calorie restriction, stress and the ubiquitin-dependent pathway in mouse livers
TRANSCRIPT

Mechanisms of Ageing and Development
105 (1998) 273–290
Calorie restriction, stress and theubiquitin-dependent pathway in mouse livers
Mona M. Scrofano a, Fu Shang a, Thomas R. Nowell Jr. a,Xin Gong a, Donald E. Smith b, Maureen Kelliher b, Jodi Dunning b,
Casilda V. Mura c, Allen Taylor a,*a Laboratory for Nutrition and Vision Research,
Jean Mayer USDA Human Nutrition Research Center on Aging at Tufts Uni6ersity,711 Washington Street, Boston, MA 02111, USA
b Comparati6e Biology and Medicine,Jean Mayer USDA Human Nutrition Research Center on Aging at Tufts Uni6ersity,
711 Washington Street, Boston, MA 02111, USAc Centro de Estudios Cientificos de Santiago, A6e Pte Errazuriz 3132, Santiago, Chile
Received 30 March 1998; received in revised form 26 July 1998; accepted 29 July 1998
Abstract
Calorie restriction (R) is the only known method to delay the aging process and extendmean and maximal lifespan in rodents. R has been shown to delay the age-relatedaccumulation of damaged proteins and to protect organisms from various stresses which canproduce damaged proteins. Such stresses include irradiation, heat shock, and oxidative stress.The ubiquitin- and ATP-dependent proteolytic pathway (UPP) has been associated with thedegradation of abnormal and/or damaged proteins. We examined the effect of diet andoxidative stress on activities of the UPP in supernatants from livers taken from 23-month-oldEmory mice which had been exposed to an in-vivo injection of paraquat. Paraquat inducesoxidative stress by generating superoxide radicals. In livers from non-stressed animals,steady-state levels of endogenous ubiquitin conjugates, de novo conjugate formation, and E1and E2 activities were significantly lower in R animals than in control (C) animals. However,after exposure to paraquat, levels of endogenous ubiquitin conjugates were significantly
* Corresponding author. Tel.: +1 617 5563155; fax +1 617 5563344; e-mail: Taylor–[email protected]
0047-6374/98/$ - see front matter © 1998 Elsevier Science Ltd. All rights reserved.
PII S0047-6374(98)00097-9

M.M. Scrofano et al. / Mechanisms of Ageing and De6elopment 105 (1998) 273–290274
higher in R versus C animals, and de novo conjugate formation and E1 and E2 activities inR animals rose to levels which were indistinguishable from levels of these activities noted inC animals. R was associated with an increased ability to degrade b-lactoglobulin by the UPPafter an oxidative stress was imposed. Ability to degrade b-lactoglobulin by the C or R liversin non-stressed animals was not significantly different. Taken together, these data indicatethat oxidative stress in R animals is associated with enhanced levels of ubiquitin conjugatesand that this enhancement may be due to an increase in UPP activity. These data alsoindicate that the ability to form ubiquitin conjugates and the UPP system does not changewith oxidative stress in C animals. The latter is consistent with prior reports that suggeststhat older C animals may already be in a state of enhanced oxidative stress and that activitiesof the UPP provide a sensitive indicator of levels of cellular redox status. © 1998 ElsevierScience Ltd. All rights reserved.
Keywords: Dietary restriction; Oxidative stress; Ubiquitin; Liver; ATP-dependent; Paraquat
1. Introduction
Oxidative stress and the accumulation of oxidatively damaged moieties have beenpostulated to play a role in the aging process. It has been demonstrated thatoxidatively damaged proteins accumulate with age (Harman, 1956; Oliver et al.,1987; Garland et al., 1988; Stadtman, 1992; Youngman et al., 1992; Sohal et al.,1993; Agarwal and Sohal, 1994; Sohal and Weindruch, 1996; Yu, 1996), and thatthis accumulation is associated with many age-related and chronic diseases includ-ing Alzheimer’s disease (Lowe et al., 1988; Vincent and Davies, 1990; Kudo et al.,1994), Parkinson’s disease (Lowe et al., 1988), cancer (Harman, 1994), and cataract(Taylor and Davies, 1987; Taylor et al., 1991; Taylor, 1992). Organisms haveprimary and secondary defenses available to protect against such accumulation.Primary defenses include antioxidants and antioxidant enzymes, some of whichhave been shown to decline with aging (Jacques and Chylack, 1991; Halliwell et al.,1992; Ames et al., 1993; Mura et al., 1996; Gong et al., 1997). The secondarydefenses are involved in remediation or repair. One such secondary defense is theubiquitin-, ATP-dependent proteolytic pathway (UPP), involved in the identifica-tion and removal of oxidatively damaged and abnormal proteins (Ciechanover etal., 1978; Hershko et al., 1979; Ciechanover et al., 1980; Hershko et al., 1980;Rechsteiner, 1987; Murakami et al., 1990; Finley and Chau, 1991; Huang et al.,1993; Peters, 1994; Huang et al., 1995; Shang et al., 1997a,b) as well as in regulationof many critical pathways by degradation of many short-lived proteins (i.e. p53 andcyclins) (Finley and Chau, 1991; Glotzer et al., 1991; Hershko and Ciechanover,1992; Hershko et al., 1994; Scheffner et al., 1994). Consistent with a protective role,the UPP appears to be up-regulated during responses to stress (Shang et al., 1997b).
Hallmarks of the UPP (Fig. 1) are: (1) the covalent attachment of ubiquitin (an8.5-kDa protein which is highly conserved among eukaryotes) to a target proteinbefore degradation, and (2) a requirement for ATP at two steps (Hershko et al.,1983, 1984a; Hershko and Ciechanover, 1992; Hershko et al., 1994; Scheffner et al.,

M.M. Scrofano et al. / Mechanisms of Ageing and De6elopment 105 (1998) 273–290 275
1994). Ubiquitin–protein conjugate formation requires the sequential action ofthree classes of enzymes: ubiquitin-activating enzyme (E1); ubiquitin-conjugatingenzymes (E2); and, for some substrates, ubiquitin–protein ligases (E3). Both E2sand E3s play roles in substrate recognition (Scheffner et al., 1994). In the reactioncatalyzed by E1, the carboxyl-terminal glycine of ubiquitin is activated by ATP toform a high-energy thiol–ester intermediate (Hershko et al., 1984b; Finley andChau, 1991; Rechsteiner, 1991; Hershko and Ciechanover, 1992; Ciechanover andSchwartz, 1994a). Ubiquitin is then transferred to E2, and E2 then transfersubiquitin to a protein substrate or to ubiquitin–protein ligase, E3, prior tocatalyzing the formation of ubiquitin–substrate bond. During the last step in theconjugation process, an isopeptide bond is formed between the carboxyl terminusof ubiquitin and e-amino groups of lysine residues of the protein substrate.Multiple ubiquitins can be attached to lower mass substrates, leading to theformation of high molecular weight ubiquitin–protein conjugates. Following for-mation of such conjugates, the ATP-dependent 26S proteasome degrades thesubstrate moiety of the conjugate. The ubiquitins are liberated and can be utilizedin further cycles of proteolysis (Finley and Chau, 1991).
Calorie restriction (R) is the only known method to delay the aging process andextend maximal lifespan (Weindruch and Walford, 1988; Weindruch, 1991; Ma-soro, 1993; Yu, 1996). Such extension of lifespan is thought to involve prolongingthe efficacy of primary and secondary defenses (Ward and Shibatani, 1994; Tayloret al., 1995; Shibatani and Ward, 1996; Yu, 1996; Gong et al., 1997; Villarreal et al.,1997), but the data to support this hypothesis are limited. Recent work in ourlaboratory indicated that levels of endogenous ubiquitin conjugates and de novoconjugate formation may change with aging in livers of control (C) but not R mice.That work also suggested that novel relationships between aging, diet and proteol-ysis might be observed if a stress was imposed. To test this possibility, C and R
Fig. 1. The ubiquitin and ATP-dependent proteolytic pathway.

M.M. Scrofano et al. / Mechanisms of Ageing and De6elopment 105 (1998) 273–290276
23-month-old Emory mice were given a single injection of paraquat to induceoxidative stress in vivo. Livers were removed 24 h later and assayed for endogenousubiquitin conjugates, ubiquitin conjugation activity and activity of the conjugationenzymes E1 and E2(s). We found that livers from non-stressed C animals had: (1)higher steady state levels of endogenous ubiquitin conjugates; (2) enhanced abilityto form conjugates with exogenous ubiquitin; and (3) higher levels of E1 and E2activities, than in the livers from non-stressed R-fed animals. However, after micewere oxidatively stressed in vivo with paraquat, R animals had higher levels ofendogenous ubiquitin conjugates than C animals. Moreover, R livers exposed tooxidative stress showed a two-fold higher rate of ATP-dependent degradation of anexogenous radiolabeled substrate (b-lactoglobulin) as compared to oxidized Clivers, or non-stressed C and R livers.
2. Materials and methods
2.1. Materials
Trizma (Tris-base), dithiothreitol, creatine phosphate, creatine phosphokinase,ATP, apyrase, Coomassie Blue R-250, paraquat (methyl viologen) and chloramineT were obtained from Sigma (St. Louis, MO). Acrylamide, N-N %-methylene-bis-acrylamide, N,-N,-N %,N %-tetramethylenediamine, 2-mercaptoethanol, sodium dode-cyl sulfate (SDS), glycine, and protein molecular mass standards were purchasedfrom Bio-Rad (Richmond, CA). Magnesium chloride was from Fisher ScientificCompany (Fairlawn, NJ). Na[125I] and [125I]protein A were obtained from DuPont/NEN (Boston, MA). Anti-ubiquitin antibody and anti-E1 antibody wereproduced, in this laboratory, in New Zealand White rabbits by injection ofSDS-denatured ubiquitin conjugated to g-globulin or synthetic peptides of E1conjugated to ovalbumin.
2.2. Animals
Male and female Emory mice were maintained in American Association for theAccreditation of Laboratory Animal Care (AAALAC)-accredited facilities in anenvironmentally controlled atmosphere (23°C, 45% humidity with 15 air changes of100% fresh hepa-filtered air per hour, and a 12-h light, 12-h dark cycle). The micewere treated in accordance with the principles identified in the NIH Guide for theCare and Use of Animals in Research.
2.3. Diets
Mice were weaned 21–35 days after birth and housed in individual cages withfree access to water. Male and female Emory mice were fed Agway ProLab 2000chow until 2 months of age and were then placed on C or 50% R diets. Dietconstituents, feeding schedules and animal handling were as described by Taylor et

M.M. Scrofano et al. / Mechanisms of Ageing and De6elopment 105 (1998) 273–290 277
al. (1995), except for the following two modifications: (1) soybean oil was used inplace of corn oil, and (2) C mice were fed an extra 0.5 g/day of diet (which resultedin a 2% as compared to a 15% restriction). Animals were killed at 23 months of age.Because there was no observed difference between sexes, data from both sexes wascombined. In order to determine the appropriate level of paraquat to be adminis-tered, levels of reduced glutathione (GSH) were measured after they had beenexposed to 5, 10 and 20 mg/kg of paraquat. The lowest dose required to decreaseGSH levels in the liver of Emory mice was 20 mg/kg. The LD50 for paraquat inmice is 25 mg/kg (Saenghirunvattana et al., 1992). We chose doses below the LD50
level so as to elicit an oxidative stress and yet avoid killing the animal. In theseexperiments, a dose of 20 mg/kg of paraquat dissolved in 0.9% saline was given toeach mouse, i.p., 24 h prior to sacrifice. Non-stressed animals were given 0.9%saline, i.p. Food was removed from the cages 12 h before the animals weresacrificed by CO2 inhalation. Livers were immediately excized and frozen in liquidnitrogen and stored at −80°C until processing.
2.4. Determination of the endogenous le6els of ubiquitin conjugates and E1 proteinle6els
One-hundred milligrams of frozen liver was cut off and immediately homoge-nized in ice-cold 50 mM Tris–Cl, pH 7.6 (1–10 w/v) using an SDT-1810 TekmarTissumizer (Cincinnati, OH). To protect the ubiquitin conjugates from degradationby the proteasome, livers were homogenized in the following cocktail of proteaseinhibitors: 1 mM EDTA, 1 mM pepstatin, 1 mM E-64, 2 mg/ml aprotinin, 10 mMiodoacetamide, and 20 nM NEM in 50 mM Tris–Cl, pH 7.6. Supernatants weremade by centrifugation of the homogenates in a Beckman microcentrifuge at3000×g for 20 min at 4°C. Protein concentrations were determined by the Lowrymethod using BSA as a standard (Lowry, 1951). Following SDS-PAGE separationof 50 mg of liver supernatant protein in 12% gels, the proteins were transferred tonitrocellulose, probed with an a polyclonal antibody raised against SDS-denaturedubiquitin conjugated to bovine g-globulin, and followed by incubation with[125I]protein A. The ubiquitin conjugates were detected by autoradiography andquantified by image analysis.
To determine E1 protein levels, liver supernatants were separated on 6% SDS-PAGE gels, blots were probed with anti-E1 antibody and protein levels weredetected using enhanced chemiluminescence (ECL, Amersham Life Science, Buck-inghamshire, UK) by autoradiography and quantified by image analysis.
2.5. Determination of ubiquitin conjugating acti6ity
Liver supernatants were prepared, as already described, but in the absence ofprotease inhibitors. Ubiquitin conjugation activity in the liver supernatants wasquantified as the ability to catalyze the formation of conjugates between endoge-nous protein substrates and 125I-labeled ubiquitin. To enhance conjugate stabilitythe b,g-nonhydrolyzable ATP analog, 5%-adenylyl imidodiphosphate (AMP-PNP),

M.M. Scrofano et al. / Mechanisms of Ageing and De6elopment 105 (1998) 273–290278
was used in the conjugating buffer. This ATP analog supports protein ubiquitinyla-tion but does not support conjugate degradation by the 26S proteasome (Johnstonand Cohen, 1991). This assay was done in a final volume of 50 m l, containing 50mM Tris–Cl (pH 7.6), 5 mM MgCl2, 1 mM DTT, 2 mM AMP-PNP, 2 mg[125I]ubiquitin (�106 cpm), and 30 m l of liver supernatants (10 mg protein/ml). Theconcentration of [125I]ubiquitin in these assays is saturating. Following incubationat 37°C for 20 min, the reaction was stopped by addition of 50 m l of 2× Laemmlibuffer. After boiling at 100°C for 5 min, the proteins in a 20 m l mixture wereseparated by SDS-PAGE using 12% gels. To assess non-specific binding of[125I]ubiquitin to liver proteins, a parallel experiment was done in which AMP-PNPwas replaced with apyrase (12 units/ml, final concentration) (Loetscher et al., 1991)to deplete endogenous ATP. After drying, the gel was exposed to film for 2–4 days.Conjugate levels were determined by scanning the autoradiogram. The difference inthe amount of conjugates formed with and without AMP-PNP reflects the ubiquitinconjugating activities.
2.6. Determination of the acti6ities of enzymes in6ol6ed in ubiquitin conjugation
The activities of the ubiquitin-activating enzyme (E1) and ubiquitin-conjugatingenzyme (E2) were determined using their property to form thiol esters withubiquitin (Ciechanover et al., 1982; Hershko et al., 1983; Haas et al., 1995). Thisassay was done in a final volume of 50 m l containing 50 mM Tris–Cl (pH 7.6), 5mM MgCl2, 1 mM DTT, 2 mM AMP-PNP, 2 mg [125I]ubiquitin (�106 cpm), and30 m l of liver supernatant (10 mg protein/ml). The reaction was started withaddition of 30 m l of supernatant to 20 m l of [125I]ubiquitin mixture. Followingincubation at 37°C for 5 min, the reaction was stopped by the addition of either 50m l of 2× Laemmli buffer (which contains b-mercaptoethanol) or 50 m l of thiolester buffer (50 mM Tris, 4% SDS, 8 M urea, pH 6.8, and no b-mercaptoethanol).After standing at room temperature for 10 min, proteins were separated bySDS-PAGE using two separate 15% gels. The two gels (with and without b-mer-captoethanol) were exposed to film after staining and drying. Levels of thiol estersof E1s and E2s are diminished due to the reduction of the thiol ester byb-mercaptoethanol in the Laemmli buffer. The activities of E1s and E2s werequantified by determining the difference in density of the thiol ester bands on theautoradiogram which was prepared from gels run in the presence and absence ofb-mercaptoethanol. The mass of the ubiquitin thiol ester of E1 is about 120 kDa(the mass of a subunit of E1 is about 110 kDa, and the mass of ubiquitin is 8.5kDa). The masses of E2–ubiquitin thiol esters are �8.5 kDa higher than themasses of the corresponding E2s (Hershko et al., 1983).
2.7. Preparation of 125I-labeled proteins
Proteins were radiolabeled essentially as described (Jahngen et al., 1986). In afinal volume of 300 m l, 200 mg of the protein in 50 mM phosphate buffer (pH 7.6)was added to 0.6–1.0 mCi of carrier-free Na[125I] which had been adjusted to pH

M.M. Scrofano et al. / Mechanisms of Ageing and De6elopment 105 (1998) 273–290 279
7.6 with 0.2 M phosphate buffer. Fifty micrograms of chloramine T was addedas an oxidizing agent, and the reaction vial swirled for 1 min at room tempera-ture. Following termination of the iodination procedure by the addition of 100mg of sodium metabisulfite and 2 mg of sodium iodide, free 125I was removedfrom the radioiodinate protein using a Centricon 10 microconcentrator (Ami-con, Beverly, MA) as per manufacturer’s instructions.
2.8. Determination of proteolytic capabilities
One-hundred milligrams of frozen liver was cut off and immediately homoge-nized in ice-cold 50 mM Tris–Cl, pH 7.6 (1–10 w/v) using an SDT-1810 Tek-mar Tissumizer (Cincinnati, OH) in the absence of protease inhibitors. Todistinguish between ATP-dependent and ATP-independent degradation, ATPwas either supplemented or removed from the reaction mixture (Evans andWilkinson, 1985; Haas et al., 1986; Costa et al., 1991; Costa and Del Monte,1994). The reaction mixture contained 300 mg of liver supernatant protein, 30mM Tris–Cl (pH 7.6), 5 mM MgCl2, 2 mM DTT, and either an ATP-generat-ing system (2 mM ATP, 10 mM creatine phosphate, and 5 mg creatine phos-phokinase) or an ATP-depletion system (apyrase, 12 units/ml) (Loetscher et al.,1991) in a final volume of 50 m l. Degradation was initiated by addition of a125I-radiolabeled b-lactoglobulin or oxidized RNase. The sample was incubatedat 37°C for 2 h and the reaction was terminated by addition of bovine serumalbumin (BSA) and then ice-cold TCA. The sample was then placed on ice toprecipitate the TCA-insoluble proteins. These proteins were then pelleted bycentrifugation. The extent of substrate degradation was determined by theamount of TCA-soluble (acid-soluble) 125I-labeled fragments, which werecounted on a gamma counter. The degradation observed in the presence ofATP reflected the total degradation and the degradation observed in the ab-sence of ATP reflected ATP-independent degradation. The difference betweentotal degradation and ATP-independent degradation is ATP-dependent degrada-tion. These assays indicate the minimum activity of the ubiquitin-dependent sys-tem since it is rarely possible to completely exhaust the ATP prior to assay.Additionally, endogenous substrates may compete with the radiolabeled exoge-nous substrates.
2.9. Statistics
Data were evaluated by two-way ANOVA using SYSTAT (Version 5.2.1, forMacintosh, Evanston, IL). Responses between male and female livers were notsignificantly different, thus results from male and female livers were combinedfor analysis. Overall effects of diet and exposure were examined individuallyonly when diet-by-exposure interactions were not statistically significant. Wheninteractions were detected, differences between treatment means were evaluatedpost hoc using the Tukey’s HSD procedure for multiple comparisons.

M.M. Scrofano et al. / Mechanisms of Ageing and De6elopment 105 (1998) 273–290280
3. Results
3.1. The effect of oxidati6e stress and diet on endogenous ubiquitin conjugates
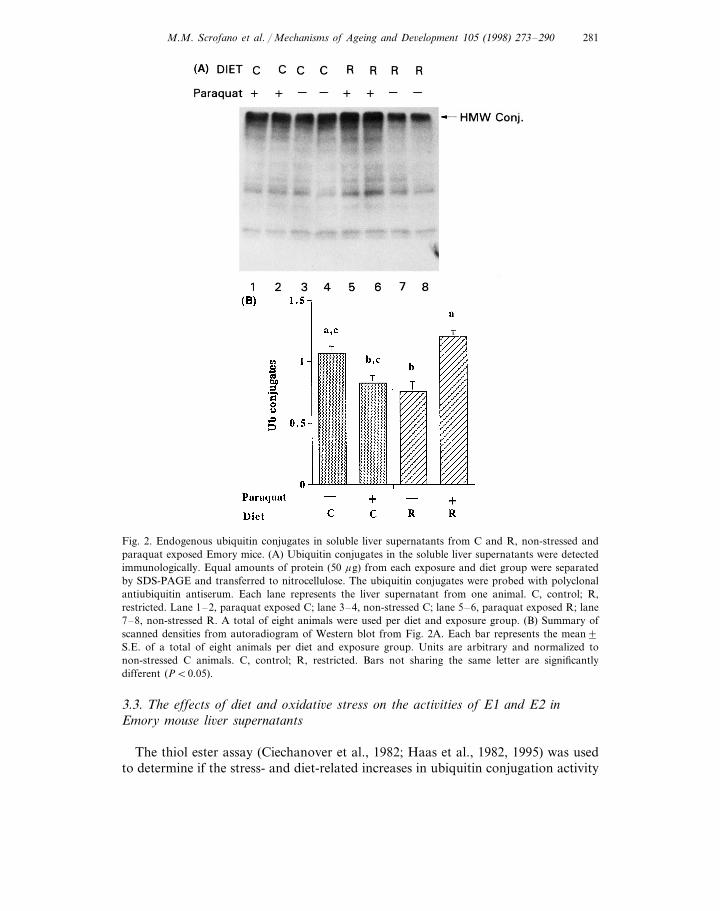
Ubiquitinylation is the first step of the ubiquitin and ATP-dependent proteolyticpathway. To determine the effect of diet and oxidative stress on the UPP, we firstmeasured the levels of endogenous ubiquitin–protein conjugates. The molecularweights of endogenous ubiquitin conjugates in the non-oxidized and oxidized liversranged from 16 kDa to greater than 200 kDa (Fig. 2A). More than 70% of theconjugates had high molecular weights (HMW) greater than 200 kDa. Withoutexposure to oxidative stress, liver supernatants from R animals had 29% lowerHMW ubiquitin protein conjugates than C animals (Fig. 2A, compare lanes 7 and8 versus lanes 3 and 4; Fig. 2B, 0.7690.08 versus 1.0790.06, P=0.01). However,in the animals which were exposed to oxidative stress, ubiquitin–protein conjugatelevels were 45% higher in R livers than in C livers (Fig. 2A, compare lanes 5 and6 versus lanes 1 and 2; Fig. 2B, 1.2090.06 versus 0.8390.06, P=0.003). Therewas a trend towards decreased liver ubiquitin–protein conjugates from C animalsexposed to oxidative stress as compared to non-stressed C animals (22%, Fig. 2B;0.8390.06 versus 1.0790.06, P=0.07). In contrast, oxidative stress in R animalswas associated with a 58% increase in ubiquitin conjugate levels as compared tonon-stressed R animals (Fig. 2A, compare lanes 5 and 6 versus lanes 7 and 8; Fig.2B, 1.2090.06 versus 0.7690.08, P=0.001).
3.2. The effect of diet and oxidati6e stress on de-no6o conjugate formation
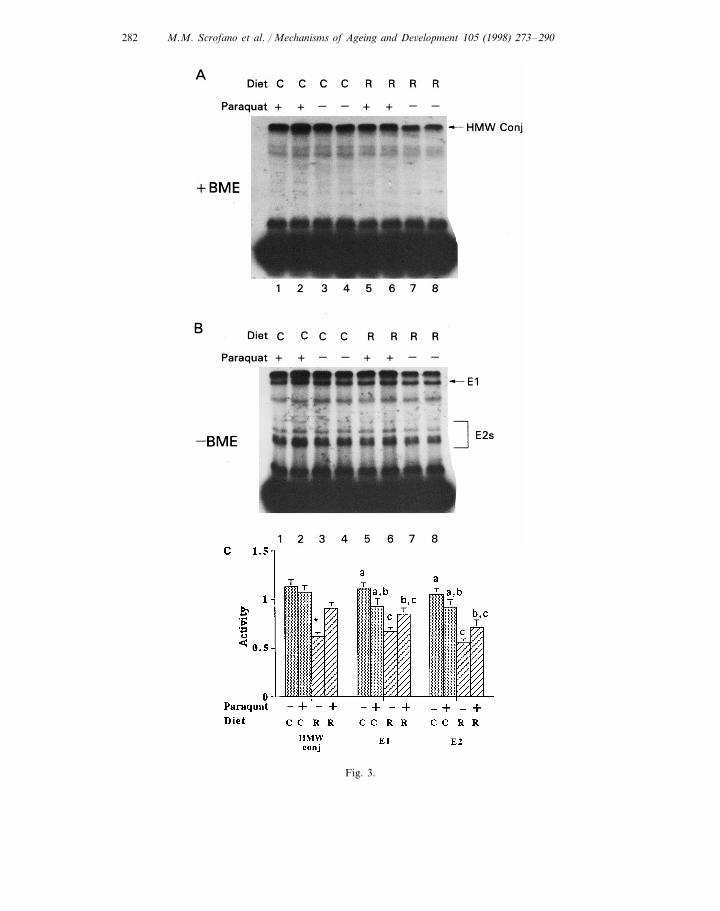
In order to determine if the stress- and/or diet-related alterations in levels ofendogenous ubiquitin conjugates were associated with altered ubiquitin conjugatingactivity, this activity was determined by incubating liver supernatants with exoge-nous 125I-labeled ubiquitin and 2 mM AMP-PNP. AMP-PNP is used because itsupports ubiquitin conjugate formation without supporting conjugate degradation.This enhances the possibility of observing ubiquitin conjugates. Fig. 3A shows thatthe liver supernatants have the ability to form ubiquitin conjugates using endoge-nous substrates and exogenous ubiquitin, with de-novo-formed conjugate molecularweights ranging from 15 to greater than 200 kDa. As was observed with endoge-nous ubiquitin conjugates, non-stressed R livers had 45% lower levels of HMWde-novo-formed ubiquitin conjugates than the non-stressed C livers (Fig. 3A,compare lanes 7 and 8 versus lanes 3 and 4; Fig. 3C, 0.6290.04 versus 1.1390.08,P=0.0002). However, after the livers were oxidatively stressed, de-novo-formedubiquitin conjugate levels in R livers increased 47% (Fig. 3A, compare lanes 5 and6 versus lanes 7 and 8; Fig. 3C, 0.9190.06 versus 0.6290.04, p=0.014). Therewas no exposure effect in the C animals. De-novo ubiquitin conjugating activities inthe liver supernatants from R animals after exposure to oxidative stress wereindistinguishable from the stressed and non-stressed C livers.
M.M. Scrofano et al. / Mechanisms of Ageing and De6elopment 105 (1998) 273–290 281
Fig. 2. Endogenous ubiquitin conjugates in soluble liver supernatants from C and R, non-stressed andparaquat exposed Emory mice. (A) Ubiquitin conjugates in the soluble liver supernatants were detectedimmunologically. Equal amounts of protein (50 mg) from each exposure and diet group were separatedby SDS-PAGE and transferred to nitrocellulose. The ubiquitin conjugates were probed with polyclonalantiubiquitin antiserum. Each lane represents the liver supernatant from one animal. C, control; R,restricted. Lane 1–2, paraquat exposed C; lane 3–4, non-stressed C; lane 5–6, paraquat exposed R; lane7–8, non-stressed R. A total of eight animals were used per diet and exposure group. (B) Summary ofscanned densities from autoradiogram of Western blot from Fig. 2A. Each bar represents the mean9S.E. of a total of eight animals per diet and exposure group. Units are arbitrary and normalized tonon-stressed C animals. C, control; R, restricted. Bars not sharing the same letter are significantlydifferent (PB0.05).
3.3. The effects of diet and oxidati6e stress on the acti6ities of E1 and E2 inEmory mouse li6er supernatants
The thiol ester assay (Ciechanover et al., 1982; Haas et al., 1982, 1995) was usedto determine if the stress- and diet-related increases in ubiquitin conjugation activity
M.M. Scrofano et al. / Mechanisms of Ageing and De6elopment 105 (1998) 273–290282
Fig. 3.

M.M. Scrofano et al. / Mechanisms of Ageing and De6elopment 105 (1998) 273–290 283
were associated with an increase in the activities of E1s or E2s. This assay is basedon the known formation of thiol esters between ubiquitin and E1s and E2s in theubiquitin conjugation process (Fig. 1). Thus, the levels of thiol esters reflect theactivities of E1s and E2s (Haas et al., 1995). Consistent with de-novo ubiquitinconjugation activity, E1 activity in the R liver was 40% lower than in C livers undernon-stressed conditions (Fig. 3B, compare lanes 7 and 8 versus lanes 3 and 4; Fig.3C, 0.6790.04 versus 1.1190.06, P=0.0003). After exposure to oxidative stress,this activity increased to the level of C livers, but the E1 activity in C livers did notchange significantly upon paraquat treatment. Thus, after exposure to paraquat, E1activity in liver supernatants from C and R animals were indistinguishable. E1activity in the oxidatively stressed R livers was significantly lower (23%, Fig. 3B,compare lanes 5 and 6 versus lanes 3 and 4; Fig. 3C, 0.8590.06 versus 1.1190.06,P=0.04) than the non-stressed C livers.
In order to determine whether the observed increase in E1 activity correspondedwith an increase in E1 protein levels, we determined E1 protein levels by Westernblots. No significant change was observed in E1 protein levels in Emory liversupernatants with oxidative stress and/or diet (data not shown).
Non-stressed R livers had 48% lower E2 activity than non-stressed C livers (Fig.3B, compare lanes 7 and 8 versus lanes 3 and 4; Fig. 3C, 0.5590.04 versus1.0590.07, P=0.0003). E2 activities in R and C livers treated with paraquat werenot significantly different. Comparable to E1 activity, there were no significantchanges in E2 activities upon imposition of oxidative stress. This obtained for bothdiet groups. However, E2 activity in supernatants from stressed R animals was 32%lower (Fig. 3B, compare lanes 5 and 6 versus lanes 3 and 4; Fig. 3C, 0.7190.08versus 1.0590.07, P=0.01) than in supernatants of the non-stressed C animals.
3.4. Effect of diet and oxidati6e stress on ATP-dependent and ATP-independentdegradation of exogenous substrates
The effects of oxidative stress on proteolysis was assessed by measuring theproteolytic capacity in liver supernatants using labeled b-lactoglobulin and oxidizedRNase as substrates. We found no difference in the ability to degrade either
Fig. 3. (see left) De novo ubiquitin conjugate formation, E1 and E2 activities in liver supernatants fromC- and R-fed non-stressed and paraquat exposed Emory mice. (A) Livers were prepared as described inSection 2. Ubiquitin conjugation, E1 and E2s activities were determined by incubation of the liversupernatant with 125I-labeled ubiquitin in the presence of ATP. Then the mixture was treated with (3A)or without (3B) b-mercaptoethanol, followed by SDS-PAGE. Equal amounts of protein (300 mg/assay)were used in each assay. The amount of de novo ubiquitin conjugates represents ubiquitin conjugationactivity. The level of thiol ester formed between ubiquitin with E1 or E2s represents activities of E1 orE2s. C, control; R, restricted. Lane 1–2, paraquat exposed C; lane 3–4, non-stressed C; lane 5–6,paraquat exposed R; lane 7–8, non-stressed R. A total of eight animals were used per diet and exposuregroup. (C) Summary of densities of autoradiogram scans. Densities were normalized to non-stressed Cliver supernatants. Each bar represents an n=8. For conjugation activity (HMW conj) and E1 activity.* Significantly different from the others (PB0.05). For E2s activities, bars not sharing the same letterare significantly different (PB0.05).

M.M. Scrofano et al. / Mechanisms of Ageing and De6elopment 105 (1998) 273–290284
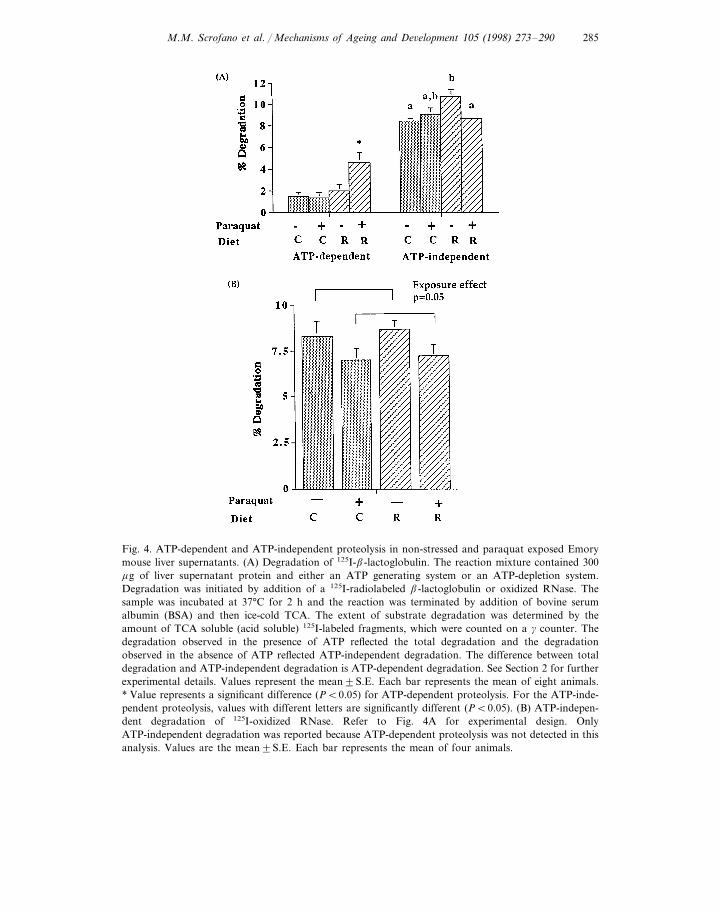
exogenous substrate in non-stressed C or R livers. However, after exposure tooxidative stress, the ATP-dependent degradation of b-lactoglobulin in the R liverswas 238% higher than was observed in paraquat-treated C livers (Fig. 4A, 4.6390.91 versus 1.3790.47, P=0.007). There was a 134% increase in ATP-dependentdegradation of b-lactoglobulin after exposure to oxidative stress in R liver prepara-tions (Fig. 4A, 4.6390.91 versus 1.9890.59, P=0.03), while oxidative stress hadno effect on ATP-dependent degradation of b-lactoglobulin in the C livers.
Non-stressed R livers had a slightly higher percentage of ATP-independentdegradation of b-lactoglobulin than non-stressed C livers (27%, Fig. 4A, 10.7690.61 versus 8.4490.25, P=0.07). There was no significant difference in the rate ofATP-independent degradation of b-lactoglobulin between oxidatively stressed C orR liver. After exposure to oxidative stress, R livers had a 20% lower ATP-indepen-dent degradation of b-lactoglobulin as compared to non-stressed R livers (Fig. 4A,8.6390.07 versus 10.7690.61, P=0.01). There was no effect of oxidative stress onATP-independent degradation in the C livers.
ATP-dependent degradation of oxidized RNase was not detected, thus onlyATP-independent degradation was reported. There was a significant effect ofparaquat exposure but not diet on ATP-independent degradation of oxidizedRNase in non-stressed livers as compared to paraquat-exposed livers, in thatstressed livers had 17% lower ATP-independent degradation of oxidized RNasethan non-stressed livers (Fig. 4B, 7.0890.65 versus 8.4890.63; P=0.05).
4. Discussion
It is well established that R is associated with enhanced ability to withstand stressincluding chronic irradiation (Youngman et al., 1992), heat shock (Heydari et al.,1993, 1995), and viral infection (Effros et al., 1991). This enhanced ability towithstand stress is particularly obvious when older C and R animals are compared(Effros et al., 1991; Youngman et al., 1992; Heydari et al., 1993, 1995). Recentbiochemical investigations indicate that ubiquitination, and/or ubiquitin-dependentproteolysis, are enhanced (perhaps during recovery) in response to oxidative stressin cultured cells or organs (Shang and Taylor, 1995; Shang et al., 1997b) and thatthis oxidation-induced enhancement of the UPP is attenuated upon aging (Shang etal., 1997a). In this work, we asked whether R is associated with an enhancedresponse of the UPP (i.e. endogenous ubiquitin conjugate levels, de-novo conjugateformation, and ubiquitin- and ATP-dependent proteolysis) after exposure to oxida-tive stress in older Emory mice. We noted that levels of endogenous ubiquitinconjugates increased in response to stress in R animals, but not in C animals (Fig.2). The enhanced levels of endogenous ubiquitin conjugates in R mice wereconsistent with enhanced ability to form de-novo ubiquitin conjugates usingexogenous radiolabeled ubiquitin (Fig. 3) and with trends toward enhanced activityof the ubiquitin conjugating enzymes in R animals. This was further supported byenhanced levels of ATP-dependent proteolysis observed in oxidatively stressed asopposed to non-stress animals (Fig. 4), since ATP-dependent proteolysis is also
M.M. Scrofano et al. / Mechanisms of Ageing and De6elopment 105 (1998) 273–290 285
Fig. 4. ATP-dependent and ATP-independent proteolysis in non-stressed and paraquat exposed Emorymouse liver supernatants. (A) Degradation of 125I-b-lactoglobulin. The reaction mixture contained 300mg of liver supernatant protein and either an ATP generating system or an ATP-depletion system.Degradation was initiated by addition of a 125I-radiolabeled b-lactoglobulin or oxidized RNase. Thesample was incubated at 37°C for 2 h and the reaction was terminated by addition of bovine serumalbumin (BSA) and then ice-cold TCA. The extent of substrate degradation was determined by theamount of TCA soluble (acid soluble) 125I-labeled fragments, which were counted on a g counter. Thedegradation observed in the presence of ATP reflected the total degradation and the degradationobserved in the absence of ATP reflected ATP-independent degradation. The difference between totaldegradation and ATP-independent degradation is ATP-dependent degradation. See Section 2 for furtherexperimental details. Values represent the mean9S.E. Each bar represents the mean of eight animals.* Value represents a significant difference (PB0.05) for ATP-dependent proteolysis. For the ATP-inde-pendent proteolysis, values with different letters are significantly different (PB0.05). (B) ATP-indepen-dent degradation of 125I-oxidized RNase. Refer to Fig. 4A for experimental design. OnlyATP-independent degradation was reported because ATP-dependent proteolysis was not detected in thisanalysis. Values are the mean9S.E. Each bar represents the mean of four animals.

M.M. Scrofano et al. / Mechanisms of Ageing and De6elopment 105 (1998) 273–290286
ubiquitin dependent. These data suggest that old R animals are able to mount aUPP response to oxidative stress, whereas old C animals lose this ability. Since theUPP is involved in maintenance of levels of many regulatory molecules (Hershkoand Ciechanover, 1992; Ciechanover and Schwartz, 1994a,b; Wilkinson, 1995;Hochstrasser, 1996) and in selective removal of damaged proteins, enhancement ofthe UPP activity in aged R animals in response to oxidative stress may affordenhanced ability to cope with oxidative stress. The accumulation of cytotoxicdamaged proteins may also be delayed by the enhancement of the UPP activity.The difference in changes in the activity of the UPP upon exposure to paraquatbetween control and calorie-restricted animals may be due to differential responseof the UPP to oxidative stress. It is also possible that levels of reactive oxygenspecies were different due to the differential metabolism of paraquat betweencontrol and calorie-restricted animals.
Together with data from previous studies, these results also suggest that levels ofendogenous conjugates and ubiquitin conjugating activities may be markers of mildoxidative stress or markers of recovery from oxidative stress. This proposal derivesfrom observations that cultured retinal pigment epithelial cells (Jahngen-Hodge etal., 1997; Obin et al., 1998), cultured whole lenses and lens cells (Shang and Taylor,1995; Shang et al., 1997a,b), and yeast (Cheng et al., 1994), also show alteredubiquitination and ubiquitin conjugation activity in response to various stresses.This is corroborated by the alterations in UPP activity in these R and C animalsupon exposure to oxidative stress.
Both enhanced and reduced UPP activity in response to oxidation can berationalized. Oxidative stress may increase the levels of damaged or abnormalproteins which are substrates for the UPP, and this increase could result in anincrease in the activity of the UPP, including elevated levels of ubiquitin conjugates.However, most of the enzymes in the UPP have essential cysteines, as such they arevulnerable to oxidative stress. Thus, severe oxidative stress might diminish theefficacy of the UPP (Shang and Taylor, 1995), presumably by oxidatively inactivat-ing the E1–ubiquitin conjugating enzyme (Shang et al., 1997b). This inactivationcould result in lower levels of ubiquitin conjugates.
It is of interest that levels of endogenous ubiquitin conjugates are as high innon-stressed C animals as in stressed R animals. This suggests that the C animalsare under chronic stress, even in the absence of an external insult. The presumptivechronic stressed state of the C animals also allows rationalization of the decrease inendogenous ubiquitin conjugates when external oxidative stress is applied to Canimals, i.e that further oxidative stress to the old C animals could overwhelm theantioxidant system and damage the UPP. This would be consistent with our priorstudies which indicate that UPP capability is diminished when the redox status ratio(GSSG/GSH) increases to greater than 0.5 in hydrogen-peroxide-treated retinas(Jahngen-Hodge et al., 1997) or in diamide-treated retinal pigment epithelial cells(Obin et al., 1998). Thus, decreased UPP activity upon oxidative stress in C animalswould be rationalized if such stress caused GSSG/GSH ratios to increase to levelswhich inactivate ubiquitin conjugating enzymes.

M.M. Scrofano et al. / Mechanisms of Ageing and De6elopment 105 (1998) 273–290 287
Although livers from non-stressed old C animals had higher levels of endogenousubiquitin conjugates and higher ubiquitin conjugating activity than livers fromnon-stressed old R animals, the ability to degrade b-lactoglobulin by the UPP wascomparable between C and R animals. Since previous studies have shown thatATP-dependent proteolysis is usually correlated with the levels of ubiquitin conju-gates (Chin et al., 1982; Jabben et al., 1989; Obin et al., 1994; Shang and Taylor,1995), the lack of a correlation between proteolytic capacity and ubiquitin conju-gating activity in the non-stressed old C animals remains an enigma. It is possiblethat the 26S proteasome which degrades proteins conjugated to ubiquitin may becompromised in the old C animals.
It has been reported that oxidized RNase is also a good substrate for the UPP(Gonda et al., 1989). However, no ATP-dependent degradation of ox RNase wasobserved in the Emory mouse liver. In contrast to the oxidation-induced increase inATP/UPP-dependent degradation of b-lactoglobulin, the ATP-independent degra-dation of oxidized Rnase and b-lactoglobulin decreased or did not change uponoxidation. This lack of an effect of oxidative stress on ATP-independent proteolyticcapacity is consistent with data by Grune et al. (Grune et al., 1995, 1996, 1997),which showed that although oxidized proteins are degraded by the ATP-indepen-dent proteolytic system (20S proteasome), ATP-independent proteolytic capacity isnot up-regulated by oxidative stress.
In summary, we demonstrated for the first time that the UPP responds tooxidative stress in old Emory mouse livers and that feeding the R diet is associatedwith an enhanced response of UPP to oxidative stress.
Acknowledgements
This work was supported in part with federal funds from the USDA undercontract c53-3K06-01, by NIH grant cEY08566 and NIH training grantcT32AG00209.
References
Agarwal, S., Sohal, R.S., 1994. Aging and proteolysis of oxidized proteins. Arch. Biochem. Biophy. 309,24–28.
Ames, B.N., Shigenaga, M.K., Hagen, T.M., 1993. Oxidants, antioxidants, and the degenerative diseasesof aging. Proc. Natl. Acad. Sci. USA 90, 7915–7922.
Cheng, L., Watt, R., Piper, P.W., 1994. Polyubiquitin gene expression contributes to oxidative stressresistance in respiratory yeast (Saccharomyces cere6isiae). Mol. Gen. Genet. 246, 358–362.
Chin, D.T., Kuehl, L., Rechsteiner, M., 1982. Conjugation of ubiquitin to denatured hemoglobin isproportional to the rate of hemoglobin degradation in HeLa cells. Proc. Natl. Acad. Sci. USA 79,5857–5861.
Ciechanover, A., Schwartz, A.L., 1994a. The ubiquitin-mediated proteolytic pathway: mechanisms ofrecognition of the proteolytic substrate and involvement in the degradation of native cellularproteins. FASEB J. 8, 182–191.
Ciechanover, A.J., Schwartz, A.L. (Eds.), 1994b. Cellular Proteolytic Systems. WileyLiss, New York.

M.M. Scrofano et al. / Mechanisms of Ageing and De6elopment 105 (1998) 273–290288
Ciechanover, A., Hod, Y., Hershko, A., 1978. A heat-stable polypeptide component of an ATP-depen-dent proteolytic system from reticulocytes. Biochem. Biophys. Res. Commun. 81, 1100–1105.
Ciechanover, A., Heller, H., Elias, S., Haas, A.L., Hershko, A., 1980. ATP-dependent conjugation ofreticulocyte proteins with the polypeptide required for protein degradation. Proc. Natl. Acad. Sci.USA 77, 1365–1368.
Ciechanover, A., Elias, S., Heller, H., Hershko, A., 1982. ‘Covalent affinity’ purlfication of ubiquitin-ac-tivating enzyme. J. Biol. Chem. 257, 2537–2542.
Costa, R.C., Del Monte, U., 1994. ATP-stimulated proteolysis by soluble extracts of liver of aged rats.In: Modern Aging Research 9: Protein Metabolism and Aging. Wiley-Liss, New York, pp. 151–158.
Costa, R.C., Bianchi, C., Del Monte, U., 1991. ATP− and ATP+ ubiquitin-stimulated proteolysis inrat liver and yoshida ascites hepatoma. Biochem. Biophys. Res. Commun. 179, 1400–1407.
Effros, R.B., Walford, R.L., Weindruch, R., Micheltree, C., 1991. Influences of dietary restriction onimmunity to influenza in aged mice. J. Gerontol. Biol. Sci. 46, B142–147.
Evans, A.C. Jr., Wilkinson, K.D., 1985. Ubiquitin-dependent proteolysis of native and alkylated bovineserum albumin: Effects of protein structure and ATP concentration on selectivity. Biochem. 24,2915–2923.
Finley, D., Chau, V., 1991. Ubiquitination. Ann. Rev. Cell Biol. 7, 25–69.Garland, D., Russel, R., Zigler, J.S. Jr., 1988. The oxidative modification of lens proteins. Basic Life Sci.
49, 347–352.Glotzer, M., Murray, A.W., Kirschner, M.W., 1991. Cyclin is degraded by the ubiquitin pathway.
Nature 349, 132–138.Gonda, D.K., Bachmair, A., Wunning, I., Tobias, J.W., Lane, W.S., Varshavsky, A., 1989. Universality
and structure of the N-end rule. J. Biol. Chem. 264, 16700–16712.Gong, X., Obin, M., Palmer, H., Scrofano, M.M., Jahngen-Hodge, J., Smith, D.E., Taylor, A., 1997.
Antioxidant enzymes in lens, liver and kidney of calorie restricted mice. Mech. Ageing Dev. 91,115–129.
Grune, T., Reinheckel, T., Joshi, M., Davies, K.J., 1995. Proteolysis in cultured liver epithelial cellsduring oxidative stress. Role of multicatalytic proteinase complex, proteasome. J. Biol. Chem. 270,2344–2351.
Grune, T., Reinheckel, T., Davies, K.J.A., 1996. Degradation of oxidized proteins in K562 humanhematopoietic cells by proteasome. J. Biol. Chem. 271, 15504–15509.
Grune, T., Reinheckel, T., Davies, K.J.A., 1997. Degradation of oxidized proteins in mammalian cells.FASEB J. 11, 526–534.
Haas, A.L., Warms, J.V.B., Hershko, A., Rose, I.A., 1982. Ubiquitin-activating enzyme: Mechanismand role in protein-ubiquitin conjugation. J. Biol. Chem. 257, 2543–2548.
Haas, A.L., Murphy, K.E., Bright, P.M., 1986. The inactivation of ubiquitin accounts for the inabilityto demonstrate ATP, ubiquitin-dependent proteolysis in liver extracts. J. Biol. Chem. 260, 4694–4703.
Haas, A.L., Baboshina, O., Williams, B., Schwartz, L.M., 1995. Coordinated induction of the ubiquitinconjugation pathway accompanies the developmentally programmed death of insect skeletal muscle.J. Biol. Chem. 270, 9407–9412.
Halliwell, B., Gutteridge, J.M.C., Cross, C.E., 1992. Free radicals, antioxidants, and human diseases:Where are we now? J. Lab. Clin. Med. 119, 598–620.
Harman, D., 1956. Aging: a theory based on free radical and radiation chemistry. J. Gerontol. 11, 289.Harman, D., 1994. Aging prospects for further increases in the functional life span. Age 17, 119–146.Hershko, A., Ciechanover, A., 1992. The ubiquitin system for protein degradation. Ann. Rev. Biochem.
61, 761–807.Hershko, A., Ciechanover, A., Rose, I.A., 1979. Resolution of the ATP-dependent proteolytic system
from reticulocytes: A component that interacts with ATP. Biochem. 76, 3107–3110.Hershko, A., Ciechanover, A., Heller, H., Haas, A.L., Rose, I.A., 1980. Proposed role of ATP in protein
breakdown: Conjugation of proteins with multiple chains of the polypeptide of ATP-dependentproteolysis. Proc. Natl. Acad. Sci. USA 77, 1783–1786.
Hershko, A., Heller, H., Elias, S., Ciechanover, A., 1983. Components of the ubiquitin-protein ligasesystem. Resolution, affinity purlfication, and role in protein breakdown. J. Biol. Chem. 258,8206–8214.

M.M. Scrofano et al. / Mechanisms of Ageing and De6elopment 105 (1998) 273–290 289
Hershko, A., Heller, H., Eytan, E., Kaklu, G., Rose, I.A., 1984a. Role of the a-amino group inubiquitin-mediated protein breakdown. Proc. Natl. Acad. Sci. USA 81, 7021–7025.
Hershko, A., Leshinsky, E., Ganoth, D., Heller, H., 1984b. ATP-dependent degradation of ubiquitin-protein conjugates. Proc. Natl. Acad. Sci. USA 81, 1619–1623.
Hershko, A., Ganoth, D., Sudakin, V., Dahan, A., Cohen, L.H., Luca, F.C., Ruderman, J.V., Eytan, E.,1994. Components of a system that ligates cyclin to ubiquitin and their regulation by the proteinkinase cdc2. J. Biol. Chem. 269, 4940–4946.
Heydari, A.R., Wu, B., Takahasi, R., Strong, R., Richardson, A., 1993. Expression of heat shockprotein 70 is altered by age and diet at the level of transcription. Mol. Cell. Biol. 13, 2909–2918.
Heydari, A.R., Conrad, G.C., Richardson, A., 1995. Expression of heat shock genes in hepatocytes isaffected by age and food restriction in rats. J. Nutr. 125, 410–418.
Hochstrasser, M., 1996. Ubiquitin-dependent protein degradation. Ann. Rev. Genet. 30, 405–439.Huang, L.L., Shang, F., Taylor, A., 1993. Ubiquitin dependent degradation of native and oxidized
alpha-crystalline by bovine lens epithelial cell. Invest. Ophthalmol. Vis. Sci. 34, 1297.Huang, L.L., Shang, F., Nowell, T.R. Jr., Taylor, A., 1995. Degradation of differentially oxidized
a-crystalline in bovine lens epithelial cells. Exp. Eye Res. 61, 45–54.Jabben, M., Shanklin, J., Vierstra, R.D., 1989. Ubiquitin-phytochrome conjugates: pool dynamics
during in vivo phytochrome degradation. J. Biol. Chem. 264, 4998–5005.Jacques, P.F., Chylack, L.T. Jr., 1991. Epidemiologic evidence of a role for the antioxidant vitamins and
carotenoids in cataract prevention. Am. J. Clin. Nutr. 53, 3520S–3525S.Jahngen, J.H., Haas, A.L., Ciechanover, A., Blondin, J., Eisenhauer, D., Taylor, A., 1986. The eye lens
has an active ubiquitin-protein conjugation system. J. Biol. Chem. 261, 13760–13787.Jahngen-Hodge, J., Obin, M., Gong, X., Shang, F., Nowell, T., Gong, J., Abasi, H., Blumberg, J.,
Taylor, A., 1997. Regulation of ubiquitin conjugating enzymes by glutathione following oxidativestress. J. Biol. Chem. 272, 28218–28226.
Johnston, N.L., Cohen, R.E., 1991. Uncoupling ubiquitin-protein conjugation from ubiquitin-dependentproteolysis by use of b,g-nonhydrolyzable ATP analogues. Biochem. 30, 7514–7522.
Kudo, T., Iqbal, K., Ravid, R., Swaab, D.F., Grundke-Iqbal, I., 1994. Alzheimer disease: correlation ofcerebro-spinal fluid and brain ubiquitin levels. Brain Res. 639, 1–7.
Loetscher, P., Pratt, G., Rechsteiner, M., 1991. The C terminus of mouse ornithine decarboxylaseconfers rapid degradation on dihydrofolate reductase. Support for the pest hypothesis. J. Biol. Chem.266, 11213–11220.
Lowe, J., Blanchard, A., Morrell, K., Lennox, G., Reynolds, L., Billet, M., Landon, M., Mayer, J.,1988. Ubiquitin is a common factor in intermediate filament inclusion bodies of diverse type in man,including those of Parkinson’s disease, Pick’s disease, and Alzheimer’s disease as well as Rosenthalfibres in cerebellar astrocytomas, cytoplasmic bodies in muscle, and Mallory bodies in alcoholic liverdisease. J. Pathol. 155, 9–15.
Lowry, O.H., 1951. J. Biol. Chem. 193, 265.Masoro, E.J., 1993. Dietary restriction and aging. J. Am. Geriat. Soc. 41, 994–999.Mura, C.V., Gong, X., Taylor, A., Villalobos-Molina, R., Scrofano, M.M., 1996. Effects of calorie
restriction and aging on the expression of antioxidant enzymes and ubiquitin in the liver of Emorymice. Mech. Ageing Dev. 91, 115–129.
Murakami, K., Jahogen, J.H., LIn, S.W., Davies, K.J.A., Taylor, A., 1990. Lens proteasome showsenhanced rates of degradation of hydroxyl radical modified alpha-crystallin. Free Rad. Biol. Med. 8,217–222.
Obin, M., Nowell, T., Taylor, A., 1994. The photoreceptor G-protein transducin (Gt) is a substrate forubiquitin-dependent proteolysis. Biochem. Biophys. Res. Commun. 200, 1169–1176.
Obin, M., Shang, F., Gong, X., Handelman, G., Blumberg, J., Taylor, A., 1998. Redox regulation ofubiquitin-conjugating enzymes: Mechanistic insights using the thiol-specific oxidant diamide. FASEBJ. 12, 561–569.
Oliver, C.N., Ahn, B., Moerman, E.J., Goldstein, S., Stadtman, E.R., 1987. Age-related changes inoxidized proteins. J. Biol. Chem. 262, 5488–5491.
Peters, J.-M., 1994. Proteasomes: protein degradation machines of the cell. Trends Biochem. Sci. 19,377–382.

M.M. Scrofano et al. / Mechanisms of Ageing and De6elopment 105 (1998) 273–290290
Rechsteiner, M., 1987. Ubiquitin-mediated pathways for intracellular proteolysis. Ann. Rev. Cell Biol.3, 1–30.
Rechsteiner, M., 1991. Natural substrates of the ubiquitin proteolytic pathway. Cell 66, 615–618.Saenghirunvattana, S., Sermswan, A., Piratchvej, V., Rochanawutanon, M., Kaojarern, S., Rat-
tananenya, T., 1992. Effect of lung irradiation on mice following paraquat intoxication. Chest 101,833–835.
Scheffner, M., Huibregtse, J.M., Howley, P.M., 1994. Identification of a human ubiquitin-conjugatingenzyme that mediates the E6-AP-dependent ubiquitination of p53. Proc. Natl. Acad. Sci. USA 91,8797–8801.
Shang, F., Taylor, A., 1995. Oxidative stress and recovery from oxidative stress are associated withaltered ubiquitin conjugating and proteolytic activities in bovine lens epithelial cells. Biochem. J. 307,297–303.
Shang, F., Gong, X., Palmer, H.J., Nowell, T.R. Jr., Taylor, A., 1997a. Age-related decline in ubiquitinconjugation in response to oxidative stress in the lens. Exp. Eye Res. 64, 21–30.
Shang, F., Gong, X., Taylor, A., 1997b. Activity of ubiquitin dependent pathway in response tooxidative stress: Ubiquitin activating enzyme is transiently upregulated. J. Biol. Chem. 272, 23086–23093.
Shibatani, T., Ward, W.F., 1996. Effect of age and food restriction on alkaline protease activity in ratliver. J. Gerontol. Biol. Sci. 51A, B175–B178.
Sohal, R.S., Agarwal, A., Dubey, A., Orr, W.C., 1993. Protein oxidative damage is associated with lifeexpectancy of houseflies. Proc. Natl. Acad. Sci. USA 90, 7255–7259.
Sohal, R.S., Weindruch, R., 1996. Oxidative stress, caloric restriction, and aging. Science 273, 59–63.Stadtman, E.R., 1992. Protein oxidation and aging. Science 257, 1220–1224.Taylor, A., 1992. Effect of photooxidation on the eye lens and role of nutrients in delaying cataract. In:
Free Radicals and Aging. Birkhauser Verlag, Basel, Switzerland, pp. 266-279.Taylor, A., Davies, K.L.A., 1987. Protein oxidation and loss of protease activity may lead to cataract
formation in the aged lens. Free Rad. Biol. Med. 3, 371–377.Taylor, A., Jahngen-Hodge, J., Huang, L., Jacques, P., 1991. Aging in the eye lens: roles for proteolysis
and nutrition in formation of cataract. Age 14, 65–71.Taylor, A., Lipman, R.D., Jahngen-Hodge, J., Smith, D., Dallal, G.E., Solomon, R., Perrone, G.,
Meydani, M., Harrison, D., Shigenaga, M., 1995. Dietary calorie restriction in the Emory mouse:effects on lifespan, eye lens cataract prevalence and progression, levels of ascorbate, glucose, andglycohemoglobin, and tail collagen breaktime, susceptibility to infection, tail necrosis, priapism,activity, oxidation of DNA and RNA, cancer and fecundity. Mech. Ageing Dev. 79, 33–57.
Villarreal, A., Villarreal, J., Ward, W.F., 1997. Alteration of proteasome structure and function by ageand food restriction. FASEB J. A426, 2464.
Vincent, I.J., Davies, P., 1990. ATP-induced loss of Alz-50 immunoreactivity with A68 proteins fromAlzheimer brain is mediated by ubiquitin. Proc. Natl. Acad. Sci. USA 87, 4840–4844.
Ward, W.F., Shibatani, T., 1994. Dietary modulation of protein turnover. In: Modulation of AgingProcesses by Dietary Restriction. CRC Press, Boca Raton, FL, pp. 121-142.
Weindruch, R., 1991. Retardation of aging by caloric restriction. In: The Potential for NutritionalModulation of Aging Processes. Food and Nutrition Press, Trumbell, CN, pp. 109-121.
Weindruch, R., Walford, R.L., 1988. The Retardation of Aging and Diseases by Dietary Restriction.Charles C. Thomas, Springfield, IL.
Wilkinson, K.D., 1995. Roles of ubiquitinylation in proteolysis and cellular regulation. Ann. Rev. Nutr.15, 161–189.
Youngman, L.D., Park, J.-L.K., Ames, B.N., 1992. Protein oxidation associated with aging is reducedby dietary restriction of protein or calories. Proc. Natl. Acad. Sci. USA 89, 9112–9116.
Yu, B.P., 1996. Aging and oxidative stress: Modulation by dietary restriction. Free Rad. Biol. Med. 21,651–668.
..