c6-ceramide inhibits the motility of anaplastic thyroid
TRANSCRIPT

95 © 2020 Tottori University Medical Press
C6-ceramide Inhibits the Motility of Anaplastic Thyroid Carcinoma Cells
Kazunori Fujiwara,* Hiroaki Yazama,* Ryohei Donishi,* Satoshi Koyama,* Takahiro Fukuhara,* Kazuyuki Kitatani,† Hideyuki Kataoka* and Hiromi Takeuchi**Division of Otolaryngology, Head and Neck Surgery, Department of Sensory and Motor Organs, School of Medicine, Tottori University Faculty of Medicine, Yonago 683-8504, Japan, and †Laboratory of Immunopharmacology, Faculty of Pharmaceutical Sciences, Setsunan University, Hirakata 573-0101, Japan
ABSTRACTBackground Anaplastic thyroid carcinoma (ATC) is an aggressive type of thyroid cancer, and its metastasis requires cell motility. Ceramide is involved in a variety of biological processes, including inflammation, cell signaling, cell motility, and induction of apoptosis, however has not previously been reported to inhibit the motility of ATC cells. We evaluated the effect of short chain C6-ceramide on motility of ATC cells.Methods Cell motility of 8305C thyroid carcinoma cell line treated with C6-ceramide was assessed using a transwell migration assay and a pseudopodia formation assay.Results Treatment with 10 µM C6-ceramide resulted in significantly fewer migratory cells than control treatment in a transwell migration assay (P < 0.002). In condition medium, 82.6% of C6-ceramide–treated cells formed lamellipodia. Importantly, treatment with 10 µM C6-ceramide drastically decreased the number of cells forming lamellipodia by 17.6% (P < 0.01).Conclusion Our results suggest that treatment with a low concentration of ceramide may prevent metastasis and recurrence of ATC by inhibiting cell motility. Further studies are necessary to investigate the mechanism of inhibition of cell motility by ceramide. Ceramide shows promise as a therapeutic treatment for ATC.
Key words anaplastic thyroid carcinoma; ceramide; glucosylceramide; migration; motility
Anaplastic thyroid carcinoma (ATC) is a rare and ag-gressive type of thyroid cancer that accounts for 2% of all thyroid carcinoma cases.1 It is associated with a median overall survival time of 3–10 months, with a median 1-year survival rate of 20%, and a median 10-year overall survival rate of less than 2%, despite
aggressive multimodal management.2, 3 Only 10% of patients present with local disease; 40% present with locoregional disease and 50% present with distant me-tastasis.4 ATC arises from follicular thyroid cells, and is the most dedifferentiated subtype of thyroid cancer; ATC cells do not retain any of the biological features of follicular cells. ATC usually disseminates and metasta-sizes widely, leading to increased mortality.
It was recently reported that the accumulation of multiple genetic alterations that can activate the PI3K/Akt and MAPK pathways promotes thyroid carcinoma aggressiveness and progression to poorly differentiated and undifferentiated (anaplastic) thyroid carcinoma.5 Establishment of cancer dissemination and metastasis requires cell motility.6, 7 Localized actin polymeriza-tion at the leading edge of cells pushes the membrane forward in finger-like structures known as filopodia and sheet-like structures known as lamellipodia.8, 9 Cancer cell motility governed by formation of pseudopodia is a strong candidate as a therapeutic target for cancer metastasis. Molecular mechanisms of regulation of ATC migration and invasion remain unclear.
Sphingolipids are a class of lipids that have a long-chain sphingoid base backbone. They exert a variety of biological functions such as mediating cell growth, dif-ferentiation, and apoptosis.10–12 Ceramide, an important sphingolipid that forms in response to treatment with chemotherapeutic reagents and cell-death ligands such as Fas ligand and tumor necrosis factor alpha, has been implicated in mediating apoptosis in cancer cells.13–15 Treatment with exogenous ceramides has been shown to induce apoptosis in human cells both in vitro and in vivo.14, 16 We reported that dietary glucosylceramides exhibit anti-tumor activity by inducing apoptosis of head and neck squamous cell carcinoma in vivo.17
Ceramide liposomes were shown to inhibit perito-neal metastasis in a murine xenograft model of human ovarian cancer, and knockdown of phosphatidylinositol 3-kinase C2β abrogates this effect.18 This suggests that ceramide may have potential as a therapy for metastatic disease, but its role in metastasis of other cancers re-mains unclear. We investigated the effect of ceramide in thyroid cancer by evaluating the motility of anaplastic thyroid cancer cells after treatment with C6-ceramide.
Original ArticleYonago Acta Medica 2020;63(2):95–98 doi: 10.33160/yam.2020.05.001
Corresponding author: Kazunori Fujiwara, MD, [email protected] 2020 January 23Accepted 2020 March 4Online published 2020 March 23Abbreviations: ATC, anaplastic thyroid carcinoma; DAPI, 4’,6- diamidino-2-phenylindole; PI3K, phosphatidylinositol 3 kinase

96
K. Fujiwara et al.
© 2020 Tottori University Medical Press
MATERIALS AND METHODSAntibodies and reagentsIsothiocyanate-conjugated phalloidin and anti–β-actin antibody (A5441) were purchased from Sigma (St. Louis, MO). C6-ceramide was obtained from Matreya (Pleasant Gap, PA).
Cell cultureUndifferentiated thyroid carcinoma cell line 8305C was obtained from RIKEN (Saitama, Japan) and grown in RPMI 1640 medium supplemented with 10% fetal bovine serum. Cells were maintained at < 80% conflu-ence under standard incubator conditions (humidified atmosphere, 95% air, 5% CO2, 37°C). No mycoplasma contamination was observed in any cell line.
Cell migration assay8305C cells (1 × 105 / well) were plated onto transwell chambers and treated with 10 µM C6-ceramide for 3 hours or PBS as a negative control. Migratory cells that passed through the membrane and attached to the lower surface were stained with 4’,6-diamidino-2-phenylindole (DAPI) and imaged using fluorescence microscopy. Cell migration was evaluated by counting the number of cells in 10 randomized visual fields.
Analysis of pseudopodium formation8305C cells (2.5 × 105 / well) were incubated on BD Falcon culture slides and treated with 10 µM C6-ceramide for 3 hours or PBS as a negative control, according to the method described by Kitatani et al. After incubation, cells were washed twice with PBS and fixed with 4% formaldehyde for 10 minutes. Fixed cells were treated with 0.1% Triton X-100 for 10 min-utes, followed by staining with tetramethylrhodamine isothiocyanate (TRITC)-conjugated phalloidin and DAPI for 5 minutes. Cells were imaged using confocal microscopy. Pseudopodia include filopodia and lamel-lipodia. Lamellipodia are thin and veil- like extensions at the edge of cells that contain a dynamic array of actin filaments. Cells were considered to have formed lamel-lipodia if F-actin enrichment was observed at the edge of cells. The rate of lamellipodium formation per cell was assessed by blinded quantification of fluorescence microscopy. The lamellipodium formation was evalu-ated by counting the rate of cells with lamellipodium in 10 randomized visual fields.
Statistical analysisValues are shown as the mean ± SE. Statistical com-parisons between control and C6-ceramide–treated cells were performed using the Mann–Whitney U test.
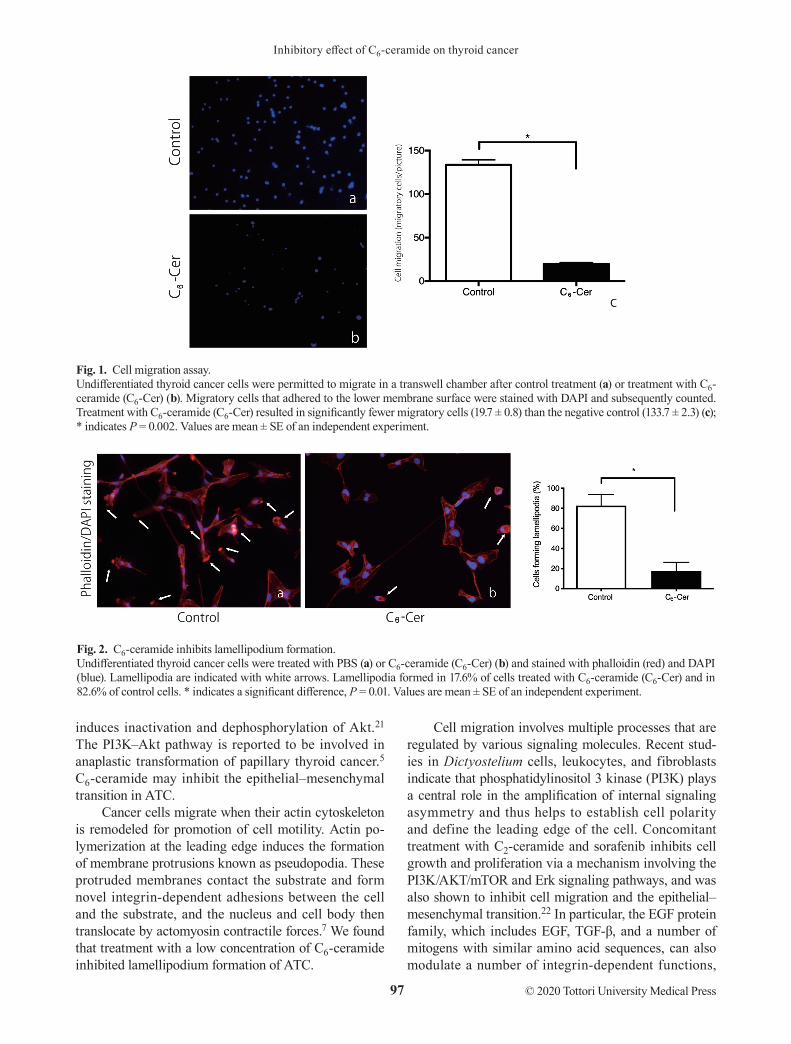
RESULTSC6-ceramide inhibits cell migrationWe evaluated the effect of C6-ceramide on 8305C cell motility using a transwell migration assay. Treatment with C6-ceramide led to significantly fewer migratory cells adhered to the lower surface (19.7 ± 0.8 vs. 133.7 ± 2.3 for the negative control; P = 0.002) (Fig. 1).
C6-ceramide inhibits lamellipodium formationThe effect of C6-ceramide on cell motility and migration was assessed by examining the formation of pseudopo-dia. Pseudopodia, such as lamellipodia and filopodia, are enriched for F-actin; thus, we stained 8305C cells with TRITC-conjugated phalloidin to visualize F-actin and DAPI to visualize nuclei in order to count the number of cells with pseudopodia. Treatment with C6-ceramide significantly inhibited lamellipodium formation; 17.6% of cells treated with C6-ceramide formed lamellipodia, whereas 82.6% of control cells formed lamellipodia (Fig. 2).
DISCUSSIONATC typically disseminates and metastasizes widely, which significantly increases mortality. The develop-ment of novel therapeutic drugs to prevent recurrence and metastasis would be very helpful for treating patients with a poor prognosis. Establishment of cancer recurrence and metastasis requires cell motility6, 7; therefore, inhibition of cell migration might prevent re-currence and metastasis. Mechanisms of cell migration in ATC are not well understood.
Ceramide is known as an intracellular mediator of apoptosis and can also exert a variety of other biological functions, including mediating inflammatory reactions, cell signaling, and cell motility. Kitatani et al. reported that ceramide inhibited cell motility in ovarian cancer.18 We evaluated the effect of ceramide on motility of ATC cells and observed that ceramide treatment significantly reduced the number of cells that could move across a membrane.
Cancer cells move within tissues during invasion and metastasis. In the course of metastasis, epithelial-derived cancer cells undergo the epithelial–mesenchy-mal transition, in which they obtain motility and lose epithelial characteristics. Intracellular adhesion is re-duced and the actin cytoskeleton is remodeled, resulting in migration through the stroma. Ceramide is known to be involved in cell motility during the epithelial–mesenchymal transition, and a decrease in C18 ceramide has been shown to increase lymph node metastasis.19
The PI3K–Akt pathway regulates cell proliferation and is also actively involved in cell motility.20 Ceramide
97
Inhibitory effect of C6-ceramide on thyroid cancer
© 2020 Tottori University Medical Press
induces inactivation and dephosphorylation of Akt.21 The PI3K–Akt pathway is reported to be involved in anaplastic transformation of papillary thyroid cancer.5 C6-ceramide may inhibit the epithelial–mesenchymal transition in ATC.
Cancer cells migrate when their actin cytoskeleton is remodeled for promotion of cell motility. Actin po-lymerization at the leading edge induces the formation of membrane protrusions known as pseudopodia. These protruded membranes contact the substrate and form novel integrin-dependent adhesions between the cell and the substrate, and the nucleus and cell body then translocate by actomyosin contractile forces.7 We found that treatment with a low concentration of C6-ceramide inhibited lamellipodium formation of ATC.
Cell migration involves multiple processes that are regulated by various signaling molecules. Recent stud-ies in Dictyostelium cells, leukocytes, and fibroblasts indicate that phosphatidylinositol 3 kinase (PI3K) plays a central role in the amplification of internal signaling asymmetry and thus helps to establish cell polarity and define the leading edge of the cell. Concomitant treatment with C2-ceramide and sorafenib inhibits cell growth and proliferation via a mechanism involving the PI3K/AKT/mTOR and Erk signaling pathways, and was also shown to inhibit cell migration and the epithelial–mesenchymal transition.22 In particular, the EGF protein family, which includes EGF, TGF-β, and a number of mitogens with similar amino acid sequences, can also modulate a number of integrin-dependent functions,
Fig. 1. Cell migration assay.Undifferentiated thyroid cancer cells were permitted to migrate in a transwell chamber after control treatment (a) or treatment with C6-ceramide (C6-Cer) (b). Migratory cells that adhered to the lower membrane surface were stained with DAPI and subsequently counted. Treatment with C6-ceramide (C6-Cer) resulted in significantly fewer migratory cells (19.7 ± 0.8) than the negative control (133.7 ± 2.3) (c); * indicates P = 0.002. Values are mean ± SE of an independent experiment.
Fig. 2. C6-ceramide inhibits lamellipodium formation.Undifferentiated thyroid cancer cells were treated with PBS (a) or C6-ceramide (C6-Cer) (b) and stained with phalloidin (red) and DAPI (blue). Lamellipodia are indicated with white arrows. Lamellipodia formed in 17.6% of cells treated with C6-ceramide (C6-Cer) and in 82.6% of control cells. * indicates a significant difference, P = 0.01. Values are mean ± SE of an independent experiment.

98
K. Fujiwara et al.
© 2020 Tottori University Medical Press
including cell adhesion, migration, and cytoskeletal organization. Ceramide was reported to impair actin polymerization through increased RhoA/Rho kinase signaling and regulate foam cell formation.23 We hy-pothesize that ceramide inhibits cytoskeletal remodeling and regulates cell migration in ATC.
In our study, we found that 10 µM C6-ceramide inhibits cell motility of ATC. However, inhibitory effect of cell motility on normal cell have not been evaluated in this study. Influence of ceramide on cell growth has not been evaluated in this study. Furthermore, C6-ceramide might inhibit cell motility for another cancers. Thus, further research is expected.
In conclusion, this study demonstrates the potential of low-concentration ceramide to prevent metastasis and recurrence of ATC. We found that ceramide inhibits formation of lamellipodia and inhibits motility of ATC cells. The relationship between ceramide and cell motil-ity should be further investigated.
Acknowledgments: This study was funded by a Grant-in-Aid for Scientific Research C of the Japan Society for the Promotion of Science (16K11233).
The authors declare no conflict of interest.
REFERENCES 1 Neff RL, Farrar WB, Kloos RT, Burman KD. Anaplastic thy-
roid cancer. Endocrinol Metab Clin North Am. 2008;37:525-38, xi. DOI: 10.1016/j.ecl.2008.02.003. PMID: 18502341
2 Smallridge RC, Copland JA. Anaplastic thyroid carci-noma: pathogenesis and emerging therapies. Clin Oncol. 2010;22:486-97. DOI: 10.1016/j.clon.2010.03.013, PMID: 20418080
3 Wendler J, Kroiss M, Gast K, Kreissl MC, Allelein S, Lichtenauer U, et al. Clinical presentation, treatment and out-come of anaplastic thyroid carcinoma: results of a multicenter study in Germany. Eur J Endocrinol. 2016;175:521-9. DOI: 10.1530/EJE-16-0574, PMID: 27926471
4 Ranganath R, Shah MA, Shah AR. Anaplastic thyroid cancer. Curr Opin Endocrinol Diabetes Obes. 2015;22:387-91. DOI: 10.1097/MED.0000000000000189, PMID: 26313900
5 Xing M. Genetic alterations in the phosphatidylinositol-3 kinase/Akt pathway in thyroid cancer. Thyroid. 2010;20:697-706. DOI: 10.1089/thy.2010.1646, PMID: 20578891
6 Wells A, Grahovac J, Wheeler S, Ma B, Lauffenburger D. Targeting tumor cell motility as a strategy against invasion and metastasis. Trends Pharmacol Sci. 2013;34:283-9. DOI: 10.1016/j.tips.2013.03.001, PMID: 23571046
7 Yamazaki D, Kurisu S, Takenawa T. Regulation of cancer cell motility through actin reorganization. Cancer Sci. 2005;96:379-86. DOI: 10.1111/j.1349-7006.2005.00062.x, PMID: 16053508
8 Lauffenburger DA, Horwitz AF. Cell migration: a physically integrated molecular process. Cell. 1996;84:359-69. DOI: 10.1016/S0092-8674(00)81280-5, PMID: 8608589
9 Nürnberg A, Kitzing T, Grosse R. Nucleating actin for inva-sion. Nat Rev Cancer. 2011;11:177-87. DOI: 10.1038/nrc3003, PMID: 21326322
10 Hannun YA. Functions of ceramide in coordinating cellular responses to stress. Science. 1996;274:1855-9. DOI: 10.1126/science.274.5294.1855, PMID: 8943189
11 Okazaki T, Bell RM, Hannun YA. Sphingomyelin turnover induced by vitamin D3 in HL-60 cells. Role in cell differen-tiation. J Biol Chem. 1989;264:19076-80. PMID: 2808413
12 Vesper H, Schmelz EM, Nikolova-Karakashian MN, Dillehay DL, Lynch DV, Merrill AH Jr. Sphingolipids in food and the emerging importance of sphingolipids to nutrition. J Nutr. 1999;129:1239-50. DOI: 10.1093/jn/129.7.1239, PMID: 10395583
13 Hannun YA, Obeid LM. Principles of bioactive lipid signal-ling: lessons from sphingolipids. Nat Rev Mol Cell Biol. 2008;9:139-50. DOI: 10.1038/nrm2329, PMID: 18216770
14 Obeid L, Linardic C, Karolak L, Hannun Y. Programmed cell death induced by ceramide. Science. 1993;259:1769-71. DOI: 10.1126/science.8456305, PMID: 8456305
15 Kolesnick RN, Krönke M. Regulation of ceramide production and apoptosis. Annu Rev Physiol. 1998;60:643-65. DOI: 10.1146/annurev.physiol.60.1.643, PMID: 9558480
16 Inamine M, Suzui M, Morioka T, Kinjo T, Kaneshiro T, Sugishita T, et al. Inhibitory effect of dietary monogluco-sylceramide 1-O-beta-glucosyl-N-2′-hydroxyarachidoyl-4,8-sphingadienine on two different categories of colon preneoplastic lesions induced by 1,2-dimethylhydrazine in F344 rats. Cancer Sci. 2005;96:876-81. DOI: 10.1111/j.1349-7006.2005.00127.x, PMID: 16367907
17 Fujiwara K, Kitatani K, Fukushima K, Yazama H, Umehara H, Kikuchi M, et al. Inhibitory effects of dietary glucosylce-ramides on squamous cell carcinoma of the head and neck in NOD/SCID mice. Int J Clin Oncol. 2011;16:133-40. DOI: 10.1007/s10147-010-0141-y, PMID: 21057846
18 Kitatani K, Usui T, Sriraman SK, Toyoshima M, Ishibashi M, Shigeta S, et al. Ceramide limits phosphatidylinositol-3-kinase C2β-controlled cell motility in ovarian cancer: potential of ceramide as a metastasis-suppressor lipid. Onco-gene. 2016;35:2801-12. DOI: 10.1038/onc.2015.330, PMID: 26364609
19 Karahatay S, Thomas K, Koybasi S, Senkal CE, ElOjeimy S, Liu X, et al. Clinical relevance of ceramide metabolism in the pathogenesis of human head and neck squamous cell carcinoma (HNSCC): attenuation of C18-ceramide in HNSCC tumors correlates with lymphovascular invasion and nodal metastasis. Cancer Lett. 2007;256:101-11. DOI: 10.1016/j.canlet.2007.06.003, PMID: 17619081
20 Xue G, Hemmings BA. PKB/Akt-dependent regulation of cell motility. JNCI Journal of the National Cancer Institute. 2013;105:393-404. DOI: 10.1093/jnci/djs648, PMID: 23355761
21 Lin C, Chen C, Lin Y. Ceramide in apoptotic signaling and anticancer therapy. Curr Med Chem. 2006;13:1609-16. DOI: 10.2174/092986706777441986, PMID: 16787207
22 Jiang S, Wang Q, Feng M, Li J, Guan Z, An D, et al. C2-ceramide enhances sorafenib-induced caspase-dependent apoptosis via PI3K/AKT/mTOR and Erk signaling pathways in HCC cells. Appl Microbiol Biotechnol. 2017;101:1535-46. DOI: 10.1007/s00253-016-7930-9, PMID: 27807662
23 Singh RK, Haka AS, Brumfield A, Grosheva I, Bhardwaj P, Chin HF, et al. Ceramide activation of RhoA/Rho kinase impairs actin polymerization during aggregated LDL catabo-lism. J Lipid Res. 2017;58:1977-87. DOI: 10.1194/jlr.M076398, PMID: 28814641