c 3 transmission genetics chromosomes … chapter 3 transmission genetics – chromosomes,...
TRANSCRIPT

35
CHAPTER 3 TRANSMISSION GENETICS – CHROMOSOMES, RECOMBINATION AND LINKAGE In Chapter 2, we learned that the genetic material is found primarily in the nucleus of the cell and that it is organized in the form of chromosomes. In this chapter, we explore how the genetic material is inherited from generation to generation and the changes in the orga-nization of the genetic material that occur between generations. This subdiscipline of ge-netics is called Transmission Genetics or Mendelian Genetics after Gregor Mendel who discovered the laws of inheritance with his experiments on garden peas. The chapter is organized into three major sections: (1) Mendelian Genetics; (2) Transmission and inheri-tance of chromosomes; and (3) Extensions of Mendel’s laws. MENDELIAN GENETICS Mendel’s Crossing Experiments with Peas In 1865, the Austrian monk Gregor Mendel began his series of crossing experiments with garden peas. By any standard, Mendel was a very gifted scientist who appreciated the im-portance of carefully designing and interpreting his experiments. Mendel sought to under-stand the patterns of inheritance of seven different characters in peas by crossing plants with identifiably different phenotypes of a particular character, and then following the presence or absence of the character in successive generations. The traits Mendel studied were morphological attributes of the seed, pod, flower and stem. The types of crosses Mendel made are still widely used in genetics today. Unfortunately, Mendel’s discoveries were largely ignored during his lifetime, and it was not until 1900 that his work was redis-covered and its importance finally appreciated. Monohybrid Cross The first crosses Mendel made were between inbred lines of peas that were true breeding for opposite types of a character, for example, tall x dwarf, yellow x green seed, round x wrin-kled seed coat, etc. These are called monohybrid crosses because the peas were inbred lines that differed for a single character (Fig. 3.1). In the progeny from the hybrid cross, called the F1 generation, he observed that all offspring were always just one of the two alternate types, e.g. tall. Never were both types observed or were offspring of some intermediate type. Mendel then self-pollinated some F1 offspring to create an F2 generation, whose plants segre-gated for both characters found in the inbred parents (Fig. 3.1). It was at this point that Mendel was at his scientific best. Not only did he observe the character types in the offspring, but he also counted them and calculated ratios of each type. In the F2 for all seven of the characters he was studying, he observed approximately three times as many plants of one character type as the other. Furthermore, the type observed in the F1 was the one in greater number in the F2.

Transmission Genetics 36
Fig. 3.1. Example of one of Mendel’s monohybrid crosses with peas. The experiments involved three generations of plants. First, the true-breeding tall and short parental plants (P1) were crossed to form the F1 generation, which were uniformly tall plants. A single F1 generation plant was then self-pollinated to form the F2 generation. The Punnett square shows the F2 offspring resulting from self-ing the F1. There are three possible genotypes (DD, Dd and dd), but only two phenotypes (tall and short) which segregate in a 3:1 ratio. Mendel interpreted the results of his monohybrid crossing experiments to develop his first three laws of inheritance. He surmised that there must be one factor for tall and one factor for dwarf, and that these factors are somehow paired (Law 1, Table 3.1). Later this became the basis of the concept of pairs of homologous chromosomes with the gene de-termining a particular character located at the same position (i.e. locus) in each homologue. A locus can have different forms of the gene, which are called alleles. For example,

Transmission Genetics 37
Table 3.1. Mendel’s laws of segregation and independent assortment. Law 1. Each genetic character is controlled by unit factors that come in pairs in indi-
vidual organisms; these unit factors are now known as alleles.
Law 2. Paired unit factors for a given character are dominant and recessive to one another.
Law 3. Unit factors for the same character segregate independently from one another during the formation of gametes.
Law 4. Pairs of unit factors for two different traits also segregate independently of one another during the formation of gametes.
in Mendel’s peas there was one allele coding for tall plants (D) and one allele coding for short plants (d) and these alleles segregated among the offspring. Plants that have the same allele at a locus on each of the homologous chromosomes are homozygous (e.g. DD and dd), whereas those with a different allele on each homologous chromosome are heterozy-gous (e.g. Dd). Mendel further hypothesized that one unit factor (i.e. allele) is dominant to the other recessive factor (i.e. the dominant allele masks the effect of the recessive allele), based on the phenotypes he found in the F1 and F2 (Law 2, Table 3.1). For example, the allele for tall (D) is dominant and the allele for dwarf (d) is recessive. This leads to the important distinction between genotype and phenotype. The three possible genotypes are DD, Dd, and dd, al-though with D being dominant to d, there are only two phenotypes. The DD and Dd geno-types are both tall phenotypes and the dd genotype is the dwarf phenotype. A simple method to interpret the results of the monohybrid cross is through the Punnett Square, named after its inventor Reginald Punnett (Fig. 3.1). It can be seen that when a heterozygous F1 is mated to itself (or crossed to an identical F1), three genotypes (DD, Dd and dd) are found in the 1:2:1 ratio, respectively. However, only two phenotypes are found, tall and dwarf, in the ratio of 3:1, respectively. Finally, Mendel proposed that the 3:1 ratio observed in the progeny of selfed F1 plants is expected if the D and d alleles are transmitted from each parent to the offspring at random (Law 3, Table 3.1). The frequencies of the offspring genotypes are the products of the fre-quencies of the alleles transmitted from each parent. The frequency of the DD genotype is 1/2 x 1/2 = 1/4, the frequency of the heterozygous Dd class is 2(1/2 x 1/2) = 1/2, and the fre-quency of dd is 1/2 x 1/2 = 1/4. Because D is dominant to d and therefore, DD and Dd have the same phenotype, the expected proportion of tall offspring is 3/4 and the dwarf offspring is 1/4 (or a 3:1 ratio). Mendel confirmed the expectations of random segregation with another type of cross, called a test cross. Here he crossed a Dd plant with a dd plant and found that approximately half of the offspring were tall and half were dwarf. Dihybrid Cross A second type of experiment Mendel performed was the dihybrid cross, which was an extension of the monohybrid cross to two characters (Fig. 3.2). Mendel crossed a yellow and round seeded type to a green and wrinkled seeded type. The F1 offspring were all yel-low and round and the alternate types of each character were found in 3:1 ratios in the F2, as observed in monohybrid crosses involving the same traits.
Mendel also counted the two-character phenotypes and these were in a 9:3:3:1 ratio in the F2. This is the expected ratio of two-character phenotypes given that both pairs of traits segregate randomly and completely independently of one another (Law 4, Table 3.1). Under independent

Transmission Genetics 38
Fig. 3.2. Example of one of Mendel’s dihybrid crosses with peas. The Punnett square shows the off-spring resulting from selfing the F1. There are nine possible genotypes but only four phenotypes (yellow and round, yellow and wrinkled, green and round, green and wrinkled) which segregate in a 9:3:3:1 ratio. assortment, the expected frequency of any two-character phenotype is the product of the fre-quencies of each component character. Therefore, the frequency of yellow round offspring is f(yellow) x f(round) = 3/4 x 3/4 = 9/16 and the frequency of yellow wrinkled is 3/4 x 1/4 = 3/16. In this case, the genes controlling color and smoothness of seed coat reside on different chromosomes. Later in this chapter, it is shown that when genes controlling different characters reside close to each other on the same chromosome, they do not assort independently (called linkage). Mendelian Inheritance of Traits in Forest Trees Simple Mendelian inheritance of morphological traits in forest trees is rarely observed. The reason for this is that forest geneticists and tree breeders have primarily concerned themselves with the genetic improvement of economic traits, such as stem volume and wood quality, which are under the control of many genes (Chapter 6). Nevertheless, ob-servant geneticists have identified a small number of traits showing Mendelian inheritance in the progeny of controlled crosses. Some interesting examples of Mendelian inheritance

Transmission Genetics 39
of morphological traits in trees include cone color in Pinus monticola (Steinhoff, 1974), seedling foliage color in Picea abies (Langner, 1953), chlorophyll deficiencies and other morphological variants in Pinus taeda (Franklin, 1969), diameter growth in Pinus patula (Barnes et al., 1987), blister rust resistance in Pinus lambertiana (Kinloch et al., 1970), and the narrow-crown phenotype in P. abies (Lepisto, 1985). Almost certainly, many other morphological traits under simple genetic control could be identified if geneticists were to invest time in observing such traits in segregating populations. Another class of traits that show Mendelian inheritance are biochemical traits, such as terpenes, isozymes and molecular markers. There are so many of these types of traits that an entire chapter (Chapter 4) is devoted to describing the various types and their applica-tions as genetic markers. Statistical Tests for Mendelian Inheritance What if Mendel had observed only a small number of F2’s instead of several hundred or even thousands of offspring? Would he have observed the 3:1 and 9:3:3:1 ratios that led him to formulate the laws of inheritance? Genetic segregation is a random process subject to chance fluctuations. Small numbers of offspring can lead to observed ratios quite differ-ent than expected. If Mendel had looked at only ten F2 plants following the crossing of a tall and a dwarf variety of peas, he might very easily have observed five tall plants and five dwarf plants just due to chance. Since the early 1900s, geneticists have used statistical methods to test whether genetic segregation ratios conform to expected Mendelian ratios. The test most commonly used is a goodness-of-fit test to determine if observed ratios are significantly different from expected ratios. The goodness-of-fit test is performed by calculating a chi-square (χ2) statistic: χ2 = Σ (o − e)2/e Equation 3.1 where o equals the observed number and e equals the expected number in each class and the summation is over all offspring classes. The larger the deviation of the observed num-ber from the expected number, the larger the magnitude of χ2. A chi-square table (e.g. Ta-ble A5, Snedecor and Cochran, 1967) must then be consulted to determine the statistical probability of the calculated value. The probability depends on the degrees of freedom (df) in the data, which when testing Mendelian segregation ratios is just the number of off-spring classes minus one. The convention is to reject the hypothesis only if the probability of the calculated χ2 is 0.05 or less. Two examples of Mendelian segregation of traits in Pinus are described in Box 3.1, along with the calculation of the χ2 statistic. TRANSMISSION AND INHERITANCE OF CHROMOSOMES In Chapter 2, we learned that in the nucleus of every cell of a diploid (2N) organism there are two sets of chromosomes, known as homologous pairs. At the time of fertilization, one set of homologues is contributed by the egg gamete (1N) and the other set by the pollen gamete (1N). The union of these two gametes forms the diploid zygote. This single-celled zygote then undergoes cell division and eventually grows into a fully developed organism. Trees grow from a single cell to very large, multi-cellular organisms by cell division in the meristematic regions of the tree. In order for every cell to have the same composition of chromosomes, there must be a mechanism whereby chromosomes are faithfully replicated during growth. This mechanism occurs through the process of mitosis. A process must also exist whereby during the formation of gametes, each gamete receives a complete haploid

Transmission Genetics 40
set of chromosomes. This process is called meiosis. Mitosis and Cell Division Mitosis is the process of nuclear division whereby replication of chromosomes is followed by separation of the replicated products such that each new daughter nucleus receives a copy of the chromosomes found in the mother cell. Before describing the individual steps in mitosis, it is important to review the cell cycle (Fig. 3.3). Cells spend most of the time in a non-dividing phase called interphase. Interphase itself has three phases: G1, S and G2. During interphase, the chromosomes consist of extended filaments that are not visible un-der a microscope. It is during this phase that DNA is transcribed and subsequently trans-lated into gene products that regulate and mediate cell function and growth. The S phase is particularly important because this is when DNA replication occurs.
Box 3.1. Statistical tests for Mendelian segregation of morphological traits in Pinus. Inheritance of cone color in Pinus monticola Steinhoff (1974) performed a series of crossing experiments to study the inheri-tance of cone color in Pinus monticola. Two color phenotypes were observed, purple and green, and Steinhoff hypothesized that cone color was controlled by a single gene, and that the allele for purple was dominant (P) and the allele for green was recessive (p). Steinhoff crossed two heterozygous trees (Pp x Pp) and expected to observe a 3:1 ratio of purple to green cones in the offspring. He observed a total of 109 offspring; 83 progeny with purple cones and 26 offspring with green cones.
Phenotype
Geno-type(s)
Expected ratio
Observed (o)
Expected (e) o-e (o-e)2/e
Purple PP, Pp 3/4 83 81.75 1.25 0.02 Green pp 1/4 26 27.25 -1.25 0.06 Total 109 109.00 0.08
Steinhoff then performed a χ2 goodness-of-fit test to determine if the observed number of offspring conformed to the expected number based on Mendelian ratios. The expected numbers were calculated by multiplying the total observations by the expected frequency in each class. Therefore, the expected number of offspring with purple cones is 109 x 0.75 = 81.75 and the expected number with green cones is 109 x 0.25 = 27.25. The χ2 is calculated as χ2 = (83 - 81.75)2/81.75 + (26 - 27.25)2/27.25 = 0.08. The probability of χ2 = 0.08, with df = 1, is greater than P = 0.75 (Table A5, Snedecor and Cochran, 1967). This means that if the true segregation ratio of purple to green cones is 3:1, then 75% of the samples of 109 offspring would give deviations of the magnitude observed or greater. A goodness-of-fit χ2 that occurs 75% of the time by chance is certainly not unreasonable, so it was declared that the observed segrega-tion ratio supports single-locus inheritance with purple being dominant to green. (Box 3.1 continued on next page)

Transmission Genetics 41
The first step in mitosis is prophase (Fig. 3.4). The beginning of this stage is marked by the condensation (i.e. shortening and thickening) of chromosomes to form visibly distinct, thin threads within the nucleus. Each chromosome is longitudinally split into duplicates called sister chromatids, which are the products of replication during the S phase of inter-phase. The sister chromatids share an attachment point called the centromere. At the end of prophase, the nuclear membrane breaks down and an oval-shaped spindle begins to form, with the poles at opposite ends of the cell and strands attached at the centromere.
The next step is metaphase, when the chromosomes move toward the center of the cell until all centromeres line up on a central plane equidistant from the spindle poles. Now the chromatids are positioned to migrate to their respective poles. At anaphase, the sister chromatids of each chromosome separate at the centromere and begin migrating toward opposite poles of the spindle. Now each sister chromatid is regarded as a separate chromo-some. When this stage is completed, the chromosomes are clustered in two groups at the opposite poles of the spindle, each group containing the same number of chromosomes that was present in the interphase nucleus of the mother cell. Finally, at telophase nuclear divi-
Box 3.1. Statistical tests for Mendelian segregation of morphological traits in Pinus. (Continued from previous page) Inheritance of cotyledon number and lethality in Pinus taeda In this example, the joint segregation of two seedling characters (normal versus bright green hypocotyl and normal versus cotyledon-stage lethal) was tested (Franklin, 1969). Franklin hypothesized that the two characters were controlled by two different loci and that there were dominant and recessive alleles at each locus. After crossing two doubly-heterozygous trees, the expected two-locus phenotypic ratios in the off-spring were 9:3:3:1.
Phenotype
Genotype(s)
Expected ratio
Observed (o)
Expected (e)
o-e
(o-e)2/e
Normal hypocotyls, non-lethal
GGLL, GgLL, GGLl, GgLl
9/16
54
60.19
-6.19
0.64 Normal hypocotyls, lethal
GGll, Ggll
3/16
22
20.06
1.94
0.19 Green hypocotyl, non-lethal
ggLL, ggLl
3/16
22
20.06
1.94
0.19 Green hypocotyl, lethal
ggll
1/16
9
6.69
2.31
0.80 Total 107 107.00 1.82
Franklin tested this hypothesis by determining the number of offspring ex-pected in each class and calculating the χ2 goodness-of-fit statistic. The χ2 = 1.82 (df = 3) is much less than the critical value at the 5% probability level (χ2 = 7.815) (Table A5, Snedecor and Cochran, 1967), supporting Mendelian inheri-tance of two independent characters, each controlled by a single locus with domi-nant and recessive alleles.

Transmission Genetics 42
sion is completed. The spindle disappears, a nuclear membrane forms around each cluster of chromosomes, and the chromosomes begin to revert to the extended, diffuse state. Cell division is completed by the process of cytokinesis, which divides the contents of the cy-toplasm to the daughter cells. The middle lamella, which eventually becomes part of the cell wall, is formed to divide the two daughter cells. The process of mitosis is repeated continuously in meristematically active regions of the plant. In fact, all cells in a tree origi-nated from the single-celled zygote. Meiosis and Sexual Reproduction Meiosis occurs only in gamete-forming cells of the tree, specifically in the nuclei of the egg and pollen mother cells, and is fundamental to inheritance (Box 3.2). Meiosis differs from mitosis in that two successive divisions (I and II) occur with only one replication of chromosomes, such that four haploid nuclei result. The same stages as in mitosis are used to describe meiosis (Fig. 3.5). Interphase I is essentially the same as interphase in mitosis when DNA replication occurs to double the amount of DNA. Like mitosis, chromosomes shorten and thicken in prophase I, such that sister chromatids are visible. Unlike mitosis, however, pairs of homologous chromosomes pair up (synapse) point for point at this stage. Each pair of chromosomes is called a bivalent (or tetrad). During synapsis,
Fig. 3.3. The cell cycle. DNA synthesis occurs during interphase. The mitotic phase is relatively short when replicated chromosomes divide to form two daughter nuclei and daughter cells, each with a complete complement of chromosomes. the chromatids are intimately intertwined and exchanges of DNA occur (crossing over) between sister and non-sister chromatids (i.e. between chromatids of the two chromo-

Transmission Genetics 43
somes). Visible evidence of crossovers are characteristic x-shaped configurations called chiasmata. Crossing over between non-sister chromatids is very important because it enables re-combination of genes on different chromosomes of a homologous pair. Therefore, crossing over breaks linkages among loci on the same chromosome and is an important mechanism for maintaining genetic variation (Box 3.2). In metaphase I, the tetrads line up on the central plane; the centromeres of each chro-mosome in a homologous pair tend to be directed at opposite poles. Homologous pairs separate at anaphase I, with one whole chromosome of each pair moving to each pole. This is called the reduction division and ensures that each daughter nucleus has a haploid set of chromosomes. Further, the chromosomes of each pair move randomly to the poles without regard to their parental origin. This is the basis of the random segregation and independent assortment of Mendelian inheritance (Table 3.1).
Fig. 3.4. Mitosis is the process whereby replicated chromosomes condense and the nuclei divide to form two new nuclei. The different steps in mitosis are illustrated here for a hypothetical cell having a single pair of homologous chromosomes (2N = 2). Note: Individual DNA strands are not actually visible during interphase, but are shown here to track the chromosome pair of interest.

Transmission Genetics 44
Fig. 3.5. Meiosis is the process of two successive divisions that occur during sexual reproduction to produce four haploid gametes from a single pollen or egg mother (diploid) cell. The series of steps in meiosis are illustrated here for a hypothetical cell with a single pair of homologous chromosomes (2N = 2). There is only a single bivalent or tetrad showing the pairing of homologous chromosomes at metaphase I. Note: Individual DNA strands are not actually visible during interphase, but are shown here to track the chromosome pair of interest. Interphase II and prophase II together constitute a brief period between the first and second divisions when chromosomes may or may not become diffuse and reform. By the beginning of the second division, the chromosomes are identical with those present at the end of the first division. In metaphase II, the haploid set of chromosomes in each nucleus lines up on the central plane, generally oriented at right angles to the first division. Sister chromatids move to opposite poles in anaphase II. In telophase II, the haploid number of daughter chromosomes arrives at the poles, chromosomes revert to their diffuse nature, nuclear membranes appear and cell walls form that separate the four haploid daughter cells, called gametes. Female gametes are called eggs; male gametes are pollen.

Transmission Genetics 45
EXTENSIONS TO MENDEL’S LAWS Mendel’s laws elegantly described the inheritance of a variety of characters in peas and provided the fundamental framework for the laws of inheritance for all characters in eu-karyotic organisms. As with most things in biology, however, there are exceptions to the basic rules. In this section, we describe these exceptions and show how all are merely ex-tensions to Mendel’s laws. Partial Dominance Mendel’s experiments established that at every locus there are two alleles, one dominant and one recessive, and that the ratio of dominant to recessive phenotypes in an F2 is 3:1 when one locus is considered. Subsequent to Mendel’s experiments with peas, the inheri-tance of characters was studied in crosses of many different organisms, and it was found that F2’s do not always segregate in a 3:1 ratio. One explanation for departure from the 3:1 ratio is incomplete or partial dominance. Consider the example of a gene controlling needle color in P. abies in a study con-ducted by Langner (1953). In P. abies there is a mutant phenotype, known as the aurea-form, that produces white needles in seedlings homozygous gg for the aurea allele. Trees heterozygous (Gg) for the aurea allele and the wild-type allele have light-green or golden needles, whereas wild-type trees (GG) have normal dark green needles. In this case, it ap- pears that the wild-type allele is not dominant to the aurea allele or else Gg trees would have normal green needles. Instead they have a golden needle color that is intermediate between green and white, a so-called blended expression. Langner established the inheritance of the locus-controlling needle color in P. abies by making three different crosses and calculating ratios of needle color phenotypes. The three crosses he made were GG x GG, GG x Gg, and Gg x Gg. As expected, he observed all green needle progeny in the GG x GG cross. In the GG x Gg cross and the Gg x Gg cross, however, he observed 1:1 and 1:2:1 ratios of green to golden and green to golden to white needles, respectively (Fig. 3.6). These phenotypic ratios are consistent only with partial dominance and not complete dominance.
Box 3.2. Significance of meiosis to heredity. 1. Meiosis ensures that each gamete has a complete haploid set of chromosomes. 2. Meiosis results in random assortment of paternal and maternal chromosomes in gam-
etes. Therefore, for each pair of homologous chromosomes, half of the gametes re-ceive the paternal chromosome of the pair and half receive the maternal chromosome.
3. Meiosis results in independent assortment of chromosomes from different homolo-
gous pairs. This means that chromosomes originating from the paternal and maternal parents randomly assort to gametes such that one gamete may receive a mix of chro-mosomes of paternal and maternal origin. Therefore, genes on non-homologous chromosomes segregate independently.
4. Through crossing over, alleles found at different loci on the same chromosome can
recombine, further enhancing genetic diversity in offspring of sexually reproducing trees and other organisms.

Transmission Genetics 46
Fig. 3.6. The partial dominance of needle color in Picea abies (Langner, 1953). Trees homozy-gous for the wild-type allele G are green and trees homozygous for the mutant aurea allele g are white. However, heterozygous trees (Gg) are not green as if G were dominant to g, but rather they are an intermediate color of light green or golden: (a) Crossing GG x Gg results in an ex-pected 1:1 ratio of green:golden progeny; and (b) Crossing two heterozygotes results in an ex-pected 1:2:1 ratio of normal:golden:white foliage in the progeny. Codominance In the previous example, which described the partial dominant inheritance of the needle color locus in P. abies, the wild-type (G) and aurea (g) alleles are said to act additively, because the phenotype of the heterozygote (Gg) is a blended result of their individual ex-pression. Alternatively, it is possible to observe a separate phenotype of each of the two alleles at a heterozygous locus, which is known as codominance. A good example of codominance occurs with allozyme genetic markers (Chapter 4). At a heterozygous al-lozyme locus, two distinct bands are observed on gels resulting from the visible and dis-tinct expression of both alleles. The importance of codominance to genetic markers is dis-cussed in more detail in Chapter 4. Epistasis Deviation from Mendelian ratios can also occur due to the interaction between alleles at different loci. This is known as epistasis. Alleles at two loci can interact in antagonistic or complementary ways to modify the expression of phenotypes. When epistasis occurs, two-locus phenotypic ratios do not conform to the standard ratios expected under independent

Transmission Genetics 47
assortment. An example of the epistatic interaction between two loci controlling flower color in pea is shown in Box 3.3; however, there are also many other types of epistatic interactions. The consequences of epistasis are more important when the inheritance of traits are controlled by many genes, as is discussed in Chapter 6. Genetic Linkage Mendel observed independent assortment among the seven characters he studied in peas, which suggests that all genes assort independently when transmitted from parent to off-spring. However, as we learned in the previous discussion about meiosis, if the chromo-some is the unit of heredity, then it would seem that all genes on a chromosome would be inherited as a unit and would not assort independently. This is known as genetic linkage, which means that there is a greater association of two or more non-allelic genes than would be expected from independent assortment. Linkage is complete if crossing over does not occur during meiosis and there is no recombination among genes on homologous chromosomes. Linkage is incomplete if crossing over and recombination occur, and the strength of the linkage is a function of the probability that a crossover will occur between two genes. This probability increases with increasing physical (i.e. map) distance between genes on a chromosome. In the early 1900s, British geneticists William Bateson and Reginald Punnett were the first to observe genetic linkage between two flower color genes in peas; however, the un-derstanding of genetic linkage was most developed by Thomas Morgan and Alfred Stur-tevant who studied the inheritance of characters in the common fruit fly, Drosophila mela-nogaster. An example of genetic linkage between allozyme loci in Pinus rigida is de-scribed in Box 3.4. The standard method for quantifying the amount of linkage between two genes is to estimate the recombination fraction (r): r = R/N Equation 3.2 where R equals the number of recombinant gametes and N is the total number of gametes in a sample of meioses. Two types of gametes can be distinguished: parental and recom-binant gametes. Before gametes can be classified, however, the linkage phase of the two genes on a chromosome must be determined. For example, for two loci with alleles A and a and B and b, respectively, coupling linkage phase is when A and B are on one homolo-gous chromosome and a and b are on the other homologous chromosome. Alternatively, repulsion linkage phase is when A and b and a and B are on opposite homologous chro-mosomes. Under coupling linkage phase, AB and ab are parental gametes and Ab and aB are recombinant gametes. Under repulsion linkage phase, the opposite is true; Ab and aB are parental gametes and AB and ab are recombinant gametes. The concepts of coupling and repulsion and the estimation of the recombination fraction are illustrated in Box 3.4; further details on estimating linkage can be found in Bailey (1961). Organelle Genome Inheritance Most of the genetic material is found in chromosomes in the nucleus of the cell (i.e. nDNA, Chapter 2). However, both chloroplasts and mitochondria have small genomes that encode genes related to their functions. The chloroplast DNA (cpDNA) and mitochondrial

Transmission Genetics 48
Box 3.3. Epistasis for flower color in pea.
Flower color in pea is a classic example demonstrating a type of epistatic interaction be-tween alleles at two loci controlling a single trait (Fig. 1). There are two loci controlling flower color, the C and P locus, each with a dominant and a recessive allele. If a doubly-heterozygous individual, CcPp, is selfed as in the normal Mendelian dihybrid cross, the ex-pected two-locus genotypes are found as shown in the Punnett Square below. However, the phenotypic ratio of colored flowers to white flowers is 9:7 instead of the 9:3:3:1 predicted by Mendel’s law in a dihybrid cross. This is because at least one dominant allele from each locus must be present to have a colored flower. The C and P alleles from the two loci do not act independently; rather, they have a dependency on one another in determining flower color.
Fig. 1. Dihybrid segregation for flower color in pea.
There is a possible model to explain the epistatic interaction between the C and P loci. Suppose the pigment producing purple flower color requires two enzymatic steps, one con-trolled by the C locus and the other by the P locus. Furthermore, suppose the dominant allele gene product is required to successfully catalyze each reaction. Then, colored flowers result only when at least one dominant allele is found at each locus, because both enzymatic reac-tions must occur to complete the biosynthesis of the colored pigment.
Transmission Genetics 49
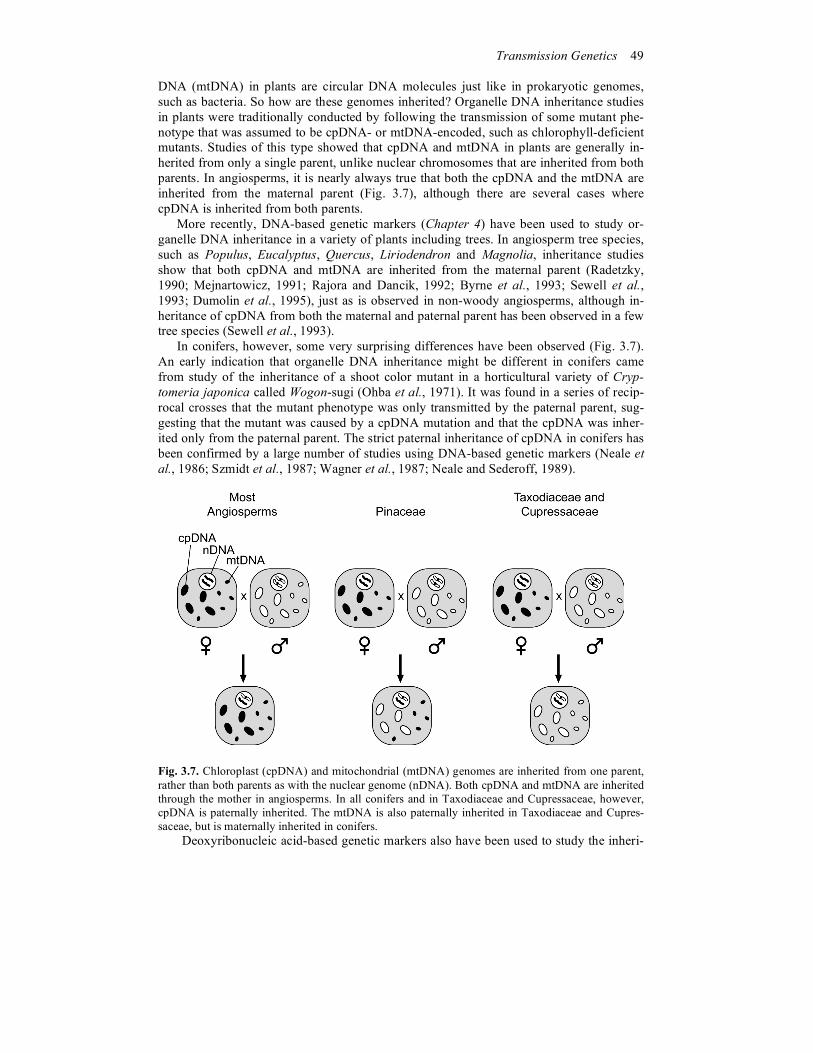
DNA (mtDNA) in plants are circular DNA molecules just like in prokaryotic genomes, such as bacteria. So how are these genomes inherited? Organelle DNA inheritance studies in plants were traditionally conducted by following the transmission of some mutant phe-notype that was assumed to be cpDNA- or mtDNA-encoded, such as chlorophyll-deficient mutants. Studies of this type showed that cpDNA and mtDNA in plants are generally in-herited from only a single parent, unlike nuclear chromosomes that are inherited from both parents. In angiosperms, it is nearly always true that both the cpDNA and the mtDNA are inherited from the maternal parent (Fig. 3.7), although there are several cases where cpDNA is inherited from both parents. More recently, DNA-based genetic markers (Chapter 4) have been used to study or-ganelle DNA inheritance in a variety of plants including trees. In angiosperm tree species, such as Populus, Eucalyptus, Quercus, Liriodendron and Magnolia, inheritance studies show that both cpDNA and mtDNA are inherited from the maternal parent (Radetzky, 1990; Mejnartowicz, 1991; Rajora and Dancik, 1992; Byrne et al., 1993; Sewell et al., 1993; Dumolin et al., 1995), just as is observed in non-woody angiosperms, although in-heritance of cpDNA from both the maternal and paternal parent has been observed in a few tree species (Sewell et al., 1993). In conifers, however, some very surprising differences have been observed (Fig. 3.7). An early indication that organelle DNA inheritance might be different in conifers came from study of the inheritance of a shoot color mutant in a horticultural variety of Cryp-tomeria japonica called Wogon-sugi (Ohba et al., 1971). It was found in a series of recip-rocal crosses that the mutant phenotype was only transmitted by the paternal parent, sug-gesting that the mutant was caused by a cpDNA mutation and that the cpDNA was inher-ited only from the paternal parent. The strict paternal inheritance of cpDNA in conifers has been confirmed by a large number of studies using DNA-based genetic markers (Neale et al., 1986; Szmidt et al., 1987; Wagner et al., 1987; Neale and Sederoff, 1989).
Fig. 3.7. Chloroplast (cpDNA) and mitochondrial (mtDNA) genomes are inherited from one parent, rather than both parents as with the nuclear genome (nDNA). Both cpDNA and mtDNA are inherited through the mother in angiosperms. In all conifers and in Taxodiaceae and Cupressaceae, however, cpDNA is paternally inherited. The mtDNA is also paternally inherited in Taxodiaceae and Cupres-saceae, but is maternally inherited in conifers. Deoxyribonucleic acid-based genetic markers also have been used to study the inheri-

Transmission Genetics 50
tance of mtDNA in conifers. In the Pinaceae, mtDNA appears to be strictly inherited from the maternal parent as in angiosperms (Neale and Sederoff, 1989; Sutton et al., 1991; DeVerno et al., 1993; David and Keathley, 1996; Fig. 3.7), although there is some evi-dence that mtDNA is occasionally inherited through the paternal parent (Wagner et al., 1991). A most surprising result, however, is that mtDNA is strictly inherited from the pa-ternal parent in Taxodiaceae and Cupressaceae (Neale et al., 1989, 1991; Kondo et al., 1998; Fig. 3.7). In no other plant or animal species has it been shown that the mtDNA is inherited strictly from the paternal parent.
Box 3.4. Estimation of genetic linkage between two allozyme loci in Pinus rigida.
Genetic linkage in forest trees can be illustrated using an example from an al-lozyme study by Guries et al. (1978). Allozymes are codominant genetic markers (Chapter 4) and the two alleles at a heterozygous locus are often designated as fast (F) and slow (S), based on their mobility difference on gels following electrophoresis. Guries et al. (1978) assayed a number of allozyme loci from seed megagametophyte tissue, which is haploid in conifers and identical to the egg gamete in genetic makeup (Chapter 4). Thus, the segregating allelic products of meiosis can be observed directly by assaying megagametophytes in a sample of seeds from a mother tree.
To illustrate the concept of genetic linkage, only two loci are considered in this example, Got-1 and Gpi-2, each heterozygous for F and S alleles in the mother (Fig. 1). If Got-1 and Gpi-2 are found on the same chromosome in the mother tree, then two possible configurations (i.e. linkage phases) of the F and S alleles at the two loci are possible. In the example below, the F alleles at each locus are on one chromosome of the mother, while the S alleles are on the homologous chromosome: the coupling link-age phase (i.e. FF/SS). The opposite configuration (FS/SF), is called the repulsion linkage phase.
Fig. 1. An example of genetic linkage for two allozyme loci.
(Box 3.4 continued on next page)

Transmission Genetics 51
SUMMARY AND CONCLUSIONS The discovery of the laws of inheritance of nuclear genetic material was made in the mid-1800s by the Austrian monk Gregor Mendel, who performed a series of crossing ex-periments in garden peas. Mendel’s monohybrid crosses between varieties of peas differ-ing in phenotype for a single character established his first three laws: (1) Unit factors come in pairs; (2) Unit factors are either dominant or recessive to one another; and (3) Unit factors segregate randomly. Mendel also studied the inheritance of pairs of factors that led to his fourth law of independent assortment. Mendel’s discoveries were ignored during his lifetime and were not rediscovered until 1900. Mendel’s laws and Charles Darwin’s theory of evolution provide the fundamental basis of the science of genetics.
Box 3.4. Estimation of genetic linkage between two allozyme loci in Pinus rigida. (Continued from previous page) Following meiosis in a doubly-heterozygous mother tree, four possible gamete types of the two-locus combinations of fast and slow alleles are expected in megagameto-phytes (FF, FS, SF, and SS). When alleles in the mother tree are in the coupling link-age phase, FF and SS gametes result if there is no crossover between the Got-1 and Gpi-2 loci; these are parental or non-crossover gametes. The FS and SF gametes result if there is a single crossover, and are called crossover or recombinant gametes.
Guries et al. (1978) assayed 160 megagametophytes for the Got-1 and Gpi-2 loci and counted the number of each of the four types. If Got-1 and Gpi-2 were located on different chromosomes and followed Mendel’s law of independent assortment, we would expect each of the four allelic combinations to occur in equal frequency, (i.e. 1:1:1:1 ratio or 40 gametes of each type). Clearly, there is a large excess of gametes in the two parental classes (FF and SS) and a corresponding deficiency in the two recom-binant classes (FS and SF).
Genotype(s)
Observed (o)
Expected (e)
(o-e)2/e
FF 90 40 62.5 FS 3 40 34.2 SF 4 40 32.4 SS 63 40 31.2 Total 160 160 142.3
Guries et al. (1978) performed a χ2 goodness-of-fit test of the 1:1:1:1 ratio and
calculated a χ2 = 142.3 (df = 1), P < 0.001, which supported the hypothesis that these loci do not assort independently and are linked. With seven recombinant gametes out of a total of 160, the recombination fraction is r = 7/160 = 0.043. The recombination fraction x 100 (called centiMorgans; cM) yields a relative measure of distance be-tween genes on a chromosome (i.e. map distance). The closer the genes are together (i.e. tighter the linkage), the smaller the cM. In this case, the linkage distance between Got-1 and Gpi-2 is 4.3 cM. Map distances range from 0 (no recombination between two genes that are extremely tightly linked) to 50 (loci that are so far apart on a chro-mosome that they assort independently).

Transmission Genetics 52
The orderly transmission of chromosomes following cell division and from generation to generation is fundamental to the inheritance of genetic material. Mitosis and meiosis are the cellular mechanisms that control the transmission of chromosomes during cell division and the formation of gametes, respectively. Mitosis occurs in vegetative plant cells during normal growth. Mitosis includes several steps where chromosomes are first replicated and then the nucleus and cell divide to form two new daughter cells each having a complete complement of chromosomes. Meiosis is similar to mitosis except that it occurs during gamete formation in pollen and egg mother cells. Two successive divisions occur following the replication of chromo-somes. The four resulting gametes each have only one homologue of a homologous pair of chromosomes. In addition, recombination can occur between non-sister chromatids, which along with mutation, are the mechanisms for creating genetic variability. In the years following the rediscovery of Mendel’s four basic laws of inheritance, sev-eral exceptions were discovered. The alleles controlling characters in peas that Mendel studied acted dominantly or recessively to one another. Incomplete or partial dominance is an exception to Mendel’s second rule, where the effects of each of the alleles are blended to produce an intermediate phenotype. Another exception to the second law is codomi-nance, where the phenotype reveals expression of both alleles, such as with allozymes. Two characters (or loci) located on the same chromosome do not always undergo inde-pendent assortment, which is a deviation from Mendel’s fourth law. This can be due to genetic linkage. The genetic distance that two loci are from each other on chromosomes is estimated by counting the number of gametes that have undergone crossing over. The pro-portion of recombinant gametes to the total is called the recombination fraction and is the standard estimator for the degree (i.e. tightness) of genetic linkage. Two-locus phenotypic ratios can also be altered by epistasis, where the phenotypic expression of an allele at one locus is modified by an allele at the other locus. One more exception to Mendel’s laws of inheritance is observed with the genetic ma-terial encoded in the chloroplast and mitochondrial genomes. These genomes are generally uniparentally inherited from either the maternal or paternal parent. In conifers, chloroplast DNA is inherited from the paternal parent and mitochondrial DNA is inherited from the maternal parent, except in the Taxodiaceae and Cupressaceae where the mitochondrial DNA is paternally inherited as well.