brain-derived neurotrophic factor promotes growth and migration of multiple myeloma cells
TRANSCRIPT

Cancer Genetics and Cytogenetics 169 (2006) 12–20
Brain-derived neurotrophic factor promotes growth and migrationof multiple myeloma cells
Yu Hua,1,*, Chun-yan Suna,1, Hua-fang Wanga, Tao Guoa, Wen-ning Weia, Ya-dan Wanga,Wen-juan Hea, Tao Wua, Hao Tanb, Tang-chun Wub
aInstitute of Hematology, Union Hospital, Tongji Medical College, Huazhong University of Science and Technology,
1277 Jiefang Dadao, Wuhan 430022, P.R. ChinabInstitute of Occupational Medicine, School of Public Health, Tongji Medical College, Huazhong University of Science and Technology,
13 Hangkong Road, Wuhan 430030, P.R. China
Received 7 September 2005; received in revised form 22 February 2006; accepted 27 February 2006
Abstract The evolution of multiple myeloma (MM) depends on complex signals from the bone marrowmicroenvironment, which support proliferation and survival of malignant plasma cells. Previousstudy defined a brain-derived neurotrophic factor–tyrosine kinase receptor B (BDNF/TrkB) axisin myeloma and autocrine growth stimulation by BDNF in various tumor cells. We examined thebiological effects of BDNF on MM cells. Using a reverse transcriptase–polymerase chain reaction,Western blotting, and immunohistochemistry, we observed that both BDNF and its high-affinity re-ceptor TrkB are expressed by MM cell lines (RPMI 8226, U266, and KM3) and primary MM cells.Functional studies revealed that BDNF was a potent growth factor for MM. BDNF (5–500 ng/mL)had strong proliferative effects on both MM cell lines and primary MM cells, shown by [3H]thymi-dine incorporation assay. BDNF (12.5–200 ng/mL) also induced migration of MM cells, as indi-cated by the Transwell migration assay. Together, our data indicate that BDNF is a potentmyeloma growth and chemotactic factor and suggest that the BDNF/TrkB pathway is a potentialtherapeutic target in MM. � 2006 Elsevier Inc. All rights reserved.
1. Introduction
Multiple myeloma (MM) is a B-cell neoplasm character-ized by bone marrow infiltration of malignant plasma cellsand accounts for ~1–2% of all human cancers. Malignanttransformation in MM evolves through different stagesfrom monoclonal gammopathy of undetermined signifi-cance to expansive and aggressive, plasmablastic MM.Most of the tumor evolution in MM takes place in the bonemarrow, indicating that cell adhesion-mediated autocrine orparacrine activation of various cytokines such as interleukin6 (IL-6), insulin-like growth factor 1 (IGF-1), hepatocytegrowth factor (HGF), and vascular endothelial growth fac-tor (VEGF) have a pivotal role in the growth, survival, drugresistance, and malignant progression of MM cells [1–4].Despite recent insights into the pathogenesis of MM, thecondition remains incurable, with a median survival of
1 These authors contributed equally to this work.
* Corresponding author. Tel.: 186-27-62766183; fax: 186-27-85776343.
E-mail address: [email protected] (Y. Hu).
0165-4608/06/$ – see front matter � 2006 Elsevier Inc. All rights reserved.
doi:10.1016/j.cancergencyto.2006.02.018
~3–4 years. New biologically based therapies are thereforeurgently needed.
Neurotrophins (NTs) are a family of related polypeptidegrowth factors, including nerve growth factor (NGF), brain-derived neurotrophic factor (BDNF), NT-3, NT-4/5, andNT-6 [5]. NTs play an important and well-described rolein the development, function, and survival of neuronal cells[6]. The activities of NTs are mediated by tyrosine kinasereceptors (TrkA, TrkB, and TrkC) and by P75NTR, a mem-ber of the death receptor family, of which the signaling maysynergize or antagonize tropomysin-related kinase signal-ing, depending on cell context [7].
Despite their well-known effects on neurons, elevatedneurotrophin concentrations have been observed under var-ious pathological conditions [8–10]. NT activation of Trkhas been implicated in the pathogenesis of a number ofneuronal and nonneuronal solid tumors [11–19]. BDNF,in particular, has been shown to have protective effectson chemotherapy-induced apoptosis [20,21]. All thesebiological effects of BDNF are transduced via TrkB, andexpression of TrkB has been shown to increase metastaticpotential by suppressing anoikis (i.e., apoptosis resulting

13Y. Hu et al. / Cancer Genetics and Cytogenetics 169 (2006) 12–20
from loss of cell–matrix interactions) [22]. Recently, it hasbeen reported that BDNF or BDNF and TrkB are also pro-duced in hematological malignancies, including multiplemyeloma [23,24]; however, the significance of this expres-sion is not completely known.
We therefore examined the biological effects of BDNFand TrkB on MM cells with the goal of clarifying thepathways involved and identifying potential therapeutictargets.
2. Materials and methods
2.1. Materials
Human MM cell lines (HMCLs) RPMI 8226 and U266were obtained from the American Type Culture Collection(ATCC, Manassas, VA). The KM3 cell line was kindly pro-vided by Jan Hou (Senond Military Medical University,Shanghai, P.R. China).
All tumor cell lines were maintained in culture in RPMI1640 supplemented with 10% heat-inactivated fetal bovineserum (FBS) and antibiotics (100 units/mL penicillin and100 mg/mL streptomycin) in an atmosphere of 5% CO2 at37�C. Recombinant human BDNF was obtained fromPeproTech (Princeton, NJ). Rabbit anti-human BDNF (SC-546), rabbit anti-human TrkB (SC-12), and rabbit anti-hu-man P75NTR antibodies (SC-8317) were obtained fromSanta Cruz Biotechnology (Santa Cruz, CA). Rabbit anti-pTrkB (9141) was obtained from Cell Signaling Technology(Beverly, MA). Goat anti-rabbit horseradish peroxidase(HRP) conjugate was purchased from Transduction Labo-ratories (Lexington, UK). S-P kit was purchased fromZhongshan Biotechnology (Beijing, China). EnhancedChemiluminescence (ECL) reagent and [3H]thymidine(25 Ci/mmol) were obtained from Amersham Biosciences(Piscataway, NJ). Penicillin, streptomycin, RPMI 1640,and FBS were obtained from Gibco Life Technologies (Bur-lington, Ontario, Canada). The TRIZOL reagent was obtainedfrom Invitrogen (Grand Island, NY). A Transwell plate wasobtained from Corning Costar (Cambridge, MA). Randomprimers were obtained from Shanghai Bioasia (Shanghai,China). RevertAid first-strand cDNA synthesis kit was pur-chased from Fermentas (Burlington, Ontario, Canada). Taqpolymerase was obtained from Shanghai CASarray (Shang-hai, China). The pharmacological inhibitor of TrkB, K252a(1 mmol/L) was obtained from Sigma (St. Louis, MO).
2.2. Isolation of primary myeloma cells
Bone marrow cells from 12 patients with active mye-loma were first separated by means of Ficoll–Hypaque den-sity centrifugation. The resulting mononuclear cells werethen further separated using anti-human CD138 microbeads(Miltenyi Biotech, Auburn, CA) according to the manufac-turer’s protocol. The separated cells consisted of O90%plasma cells.
2.3. Reverse-transcriptase polymerase chain reactionanalyses of BDNF and BDNF receptors
Total RNA was prepared with TRIZOL reagent accordingto manufacturer’s instructions. Complementary DNA(cDNA) was subsequently synthesized from total RNA us-ing RevertAid first-strand cDNA synthesis kit, and reversetranscriptase–polymerase chain reaction (RT-PCR) wasperformed using a PCR thermal cycler (Eppendorf, Ham-burg, Germany). The PCR program consisted of denaturat-ing at 94�C for 1 min, annealing at 55�C for 1 min, andextension at 72�C for 1 min. The samples were amplifiedfor 35 cycles. b-actin was used as an internal control.The primer sequence were as follows: for BDNF, forwardprimer 50-GCA GCC TTC TTT TGT GTA ACC-30, reverseprimer 50-AGA GTG ATG ACC ATC CTT TTC-30; forTrkB, forward primer 50-GTT TCA TAA GAT CCC ACTGGA TGG-30, reverse primer 50-TGC TGC TTA GCTGCC TGA GAG TTA-30; and for b-actin, forward primer50-TGA GAC CTT CAA CAC CCC AG-30, reverse primer50-GCC ATC TCT TGC TCG AAG TC-30. PCR productswere separated on a 1.5% agarose and photographed.
2.4. Western blotting
MM cells were harvested and washed three times withphosphate-buffered saline and resuspended in NP40 lysisbuffer (50 mmol/L Tris-HCl, pH 7.4; 150 mmol/L NaCl;1% NP40; 5 mmol/L EDTA; 5 mmol/L NaF; 2 mmol/L so-dium vanadate; 1 mmol/L phenylmethylsulfonyl fluoride;5 mg/mL leupeptin; and 5 mg/mL aprotinin) and extractedfor 20 min on ice. The protein content was quantitated byBio-Rad protein assay. After thawing and boiling for 5 minin Laemmli buffer, 30–100 mg protein extracts were sepa-rated by means of sodium dodecyl sulfate–polyacrylamidegel electrophoresis and were transferred onto nitrocellulosemembranes. After blocking with 5% nonfat milk (or 1% bo-vine serum albumin for phospho-specific antibodies), mem-branes were incubated overnight at 4�C with respectiveprimary antibody. After washing, membranes were incubatedwith HRP-conjugated secondary antibodies at room temper-ature for 1 hour. Blots were then developed using ECL.
2.5. Enzyme-linked immunosorbent assay
Cell were seeded onto six-well plates at a concentrationof 1 � 106 cells per well in RPMI 1640 with 1% FBS. After48 hours, the supernatants were collected. Aliquots of 100mL were removed, and BDNF concentrations were ana-lyzed using a commercial enzyme-linked immunosorbentassay kit (R&D Systems, Minneapolis, MN). Each wellwas analyzed in triplicate.
2.6. Immunohistochemistry for BDNF and TrkB
Bone marrow biopsies were obtained from patients withdiagnosis of MM. Bone marrow specimens were fixed

14 Y. Hu et al. / Cancer Genetics and Cytogenetics 169 (2006) 12–20
in paraformaldehyde, decalcified with EDTA, and embed-ded in paraffin. Serial sections (4 mm thick) of each samplewere processed immunohistochemically for the expressionof BDNF and its receptors with rabbit polyclonal anti-human BDNF (1:100), anti-human TrkB (1:500), andanti-human P75NTR antibodies (1:200). Controls forimmunostaining using nonimmune rabbit IgG in substitu-tion for the specific first antibodies were consistentlynegative.
Immunohistochemical staining was performed using a la-beled streptavidin–biotin peroxidase method (S-P kit;Zhongshan Biotech, Beijing, China). Briefly, tissue sectionswere deparaffinized in xylene and rehydrated in a gradedethanol series. The sections were incubated with 1%
H2O2 diluted in methanol for 20 min. Slides were pre-treated to promote antigen retrieval in a microwave ovenat 650 W for 10 min in 10 mmol/L sodium citrate, pH6.0. The primary antibodies were applied overnight at4�C. Subsequent steps were performed according to themanufacturer’s instructions. 3,30-diaminobenzidine wasused as chromogen, and sections were counterstained withhematoxylin. The degree of expression in tumor cells wasjudged at 400� magnification as 41 (very intensely posi-tive), 31 (moderately intensely positive), 21 (moderate),11 (faint), or 0 (completely negative) throughout the sam-ple. Samples were judged by two independent reviewers.
All patients had given written informed consent prior tothe sampling procedure.
Fig. 1. Expression of BDNF in myeloma cells. (A) BDNF transcripts (593 bp) in human myeloma cell lines and in 12 sorted myeloma bone marrow samples
(MM 1–12) were analyzed with RT-PCR. Peripheral blood B lymphocytes from healthy volunteers served as controls (PBB-L 1–3).b-actin (312 bp) was used
as a control for RNA integrity. C, no template PCR control, M, marker. (B) BDNF protein expression was revealed with Western blot analysis. Whole-cell
lysates were immunoblotted using a rabbit anti-BDNF antibody. Homogenate of rat brain were used as positive control for the expression of BDNF. (C)
Secretion of BDNF was determined in supernatants of myeloma cell lines with ELISA after 48 hours of culture. Data represent means for at least triplicate
samples; error bars indicate standard deviation. (D) Representative pattern of immunohistochemical staining of bone marrow section obtained at presentation
from MM and control patient for BDNF. Immunohistochemical localization was performed with the anti-BDNF antibody. Note the strong expression of
BDNF in the bone marrow of MM patient (1) compared to the faint BDNF signal observed in occasional metamyelocytes (open arrows) and in scattered
megakaryocytes (solid arrows) in the marrow section of control (2) (400� magnification).
15Y. Hu et al. / Cancer Genetics and Cytogenetics 169 (2006) 12–20
2.7. Cell survival assay
For cell viability assay, MM cells were plated in RPMI1640 supplemented with 1% FBS at a density of 1 � 105/mL in 96-well plates. Various concentrations of BDNFwere added to the medium and were replenished every 2days. After 4–8 days incubation, cell viability was deter-mined using a trypan blue dye-exclusion method.
2.8. Thymidine incorporation assay
Cell growth was determined by [3H]thymidine incorpo-ration. Briefly, cells were starved for 3 hours prior to stim-ulation with different concentrations of BDNF in mediumwith 1% FBS and cultured in starvation medium for 72hours. During the last 6 hours of incubation before har-vesting, 0.5 mCi [3H]thymidine was added to each well,and the radioactive labeling was monitored by Direct Beta
Counter (Matrix 9600; Packard Instruments, Meriden,CT).
2.9. Transwell migration assay
Cell migration was assayed using Transwell (Invitrogen)with 6.5-mm-diameter polycarbonate filters (pore size8.0 mm), as described previously [25]. Briefly, the lowersurface of the filter was coated with 10 mg of gelatin. BDNF(12.5–200 ng/mL) with or without the TrkB inhibitorK252a was added to the lower chamber. RPMI 8226 cellswere serum starved overnight and then resuspended at a finalconcentration of 1� 106 cells/mL in RPMI 1640 containing1% FBS. Next, 100 mL of the cell suspension was loadedinto each of the upper chambers. The chamber was incu-bated at 37�C for 6 hours. Cells that migrated into the lowercompartment were counted using a Coulter counter (modelZBII; Coulter Electronics, Bedfordshire, UK).
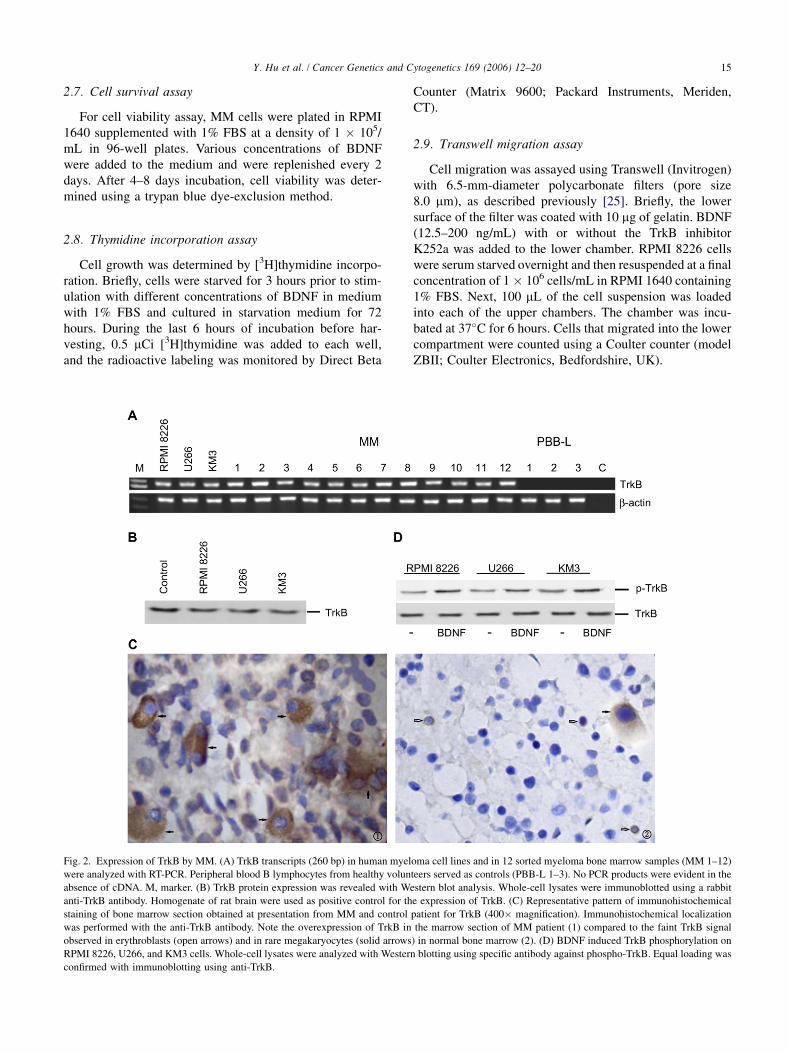
Fig. 2. Expression of TrkB by MM. (A) TrkB transcripts (260 bp) in human myeloma cell lines and in 12 sorted myeloma bone marrow samples (MM 1–12)
were analyzed with RT-PCR. Peripheral blood B lymphocytes from healthy volunteers served as controls (PBB-L 1–3). No PCR products were evident in the
absence of cDNA. M, marker. (B) TrkB protein expression was revealed with Western blot analysis. Whole-cell lysates were immunoblotted using a rabbit
anti-TrkB antibody. Homogenate of rat brain were used as positive control for the expression of TrkB. (C) Representative pattern of immunohistochemical
staining of bone marrow section obtained at presentation from MM and control patient for TrkB (400� magnification). Immunohistochemical localization
was performed with the anti-TrkB antibody. Note the overexpression of TrkB in the marrow section of MM patient (1) compared to the faint TrkB signal
observed in erythroblasts (open arrows) and in rare megakaryocytes (solid arrows) in normal bone marrow (2). (D) BDNF induced TrkB phosphorylation on
RPMI 8226, U266, and KM3 cells. Whole-cell lysates were analyzed with Western blotting using specific antibody against phospho-TrkB. Equal loading was
confirmed with immunoblotting using anti-TrkB.
16 Y. Hu et al. / Cancer Genetics and Cytogenetics 169 (2006) 12–20
2.10. Statistical analysis
Statistical significance of difference observed in BDNF-treated versus control cultures was determined by means ofan unpaired Student’s t-test. The minimal level of signifi-cance was P ! 0.05.
3. Results
3.1. Expression and secretion of BDNF by MM
The expression of BDNF transcripts was first determinedwith RT-PCR in marrow myeloma cells from 12 patientsand HMCLs (RPMI 8226, U266, and KM3). Peripheralblood B lymphocytes from healthy individuals (n 5 3)served as controls. All tested MM cells showed the pres-ence of BDNF mRNA. In contrast, BDNF mRNA was notdetected in normal peripheral blood B lymphocytes(Fig. 1A). With Western blot analysis, we confirmed thatHMCLs expressed BDNF protein (Fig. 1B), identified asa band of ~14 kDa (mature BDNF); a band of 35 kDa(uncleaved pro-BDNF) was also observed. BDNF proteinsecretion by different myeloma cell lines (RPMI 8226,U266, and KM3) was examined by enzyme-linked immu-nosorbent assay (ELISA) on the cell culture supernatant(Fig. 1C); median BDNF concentrations were 14.73, 8.23,and 13.2 ng/mL, respectively. BDNF secretion in the mye-loma cell lines were significantly higher than in the normalhuman plasma (median 2.03 ng/mL).
The expression of BDNF by myeloma cells was furtherstudied using immunohistochemistry methods in bone mar-row biopsy samples from 16 patients with MM. Resultsfrom these MM patients were compared with the degreeof protein expression in bone marrow biopsy obtained at di-agnosis from nine patients with different diseases but allwith normal bone marrow morphology. In normal bonemarrow, the detection of BDNF expression in hematopoi-etic elements was uncommon. Under immunohistochemicalanalysis, BDNF protein expression was not observed inerythroblasts, myeloblasts, or plasma cells. A faint BDNFsignal was observed in occasional metamyelocytes and inscattered megakaryocytes (Fig. 1D, right panel). BDNF ex-pression in MM specimens was detected in a diffuse cyto-plasmic pattern in malignant plasma cells with varied signalintensity. In bone marrow specimens from 12 of 16 patientswith MM, cytoplasmic expression of BDNF was detectedin malignant plasma cells, ranging from faint (11) to in-tensely positive (14); no signal or only a weakly positivesignal was detected in erythroblasts and myeloblasts(Fig. 1D, left panel).
3.2. TrkB is expressed and activatedby BDNF in MM cells
Given that BDNF is a growth and chemotactic factor forTrkB-expressing neurons and neurogenic malignancies, itmight also provide autocrine support for TrkB-expressing
MM cells. We therefore further evaluated the expressionof both BDNF receptors, TrkB and P75NTR, in HMCLsand in primary myeloma cells. Comparable transcript andprotein expression of high-affinity BDNF receptor TrkBwere detected by RT-PCR (Fig. 2A) and Western blotting(Fig. 2B), respectively, in 3 of 3 HMCLs and in 12 sortedmyeloma cells from the bone marrow of patients with mul-tiple myeloma. TrkB was also detected immunohistochemi-cally in 15 of 16 bone marrow biopsy samples, where
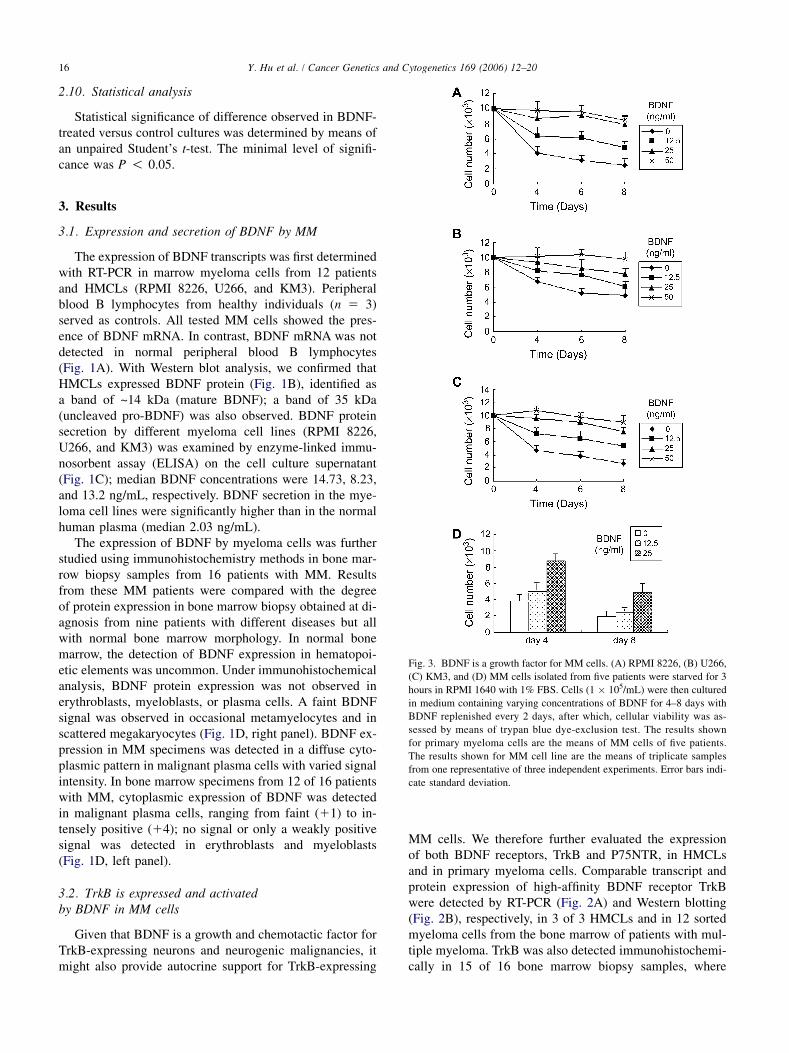
Fig. 3. BDNF is a growth factor for MM cells. (A) RPMI 8226, (B) U266,
(C) KM3, and (D) MM cells isolated from five patients were starved for 3
hours in RPMI 1640 with 1% FBS. Cells (1 � 105/mL) were then cultured
in medium containing varying concentrations of BDNF for 4–8 days with
BDNF replenished every 2 days, after which, cellular viability was as-
sessed by means of trypan blue dye-exclusion test. The results shown
for primary myeloma cells are the means of MM cells of five patients.
The results shown for MM cell line are the means of triplicate samples
from one representative of three independent experiments. Error bars indi-
cate standard deviation.
17Y. Hu et al. / Cancer Genetics and Cytogenetics 169 (2006) 12–20
malignant plasma cells, but not erythrocyte or myeloid pre-cursors, showed immunoreactivity (Fig. 2C, left panel). Afaint TrkB signal was observed in erythroblasts and in raremegakaryocytes in normal bone marrow (Fig. 2C, rightpanel). In contrast, P75NTR, the low-affinity receptor forBDNF, was not detected on HMCL or MM plasma cellswith either RT-PCR or immunohistochemistry.
To determine whether the TrkB present in MM cells isactivated, we performed Western blots using anti-pTrkBfollowed by Western blotting analysis using anti-TrkB.We found that a fraction of TrkB was tyrosine-phosphory-lated under baseline culture conditions, and phosphorylatedTrkB levels were increased following the addition of exog-enous BDNF (Fig. 2D). Together, these data suggest thatTrkB is a functional signaling receptor for BDNF in MMcells.
3.3. BDNF promotes growth and survival of MM cells
Having established that BDNF and TrkB are expressedon MM cell lines and patient MM cells, we next examinedwhether BDNF was biologically significant in terms of tu-mor cell growth. In RPMI 8226 cultures, serum deprivationresulted in a strong reduction in the number of viable cells,whereas BDNF stimulation resulted in a rescue with stablecell number (Fig. 3A). Also, U266 and KM3 cultures re-sponded to BDNF treatment. Like RPMI 8226, culturingthese MM cells in the presence of BDNF resulted in a res-cue of cell numbers (Figs. 3B,C). The prosurvival activityof BDNF can also be demonstrated using patient MM cells.As seen in Fig. 3D, BDNF prolonged the survival of pri-mary myeloma cells at concentrations as low as 25 ng/mL (P ! 0.05), confirming a potent prosurvival effect ofBDNF on MM cells.
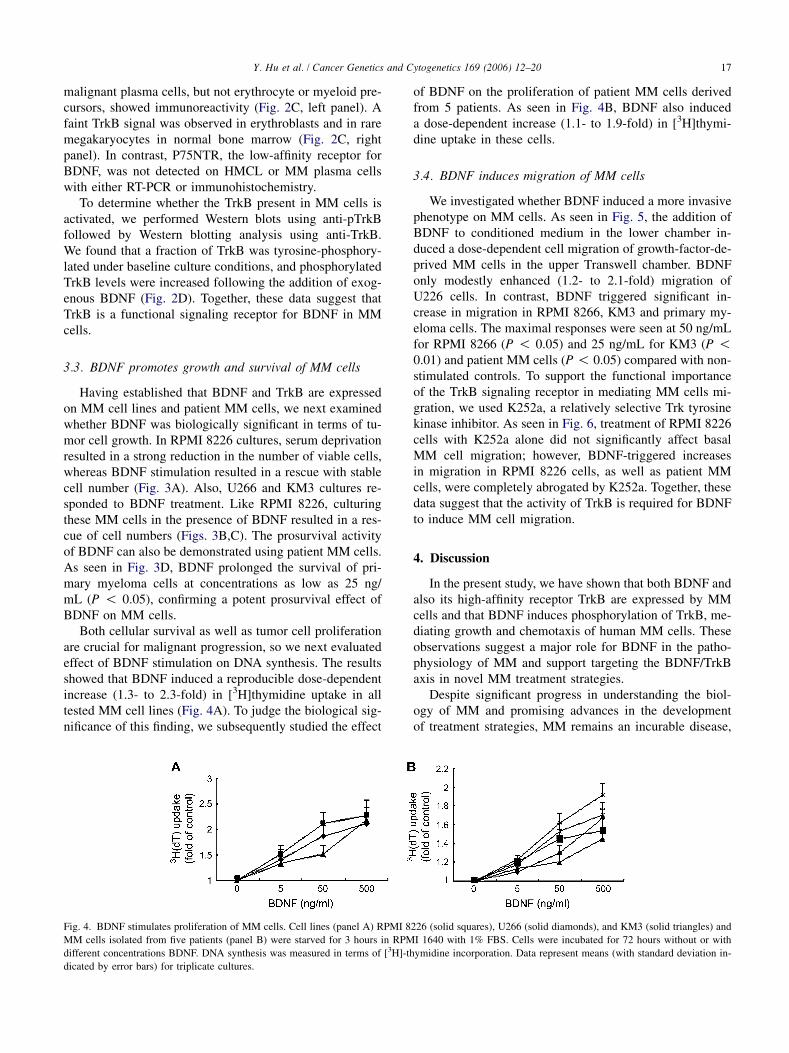
Both cellular survival as well as tumor cell proliferationare crucial for malignant progression, so we next evaluatedeffect of BDNF stimulation on DNA synthesis. The resultsshowed that BDNF induced a reproducible dose-dependentincrease (1.3- to 2.3-fold) in [3H]thymidine uptake in alltested MM cell lines (Fig. 4A). To judge the biological sig-nificance of this finding, we subsequently studied the effect
of BDNF on the proliferation of patient MM cells derivedfrom 5 patients. As seen in Fig. 4B, BDNF also induceda dose-dependent increase (1.1- to 1.9-fold) in [3H]thymi-dine uptake in these cells.
3.4. BDNF induces migration of MM cells
We investigated whether BDNF induced a more invasivephenotype on MM cells. As seen in Fig. 5, the addition ofBDNF to conditioned medium in the lower chamber in-duced a dose-dependent cell migration of growth-factor-de-prived MM cells in the upper Transwell chamber. BDNFonly modestly enhanced (1.2- to 2.1-fold) migration ofU226 cells. In contrast, BDNF triggered significant in-crease in migration in RPMI 8266, KM3 and primary my-eloma cells. The maximal responses were seen at 50 ng/mLfor RPMI 8266 (P ! 0.05) and 25 ng/mL for KM3 (P !0.01) and patient MM cells (P ! 0.05) compared with non-stimulated controls. To support the functional importanceof the TrkB signaling receptor in mediating MM cells mi-gration, we used K252a, a relatively selective Trk tyrosinekinase inhibitor. As seen in Fig. 6, treatment of RPMI 8226cells with K252a alone did not significantly affect basalMM cell migration; however, BDNF-triggered increasesin migration in RPMI 8226 cells, as well as patient MMcells, were completely abrogated by K252a. Together, thesedata suggest that the activity of TrkB is required for BDNFto induce MM cell migration.
4. Discussion
In the present study, we have shown that both BDNF andalso its high-affinity receptor TrkB are expressed by MMcells and that BDNF induces phosphorylation of TrkB, me-diating growth and chemotaxis of human MM cells. Theseobservations suggest a major role for BDNF in the patho-physiology of MM and support targeting the BDNF/TrkBaxis in novel MM treatment strategies.
Despite significant progress in understanding the biol-ogy of MM and promising advances in the developmentof treatment strategies, MM remains an incurable disease,
Fig. 4. BDNF stimulates proliferation of MM cells. Cell lines (panel A) RPMI 8226 (solid squares), U266 (solid diamonds), and KM3 (solid triangles) and
MM cells isolated from five patients (panel B) were starved for 3 hours in RPMI 1640 with 1% FBS. Cells were incubated for 72 hours without or with
different concentrations BDNF. DNA synthesis was measured in terms of [3H]-thymidine incorporation. Data represent means (with standard deviation in-
dicated by error bars) for triplicate cultures.

18 Y. Hu et al. / Cancer Genetics and Cytogenetics 169 (2006) 12–20
leading ultimately to death of all patients. For the past sev-eral years, considerable effort has been directed towardidentifying the biochemical lesions and signaling pathwaysthat contribute to either survival or growth of myelomacells. Recently, Pearse et al. [23] showed that a BDNF/TrkBaxis in MM and expression of BDNF and TrkB are likely tocontribute to malignant plasma cell survival. Our data showthat, in addition to these effects, BDNF also has directeffects on MM cells, and suggest that BDNF might be
Fig. 5. BDNF induces migration of MM cells. (A) RPMI 8226, (B) U266,
(C) KM3 and (D) primary myeloma cells starved overnight in medium
containing 1% FBS were plated on polycarbonate membranes (pore size
8 mm) in a modified Boyden chamber and exposed to BDNF (12.5–200
ng/mL) added to the lower chamber. After 6 hours of incubation, cells
in the lower chamber were harvested and counted with a Coulter counter
ZBII. Data represent means for triplicate cultures; error bars indicate stan-
dard deviation. The results shown for MM cell line are representative of
three independent experiments. *, P ! 0.05; **, P ! 0.01.
an important component in the pathophysiology of MMby promoting growth and migration of MM cells in an auto-crine mechanism.
BDNF is a member of the neurotrophin superfamily andinteracts with two classes of membrane receptor: the proto-oncogene product TrkB and a secondary receptor P75NTR[7]. In the present study, we found that BDNF is expressedand secreted by MM cells in high concentrations (<14.7ng/mL). In accordance with our data is the finding thatBDNF levels are elevated in the plasma of patients with ac-tive myeloma compared with normal control [24]. Further-more, expression of BDNF receptor TrkB is observed inMM cells.
These observations suggest an autocrine loop in MM.Expression of BDNF and its receptor TrkB has been foundin various human malignancies, of both neurogenic andnonneurogenic origin [11,12,15,26,27], consistent with anautocrine BDNF pathway. BDNF and its receptor TrkB
Fig. 6. Activation of TrkB is responsible for BDNF-induced migration.
Starved RPMI 8226 (panel A) and patient MM cells (panel B) were pre-
treated with or without K252a (500 mmol/L), plated on a polycarbonate
membranes (pore size 8 mm) in a modified Boyden chamber, and exposed
to BDNF (25 and 50 ng/mL) added to the lower chamber. After 6 hours of
incubation, cells in the lower chamber were harvested and counted with
a Coulter counter ZBII. Results are shown as means (n 5 3), with error
bars indicating standard deviation, and are representative of two separate
experiments.

19Y. Hu et al. / Cancer Genetics and Cytogenetics 169 (2006) 12–20
induce complex biological responses in target cells includ-ing growth, survival, and migration. In mice, BDNF defi-ciency results in early postnatal death, with severe defectsin the development of endothelial cells [28]. BDNF overex-pression, in both mouse and human studies, has been impli-cated in tumor growth, survival, and invasion [11–13,18–21,29]. Of note, BDNF has been found to act as a growthfactor for several malignant cell types, including medullo-blastoma [30], retinoblastoma [27], neuroblastoma [31],and hepatocellular carcinoma cells [32]. It is conceivable,therefore, that during progression of MM the coexpressionof BDNF and TrkB may lead to autonomous growth and todissemination to extramedullary sites.
Initial studies revealed that BDNF was a potent growthfactor for MM. Addition of exogenous BDNF at a concen-tration of 25 ng/mL increased survival of MM cells in cul-ture. In both MM cell lines and primary MM cells, BDNFstimulation induced a strong dose-dependent increase ofDNA synthesis. Moreover, it had potent chemotactic prop-erties. One aspect of myeloma biology that has received rel-atively little attention is that of the signals and biochemicalpathways that contribute to the important properties relat-ing to myeloma cell motility and ability to invade other tis-sues. In the initial stage of this disease, circulatingmyeloma cells specifically home to the bone marrow byprocesses wherein they must bind to the surface of bloodvessel endothelium and penetrate the endothelial membraneto reach the bone marrow compartment. Once in the bonemarrow, these cells must interact with, and likely movethrough, bone marrow stromal cells from primary to sec-ondary sites. Finally, in late-stage disease myeloma cellsagain extravasate through blood vessels and invade multipletissues to produce disseminated disease [33].
Our data showed that BDNF exerts its chemotactic prop-erties on MM cells and that BDNF is also secreted by MMcells, which could allow the hypothesis that both homing ofMM cells to the bone marrow and MM cluster cell forma-tion in the bone marrow are mediated, at least in part, byBDNF.
Having established these distinct biologic sequelae ofBDNF in MM cells, we began to delineate the responsiblesignaling pathways. BDNF mediates its activity via two re-ceptors with distinct ligand affinity and specificity: TrkBand P75NTR. We found the high-affinity BDNF receptorTrkB in HMCLs and primary myeloma cells; in contrast,P75NTR transcript and protein expression were notdetected under RT-PCR, Western blot, and immunohisto-chemical analysis, respectively. Activation of BDNF–TrkBoccurs through ligand binding, which facilitates receptordimerization and autophosphorylation of tyrosine residuesin the cytoplasmic portion. We subsequently showed, withWestern blot analysis, that BDNF triggered tyrosine-spe-cific phosphorylation of the TrkB receptor and that migra-tion induced by BDNF was completely abrogated by theTrk tyrosine kinase inhibitor K252a, showing that thisreceptor is functional in MM cells.
The interaction of MM cells with the microenvironmenthas been postulated to be critical in the pathogenesis ofMM [34]. Apart from the autocrine growth stimulation byBDNF in MM, BDNF is also expressed by a number of celltypes in the bone marrow microenvironment, including os-teoblast, megakaryocytes, endothelial cells, and adventitialreticular cells [35,36]. A paracrine function of BDNF maythus contribute additional support for myeloma cell growth.
In summary, BDNF appears to have a major contributionto the pathophysiology of MM, leading to growth and mi-gration of MM cells. These findings provide the frameworkfor novel therapeutic strategies targeting BDNF/TrkBsignaling in MM. We have further studies under way todelineate the precise mechanisms underlying the BDNF-dependent direct effects.
Acknowledgments
This work was supported by Youth Talent Foundation ofHubei Province to Y.H. We thank Dr. Jan Hou (Senond Mil-itary Medical University, Shanghai, P.R. China) for provid-ing the multiple myeloma cell line and Dr. Zhong Chen(University of Utah, Salt Lake City, Utah) for assistancein manuscript preparation.
References
[1] Klein B, Zhang XG, Lu ZY, Bataille R. Interleukin-6 in human mul-
tiple myeloma. Blood 1995;85:863–72.
[2] Kumar S, Witzig TE, Timm M, Haug J, Wellik L, Fonseca R,
Greipp PR, Rajkumar SV. Expression of VEGF and its receptors by
myeloma cells. Leukemia 2003;17:2025–31.
[3] Tai YT, Podar K, Catley L, Tseng YH, Akiyama M, Shringarpure R,
Burger R, Hideshima T, Chauhan D, Mitsiades N, Richardson P,
Munshi NC, Kahn CR, Mitsiades C, Anderson KC. Insulin-like
growth factor-1 induces adhesion and migration in human multiple
myeloma cells via activation of b1-integrin and phosphatidylinositol
30-kinase/AKT signaling. Cancer Res 2003;63:5850–8 [Erratum in:
Cancer Res 2003;63:7543].
[4] Derksen PW, de Gorter DJ, Meijer HP, Bende RJ, van Dijk M,
Lokhorst HM, Bloem AC, Spaargaren M, Pals ST. The hepatocyte
growth factor/Met pathway controls proliferation and apoptosis in
multiple myeloma. Leukemia 2003;17:764–74.
[5] Lewin GR, Barde YA. Physiology of the neurotrophins. Annu Rev
Neurosci 1996;19:289–317.
[6] Levi-Montalcini R, Dal Toso R, della Valle F, Skaper SD, Leon A.
Update of the NGF saga. J Neurol Sci 1995;130:119–27.
[7] Miller FD, Kaplan DR. Neurotrophin signalling pathways regulating
neuronal apoptosis. Cell Mol Life Sci 2001;58:1045–53.
[8] Noga O, Hanf G, Schaper C, O’Connor A, Kunkel G. The influence
of inhalative corticosteroids on circulating nerve growth factor, brain-
derived neurotrophic factor and neurotrophin-3 in allergic asthmatics.
Clin Exp Allergy 2001;31:1906–12.
[9] Hoyle GW. Neurotrophins and lung disease. Cytokine Growth Factor
Rev 2003;14:551–8.
[10] Botchkarev VA. Neurotrophins and their role in pathogenesis of
alopecia areata. J Investig Dermatol Symp Proc 2003;8:195–8.
[11] Ricci A, Greco S, Mariotta S, Felici L, Bronzetti E, Cavazzana A,
Cardillo G, Amenta F, Bisetti A, Barbolini G. Neurotrophins and neu-
rotrophin receptors in human lung cancer. Am J Respir Cell Mol Biol
2001;25:439–46.

20 Y. Hu et al. / Cancer Genetics and Cytogenetics 169 (2006) 12–20
[12] Satoh F, Mimata H, Nomura T, Fujita Y, Shin T, Sakamoto S,
Hamada Y, Nomura Y. Autocrine expression of neurotrophins and
their receptors in prostate cancer. Int J Urol 2001;8:S28–34.
[13] Feng X, Jiang H, Baik JC, Edgar C, Eide FF. BDNF dependence in
neuroblastoma. J Neurosci Res 2001;64:355–63.
[14] Sakamoto Y, Kitajima Y, Edakuni G, Sasatomi E, Mori M,
Kitahara K, Miyazaki K. Expression of Trk tyrosine kinase receptor
is a biologic marker for cell proliferation and perineural invasion of-
human pancreatic ductal adenocarcinoma. Oncol Rep 2001;8:477–84.
[15] Aoyama M, Asai K, Shishikura T, Kawamoto T, Miyachi T, Yokoi T,
Togari H, Wada Y, Kato T, Nakagawara A. Human neuroblastomas
with unfavorable biologies express high levels of brain-derived neu-
rotrophic factor mRNA and a variety of its variants. Cancer Lett
2001;164:51–60.
[16] Schneider MB, Standop J, Ulrich A, Wittel U, Friess H, Andren-
Sandberg A, Pour PM. Expression of nerve growth factors in pancre-
atic neural tissue and pancreatic cancer. J Histochem Cytochem
2001;49:1205–10.
[17] Descamps S, Toillon RA, Adriaenssens E, Pawlowski V, Cool SM,
Nurcombe V, Le Bourhis X, Boilly B, Peyrat JP, Hondermarck H.
Nerve growth factor stimulates proliferation and survival of human
breast cancer cells through two distinct signaling pathways. J Biol
Chem 2001;276:17864–70.
[18] Kowalski PJ, Paulino AF. Perineural invasion in adenoid cystic carci-
noma: its causation/promotion by brain-derived neurotrophic factor.
Hum Pathol 2002;33:933–6.
[19] Matsumoto K, Wada RK, Yamashiro JM, Kaplan DR, Thiele CJ. Ex-
pression of brain-derived neurotrophic factor and p145trkB affects
survival, differentiation, and invasiveness of human neuroblastoma
cells. Cancer Res 1995;55:1798–806.
[20] Jaboin J, Kim CJ, Kaplan DR, Thiele CJ. Brain-derived neurotrophic
factor activation of TrkB protects neuroblastoma cells from chemo-
therapy-induced apoptosis via phosphatidylinositol 30-kinase path-
way. Cancer Res 2002;62:6756–63.
[21] Jaboin J, Hong A, Kim CJ, Thiele CJ. Cisplatin-induced cytotoxicity is
blocked by brain-derived neurotrophic factor activation of TrkB signal
transduction path in neuroblastoma. Cancer Lett 2003;193:109–14.
[22] Douma S, Van Laar T, Zevenhoven J, Meuwissen R, Van Garderen E,
Peeper DS. Suppression of anoikis and induction of metastasis by the
neurotrophic receptor TrkB. Nature 2004;430:1034–9.
[23] Pearse RN, Swendeman SL, Li Y, Rafii D, Hempstead BL. A neuro-
trophin axis in myeloma: TrkB and BDNF promote tumor cell sur-
vival. Blood 2005;105:4429–36.
[24] Hu Y, Sun CY, Wang YD, Wei WN, Wu T, He WJ, Zhao S. Study on
the high expression of brain-derived neurotrophic factor in multiple
myeloma patients and its possible mechanism [In Chinese; English
abstract]. Zhongguo Shi Yan Xue Ye Xue Za Zhi [J Exp Hematol]
2005;13:104–9.
[25] Lee OH, Kim YM, Lee YM, Moon EJ, Lee DJ, Kim JH, Kim KW,
Kwon YG. Sphingosine 1-phosphate induces angiogenesis: its angio-
genic action and signaling mechanism in human umbilical vein endo-
thelial cells. Biochem Biophys Res Commun 1999;264:743–50.
[26] Eggert A, Grotzer MA, Ikegaki N, Zhao H, Cnaan A, Brodeur GM,
Evans AE. Expression of the neurotrophin receptor TrkB is associ-
ated with unfavorable outcome in Wilms’ tumor. Clin Oncol
2001;19:689–96.
[27] Wagner N, Wagner KD, Sefton M, Rodriguez-Tebar A, Grantyn R.
An abnormal response of retinoblastoma cells (Y-79) to neurotro-
phins. Invest Ophthalmol Vis Sci 2000;41:1932–9.
[28] Donovan MJ, Lin MI, Wiegn P, Ringstedt T, Kraemer R, Hahn R,
Wang S, Ibanez CF, Rafii S, Hempstead BL. Brain derived neurotro-
phic factor is an endothelial cell survival factor required for intra-
myocardial vessel stabilization. Development 2000;127:4531–40.
[29] Blanco V, Lopez Camelo J, Carri NG. Growth inhibition, morpholog-
ical differentiation and stimulation of survival in neuronal cell type
(Neuro-2a) treated with trophic molecules. Cell Biol Int 2001;25:
909–17.
[30] Chou TT, Trojanowski JQ, Lee VM. Neurotrophin signal transduc-
tion in medulloblastoma. J Neurosci Res 1997;49:522–7.
[31] Girgert R, Wittrock J, Pfister S, Schweizer P. Farnesyltransferase in-
hibitor FTI-277 prevents autocrine growth stimulation of neuroblas-
toma by BDNF. J Cancer Res Clin Oncol 2003;129:227–33.
[32] Yang ZF, Ho DW, Lam CT, Luk JM, Lum CT, Yu WC, Poon RT,
Fan ST. Identification of brain-derived neurotrophic factor as a novel
functional protein in hepatocellular carcinoma. Cancer Res 2005;65:
219–25.
[33] Qiang YW, Yao L, Tosato G, Rudikoff S. Insulin-like growth factor I
induces migration and invasion of human multiple myeloma cells.
Blood 2004;103:301–8.
[34] Hideshima T, Chauhan D, Podar K, Schlossman RL, Richardson P,
Anderson KC. Novel therapies targeting the myeloma cell and its
bone marrow microenvironment. Semin Oncol 2001;28:607–12.
[35] Cattoretti G, Schiro R, Orazi A, Soligo D, Colombo MP. Bone mar-
row stroma in humans: anti-nerve growth factor receptor antibodies
selectively stain reticular cells in vivo and in vitro. Blood 1993;81:
1726–38.
[36] Labouyrie E, Dubus P, Groppi A, Mahon FX, Ferrer J, Parrens M,
Reiffers J, de Mascarel A, Merlio JP. Expression of neurotrophins
and their receptors in human bone marrow. Am J Pathol 1999;154:
405–15.