cell fate determination during tooth development and ... and graf 2009 (tooth development...lecular...
TRANSCRIPT

Cell Fate Determination During ToothDevelopment and Regeneration
Thimios A. Mitsiadis* and Daniel Graf
Teeth arise from sequential and reciprocal interactions between the oralepithelium and the underlying cranial neural crest-derived mesen-chyme. Their formation involves a precisely orchestrated series of mo-lecular and morphogenetic events, and gives us the opportunity to dis-cover and understand the nature of the signals that direct cell fates andpatterning. For that reason, it is important to elucidate how signalingfactors work together in a defined number of cells to generate thediverse and precise patterned structures of the mature functional teeth.Over the last decade, substantial research efforts have been directedtoward elucidating the molecular mechanisms that control cell fate deci-sions during tooth development. These efforts have contributed towardthe increased knowledge on dental stem cells, and observation ofthe molecular similarities that exist between tooth developmentand regeneration. Birth Defects Research (Part C) 87:199–211,2009. VC 2009 Wiley-Liss, Inc.
Key words: tooth; incisor; cell fates; ameloblasts; odontoblasts; notchsignaling; bone morphogenetic protein (BMP); craniofacial develop-ment; dental defects; islet1; Tbx1; Pitx1
INTRODUCTIONMammalian tooth developmentproceeds through a series of well-defined morphological stages thatnecessitate sequential and recipro-cal interactions between the epi-thelium and mesenchyme (seeFig. 1). The mesenchyme isderived from cranial neural crestcells that form a uniform pool ofmultipotent progenitors. Thesecells migrate from the dorsal partof the neural tube and subse-quently acquire diverse cell fatesto generate structures of uniquemorphology and function, such asbone, cartilage, smooth muscle,neurons, and teeth (Anderson,1993; Weissman et al., 2001). In
mice, the first sign of tooth devel-opment is seen as a thickening ofthe oral epithelium at embryonicday 10.5 (E10.5), which invagi-nates the underlying mesenchymeand progressively forms the toothbud (E12.5–E13.5). The epithe-lium continues to grow and adoptthe cap (E14.5–E15.5) and bellconfigurations (E16.5–E18.5). Atthe late bell stage (E18.5), themesenchymal cells form the dentalfollicle and dental pulp. Pulp cellsadjacent to the dental epitheliumdifferentiate into odontoblasts andthe epithelial cells next to dentalpulp differentiate into ameloblasts(Bluteau et al., 2008). Odonto-blasts are columnar postmitotic
cells that form a layer in contactwith the dentin. Odontoblasticprocesses are formed at their dis-tal part, penetrate the dentin, andparticipate in the secretion of den-tin matrix and minerals. The ma-trix is composed of collagens(90%) and noncollagenous pro-teins, such as dentin sialophos-phoprotein and dentin matrix pro-tein 1. The deposition of apatiteminerals on this matrix gives riseto the mature calcified dentin.Enamel is secreted by ameloblastsalong the dentinoenamel junction.Enamel is mainly composed ofhydrophobic proteins, such asamelogenin, ameloblastin, ename-lin, amelotin, tuftelin, and odonto-genic ameloblast-associated pro-teins (Sire et al., 2007).Tissue recombination experi-
ments have shown that a molecu-lar dialogue starts once the neuralcrest-derived cells contact the oralepithelium (Fig. 2). The sameexperiments have also demon-strated that the inductive capacityfor mouse tooth formation residesin the epithelium until E12, afterwhich it shifts to the condensingmesenchyme (Mina and Kollar,1987). The importance of cranialneural crest-derived cells in odon-togenesis has been shown inexperiments where transplantationof mouse neural crest cells intochick embryos allowed growth oftooth germs (Mitsiadis et al.,
REVIE
W
VC 2009 Wiley-Liss, Inc.
Birth Defects Research (Part C) 87:199–211 (2009)
Thimios A. Mitsiadis and Daniel Graf are from the Institute of Oral Biology, ZZMK, Faculty of Medicine, University of Zurich, 8032Zurich, Switzerland
Supported by grants from the University of Zurich and the Swiss National Foundation.
*Correspondence to: Thimios A. Mitsiadis, Institute of Oral Biology, ZZMK, Faculty of Medicine, University of Zurich, Plattenstrasse11, 8032 Zurich, Switzerland. E-mail: [email protected]
Published online in Wiley InterScience (www.interscience.wiley.com). DOI: 10.1002/bdrc.20160

2003b, 2006). Molecular and cel-lular interactions control all stepsof tooth formation by coordinatingthe diverse cellular processes,such as cell proliferation, apopto-
sis, cytoskeleton modifications,terminal differentiation, and extra-cellular matrix synthesis. Thesame signals are repetitively usedduring the different stages of
odontogenesis. These signalsemerge in various developmentalcontexts: for example, the samesignal that activates proliferationin a cell type elsewhere will leadanother cell type to differentiateand in a third cell type will triggerapoptosis. The result is deter-mined by the developmental stateof a cluster of cells and dependson various cellular targets, such astranscription factors and cytoskel-eton proteins.
TIMING IN TOOTH
DEVELOPMENT
These developmental signalingevents must be precisely regu-lated. A signal that is produced atthe wrong time will lead to inap-propriate developmental respon-ses. Cell division, apoptosis, celldifferentiation, tissue growth andpatterns, and organ formationrequire proper timing. A moleculartiming mechanism exists, and itsfunction is to specify cell fates inall tissues and organs of the devel-oping embryo (Dorsky et al.,2000; Moss, 2007). Timing couldbe considered as the fourthdimension of embryonic develop-ment, but it is very difficult toobserve in vertebrates becausetheir developmental stages are notreally disconnected.Timing is thus an important
issue in tooth development and itsderegulation may have remarkableconsequences with a range ofeffects, from the generation ofpathologies (e.g., dental defects)to evolution. Changes in timingcould be one of the most impor-tant reasons in the evolution oftooth morphology. Although differ-ences in size and shape can beprecisely measured in teeth ofvarious species, the causal molec-ular mechanisms are complex anddifficult to define.
DENTAL CELL DIVERSITY—
CELL FATE SPECIFICATION
The generation of cell diversity is acentral issue in developmentalbiology. This process requiresnumerous cellular and geneticinteractions that result in the pro-
Figure 1. The different stages of human embryonic tooth development. In red the epi-thelium and its derivatives, in blue the mesenchyme and its derivatives. This figurewas obtained from an article by Bluteau et al., 2008.
Figure 2. (A) Schematic representation of the migration of cranial neural crest cellstoward the facial region and the oral cavity. (B) Section showing neural crest-derivedcells (red arrows) in contact with the oral epithelium. (C) Section showing exclusiveexpression of the midkine protein in dental tissues (red color). (D) Section demon-strating expression of Tbx1 in cells of the inner dental epithelium (violet color). r1,rhombomere 1; r2, rhombomere 2; r3, rhombomere 3.
200 MITSIADIS AND GRAF
Birth Defects Research (Part C) 87:199–211, (2009)

gressive restriction of cells to par-ticular cell phenotypes. Technicalprogress in biology, during recentyears, has allowed the analysis of
cell lineage relationships in thedeveloping teeth. These techni-ques include the genetic markingof cell fates (Trainor et al., 1999;
Chai et al., 2000) and lineagetracers for microinjection of dentalprogenitor cells (Mitsiadis et al.,2008b; Diep et al., 2009). Theresults have shown that dentalprecursor cells cause a variety ofcell types (Fig. 3). However, it isimportant to understand how thefate of uncommitted dental pro-genitor cells is influenced by fac-tors in their local environment.Factors expressed in early stagesof odontogenesis, such as bonemorphogenetic proteins (BMPs),fibroblast growth factors (FGFs),and Wnt molecules, may influencelineage commitment in two differ-ent ways: precursor cells are ei-ther instructed to choose a partic-ular fate, at the expense of others,in response to a given signal, orchoose their fate stochastically,and factors simply support theirsurvival and/or proliferation.Growth factor concentrations varyduring development and theresponse of the progenitors differsaccording to the concentration of agiven factor (Lillien and Gulacsi,2006). Tissue recombination andtransplantation experiments haveindicated that signals from theoral epithelium influence neuralcrest-derived mesenchyme cells toadopt a dental fate (Mina andKollar, 1987; Lumsden, 1988;Mitsiadis et al., 2003b). Indeed, inmany systems, developmentaldecisions involve one fate that ispromoted by a specific signal andan alternative fate that is assumedin the absence of that signal (Kellyand Melton, 1995) (Fig. 4).
THE FATE OF DENTAL
EPITHELIUM: GRADIENTS
AND SHAPES
The fate of the presumptive dentalepithelium is defined early duringembryonic development (E8.5) bythe bicoid-related transcriptionfactor, Pitx2 (Mucchielli et al.,1997; Mitsiadis et al., 1998b). Atmore advanced stages, Pitx2expression becomes restricted tothe epithelium of the tooth germs(Mucchielli et al., 1997). The pat-tern of the mammalian dentition isalso defined early, before any
Figure 3. Schematic representation of the acquisition of diverse cell fates during toothdevelopment.
Figure 4. Mesenchymal cell fates during odontogenesis (from cranial neural crest cellsto odontoblasts). Factors expressed during odontogenesis influence lineage commit-ment and instruct them to choose a particular fate, at the expense of others.
CELL FATE DETERMINATION IN ODONTOGENESIS 201
Birth Defects Research (Part C) 87:199–211, (2009)

obvious sign of tooth develop-ment. Parallel independent geneticpathways control development ofthe various groups of teeth: inci-sors, canines, premolars, andmolars. Dental diversity may bethe result of prepatterned cranialneural crest cell populations thatcontribute to the formation of pre-cise and specific tooth shapes.Alternatively, the morphologicalvariety may reflect the differentresponses of the neural crest-derived cells that colonize themandible and maxilla to signalsoriginated by the oral epithelium.Thus, rodent dentition patternmust be determined by a prepat-terning of either the oral epithe-lium into molar and incisor fieldsor the cranial neural crest-derivedcells as incisor and molar popula-tions.It is widely accepted that the
dental fields that define the num-ber and shape of teeth in the oralcavity are established by signalsoriginating from the oral epithe-lium. Their degree of concentra-tion determines the arrangementand fate of the responding cells.Signals emanating from a localizedsource may form morphogenicgradients that provide positionalinformation to populations ofuncommitted cells to decide theirdevelopmental fate accordingly(Green, 2002). This mechanismleads to the formation of distinc-tive cell types and consequently tothe generation of complex pat-terns and structures. Members ofthe transforming growth factor-b(TGFb), hedgehog (Hh), and Wntfamilies function as morphogens(Charron and Tessier-Lavigne,2007). In contrast, members ofthe FGF family have permissiverather than concentration-relatedeffects.Numerous signaling factors are
involved in different stages of em-bryonic tooth development. Forexample, BMP2, BMP4, and Mid-kine (MK) regulate epithelial–mes-enchymal interactions (Vainioet al., 1993; Kratochwil et al.,1996; Nadiri et al., 2004; Bluteauet al., 2008), FGF3, FGF4, FGF8,FGF9, and FGF10 are involved incell proliferation and regulate
expression of specific target genes(Bei and Maas, 1998; Kettunenet al., 1998, 2000). Wnt3, Wnt7b,Wnt10a, and Wnt10b are regula-tors of cell proliferation, migration,and differentiation during toothinitiation and morphogenesis(Dassule and McMahon, 1998),and sonic hedgehog (shh) contrib-utes to tooth initiation and mor-phogenesis (Khan et al., 2007).From all these molecules, BMP4and FGF8 constitute essentialearly oral epithelial signals thathave a crucial role in activatingspecific homeobox genes inthe underlying mesenchyme(Mitsiadis, 2001). It has been pro-posed that these two signals couldcontrol tooth patterning inrodents: BMP4 directs the shapeof incisors and FGF8 the shape ofmolars (Mitsiadis and Smith,2006). The mesenchyme of thedeveloping incisors expresses aspecific complement of genes(Msx1, Msx2), whereas the mes-enchyme of the molars possessesa different complement of genes(Dlx1, Dlx2, Barx1). The specificcomplement of these transcriptionfactors dictates the developmentof the tooth germs toward an inci-sorform or molariform shape(Cobourne and Mitsiadis, 2006).Based on the restricted expressiondomains of signaling moleculesand homeobox genes in thecranial neural crest cell-derivedmesenchyme of the maxilla andmandible, a ‘‘cooperative geneticinteraction’’ model has been pro-posed (Mitsiadis and Smith,2006).Expression of BMP4 in oral epi-
thelium is required for the expres-sion of Msx1 and Msx2 in theunderlying mesenchyme. Msx1has an important role in directingthe development of incisor mor-phogenesis and forms part of fewhomeobox genes that provide thespatial information for dental pat-terning (Cobourne and Mitsiadis,2006; Mitsiadis and Smith, 2006).On the other hand, FGF8 positivelyregulates the mesenchymalexpression of Barx1 and Dlx2 thatare essential molecules for molarmorphogenesis (Cobourne andMitsiadis, 2006). Previous studies
have suggested that Barx1 isinvolved in determining the iden-tity of neural crest-derived mesen-chymal cells in both maxilla andmandible (Tissier-Seta et al.,1995; Mucchielli et al., 1997;Mitsiadis et al., 1998b). Pax9 isalso expressed in the mesenchymethat underlies the locations of theepithelial invaginations (Neubuseret al., 1997). The presence of allthese transcription factors appearsto be required for a transcriptionalprogram responsible for the char-acteristic growth and morphologyof teeth (Cobourne and Mitsiadis,2006).
BMP Signaling Network
The reiterated requirement ofBMP signaling indicates the contin-uous need to probe cellular andmolecular interactions to directappropriate tooth development.BMP molecules encompass a largesubgroup of phylogenetically con-served signaling proteins belong-ing to the TGFb superfamily. Morethan 20 BMP-like molecules havebeen identified in vertebrates andinvertebrates. Based on thedegree of amino acid sequencehomology, they can be classifiedinto several subgroups. Two im-portant clusters are formed byBMP2/BMP4 and BMP5/BMP6/BMP7/BMP8. Although BMP mole-cules were originally identified asmolecules that induce bone andcartilage formation, they exert awide range of biological functions(e.g. cell differentiation, apopto-sis) in a variety of cell types dur-ing embryonic development. Infact, BMP signaling plays a pivotalrole in morphogenesis of most tis-sues and organs (Hogan, 1996;Zhao, 2003; Chen et al., 2004;Kishigami and Mishina, 2005).BMP molecules signal through
two different types of serine–thre-onine kinase receptors, termedBMPR type I and type II. Type Ireceptors encompass BMPRIA(Alk3), BMPRIB (Alk6), Activin re-ceptor type IA (ActRIA or Alk2),and Alk1 (Koenig et al., 1994; tenDijke et al., 1994b,c; Macias-Silvaet al., 1998; David et al., 2007),whereas BMPRII, ActRII, and
202 MITSIADIS AND GRAF
Birth Defects Research (Part C) 87:199–211, (2009)

ActRIIB are type II receptors(Kawabata et al., 1995; Rose-nzweig et al., 1995; Yamashitaet al., 1995). Individual BMP mole-cules have different affinities forthe various type I receptors (tenDijke et al., 1994a; Ebisawa et al.,1999), which are also influencedby the type II receptors (Yu et al.,2005). For this reason, the variousBMP proteins are not redundant,although it is not well understoodhow they achieve their distinctactivities in vivo.The BMP-associated signaling
cascades are divided into Smad-dependent and Smad-independentsignaling. Binding of BMP mole-cules to a preformed type I/typeII receptor heterodimer complexinduces Smad-dependent signal-ing. Smad-independent signalingis induced when the BMP firstbinds to an isolated type I recep-tor, with subsequent recruitmentof the type II receptor to the BMP/BMPRI complex (Nohe et al.,2002; Hassel et al., 2003; Hartunget al., 2006).The fine-tuning of BMP signaling
is in part achieved by interactionof BMP molecules with a variety ofsecreted proteins, mostly antago-nists, which are located in theextracellular space. The impor-tance of this extracellular regula-tion is illustrated by the severeand complex phenotypes observedin the absence of those antago-nists (Zhao, 2003).BMP antagonists can be classi-
fied into three subfamilies: theDAN family (cerberus, coco, grem-lin, ectodin, etc.), twisted gastru-lation, and the chordin and nogginsubgroup (Avsian-Kretchmer andHsueh, 2004). The different typesof BMP antagonists can bind toBMP molecules in diverse ways.For example, noggin binds BMP7in a clamp-like fashion and blocksbinding to both the BMPRI andBMPRII epitopes (Groppe et al.,2002), whereas chordin binds syn-ergistically with twisted gastrula-tion to form a ternary complex forefficient blocking (Graf et al.,2002; Yamamoto and Oelgeschl-ager, 2004). As the BMP networkis very complex, our understand-ing of its involvement in tooth de-
velopment is not well known andyet incomplete.Functional involvement of BMP
signaling in tooth developmentcould be studied in gene deletionmutants. Conditional deletion ofAlk3 (BMPR1A) in the epitheliumarrests tooth development at thebud stage, thus indicating the im-portance of mesenchyme-derivedBMP signals for the further devel-opment of tooth epithelium (Andlet al., 2004). This early arrest,however, obscures insight of Alk3function in the development oflater structures, such as the root,crown, enamel, or dentin. Bmp7-deficient mouse embryos oftenlack incisors, and occasionallymolars (Zouvelou et al., 2009). Incontrast, mutants deficient forthe BMP antagonist, ectodin, ex-hibit supernumerary teeth in theiroral cavity (Murashima-Suginamiet al., 2007). Overexpression ofnoggin in the epithelium results invarious phenotypic alterationsincluding lack of mandibularmolars, reduced number of maxil-lary molars, disrupted root sizeand pattern, and poorly mineral-ized enamel (Plikus et al., 2005).These studies reveal the involve-
ment of BMP signaling in the for-mation of the various dental struc-tures during the different stages ofodontogenesis. However, there isa clear lack of information con-cerning the roles of BMP signalingduring the establishment of cuspshapes, root formation, and gen-eration and maintenance of stemcells and/or progenitors in dentaltissues. A promising approach toaddress these issues is offered bystudies combining gene reportingwith cell-specific gene deletion(Graf and Economides, 2008; Zou-velou et al., 2009).
Islet1
Islet1 is a transcriptional regula-tor that plays critical roles in thecontrol of pattern formation andcell fate decisions of many varioustissues (Laugwitz et al., 2008). Is-let1 is expressed only in the inci-sor field of the oral epithelium,before the initiation of tooth for-mation (Mitsiadis et al., 2003a).
Expression of Islet1 during odon-togenesis is detected only in theepithelium of the incisors. Islet1expression in both proliferatingand differentiating epithelial cellsof the incisor indicates a potentialrole of Islet1 in the progression ofprogenitor cells from the dividing,undifferentiated state, to that ofpostmitotic ameloblasts. Theinduction of Islet1 in epithelium isintrinsic and not induced by theunderlying mesenchyme. Islet1and Bmp4 appear to have a posi-tive autoregulatory relationship(Mitsiadis et al., 2003a). Forcedexpression of Islet1 in the molarfield of the oral epithelium resultsin the loss of Barx1 expression inmesenchyme, leading to inhibitionof molar development (Mitsiadiset al., 2003a). Thus, Islet1 is partof the genetic program thatdefines territories associated withincisor formation in the oral epi-thelium.
THE FATE OF TOOTH
NUMBER
The size of the dental fields in theoral epithelium, and thus the pro-portion of the number of teeth, isestablished at the very earlystages of embryonic development.Ectodysplasin (Eda) signaling mol-ecules have been shown to beinvolved in the determinationof the dental fields (reviewed inMikkola, 2008). Indeed, disruptionof genes encoding for the Eda,Edar (Eda receptor), and Edaradd(an intracellular adaptor protein)proteins leads to mice with anirregular number of teeth.Increased Eda signaling has as aconsequence the formation ofsupernumerary teeth. In contrast,loss of function of Eda or Edarquite often leads to a reduction intooth number.
SIGNALS SPECIFYING
DENTAL CELL FATES
Signaling molecules and transcrip-tion factors are involved inthe determination of specific cellpopulations within dental tissues(Mitsiadis, 2001). During embryo-genesis, a subpopulation of oral
CELL FATE DETERMINATION IN ODONTOGENESIS 203
Birth Defects Research (Part C) 87:199–211, (2009)
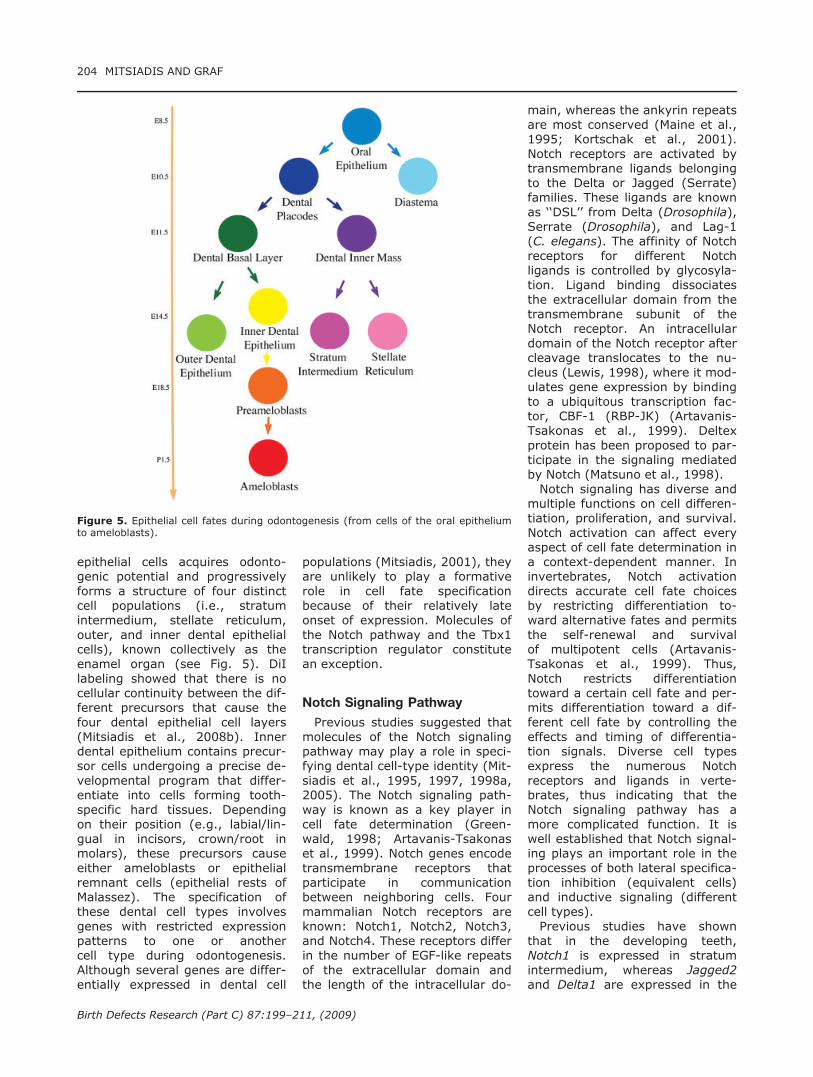
epithelial cells acquires odonto-genic potential and progressivelyforms a structure of four distinctcell populations (i.e., stratumintermedium, stellate reticulum,outer, and inner dental epithelialcells), known collectively as theenamel organ (see Fig. 5). DiIlabeling showed that there is nocellular continuity between the dif-ferent precursors that cause thefour dental epithelial cell layers(Mitsiadis et al., 2008b). Innerdental epithelium contains precur-sor cells undergoing a precise de-velopmental program that differ-entiate into cells forming tooth-specific hard tissues. Dependingon their position (e.g., labial/lin-gual in incisors, crown/root inmolars), these precursors causeeither ameloblasts or epithelialremnant cells (epithelial rests ofMalassez). The specification ofthese dental cell types involvesgenes with restricted expressionpatterns to one or anothercell type during odontogenesis.Although several genes are differ-entially expressed in dental cell
populations (Mitsiadis, 2001), theyare unlikely to play a formativerole in cell fate specificationbecause of their relatively lateonset of expression. Molecules ofthe Notch pathway and the Tbx1transcription regulator constitutean exception.
Notch Signaling Pathway
Previous studies suggested thatmolecules of the Notch signalingpathway may play a role in speci-fying dental cell-type identity (Mit-siadis et al., 1995, 1997, 1998a,2005). The Notch signaling path-way is known as a key player incell fate determination (Green-wald, 1998; Artavanis-Tsakonaset al., 1999). Notch genes encodetransmembrane receptors thatparticipate in communicationbetween neighboring cells. Fourmammalian Notch receptors areknown: Notch1, Notch2, Notch3,and Notch4. These receptors differin the number of EGF-like repeatsof the extracellular domain andthe length of the intracellular do-
main, whereas the ankyrin repeatsare most conserved (Maine et al.,1995; Kortschak et al., 2001).Notch receptors are activated bytransmembrane ligands belongingto the Delta or Jagged (Serrate)families. These ligands are knownas ‘‘DSL’’ from Delta (Drosophila),Serrate (Drosophila), and Lag-1(C. elegans). The affinity of Notchreceptors for different Notchligands is controlled by glycosyla-tion. Ligand binding dissociatesthe extracellular domain from thetransmembrane subunit of theNotch receptor. An intracellulardomain of the Notch receptor aftercleavage translocates to the nu-cleus (Lewis, 1998), where it mod-ulates gene expression by bindingto a ubiquitous transcription fac-tor, CBF-1 (RBP-JK) (Artavanis-Tsakonas et al., 1999). Deltexprotein has been proposed to par-ticipate in the signaling mediatedby Notch (Matsuno et al., 1998).Notch signaling has diverse and
multiple functions on cell differen-tiation, proliferation, and survival.Notch activation can affect everyaspect of cell fate determination ina context-dependent manner. Ininvertebrates, Notch activationdirects accurate cell fate choicesby restricting differentiation to-ward alternative fates and permitsthe self-renewal and survivalof multipotent cells (Artavanis-Tsakonas et al., 1999). Thus,Notch restricts differentiationtoward a certain cell fate and per-mits differentiation toward a dif-ferent cell fate by controlling theeffects and timing of differentia-tion signals. Diverse cell typesexpress the numerous Notchreceptors and ligands in verte-brates, thus indicating that theNotch signaling pathway has amore complicated function. It iswell established that Notch signal-ing plays an important role in theprocesses of both lateral specifica-tion inhibition (equivalent cells)and inductive signaling (differentcell types).Previous studies have shown
that in the developing teeth,Notch1 is expressed in stratumintermedium, whereas Jagged2and Delta1 are expressed in the
Figure 5. Epithelial cell fates during odontogenesis (from cells of the oral epitheliumto ameloblasts).
204 MITSIADIS AND GRAF
Birth Defects Research (Part C) 87:199–211, (2009)
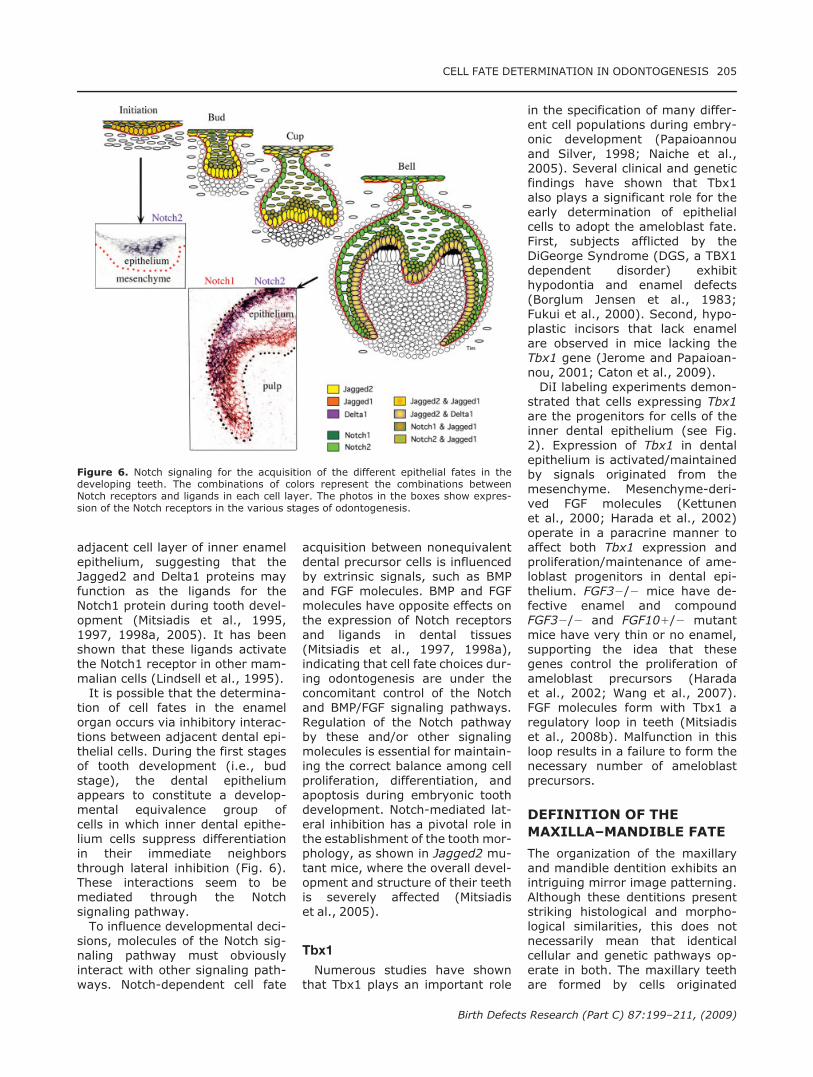
adjacent cell layer of inner enamelepithelium, suggesting that theJagged2 and Delta1 proteins mayfunction as the ligands for theNotch1 protein during tooth devel-opment (Mitsiadis et al., 1995,1997, 1998a, 2005). It has beenshown that these ligands activatethe Notch1 receptor in other mam-malian cells (Lindsell et al., 1995).It is possible that the determina-
tion of cell fates in the enamelorgan occurs via inhibitory interac-tions between adjacent dental epi-thelial cells. During the first stagesof tooth development (i.e., budstage), the dental epitheliumappears to constitute a develop-mental equivalence group ofcells in which inner dental epithe-lium cells suppress differentiationin their immediate neighborsthrough lateral inhibition (Fig. 6).These interactions seem to bemediated through the Notchsignaling pathway.To influence developmental deci-
sions, molecules of the Notch sig-naling pathway must obviouslyinteract with other signaling path-ways. Notch-dependent cell fate
acquisition between nonequivalentdental precursor cells is influencedby extrinsic signals, such as BMPand FGF molecules. BMP and FGFmolecules have opposite effects onthe expression of Notch receptorsand ligands in dental tissues(Mitsiadis et al., 1997, 1998a),indicating that cell fate choices dur-ing odontogenesis are under theconcomitant control of the Notchand BMP/FGF signaling pathways.Regulation of the Notch pathwayby these and/or other signalingmolecules is essential for maintain-ing the correct balance among cellproliferation, differentiation, andapoptosis during embryonic toothdevelopment. Notch-mediated lat-eral inhibition has a pivotal role inthe establishment of the tooth mor-phology, as shown in Jagged2 mu-tant mice, where the overall devel-opment and structure of their teethis severely affected (Mitsiadiset al., 2005).
Tbx1
Numerous studies have shownthat Tbx1 plays an important role
in the specification of many differ-ent cell populations during embry-onic development (Papaioannouand Silver, 1998; Naiche et al.,2005). Several clinical and geneticfindings have shown that Tbx1also plays a significant role for theearly determination of epithelialcells to adopt the ameloblast fate.First, subjects afflicted by theDiGeorge Syndrome (DGS, a TBX1dependent disorder) exhibithypodontia and enamel defects(Borglum Jensen et al., 1983;Fukui et al., 2000). Second, hypo-plastic incisors that lack enamelare observed in mice lacking theTbx1 gene (Jerome and Papaioan-nou, 2001; Caton et al., 2009).DiI labeling experiments demon-
strated that cells expressing Tbx1are the progenitors for cells of theinner dental epithelium (see Fig.2). Expression of Tbx1 in dentalepithelium is activated/maintainedby signals originated from themesenchyme. Mesenchyme-deri-ved FGF molecules (Kettunenet al., 2000; Harada et al., 2002)operate in a paracrine manner toaffect both Tbx1 expression andproliferation/maintenance of ame-loblast progenitors in dental epi-thelium. FGF32/2 mice have de-fective enamel and compoundFGF32/2 and FGF101/2 mutantmice have very thin or no enamel,supporting the idea that thesegenes control the proliferation ofameloblast precursors (Haradaet al., 2002; Wang et al., 2007).FGF molecules form with Tbx1 aregulatory loop in teeth (Mitsiadiset al., 2008b). Malfunction in thisloop results in a failure to form thenecessary number of ameloblastprecursors.
DEFINITION OF THE
MAXILLA–MANDIBLE FATE
The organization of the maxillaryand mandible dentition exhibits anintriguing mirror image patterning.Although these dentitions presentstriking histological and morpho-logical similarities, this does notnecessarily mean that identicalcellular and genetic pathways op-erate in both. The maxillary teethare formed by cells originated
Figure 6. Notch signaling for the acquisition of the different epithelial fates in thedeveloping teeth. The combinations of colors represent the combinations betweenNotch receptors and ligands in each cell layer. The photos in the boxes show expres-sion of the Notch receptors in the various stages of odontogenesis.
CELL FATE DETERMINATION IN ODONTOGENESIS 205
Birth Defects Research (Part C) 87:199–211, (2009)

from midbrain and forebrain neu-ral crest, whereas the mandibleteeth receive neural crest cellsderived from hindbrain (rhombo-meres 1 and 2) and midbrain(Osumi-Yamashita et al., 1994;Trainor and Tam, 1995; Imaiet al., 1996; Cobourne and Mitsia-dis, 2006) (Fig. 2). Mutations intranscription factors, such as Lef1,Msx1, Pitx2, and Pax9, result indevelopmental arrest of all teeth,indicating that these genes partici-pate in processes common to de-velopment of all teeth (reviewed inMitsiadis, 2001). In contrast, sev-eral transcription factors, such asDlx, are differentially expressed inthe maxillary and mandible proc-esses (Zhao et al., 2000), thusindicating a basic genetic differ-ence between upper and lowermolar specification. The failure ofmaxillary molar development indouble mutants that lack bothDlx1 and Dlx2 (Dlx1/2) genes sug-gests a specific role for thesegenes in regional specification of
the odontogenic mesenchyme (Qiuet al., 1997).Pitx1 is another gene controlling
tooth identity in the mandible andmaxilla. Pitx1 is initially expressedin the proximal mesenchyme ofthe developing mandible wheremolars will develop. During odon-togenesis, Pitx1 is exclusivelyexpressed in dental epithelium(Mitsiadis and Drouin, 2008). Theexpression of Pitx1 in both man-dibular mesenchyme and dentalepithelium is negatively regulatedby BMP4 (St Amand et al., 2000;Mitsiadis and Drouin, 2008). InPitx1 mutant mice, mandiblemolars are smaller than in wild-type mice, their number of cuspsis reduced, and first and secondmolars fuse, most probablybecause of the delayed develop-ment of the second molars(Mitsiadis and Drouin, 2008). Themorphological change in thePitx12/2 mandible molars couldbe attributed to the shortening ofthe mandible and the consequent
narrowing of the field wheremolars develop. Disruption inexpansion and growth of the toothprimordia could reflect an alteredpatterning, resulting in a newshape and size of the teeth.Although a part of the genetic
code controlling tooth develop-ment (i.e., Fgf8, Bmp4, Shh,Msx1 and Msx2) is unaltered inPitx12/2 mice (St Amand et al.,2000), Tbx1 expression is com-pletely suppressed in the epithe-lium of the mutant teeth (Mitsiadisand Drouin, 2008). Pitx1 seems toplay a role in specifying a subpo-pulation of neural crest-derivedmesenchymal cells in the mandi-ble. In Pitx1 mutants, Barx1expression is only downregulatedin the mesenchyme of the mandib-ular molars (Mitsiadis and Drouin,2008). Thus, the odontogenicspecification of neural crest-derived mesenchymal cells may bepartly controlled by Pitx1 in themandible.Activin is one of the earliest
mesenchymal signals that conferpositional information for theexact location where the teeth willdevelop. Activin ßA, activin recep-tor ActR[IIA1/2;IIB2/2], andSmad21/2 mice also exhibit atooth phenotype, in which the inci-sors and mandibular molars aremissing but the maxillary molarsdevelop normally (Ferguson et al.,1998, 2001).
FATES OF DENTAL
STEM CELLS
The rodent incisor has a cylindricalshape and differs from other teethin that it is a continuously growingorgan that erupts throughout thelife of the animal (Mitsiadis et al.,2007). The incisor is formed bytwo distinct epithelia, the lingualand labial epithelia, which enclosethe mesenchyme of the pulp (seeFig. 7). The lingual side of the inci-sor is composed of two epithelialcell layers, the inner and the outerdental epithelia. Cementum is de-posited on this side of the incisorthat is known as the root analogueof the molar. The labial side of theincisor is comprised of four epithe-lial cell layers: the inner and outer
Figure 7. Sections through mouse incisors show the creation of different cell layers inepithelium (pseudocolors) and the expression of the Notch receptors in distinct cellpopulations (violet color).
206 MITSIADIS AND GRAF
Birth Defects Research (Part C) 87:199–211, (2009)

dental epithelia that surround thecore of stellate reticulum and stra-tum intermedium. The inner andouter enamel epithelia are in con-tact with the mesenchyme of thepulp and the follicle, respectively.Cells of the stratum intermediumare compressed, flattened cuboi-dal cells in contact with the innerdental epithelium. Enamel is de-posited on the labial side of the in-cisor, which is thus analogous tothe crown of a molar. The cervicalloop is located at the posterior endof the epithelium and consists of acore of undifferentiated cells sur-rounded by the inner and outerdental epithelia. Cells of the coreof this area proliferate and gener-ate the transit amplifying progeni-tor cells, which then differentiateinto all of the cells of the incisor,including the terminally differenti-ated ameloblasts (Mitsiadis et al.,2007; Bluteau et al., 2008).Cells responsible for the contin-
ual growth of the incisor arelocated in the cervical loop, sug-gesting that this area is the puta-tive stem cell niche (Mitsiadiset al., 2007). Cell tracking experi-ments have demonstrated that thecells from the cervical loop aremaintained in this area, whereasothers migrate toward the anteriorpart of the incisor where theydifferentiate into ameloblasts(Harada et al., 1999). A character-istic feature of stem cells is theirslow cycling nature. BrdU incorpo-ration studies have demonstratedthat the cells of the cervical loopregion are cycling slowly. Shortperiods of BrdU incubation haveshown that the cells of the innerdental epithelium are highly prolif-erative and cycling very quickly.In contrast, other cells of the cer-vical loop are labeled only verysparsely and divide slowly. Pulse-chase experiments have demon-strated that BrdU labeling stillremains in cells located in the in-ternal part of the cervical loop epi-thelium. These cells divide slowly,remain in the cervical loop, andmaintain the stem cell populationthrough self-renewal. Cells origi-nated from the cervical loop regionexhibit a clonal potential and havethe ability to differentiate into cells
expressing markers of the innerdental epithelium and amelo-blasts, such as the low-affinityneurotrophin receptor, P75NTFR,and amelogenin.In incisors, Notch receptors
and ligands are expressed inadjacent cell populations: Notch1 isexpressed in the stratum inter-medium, whereas Delta1, Jagged1,and Jagged2 are expressed in cellsof the inner dental epithelium andameloblasts (Mitsiadis et al., 1998a,2005, 2007; Harada et al., 1999).Notch receptors are also expressedin the cells of the cervical loop area,but their expression remains absentin cells of the inner dental epithe-lium and ameloblasts (see Fig. 7).Notch2 and lunatic fringe (Lnf), amodulator of Notch signaling, havebeen implicated in the rotation ofthe incisor that determines the lin-gual/labial asymmetry (Mucchielliand Mitsiadis, 2000; Pouyet andMitsiadis, 2000). These patterns ofexpression indicate that the Notchsignaling plays a role in the mainte-nance and determination of thedental stem cell fates.FGF molecules, such as FGF3
and FGF10, are important mesen-chymal signals involved in themaintenance of epithelial cell pop-ulations within the cervical loop(Harada et al., 2002). FGF10 isexpressed in the mesenchymethat surrounds the cervical loopepithelium and FGF3 expression isrestricted to the mesenchyme thatunderlies the inner dental epithe-lium. FGF10 seems to be involvedin the creation of the adult stemcell compartment in the cervicalloop region. Indeed, incisors fromthe FGF10 null mice are smallerand the cervical loop is missing.The reduced size of the mutant in-cisor is due to the lack of the stemcell compartment in the cervicalloop, rather than an overalldecrease of cell proliferation in theincisor. FGF10 has been shown toprevent apoptosis in the cervicalloop epithelium and induces differ-entiation of progenitor cells intocells of the stratum intermedium.Bead implantation experimentshave shown that FGF10 stimulatesLnf expression, thereby modulat-ing Notch signaling in the cervical
loop region (Harada et al., 2002).These findings suggest interac-tions between the FGF and Notchsignaling pathways for the mainte-nance of stem cells of the cervicalloop in an undifferentiated state(reviewed in Mitsiadis et al.,2007).
STEM CELL FATES DURING
TOOTH REPAIR AND
REGENERATION
Stem cells play a critical role in tis-sue homeostasis and repairthroughout life. Their fate betweenself-renewal and differentiation isregulated by both cell intrinsicdeterminants and signals from aspecialized microenvironment(Moore and Lemischka, 2006).Physical interactions and molecu-lar cross talk with stem cells, aswell as the orientation of thecleavage plane during their mito-sis, determine the fate of thesecells. Molecules of the Wnt, Notch,and BMP pathways have beenshown to control the balancebetween symmetric and asymmet-ric division of stem cells andrelated cell fate specification out-comes (Betschinger and Knoblich,2004; Lechler and Fuchs, 2005). Aderegulation in the control ofthese events could lead to anabnormal tissue formation and areduced tissue repair.The growing interest in the mo-
lecular regulation of stem cellsarises from the potential to influ-ence their fate and consequentlytheir functions during tissue repairand/or regeneration. The repara-tive mechanisms that operate fol-lowing dental injuries involve a se-ries of highly conserved processesthat share genetic programs thatoccur throughout embryogenesis(reviewed in Mitsiadis andRahiotis, 2004). In a severewound, necrotic odontoblasts arereplaced through a complex pro-cess by cells having stem cellproperties, to differentiate into anew generation of odontoblaststhat produce a novel dentin ma-trix, called reparative dentin (Mit-siadis and Rahiotis, 2004; Mitsia-dis et al., 2008a).
CELL FATE DETERMINATION IN ODONTOGENESIS 207
Birth Defects Research (Part C) 87:199–211, (2009)

Signaling molecules that areexpressed by pulp cells play a rolein both tooth homeostasis andrepair (reviewed in Smith andLesot, 2001). Signals present atthe injury site may act both as mi-togens or chemotactic factors forpulp cells and initiate the healingprocess. Notch, nestin, and cad-herins are also involved in thedynamic processes triggered bypulp injury and contribute to thesignaling cascade by coordinatingcell fate decisions, proliferative,migratory, and differentiationactivities (Mitsiadis et al., 1999;Mitsiadis and Rahiotis, 2004). Ininjured teeth, Notch receptors areexpressed by progenitors and un-differentiated cells of the subodon-toblastic layer, but not by newlyformed odontoblasts (see Fig. 8).Notch signaling reactivation duringpulp healing might enhance sur-vival of uncommitted precursors,while preserving multilineagepotential. Notch expression is acti-vated in cells close to the injurysite, as well as in cells located atthe apex of the roots, suggestingthat these sites represent impor-tant pools of cells from which dif-ferent cell types could derive afterinjury (Mitsiadis et al., 1999).Bone marrow stem cells expressNotch receptors (Calvi et al.,2003), suggesting that the Notch-positive cells of the pulp have atleast some stem cell properties.Activation of the Notch moleculesin endothelial cells after injurymay reflect another pool of stemcells (Lovschall et al., 2007). Acti-vation of Notch signaling duringtooth regeneration may ensure a
continuous balance between odon-toblasts and dental pulp progeni-tor cells. Aggregates of Notch-pos-itive cell populations could be usedfor transplantation in the injuredteeth. Local transplantation ofthese cells for therapeutic applica-tions may permit a more extensivereconstruction than in those inju-ries that would spontaneouslyheal.
CONCLUSIONS
To understand the control of cellfate decisions, cytodifferentiation,and morphogenesis is a challengethat necessitates a thorough eluci-dation of the cellular and molecu-lar events involved in initiation,development, repair, and regener-ation of teeth. Understandingwhen and how signaling moleculescontrol cell fate outcomes will pro-vide new tools with which to directthe differentiation of cells into par-ticular cell types. The identificationof dental epithelial and mesenchy-mal stem cells and the knowledgeof molecules involved in their fatedetermination is a considerableaccomplishment. The engineeringof tridimensional scaffolds with acomposition and shape more orless similar to that of the teeth toreconstruct and the addition ofsignaling molecules might facili-tate the transplantation and thedifferentiation of stem cells towardthe desired fate. This scientificknowledge is likely to instruct de-velopment of novel therapies inthe near future. However, themain challenge in cell therapy is tofind a compromise between the
benefits to the patients, ethicalissues, and state regulations.
ACKNOWLEDGMENTS
We are grateful to ProfessorHansueli Luder (Institute of OralBiology, ZZMK, University of Zur-ich) for providing the histologypictures of Figure 7.
REFERENCES
Anderson DJ. 1993. Molecular controlof cell fate in the neural crest: thesympathoadrenal lineage. Annu RevNeurosci 16:129–158.
Andl T, Ahn K, Kairo A, et al. 2004.Epithelial Bmpr1a regulates differen-tiation and proliferation in postnatalhair follicles and is essential for toothdevelopment. Development 131:2257–2268.
Artavanis-Tsakonas S, Rand MD, LakeRJ. 1999. Notch signaling: cell fatecontrol and signal integration indevelopment. Science 284:770–776.
Avsian-Kretchmer O, Hsueh AJ. 2004.Comparative genomic analysis of theeight-membered ring cystine knot-containing bone morphogenetic pro-tein antagonists. Mol Endocrinol 18:1–12.
Bei M, Maas R. 1998. FGFs and BMP4induce both Msx1-independent andMsx1-dependent signaling pathwaysin early tooth development. Develop-ment 125:4325–4333.
Betschinger J, Knoblich JA. 2004. Dare tobe different: asymmetric cell division inDrosophila, C. elegans and vertebrates.Curr Biol 14:R674–R685.
Bluteau G, Luder HU, De Bari C, Mitsia-dis TA. 2008. Stem cells for tooth en-gineering. Eur Cell Mater 16:1–9.
Borglum Jensen S, Jacobsen P, RotneL, et al. 1983. Oral findings inDiGeorge syndrome. Int J Oral Surg12:250–254.
Calvi LM, Adams GB, Weibrecht KW,et al. 2003. Osteoblastic cells regu-late the haematopoietic stem cellniche. Nature 425:841–846.
Caton J, Luder HU, Zoupa M, et al.2009. Enamel-free teeth: Tbx1 dele-tion affects amelogenesis in rodentincisors. Dev Biol 328:493–505.
Chai Y, Jiang X, Ito Y, et al. 2000. Fateof the mammalian cranial neuralcrest during tooth and mandibularmorphogenesis. Development 127:1671–1679.
Charron F, Tessier-Lavigne M. 2007.The Hedgehog, TGF-beta/BMP andWnt families of morphogens inaxon guidance. Adv Exp Med Biol621:116–133.
Chen D, Zhao M, Mundy GR. 2004.Bone morphogenetic proteins. GrowthFactors 22:233–241.
Cobourne MT, Mitsiadis T. 2006. Neuralcrest cells and patterning of the
Figure 8. Sections of an injured area of the tooth show the formation of a reparativedentin (arrows), and reexpression of nestin (brown color) in the newly formed odonto-blasts (o) and Notch2 (red color) in cells of the subodontoblastic layer (soc).
208 MITSIADIS AND GRAF
Birth Defects Research (Part C) 87:199–211, (2009)

mammalian dentition. J Exp Zoolog BMol Dev Evol 306:251–260.
Dassule HR, McMahon AP. 1998. Anal-ysis of epithelial-mesenchymal inter-actions in the initial morphogenesisof the mammalian tooth. Dev Biol202:215–227.
David L, Mallet C, Mazerbourg S, et al.2007. Identification of BMP9 andBMP10 as functional activators of theorphan activin receptor-like kinase 1(ALK1) in endothelial cells. Blood109:1953–1961.
Diep L, Matalova E, Mitsiadis TA,Tucker AS. 2009. Contribution of thetooth bud mesenchyme to alveolarbone. J Exp Zoolog B Mol Dev Evol312:510–517.
Dorsky RI, Moon RT, Raible DW. 2000.Environmental signals and cell fatespecification in premigratory neuralcrest. Bioessays 22:708–716.
Ebisawa T, Tada K, Kitajima I, et al.1999. Characterization of bone mor-phogenetic protein-6 signaling path-ways in osteoblast differentiation.J Cell Sci 112 (Part 20):3519–3527.
Ferguson CA, Tucker AS, ChristensenL, et al. 1998. Activin is an essentialearly mesenchymal signal in toothdevelopment that is required for pat-terning of the murine dentition.Genes Dev 12:2636–2649.
Ferguson CA, Tucker AS, HeikinheimoK, et al. 2001. The role of effectorsof the activin signaling pathway, acti-vin receptors IIA and IIB, andSmad2, in patterning of tooth devel-opment. Development 128:4605–4613.
Fukui N, Amano A, Akiyama S, et al.2000. Oral findings in DiGeorge syn-drome: clinical features and histo-logic study of primary teeth. OralSurg Oral Med Oral Pathol Oral RadiolEndod 89:208–215.
Graf D, Economides AN. 2008. Dissec-tion of BMP signaling using genomeengineering tools. In: Vukicevic S,Sampath KT, editors. Bone morpho-genetic proteins: from local to sys-temic therapeutics. Basel/Switzer-land: Birkhauser. pp. 115–139.
Graf D, Nethisinghe S, Palmer DB,et al. 2002. The developmentallyregulated expression of twisted gas-trulation reveals a role for bone mor-phogenetic proteins in the control ofT cell development. J Exp Med 196:163–171.
Green J. 2002. Morphogen gradients,positional information, and Xenopus:interplay of theory and experiment.Dev Dyn 225:392–408.
Greenwald I. 1998. LIN-12/Notch sig-naling: lessons from worms and flies.Genes Dev 12:1751–1762.
Groppe J, Greenwald J, Wiater E, et al.2002. Structural basis of BMP signal-ing inhibition by the cystine knot pro-tein Noggin. Nature 420:636–642.
Harada H, Kettunen P, Jung HS, et al.1999. Localization of putative stemcells in dental epithelium and their
association with Notch and FGF sig-naling. J Cell Biol 147:105–120.
Harada H, Toyono T, Toyoshima K,et al. 2002. FGF10 maintains stemcell compartment in developingmouse incisors. Development 129:1533–1541.
Hartung A, Bitton-Worms K, RechtmanMM, et al. 2006. Different routes ofbone morphogenic protein (BMP) re-ceptor endocytosis influence BMPsignaling. Mol Cell Biol 26:7791–7805.
Hassel S, Schmitt S, Hartung A, et al.2003. Initiation of Smad-dependentand Smad-independent signaling viadistinct BMP-receptor complexes. JBone Joint Surg Am 85 (Suppl 3):44–51.
Hogan BL. 1996. Bone morphogeneticproteins: multifunctional regulatorsof vertebrate development. GenesDev 10:1580–1594.
Imai H, Osumi-Yamashita N, NinomiyaY, Eto K. 1996. Contribution of early-emigrating midbrain crest cells to thedental mesenchyme of mandibularmolar teeth in rat embryos. Dev Biol176:151–165.
Jerome LA, Papaioannou VE. 2001.DiGeorge syndrome phenotype inmice mutant for the T-box gene,Tbx1. Nat Genet 27:286–291.
Kawabata M, Chytil A, Moses HL. 1995.Cloning of a novel type II serine/threonine kinase receptor throughinteraction with the type I transform-ing growth factor-beta receptor.J Biol Chem 270:5625–5630.
Kelly OG, Melton DA. 1995. Induction andpatterning of the vertebrate nervoussystem. Trends Genet 11: 273–278.
Kettunen P, Karavanova I, Thesleff I.1998. Responsiveness of developingdental tissues to fibroblast growthfactors: expression of splicing alter-natives of FGFR1, -2, -3, and ofFGFR4; and stimulation of cell prolif-eration by FGF-2, -4, -8, and -9. DevGenet 22:374–385.
Kettunen P, Laurikkala J, Itaranta P,et al. 2000. Associations of FGF-3and FGF-10 with signaling networksregulating tooth morphogenesis. DevDyn 219:322–332.
Khan M, Seppala M, Zoupa M,Cobourne MT. 2007. Hedgehog path-way gene expression during early de-velopment of the molar tooth root inthe mouse. Gene Expr Patterns 7:239–243.
Kishigami S, Mishina Y. 2005. BMP sig-naling and early embryonic pattern-ing. Cytokine Growth Factor Rev 16:265–278.
Koenig BB, Cook JS, Wolsing DH, et al.1994. Characterization and cloning ofa receptor for BMP-2 and BMP-4 fromNIH 3T3 cells. Mol Cell Biol 14:5961–5974.
Kortschak RD, Tamme R, Lardelli M.2001. Evolutionary analysis of verte-brate Notch genes. Dev Genes Evol211:350–354.
Kratochwil K, Dull M, Farinas I, et al.1996. Lef1 expression is activated byBMP-4 and regulates inductive tissueinteractions in tooth and hair devel-opment. Genes Dev 10:1382–1394.
Laugwitz KL, Moretti A, Caron L, et al.2008. Islet1 cardiovascular progeni-tors: a single source for heart line-ages? Development 135:193–205.
Lechler T, Fuchs E. 2005. Asymmetriccell divisions promote stratificationand differentiation of mammalianskin. Nature 437:275–280.
Lewis J. 1998. Notch signaling and thecontrol of cell fate choices in verte-brates. Semin Cell Dev Biol 9:583–589.
Lillien L, Gulacsi A. 2006. Environmen-tal signals elicit multiple responses indorsal telencephalic progenitors bythreshold-dependent mechanisms.Cereb Cortex 16(Suppl 1):i74–i81.
Lindsell CE, Shawber CJ, Boulter J,Weinmaster G. 1995. Jagged: amammalian ligand that activatesNotch1. Cell 80:909–917.
Lovschall H, Mitsiadis TA, Poulsen K,et al. 2007. Coexpression of Notch3and Rgs5 in the pericyte-vascularsmooth muscle cell axis in responseto pulp injury. Int J Dev Biol 51:715–721.
Lumsden AG. 1988. Spatial organiza-tion of the epithelium and the role ofneural crest cells in the initiation ofthe mammalian tooth germ. Devel-opment 103(Suppl):155–169.
Macias-Silva M, Hoodless PA, Tang SJ,et al. 1998. Specific activation ofSmad1 signaling pathways by theBMP7 type I receptor, ALK2. J BiolChem 273:25628–25636.
Maine EM, Lissemore JL, Starmer WT.1995. A phylogenetic analysis ofvertebrate and invertebrate Notch-related genes. Mol Phylogenet Evol4:139–149.
Matsuno K, Eastman D, Mitsiades T,et al. 1998. Human deltex is a con-served regulator of Notch signaling.Nat Genet 19:74–78.
Mikkola ML. 2008. TNF superfamily inskin appendage development. Cyto-kine Growth Factor Rev 19:219–230.
Mina M, Kollar EJ. 1987. The inductionof odontogenesis in non-dental mes-enchyme combined with early murinemandibular arch epithelium. ArchOral Biol 32:123–127.
Mitsiadis T. 2001. Bases moleculairesdu developpement dentaire. In:Piette E, Goldberg M, editors. La dentnormale et pathologique. De Boeck-Universite Press: Brussels. pp. 19–38.
Mitsiadis TA, Drouin J. 2008. Deletionof the Pitx1 genomic locus affectsmandibular tooth morphogenesis andexpression of the Barx1 and Tbx1genes. Dev Biol 313:887–896.
Mitsiadis TA, Rahiotis C. 2004. Parallelsbetween tooth development andrepair: conserved molecular mecha-nisms following carious and dentalinjury. J Dent Res 83:896–902.
CELL FATE DETERMINATION IN ODONTOGENESIS 209
Birth Defects Research (Part C) 87:199–211, (2009)

Mitsiadis TA, Smith MM. 2006. How dogenes make teeth to order throughdevelopment? J Exp Zoolog B MolDev Evol 306:177–182.
Mitsiadis TA, Lardelli M, Lendahl U,Thesleff I. 1995. Expression of Notch1, 2, and 3 is regulated by epithelial-mesenchymal interactions and reti-noic acid in the developing mousetooth and associated with determina-tion of ameloblast cell fate. J Cell Biol130:407–418.
Mitsiadis TA, Henrique D, Thesleff I,Lendahl U. 1997. Mouse Serrate-1(Jagged-1): expression in the devel-oping tooth is regulated by epithelial-mesenchymal interactions and fibro-blast growth factor-4. Development124:1473–1483.
Mitsiadis TA, Hirsinger E, Lendahl U,Goridis C. 1998a. Delta-notch signal-ing in odontogenesis: correlationwith cytodifferentiation and evidencefor feedback regulation. Dev Biol204:420–431.
Mitsiadis TA, Mucchielli ML, Raffo S,et al. 1998b. Expression of the tran-scription factors Otlx2, Barx1 andSox9 during mouse odontogenesis.Eur J Oral Sci 106 (Suppl 1):112–116.
Mitsiadis TA, Fried K, Goridis C. 1999.Reactivation of Delta-Notch signalingafter injury: complementary expres-sion patterns of ligand and receptorin dental pulp. Exp Cell Res 246:312–318.
Mitsiadis TA, Angeli I, James C, et al.2003a. Role of Islet1 in the pattern-ing of murine dentition. Development130:4451–4460.
Mitsiadis TA, Cheraud Y, Sharpe P,Fontaine-Perus J. 2003b. Develop-ment of teeth in chick embryos aftermouse neural crest transplantations.Proc Natl Acad Sci USA 100:6541–6545.
Mitsiadis TA, Regaudiat L, Gridley T.2005. Role of the Notch signalingpathway in tooth morphogenesis.Arch Oral Biol 50:137–140.
Mitsiadis TA, Caton J, Cobourne M.2006. Waking-up the sleepingbeauty: recovery of the ancestralbird odontogenic program. J ExpZoolog B Mol Dev Evol 306:227–233.
Mitsiadis TA, Barrandon O, Rochat A,et al. 2007. Stem cell niches in mam-mals. Exp Cell Res 313:3377–3385.
Mitsiadis TA, De Bari C, About I.2008a. Apoptosis in developmentaland repair-related human toothremodeling: a view from the inside.Exp Cell Res 314:869–877.
Mitsiadis TA, Tucker AS, De Bari C,et al. 2008b. A regulatory relation-ship between Tbx1 and FGF signalingduring tooth morphogenesis andameloblast lineage determination.Dev Biol 320:39–48.
Moore KA, Lemischka IR. 2006. Stemcells and their niches. Science 311:1880–1885.
Moss EG. 2007. Heterochronic genesand the nature of developmentaltime. Curr Biol 17:R425–R434.
Mucchielli ML, Mitsiadis TA. 2000. Cor-relation of asymmetric Notch2expression and mouse incisor rota-tion. Mech Dev 91:379–382.
Mucchielli ML, Mitsiadis TA, Raffo S,et al. 1997. Mouse Otlx2/RIEGexpression in the odontogenic epi-thelium precedes tooth initiation andrequires mesenchyme-derived sig-nals for its maintenance. Dev Biol189:275–284.
Murashima-Suginami A, Takahashi K,Kawabata T, et al. 2007. Rudimentincisors survive and erupt as super-numerary teeth as a result of USAG-1 abrogation. Biochem Biophys ResCommun 359:549–555.
Nadiri A, Kuchler-Bopp S, Haikel Y,Lesot H. 2004. Immunolocalization ofBMP-2/-4, FGF-4, and WNT10b inthe developing mouse first lowermolar. J Histochem Cytochem 52:103–112.
Naiche LA, Harrelson Z, Kelly RG,Papaioannou VE. 2005. T-box genesin vertebrate development. Annu RevGenet 39:219–239.
Neubuser A, Peters H, Balling R, MartinGR. 1997. Antagonistic interactionsbetween FGF and BMP signalingpathways: a mechanism for position-ing the sites of tooth formation. Cell90:247–255.
Nohe A, Hassel S, Ehrlich M, et al.2002. The mode of bone morphoge-netic protein (BMP) receptor oligo-merization determines different BMP-2 signaling pathways. J Biol Chem277:5330–5338.
Osumi-Yamashita N, Ninomiya Y, DoiH, Eto K. 1994. The contribution ofboth forebrain and midbrain crestcells to the mesenchyme in the fron-tonasal mass of mouse embryos. DevBiol 164:409–419.
Papaioannou VE, Silver LM. 1998. TheT-box gene family. Bioessays 20:9–19.
Plikus MV, Zeichner-David M, MayerJA, et al. 2005. Morphoregulation ofteeth: modulating the number, size,shape and differentiation by tuningBmp activity. Evol Dev 7:440–457.
Pouyet L, Mitsiadis TA. 2000. DynamicLunatic fringe expression is corre-lated with boundaries formation indeveloping mouse teeth. Mech Dev91:399–402.
Qiu M, Bulfone A, Ghattas I, et al.1997. Role of the Dlx homeoboxgenes in proximodistal patterning ofthe branchial arches: mutations ofDlx-1, Dlx-2, and Dlx-1 and -2 altermorphogenesis of proximal skeletaland soft tissue structures derivedfrom the first and second arches.Dev Biol 185:165–184.
Rosenzweig BL, Imamura T, OkadomeT, et al. 1995. Cloning and character-ization of a human type II receptorfor bone morphogenetic proteins.
Proc Natl Acad Sci USA 92:7632–7636.
Sire JY, Davit-Beal T, Delgado S, Gu X.2007. The origin and evolution ofenamel mineralization genes. CellsTissues Organs 186:25–48.
Smith AJ, Lesot H. 2001. Induction andregulation of crown dentinogenesis:embryonic events as a template fordental tissue repair? Crit Rev OralBiol Med 12:425–437.
St Amand TR, Zhang Y, Semina EV,et al. 2000. Antagonistic signalsbetween BMP4 and FGF8 define theexpression of Pitx1 and Pitx2 inmouse tooth-forming anlage. DevBiol 217:323–332.
ten Dijke P, Franzen P, Yamashita H,et al. 1994a. Serine/threonine kinasereceptors. Prog Growth Factor Res5:55–72.
ten Dijke P, Yamashita H, Ichijo H,et al. 1994b. Characterization of typeI receptors for transforming growthfactor-beta and activin. Science264:101–104.
ten Dijke P, Yamashita H, Sampath TK,et al. 1994c. Identification of type Ireceptors for osteogenic protein-1and bone morphogenetic protein-4.J Biol Chem 269:16985–16988.
Tissier-Seta JP, Mucchielli ML, Mark M,et al. 1995. Barx1, a new mousehomeodomain transcription factorexpressed in cranio-facial ectome-senchyme and the stomach. MechDev 51:3–15.
Trainor PA, Tam PP. 1995. Cranial par-axial mesoderm and neural crestcells of the mouse embryo: co-distri-bution in the craniofacial mesen-chyme but distinct segregation inbranchial arches. Development 121:2569–2582.
Trainor PA, Zhou SX, Parameswaran M,et al. 1999. Application of lacZ trans-genic mice to cell lineage studies.Methods Mol Biol 97:183–200.
Vainio S, Karavanova I, Jowett A,Thesleff I. 1993. Identification ofBMP-4 as a signal mediating second-ary induction between epithelialandmesenchymal tissues during earlytooth development. Cell 75:45–58.
Wang XP, Suomalainen M, FelszeghyS, et al. 2007. An integrated generegulatory network controls stem cellproliferation in teeth. PLoS Biol 5:e159.
Weissman IL, Anderson DJ, Gage F.2001. Stem and progenitor cells: ori-gins, phenotypes, lineage commit-ments, and transdifferentiations.Annu Rev Cell Dev Biol 17:387–403.
Yamamoto Y, Oelgeschlager M. 2004.Regulation of bone morphogeneticproteins in early embryonic develop-ment. Naturwissenschaften 91:519–534.
Yamashita H, ten Dijke P, HuylebroeckD, et al. 1995. Osteogenic protein-1binds to activin type II receptors andinduces certain activin-like effects.J Cell Biol 130:217–226.
210 MITSIADIS AND GRAF
Birth Defects Research (Part C) 87:199–211, (2009)

Yu PB, Beppu H, Kawai N, et al. 2005.Bone morphogenetic protein (BMP)type II receptor deletion revealsBMP ligand-specific gain of signalingin pulmonary artery smoothmuscle cells. J Biol Chem 280: 24443–24450.
Zhao GQ. 2003. Consequences ofknocking out BMP signaling in themouse. Genesis 35:43–56.
Zhao Z, Stock D, Buchanan A, Weiss K.2000. Expression of Dlx genes duringthe development of the murine denti-tion. Dev Genes Evol 210: 270–275.
Zouvelou V, Luder HU, Mitsiadis TA,Graf D. 2009. Deletion of BMP7affects the development of bones,teeth, and other ectodermal appen-dages of the orofacial complex.J Exp Zoolog B Mol Dev Evol 312:361–374.
CELL FATE DETERMINATION IN ODONTOGENESIS 211
Birth Defects Research (Part C) 87:199–211, (2009)