ccl21 transgenic expression of the cc chemokine of the thyroid
TRANSCRIPT

of April 4, 2018.This information is current as
CCL21Transgenic Expression of the CC Chemokineof the Thyroid Gland Generated by A Novel Model for Lymphocytic Infiltration
Martin Lipp, Glaucia C. Furtado and Sergio A. LiraCanasto-Chibuque, Jonathon D. Sedgwick, Paul S. Frenette,Shu-Cheng Chen, Galya Vassileva, Claudia Andrea P. Martin, Elizabeth C. Coronel, Gen-ichiro Sano,
http://www.jimmunol.org/content/173/8/4791doi: 10.4049/jimmunol.173.8.4791
2004; 173:4791-4798; ;J Immunol
Referenceshttp://www.jimmunol.org/content/173/8/4791.full#ref-list-1
, 23 of which you can access for free at: cites 61 articlesThis article
average*
4 weeks from acceptance to publicationFast Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
Submit online. ?The JIWhy
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved.Copyright © 2004 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on April 4, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
by guest on April 4, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

A Novel Model for Lymphocytic Infiltration of the ThyroidGland Generated by Transgenic Expression of the CCChemokine CCL211
Andrea P. Martin,2* Elizabeth C. Coronel,2† Gen-ichiro Sano,* Shu-Cheng Chen,†
Galya Vassileva,† Claudia Canasto-Chibuque,* Jonathon D. Sedgwick,‡ Paul S. Frenette,*Martin Lipp,§ Glaucia C. Furtado,2* and Sergio A. Lira3*
Lymphocytic infiltrates and lymphoid follicles with germinal centers are often detected in autoimmune thyroid disease (AITD), butthe mechanisms underlying lymphocyte entry and organization in the thyroid remain unknown. We tested the hypothesis thatCCL21, a chemokine that regulates homeostatic lymphocyte trafficking, and whose expression has been detected in AITD, isinvolved in the migration of lymphocytes to the thyroid. We show that transgenic mice expressing CCL21 from the thyroglobulinpromoter (TGCCL21 mice) have significant lymphocytic infiltrates, which are topologically segregated into B and T cell areas.Although high endothelial venules expressing peripheral lymph node addressin were frequently observed in the thyroid tissue,lymphocyte recruitment was independent of L-selectin or lymphotoxin-� but required CCR7 expression. Taken together, theseresults indicate that CCL21 is sufficient to drive lymphocyte recruitment to the thyroid, suggest that CCL21 is involved in AITDpathogenesis, and establish TGCCL21 transgenic mice as a novel model to study the formation and function of lymphoid folliclesin the thyroid. The Journal of Immunology, 2004, 173: 4791–4798.
T he autoimmune thyroid diseases (AITD)4 are the mostcommon autoimmune endocrine diseases, affecting�1.5% of the general population (1). The major forms of
AITD are Hashimoto’s thyroiditis (HT), causing hypothyroidism,and Graves’ disease (GD), causing hyperthyroidism. The hallmarkof these conditions is thyroid dysfunction associated with the pres-ence of lymphocytic infiltrates in the thyroid (2, 3). In HT, there isa diffuse lymphocytic infiltration with the presence of macro-phages and destroyed thyrocytes. The lymphocytes often cluster toform lymphoid follicles with germinal centers. Lymphocytic infil-trates, usually without germinal center formation, are also seen inGD (4), but diffuse follicular cell hyperplasia and increased vas-cularity, rather than thyrocyte destruction, are observed (3).
The etiopathogenic role of lymphocytes in HT and GD has beenamply studied and a variety of effector mechanisms have beendefined for both T and B cells (2, 5). T cells cause thyroid de-struction directly, via cytotoxicity, or indirectly, through cytokinesthat induce apoptosis of thyroid follicle cells (5). B cells produce
autoantibodies directed against thyroid Ags such as thyroglobulin(TG), thyroid peroxidase, and the thyroid-stimulating hormone(TSH) receptor (TSH-R) (2). These autoantibodies can either stim-ulate or inhibit thyroid function.
Despite substantial clinical and experimental data supporting arole for lymphocytes in triggering and sustaining AITD, factorsresponsible for their recruitment and retention in the thyroid remainuncertain. Migration of T cells appears to be regulated by inflamma-tory (inducible) and constitutive chemokines. Examples of inflamma-tory chemokines include CXCL8, CCL2, CCL3, CCL4, andCXCL10. Expression of these chemokines occurs during inflam-mation and is required for attraction of specific leukocyte subsetsmediating inflammatory reactions (6, 7). CXCL10, the best studiedof the inflammatory chemokinesregulating T cell migration, is in-duced by IFN-�, a key cytokine involved in inflammation and im-mune regulation (8). CXCL10 interacts with CXCR3, a chemokinereceptor expressed by effector T cells and endothelial cells (9).
Chemokines constitutively produced by lymphoid organs(CCL19, CCL21, CXCL12, and CXCL13) are involved in physi-ological trafficking of leukocytes and their segregation into spe-cialized compartments. Among these, CCL19 and CCL21 are thebest-characterized chemokines regulating homeostatic T cell mi-gration. Both chemokines interact with a common receptor, CCR7,which is expressed by naive and memory T cells (10). CCL21 andCCL19 are displayed by high endothelial venules (HEVs) and ac-tivate integrins after interaction with CCR7 present on circulatinglymphocytes (11). The functional consequence of integrin activa-tion is increased adhesiveness of lymphocytes to the endothelium,a key step in lymphocyte transendothelial migration. Accordingly,mice lacking CCR7, CCL19, and CCL21 have defective homing ofT cells into lymphoid tissue (12–14). In addition to controlling theinitial aspects of lymphocyte migration, chemokines also controltargeted homing of T and B cells within lymphoid organs. Thespecific segregation of B and T cells in secondary lymphoid organs
*Immunobiology Center, Mount Sinai School of Medicine, New York, NY 10029;†Schering-Plough Research Institute, Kenilworth, NJ 07033;‡DNAX Research, PaloAlto, CA 94304; and§Max-Delbruck-Center for Molecular Medicine, Berlin, Germany
Received for publication June 4, 2004. Accepted for publication July 30, 2004.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby markedadvertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 S.A.L. is an Irene Diamond Associate Professor of Immunology. This work wassupported in part by a grant from the Irene Diamond Fund, and by Grants DK 067989(to S.A.L.) and HL 69438 (to P.S.F.) from the National Institutes of Health.2 A.P.M., E.C.C., and G.C.F. contributed equally to the manuscript.3 Address correspondence and reprint requests to Dr. Sergio A. Lira, ImmunobiologyCenter, Mount Sinai School of Medicine, 1425 Madison Avenue, Box 1630, NewYork, NY, 10029-6574. E-mail address: [email protected] Abbreviations used in this paper: AITD, autoimmune thyroid diseases; HT, Hashi-moto’s thyroiditis; GD, Graves’ disease; TG, thyroglobulin; TSH, thyroid-stimulatinghormone; HEV, high endothelial venule; Lt�, lymphotoxin �; PNAd, peripherallymph node addressin; LDL, low density lipoprotein.
The Journal of Immunology
Copyright © 2004 by The American Association of Immunologists, Inc. 0022-1767/04/$02.00
by guest on April 4, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

is dependent on multiple chemokine gradients established by den-dritic cells, stromal cells, and lymphocytes (15).
Chemokines, with the exception of CXCL12 (16), are not ex-pressed by the normal human thyroid at appreciable levels. How-ever, most chemokines studied to date are expressed within thethyroids of GD and HT patients. These include CXC chemokines,CXCL1 (17), the IFN-� inducible chemokines CXCL9, andCXCL10 (9, 18, 19), CXCL12 (16), and CXCL13 (20). The CCchemokines CCL2, CCL3, CCL4, and CCL5 are also reported tobe expressed in thyroid disease (19, 21, 22). Thyroid expression ofthese chemokines may be critical for the recruitment of chemokinereceptor-expressing leukocytes to the thyroid. Indeed, CXCR3,CCR2, and CCR5 expression is elevated on infiltrating inflamma-tory cells relative to peripheral blood leukocytes (9, 18, 21, 23).The cellular source of these chemokines in the thyroid is usuallythe infiltrating lymphocytes, but thyrocytes and fibroblasts alsoexpress chemokines in diseased thyroid (17). Different proinflam-matory stimuli such as IL-1, IFN-�, and TNF-� increase expres-sion of chemokines by thyroid follicular cells in culture (17, 18,24, 25), suggesting that inflammatory cytokines may trigger ex-pression of chemokines by thyrocytes and other resident thyroidcells in vivo. Interestingly, recent studies have shown that chemo-kines required for the formation and maintenance of lymphoid fol-licles (i.e., CXCL12, CCL21, and CXCL13) are expressed in thethyroids of AITD patients (20, 26). The expression of these mol-ecules in the thyroid suggests that chemokines may be necessaryfor recruitment and subsequent organization of lymphoid folliclesin the thyroid. CCL21 and CXCL13 have been shown to induceformation of lymphoid structures when expressed in the pancreas(27–29). However, this property does not seem to be general, asexpression of CCL21 in brain and skin of transgenic mice fails toinduce development of such structures (30). Here, we directly testthe hypothesis that the chemokine CCL21 may be directly impli-cated in regulating influx of lymphocytes into the thyroid. Weshow that CCL21 is sufficient to drive not only influx, but alsoformation of lymph node-like structures in the thyroid of trans-genic animals, suggesting that this chemokine and its receptor,CCR7, may be involved in the formation of lymphoid follicles inthe thyroid of patients with AITD. Additionally, we show thatinfiltration of the thyroid by lymphocytes under these conditions isindependent of L-selectin and lymphotoxin � (LT�).
Materials and MethodsConstruction of the TGCCL21 transgene
The rat TG promoter (GenBank X06162) was amplified by PCR from ratgenomic DNA using recognition primers that contained sites for HindIII atthe 5� end and for EcoRI at the 3� end to include nucleotides �868 to �1from the transcription start site. The rat TG promoter was subcloned intoHindIII/EcoRI sites of pBS plasmid (Stratagene, La Jolla, CA) to generatethe TG/pBS plasmid.
A BstX I fragment (2.4 kb) containing CCL21 genomic DNA encodingCCL21a (6Ckine serine) was isolated from a BAC clone (31) and blunt-endligated into the EcoRI site of the TG/pBS plasmid to generate a vectorencoding the TGCCL21 transgene. The TGCCL21 transgene (3.3 kb) wasreleased from the TG/pBS vector by digestion with XhoI/XbaI restrictionenzymes. Separation of the transgene DNA from vector DNA was accom-plished by zonal sucrose gradient centrifugation as described (32). Thefractions containing the transgene DNA were pooled, microcentrifugedthrough Microcon-100 filters (Millipore, Bedford, MA), and washed fivetimes with microinjection buffer (5 mM Tris-HCl, pH 7.4, 5 mM NaCl, 0.1mM EDTA).
Mice
The TGCCL21 transgene DNA was resuspended in microinjection bufferto a final concentration of 1–5 ng/�l, microinjected into fertilized eggsfrom (C57BL/6J � DBA/2)F2 females (The Jackson Laboratory, Bar Har-bor, ME), and transferred into oviducts of ICR foster mothers (Charles
River Laboratories, Wilmington, MA) according to published procedures(33). Genotyping of the TGCCL21 transgenic founders and progeny wasconducted by PCR analysis of mouse tail DNA using recognition primersfor the rat TG promoter 5�-CTG CAG ACA AGC AGG CAT GCA-3�(forward) and 5�-CAC ACA TGG CAC ATA TGC-3� (reverse) as previ-ously described (34). The endogenous low density lipoprotein gene (LDL)was used as an internal control using primers LDLL 5�-CGC AGT GCTCCT CAT CTG ACT TGT-3� (forward) and LDLU 5�-ACC CCA AGACGT GCT CCC AGG ATG A-3� (reverse). PCR conditions were 94°C,30 s; 60°C, 30 s; 72°C, 60 s for 30 cycles.
TGCCL21 transgenic mice (line 24) were crossed to CCR7�/� mice,described by Forster et al. (12), to LT��/� (35) and to RAG-2�/� mice(36). Mice hemizygous for CCL21 and null for RAG or CCR7 or LT�(referred to as TGCCL21/RAG�/� mice, TGCCL21/CCR7�/� mice, andTGCCL21/LT��/� mice, respectively) and littermate controls were usedfor the experiments described here. Transgenic mice expressing GFP (37)and L-selectin�/� mice (38) were backcrossed over 10 generations in theC57BL/6 background. All mice were housed under specific-pathogen-freeconditions in individually ventilated cages at the Mount Sinai School ofMedicine Animal Facility. All experiments were performed following in-stitutional guidelines.
Histology
Tissues for light microscopic examination were fixed by immersion in 10%phosphate-buffered formalin and then processed for paraffin sections. Rou-tinely, 5-�m sections were cut and stained with H&E. For immunohisto-chemical staining, fresh frozen sections were first fixed with ice-cold ac-etone for 20 min, dried, and stored at �20°C. Slides were stained andanalyzed as described previously (39). Slides were incubated for 1 h atroom temperature with purified primary Abs followed by incubation withthe appropriate labeled secondary Abs for 30 min. Primary Abs used wereanti-Thy1.2 (53-2.1), CD3 (145-2C11), CD4 (H129.19), B220 (RA3-6B2),peripheral lymph node addressin (PNAd) (MECA79) from BD Biosciences(San Diego, CA), and anti-6Ckine/CCL21 (no. AF457) from R&D Systems(Minneapolis, MN). Secondary Abs used were Alexa Fluor 594 donkeyanti-goat (no. A-11058), Alexa Fluor 488 and 594 goat anti-rat IgM (nos.A-21212 and A-21213) and anti-rat IgG (nos. A-11006 and A-11007) fromMolecular Probes (Eugene, OR) and FITC and rhodamine red-X goat anti-Armenian hamster (nos. 127-095-160 and 127-295-160) and Cy5 goat anti-rat (no. 112-175-167) from Jackson ImmunoResearch Laboratories (WestGrove, PA).
Flow cytometry
To prepare single cell suspension, individual thyroids from TGCCL21 an-imals (6 wk of age) were minced in serum-free RPMI 1640 media con-taining 0.0012 U/ml dispase I (Sigma-Aldrich, St. Louis, MO) and 0.25U/ml collagenase type II (Worthington Biochemical/Invitrogen Life Tech-nologies, Rockville, MD), incubated for 2 h at 4°C followed by an addi-tional incubation of 10 min at 37°C. Thyroid cell suspensions obtainedfrom both lobes were centrifuged at 250 � g for 10 min at 4°C and re-suspended in FACS staining buffer (PBS containing 2% FCS and 0.01%sodium azide).
Cells were incubated for 20 min at 4°C with 5 �g/ml Fc block (BDPharmingen, San Diego, CA) and then stained with directly conjugatedprimary mAbs. mAbs to the following mouse leukocyte surface markerswere purchased from BD Pharmingen: CD45 (30-F11), CD3e (145-2C11),CD4 (RM4-5), B220 (RA3-6B2), CD62L (MEL-14), CD11c (HL-3), andCD11b (M1/70). To determine viability, samples were subsequentlystained with 20 �l of 5 �g/ml propidium iodide (Calbiochem, La Jolla,CA). Events were acquired on a BD Biosciences FACScan and analyzedusing CellQuest software. To rule out the presence of rare thymic contam-inants in the thyroid single cell preparations, we routinely determined therelative proportion of thymocytes or immature T cells (CD4�/CD8� cells).Samples that contained �2% of this cell population were excluded fromthe analysis.
Adoptive cell transfers
Spleen cells (107) were resuspended in PBS and injected i.v. Lymph nodesand thyroids from recipient animals were collected at various points (24,48, 72, and 96 h) and fixed in 1.5% paraformaldehyde containing 20%sucrose for 24 h at 4°C. Fixed tissues were frozen in OCT compound(Tissue Tek, Torrance, CA), cryosectioned (8 mm), and examined under afluorescent microscope. In some instances, images were overlaid and pro-cessed using Adobe Photoshop.
4792 TRANSGENIC EXPRESSION OF CCL21 IN THE THYROID
by guest on April 4, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
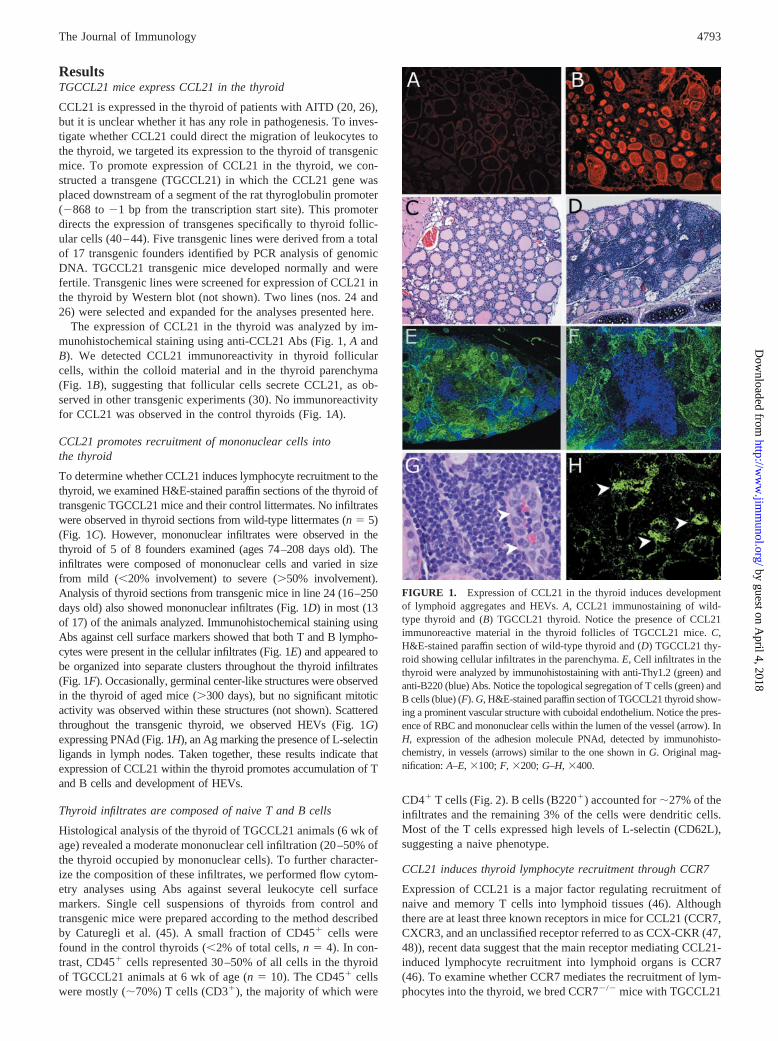
ResultsTGCCL21 mice express CCL21 in the thyroid
CCL21 is expressed in the thyroid of patients with AITD (20, 26),but it is unclear whether it has any role in pathogenesis. To inves-tigate whether CCL21 could direct the migration of leukocytes tothe thyroid, we targeted its expression to the thyroid of transgenicmice. To promote expression of CCL21 in the thyroid, we con-structed a transgene (TGCCL21) in which the CCL21 gene wasplaced downstream of a segment of the rat thyroglobulin promoter(�868 to �1 bp from the transcription start site). This promoterdirects the expression of transgenes specifically to thyroid follic-ular cells (40–44). Five transgenic lines were derived from a totalof 17 transgenic founders identified by PCR analysis of genomicDNA. TGCCL21 transgenic mice developed normally and werefertile. Transgenic lines were screened for expression of CCL21 inthe thyroid by Western blot (not shown). Two lines (nos. 24 and26) were selected and expanded for the analyses presented here.
The expression of CCL21 in the thyroid was analyzed by im-munohistochemical staining using anti-CCL21 Abs (Fig. 1, A andB). We detected CCL21 immunoreactivity in thyroid follicularcells, within the colloid material and in the thyroid parenchyma(Fig. 1B), suggesting that follicular cells secrete CCL21, as ob-served in other transgenic experiments (30). No immunoreactivityfor CCL21 was observed in the control thyroids (Fig. 1A).
CCL21 promotes recruitment of mononuclear cells intothe thyroid
To determine whether CCL21 induces lymphocyte recruitment to thethyroid, we examined H&E-stained paraffin sections of the thyroid oftransgenic TGCCL21 mice and their control littermates. No infiltrateswere observed in thyroid sections from wild-type littermates (n � 5)(Fig. 1C). However, mononuclear infiltrates were observed in thethyroid of 5 of 8 founders examined (ages 74–208 days old). Theinfiltrates were composed of mononuclear cells and varied in sizefrom mild (�20% involvement) to severe (�50% involvement).Analysis of thyroid sections from transgenic mice in line 24 (16–250days old) also showed mononuclear infiltrates (Fig. 1D) in most (13of 17) of the animals analyzed. Immunohistochemical staining usingAbs against cell surface markers showed that both T and B lympho-cytes were present in the cellular infiltrates (Fig. 1E) and appeared tobe organized into separate clusters throughout the thyroid infiltrates(Fig. 1F). Occasionally, germinal center-like structures were observedin the thyroid of aged mice (�300 days), but no significant mitoticactivity was observed within these structures (not shown). Scatteredthroughout the transgenic thyroid, we observed HEVs (Fig. 1G)expressing PNAd (Fig. 1H), an Ag marking the presence of L-selectinligands in lymph nodes. Taken together, these results indicate thatexpression of CCL21 within the thyroid promotes accumulation of Tand B cells and development of HEVs.
Thyroid infiltrates are composed of naive T and B cells
Histological analysis of the thyroid of TGCCL21 animals (6 wk ofage) revealed a moderate mononuclear cell infiltration (20 –50% ofthe thyroid occupied by mononuclear cells). To further character-ize the composition of these infiltrates, we performed flow cytom-etry analyses using Abs against several leukocyte cell surfacemarkers. Single cell suspensions of thyroids from control andtransgenic mice were prepared according to the method describedby Caturegli et al. (45). A small fraction of CD45� cells werefound in the control thyroids (�2% of total cells, n � 4). In con-trast, CD45� cells represented 30–50% of all cells in the thyroidof TGCCL21 animals at 6 wk of age (n � 10). The CD45� cellswere mostly (�70%) T cells (CD3�), the majority of which were
CD4� T cells (Fig. 2). B cells (B220�) accounted for �27% of theinfiltrates and the remaining 3% of the cells were dendritic cells.Most of the T cells expressed high levels of L-selectin (CD62L),suggesting a naive phenotype.
CCL21 induces thyroid lymphocyte recruitment through CCR7
Expression of CCL21 is a major factor regulating recruitment ofnaive and memory T cells into lymphoid tissues (46). Althoughthere are at least three known receptors in mice for CCL21 (CCR7,CXCR3, and an unclassified receptor referred to as CCX-CKR (47,48)), recent data suggest that the main receptor mediating CCL21-induced lymphocyte recruitment into lymphoid organs is CCR7(46). To examine whether CCR7 mediates the recruitment of lym-phocytes into the thyroid, we bred CCR7�/� mice with TGCCL21
FIGURE 1. Expression of CCL21 in the thyroid induces developmentof lymphoid aggregates and HEVs. A, CCL21 immunostaining of wild-type thyroid and (B) TGCCL21 thyroid. Notice the presence of CCL21immunoreactive material in the thyroid follicles of TGCCL21 mice. C,H&E-stained paraffin section of wild-type thyroid and (D) TGCCL21 thy-roid showing cellular infiltrates in the parenchyma. E, Cell infiltrates in thethyroid were analyzed by immunohistostaining with anti-Thy1.2 (green) andanti-B220 (blue) Abs. Notice the topological segregation of T cells (green) andB cells (blue) (F). G, H&E-stained paraffin section of TGCCL21 thyroid show-ing a prominent vascular structure with cuboidal endothelium. Notice the pres-ence of RBC and mononuclear cells within the lumen of the vessel (arrow). InH, expression of the adhesion molecule PNAd, detected by immunohisto-chemistry, in vessels (arrows) similar to the one shown in G. Original mag-nification: A–E, �100; F, �200; G–H, �400.
4793The Journal of Immunology
by guest on April 4, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

mice to generate animals that expressed CCL21 in the thyroid, butlacked CCR7 expression (TGCCL21/CCR7�/� mice). Histologi-cal examination of thyroids from TGCCL21/CCR7�/� orTGCCL21/CCR7�/� mice (n � 10 mice/group) revealed infiltra-tion by mononuclear cells (Fig. 3A). In contrast, none of the thy-roids from TGCCL21/CCR7�/� mice (n � 10) examined had leu-kocytic infiltrates when examined by light (Fig. 3B) orimmunofluorescence microscopy using Abs against the pan-leu-kocyte marker CD45 (not shown). These findings indicate thatCCR7 is the main chemokine receptor mediating CCL21-inducedlymphocyte recruitment into the thyroid.
CCL21 is not sufficient to promote HEV development in thethyroid
HEVs, specialized blood vessels used by circulating lymphocytesto migrate into lymphoid organs, were present in the thyroid pa-renchyma of TGCCL21 mice. Next, we asked whether CCL21could directly promote the development of HEVs in this tissue. Tothis end, we crossed the TGCCL21 mice with RAG-2�/� mice,which are devoid of mature T and B cells and examined the pres-ence of HEVs in the thyroid (Fig. 4). As expected, significantinfiltrates were observed in the thyroid of TGCCL21/RAG�/�
mice (n � 5) whereas no infiltrates were observed in the thyroid ofTGCCL21/RAG�/� mice (n � 8), by H&E staining (Fig. 4, A andB, respectively) or CD45 staining (not shown). HEVs expressingPNAd (Fig. 4C), were found within TGCCL21/RAG�/� thyroids.In contrast, no HEVs were observed in the TGCCL21/RAG�/�
thyroid (Fig. 4D), suggesting that CCL21 is not sufficient to drive
HEV formation and that factors produced by the infiltrating lym-phocytes and/or dendritic cells may be important for the develop-ment of these structures in the tissue.
Expression of CCL21 promotes recruitment of adoptivelytransferred lymphocytes to the adult thyroid
To test whether CCL21 expression could promote recruitment ofdonor lymphocytes to the thyroid, we transferred 107 GFP�
splenocytes (i.v.) into the TGCCL21/RAG�/� animals describedabove (n � 8). GFP� splenocytes were also injected into controlRAG�/� mice (n � 5) that did not express CCL21 in the thyroid(Fig. 5B). Five days later we examined the presence of GFP� cellsin frozen sections of lymph nodes, spleen, and thyroids ofTGCCL21/RAG�/� and RAG�/� mice. As expected, GFP� cellsaccumulated in lymph nodes (Fig. 5A, a and b) and spleens (notshown) of both TGCCL21/RAG�/� and RAG�/� mice. Interest-ingly, clusters of GFP� cells were visualized in the thyroid of theTGCCL21/RAG�/� mice (Fig. 5C), but not in the thyroid ofRAG�/� mice (Fig. 5B) 5 days after transfer. Most of the infil-trating GFP� cells at this time point were CD4� T cells (notshown). Sixty days after transfer, large clusters of GFP� cells wereobserved in the thyroid (Fig. 5D) (n � 2). These cells were mostlyT and B cells that appeared to segregate into specific areas (Fig.5E). Within these clusters we observed PNAd� vessels (Fig. 5F).These results indicate that expression of CCL21 by thyroid follic-ular cells promotes mobilization of adoptively transferred lympho-cytes into the thyroid parenchyma.
Expression of CCL21 promotes the recruitment of adoptivelytransferred lymphocytes independently of HEV formation andL-selectin expression
The absence of both lymphoid infiltrates and HEVs in TGCCL21/RAG�/� suggested that the presence of HEVs was important forlymphocytic recruitment. To test this possibility, we transferreddonor wild-type splenocytes into TGCCL21/RAG�/� mice andexamined thyroid specimens for the presence of HEVs and expres-sion of PNAd. We found no HEV-like structures, or PNAd ex-pression up to 96 h after adoptive transfer, indicating that HEVs
FIGURE 2. Naive lymphocytes infiltrate the thyroid of TGCCL21 ani-mals. Single cell suspensions of the thyroid of TGCCL21 mice (6 wk old)were stained with the indicated Abs. Cells were gated on the viable leu-kocyte population (CD45�/PI�). Cells in the histogram plot were gated onthe CD45�/PI�/CD3� subset. Results are representative of several inde-pendent experiments (n � 10 mice); PI, propidium iodide.
FIGURE 3. Absence of cellular infiltrates in TGCCL21/CCR7�/� mice.A, Large cellular infiltrates in TGCCL21/CCR7�/� thyroid. B, No infil-trates are observed in TGCCL21/CCR7�/� thyroid. Original magnifica-tion: A–B, �200.
FIGURE 4. Absence of cellular infiltrates and HEV structures inTGCCL21/RAG�/� mice. A, Large cellular infiltrates in TGCCL21/RAG�/� thyroid. B, No infiltrates are observed in TGCCL21/RAG�/�
thyroid. PNAd staining is observed in HEVs in the thyroid of TGCCL21/RAG�/� (C), but absent in the thyroid of TGCCL21/RAG�/� mice (D).Original magnification: A–B, �200; C–D, �400.
4794 TRANSGENIC EXPRESSION OF CCL21 IN THE THYROID
by guest on April 4, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

are not required for initial colonization of the thyroid by lympho-cytes (data not shown).
The absence of PNAd expression suggested that the initial lym-phocyte infiltration might have occurred through an L-selectin-independent pathway. To test this hypothesis, we transferred L-se-lectin�/� and L-selectin�/� splenocytes into TGCCL21/RAG�/�
mice and examined the thyroids 5 days later (n � 5 mice/group).As shown in Fig. 6, L-selectin�/� lymphocytes were found in thethyroid in a pattern indistinguishable from that observed for theL-selectin�/� lymphocytes. To further analyze whether L-selectinplayed a major role in migration of cells to the thyroid we quan-tified the infiltrates by FACS. The number of CD45� cells foundin the thyroid of TGCCL21/RAG�/� mice after transfer of L-se-lectin�/� and L-selectin�/� splenocytes was similar (average
2.8 � 103 cells vs 2.2 � 103 cells, n � 3). These results indicatethat L-selectin is not required for initial entry of lymphocytes intothe thyroid of TGCCL21/RAG�/� mice.
Lymphocyte infiltration and topological arrangement inTGCCL21 mice is independent of LT�
Genetic studies in mice have shown that members of the TNF/lymphotoxin family are important for lymphoid organ develop-ment (49). Furthermore, expression of LT� has been documentedin AITD (26). However, no information exists to date on the roleof the lymphotoxin system in the generation of lymphoid aggre-gates in the thyroid. To test the hypothesis that the formation oflymphoid aggregates induced by CCL21 requires LT�, we crossedthe TGCCL21 with LT��/� mice to generate mice deficient inLT�, expressing CCL21 in the thyroid (TGCCL21/LT��/�).When examined at 6 wk of age TGCCL21/LT��/� mice (n � 8)and LT��/� mice (n � 3) had no peripheral lymph nodes. How-ever, TGCCL21/LT��/�, but not LT��/� mice, had thyroidmononuclear infiltrates composed of B and T cells that were or-ganized in distinct areas (Fig. 7B) similar to that observed in theTGCCL21/LT��/� mice (Fig. 7A). In addition, PNAd-positiveHEVs were found within both TGCCL21/LT��/� (Fig. 7C) andTGCCL21/LT��/� (Fig. 7D) thyroids. Thus, LT� is not requiredfor accumulation of T, B cells, or the formation of HEVs inducedby CCL21 in the thyroid.
DiscussionDespite the existence of solid experimental data linking humanthyroid disease and expression of chemokines, the studies to datehave been mostly descriptive. Little is know about the mechanisms
FIGURE 5. Selective migration of adoptively transferred GFP� splenocytes into the thyroid of mice expressing CCL21. GFP� splenocytes weretransferred i.v. into RAG�/� mice and into TGCCL21/RAG�/� mice. GFP� splenocytes migrated to the lymph nodes of RAG�/� (Aa) and TGCCL21/RAG�/�(Ab) mice with similar efficiency. RAG�/� mice do not express CCL21 in the thyroid (B), but the TGCL21/RAG�/� mice do express it (red, C).Five days after the adoptive transfer, GFP� cells (green) colocalized with CCL21 expressing cells (red) in the thyroid of the TGCL21/RAG�/� mice (C),but not in the thyroid of RAG�/� mice (B). A large cluster of GFP� cells 60 days after transfer (D), with T cells present predominantly in the right partof the infiltrate (yellow) and B cells (blue) in the left side (E). Expression of PNAd (red) is associated with the presence of large clusters of GFP� cells(F). Shown in the inset of F is a higher magnification of the PNAd-positive vessel marked by the arrow. Original magnification: A, �40; B–F, �100; insetin F, �400.
FIGURE 6. L-selectin�/� T cells can enter the thyroid of TGCCL21/RAG�/� mice. Immunohistological analysis of the thyroid of TGCCL21/RAG�/� mice transferred with 107 L-selectin�/� (A) and L-selectin�/� (B)splenocytes. Thyroids were collected 5 days after cell transfer and stainedwith anti-CD3 Ab. Original magnification: A–B, �100.
4795The Journal of Immunology
by guest on April 4, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

or the nature of the factors regulating the influx and the fate of thecells infiltrating the thyroid gland. In the present report we testedthe hypothesis that expression of CCL21 is sufficient to promoteleukocyte entry into the thyroid. CCL21 is expressed in autoim-mune thyroid disease (26) and has been implicated in the devel-opment of ectopic lymphoid tissues in this organ (20, 26, 50). Weshow that expression of CCL21 in the thyroid of transgenic micepromotes recruitment of T and B lymphocytes to the thyroid. Mostof the cells are naive, and are arranged in discrete compartments,resembling those observed in lymph nodes.
In lymphoid tissue, trafficking of naive T and B cells is regulatedby chemokines acting at the level of specialized endothelial struc-tures known as HEV. Chemokines such as CCL21 (51, 52) andCXCL13 (53) are expressed in the HEVs of lymph nodes andPeyer’s patches and trigger the activation of integrins present inlymphocytes, a process that depends on L-selectin and CCR7 (54,55). Because our histological analysis showed the presence ofHEVs within the thyroid of TGCCL21 mice, we initially askedwhether these structures were directly induced by CCL21 and es-sential for the influx of lymphocytes into the thyroid. Our studiesshowed that HEVs are not formed within the thyroid of TGCCL21mice that lack T and B cells (TGCCL21/RAG�/� mice), indicat-ing that CCL21 expressed in the thyroid does not directly inducethe differentiation of thyroid endothelial cells. Our studies alsoshow that adoptively transferred lymphocytes infiltrate theTGCCL21/RAG�/� thyroids, suggesting that infiltration of lym-phocytes is independent of HEV formation. As shown here, 60days after transfer, vessels expressing PNAd are found within thelarge clusters of adoptively transferred cells. These results suggestthat HEV formation may depend on the infiltrating cells and, thus,may be a later event on the formation of the lymphoid aggregates.The exact contribution of the incoming cells to the formation of theHEVs remains to be determined.
Unexpectedly, we found that the initial recruitment of lympho-cyte in CCL21 expressing thyroid was L-selectin independent. Al-though this finding is consistent with the observation that lympho-cytes can infiltrate the thyroids of TGCCL21/RAG�/� mice,
which lack expression of the L-selectin ligand PNAd, it under-scores a major difference between the thyroid gland and peripherallymph nodes in which the recruitment of naive lymphocyteslargely depends on L-selectin. Whether �4 integrins and their li-gands, which can mediate rolling interactions similar to the selec-tins (56), contribute to lymphocyte recruitment is a likely possi-bility that will be tested in future studies.
We also show that the main chemokine receptor mediatingCCL21 activity in the thyroid is CCR7 because mice that lackCCR7 expression did not exhibit thyroid infiltrates. Our resultssuggest that expression of the other murine CCL21 receptors(CXCR3 and CCX-CKR) is not sufficient to promote lymphocytemigration to the thyroid. However, whether these receptors haveany role in migration of lymphocytes to the thyroid during inflam-matory conditions remains to be tested.
Given the central role of LT� in lymphoid tissue development(49, 59–61), we wondered whether LT� was required for CCL21-induced lymphoid neogenesis. We have evaluated this hypothesisby crossing TGCCL21 mice with mice deficient in LT� (35). In-terestingly, mice deficient in LT�, expressing CCL21 in the thy-roid (TGCCL21/LT��/�) had mononuclear infiltrates with sepa-rate B and T cells areas and HEV formation that wasindistinguishable from TGCCL21/LT��/� mice. Similarly, micedeficient in LT�, expressing CCL21 in the pancreas also had in-filtrates with clear segregation of T and B cells (our unpublishedresults). Finally, treatment of RIP-CCL21 mice with LT�R-Fc af-fected the expression of PNAd and mucosal addressin cell adhe-sion molecule-1 on HEV but had a minor effect in reducing thecellularity of pancreatic infiltrates induced by CCL21 (29). In lightof these combined results, we suggest that LT�-independent path-ways control development and organization of specific lymphoidaggregates induced by CCL21. The existence of LT�-independentpathways has been invoked to explain development of nasopha-ryngeal-associated lymphoid tissue (62, 63). Initiation of nasopha-ryngeal-associated lymphoid tissue development happens mostlyafter birth and, similar to what was observed here, it depends onLT� and L-selectin. Surprisingly, ectopic lymphoid neogenesis in-duced by another homeostatic chemokine (CXCL13) clearly re-quires LT� (64). Our results raise the intriguing possibility that therequirements for induction of ectopic lymphoid neogenesis in-duced by these two chemokines may involve different signalingpathways.
Although it is indisputable that lymphocytes play a role in thepathogenesis of AITD, the function of the lymph node-like struc-tures often found in the thyroid in AITD remains unresolved. It hasbeen proposed that the presence of lymphoid follicles in areas richin Ag may represent a site for magnification of the immune re-sponse (20). We observed the presence of organized lymphoidaggregates in the thyroid of TGCCL21 mice that resemble thosefound in the thyroids of patients with AITD. Despite this similar-ity, none of the transgenic mice examined (n � 50) developedsigns of thyroid dysfunction (growth abnormalities and infertility)within the first year of life. Serum thyroid hormone levels, as wellas levels of TSH (which are elevated in hypothyroidism), did notdiffer between control and transgenic mice (data not shown). Wealso screened for TG autoantibodies in the serum of nontreatedtransgenic (n � 15) and wild-type littermates (n � 12) (ages 65–381 days), and found no differences between these groups. Thus,the presence of lymphocytes organized into follicular structureswithin the thyroid was not sufficient to initiate autoimmunity inTGCCL21 mice generated in the B6D2 background (MHCH-2b,d). Failure to develop autoimmunity may be due to multiplecauses, including lack of proper lymphocyte activation and/or ac-tive lymphocyte suppression. Studies to examine the susceptibility
FIGURE 7. Lymphocyte infiltration in the thyroid of TGCCL21 ani-mals is independent of LT�. Cell infiltrates in the thyroid of TGCCL21/LT��/� (A) and TGCCL21/LT��/� mice (B). Notice the presence andtopological segregation of T cells (CD3�, green) and B cells (B220�, red).PNAd-positive HEVs were present in both TGCCL21/LT��/� (C) andTGCCL21/LT��/� (D) thyroids. Original magnification: A–B, �100;C–D, �200.
4796 TRANSGENIC EXPRESSION OF CCL21 IN THE THYROID
by guest on April 4, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

of the TGCCL21 mice to autoimmunity and their response to con-ventional immunization will require backcrossing of the TG-GCCL21 mice into susceptible genetic backgrounds.
The mechanisms whereby CCL21 induces lymphoid neogenesisare not known, but appear to be context dependent. When over-expressed in the skin (27) or brain (30), CCL21 does not promotelymphocyte recruitment and organization. However, when ex-pressed in the thyroid or pancreatic islets (27–29), CCL21 inducesdevelopment of infiltrates rich in T and B cells, which segregateinto separate compartments, with T cells at the center of the infil-trates. These infiltrates also contain CD11c� dendritic cells andvery small numbers of macrophages (F4/80� cells) (27). The rea-son for this apparent tissue specificity is not clear, and may includeprocessing of CCL21, presence of CCL21-responding cells thatcould promote presentation or processing of CCL21 in the endo-thelium of “susceptible tissues”, or the responsiveness of the localendothelium.
The development of the models described here (in particular, theTGCCL21/RAG�/� mice) should now facilitate the characterizationof the mechanisms used by CCL21 to promote lymphocyte entry intononlymphoid tissue. For instance, it will now be possible to define thephenotype and developmental properties of the cells initially recruitedinto the thyroid. Fetal lymphoid CD4�CD3�ROR�t�IL-7r�� cells,also known as lymphoid tissue inducer cells have been implicated inthe development of lymphoid structures during embryogenesis (65,66). Cells with similar properties may exist at reduced numbers inadult mice, or may be produced in specific inflammatory settings. Themodel described here will allow us to define if lymphoid tissue in-ducer cells are functionally relevant for the CCL21-induced formationof ectopic lymphoid tissue in adult mice.
In summary, we have shown that CCL21, a chemokine ex-pressed in AITD, is sufficient to drive naive T and B cell recruit-ment into the thyroid. This process is mediated by CCR7 and re-sults in accumulation and topological arrangement of lymphocyteswithin the thyroid. Adoptive transfer experiments indicate that theinitial recruitment of lymphocytes is independent of L-selectin andthe presence of HEVs. Once the cells infiltrate the adult thyroid,aggregates are formed with segregation of T and B cells into spe-cific compartments, resembling the ectopic, lymph node-like struc-tures observed in many autoimmune and chronic inflammatoryconditions, including AITD. The generation of the genetic modelsdescribed here will facilitate the analysis of the mechanisms lead-ing to the formation of these lymphoid structures in adult animalsand to the definition of their role in disease.
AcknowledgmentsWe thank P. Zalamea, Andrei Golovko, and David Kinsley for expert tech-nical assistance. We are grateful to Jay Unkeless and Terry Davies forcritical comments to the manuscript.
References1. Juan, C. J. 2003. Endocrine autoimmunity. In Basic and Clinical Endocrinology.
F. S. Greenspan, ed. Lange, New York.2. Prummel, M. F., and W. M. Wiersinga. 2002. Autoimmune thyroid diseases. In
Immmunologically Mediated Endocrine Diseases. R.G. Gill, J. T. Harmon, andN. K. Maclaren, eds. Lippincott Williams & Wilkins, Philadelphia, p. 373.
3. Davies, T. F. 2000. The pathogenesis of Graves’ disease. In Wernere and Ing-bar’s The Thyroid. L. E. Baverman and R. D. Utiger, eds. Lippincott, Philadel-phia, p. 518.
4. Barbesino, G., and T. F. Davies. 2002. The immunological mediation of RobertGraves’ disease. In Immmunologically Mediated Endocrine Diseases. R. G. Gill,J. T. Harmon, and N. K. Maclaren, eds. Lippincott Williams & Wilkins, Phila-delphia, p. 397.
5. Stassi, G., and R. De Maria. 2002. Autoimmune thyroid disease: new models ofcell death in autoimmunity. Nat. Rev. Immunol. 2:195.
6. Baggiolini, M. 1998. Chemokines and leukocyte traffic. Nature 392:565.7. Zlotnik, A., and O. Yoshie. 2000. Chemokines: a new classification system and
their role in immunity. Immunity 12:121.
8. Farber, J. M. 1997. Mig and IP-10: CXC chemokines that target lymphocytes.J. Leukocyte Biol. 61:246.
9. Romagnani, P., M. Rotondi, E. Lazzeri, L. Lasagni, M. Francalanci,A. Buonamano, S. Milani, P. Vitti, L. Chiovato, M. Tonacchera, et al. 2002.Expression of IP-10/CXCL10 and MIG/CXCL9 in the thyroid and increasedlevels of IP-10/CXCL10 in the serum of patients with recent-onset Graves’ dis-ease. Am. J. Pathol. 161:195.
10. Bjorkdahl, O., K. A. Barber, S. J. Brett, M. G. Daly, C. Plumpton,N. A. Elshourbagy, J. P. Tite, and L. L. Thomsen. 2003. Characterization ofCC-chemokine receptor 7 expression on murine T cells in lymphoid tissues.Immunology 110:170.
11. Constantin, G., M. Majeed, C. Giagulli, L. Piccio, J. Y. Kim, E. C. Butcher, andC. Laudanna. 2000. Chemokines trigger immediate �2 integrin affinity and mo-bility changes: differential regulation and roles in lymphocyte arrest under flow.Immunity 13:759.
12. Forster, R., A. Schubel, D. Breitfeld, E. Kremmer, I. Renner-Muller, E. Wolf, andM. Lipp. 1999. CCR7 coordinates the primary immune response by establishingfunctional microenvironments in secondary lymphoid organs. Cell 99:23.
13. Luther, S. A., H. L. Tang, P. L. Hyman, A. G. Farr, and J. G. Cyster. 2000.Coexpression of the chemokines ELC and SLC by T zone stromal cells anddeletion of the ELC gene in the plt/plt mouse. Proc. Natl. Acad. Sci. USA97:12694.
14. Gunn, M. D., S. Kyuwa, C. Tam, T. Kakiuchi, A. Matsuzawa, L. T. Williams, andH. Nakano. 1999. Mice lacking expression of secondary lymphoid organ che-mokine have defects in lymphocyte homing and dendritic cell localization.J. Exp. Med. 189:451.
15. Muller, G., U. E. Hopken, H. Stein, and M. Lipp. 2002. Systemic immunoregu-latory and pathogenic functions of homeostatic chemokine receptors. J. Leuko-cyte Biol. 72:1.
16. Aust, G., M. Steinert, S. Kiessling, M. Kamprad, and C. Simchen. 2001. Reducedexpression of stromal-derived factor 1 in autonomous thyroid adenomas and itsregulation in thyroid-derived cells. J. Clin. Endocrinol. Metab. 86:3368.
17. Aust, G., M. Steinert, C. Boltze, S. Kiessling, and C. Simchen. 2001. GRO-� innormal and pathological thyroid tissues and its regulation in thyroid-derived cells.J. Endocrinol. 170:513.
18. Garcia-Lopez, M. A., D. Sancho, F. Sanchez-Madrid, and M. Marazuela. 2001.Thyrocytes from autoimmune thyroid disorders produce the chemokines IP-10and Mig and attract CXCR3� lymphocytes. J. Clin. Endocrinol. Metab. 86:5008.
19. Kemp, E. H., R. A. Metcalfe, K. A. Smith, M. N. Woodroofe, P. F. Watson, andA. P. Weetman. 2003. Detection and localization of chemokine gene expressionin autoimmune thyroid disease. Clin. Endocrinol. 59:207.
20. Armengol, M. P., M. Juan, A. Lucas-Martin, M. T. Fernandez-Figueras,D. Jaraquemada, T. Gallart, and R. Pujol-Borrell. 2001. Thyroid autoimmunedisease: demonstration of thyroid antigen-specific B cells and recombination-activating gene expression in chemokine-containing active intrathyroidal germi-nal centers. Am. J. Pathol. 159:861.
21. Simchen, C., I. Lehmann, D. Sittig, M. Steinert, and G. Aust. 2000. Expressionand regulation of regulated on activation, normal T cells expressed and secretedin thyroid tissue of patients with Graves’ disease and thyroid autonomy and inthyroid-derived cell populations. J. Clin. Endocrinol. Metab. 85:4758.
22. Ashhab, Y., O. Dominguez, M. Sospedra, C. Roura-Mir, A. Lucas-Martin, andR. Pujol-Borrell. 1999. A one-tube polymerase chain reaction protocol demon-strates CC chemokine overexpression in Graves’ disease glands. J. Clin. Endo-crinol. Metab. 84:2873.
23. Aust, G., D. Sittig, M. Steinert, P. Lamesch, and T. Lohmann. 2002. Graves’disease is associated with an altered CXCR3 and CCR5 expression in thyroid-derived compared to peripheral blood lymphocytes. Clin. Exp. Immunol.127:479.
24. Weetman, A. P., G. L. Bennett, and W. L. Wong. 1992. Thyroid follicular cellsproduce interleukin-8. J. Clin. Endocrinol. Metab. 75:328.
25. Kasai, K., N. Banba, S. Motohashi, Y. Hattori, K. Manaka, and S. I. Shimoda.1996. Expression of monocyte chemoattractant protein-1 mRNA and protein incultured human thyrocytes. FEBS Lett. 394:137.
26. Armengol, M. P., C. B. Cardoso-Schmidt, M. Fernandez, X. Ferrer,R. Pujol-Borrell, and M. Juan. 2003. Chemokines determine local lymphoneo-genesis and a reduction of circulating CXCR4� T and CCR7 B and T lympho-cytes in thyroid autoimmune diseases. J. Immunol. 170:6320.
27. Chen, S. C., G. Vassileva, D. Kinsley, S. Holzmann, D. Manfra,M. T. Wiekowski, N. Romani, and S. A. Lira. 2002. Ectopic expression of themurine chemokines CCL21a and CCL21b induces the formation of lymph node-like structures in pancreas, but not skin, of transgenic mice. J. Immunol.168:1001.
28. Fan, L., C. R. Reilly, Y. Luo, M. E. Dorf, and D. Lo. 2000. Cutting edge: ectopicexpression of the chemokine TCA4/SLC is sufficient to trigger lymphoid neo-genesis. J. Immunol. 164:3955.
29. Luther, S. A., A. Bidgol, D. C. Hargreaves, A. Schmidt, Y. Xu, J. Paniyadi,M. Matloubian, and J. G. Cyster. 2002. Differing activities of homeostatic che-mokines CCL19, CCL21, and CXCL12 in lymphocyte and dendritic cell recruit-ment and lymphoid neogenesis. J. Immunol. 169:424.
30. Chen, S. C., M. W. Leach, Y. Chen, X. Y. Cai, L. Sullivan, M. Wiekowski,B. J. Dovey-Hartman, A. Zlotnik, and S. A. Lira. 2002. Central nervous systeminflammation and neurological disease in transgenic mice expressing the CC che-mokine CCL21 in oligodendrocytes. J. Immunol. 168:1009.
31. Vassileva, G., H. Soto, A. Zlotnik, H. Nakano, T. Kakiuchi, J. A. Hedrick, andS. A. Lira. 1999. The reduced expression of 6Ckine in the plt mouse results fromthe deletion of one of two 6Ckine genes. J. Exp. Med. 190:1183.
4797The Journal of Immunology
by guest on April 4, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

32. Mann, J. R., and A. P. McMahon. 1993. Factors influencing production frequencyof transgenic mice. Methods Enzymol. 225:771.
33. Hogan, B., F. Constantini, and L. Lacy. 1986. Manipulating the Mouse Embryo.Cold Spring Harbor Laboratory Press, Cold Spring Harbor.
34. Lira, S. A., R. A. Kinloch, S. Mortillo, and P. M. Wassarman. 1990. An upstreamregion of the mouse ZP3 gene directs expression of firefly luciferase specificallyto growing oocytes in transgenic mice. Proc. Natl. Acad. Sci. USA 87:7215.
35. Korner, H., M. Cook, D. S. Riminton, F. A. Lemckert, R. M. Hoek,B. Ledermann, F. Kontgen, B. Fazekas de St. Groth, and J. D. Sedgwick. 1997.Distinct roles for lymphotoxin-� and tumor necrosis factor in organogenesis andspatial organization of lymphoid tissue. Eur. J. Immunol. 27:2600.
36. Shinkai, Y., G. Rathbun, K. P. Lam, E. M. Oltz, V. Stewart, M. Mendelsohn,J. Charron, M. Datta, F. Young, A. M. Stall, et al. 1992. RAG-2-deficient micelack mature lymphocytes owing to inability to initiate V(D)J rearrangement. Cell68:855.
37. Manfra, D. J., S. C. Chen, T. Y. Yang, L. Sullivan, M. T. Wiekowski,S. Abbondanzo, G. Vassileva, P. Zalamea, D. N. Cook, and S. A. Lira. 2001.Leukocytes expressing green fluorescent protein as novel reagents for adoptivecell transfer and bone marrow transplantation studies. Am. J. Pathol. 158:41.
38. Robinson, S. D., P. S. Frenette, H. Rayburn, M. Cummiskey, M. Ullman-Cullere,D. D. Wagner, and R. O. Hynes. 1999. Multiple, targeted deficiencies in selectinsreveal a predominant role for P-selectin in leukocyte recruitment. Proc. Natl.Acad. Sci. USA 96:11452.
39. Yang, T. Y., S. C. Chen, M. W. Leach, D. Manfra, B. Homey, M. Wiekowski,L. Sullivan, C. H. Jenh, S. K. Narula, S. W. Chensue, and S. A. Lira. 2000.Transgenic expression of the chemokine receptor encoded by human herpesvirus8 induces an angioproliferative disease resembling Kaposi’s sarcoma. J. Exp.Med. 191:445.
40. Chiappetta, G., N. Fabien, A. Picone, D. Califano, C. Monaco, V. de Franciscis,G. Vecchio, and G. Santelli. 1996. Transgenic mice carrying the human KRASoncogene under the control of a thyroglobulin promoter: KRAS expression inthyroids analyzed by in situ hybridization. Oncol. Res. 8:85.
41. Rochefort, P., B. Caillou, F. M. Michiels, C. Ledent, M. Talbot,M. Schlumberger, F. Lavelle, R. Monier, and J. Feunteun. 1996. Thyroid pathol-ogies in transgenic mice expressing a human activated Ras gene driven by athyroglobulin promoter. Oncogene 12:111.
42. Santoro, M., G. Chiappetta, A. Cerrato, D. Salvatore, L. Zhang, G. Manzo,A. Picone, G. Portella, G. Santelli, G. Vecchio, and A. Fusco. 1996. Developmentof thyroid papillary carcinomas secondary to tissue-specific expression of theRET/PTC1 oncogene in transgenic mice. Oncogene 12:1821.
43. Wallace, H., A. R. Clarke, D. J. Harrison, M. L. Hooper, and J. O. Bishop. 1996.Ganciclovir-induced ablation non-proliferating thyrocytes expressing herpesvirusthymidine kinase occurs by p53-independent apoptosis. Oncogene 13:55.
44. Caturegli, P., M. Hejazi, K. Suzuki, O. Dohan, N. Carrasco, L. D. Kohn, andN. R. Rose. 2000. Hypothyroidism in transgenic mice expressing IFN-� in thethyroid. Proc. Natl. Acad. Sci. USA 97:1719.
45. Caturegli, P., N. R. Rose, M. Kimura, H. Kimura, and S. C. Tzou. 2003. Studieson murine thyroiditis: new insights from organ flow cytometry. Thyroid 13:419.
46. Muller, G., U. E. Hopken, and M. Lipp. 2003. The impact of CCR7 and CXCR5on lymphoid organ development and systemic immunity. Immunol. Rev. 195:117.
47. Gosling, J., D. J. Dairaghi, Y. Wang, M. Hanley, D. Talbot, Z. Miao, andT. J. Schall. 2000. Cutting edge: identification of a novel chemokine receptor thatbinds dendritic cell- and T cell-active chemokines including ELC, SLC, andTECK. J. Immunol. 164:2851.
48. Townson, J. R., and R. J. Nibbs. 2002. Characterization of mouse CCX-CKR, areceptor for the lymphocyte-attracting chemokines TECK/mCCL25, SLC/mCCL21 and MIP-3�/mCCL19: comparison to human CCX-CKR. Eur. J. Im-munol. 32:1230.
49. Mebius, R. E. 2003. Organogenesis of lymphoid tissues. Nat. Rev. Immunol.3:292.
50. Hjelmstrom, P., J. Fjell, T. Nakagawa, R. Sacca, C. A. Cuff, and N. H. Ruddle.2000. Lymphoid tissue homing chemokines are expressed in chronic inflamma-tion. Am. J. Pathol. 156:1133.
51. Nagira, M., T. Imai, R. Yoshida, S. Takagi, M. Iwasaki, M. Baba, Y. Tabira,J. Akagi, H. Nomiyama, and O. Yoshie. 1998. A lymphocyte-specific CC che-mokine, secondary lymphoid tissue chemokine (SLC), is a highly efficient che-moattractant for B cells and activated T cells. Eur. J. Immunol. 28:1516.
52. Gunn, M. D., K. Tangemann, C. Tam, J. G. Cyster, S. D. Rosen, andL. T. Williams. 1998. A chemokine expressed in lymphoid high endothelialvenules promotes the adhesion and chemotaxis of naive T lymphocytes. Proc.Natl. Acad. Sci. USA 95:258.
53. Ebisuno, Y., T. Tanaka, N. Kanemitsu, H. Kanda, K. Yamaguchi, T. Kaisho,S. Akira, and M. Miyasaka. 2003. Cutting edge: the B cell chemokine CXCchemokine ligand 13/B lymphocyte chemoattractant is expressed in the high en-dothelial venules of lymph nodes and Peyer’s patches and affects B cell traffick-ing across high endothelial venules. J. Immunol. 171:1642.
54. Campbell, J. J., K. E. Murphy, E. J. Kunkel, C. E. Brightling, D. Soler, Z. Shen,J. Boisvert, H. B. Greenberg, M. A. Vierra, S. B. Goodman, et al. 2001. CCR7expression and memory T cell diversity in humans. J. Immunol. 166:877.
55. Stein, J. V., S. F. Soriano, C. M’Rini, C. Nombela-Arrieta, G. G. de Buitrago,J. M. Rodriguez-Frade, M. Mellado, J. P. Girard, and A. C. Martinez. 2003.CCR7-mediated physiological lymphocyte homing involves activation of a ty-rosine kinase pathway. Blood 101:38.
56. Berlin, C., R. F. Bargatze, J. J. Campbell, U. H. von Andrian, M. C. Szabo,S. R. Hasslen, R. D. Nelson, E. L. Berg, S. L. Erlandsen, and E. C. Butcher. 1995.�4 integrins mediate lymphocyte attachment and rolling under physiologic flow.Cell 80:413.
57. Stein, J. V., A. Rot, Y. Luo, M. Narasimhaswamy, H. Nakano, M. D. Gunn,A. Matsuzawa, E. J. Quackenbush, M. E. Dorf, and U. H. von Andrian. 2000. TheCC chemokine thymus-derived chemotactic agent 4 (TCA-4, secondary lym-phoid tissue chemokine, 6Ckine, exodus-2) triggers lymphocyte function-associ-ated antigen 1-mediated arrest of rolling T lymphocytes in peripheral lymph nodehigh endothelial venules. J. Exp. Med. 191:61.
58. Soto, H., W. Wang, R. W. Strieter, N. G. Copeland, D. J. Gilbert, N. A. Jenkins,J. Hedrick, and A. Zlotnik. 1998. The CC chemokine 6Ckine binds the CXCchemokine receptor CXCR3. Proc. Natl. Acad. Sci. USA 95:8205.
59. Matsumoto, M., Y. X. Fu, H. Molina, and D. D. Chaplin. 1997. Lymphotoxin-�-deficient and TNF receptor-I-deficient mice define developmental and func-tional characteristics of germinal centers. Immunol. Rev. 156:137.
60. Ruddle, N. H. 1999. Lymphoid neo-organogenesis: lymphotoxin’s role in inflam-mation and development. Immunol. Res. 19:119.
61. Tumanov, A. V., D. V. Kuprash, and S. A. Nedospasov. 2003. The role of lym-photoxin in development and maintenance of secondary lymphoid tissues. Cyto-kine Growth Factor Rev. 14:275.
62. Harmsen, A., K. Kusser, L. Hartson, M. Tighe, M. J. Sunshine, J. D. Sedgwick,Y. Choi, D. R. Littman, and T. D. Randall. 2002. Cutting edge: organogenesis ofnasal-associated lymphoid tissue (NALT) occurs independently of lympho-toxin-� (LT �) and retinoic acid receptor-related orphan receptor-�, but the or-ganization of NALT is LT � dependent. J. Immunol. 168:986.
63. Fukuyama, S., T. Hiroi, Y. Yokota, P. D. Rennert, M. Yanagita, N. Kinoshita,S. Terawaki, T. Shikina, M. Yamamoto, Y. Kurono, and H. Kiyono. 2002. Ini-tiation of NALT organogenesis is independent of the IL-7R, LT�R, and NIKsignaling pathways but requires the Id2 gene and CD3�CD4�CD45� cells. Im-munity 17:31.
64. Luther, S. A., T. Lopez, W. Bai, D. Hanahan, and J. G. Cyster. 2000. BLCexpression in pancreatic islets causes B cell recruitment and lymphotoxin-depen-dent lymphoid neogenesis. Immunity 12:471.
65. Eberl, G., and D. R. Littman. 2003. The role of the nuclear hormone receptorROR�t in the development of lymph nodes and Peyer’s patches. Immunol. Rev.195:81.
66. Eberl, G., S. Marmon, M. J. Sunshine, P. D. Rennert, Y. Choi, and D. R. Littman.2004. An essential function for the nuclear receptor ROR�(t) in the generation offetal lymphoid tissue inducer cells. Nat. Immunol. 5:64.
4798 TRANSGENIC EXPRESSION OF CCL21 IN THE THYROID
by guest on April 4, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from