calorimetry and thermodynamics in drug design - …...enthalpy values may be temperature de-pendent,...
TRANSCRIPT

ANRV343-BB37-07 ARI 24 April 2008 15:14
Calorimetry andThermodynamicsin Drug DesignJonathan B. ChairesJames Graham Brown Cancer Center, University of Louisville, Louisville,Kentucky 40202; email: [email protected]
Annu. Rev. Biophys. 2008. 37:135–51
First published online as a Review in Advance onJanuary 24, 2008
The Annual Review of Biophysics is online atbiophys.annualreviews.org
This article’s doi:10.1146/annurev.biophys.36.040306.132812
Copyright c© 2008 by Annual Reviews.All rights reserved
1936-122X/08/0609-0135$20.00
Key Words
free energy, enthalpy, entropy, binding, rational drug design
AbstractModern instrumentation for calorimetry permits direct determina-tion of enthalpy values for binding reactions and conformationaltransitions in biomolecules. Complete thermodynamic profiles con-sisting of free energy, enthalpy, and entropy may be obtained forreactions of interest in a relatively straightforward manner. Suchprofiles are of enormous value in drug design because they pro-vide information about the balance of driving forces that cannotbe obtained from structural or computational methods alone. Thisperspective shows several examples of the insight provided by ther-modynamic data in drug design.
135
Ann
u. R
ev. B
ioph
ys. 2
008.
37:1
35-1
51. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsite
Lou
is P
aste
ur -
U. S
tras
bour
g 1,
2 an
d 3
on 0
5/20
/08.
For
per
sona
l use
onl
y.

ANRV343-BB37-07 ARI 24 April 2008 15:14
Contents
INTRODUCTION. . . . . . . . . . . . . . . . . 136BASIC THERMODYNAMIC
RELATIONSHIPS . . . . . . . . . . . . . . 136MODERN
MICROCALORIMETRY . . . . . . . . 138HIGH-THROUGHPUT
THERMODYNAMICS ANDCALORIMETRY . . . . . . . . . . . . . . . . 140
CASE STUDIES: BINDINGTO NUCLEIC ACIDS . . . . . . . . . . 141Isostructural Is Not Isoenergetic . . 141A Thermodynamic Signature
for DNA Binding Mode . . . . . . . 142Measuring Ultratight DNA
Binding by DSC. . . . . . . . . . . . . . . 143CASE STUDIES: PROTEIN
BINDING INTERACTIONS . . . 144Optimizing Affinity in HIV-1
Protease Inhibitors . . . . . . . . . . . . 144The Enthalpy Funnel . . . . . . . . . . . . . 145
CASE STUDY: HIGHSELECTIVITY DOES NOTDEMAND HIGH AFFINITY . . . 145
CONCLUSIONS. . . . . . . . . . . . . . . . . . . 147
INTRODUCTION
Biological calorimetry dates from the 1780s,when Lavoisier and Laplace used their icecalorimeter to measure the metabolic heatsof live animals (35, 36, 40). They studied,for example, the metabolism of a guinea pigby weighing the ice melted in the calorime-ter over a given observation period, a valuethat could then be converted to energy fromthe knowledge of the latent enthalpy of fu-sion of water. This metabolic heat was cor-related with CO2 production over the sametime period to provide some of the first quan-titative data in physiology. Until fairly re-cently, the use of calorimetry in biophysicsand molecular biology was limited to a fewexpert laboratories that were able to buildtheir own instruments with the requisite sen-sitivity to measure the heats of reactions of
biomolecules that are often only available inlimited quantities. This era when calorime-try was in the hands of a few and largely in-accessible to many has been characterized byAckers & Bolen (1) as the time of the “cult ofcalorimetry.” The cult has expanded. Highlysensitive microcalorimeters for biochemicaland biophysical applications became commer-cially available, leading to a fairly widespreaduse of the calorimetric methods in biophysicsand molecular biology. Such widespread useis perhaps inevitably accompanied by someabuse, a cause for some concern (70).
Recent drug discovery efforts have beendominated by structure-based design con-cepts. In structure-based design, lead com-pounds are sought by attempting to matchtheir shapes with the complementary shapesof active sites of receptors using known struc-tures obtained by X-ray crystallography orNMR. It was noted previously, however, thatthermodynamic studies are an essential andnecessary complement to structural studies indrug design (25). Structural data alone, evenwhen coupled with the most sophisticated cur-rent computational methods, cannot fully de-fine the driving forces for binding interac-tions or even accurately predict their bindingaffinities. Thermodynamics provides quanti-tative data of use in elucidating these driv-ing forces and for evaluating and understand-ing at a deeper level the effects of substituentchanges on binding affinity (74).
This review explores several examples inwhich thermodynamic studies have providedcomplementary data that serve to enhancedrug design efforts. Space limitations do notpermit a comprehensive review of thermo-dynamics in drug discovery, so the exampleswere selected somewhat arbitrarily. I apolo-gize in advance to those authors whose im-portant work I have neglected.
BASIC THERMODYNAMICRELATIONSHIPS
Basic thermodynamic relationships (15) aresummarized in the sidebar. Equilibrium
136 Chaires
Ann
u. R
ev. B
ioph
ys. 2
008.
37:1
35-1
51. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsite
Lou
is P
aste
ur -
U. S
tras
bour
g 1,
2 an
d 3
on 0
5/20
/08.
For
per
sona
l use
onl
y.

ANRV343-BB37-07 ARI 24 April 2008 15:14
constants may be used to calculate standardfree energy changes (�G◦) using the Gibbsrelationship (Equation 1.1). The standard freeenergy change refers to an arbitrary and oftenunrealistic standard state, typically 1 M reac-tant concentrations. The free energy change(�G) may be calculated from �G◦ for a par-ticular set of reaction conditions defined bythe ratio Q (Equation 1.2). The ratio Q is afunction of the ratio of actual product and re-actant concentrations, the exact mathematicalform of which depends on the exact reactionmechanism. Enthalpy values (�H) may be de-termined directly by calorimetry or indirectlyfrom the temperature dependence of equilib-rium constants and application of the van’tHoff relationship (�HVH; Equation 1.4). Freeenergy may be parsed into its enthalpic andentropic (�S) components (Equation 1.3).Enthalpy values may be temperature de-pendent, signifying a nonzero heat capacitychange (�Cp; Equation 1.5). If the heat ca-pacity change is not zero, a more compli-cated expression for the free energy is needed(Equation 1.8). A complete thermodynamicprofile for a reaction of interest requires de-termination of the free energy, enthalpy, andentropy at a given temperature, and the heatcapacity change if the enthalpy varies withtemperature.
The free energy �G is the key parameter,because its value under a particular set of re-actant concentrations dictates the direction ofbiomolecular equilibria. If its sign is negative,the binding reaction or conformational tran-sition will proceed spontaneously to an extentgoverned by the magnitude of �G. If its signis positive, the magnitude of �G specifies theenergy needed to drive the reaction to formproduct. The free energy is a balance betweenenthalpy and entropy. The enthalpy changereflects the amount of heat energy requiredto achieve a particular state, and the entropymeasures how easily that energy might be dis-tributed among various molecular energy lev-els. For binding reactions, negative enthalpyvalues are common (but not omnipresent), re-flecting a tendency for the system to fall to
BASIC THERMODYNAMIC RELATIONSHIPS
�G0 = −RT ln Ka 1.1
�G = �G0 + RT ln Q 1.2
Q = f (product concentrations)f (reactant concentrations)
�G = �H − T�S 1.3
�HV H = −R(
δ ln Ka
δ(1/T)
)1.4
�Cp =(
δ�HδT
)p
1.5
�HT = �Hr + �Cp (T − Tr ) 1.6
�ST = �Sr + �Cp ln(
TTr
)1.7
�G = �Hr − T�Sr + �Cp
[(T − Tr ) − T ln
(TTr
)]1.8
Notes: �G, free energy change; �H, enthalpy change; �S,entropy change; �Cp, heat capacity change. T and Tr refer tothe temperature and an arbitrary reference temperature, re-spectively. Thermodynamic parameters subscripted with “T”or “r” refer to those temperatures.
lower energy levels by bond formation. Pos-itive entropy values are common for bind-ing reactions, reflecting a natural tendencyfor disruption of order, commonly resultingfrom disruption of solvation interactions andthe concomitant release of bound water.
All binding reactions must overcome in-escapable entropic penalties (2, 3, 6, 20, 33,42, 60). The first penalty results from the lossof rotational and translational degrees of free-dom as two reacting partners combine to be-come one complex. At least six degrees of free-dom are lost upon complex formation owingto restriction of translational and rotationalmotions. This penalty may result in an en-ergy cost of up to +15 kcal mol−1, althoughthe exact value is subject to debate (2, 3, 6,20, 33, 42, 60). The second penalty arisesfrom the restriction of individual bond ro-tation upon complex formation, for example,
www.annualreviews.org • Calorimetry and Thermodynamics in Drug Design 137
Ann
u. R
ev. B
ioph
ys. 2
008.
37:1
35-1
51. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsite
Lou
is P
aste
ur -
U. S
tras
bour
g 1,
2 an
d 3
on 0
5/20
/08.
For
per
sona
l use
onl
y.

ANRV343-BB37-07 ARI 24 April 2008 15:14
DSC: differentialscanning calorimetry
ITC: isothermalscanning calorimetry
when a small molecule is locked into a partic-ular conformation when bound to its receptorsite. The penalty for restriction of bond rota-tion is estimated to be about 0.6 kcal mol−1
per bond (44). This penalty might be reducedby engineering rigid bonds into ligand drugcandidates if sufficient structural data exist toguide the design to ensure a precise fit intothe receptor site. These inescapable entropiccosts must be balanced by, and overcome by,other favorable interactions if binding is to oc-cur. That is one fundamental consideration indrug design.
Although it would appear easy to assign en-thalpy and entropy contributions to the smallcatalog of noncovalent interactions importantin biological binding interactions (hydrogenbonding, hydrophobic and electrostatic in-teractions, and van der Waals forces), it isdifficult to disentangle their separate contri-butions to the energetics of biomolecular in-teractions and transitions (14). Nonetheless,some tenuous attempts have been made toparse the free energy of drug-DNA bindinginteractions into component contributions (9,22, 23, 55). Perhaps a better, and safer, pointof view is one proposed by Freire and cowork-ers, who view binding enthalpy as a measure ofbond formation (hydrogen and van der Waalsbonding) and therefore of binding specificity,while binding entropy is a measure primar-ily of the nonspecific hydrophobic force (56,66, 68). Examples of the utility of this vieware discussed below. The interpretation ofthermodynamic data in term of componentmolecular interactions is discussed in severalpublications (9, 14, 20, 22, 27, 28, 33, 60, 72).
Enthalpy changes can be obtained by ei-ther calorimetry or, in principle, from thevan’t Hoff relationship using equilibrium con-stants measured as a function of temperature.In an ideal world measured calorimetric andvan’t Hoff enthalpy values will be equivalentfor a given reaction if the proper reactionmechanism is used in data reduction and anal-ysis. The proper reaction mechanism may in-clude coupled processes such as protonation,ion binding and release, or conformational
changes. There are reports of discrepanciesbetween calorimetric and van’t Hoff enthalpyvalues (39, 43). The use of van’t Hoff analysisto obtain enthalpy values, however, is fraughtwith error and difficulty (10, 29). A funda-mental problem is that, given even modest er-rors in equilibrium constant determinations,the presence of curvature in van’t Hoff plotsthat would result from nonzero heat capacitychanges may not be apparent. In such cases,linear fits to the data that neglect heat capac-ity changes would lead to systematically biasedenthalpy values that would differ significantlyfrom calorimetric values (10).
MODERNMICROCALORIMETRY
The modern microcalorimetry revolution wasfueled by the availability of sensitive commer-cial instruments. Two calorimetric methods,differential scanning calorimetry (DSC) andisothermal scanning calorimetry (ITC), dom-inate in biophysics and biochemistry (15).
Instruments for both approaches mostcommonly use a power compensation design.In DSC, the temperature difference betweensample and reference cells is monitored asthe temperature of both is increased in a pre-cisely controlled fashion. A thermally drivenreaction in the sample cell leads to a tem-perature imbalance between the cells. Whenthat occurs, power to the heaters on the cellsis modulated to bring the sample and ref-erence back into balance. The primary datarecorded is compensation power as a func-tion of temperature. Power, as a fundamentalphysical quantity, is readily converted to en-ergy units of joules or calories. Descriptions ofthe most recent DSC instrument design havebeen published (49, 54).
Representative DSC data are shown inFigure 1. The area under the transition curvedefines the enthalpy of the reaction. A dif-ference between the pre- and post-transitionbaselines results if there is a change in heatcapacity. The distance between the two base-lines in such cases defines the value of the heat
138 Chaires
Ann
u. R
ev. B
ioph
ys. 2
008.
37:1
35-1
51. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsite
Lou
is P
aste
ur -
U. S
tras
bour
g 1,
2 an
d 3
on 0
5/20
/08.
For
per
sona
l use
onl
y.

ANRV343-BB37-07 ARI 24 April 2008 15:14
0.0 0.5 1.0 1.5 2.0 2.5 3.0 3.5
-16
-12
-8
-4
0
40 50 60 70 80 90
0
20
40
60
80
-5
-4
-3
-2
-1
0
10 20 40 60 80 100 120
Time (min)
μca
l/sec
Molar ratio
kcal
/mo
le o
f lig
and Exc
ess
he a
t ca
pac
ity
Temperature (°C)
∆Cp
Tm
a b
Figure 1Representative ITC (a) or DSC (b) data. The ITC data are for the binding of 2′CMP to RNaseA. The toppanel shows the primary titration data, and the lower panel shows the binding isotherm constructed fromthe primary data. The DSC transition shown is for the thermal denaturation of the protein ubiquitin.The diagram indicates the melting transition temperature Tm and the heat capacity change �Cp.
capacity change. The degree of the transitionat any temperature may be defined by inte-gration of the transition curve. Recommenda-tions for the proper presentation and process-ing of DSC data were recently published (26).
In ITC, again the temperature differencebetween a sample cell and a reference cell ismonitored. In contrast to DSC, both cells inthis case are maintained at a constant temper-ature. A titration system delivers a reactant tothe sample cell. Any heat of reactant bindingresults in an imbalance between the referenceand sample cells, which is compensated for bymodulating the power applied to cell heaters.
A description of an ITC instrument was pub-lished (77).
Representative ITC data are shown inFigure 1. The top panel shows primary data,in which power is displayed as a functionof time. The programmed titration deliverysteps are evident by the series of peaks thatreturn to baseline. The area of each peak isthe heat of reaction for each reactant addi-tion. As the receptor binding site becomessaturated, the magnitude of the peak area de-creases. The bottom panel shows transforma-tion of the primary ITC data into the form ofa binding isotherm. Such a binding isotherm
www.annualreviews.org • Calorimetry and Thermodynamics in Drug Design 139
Ann
u. R
ev. B
ioph
ys. 2
008.
37:1
35-1
51. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsite
Lou
is P
aste
ur -
U. S
tras
bour
g 1,
2 an
d 3
on 0
5/20
/08.
For
per
sona
l use
onl
y.

ANRV343-BB37-07 ARI 24 April 2008 15:14
may be fit to a particular binding model toobtain a binding constant, binding enthalpyand the stoichiometry of binding. Some nec-essary considerations in fitting ITC data canbe found in references (18, 32, 61–64).
HIGH-THROUGHPUTTHERMODYNAMICSAND CALORIMETRY
DSC and ITC are somewhat laborious andtime-consuming methods. Even a practicedinvestigator can usually complete only a fewexperiments per day. Demands of the pharma-ceutical industry require higher throughputthan is typically possible with available singlesample cell DSC and ITC instruments, whichoften relegates calorimetry to be used as a sec-ondary screening method or for validation ofother assays. To overcome these limitations,several attempts have been made to improvethe throughput of calorimetry and thermody-namic measurements.
Perhaps the most impressive attempt is thedevelopment of ThermoFluorTM technology(47) by 3D Pharmaceuticals, Inc., now ownedby Johnson & Johnson. ThermoFluorTM maybe used to conduct high-density miniaturizedthermal shift assays as a screen in drug discov-ery. The technique measures the fluorescenceof fluorescent dyes that sense protein un-folding. A 384-well microplate that can beheated at a defined rate is used. Libraries ofsmall molecules can be screened for bindingto a particular protein target of interest,exploiting firm underlying thermodynamicprinciples. Ligand binding is thermody-namically coupled to protein denaturationreactions. Ligands that bind preferentially tothe native folded state will stabilize the pro-tein, elevating its melting temperature. Themagnitude of the melting temperature shiftis a function of the binding affinity, althoughother binding properties also exert an influ-ence (71). Thus, ThermoFluorTM technologyallows characterization of the thermal denat-uration of the target protein of interest aloneor in the presence of drug candidates, provid-
ing a quantitative high-throughput screeningmethod that yields accurate thermodynamicinformation. By this approach, bindingto hundreds of different protein targets bylibraries containing tens of thousands of com-pounds was made possible, greatly facilitatingthe drug discovery process. An additionalnovel use of the ThermoFluorTM technologyis to identify biological function. For example,a screen for ligand binding to a protein prod-uct of an essential gene of unknown functionwas devised (8). ThermoFluorTM was used toscreen the protein against a library of 3000compounds that were specifically selected toprovide information about possible biologicalfunctions, and identified binding of only par-ticular ligands from that set, which providedclues to the biological role of the protein.
ThermoFluorTM technology, as powerfulas it is, is not true calorimetry and can pro-vide only van’t Hoff enthalpy estimates. High-throughput calorimeters are still an urgentneed. The best available approach to date isone in which a sensitive DSC instrument iscoupled to a robotic system for sample han-dling (48). This approach remains limitedby serial calorimetric determinations, but therobotic system automates cell loading, data ac-quisition, and cleaning between experiments.Samples are placed in a 96-well plate and keptin a temperature-controlled environment un-til they are loaded in the calorimeter. The sys-tem runs unattended after the initial setup andallows 10 to 50 experiments to be run per day.
There are two reported attempts to pro-duce miniature calorimeters using microscaletechnology. An enthalpy array device allowsmixing of two 250 nanoliter drops, facili-tating the measurement of heats of bind-ing or of enzymatic reactions (65). Arraysof such reactors could be fabricated, al-lowing parallel enthalpy measurements fortrue high-throughput screening. A microma-chined thermoelectric flow calorimeter witha sample volume of 5 microliters was de-scribed (69), a design that would be amenableto the fabrication of an array for parallel dataacquisition. Although both of these devices
140 Chaires
Ann
u. R
ev. B
ioph
ys. 2
008.
37:1
35-1
51. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsite
Lou
is P
aste
ur -
U. S
tras
bour
g 1,
2 an
d 3
on 0
5/20
/08.
For
per
sona
l use
onl
y.

ANRV343-BB37-07 ARI 24 April 2008 15:14
represent promising initial steps toward high-throughput calorimetry and can operate withgreatly reduced sample volumes, their perfor-mance and sensitivity appear to be mediocreat best when compared with existing commer-cial microcalorimeters.
CASE STUDIES: BINDINGTO NUCLEIC ACIDS
Isostructural Is Not Isoenergetic
The anthracycline antibiotics doxorubicinand daunorubicin are mainstays of cancerchemotherapy, having been in clinical usefor over three decades (52, 75). The twocompounds differ only by the presence ofa hydroxyl group present in doxorubicin atthe C14 position. Doxorubicin binds moretightly to DNA than does daunorubicin,with about 1 kcal mol−1 more favorablebinding free energy (11). These compoundspresent a quandary for structure-based design.High-resolution X-ray crystallographic stud-ies show that doxorubicin and daunorubicinare essentially isostructural when bound totheir DNA binding site (19, 30). The molec-
ular interactions visualized in their respectivecomplexes are identical, and the added hy-droxyl group in doxorubicin does not appearto participate in any additional specific inter-actions. The structures of the two compoundsbound to DNA are shown in Figure 2. Wheredoes the additional binding free energy comefrom in the doxorubicin complex? The ther-modynamic profiles for the two compounds(Figure 2c) shed light on the problem. Whenthe binding free energies are parsed into theirenthalpic and entropic components, distinc-tive patterns are revealed (11). Doxorubicinbinding results from favorable contributionsfrom both enthalpy and entropy. In contrast,daunorubicin binding contains an unfavor-able entropic component. The distinctivethermodynamic profiles reveal subtle differ-ences not evident in their high-resolutionstructures. The difference in entropy betweentwo compounds points toward important con-tributions that are not readily visualized, per-haps differences in solvent or ion organizationthat cannot be seen even in high-resolutionstructures (30). Even though the doxoru-bicin and daunorubicin DNA complexes areisostructural, they are not isoenergetic. That
a b c
kcal mol-1
∆G
∆H
-T∆S
-8 -6 -4 -2 0 2
Doxorubicin Daunorubicin
Doxorubicin
Daunorubicin
C14 hydroxyl group
C14 hydroxyl group
Figure 2Structures of the DNA complexes of (a) doxorubicin and (b) daunorubicin. The arrows in panel a indicatethe C14 hydroxyl group present in doxorubicin. Panel c shows the thermodynamic profiles fordoxorubicin and daunorubicin binding to DNA.
www.annualreviews.org • Calorimetry and Thermodynamics in Drug Design 141
Ann
u. R
ev. B
ioph
ys. 2
008.
37:1
35-1
51. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsite
Lou
is P
aste
ur -
U. S
tras
bour
g 1,
2 an
d 3
on 0
5/20
/08.
For
per
sona
l use
onl
y.
ANRV343-BB37-07 ARI 24 April 2008 15:14
Groove binding Intercalation
Groove binders Intercalators
-20 -15 -10 -5 0 5 10 15
-30
-25
-20
-15
-10
-5
0
5
10
∆H
, kca
l mo
l-1
-T∆S, kcal mol-1
a
b
c
Hoechst 33258 Daunorubicin
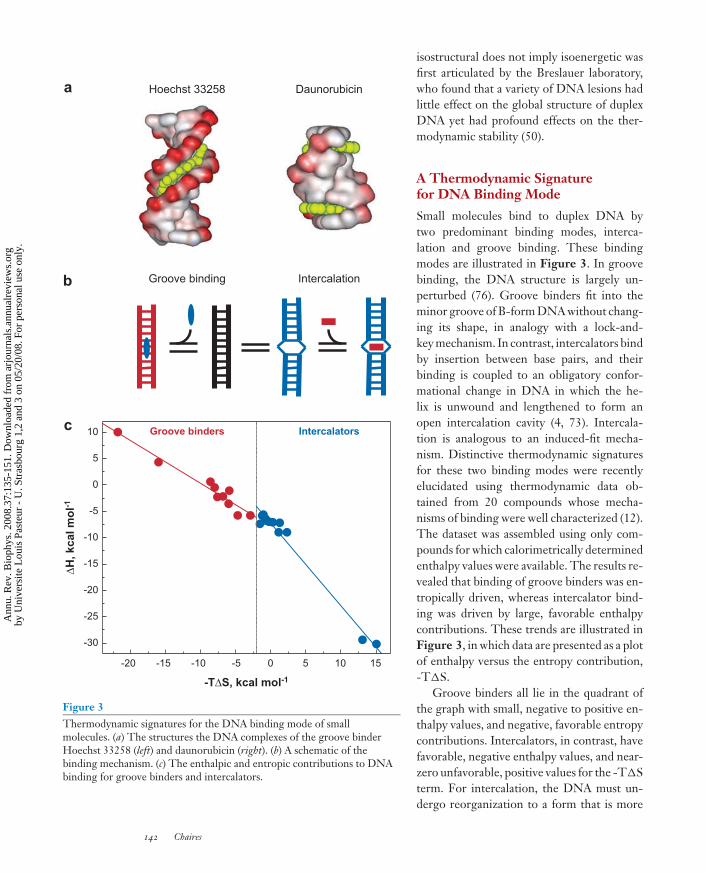
Figure 3Thermodynamic signatures for the DNA binding mode of smallmolecules. (a) The structures the DNA complexes of the groove binderHoechst 33258 (left) and daunorubicin (right). (b) A schematic of thebinding mechanism. (c) The enthalpic and entropic contributions to DNAbinding for groove binders and intercalators.
isostructural does not imply isoenergetic wasfirst articulated by the Breslauer laboratory,who found that a variety of DNA lesions hadlittle effect on the global structure of duplexDNA yet had profound effects on the ther-modynamic stability (50).
A Thermodynamic Signaturefor DNA Binding Mode
Small molecules bind to duplex DNA bytwo predominant binding modes, interca-lation and groove binding. These bindingmodes are illustrated in Figure 3. In groovebinding, the DNA structure is largely un-perturbed (76). Groove binders fit into theminor groove of B-form DNA without chang-ing its shape, in analogy with a lock-and-key mechanism. In contrast, intercalators bindby insertion between base pairs, and theirbinding is coupled to an obligatory confor-mational change in DNA in which the he-lix is unwound and lengthened to form anopen intercalation cavity (4, 73). Intercala-tion is analogous to an induced-fit mecha-nism. Distinctive thermodynamic signaturesfor these two binding modes were recentlyelucidated using thermodynamic data ob-tained from 20 compounds whose mecha-nisms of binding were well characterized (12).The dataset was assembled using only com-pounds for which calorimetrically determinedenthalpy values were available. The results re-vealed that binding of groove binders was en-tropically driven, whereas intercalator bind-ing was driven by large, favorable enthalpycontributions. These trends are illustrated inFigure 3, in which data are presented as a plotof enthalpy versus the entropy contribution,-T�S.
Groove binders all lie in the quadrant ofthe graph with small, negative to positive en-thalpy values, and negative, favorable entropycontributions. Intercalators, in contrast, havefavorable, negative enthalpy values, and near-zero unfavorable, positive values for the -T�Sterm. For intercalation, the DNA must un-dergo reorganization to a form that is more
142 Chaires
Ann
u. R
ev. B
ioph
ys. 2
008.
37:1
35-1
51. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsite
Lou
is P
aste
ur -
U. S
tras
bour
g 1,
2 an
d 3
on 0
5/20
/08.
For
per
sona
l use
onl
y.

ANRV343-BB37-07 ARI 24 April 2008 15:14
rigid and entropically less favorable. Theentropic penalty must be overcome by en-thalpy, arising at least in part from favorablevan der Waals stacking interactions betweenthe base pairs and the intercalated chro-mophore. For the case of DNA binding pro-teins, a different trend was elucidated (34).Proteins that bend or distort DNA were en-tropically driven, whereas proteins that boundwithout major alteration of the DNA struc-ture were enthalpically driven.
Measuring Ultratight DNABinding by DSC
One advantage of DSC is its capability tomeasure association constants for ultratightbinding reactions (7). Thermal denaturationmethods offer a particularly powerful ap-proach for studying both the specificity andaffinity of nucleic acid binding ligands (58,59). We exploited this capability in studies of anew class of bisintercalating anthracycline an-tibiotics (31, 37, 51, 53). The novel bisanthra-cyclines were designed on the basis of struc-tures such as those shown in Figure 2 (13). Inthose structures, anthracycline monomers areseen to bind to DNA in a tail-to-tail orien-tation, with reactive amine groups separatedby only 7 angstroms in the minor groove.The simple design strategy was to covalentlylink those amine groups with a moiety thatwould fit precisely into the minor groove andwould not interfere with intercalation of theanthraquinone rings between base pairs or ofthe fit of the daunosamine moiety in the minorgroove. The structure of one such bisinterca-lating anthracycline is shown in Figure 4 (51).The binding affinity of such a bisintercalatorwas expected to be approximately the squareof the binding constant of the monomer (17).Because the binding constant of daunorubicinis approximately 106 M−1, bisdaunorubicinwould be expected to bind with picomolaraffinity. For such tight binding, traditionalspectroscopic approaches to measuring bind-ing become prohibitively difficult for a varietyof reasons, chief among them is the difficulty
a
b c
40 50 60 70 80 90 100 110
250
500
750
1000
1250
1500
∆H2
Exc
ess
hea
t ca
pac
ity
(kca
l mo
l-1)
Temperature (°C)
∆Tm
∆H1
Daunorubicin
WP631
WP762
-30 -20 -10 0 10 20
kcal mol-1
BisanthracyclineWP762 saturated
BisanthracyclineWP762 unsaturated
Freeenergy
Enthalpy
-T∆SG1G1
G3G3
T4T4
A5A5
C6C6
G7G7
C8C8
C10C10
G9G9
G15G15
C14C14
A13A13
T12T12
i11i11
C2C2
C16C16
G1
G3
T4
A5
C6
G7
C8
C10
G9
G15
C14
A13
T12
i11
C2
C16
Figure 4Ultratight binding of a bisanthracycline to DNA. (a) A schematic of theDSC thermograms observed for DNA in the absence and presence ofsaturating amounts of the bisanthracycline WP762. (b) The structure of theWP762-DNA complex. (c) The thermodynamic profiles for the binding toDNA of the parent monomer daunorubicin and two bisanthracyclines,WP631 and WP762, which differ in their linker geometry. Free energy,enthalpy, and -T�S are shown.
of working at low enough ligand concentra-tions to avoid stoichiometric binding yet stillretaining a measurable signal. A number ofwell-developed statistical mechanical theoriesshow that binding constants can be reliably
www.annualreviews.org • Calorimetry and Thermodynamics in Drug Design 143
Ann
u. R
ev. B
ioph
ys. 2
008.
37:1
35-1
51. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsite
Lou
is P
aste
ur -
U. S
tras
bour
g 1,
2 an
d 3
on 0
5/20
/08.
For
per
sona
l use
onl
y.

ANRV343-BB37-07 ARI 24 April 2008 15:14
measured from shifts in melting temperaturesof the receptor macromolecule (7, 16, 41, 57),although caution is needed in the applicationof these approaches (71).
Figure 4 shows the application of that ap-proach to the characterization of bisanthra-cycline binding to DNA. Upon binding, theDSC thermogram is shifted upward by over30◦. That Tm shift may be used to calculatea binding constant for the bisanthracycline atthe elevated Tm. DSC offers the added ad-vantage that the binding enthalpy may also beestimated from the difference in the areas ofthe thermograms in the presence and absenceof bound ligand. The enthalpy obtained fromthe area of the thermogram for DNA alone,along with the total enthalpy for melting ofthe bisanthracycline-DNA complex may beused with application of Hess’s law to inferthe enthalpy of the bisanthracycline bindingreaction. A complete thermodynamic profilemay then be derived for the binding reaction.
Comparative binding profiles are shownin Figure 4 for daunorubicin and two bis-daunorubicins that differ only in their linkergeometry. The binding free energy for thebisdaunorubicin molecules is nearly double
Indinavir
Nelfinavir
Saquinavir
Ritonavir
KNI-764
-16 -14 -12 -10 -8 -6 -4 -2 0 2 4
kcal mol-1
Free energy
Enthalpy-T∆S
Figure 5Enthalpy optimization of HIV protease inhibitors. Thermodynamicprofiles for the binding of first- and second-generation (KNI-764) HIVprotease inhibitors. Free energy, enthalpy, and -T�S are shown.
that of the parent monomer, as predicted andintended. The thermodynamic profiles, how-ever, show that the bisanthracyclines sufferfrom a large, unfavorable entropic contribu-tion. The source of this unfavorable entropy islikely due to restriction of rotation around thelinker bonds upon binding (44). The thermo-dynamic data suggest an improved design inwhich rotation around the linker bonds is re-stricted, while retaining the correct geometryfor bisintercalation into DNA. If such a designwere to be realized, binding affinity would bepredicted to increase from the current pico-molar range to femtomolar or greater.
CASE STUDIES: PROTEINBINDING INTERACTIONS
Optimizing Affinity in HIV-1Protease Inhibitors
HIV protease inhibitors are competitive in-hibitors, so their potency is directly correlatedwith their binding affinity. The Freire labora-tory has pursued a program to optimize HIVinhibitor binding by consideration of the ther-modynamics of the binding interactions (45,46, 67, 68). Their starting point is the real-ization that the binding free energy is a func-tion of both enthalpy and entropy, and thatmany combinations of �H and T�S valuescan yield the same value of �G. Enthalpicallyor entropically optimized inhibitors, however,might have different specificities or differ-ent pharmacological properties, even thoughtheir interaction with the target protease mayhave similar free energies. Enthalpic contri-butions reflect mostly the strength of inhibitorbonding interactions with protease (hydrogenbonds; van der Waals interactions) relative tosolvent bonding. Entropic contributions tothe free energy reflect mostly changes in sol-vent entropy arising from the hydrophobictransfer of inhibitor to its binding site.
Figure 5 shows thermodynamic pro-files for the first-generation HIV proteaseinhibitors indinavir, nelfinavir, saquinavir,and ritonavir. For all of these, binding is
144 Chaires
Ann
u. R
ev. B
ioph
ys. 2
008.
37:1
35-1
51. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsite
Lou
is P
aste
ur -
U. S
tras
bour
g 1,
2 an
d 3
on 0
5/20
/08.
For
per
sona
l use
onl
y.

ANRV343-BB37-07 ARI 24 April 2008 15:14
entropically driven, and for three of the four,binding is accompanied by an unfavorablepositive enthalpy change. Because these in-hibitors are conformationally constrained, thebinding entropy reflects primarily a large sol-vation entropy gain due to a hydrophobictransfer. Differences in the thermodynamicbinding profile of the newer-generation in-hibitors are evident, as shown by one exam-ple in Figure 5. KNI-764 binds more tightlythan any of the first-generation inhibitors, byat least 1 kcal mol−1 in free energy. The fa-vorable free energy results from almost equalfavorable enthalpic and entropic contribu-tions. KNI-764 thus has improved bondinginteractions with the protease. Analysis of thethermodynamic profiles of a series of HIVprotease inhibitors led to the conclusion thatextremely high binding affinity requires a fa-vorable binding enthalpy, and to the con-cept of enthalpic optimization as a drug de-sign principle (46). Optimal binding enthalpydoes not correlate simply with the number ofhydrogen bond donors or acceptors, but in-stead results from the quality of the hydrogenbond formed within the binding pocket rel-ative to inhibitor-solvent hydrogen bondinginteractions.
The Enthalpy Funnel
The value of enthalpy optimization was rein-forced by additional studies from the Freirelaboratory on inhibitors of plasmepsin II, ahemoglobin-degrading enzyme that is a keycomponent in the life cycle of the Plasmodiumparasites responsible for malaria (56). Plas-mepsin II represents a novel target for anti-malarial drug development. A lead compoundwas developed, based on an allophenylnorsta-tine scaffold that mimics the main cleavagesite in the hemoglobin molecule of infectedvictims. Compounds evolved from the leadthat optimized the contribution of the en-thalpy to the binding free energy. The affin-ity of 71 allophenylnorstatine inhibitors wasmeasured and parsed into enthalpic and en-tropic components, with the results shown in
5
6
7
8
9
10
-0.25 0.00 0.25 0.50 0.75 1.00
∆H/∆G
log
Ka
Figure 6The enthalpy funnel. Thermodynamic data for the binding of 71plasmepsin inhibitors are shown. The ratio of the binding enthalpy to freeenergy is shown as a function of the logarithm of the association constant.
Figure 6. The results define a rather dramaticand distinctive enthalpy funnel, in whichthe enthalpy/entropy combinations narrow asaffinity (expressed as log Ka) increases. Theoptimal balance in this case appears to arisewith the free energy partitioned as one-thirdenthalpy and two-thirds entropy. Being mind-ful of the need to achieve such a thermody-namic balance throughout the optimizationprocess is crucial for a successful design effort.
CASE STUDY: HIGHSELECTIVITY DOES NOTDEMAND HIGH AFFINITY
Although high affinity is often a major goal indrug design, it is not an absolute requirementfor selectivity and specificity. It is instructiveto review the results obtained from a DNAaptamer that illustrates this point. A DNAaptamer that recognized l-argininamide wasdiscovered (24) by the SELEX (SystematicEvolution of Ligands by Exponential en-richment system) (21) approach. The ap-tamer was approximately 100-fold more se-lective for l-argininamide over several otherarginine analogs and other amino acids. A
www.annualreviews.org • Calorimetry and Thermodynamics in Drug Design 145
Ann
u. R
ev. B
ioph
ys. 2
008.
37:1
35-1
51. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsite
Lou
is P
aste
ur -
U. S
tras
bour
g 1,
2 an
d 3
on 0
5/20
/08.
For
per
sona
l use
onl
y.

ANRV343-BB37-07 ARI 24 April 2008 15:14
high-resolution NMR structure of a truncatedversion of the aptamer showed that a largeportion of the DNA was unstructured in theabsence of argininamide, but in the presenceof argininamide a highly ordered structureformed in which the amino acid was encapsu-lated into a unique DNA fold (38). Our labo-
Figure 7Binding of l-argininamide to its DNA aptamer. (a) The structures of the(left) ligated and (right) unligated DNA aptamers. (b) Thermodynamicprofiles (i) for the binding of l-argininamide to the aptamer, (ii) forethidium intercalation into DNA, (iii) for daunorubicin intercalation intoDNA, and (iv) for the groove binding of Hoechst 33258 to DNA.
ratory attempted the first detailed thermody-namic characterization of aptamer binding forthis system, coupled with molecular dynamicssimulations (5).
Figure 7 shows the structures of the lig-ated and unligated argininamide aptamer. Inthe absence of argininamide, our thermody-namic and molecular dynamics simulationsindicated that the DNA formed a classichairpin-loop structure, with the loop contain-ing 10 unpaired bases. The highly selectivebinding of l-argininamide to the DNA had afree energy change of only −5.1 kcal mol−1,corresponding to an association constant ofonly 6000 M−1. Parsing the free energy intoits component enthalpy and entropy contri-butions showed that binding was accompa-nied by a large, favorable enthalpy of −8.7 to−9.2 kcal mol−1. Binding was opposed by anunfavorable -T�S contribution of +3.6 to+4.1 kcal mol−1 (Figure 7). This case rep-resents a clear illustration of an induced-fitmechanism, in which binding is tightly cou-pled to a large conformational change in thereceptor DNA. The otherwise disordered anddynamic loop structure undergoes a majorconformational change to form a stable bind-ing pocket to which the argininamide is an-chored. Such ordering is entropically unfavor-able, as was measured, and must be overcomeby specific bonding interactions that are en-thalpically favorable. The opposing enthalpyand entropy contributions yield a modest fa-vorable free energy change. High specificityresults, without high affinity.
Figure 7 shows a progression in the ther-modynamic profiles of small molecules thatbind to DNA. The progression begins with anentropically driven groove-binder that doesnot perturb the B-DNA structure and thatbinds with positive enthalpy. Two DNA in-tercalators with different unwinding anglesshow proportionally larger unfavorable en-tropic contributions, with roughly equal bind-ing enthalpy values. Finally, the argininamideaptamer with its massive conformationalchange shows the largest opposing entropycontribution.
146 Chaires
Ann
u. R
ev. B
ioph
ys. 2
008.
37:1
35-1
51. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsite
Lou
is P
aste
ur -
U. S
tras
bour
g 1,
2 an
d 3
on 0
5/20
/08.
For
per
sona
l use
onl
y.

ANRV343-BB37-07 ARI 24 April 2008 15:14
CONCLUSIONS
These few examples illustrate how thermo-dynamic data complement structural data, ifthey exist for a particular system. The ther-modynamic profile shows the enthalpic and
entropic contributions to the binding free en-ergy and indicates the nature of forces thatdrive complex formation. Such informationcan guide drug development by suggesting thenature of modification that could most opti-mize binding to the macromolecular target.
SUMMARY POINTS
1. Thermodynamic data are an essential complement to structural data in drug devel-opment and for the optimization of lead compounds.
2. A complete thermodynamic profile for a binding interaction includes the binding freeenergy (�G), enthalpy (�H), entropy (�S), and heat capacity change (�Cp).
3. The thermodynamic profile indicates the predominant forces that drive the bindinginteraction.
4. Isostructural complexes need not be isoenergetic.
5. Favorable enthalpy contributions arise from favorable hydrogen bonding and vander Waals interactions, and favorable entropic contributions arise primarily fromhydrophobic interactions and desolvation.
6. Thermal denaturation methods provide valuable tools for rapid screening of the bind-ing compound libraries and for quantitative measurement of ultratight binding inter-actions.
7. High specificity does not demand high affinity.
FUTURE ISSUES
1 Full integration of calorimetry and thermodynamics into the drug discovery processrequires development of high-throughput calorimeters to facilitate the acquisition ofquantitative data.
2. A more fundamental thermodynamic understanding of the molecular interactions(e.g., hydrogen bonds, van der Waals forces, hydrophobic forces, water and ion bind-ing interactions, electrostatic forces) that govern noncovalent binding of drugs totheir receptor targets is urgently needed.
DISCLOSURE STATEMENT
The author is not aware of any biases that might be perceived as affecting the objectivity ofthis review.
ACKNOWLEDGMENTS
Work in my laboratory is supported by grants from the National Cancer Institute (CA35635),the National Institute for General Medical Sciences (GM077422), and the James GrahamBrown Foundation.
www.annualreviews.org • Calorimetry and Thermodynamics in Drug Design 147
Ann
u. R
ev. B
ioph
ys. 2
008.
37:1
35-1
51. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsite
Lou
is P
aste
ur -
U. S
tras
bour
g 1,
2 an
d 3
on 0
5/20
/08.
For
per
sona
l use
onl
y.

ANRV343-BB37-07 ARI 24 April 2008 15:14
LITERATURE CITED
1. Ackers GK, Bolen DW. 1997. The Gibbs conference on biothermodynamics: origins andevolution. Biophys. Chem. 64:3–5
2. Amzel LM. 1997. Loss of translational entropy in binding, folding, and catalysis. Proteins28:144–49
3. Amzel LM. 2000. Calculation of entropy changes in biological processes: folding, binding,and oligomerization. Methods Enzymol. 323:167–77
4. Berman HM, Young PR. 1981. The interaction of intercalating drugs with nucleic acids.Annu. Rev. Biophys. Bioeng. 10:87–114
5. Bishop GR, Ren J, Polander BC, Jeanfreau BD, Trent JO, Chaires JB. 2007. Energeticbasis of molecular recognition in a DNA aptamer. Biophys. Chem. 126:165–75
6. Brady GP, Sharp KA. 1997. Entropy in protein folding and in protein-protein interactions.Curr. Opin. Struct. Biol. 7:215–21
7. Brandts JF, Lin L-N. 1990. Study of strong to ultratight protein interactions using differ-ential scanning calorimetry. Biochemistry 29:6927–40
8. Carver TE, Bordeau B, Cummings MD, Petrella EC, Pucci MJ, et al. 2005. Decrypting thebiochemical function of an essential gene from Streptococcus pneumoniae using ThermoFluortechnology. J. Biol. Chem. 280:11704–12
9. Chaires JB. 1997. Energetics of drug-DNA interactions. Biopolymers 44:201–1510. Chaires JB. 1997. Possible origin of differences between van’t Hoff and calorimetric en-
thalpy estimates. Biophys. Chem. 64:15–2311. Chaires JB. 2003. Energetics of anthracycline-DNA interactions. In DNA and RNA Binders,
ed. M Demeunynck, C Bailly, WD Wilson, pp. 461–81. Weinheim: Wiley-VCH12. Chaires JB. 2006. A thermodynamic signature for drug-DNA binding mode. Arch. Biochem.
Biophys. 453:26–3113. Chaires JB, Leng F, Przewloka T, Fokt I, Ling YH, et al. 1997. Structure-based design of
a new bisintercalating anthracycline antibiotic. J. Med. Chem. 40:261–6614. Cooper A. 1999. Thermodynamic analysis of biomolecular interactions. Curr. Opin. Chem.
Biol. 3:557–6315. Cooper A, Johnson CM. 1994. Introduction to microcalorimetry and biomolecular ener-
getics. Methods Mol. Biol. 22:109–2416. Crothers DM. 1971. Statistical thermodynamics of nucleic acid melting transitions with
coupled binding equilibria. Biopolymers 10:2147–6017. Crothers DM, Metzger H. 1972. The influence of polyvalency on the binding properties
of antibodies. Immunochemistry 9:341–5718. Fisher HF, Singh N. 1995. Calorimetric methods for interpreting protein-ligand interac-
tions. Methods Enzymol. 259:194–22119. Frederick CA, Williams LD, Ughetto G, Van Der Marel GA, van Boom JH, et al. 1990.
Structural comparison of anticancer drug-DNA complexes: adriamycin and daunomycin.Biochemistry 29:2538–49
20. Gilson MK, Given JA, Bush BL, McCammon JA. 1997. The statistical-thermodynamicbasis for computation of binding affinities: a critical review. Biophys. J. 72:1047–69
21. Gold L. 1995. The SELEX process: a surprising source of therapeutic and diagnosticcompounds. Harvey Lect. 91:47–57
22. Haq I, Jenkins TC, Chowdhry BZ, Ren J, Chaires JB. 2000. Parsing the free energy ofdrug-DNA interactions. Methods Enzymol. 323:373–405
23. Haq I, Ladbury JE, Chowdhry BZ, Jenkins TC, Chaires JB. 1997. Specific binding ofHoechst 33258 to the d(CGCAAATTTGCG)2 duplex: calorimetric and spectroscopicstudies. J. Mol. Biol. 271:244–57
148 Chaires
Ann
u. R
ev. B
ioph
ys. 2
008.
37:1
35-1
51. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsite
Lou
is P
aste
ur -
U. S
tras
bour
g 1,
2 an
d 3
on 0
5/20
/08.
For
per
sona
l use
onl
y.

ANRV343-BB37-07 ARI 24 April 2008 15:14
24. Harada K, Frankel AD. 1995. Identification of two novel arginine binding DNAs. EMBOJ. 14:5798–811
25. Henry CM. 2001. Structure-based drug design. Chem. Eng. News 79:69–7426. Hinz H-J, Schwarz FP. 2001. Measurement and analysis of results obtained on biological
substances with differential scanning calorimetry. Pure Appl. Chem. 73:745–5927. Holdgate GA. 2001. Making cool drugs hot: isothermal titration calorimetry as a tool to
study binding energetics. Biotechniques 31:164–6628. Holdgate GA, Ward WH. 2005. Measurements of binding thermodynamics in drug dis-
covery. Drug Discov. Today 10:1543–5029. Horn JR, Russell D, Lewis EA, Murphy KP. 2001. Van’t Hoff and calorimetric enthalpies
from isothermal titration calorimetry: Are there significant discrepancies? Biochemistry40:1774–78
30. Howerton SB, Nagpal A, Williams LD. 2003. Surprising roles of electrostatic interactionsin DNA-ligand complexes. Biopolymers 69:87–99
31. Hu GG, Shui X, Leng F, Priebe W, Chaires JB, Williams LD. 1997. Structure of a DNA-bisdaunomycin complex. Biochemistry 36:5940–46
32. Indyk L, Fisher HF. 1998. Theoretical aspects of isothermal titration calorimetry. MethodsEnzymol. 295:350–64
33. Janin J. 1995. Elusive affinities. Proteins 21:30–3934. Jen-Jacobson L, Engler LE, Jacobson LA. 2000. Structural and thermodynamic strategies
for site-specific DNA binding proteins. Structure 8:1015–2335. Lavoisier AL, Laplace PS. 1783. Memoir on Heat. Transl. H Guerlac, 1982, New York:
Neale Watson (From French)36. Lavoisier AL, Laplace PS. 1784. Memoire sur la chaleur. In Memoir Academie Royale Scien-
tifique 1784, pp. 355–40837. Leng F, Priebe W, Chaires JB. 1998. Ultratight DNA binding of a new bisintercalating
anthracycline antibiotic. Biochemistry 37:1743–5338. Lin CH, Patel DJ. 1996. Encapsulating an amino acid in a DNA fold. Nat. Struct. Biol.
3:1046–5039. Liu Y, Sturtevant JM. 1995. Significant discrepancies between van’t Hoff and calorimetric
enthalpies. II. Protein Sci. 4:2559–6140. Lodwig TH, Smeaton WA. 1974. The ice calorimeter of Lavoisier and Laplace and some
of its critics. Ann. Sci. 31:1–1841. McGhee JD. 1976. Theoretical calculations of the helix-coil transition of DNA in the
presence of large, cooperatively binding ligands. Biopolymers 15:1345–7542. Murphy KP, Xie D, Thompson KS, Amzel LM, Freire E. 1994. Entropy in biological
binding processes: estimation of translational entropy loss. Proteins 18:63–6743. Naghibi H, Tamura A, Sturtevant JM. 1995. Significant discrepancies between van’t Hoff
and calorimetric enthalpies. Proc. Natl. Acad. Sci. USA 92:5597–9944. Novotny J, Bruccoleri RE, Davis M, Sharp KA. 1997. Empirical free energy calculations:
a blind test and further improvements to the method. J. Mol. Biol. 268:401–1145. Ohtaka H, Freire E. 2005. Adaptive inhibitors of the HIV-1 protease. Prog. Biophys. Mol.
Biol. 88:193–208
46. Articulation ofthe enthalpicoptimizationconcept.
46. Ohtaka H, Muzammil S, Schon A, Velazquez-Campoy A, Vega S, Freire E. 2004.Thermodynamic rules for the design of high affinity HIV-1 protease inhibitorswith adaptability to mutations and high selectivity towards unwanted targets. Int. J.
Biochem. Cell Biol. 36:1787–99
www.annualreviews.org • Calorimetry and Thermodynamics in Drug Design 149
Ann
u. R
ev. B
ioph
ys. 2
008.
37:1
35-1
51. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsite
Lou
is P
aste
ur -
U. S
tras
bour
g 1,
2 an
d 3
on 0
5/20
/08.
For
per
sona
l use
onl
y.

ANRV343-BB37-07 ARI 24 April 2008 15:14
47. A description ofThermoFluorTM
technology and itsapplication.
47. Pantoliano MW, Petrella EC, Kwasnoski JD, Lobanov VS, Myslik J, et al. 2001. High-density miniaturized thermal shift assays as a general strategy for drug discovery.J. Biomol. Screen. 6:429–40
48. Plotnikov V, Rochalski A, Brandts M, Brandts JF, Williston S, et al. 2002. An autosam-pling differential scanning calorimeter instrument for studying molecular interactions. As-say Drug Dev. Technol. 1:83–90
49. Plotnikov VV, Brandts JM, Lin LN, Brandts JF. 1997. A new ultrasensitive scanningcalorimeter. Anal. Biochem. 250:237–44
50. Plum GE, Breslauer KJ. 1994. DNA lesions. A thermodynamic perspective. Ann. N. Y.Acad. Sci. 726:45–55
51. Portugal J, Cashman DJ, Trent JO, Ferrer-Miralles N, Przewloka T, et al. 2005. A new bis-intercalating anthracycline with picomolar DNA binding affinity. J. Med. Chem. 48:8209–19
52. Priebe W, ed. 1995. Anthracycline Antibiotics: New Analogues, Methods of Delivery and Mech-anisms of Action, Vol. 574. Washington, DC: Am. Chem. Soc.
53. Priebe W, Fokt I, Przewloka T, Chaires JB, Portugal J, Trent JO. 2001. Exploiting anthra-cycline scaffold for designing DNA-targeting agents. Methods Enzymol. 340:529–55
54. Privalov G, Kavina V, Freire E, Privalov PL. 1995. Precise scanning calorimeter for study-ing thermal properties of biological macromolecules in dilute solution. Anal. Biochem.232:79–85
55. Ren J, Jenkins TC, Chaires JB. 2000. Energetics of intercalation reactions. Biochemistry39:8439–47
56. Presentation ofthe enthalpy funnelconcept.
56. Ruben AJ, Kiso Y, Freire E. 2006. Overcoming roadblocks in lead optimization: athermodynamic perspective. Chem. Biol. Drug. Des. 67:2–4
57. Schellman JA. 1958. The factors affecting the stability of hydrogen-bonded polypeptidestructures in solution. J. Phys. Chem. 62:1485–94
58. Shi X, Chaires JB. 2006. Sequence- and structural-selective nucleic acid binding revealedby the melting of mixtures. Nucleic Acids Res. 34:e14
59. Shi X, Chaires JB. 2006. Thermal denaturation of drug-DNA complexes: tools and tricks.In Sequence-Specific DNA Binding Agents, ed. M Waring, pp. 130–51. Cambridge, UK: RSCPubl.
60. Spolar RS, Record MT Jr. 1994. Coupling of local folding to site-specific binding of proteinsto DNA. Science 263:777–84
61. Tellinghuisen J. 2003. A study of statistical error in isothermal titration calorimetry. Anal.Biochem. 321:79–88
62. Tellinghuisen J. 2004. Statistical error in isothermal titration calorimetry. Methods Enzymol.383:245–82
63. Tellinghuisen J. 2004. Volume errors in isothermal titration calorimetry. Anal. Biochem.333:405–6
64. Tellinghuisen J. 2005. Statistical error in isothermal titration calorimetry: variance functionestimation from generalized least squares. Anal. Biochem. 343:106–15
65. Torres FE, Kuhn P, De Bruyker D, Bell AG, Wolkin MV, et al. 2004. Enthalpy arrays.Proc. Natl. Acad. Sci. USA 101:9517–22
66. Velazquez Campoy A, Freire E. 2005. ITC in the postgenomic era. . .? Priceless. Biophys.Chem. 115:115–24
67. Velazquez-Campoy A, Luque I, Todd MJ, Milutinovich M, Kiso Y, Freire E. 2000. Ther-modynamic dissection of the binding energetics of KNI-272, a potent HIV-1 proteaseinhibitor. Protein Sci. 9:1801–9
150 Chaires
Ann
u. R
ev. B
ioph
ys. 2
008.
37:1
35-1
51. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsite
Lou
is P
aste
ur -
U. S
tras
bour
g 1,
2 an
d 3
on 0
5/20
/08.
For
per
sona
l use
onl
y.

ANRV343-BB37-07 ARI 24 April 2008 15:14
68. Velazquez-Campoy A, Todd MJ, Freire E. 2000. HIV-1 protease inhibitors: enthalpic versusentropic optimization of the binding affinity. Biochemistry 39:2201–7
69. Verhaegen K, Van Gerven P, Baert K, Hermans L, Mertens R, Luyten W. 1998. Designof a high-throughput microphysiometer. In Biocalorimetry: Applications of Calorimetry in theBiological Sciences, ed. JE Ladbury, BZ Chowdhry, pp. 227–31. New York: Wiley
70. Wadso I. 1997. Neither calorimeters nor colorimetrists are what they used to be.Thermochim. Acta 300:1–5
71. Warnings aboutthe proper use ofthermaldenaturationmethods todetermine bindingenergetics.
71. Waldron TT, Murphy KP. 2003. Stabilization of proteins by ligand binding: appli-cation to drug screening and determination of unfolding energetics. Biochemistry
42:5058–6472. Ward WH, Holdgate GA. 2001. Isothermal titration calorimetry in drug discovery. Prog.
Med. Chem. 38:309–7673. Waring MJ. 1981. DNA modification and cancer. Annu. Rev. Biochem. 50:159–92
74. A useful concisesummary of therole of calorimetryin drug discovery.
74. Weber PC, Salemme FR. 2003. Applications of calorimetric methods to drug dis-covery and the study of protein interactions. Curr. Opin. Struct. Biol. 13:115–21
75. Weiss RB. 1992. The anthracyclines: Will we ever find a better doxorubicin? Semin. Oncol.19:670–86
76. Wemmer DE. 2000. Designed sequence-specific minor groove ligands. Annu. Rev. Biophys.Biomol. Struct. 29:439–61
77. Wiseman T, Williston S, Brandts JF, Lin LN. 1989. Rapid measurement of binding con-stants and heats of binding using a new titration calorimeter. Anal. Biochem. 179:131–37
RELATED RESOURCES
Hinz HJ. 1983. Thermodynamics of protein-ligand interactions: calorimetric approaches.Annu. Rev. Biophys. Bioeng. 12:285–317
Jelesarov I, Bosshard HR. 1999. Isothermal titration calorimetry and differential scanningcalorimetry as complementary tools to investigate the energetics of biomolecular recog-nition. J. Mol. Recognit. 12:3–18
Ladbury JE, Chowdhry BZ, eds. 1998. Biocalorimetry: Applications of Calorimetry in the BiologicalSciences. New York: Wiley
Luque I, Leavitt SA, Freire E. 2002. The linkage between protein folding and functionalcooperativity: two sides of the same coin? Annu. Rev. Biophys. Biomol. Struct. 31:235–56
Sturtevant JM. 1974. Some applications of calorimetry in biochemistry and biology. Annu. Rev.Biophys. Bioeng. 3:35–51
Sturtevant JM. 1987. Biochemical applications of differential scanning calorimetry. Annu. Rev.Phys. Chem. 38:463–88
www.annualreviews.org • Calorimetry and Thermodynamics in Drug Design 151
Ann
u. R
ev. B
ioph
ys. 2
008.
37:1
35-1
51. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsite
Lou
is P
aste
ur -
U. S
tras
bour
g 1,
2 an
d 3
on 0
5/20
/08.
For
per
sona
l use
onl
y.

AR343-FM ARI 10 April 2008 7:2
Annual Review ofBiophysics
Volume 37, 2008
Contents
FrontispieceRobert L. Baldwin � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �xiv
The Search for Folding Intermediates and the Mechanismof Protein FoldingRobert L. Baldwin � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �1
How Translocons Select Transmembrane HelicesStephen H. White and Gunnar von Heijne � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 23
Unique Rotary ATP Synthase and Its Biological DiversityChristoph von Ballmoos, Gregory M. Cook, and Peter Dimroth � � � � � � � � � � � � � � � � � � � � � � � � 43
Mediation, Modulation, and Consequencesof Membrane-Cytoskeleton InteractionsGary J. Doherty and Harvey T. McMahon � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 65
Metal Binding Affinity and Selectivity in Metalloproteins:Insights from Computational StudiesTodor Dudev and Carmay Lim � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 97
Riboswitches: Emerging Themes in RNA Structure and FunctionRebecca K. Montange and Robert T. Batey � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �117
Calorimetry and Thermodynamics in Drug DesignJonathan B. Chaires � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �135
Protein Design by Directed EvolutionChristian Jäckel, Peter Kast, and Donald Hilvert � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �153
PIP2 Is A Necessary Cofactor for Ion Channel Function:How and Why?Byung-Chang Suh and Bertil Hille � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �175
RNA Folding: Conformational Statistics, Folding Kinetics,and Ion ElectrostaticsShi-Jie Chen � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �197
Intrinsically Disordered Proteins in Human Diseases: Introducingthe D2 ConceptVladimir N. Uversky, Christopher J. Oldfield, and A. Keith Dunker � � � � � � � � � � � � � � � �215
Crowding Effects on Diffusion in Solutions and CellsJames A. Dix and A.S. Verkman � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �247
vii
Ann
u. R
ev. B
ioph
ys. 2
008.
37:1
35-1
51. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsite
Lou
is P
aste
ur -
U. S
tras
bour
g 1,
2 an
d 3
on 0
5/20
/08.
For
per
sona
l use
onl
y.

AR343-FM ARI 10 April 2008 7:2
Nanobiotechnology and Cell Biology: Micro- and NanofabricatedSurfaces to Investigate Receptor-Mediated SignalingAlexis J. Torres, Min Wu, David Holowka, and Barbara Baird � � � � � � � � � � � � � � � � � � � � � �265
The Protein Folding ProblemKen A. Dill, S. Banu Ozkan, M. Scott Shell, and Thomas R. Weikl � � � � � � � � � � � � � � � � � �289
Translocation and Unwinding Mechanisms of RNAand DNA HelicasesAnna Marie Pyle � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �317
Structure of Eukaryotic RNA PolymerasesP. Cramer, K.-J. Armache, S. Baumli, S. Benkert, F. Brueckner, C. Buchen,G.E. Damsma, S. Dengl, S.R. Geiger, A.J. Jasiak, A. Jawhari, S. Jennebach,T. Kamenski, H. Kettenberger, C.-D. Kuhn, E. Lehmann, K. Leike, J.F. Sydow,and A. Vannini � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �337
Structure-Based View of Epidermal Growth Factor ReceptorRegulationKathryn M. Ferguson � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �353
Macromolecular Crowding and Confinement: Biochemical,Biophysical, and Potential Physiological ConsequencesHuan-Xiang Zhou, Germán Rivas, and Allen P. Minton � � � � � � � � � � � � � � � � � � � � � � � � � � � � �375
Biophysics of Catch BondsWendy E. Thomas, Viola Vogel, and Evgeni Sokurenko � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �399
Single-Molecule Approach to Molecular Biology in Living BacterialCellsX. Sunney Xie, Paul J. Choi, Gene-Wei Li, Nam Ki Lee, and Giuseppe Lia � � � � � � � � �417
Structural Principles from Large RNAsStephen R. Holbrook � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �445
Bimolecular Fluorescence Complementation (BiFC) Analysisas a Probe of Protein Interactions in Living CellsTom K. Kerppola � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �465
Multiple Routes and Structural Heterogeneity in Protein FoldingJayant B. Udgaonkar � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �489
Index
Cumulative Index of Contributing Authors, Volumes 33–37 � � � � � � � � � � � � � � � � � � � � � � � �511
Errata
An online log of corrections to Annual Review of Biophysics articles may be found athttp://biophys.annualreviews.org/errata.shtml
viii Contents
Ann
u. R
ev. B
ioph
ys. 2
008.
37:1
35-1
51. D
ownl
oade
d fr
om a
rjou
rnal
s.an
nual
revi
ews.
org
by U
nive
rsite
Lou
is P
aste
ur -
U. S
tras
bour
g 1,
2 an
d 3
on 0
5/20
/08.
For
per
sona
l use
onl
y.