M)and several lipid globules (L). Dense granular nuclear fragments (N) in extracellular bodies aredepicted in Fig. 12. A well-preserved apoptotic body within a " histiocyte " (H) is shown inFig. 13: the long arrow points to the membrane of the body and the short arrow to the phagosomemembrane. Fig. 14 illustrates a partly degraded apoptotic body in a " histiocyte ", which liesbetween an epithelial cell (E) and collagen fibres (C). Fig. 11: X 17,000; Fig. 12: x 14,000;Fig. 13: x 13,000; Fig. 14: x 10,500.

A BASIC BIOLOGICAL PHENOMENON WITH WIDE-RANGING IMPLICATIONS 247
Fig. 15 Fig. HI6
,I It¶
Fig. 17 Fig. IX
FIG. 15-18.-Paracordal mesoderm of a rat embryo killed 24 hours after administration of the terato-gen 7-hydroxymethyl-12-methylbenz(a)-anthracene to the mother on day 13 of pregnancy. Twoextracellular apoptotic bodies are shown in Fig. 15: note their compact cytoplasm and contrast thedensity of their nuclear remnants with the loose texture of the chromatin in the nucleus (N) of theadjacent intact mesenchymal cell. Fig. 16 and 17 show apoptotic bodies within the cytoplasm ofmesencyhmal cells: their ribosomes are closely aggregated and mitochondria (M) display autolyticchanges. A large complex lysosomal residual body, which is the result of fusion of a number oflysosomes containing remnants of phagocytosed apoptotic bodies in a mesenchymal cell, is illus-trated in Fig. 18. Fig. 15: x 9200; Fig. 16: x 18,000; Fig. 17: x 23,500; Fig. 18: x 9500.
.4 4.:
-.., I
J,:w: .4

248 J. F. R. KERR, A. H. WYLLIE AND A. R. CURRIE
.4AA .t
Fim. 19.-Adrenal cortex of a rat foetus killed after thrice-daily administration of 1 mg prednisolene tothe mother for 3 days, starting on the 17th day of gestation. An apoptotic body derived from anepithelial cell lies within the cytoplasm of a sinusoid-lining cell. Its mitochondria show earlyautolytic changes. x 16,000. Inset: x 25,500.
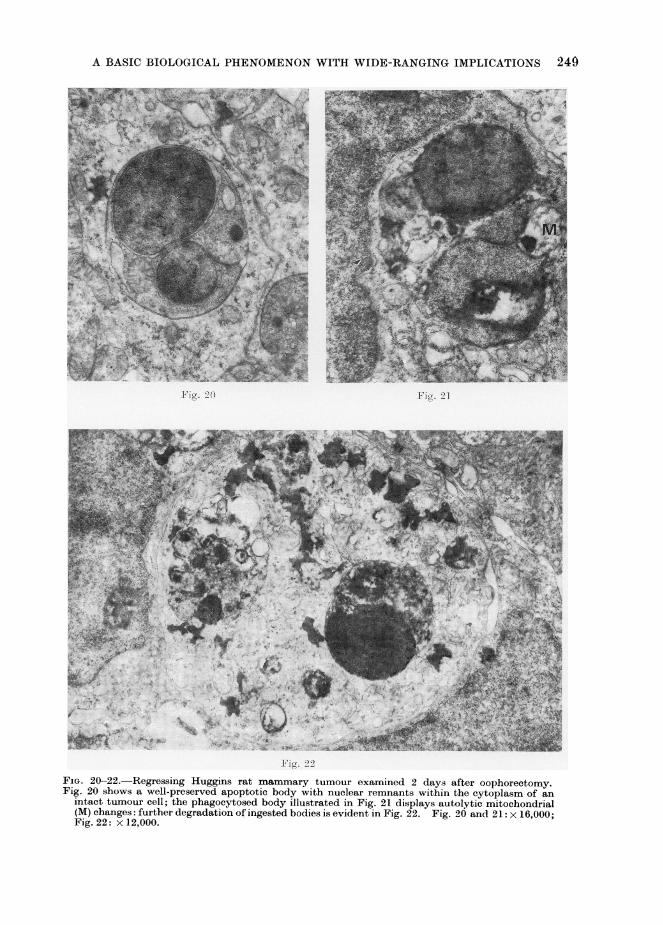
A BASIC BIOLOGICAL PHENOMENON WITH WIDE-RANGING IMPLICATIONS 249
Fig. 20 1'ig. 21
Fig. 22
FiG. 20-22. Regressing Huggins rat mammary tumour examined 2 days after oophorectomy.Fig. 20 shows a well-preserved apoptotic body with nuclear remnants within the cytoplasm of an
intact tumour cell; the phagocytosed body illustrated in Fig. 21 displays autolytic mitochondrial(M) changes: further degradation of ingested bodies is evident in Fig. 22. Fig. 20 and 21: x 16,000;Fig. 22: x 12,000.

J. F. R. KERR, A. H. WYLLIE AND A. R. CURRIE
take up large structures such as apoptoticbodies under physiological conditions.
Subsequent to their ingestion by othercells whether these be embryonic oradult, " histiocytic " or epithelial, normalor neoplastic apoptotic bodies undergoa process within phagosomes (Fig. 8 and10; Kerr, 1971, 1972a; Currie et al., 1972;Wyllie et al., 1972a) that is ultrastructur-ally very similar to ischaemic coagulativenecrosis (Kerr, 1970) and in vitro auto-lysis (Trump et al., 1965) of whole cells.The matrix of mitochondria becorneselectron-lucent and displays focal floccu-lent densities, the membranes of organellesand those bounding the bodies themselvesbreak down and ribosomes become swollenand indistinct. It should be noted thatthis is the first stage at which the bodiesexhibit changes that indubitably indicatecessation of co-ordinated metabolic acti-vity (Trump and Ginn, 1]969). The evi-dence suggests that lysosomes are notinvolved in the genesis of this degeneration(Fig. 8; Kerr, 1971, 1972a), and it seemslikely that autolysis of phagocytosedapoptotic bodies is a result of their inabilityto maintain chemical homoeostasis withinphagosomes: whether they are capable ofprolonged " survival " in the extracellularspace is unknown, since they are probablyalways either rapidly ingested or " sloughedoff" from mucosal surfaces soon aftertheir formation. Lysosomal enzymes do,however, play a vital role in the furtherdegradation of phagocytosed bodies(Ballard and Holt, 1968; Kerr, 1971;Kerr and Searle, 1972b), and these arerapidly reduced to electron-dense lyso-somal residual bodies (Fig. 9; Klion andSchaffner, 1966; Saunders, 1966; Saundersand Fallon, 1966; Farbman, 1968; Kerr,1971; Currie et al., 1972; Kerr and Searle,1972a and b; Wyllie et al., 1972a). Theacquisition of lysosomal hydrolases bythe phagosomes is associated with deple-tion of pre-existing secondary lysosomesin the cytoplasm of the ingesting cells(Fig. 9; Kerr, 1971; Kerr and Searle,1972b), indicating that they fuse with the
phagosomes. But there is little doubtthat the presence of these phagosomesalso evokes new synthesis of hydrolases(Ballard and Holt, 1968), and an increasein hydrolase content in response to theingestion of apoptotic bodies has alsobeen reported in neoplastic epithelialcells (Kerr and Searle, 1972b).
It is difficult to determine preciselythe time taken for the sequence of eventsdescribed above, since apoptosis is goingon continuously in normal foetal andadult tissues and growing neoplasms;even when augmented by various stimuli,the process appears to start in individualcells of the same organ or tissue atdifferent times. However, examinationof the serial changes that take place inseveral experimental models (Kerr, 1971;Crawford et al., 1972; Wyllie et al., 1972b)suggests that the process is completedfairly rapidly: bodies may form anddisappear within 24 hours. Partly de-graded remnants of apoptotic bodies aredifficult to discern histologically andelectron microscopy shows that bodiesthat can be detected with the light micro-scope comprise only a small fraction ofthe total number of cell remnants present(Kerr and Searle, 1972b).
It is important to appreciate that thefinding of relatively few apoptotic bodiesin histological sections of a tissue meansthat quite extensive cell " drop-out " istaking place: mitotic figures have ananalagous significance for cell prolifera-tion.
Apoptosis is well suited to a role intissue homoeostasis, since it can result inextensive deletion of cells with littletissue disruption. Following fragmenta-tion of an affected cell, the remains arerapidly disposed of by nearby intact cells.There is no inflammation, as is elicited bycoagulative necrosis, and even the lyso-somal residual bodies soon disappear,possibly as a result of cell defaecation(Kerr, 1971, 1972a). Moreover, the pro-cess is economical in terms of re-utilizationof cell components.
250

A BASIC BIOLOGICAL PHENOMENON WITH WIDE-RANGING IMPLICATIONS 251
Fig. 23
Fig. 24
Fia. 23 and 24. Untreated squamous cell carcinoma of the human cervix uteri. Apoptotic bodieswith nuclear remnants are seen within carcinoma cells, which can readily be identified by thepresence of tonofibrils (T). Autolytic changes are evident in the body illustrated in Fig. 23: moreadvanced degradation is shown in Fig. 24. Fig. 23: x 14,200; Fig. 24: x 16,000.

J. F. R. KERR, A. H. WYLLIE AND A. R. CURRIE
THE OCCURRENCE AND IMPLICATIONSOF APOPTOSIS
The size of a cell population, whetherneoplastic or not, depends on the balancebetween cell production and cell loss.An enormous amount of work has beendone over many years on multiplicationof cells under various circumstances; bycontrast, relatively little attention hasbeen paid to controlled cell deletion. Webelieve that enough is now known aboutthe occurrence of apoptosis to establishit as an important, and possibly the onlymode of controlled cell death, whichcontributes to the regulation of cellpopulations in a variety of mammaliantissues under many different conditions.
Small numbers of apoptotic bodiescan be found in histological sections ofmany healthy tissues (Fig. 1 and 3a), andthere seems little doubt that apoptosisplays an important role in the regulationof normal cell populations. The sparse-ness of the bodies in most tissues makesultrastructural studies very difficult, butelectron microscopy of the normal neo-natal rat adrenal cortex, where apoptosisis enhanced as a result of temporaryphysiological ACTH-deprivation (Wyllieet al., 1972a), shows the process to conformexactly (Fig. 11-14) to the general des-cription already given. Much more workis needed to determine the extent andfrequency of apoptosis in the organs andtissues of healthy adult animals, andpractically nothing is known about thefactors that determine which cells willbe affected.
An interesting though poorly under-stood manifestation of apoptosis in healthyanimals is the occurrence of so-calledtingible bodies in the germinal centres oflymphoid follicles. They show the typicalhistological features of apoptotic bodies,and examination of electron micrographspublished by Swartzendruber and Congdon(1963) indicates to us that they undergothe classic sequence of changes followingphagocytosis. Many of them have beenshown to be derived from cells that haverecently synthesized DNA (Fliedner, 1967;
Odartchenko et al., 1967) and it has beensuggested that cell death in lymphoidfollicles might be an inevitable conse-quence of rapid cell proliferation (Yoffeyand Courtice, 1970). However, we havenot been able to detect an increase inapoptosis in the regenerating liver rem-nant examined at varying times afterpartial hepatectomy in the rat, and in theembryo the distribution of apoptotic celldeletion has been shown to be focal andhighly specific, and is certainly notmerely a manifestation of rapid cellmultiplication (Gluicksmann, 1951;Saunders, 1966; Menkes, Sandor and Ilies,1970).
Perhaps because of its frequentlymassive dimensions in the embryo, thesignificance of apoptosis in vertebrateontogeny was recognized early, and recentelectron microscope studies (Fig. 15-18;Saunders and Fallon, 1966; Farbman,1968; Webster and Gross, 1970; Crawfordet al., 1972) have shown that the ultra-structural features of the embryonicprocess are essentially the same as thoseobserved during post-natal life. The mor-phological appearances have not, however,always been correctly interpreted: asrecently as 1970, Menkes and his colleaguessuggested that Feulgen-positive, baso-philic " necrospherules " within the cyto-plasm of other embryonic cells mightrepresent signs of early degeneration ofthe latter, whereas their structure seemsto us typical of phagocytosed apoptoticbodies. A similar misconception may tosome extent account for the delay in theappreciation of the significance of apop-tosis in adult tissues and neoplasms: aswill be emphasized later, it is likely thatapoptotic bodies within heterophagosomeshave sometimes been mistakenly identifiedas autophagic vacuoles.
Focal apoptosis plays a vital role inmany normal embryonic processes suchas the development of the lumina oftubular structures, the fashioning oflimbs, the formation of interdigital clefts,and the involution of phylogenetic vestiges(Glticksmann, 1951; Saunders, 1966;
252

A BASIC BIOLOGICAL PHENOMENON WITH WIDE-RANGING IMPLICATIONS 253
Menkes et al., 1970). Its appearance isprecisely controlled, probably by diffusiblesubstances: the susceptibility of groups ofcells depends on their position in theembryo and the stage of developmentthat has been reached (Saunders, 1966;Saunders and Fallon, 1966). It is inter-esting that hormones that cause prolifera-tion and differentiation of cells in some
situations may cause, at the same time,apoptosis of others: for example, indeveloping amphibians, thyroid hormonestimulates both general body growth andwidespread cell death in the gills andtail (Saunders, 1966).
Apart from its role in normal onto-genesis, apoptosis is also important interatogenesis. A number of teratogenicagents have been found to producemassive apoptosis at their site of action(Fig. 15-18; Menkes et al., 1970; Crawfordet al., 1972), and in at least some cases,the subsequent congenital malformationshave been fully accounted for by thedistribution of focal apoptosis observedwithin a few hours of treatment (Menkeset al., 1970; Crawford et al., 1972).
In the pathogenesis of such congenitalmalformations, apoptosis greatly exceedsmitosis in localized areas, effecting a netreduction in cell numbers. By contrast,in growing malignant neoplasms cell num-bers progressively increase. Nevertheless,as has already been pointed out, manyrecent studies have shown that spon-taneous and continuous death of cells isan inherent property of malignant neo-plasms. However, there has been littledetailed investigation of the associatedmorphology; the foci of coagulativenecrosis seen in some lesions obviouslycould not account for the kinetic data.
Apoptotic bodies have been foundhistologically in all the malignant neo-
plasms we have examined (Fig. 4) andthey are frequently extremely numerous.Many are rather inconspicuous and somepractice is needed to identify the smallerbodies with the light microscope, especi-ally those without a nuclear component.
Electron microscope studies, so far19
performed on human basal (Kerr andSearle, 1972a and b) and squamous(Searle, unpublished observations) cellcarcinomata, and on Huggins rat mam-mary tumours induced by 7,12-dimethyl-benz(a)anthracene (DMBA) (Currie et al.,1972), have shown that the apoptoticbodies are essentially the same in struc-ture as those derived from non-neoplasticcells and that whilst a few of the bodiesare taken up by " histiocytes ", themajority are rapidly phagocytosed byintact tumour cells (Fig. 20 and 23).Progressive stages in the degradation ofthe bodies by tumour cells can be tracedwith the electron microscope (Fig. 21,22 and 24) and the participation of lyso-somal hydrolases synthesized in the tum-our cells has been substantiated in thecase of basal cell carcinomata by theapplication of electron histochemical tech-niques (Kerr and Searle, 1972b).
The kinetic significance of findingmoderate numbers of apoptotic bodies inhistological sections of a tissue hasalready been stressed, and electron micro-scopy of basal cell carcinomata leaves nodoubt that apoptosis accounts for exten-sive and continuous deletion of cells fromthese tumours (Kerr and Searle, 1 972aand b). Both apoptotic bodies and mito-tic figures are sometimes numerous inrapidly growing tumours; it is the balancebetween the two processes that determinesthe rate of enlargement.
The spontaneous occurrence of apop-tosis in growing malignant neoplasmssuggests that it might also be implicatedin some types of therapeutically inducedtumour regression, but few investigationshave so far been undertaken. Prelim-inary studies of human squamous cellcarcinomata indicate that there is anincrease in apoptosis after irradiation(Kerr and Searle, 1972b), and the regres-sion of Huggins rat mammary tumoursthat follows oophorectomy has beenshown to be associated with extensiveand diffuse apoptotic deletion of tumourcells (Currie et al., 1972). A decrease inthe size of individual cells is, of course,

J. F. R. KERR, A. H. WYLLIE AND A. R. CURRIE
also often of great importance (Scott,Christian and Currie, 1967) and auto-phagocytosis probably plays a part in itsgenesis (Scott et al., 1967; Anton andBrandes, 1968; Paris and Brandes, 1971).However, it seems to us likely that someof the structures that have been identifiedas autophagic vacuoles in the past mightreally have been ingested apoptotic bodies.It is possible that histological assessmentof the " apoptotic index " of a tumourseveral days after the commencementof therapy might, in some cases, provide auseful measure of its response, but moreexperimental work is needed before thissuggestion is applied to man for thesituation is complex and cells may alsobe deleted in other ways (Scott et al.,1967).A rather similar situation pertains in
the atrophy and involution of non-neo-plastic tissues and organs, where adecrease in cell numbers is probably oftenas important as a decrease in the size ofindividual cells. Whilst it has beensuggested that autophagocytosis mightcontribute to the development of thelatter (Helminen, Ericsson and Niemi,1970; Cole, Matter and Karnovsky, 1971;Helminen and Ericsson, 1971), little orno attention has been paid to the way inwhich cells are deleted.
We have found that apoptosis plays asignificant part in at least some types ofatrophy and involution. Perhaps themost thoroughly studied is the gross andrapid reduction in size of rat liver lobesthat follows ligation of their portal bloodsupply; here there is enhancement ofautophagocytosis associated with diffuseshrinkage of hepatocytes within hours ofoperation (Cole et al., 1971), and this isfollowed by massive apoptotic deletion ofliver cells, which reaches a peak severaldays later (Fig. 6-10; Kerr, 1971).Apoptosis is also prominent in certaindifferentiated tissues undergoing atrophyas a result of withdrawal or administra-tion of hormones. Thus, following areduction in the blood concentration ofACTH in rats, a shower of apoptotic
bodies appears in the adrenal cortex(Fig. 19), an occurrence that can beprevented by administration of exogenousACTH (Wyllie et al., 1972b); several daysafter orchidectomy in the rat, numerousapoptotic bodies are seen amongst thesomewhat shrunken epithelial cells thatline the prostatic glandular acini (Fig. 2);published descriptions of the processresponsible for the deletion of lymphoidcells that follows large doses of gluco-corticoids (Haelst, 1967; Makman,Nakagawa and White, 1967; Abraham,Morris and Hendy, 1969; La Pushin andde Harven, 1971) indicate to us that thisis also an example of apoptosis. Finally,apoptotic bodies are quite numerous inthe involuting human corpus luteum(Searle, unpublished observations).
There is little doubt that phagocytosedapoptotic bodies have sometimes beenmistaken for autophagic vacuoles inelectron microscopic studies of atrophyand involution (see, for example, Fig. 6in Helminen and Ericsson, 1971). Thedistinction may be difficult unless recog-nizable nuclear remnants are present inthe ingested apoptotic bodies (Kerr,1972a), but it is crucial to the correctunderstanding of many problems in cellpopulation kinetics.
The evidence for the participation ofapoptosis in the various types of cellpopulation change considered so far hasbeen founded on detailed observation.We should now like to speculate thathyperplasia might sometimes result fromdecreased apoptosis rather than increasedmitosis, although we emphasize that weknow of no definitive studies that supportsuch an hypothesis. It seems to uspossible that focal hyperplasia in tissuessubject to cyclic hormonal stimulationsuch as the breast might be due to failureof clones of cells to respond in the normalway to falling hormone concentrationsby undergoing apoptosis.
We have an impression that apoptoticbodies are rare in benign neoplasms, butmuch more work is required to showwhether this is a constant finding.
254

A BASIC BIOLOGICAL PHENOMENON WITH WIDE-RANGING IMPLICATIONS 255
In view of what has been said aboutthe occurrence of apoptosis under physio-logical conditions, it is perhaps somewhatsurprising to find that it is also augmen-ted in " tissue injury ", often developingin association with focal coagulativenecrosis, both in vivo (Kerr, 1971) andin vitro (L. T. Hou, personal communica-tion). Electron microscopic studies ofthe augmented apoptosis occurring in avariety of types of liver injury (Biavaand Mukhlova-Montiel, 1965; Klion andSchaffner, 1966; Moppert, Ekesparre andBianchi, 1967; Kerr, 1969, 1970, 1971)show that the morphological changesconform to the usual stereotyped pattern,and ultrastructurally typical apoptosiscan be induced in the rat adrenal cortexby administration ofthe adrenocorticolyticagent DMBA (Kerr, 1972b). Examina-tion of published electron micrographsof so-called individual-cell dyskeratosisproduced in the epidermis by ultravioletirradiation (Wilgram et al., 1970) suggeststo us that this is also an example ofapoptosis.
It is evident that certain agents(hepatotoxins, electromagnetic radiation,DMBA) are capable of inducing eithercoagulative necrosis or apoptosis of cells,as well as having both carcinogenic andteratogenic properties. However, the sig-nificance of this observation is obscure.
FACTORS INITIATING APOPTOSIS
Little is known of the factors thatinitiate apoptosis or of the nature of thecellular mechanisms activated before theappearance of the characteristic mor-phological changes. It seems clear, how-ever, that in certain circumstances apop-tosis is an inherently programmed event,determined by intrinsic " clocks " specificfor the cell type involved. Thus in avianembryonic tissue explants apoptosis oc-curred in susceptible zones " on schedule "(Saunders and Fallon, 1966). But evenhere, some degree of environmental con-trol is evident; diffusible substances fromadjacent tissues are capable of delaying the
process, sometimes indefinitely (Saundersand Fallon, 1966).
Although the nature of many of thesediffusible substances is still obscure, ster-oid hormones are known to affect apoptosisin the Mullerian ducts of chick embryos,oestrogenic steroids inhibiting the expec-ted Mullerian regression in genetic males,and androgenic steroids promoting regres-sion in genetic females (Menkes et al.,1970). As we have already shown, therole of hormones in modulating apoptosisis not restricted to ontogenesis; apoptosiscan be promoted or inhibited in certaindifferentiated mammalian tissues by hor-mone withdrawal or stimulation.
The mode of action of triggers andinhibitors of apoptosis is unknown, butit is tempting to speculate that it mightinvolve stimulation of messenger RNAand protein synthesis. If indeed apop-tosis depends on expression of part of thegenome, which is normally repressed inviable cells, the initiation of such a stereo-typed series of changes by a wide varietyof stimuli would be understandable.
It is obvious that much work has stillto be done on the factors that determinethe occurrence and extent of apoptosis inmalignant neoplasms. Ischaemia probablyaccounts for the dense clusters of apopto-tic bodies often seen in the neighbourhoodof coagulative necrosis. However, weknow little about the aetiology of diffuseapoptosis: nutritional, hormonal and im-mune factors, and intrinsic controls,perhaps including cell ageing, may beinvolved.
THE CONCEPT OF APOPTOSIS:CONCLUSIONS
Though certain of the morphologicalmanifestations of apoptosis have longbeen recognized at the light microscopelevel, its ultrastructural features havebeen described only recently. The impli-cations of the process in tissue kineticshave not hitherto been appreciated, exceptby embryologists. We believe that theevidence presented here establishes itnot only as a distinctive morphological

256 J. F. R. KERR, A. H. WYLLIE AND A. R. CURRIE
process but also as an important basicbiological phenomenon, which plays acomplementary but opposite role to mito-sis in the regulation of animal cell popula-tions. Thus, it is involved in cell turn-over in many normal tissues and accountsfor extensive spontaneous cell loss inmalignant neoplasms; it is implicated intissue atrophy and involution and itplays a part in at least some types oftumour regression; it is of major impor-tance in both normal ontogenesis andteratogenesis. The ultrastructural fea-tures of apoptosis and its initiation andinhibition by a variety of environmentalstimuli suggest to us that it is an active,controlled process.
Many challenging problems obviouslyremain to be solved, the factors thatdetermine which cells will be affected,the role of ageing, the mode of action ofinitiators and inhibitors, the earliest bio-chemical and morphological events, andthe mechanism of the cellular condensationbeing but a few.
This work was supported in part bythe Scottish Hospital Endowments Re-search Trust and in by part the NationalHealth and Medical Research Council ofAustralia. A. H. W. held a JuniorResearch Fellowship of the MedicalResearch Council. We are grateful toDr J. Searle and Mr D. Collins for pro-viding Fig. 1, 23 and 24, and to Dr D. N.Wheatley for the histological sections ofregenerating rat liver. We thank thefollowing for their technical assistance:Mrs Marget Inglis, Mr Peter MacLennan,Mr George Milne, Mr Alastair McKinnon,Mr Robert Cardno, Miss Lynda Hardyand Mr Brian Harmon.
REFERENCESABRAHAM, R., MORRIS, M. & HENDY, R. (1969)
Lysosomal Changes in Epithelial Cells of theMouse Thymus after Hydrocortisone Treatment.Hi8tochemie, 17, 295.
ANTON, E. & BRANDES, D. (1968) Lysomes in MiceMammary Tumors Treated with Cyclophos-phamide. Cancer, N.Y., 21, 483.
BALLARD, K. J. & HOLT, S. J. (1968) Cytologicaland Cytochemical Studies on Cell Death and
Digestion in the Foetal Rat Foot: the Role ofMacrophages and Hydrolytic Enzymes. J. CellSci., 3, 245.
BIAVA, C. & MUKHLOVA-MONTIEL, M. (1965) Elec-tron Microscopic Observations on Councilman-like Acidophilic Bodies and Other Forms ofAcidophilic Changes in Human Liver Cells.Am. J. Path., 46, 775.
CLIFTON, K. H. & YATVIN, M. B. (1970) Cell Popula-tion Growth and Cell Loss in the MTG-B MouseMammary Carcinoma. Cancer Res., 30, 658.
COLE, S., MATTER, A. &IKARNOVSKY, M. J. (1971)Autophagic Vacuoles in Experimental Atrophy.Exp. mol. Pathol., 14, 158.
CRAWFORD, A. M., KERR, J. F. R. & CURRIE, A. R.(1972) The Relationship of Acute MesodermalCell Death to the Teratogenic Effects of 7-OHM-12-MBA in the Fetal Rat. Br. J. Cancer. Inpress.
CURRIE, A. R., KERR, J. F. R., SCOTT, G. B. &INGLIS, M. S. (1972) Regression of Endocrine-dependent Mammary Tumours: the Role ofApoptosis. Br. J. Cancer. In press.
FARBMAN, A. I. (1968) Electron Microscope Studyof Palate Fusion in Mouse Embryos. Devl Biol.,18, 93.
FLIEDNER, T. M. (1967) On the Origin of TingibleBodies in Germinal Centers. In Germinal Centersin Immune Responses. Ed. H. Cottier, N.Odartchenko, R. Schindler and C. C. Congdon.Berlin: Springer. p. 218.
FRINDEL, E., MALAISE, E. & TUBIANA, M. (1968)Cell Proliferation Kinetics in Five Human SolidTumors. Cancer, N. Y., 22, 611.
GLtCKSMANN, A. (1951) Cell Deaths in NormalVertebrate Ontogeny. Biol. Rev., 26, 59.
GOLDSMITH, M. (1966) The Anatomy of Cell Death.J. Cell Biol., 31, 41A.
HAELST, U. VAN (1967) Light and Electron Micro-scopic Study of the Normal and PathologicalThymus of the Rat. II The Acute ThymicInvolution. Z. Zellforsch. mikrosk. Anat., 80, 153.
HELMINEN, H. J. & ERICSSON, J. L. E. (1971)Ultrastructural Studies on Prostatic Involutionin the Rat. Mechanism of Autophagy in Epithel-ial Cells, with Special Reference to the Rough-surfaced Endoplasmic Reticulum. J. Ultra-strUCt. Res., 36, 708.
HELMINEN, H. J., ERICSSON, J. L. E. & NIEMI, M.(1970) Lysosomal Changes during Castration-induced Prostatic Involution in the Rat. Actapath. microbiol. scand., 78A, 493.
IVERSEN, 0. H. (1967) Kinetics of Cellular Prolifera-tion and Cell Loss in Human Carcinomas. ADiscussion of Methods Available for in vivoStudies. Eur. J. Cancer, 3, 389.
JUDAH, J. D., AHMED, K. & McLEAN, A. E. M.(1965) Pathogenesis of Cell Necrosis. FednProc. Fedn Am. Socs exp. Biol., 24, 1217.
KERR, J. F. R. (1965) A Histochemical Study ofHypertrophy and Ischaemic Injury of Rat Liverwith Special Reference to Changes in Lysosomes.J. Path. Bact., 90, 419.
KERR, J. F. R. (1967) Lysosome Changes in AcuteLiver Injury due to Heliotrine. J. Path. Bact.,93, 167.
KERR, J. F. R. (1969) An Electron-microscope Studyof Liver Cell Necrosis due to Heliotrine. J. Path.,97, 557.
KERR, J. F. R. (1970) An Electron Microscopic

A BASIC BIOLOGICAL PHENOMENON WITH WIDE-RANGING IMPLICATIONS 257
Study of Liver Cell Necrosis due to Albitocin.Pathology,2, 251.
KERR, J. F. R. (1971) Shrinkage Necrosis: a DistinctMode of Cellular Death. J. Path., 105, 13.
KERR, J. F. R. (1972a) Some Lysosome Functionsin Liver Cells Reacting to Sublethal Injury. InLysosomes in Biology and Pathology, Vol. 3.Ed. J. T. Dingle and Honor B. Fell. Amsterdam:North-Holland. In press.
KERR, J. F. R. (1972b) Shrinkage Necrosis of Adre-nal Cortical Cells. J. Path. In press.
KERR, J. F. R. & SEARLE, J. (1972a) A SuggestedExplanation for the Paradoxically Slow GrowthRate of Basal-cell Carcinomas that ContainNumerous Mitotic Figures. J. Path. In press.
KERR, J. F. R. & SEARLE, J. (1972b) The Digestionof Cellular Fragments within Phago-lysosomes inCarcinoma Cells. J. Path. In press.
KLION, F. M. & SCHAFFNER, F. (1966) The Ultra-structure of Acidophilic " Councilman-like"Bodies in the Liver. Am. J. Path., 48, 755.
LAIRD, A. K. (1969) Dynamics of Growth in Tumorsand in Normal Organisms. In Human TumorCell KinetiCs. Ed. i. Perry. Natn. Cancer Inst.Monogr., No. 30. p. 15.
LALA, P. K. (1971) Studies on Tumor Cell Popula-tion Kinetics. In Methods in Cancer Research,Vol. 6. Ed. H. Busch. New York: AcademicPress. p. 3.
LALA, P. K. (1972) Evaluation of the Mode of CellDeath in Ehrlich Ascites Tumor. Cancer, N.Y.,29, 261.
LA PUSHIN, R. W. & DE HARVEN, E. (1971) AStudy of Gluco-corticosteroid-induced Pyknosisin the Thymus and Lymph Node of the Adrenal-ectomized Rat. J. Cell Biol., 50, 583.
MAKMAN, M. H., NAKAGAwA, S. & WHITE, A. (1967)Studies of the Mode of Action of Adrenal Steroidson Lymphocytes. Recent Prog. Horm. Res., 23,195.
MENKES, B., SANDOR, S. & ILIES, A. (1970) CellDeath in Teratogenesis. In Advances In Tera-tology, Vol. 4. Ed. D. H. M. Woollam. London:Logos Press. p. 169.
MOPPERT, J., EKESPARRE, D. V. & BIANCHI, L.(1967) Zur Morphogenese der eosinophilen Ein-zelzellnekros im Leberparenchym des Menschen.Eine licht-und elektronenoptisch korrelierte Unter-suchung. Virchows Arch. path. Anat. Physiol.,342,210.
ODARTCHENKO, N., LEWERENZ, M., SORDAT, B.,Roos, B. & COTTIER, H. (1967) Kinetics ofCellular Death in Germinal Centers of MouseSpleen. In Germinal Centers in Immune Respon-ses. Ed. H. Cottier, N. Odartchenko, R.Schindler and C. C. Congdon. Berlin: Springer.p. 212.
PARIS, J. E. & BRANDEs, D. (1971) Effect of X-
irradiation on the Functional Status of LysosomalEnzymes of Mouse Mammary Gland Carcinomas.Cancer Res., 31, 392.
REFSUM, S. B. & BERDAL, P. (1967) Cell Loss inMalignant Tumours in Man. Eur. J. Cancer, 3,235.
SAUNDERS, J. W. (1966) Death in EmbryonicSystems. Science, N. Y., 154, 604.
SAUNDERS, J. W. & FALLON, J. F. (1966) CellDeath in Morphogenesis. In Major Problemsin Developmental Biology. Ed. M. Locke. NewYork: Academic Press. p. 289.
SCOTT, G. B., CHRISTIAN, H. J. & CURRIE, A. R.(1967) The Huggins Rat Mammary Tumors:Cellular Changes Associated with Regression. InEndogenous Factors Influencing Host-Tumor Bal-ance. Ed. R. W. Wissler, T. L. Dao and S.Wood, Jr. Chicago: University of Chicago Press.p. 99.
STEEL, G. G. (1967) Cell Loss as a Factor in theGrowth Rate of Human Tumours. Eur. J.Cancer, 3, 381.
SWARTZENDRUBER, D. C. & CONGDON, C. C. (1963)Electron Microscope Observations on TingibleBody Macrophages in Mouse Spleen. J. CellBiol., 19, 641.
TRUMP, B. F. & GINN, F. L. (1969) The Pathogenesisof Subcellular Reaction to Lethal Injury. InMethods and Achievements in ExperimentalPathology, Vol. 4. Ed. E. Bajusz and G. Jasmin.Basel: Karger, p. 1.
TRUMP, B. F., GOLDBLATT, P. J. & STOWELL, R. E.(1965) Studies on Necrosis of Mouse Liver invitro. Ultrastructural Alterations in the Mito-chondria of Hepatic Parenchymal Cells. Lab.Invest., 14, 343.
WEBSTER, D. A. & GRoss, J. (1970) Studies onPossible Mechanisms of Programmed Cell Deathin the Chick Embryo. Devl Biol., 22, 157.
WEINSTEIN, G. D. & FROST, P. (1970) Cell Prolifera-tion in Human Basal Cell Carcinoma. CancerRes., 30, 724.
WILGRAM, G. F., KIDD, R. L., KRAWCZYK, W. S.& COLE, P. L. (1970) Sunburn Effect on Keratino-somes. A Report with Special Note of Ultra-violet-induced Dyskeratosis. Archs Derm., 101,505.
WYLLIE, A. H., KERR, J. F. R. & CURRIE, A. R.(1972a) Cell Death in the Normal Neonatal RatAdrenal Cortex. J. Path. (To be published).
WYLLIE, A. H., KERR, J. F. R., MACASKILL, I. A. M.& CURRIE, A. R. (1972b) Adrenocortical CellDeletion: the Role of ACTH. J. Path. (To bepublished).
YOFFEY, J. M. & COURTICE, F. C. (1970) Lympha-tics, Lymph and the Lymphomyeloid Complex.New York: Academic Press. p. 534.