carier prenatal dmd 1991
DESCRIPTION
Prenatal Diagnosis for carrier of DMDTRANSCRIPT

Am. J. Hum. Genet. 49:951-960, 1991
Carrier Detection and Prenatal Diagnosis in Duchenne andBecker Muscular Dystrophy Families, Using DinucleotideRepeat PolymorphismsP. R. Clemens,* R. G. Fenwick,* J. S. Chamberlain,§ R. A. Gibbs,* M. de AndradetR. Chakrabortyt and C. T. Caskey*,t
*Institute for Molecular Genetics and tHoward Hughes Medical Institute, Baylor College of Medicine, Houston; tGenetics Centers, Universityof Texas Graduate School of Biomedical Sciences, Houston; and §Department of Human Genetics, University of Michigan Medical School, AnnArbor
Summary
To improve carrier detection and prenatal diagnosis for Duchenne and Becker muscular dystrophy families,we determined allele frequencies and measures of variation for four (dC-dA)n * (dG-dT)n loci identified withina deletion-prone region of the human dystrophin gene. The loci are highly polymorphic, with predictedheterozygosities of 71.6%-93.3%. Direct DNA sequence analysis of the (dC-dA)n * (dG-dT)n locus in intron49 revealed an additional length polymorphism which varies by single-basepair increments, is adjacent tothe dinucleotide repeat block, and enhances the polymorphic content of this marker. The four(dC-dA)n * (dG-dT)n loci are each easily amplified by PCR in two diplex reactions. The variability of allelelengths at these loci makes them ideal for carrier detection and prenatal diagnosis, often providing diagnosticinformation when RFLP analysis is uninformative. These markers have aided in identification of deletionmutations, exclusion of maternal cell contamination of chorionic villus samples, confirmation of paternity,and mapping of gene recombinations. The allele identification of these loci can be performed either witha radiolabel or with an automated, nonradioactive, fluorescent gel detection system.
Introduction
Duchenne and Becker muscular dystrophies (D/BMD)are allelic forms of an X-linked recessive disease char-acterized by absent or abnormal dystrophin in skeletalmuscle, resulting in early muscle degeneration anddeath. DMD is a common disease that affects 1 /3,500male births, which has led to an emphasis on the devel-opment of simplified tools for carrier detection andprenatal diagnosis (Chamberlain and Caskey 1990).Direct mutation identification in the 2.3 x 1 06-bpdystrophin gene is possible in 65%-70% of cases ofD/BMD by multiplex exon amplification using PCR(Chamberlain et al. 1988; Beggs et al. 1990) and
Received April 8, 1991; revision received July 3, 1991.Address for correspondence and reprints: P. R. Clemens, Insti-
tute for Molecular Genetics, Baylor College of Medicine, Houston,TX 77030.i 1991 by The American Society of Human Genetics. All rights reserved.0002-9297/91 /4905-0005$02.00
Southern-based analysis (Koenig et al. 1987; DenDunnen et al. 1989; Hu et al. 1990). In the remainderof cases without an identifiable mutation, however,carrier detection and prenatal diagnosis depend solelyon DNA linkage studies. To date, linkage analysisin D/BMD has been performed using diallelic RFLPmarkers which may be limited by small family size,inadequate family-member cooperation, and deceasedDMD males (Ward et al. 1989).There are approximately 50,000-100,000 (dC-
dA)n (dG-dT)n (hereafter designated (CA)W) loci inthe human genome (Hamada and Kakunaga 1982;Tautz and Renz 1984). (CA)n loci are a subclass of allshort tandem repeat (STR) sequences. The presenceof these frequently polymorphic loci can be exploitedto improve DNA linkage studies. In addition, STRloci are of great diagnostic utility because they canbe easily assayed by PCR. One (CA)n locus has beendescribed in the 3' untranslated region of the humandystrophin gene (Beggs and Kunkel 1990; Oudet et al.
951

Clemens et al.
Table I
Oligonucleotide Primers for PCR Amplification and Direct DNA SequenceAnalysis of (CA)n Loci in Human Dystrophin Gene (5' to 3')
Intron and Primer (type) Oligonucleotide Sequence
44:DMD-44/A (PCR) .......................... TCCAACATTGGAAATCACATTTCAADMD-44/B (PCR) .......................... TCATCACAAATAGATGTTTCACAG
45:DMD-45/A (PCR) .......................... GAGGCTATAATTCTTTAACTTTGGCDMD-45/B (PCR) .......................... CTCTTTCCCTCTTTATTCATGTTAC
49:DMD-49/A (PCR) .......................... CGTTTACCAGCTCAAAATCTCAACDMD-49/B (PCR) .......................... CATATGATACGATTCGTGTTTTGCDMD-49/C (sequencing) ................. GAGGCTTTATCAGAAGGGTGCAGA
50:DMD-50/A (PCR) .......................... AAGGTTCCTCCAGTAACAGATTTGGDMD-50/B (PCR) .......................... TATGCTACATAGTATGTCCTCAGAC
1990), and four others have been identified in the 5'terminus (Feener et al. 1991). J. S. Chamberlain (un-published data) has identified and sequenced six othersand has designed flanking oligonucleotide PCR prim-ers for their amplification. We describe here (a) allelefrequencies and measures of variation of four of thesesix (CA)n loci (these four being located in introns 44,45, 49, and 50 of the human dystrophin gene) and (b)the use of these markers to improve the accuracy oflinkage analysis for D /BMD carrier detection and pre-natal diagnosis.
Methods
DNA Samples
Samples for DNA extraction included 15-20 cc ofperipheral blood collected in sodium EDTA, culturedamniotic fluid cells, and chorionic villus samples.DNA was isolated using the ABI 340 DNA extractor(Applied Biosystems, Foster City, CA).
Oligonucleotide Primers
Primers for PCR and direct DNA sequencing weresynthesized on an ABI 380B DNA synthesizer (Ap-plied Biosystems, Foster City, CA), deprotected, evap-orated to dryness, and used for PCR and sequencingwithout further purification. Oligonucleotide primersequences are shown in table 1. For detection usingan automated fluorescent gel detection system, oneprimer from each PCR primer pair was fluorescein-labeled at the 5' terminus by a slight modification ofthe method described by Gibbs et al. (1990). Oligonu-
cleotides to be labeled were synthesized using cyano-ethyl phosphoramidites (Applied Biosystems) andAminolink II (Applied Biosystems). Following depro-tection and evaporation to dryness, 0.2 gmol of theamino-modified oligonucleotide was dissolved in 200il of 150 mM sodium carbonate/bicarbonate, pH9.0. After addition of 1 mg of the succinimidyl esterof 5- (and 6-) carboxyfluorescein (Molecular Probes,Inc) mixed with 100 gl dimethylformamide, the oligo-nucleotide-dye mixture was held at room temperaturefor 16 h. Following purification by sodium acetate/ethanol precipitation, the oligonucleotide was dis-solved in 50% formamide, denatured by heating to90°C for 2 min, and further purified by electrophore-sis through a 20% denaturing polyacrylamide gel.Bands containing labeled oligonucleotide were excisedfrom the gel and soaked in 1.5 ml water at 37°C for16 h. The supernatant was aspirated and concentratedto dryness.
In Vitro DNA Amplification and Electrophoresis
PCR was performed essentially according to themethod of Saiki et al. (1988). Twenty nanograms ofgenomic DNA was mixed with 15 pmol of each PCRprimer in a total volume of 15 pl containing 10 mMTris-HCl, pH 8.3, 50 mM KCl, 1.5 mM MgCl2, 0.2mM each deoxyribonucleotide triphosphate, 2 iCideoxycytidine 5'-[a-32P] triphosphate, and 0.75 unitsof Amplitaq DNA polymerase (Cetus). Samples wereheated to 94°C for 4 min, followed by 18-25 cyclesof DNA denaturation (940C for 30 s), annealing(620C for 30 s), and polymerization (650C for 2 min).The final 650C incubation was extended to 7 min.
9S2

(CA), Polymorphisms in Dystrophin
The (CA). blocks in introns 44 and 49 can be amplifiedin the same reaction. The (CA)n loci in introns 45and 50 can also be amplified simultaneously, with theexception that the amount of each DMD-50 primer isdecreased to 2.5 pmol. Aliquots ofPCR products weremixed with one-half volume of formamide dye solu-tion (98% formamide, 10 mM EDTA, 0.1% bromo-phenol blue, and 0.1% xylene cyanol), heated to 90'Cfor 3 min, and electrophoresed on a 6% denaturingpolyacrylamide sequencing gel. Following electropho-resis, each gel was fixed for 30 min in a bath of 10%methanol and 10% acetic acid. Autoradiography ofthe dried gels with an intensifying screen was per-formed for 12-72 h. Alleles were sized by comparisonwith an M13mp18 sequencing standard electropho-resed on the same gel.
Direct DNA Sequencing
DNA sequencing of PCR products was performedessentially according to the method of Gibbs et al.(1989). Primers DMD-49A and DMD-49B were usedto amplify genomic DNA in a reaction volume of 50jil by using the PCR conditions described above andomitting deoxycytidine 5'-[a-32P] triphosphate. Forthe single-strand-producing reaction, 20 pmoles ofprimer DMD-49A was used to amplify 1 11 of thedouble-strand PCR product in a total volume of 100p1. The annealing and elongation times were increasedto 1 and 4 min, respectively. DMD-49C was used asthe sequencing primer.
Fluorescent PCR Product Detection
PCR was performed essentially as described above,except that one of the oligonucleotide primers used foreach amplification was labeled with fluorescein. AnM13mpl8 DNA sequencing standard was generatedusing a fluorescein-labeled Universal sequencingprimer. Aliquots ofPCR products were mixed with anequal volume of deionized formamide with 8.3 mMEDTA, pH 8.0. Following denaturation for 3 min at90°C, samples were electrophoresed on a 6% dena-turing polyacrylamide sequencing gel by using an au-tomated laser fluorescent (ALF)DNA sequencer detec-tion system (Pharmacia LKB Biotechnology).
Results
In Vitro (CA)n Amplification
To facilitate the rapid assignment of alleles, theprimers were designed such that the alleles of the in-tron 45 (CA)n block migrated to a different region of
C C?_ I
Cc_
STR-
STR-4!
STR-49
STR-44
Figure I Autoradiograph showing alleles at 4 (CA)n loci inhuman dystrophin gene for members ofDMD family (pedigree andallele assignments are shown in fig. 8). Each locus is labeled as aSTR followed by the number of the intron in which the (CA)n blockis found.
the gel than did the alleles of the intron 50 (CA)n block.These two loci could then be amplified simultane-ously. Similarly, the primers for the (CA)n loci in in-trons 44 and 49 were designed such that both locicould be amplified in the same reaction. An autoradio-gram of the electrophoretic pattern ofDNA from fourmembers of a DMD family amplified at the four (CA)nloci in two diplex reactions is illustrated in figure 1.The electrophoretic pattern of each allele consists oftwo or more bands spaced at 1- or 2-bp intervals.The complexity of the electrophoretic band pattern isminimized by reducing the number of PCR amplifica-tion cycles to the smallest number required to detectthe product. The band pattern produced for a givenallele is highly reproducible, and one band is mostintense in a homozygote or hemizygote (or two in aheterozygote). Different alleles can be easily distin-guished and assigned a size by comparison with anMl3mpl 8 sequencing standard. We have assigned let-ters for the alleles at each locus, beginning with A andincrementing at 2-bp intervals. Rare alleles of STR-49that occur between the 2-bp interval alleles are indi-cated by the letter of the preceding allele 1 bp smallerfollowed by "1." Allele labeling begins at 174 bp forSTR-44, at 156 bp for STR-45, at 227 bp for STR-49,and at 233 bp for STR-50.
953

Table 2
Allele Frequencies and Predicted Heterozygosity
To determine the informativeness of each (CA). lo-cus, we analyzed 57 unrelated Caucasian males un-affected by D/BMD. For this segment of the popula-tion, we determined frequencies of observed allelesfor each of the (CA)n loci (table 2). Since males arehemizygous for these loci, the allele frequencies aredirectly obtained from their relative counts (yieldingmaximum likelihood estimates). Measures of varia-tion for the four loci are summarized in table 3, inwhich (a) the heterozygosity is estimated by Nei's(1978) method and (b) the numbers of alleles expectedon the basis of estimates of heterozygosity are com-puted according to the method of Chakraborty andGriffiths (1982).
Direct DNA Sequence Analysis
All alleles observed for each of the (CA)n loci inintrons 44, 45, and 50 varied in length by 2-bp inter-vals, presumably because of a variable number of tan-dem CA blocks. However, alleles at 1-bp intervalswere found for the intron 49 (CA)n locus. To explorethis observation further, we sequenced several differ-ent alleles at this locus by using a primer complemen-tary to DNA sequences between the PCR primers forthe sequencing primer and single-strand PCR productgenerated from the double-strand PCR amplificationfor the template. We identified polymorphism in thenumber of C's adjacent to the (CA)n block in intron 49.An example of the analysis for three males is shown infigure 2.
DIBMD Carrier Detection and Prenatal DiagnosisHaplotypes of allele lengths at four (CA)n loci in the
human dystrophin gene were determined in selectedD /BMD familieis in order to perform linkage analysis.Standard Southern-based RFLP haplotyping for link-age analysis and both Southern analysis using cDNAprobes and exon amplification analysis for deletiondetection were also performed in each family, for con-firmation of the STR analysis data and for comparisonof the techniques. In all cases, results of STR analysiswere compatible with results of both Southern-basedRFLP haplotyping and presence of the adjacent exons.Of 38 affected males studied thus far, failure of DNAamplification at one or more of the (CA)n loci hasalways correlated with a deletion of the region (P. R.Clemens and R. G. Fenwick, unpublished observa-tions). STR-based haplotypes for carrier detection(figs. 3-6), prenatal diagnosis (fig. 7), and gene recoin-
Allele Frequencies at (CA), Loci in DystrophinGene
Frequency ± SEAllele Label (sizea) (%)
STR-44:A (174) .............................E (182) .............................G (186) .............................H (188) .............................I (190) ..............................J (192) ..............................K (194) .............................L (196) ..............................M (198) ............................N (200) .............................0 (202) .............................P (204) ..............................
STR-45:A (156) .............................B (158) ..............................E (164) .............................F (166) ..............................G (168) .............................H (170) .............................1 (172) ..............................J (174) ..............................K (176) .............................L (178) ..............................M (180) ............................N (182) .............................0 (184) .............................
STR-49:A (227) .............................B (229) ..............................C (231) .............................D (233) .............................E (235) .............................F (237) ....:.........................G (239) .............................G1 (240) ...........................H (241) .............................I (243) ..............................J (245) ..............................K (247)..............................L (249)...............................Li (250).............................M (251) .............................Ml (252)............................N (253)..............................0 (255)..............................P (257)...............................
STR-50:A (233)..............................E (241)..............................F (243)...............................G (245)..............................I (249) ...............................J (251) ...............................
a Determined by comparison wUniversal sequencing primer.
1.85.35.33.5
19.33.5
14.022.81.83.53.5
15.8
± 1.7+ 2.9± 2.9± 2.4± 5.2± 2.4± 4.6± 5.5± 1.7± 2.4± 2.4± 4.8
5.3 ± 2.912.3 ± 4.35.3 ± 2.93.5 ± 2.4
12.3 ± 4.310.5 ± 4.022.8 ± 5.515.8 ± 4.83.5 ± 2.41.8 ± 1.73.5 ± 2.41.8 ± 1.71.8 ± 1.7
3.5 ± 2.41.8 ± 1.71.8 ± 1.77.0 ± 3.31.8 ± 1.71.8 ± 1.7
10.5 ± 3.71.8 ± 1.78.8 ± 3.73.5 ± 2.4
12.3 ± 4.37.0 ± 3.3
14.0 ± 4.61.8 ± 1.7
10.5 ± 4.01.8 ± 1.71.8 ± 1.73.5 ± 2.45.3 ± 2.9
17.5 ± 5.047.4 ± 6.610.5 ± 4.07.0 ± 3.3
15.8 ± 4.81.8 ± 1.7
rith M13mpl8 sequenced with

(CA). Polymorphisms in Dystrophin
Table 3
Summary of Measures of Variation
Locus Label Heterozygosity ± SE No. of Alleles No. of Alleles(sample size) (%) Observed Expecteda
STR-44 (57) ........ 87.0 ± 2.1 12 13.96STR-45 (57) ........ 88.7 ± 1.9 13 15.48STR-49 (57) ........ 93.3 ± 1.3 19 21.79STR-50 (57) ........ 71.6 ± 4.6 6 7.14
a Based on heterozygosity and sample size and obtained by following Chakraborty and Griffiths' (1982)method.
bination studies (fig. 8) in D/BMD families are illus-trated.
In the DMD family illustrated in figure 3, theaffected male (III-4) was known to be deleted for exons46-50 (data not shown). STR haplotyping confirmedthe deletion, since STR-49 failed to amplify whileSTR-50 (3' to the exon) was detected. Six females inthis pedigree have the at-risk STR haplotype. For eachof these females, apparent homozygosity at STR-49reflects deletion of one allele; densitometry of South-ern analysis data (not shown) confirmed the deletionin each case.The pedigrees shown in figures 4 and 5 illustrate
carrier detection in families in which all DMD maleswere deceased. In figure 4, the carrier status for II-1was not resolved by two-allele RFLP haplotyping ofthe available family members. However, on STR anal-ysis, II-1 inherits the maternal haplotype which is op-posite from the haplotype inherited by II-3, her un-affected brother. In figure 5, 1-2, 11-2, and III-1 eachhave the same deletion beginning in exon 12 and ex-tending through exon 52 (determined by densitometry
of Southern analysis; data not shown). Therefore,each female is hemizygous for the STR haplotype.
Figure 6 shows the use of STR haplotypes to detectgonadal mosaicism. A female sibling (IT-1) of a de-ceased affected DMD male failed to inherit either ma-ternal STR-45 allele. A deletion in this region wasconfirmed by densitometry of the Southern analysis,which shows hemizygosity.The pedigrees of two DMD families requesting pre-
natal diagnosis are illustrated in figure 7. In family 1the affected male (II-1) was found to have DMD whenhis mother (1-2) was in the second trimester of preg-nancy. On STR assay, the male fetus had the oppositemateral haplotype from his affected brother, a rapidindicator that the fetus was unlikely to be affected.Subsequent studies, using Southern analysis and mul-tiplex exon amplification, showed a deletion mutationremoving exons 52-54 of dystrophin in 1-2 and II-1but not in 11-2. In family 2, carrier testing and preg-nancy risk were also performed simultaneously be-
T C AG T C AG
GGG
,~G
G/GGG
N
III
Allele Gi Allele G1l Allele JFigure 2 Direct DNA sequence analysis of amplified allelesof STR-49. Note variation in number of C's adjacent to the (CA)nrepeat (opposite strand shown).
IL t I a I I k 'A IE I ENE F E E E E E
Figure 3 Pedigree ofDMD family showing STR haplotypes.DLN = deletion. The affected male is deleted for the STR-49 locus.Females with the at-risk haplotype are hemizygous for the STR-49locus. Note that the daughters of II-1 have different paternal haplo-types, demonstrating that STR haplotyping is an excellent methodfor confirming paternity within a family.
1,
955
AN.. ..A-%
Clemens et al.
PROBE ORDERSTR-44STR-45STR-49STR-50
RFLPPROBE ORDER
87.15 TaqlC7 EcoRV
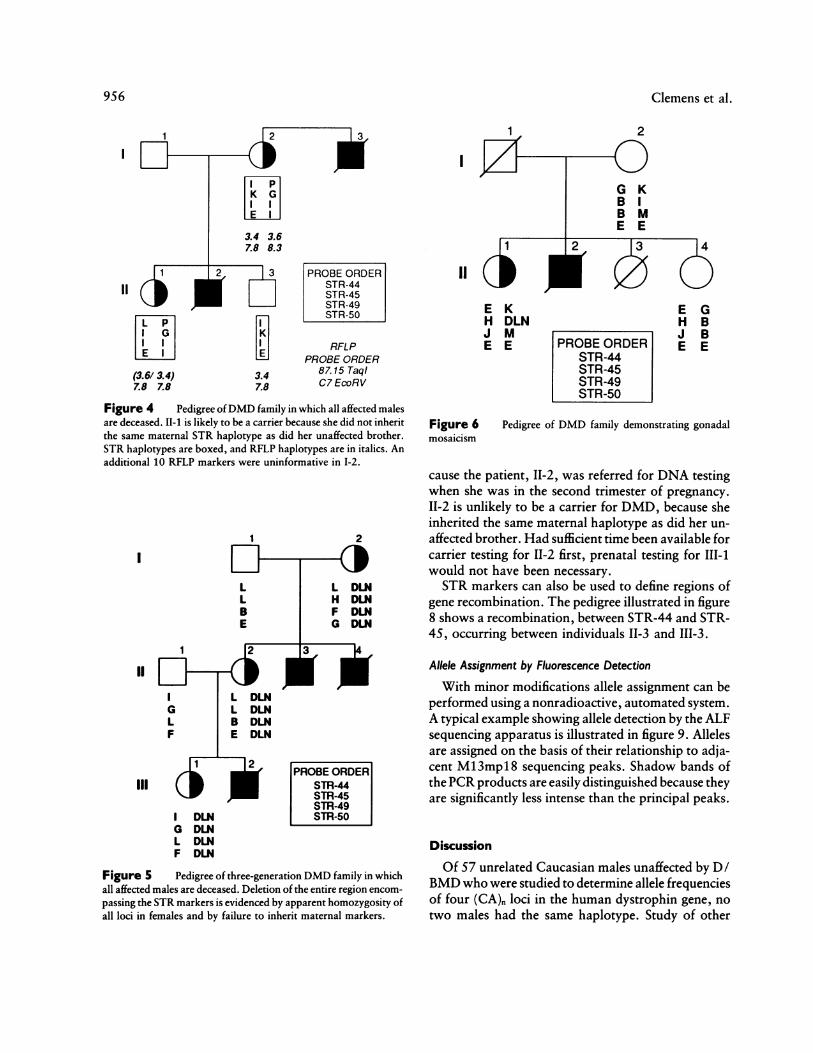
Figure 4 Pedigree ofDMD family in which all affected malesare deceased. II-1 is likely to be a carrier because she did not inheritthe same maternal STR haplotype as did her unaffected brother.STR haplotypes are boxed, and RFLP haplotypes are in italics. Anadditional 10 RFLP markers were uninformative in 1-2.
1
I
III
I DLNG DLNL DLNF DLN
2
PROBE ORDERSTR-44STR-45STR-49STR-50
Figure 5 Pedigree of three-generation DMD family in whichall affected males are deceased. Deletion of the entire region encom-passing the STR markers is evidenced by apparent homozygosity ofall loci in females and by failure to inherit maternal markers.
1 ' 4
E KH DLNJ ME E
I~~~1-~ I
2
G KB IB ME E
2 3 4
E GH B
PROBE ORDER E ESTR-44STR-45STR-49STR-50
Figure 6 Pedigree of DMD family demonstrating gonadalmosaicism
cause the patient, 11-2, was referred for DNA testingwhen she was in the second trimester of pregnancy.II-2 is unlikely to be a carrier for DMD, because sheinherited the same maternal haplotype as did her un-affected brother. Had sufficient time been available forcarrier testing for 11-2 first, prenatal testing for 111-1would not have been necessary.STR markers can also be used to define regions of
gene recombination. The pedigree illustrated in figure8 shows a recombination, between STR-44 and STR-45, occurring between individuals 11-3 and III-3.
Allele Assignment by Fluorescence Detection
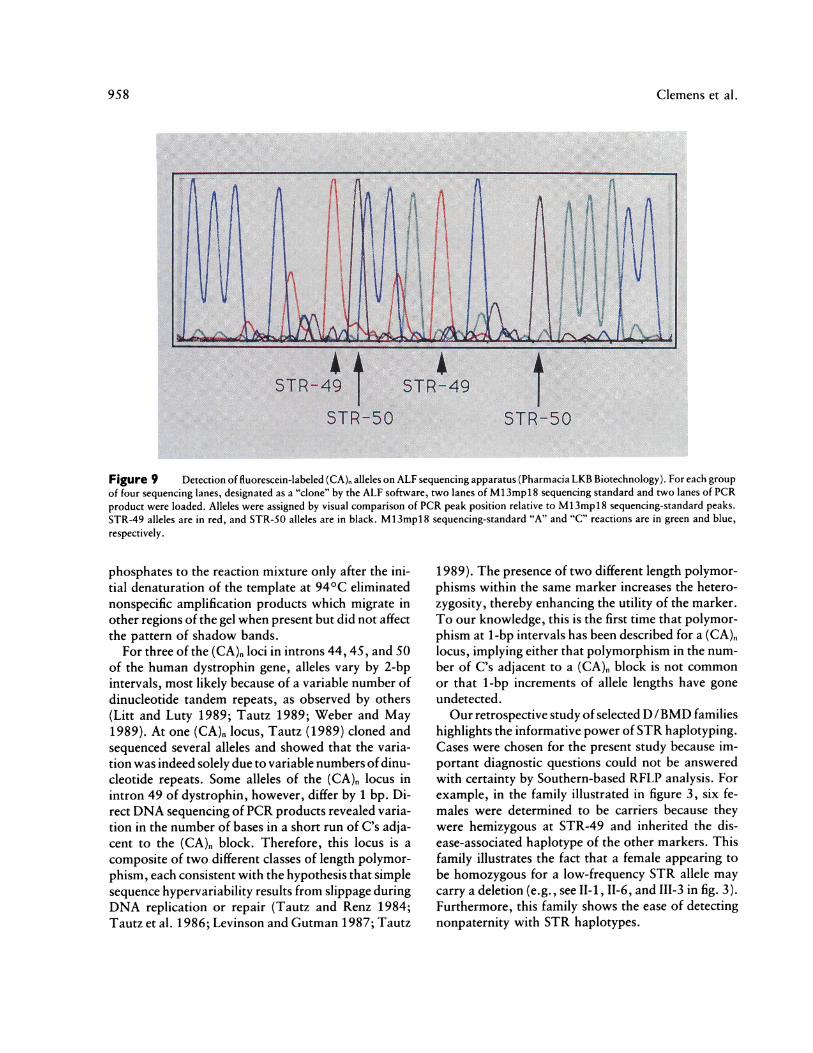
With minor modifications allele assignment can beperformed using a nonradioactive, automated system.A typical example showing allele detection by the ALFsequencing apparatus is illustrated in figure 9. Allelesare assigned on the basis of their relationship to adja-cent M13mpl8 sequencing peaks. Shadow bands ofthePCR products are easily distinguished because theyare significantly less intense than the principal peaks.
Discussion
Of 57 unrelated Caucasian males unaffected by D/BMD who were studied to determine allele frequenciesof four (CA). loci in the human dystrophin gene, notwo males had the same haplotype. Study of other
1 L
l lE I3.4 3.67.8 8.3
Ii-}1'
11 l I
I E I I(3.6/ 3.4)7.8 7.8
2 -~3
1'
3.47.8
956

(CA)n Polymorphisms in Dystrophin
Family 1
F
PROBE ORDERSTR-44STR-45STR-49STR-50
11
11
Family 2
ll
Ill
ADi
F
L
JJI
E
Figure 7 Prenatal diagnosis in two DMD families. The malefetus is unlikely to be affected in either family; in family 1 the fetusdid not inherit the same haplotype as did his affected brother, andin family 2 the fetus inherited the grandpaternal haplotype.
ethnic groups may show differences in degree or distri-bution of polymorphism of these (CA)n loci. The most
polymorphic locus in our study population is STR-49,with 19 alleles and a predicted heterozygosity fre-quency of 93.3%. The least polymorphic locus isSTR-5O, with six alleles and a predicted heterozygos-ity frequency of 71.6%. The agreement between theobserved and expected numbers of alleles at each ofthefour (CA)n loci indicates that the population geneticassumptions of homogeneity and genetic equilibriumthat are required for using these markers for linkageanalyses are adequately justified by these data.STR locus amplification can be used to identify dele-
tions in males affected by D/BMD. To date, we haveidentified and characterized 181 dystrophin deletionmutations by multiplex exon amplification and South-ern analysis (R. G. Fenwick, unpublished observa-tions). The four (CA)n markers described here are lo-cated within a dystrophin gene region frequently
H J G JI AH L MH D D GA E E A
Figure 8 Pedigree of DMD family showing recombinationbetween II-3 and 111-3. The arrow marks the site of recombination.
involved in deletion mutation (Chamberlain et al.,submitted), and so a high proportion (71/181, or39% of the total deletions) would be predicted to bedetected by failure of amplification of one or more ofthe four (CA)n loci.PCR artifacts create a potential problem for deter-
mination ofSTR haplotypes. Other investigators havereported two bands for each (CA)n allele and havedemonstrated differing electrophoretic mobilities ofthe complementary strands of amplifiedDNA by usingend-labeled primers (Weber and May 1989). We didnot observe a significant improvement in the complexband pattern when we used end-labeled primers asan alternative to deoxycytidine 5'-[a-32P] triphosphateincorporation in the assay. This finding, together withour observation that the complexity of the band pat-tern is diminished by performing fewer cycles ofDNAamplification, suggests that there are in vitro-gener-ated minor sequence variants (shadow bands), possi-bly due to replication slippage and 3' base additionby the thermostable DNA polymerase. The shadowbands created during PCR amplification were noteliminated by a variety of other changes in reactionconditions. The shadow band pattern is relatively con-stant at a given locus and is reproducible and, there-fore, did not interfere with allele assignment. Theaddition of the unlabeled deoxyribonucleotide tri-
957
I
Clemens et al.
Figure 9 Detection of fluorescein-labeled (CA), alleles on ALF sequencing apparatus (Pharmacia LKB Biotechnology). For each groupof four sequencing lanes, designated as a "clone" by the ALF software, two lanes of M13mpl8 sequencing standard and two lanes of PCRproduct were loaded. Alleles were assigned by visual comparison of PCR peak position relative to M13mpl8 sequencing-standard peaks.STR-49 alleles are in red, and STR-50 alleles are in black. M13mpl8 sequencing-standard "A" and "C" reactions are in green and blue,respectively.
phosphates to the reaction mixture only after the ini-tial denaturation of the template at 940C eliminatednonspecific amplification products which migrate inother regions of the gel when present but did not affectthe pattern of shadow bands.
For three of the (CA), loci in introns 44, 45, and 50of the human dystrophin gene, alleles vary by 2-bpintervals, most likely because of a variable number ofdinucleotide tandem repeats, as observed by others(Litt and Luty 1989; Tautz 1989; Weber and May1989). At one (CA)n locus, Tautz (1989) cloned andsequenced several alleles and showed that the varia-tion was indeed solely due to variable numbers of dinu-cleotide repeats. Some alleles of the (CA)n locus inintron 49 of dystrophin, however, differ by 1 bp. Di-rect DNA sequencing ofPCR products revealed varia-tion in the number of bases in a short run of C's adja-cent to the (CA)n block. Therefore, this locus is a
composite of two different classes of length polymor-phism, each consistent with the hypothesis that simplesequence hypervariability results from slippage duringDNA replication or repair (Tautz and Renz 1984;Tautz et al. 1986; Levinson and Gutman 1987; Tautz
1989). The presence of two different length polymor-phisms within the same marker increases the hetero-zygosity, thereby enhancing the utility of the marker.To our knowledge, this is the first time that polymor-phism at 1-bp intervals has been described for a (CA)nlocus, implying either that polymorphism in the num-ber of C's adjacent to a (CA), block is not commonor that 1-bp increments of allele lengths have goneundetected.Our retrospective study of selected D/BMD families
highlights the informative power of STR haplotyping.Cases were chosen for the present study because im-portant diagnostic questions could not be answeredwith certainty by Southern-based RFLP analysis. Forexample, in the family illustrated in figure 3, six fe-males were determined to be carriers because theywere hemizygous at STR-49 and inherited the dis-ease-associated haplotype of the other markers. Thisfamily illustrates the fact that a female appearing tobe homozygous for a low-frequency STR allele maycarry a deletion (e.g., see II-1, 11-6, and 111-3 in fig. 3).Furthermore, this family shows the ease of detectingnonpaternity with STR haplotypes.
958

(CA)n Polymorphisms in Dystrophin
In D /BMD families in which affected individualsare unavailable, carrier detection or exclusion fre-quently relies on linkage analysis to determine whetherthe female family members have the same or oppositehaplotype as do unaffected male family members. Inthis setting, STR haplotypes are more likely to be in-formative than are RFLP haplotypes. If the sister of a
deceased affected male is found to be at high risk tobe a carrier for DMD (e.g., see fig. 4), then she has theoption of prenatal diagnosis via STR haplotyping ofa chorionic villus sample or cultured amniocentesiscells in the future.
Figure 5 illustrates a DMD family with no livingaffected males in which we accurately identified dele-tion carriers by apparent homozygosity of the paternalalleles for the four (CA)n loci. The high degree of poly-morphism of these loci decreases the likelihood of ho-mozygosity at all four loci in a normal female. Asdemonstrated in this family, failure to inherit a mater-nal STR marker is readily apparent and providesstrong evidence for a deletion mutation in a carrierdaughter.The high degree of heterozygosity of STR loci also
increases the likelihood for identification of gonadalmosaicism. An example is illustrated in figure 6. Al-though the male with DMD is deceased, a female sib-ling inherits the paternal allele (but neither maternalallele) of STR-45. The simplest interpretation of thesedata is that the mother is a gonadal mosaic for thedeletion which was transmitted to her affected son andcarrier daughter.
For prenatal diagnosis, STR haplotyping providesa marked improvement over Southern-based RFLPanalysis, because of the informativeness and speed ofthe assay. Crucial diagnostic data can be obtainedeven before extracting DNA, because DNA amplifi-cation of (CA)n loci can be performed on boiled celllysates. Furthermore, the STR assay can often excludematernal cell contamination of a chorionic villus sam-ple, because of differences in the STR haplotype be-tween mother and fetus. We have observed one familyin which a female fetus was predicted to carry a dele-tion mutation which included the STR-49 and STR-50loci, because of failure to inherit the maternal allelesfor these markers (data not shown).STR analysis permits more precise localization of
some recombination events within the dystrophin genethan can be achieved with RFLP markers alone. Of 13cases of gene recombination distal to pERT87 thatwere identified by ourDNA diagnostic laboratory, therecombination was localized between intron 44 and
intron 50 in three cases by using STR analysis (P. R.Clemens, unpublished observations). An example ofa recombination between STR-44 and STR-45 is illus-trated in Figure 8. We do not propose that the recom-bination is the cause of the disease in the affected male(111-3), because his mother and sister, who do not haverecombinant haplotypes, have elevated levels of serumcreatine kinase and mild myopathy.We performed allele identification by electrophore-
sis either of radiolabeled PCR product on standarddenaturing sequencing gels or of fluorecein-labeledproduct on an automated, fluorescent detection sys-tem. The principal advantages of the fluorescent detec-tion system are use of nonradioactive substrates andelimination of the need to fix, dry, and expose gelsafter electrophoresis. Currently available computersoftware for the automated, fluorescent system, whichis designed to analyze raw sequencing data generatedby the ALF sequencer, can perform only a crude analy-sis of STR amplification products. As additional soft-ware is developed to size alleles directly and to enterallele assignments into a data base, the labor requiredto interpret the gels and assemble the data for linkageanalysis could be markedly reduced.
In summary, STR analysis using (CA)n repeatswithin the human dystrophin gene is well suited forroutine use in clinical laboratories engaged in linkagestudies for carrier detection and prenatal diagnosis inD/BMD families, because the STR assay relies onPCR amplification, which is simple to perform, pro-vides reproducible results, and is highly informative.With the addition of other hypervariable loci, the de-velopment of software to facilitate allele assignmenton an automated fluorescence sequencing apparatus,and the capability to determine dosage, STR haplotyp-ing may well replace the more cumbersome Southern-based studies as the standard technique for linkageanalysis in D/BMD families.
AcknowledgmentsWe thank Pat Ward, Nikki Nguyen, Maria Saluta, and
Al Edwards for valuable discussions; Susan Smith and RezaMalek for technical assistance; and Pharmacia LKB Biotech-nology for loan of the ALFDNA sequencer detection system.P.R.C. is a recipient of a Muscular Dystrophy Associationpostdoctoral research fellowship, and C.T.C. is an investi-gator of the Howard Hughes Medical Institute. This workwas supported by a Task Force on Genetics grant from theMuscular Dystrophy Association.
959

960 Clemens et al.
ReferencesBeggs AH, Koenig M, Boyce FM, Kunkel LM (1990) Detec-
tion of 98% of DMD/BMD deletions by PCR. HumGenet 86:45-48
Beggs AH, Kunkel LM (1990) A polymorphic CACA repeatin the 3' untranslated region of dystrophin. Nucleic AcidsRes 18:1931
Chakraborty R, Griffiths RC (1982) Correlation of hetero-zygosity and the number of alleles in different frequencyclasses. Theor Popul Biol 21:205-218
Chamberlain JS, Caskey CT (1990) Duchenne musculardystrophy. In: Appel SH (ed) Current neurology, vol 10.Yearbook Medical, Chicago, pp 65-103
Chamberlain JS, Chamberlain JR, Fenwick RG, Ward PA,Dimnik LS, Bech-Hansen NT, Tantravahi U, et al. Diag-nosis of Duchenne's and Becker's muscular dystrophy bypolymerase chain reaction: a multicenter study (sub-mitted)
Chamberlain JS, Gibbs RA, Ranier JE, Nguyen PN, CaskeyCT (1988) Deletion screening of the Duchenne musculardystrophy locus via multiplex DNA amplification. Nu-cleic Acids Res 16:11141-11156
Den Dunnen JT, Grootscholten PM, Bakker E, BlondenLAJ, Ginjaar HB, Wapenaar MC, van Paassen HMB, et al(1989) Topography of the Duchenne muscular dystrophy(DMD) gene: FIGE and cDNA analysis of 194 cases re-veals 115 deletions and 13 duplications. AmJ Hum Genet45:835-847
Feener CA, Boyce FM, Kunkel LM (1991) Rapid detectionof CA polymorphisms in cloned DNA: application to the5' region of the dystrophin gene. Am J Hum Genet 48:621-627
Gibbs RA, Chamberlain JS, Caskey CT (1989) Diagnosis ofnew mutation diseases using the polymerase chain reac-tion. In: Erlich HA (ed) PCR technology: principles andapplications for DNA amplification. Stockton, NewYork, pp 171-191
Gibbs RA, Nguyen P-N, Edwards A, Civitello AB, CaskeyCT (1990) Multiplex DNA deletion detection and exonsequencing of the hypoxanthine phosphoribosyltransfer-ase gene in Lesch-Nyhan families. Genomics 7:235-244
Hamada H. Kakunaga T (1982) Potential Z-DNA formingsequences are highly dispersed in the human genome. Na-ture 298:396-398
Hu X, Ray PN, Murphy EG, Thompson MW, Worton RG(1990) Duplicational mutation at the Duchenne musculardystrophy locus: its frequency, distribution, origin, andphenotype/genotype correlation. Am J Hum Genet 46:682-695
Koenig M, Hoffman EP, Bertelson CJ, Monaco AP, FeenerC, Kunkel LM (1987) Complete cloning of the Duchennemuscular dystrophy gene. Cell 50:509-517
Levinson G, Gutman GA (1987) Slipped-strand mispairing:a major mechanism for DNA sequence evolution. MolBiol Evol 4:203-221
Litt M, Luty JA (1989) A hypervariable microsatellite re-vealed by in vitro amplification of a dinucleotide repeatwithin the cardiac muscle actin gene. Am J Hum Genet44:397-401
Nei M (1978) Estimation of average heterozygosity and ge-netic distance from a small number of individuals. Genet-ics 89:583-590
Oudet C, Heilig R, Mandel JL (1990) An informative poly-morphism detectable by polymerase chain reaction at the3' end of the dystrophin gene. Hum Genet 84:283-285
Saiki RK, Gelfand DH, Stoffel S, Scharf SJ, Higuchi R, HornGT, Mullis KB, et al (1988) Primer-directed enzymaticamplification ofDNA with a thermostable DNA polymer-ase. Science 239:487-491
Tautz D (1989) Hypervariability of simple sequences as ageneral source for polymorphic DNA markers. NucleicAcids Res 17:6463-6471
Tautz D, Renz M (1984) Simple sequences are ubiquitousrepetitive components of eukaryotic genomes. NucleicAcids Res 12:4127-4138
Tautz D, Trick M, Dover GA (1986) Cryptic simplicity inDNA is a major source of genetic variation. Nature 322:652-656
Ward PA, Hejtmancik JF, Witkowski JA, Baumbach LL,Gunnell S, SpeerJ, Hawley P, et al (1989) Prenatal diagno-sis of Duchenne muscular dystrophy: prospective linkageanalysis and retrospective dystrophin cDNA analysis. AmJ Hum Genet 44:270-281
Weber JL, May PE (1989) Abundant class of human DNApolymorphisms which can be typed using the polymerasechain reaction. Am J Hum Genet 44:388-396