cells by mglur6, cb1, and gabac receptors an abstract …
TRANSCRIPT

shua Nathaniel hng
MODULATION OF OFF RESPONSE OUTPUT FROM MOUSE RETINAL GANGLION
CELLS BY MGLUR6, CB1, AND GABAc RECEPTORS
AN ABSTRACT
SUBMITTED ON THE SECOND DAY OF APRIL 2013
TO THE NEUROSCIENCE PROGRAM
IN PARTIAL FULFILLMENT OF THE REQUIREMENTS
OF THE SCHOOL OF SCIENCE AND ENGINEERING
OF TULANE UNIVERSITY
FOR THE DEGREE
OF
DOCTOR OF PHILOSOPHY
BY
APPROVED: 1,6
Guoyo .D., Ph.D Director
/7
Jeffrey G. Taster, Ph.D.
aura A. Schra , .D.
FionaM. Inglis, Ph.D.
Beamin J. Hall, Ph.D.

ABSTRACT
The retina is a sensory tissue that converts optical images into neural signals
known as light responses. Light responses are transmitted from photoreceptors to
bipolar cells to retinal ganglion cells (RGCs) in parallel pathways specific for either
light increments or light decrements. This improves vision by doubling the retina’s
dynamic range and increasing contrast sensitivity. Research has shown that Off
pathways, which are sensitive to light decrements, are likely modulated by the
activity of metabotropic glutamate receptor 6 (mGluR6) receptors, cannabinoid 1
receptors (CB1Rs), and -aminobutyric acid C (GABAC) receptors. In this
dissertation, I investigate how these neurotransmitter receptors modulate Off
responses in the retina by performing whole-cell recordings of mouse RGCs.
On bipolar cells express mGluR6 receptors, a type of glutamate receptor that
hyperpolarizes bipolar cells when bound to glutamate. Previous research has shown
that these receptors modulate Off responses under dark adaptation, but effects
under light adaptation were unclear. My research has shown that mGluR6 receptor
agonist DL-2-amino-4-phosphonobutyric acid (APB) decreases light-evoked Off
responses under light adaptation by disrupting dopaminergic transmission between
amacrine cells and Off bipolar cells.

CB1Rs are localized to many cell types including cone and bipolar cell axon
terminals, each of which release glutamate. Research primarily in brain has shown
that cannabinoid receptor activation prevents neurotransmitter release from the
presynapse. This has led to the hypothesis that CB1R activation would decrease
glutamate release in Off pathways and attenuate Off responses. My research shows
that CB1R agonists differentially modulate Off responses. Based on my results, I
suggest that CB1R agonists increase light-evoked Off responses in one population of
RGCs by reducing GABA transmission between GABAergic amacrine cells and Off
bipolar cells.
GABAergic amacrine cells feed back onto bipolar cell axon terminals that
express GABAC receptors. Previous research has shown that GABAC receptor
antagonist (1,2,5,6-tetrahydropyridin-4-yl)methylphosphinic acid (TPMPA) alters
On responses, but effects on Off responses are unclear. I show that TPMPA
modulates kinetics of both On and Off responses recorded from On-Off RGCs. All
together, the results in this dissertation indicate that mGluR6 receptors, CB1Rs, and
GABAC receptors modulate Off responses, and therefore vision.

oshua Nathaniel P ng
APPROVED: Guoy • ng W ng, M.D., Director
MODULATION OF OFF RESPONSE OUTPUT FROM MOUSE RETINAL GANGLION
CELLS BY MGLUR6, CB1, AND GABAc RECEPTORS
A DISSERTATION
SUBMITTED ON THE SECOND DAY OF APRIL 2013
TO THE NEUROSCIENCE PROGRAM
IN PARTIAL FULFILLMENT OF THE REQUIREMENTS
OF THE SCHOOL OF SCIENCE AND ENGINEERING
OF TULANE UNIVERSITY
FOR THE DEGREE
OF
DOCTOR OF PHILOSOPHY
BY
Jeffrey„p. Tasker, Ph.D.
Schrader, Ph.D.
61, ( S
Fion . Inglis, Phi.
Benj in J. Hall, Ph.D.
aura


ii
ACKNOWLEDGMENTS
Completing this dissertation would not have been possible without the
support of many people. First, I thank my parents for instilling a life-long interest in
science and for nurturing a curious mind. Especially, I want to thank my mom,
Colleen Marvin, who has made many personal sacrifices for my education and
happiness. I am exceedingly grateful for my fiancée, Amanda Hill, for her love and
support. She has often been a sounding board for the ideas and experiments in this
dissertation. I also want to express my deep appreciation for my advisor, Dr.
Guoyong Wang. Dr. Wang has been a true mentor for me the past five years.
Whenever I needed assistance with experiments, understanding scientific concepts,
or even life advice, Dr. Wang was available and eager to help. Under Dr. Wang’s
direction, I have grown both as a scientist and as a young man. In addition, I want to
thank my committee: Dr. Jeffrey Tasker, Dr. Laura Schrader, Dr. Fiona Inglis, and Dr.
Ben Hall. I am very grateful for their guidance and mentorship over the years.
I deeply appreciate all of my fellow Wang Lab members. Patch clamp
electrophysiology is an extremely challenging technique. I attribute much of my
success with the technique to Dr. Joey Nemargut who trained me on the patch clamp
rig. Also, I am very appreciative of Joey’s support and friendship outside of the lab.
In addition, I want to thank our post-doctoral fellow Dr. Jinnan Yang and our lab
technician Wei Huang for support in the lab. Finally, I would like to extend my

iii
thanks to our lab undergraduates Nathan Pham and Carter Kelly for all their
assistance inside and outside of lab.
Finally, I want to thank my many friends in the Neuroscience Program at
Tulane. I especially want to thank Sherrie Calogero for guiding me through the
program. To all those I have mentioned here and countless others I have shared
experiences with at Tulane, thank you for helping to make the last five years so
enjoyable.

iv
FOREWARD
Joshua Nathaniel Pahng has conducted many experiments to examine the
roles of mGluR6, CB1, and GABAC receptors in shaping light-evoked Off responses in
the mouse retina. His research is primarily focused on synaptic circuitry in the light-
adapted retina. Over the course of his graduate career, he has co-authored a peer-
reviewed journal article and published two abstracts at the Society for Neuroscience
conference. Much of the research presented in this dissertation is also presented in
the following:
Yang J, Pahng J, Wang GY (2012) Dopamine modulates the Off pathway in light-adapted mouse retina. J Neurosci Res. 91 (1), 138-150.
Pahng J, Yang J, Wang GY. Cannabinoid receptor 1 agonist 2-arachidonoylglycerol
differentially affected light-evoked responses in mouse retinal ganglion cells under light adaptation. Program No. 775.10. Neuroscience Meeting Planner. San Diego, CA: Society for Neuroscience, 2010. Online.
Pahng J, Yang J, Wang GY. Cannabinoid antagonist differentially modulated light-
evoked responses of mouse retinal ganglion cells under light adaptation. Program No. 557.3. Neuroscience Meeting Planner. Chicago, IL: Society for Neuroscience, 2009. Online.

v
TABLE OF CONTENTS
ACKNOWLEDGMENTS............................................................................................................................ii FOREWARD………………………………………………………………………………………………………..iv LIST OF FIGURES………………………………………………………………………………………………vii INTRODUCTION………………………………………………………………………………………………….1 Retina structure and organization……………………………………………………………...3 On and Off pathways………………………………………………………………………………….6 mGluR6 receptors in the retina………………………………………………………...………10 CB1 receptors in the retina………………………………………………………………………11 GABAC receptors in the retina…………………………………………………………………..14 Hypothesis………………………………………………………………………………………………15 Statement of specific aims………………………………………………………………………..16 CHAPTER 1: Dopamine modulates the Off pathway in light-adapted mouse
retina Introduction……...…………………………………………………………………………………….19 Materials and Methods…………………………………………………………………………….21 Results……………………………………………………………………………………………………27

vi
Discussion………………………………………………………………………………………………33 CHAPTER 2: CB1Rs modulate Off responses in light-adapted mouse retina Introduction……...…………………………………………………………………………………….54
Materials and Methods…………………………………………………………………………….56 Results……………………………………………………………………………………………………58 Discussion…………………………………………………………………………………….…………63 CHAPTER 3: GABAC receptor antagonist TPMPA modulates light-evoked Off
responses in light-adapted mouse retina Introduction……...…………………………………………………………………………………….82 Materials and Methods…………………………………………………………………………….85 Results……………………………………………………………………………………………………87 Discussion………………………………………………………………………………………………90 CONCLUSION…………………………………………………………………………………………………..102 LIST OF ABBREVIATIONS……………………………………………………………………………….107 LIST OF REFERENCES……………………………………………………………………………………..109

vii
LIST OF FIGURES 1. Diagram of rod and cone signaling pathways………………………………………………9 2. The effects of APB on RGC Off responses under different ambient light
conditions……………………………………………………………………………………………….39 3. The effects of L-AP4 and DCPG on RGC Off responses under different ambient
light conditions………………………………………………………………………………………..41 4. Antagonists of glycine, GABAA, and GABAC receptors did not eliminate the
APB-induced reductions of Off responses of RGCs in light-adapted mouse retinas…………………………………………………………………………………………………….43
5. Effects of dopamine receptor antagonists on the APB-induced reductions of
Off responses of RGCs in light-adapted mouse retinas……………………………….45 6. Effects of APB on light-evoked Off responses when D1 receptors and
inhibitory receptors are blocked………………………………………………………………47 7. HCN channel antagonist ZD 7288 prevented the APB-induced reduction in Off
responses under light adaptation……………………………………………………………..49 8. A depolarized holding potential of -40 mV, which inactivates HCN channels in
the recorded RGCs, did not prevent the APB-induced reduction of Off responses under light adaptation……………………………………………………………..51
9. A diagram showing a functional circuit from On pathways to Off pathways
under light adaptation……………………………………………………………………………..53 10. DSE of light-evoked Off EPSCs recorded from RGCs…………………………………..71 11. Effects of CB1R antagonist O-2050 on light-evoked Off responses of RGCs…73 12. Effects of CB1R agonist WIN on light-evoked Off responses of RGCs…………..75 13. Effects of CB1R agonist 2-AG when crossover inhibition is blocked with
mGluR6 agonist APB………………………………………………………………………………..77

viii
14. Increases in Off response amplitudes are prevented when GABAC receptors are blocked by TPMPA……………………………………………………………………………..79
15. Diagram showing a functional circuit for CB1R-mediated modulation of light-
evoked Off response amplitudes………………………………………………………………81 16. GABAC antagonist TPMPA increases the decay time of On and Off responses
in On-Off RGCs under low photopic conditions…………………………………………95 17. GABAC antagonist TPMPA increases the decay time of On and Off responses
in On-Off RGCs under high photopic conditions………………………………………..97 18. Blocking crossover inhibition with mGluR6 agonist APB does not prevent the
TPMPA-induced increase in Off response decay time under high photopic conditions………………………………………………………………………………………………99
19. Diagram showing a functional circuit for GABAC receptor-mediated
modulation of Off responses…………………………………………………………………..101

1
INTRODUCTION
The retina is a thin piece of neural tissue that senses photons and is crucial
for vision (1). It develops from the maturing diencephalon and, as such, is
considered part of the brain. Optical images are initially transformed into neural
signals by retinal photoreceptors that are sensitive to photons. The photoreceptors
can be subdivided into rods and cones that are active in dim and bright ambient
light intensities, respectively. The neural signal, also known as a light response, is
then transferred to interneurons known as bipolar cells, and ultimately to ganglion
cells whose axons terminate in the lateral geniculate nucleus of the thalamus. This
simple circuit uses glutamate as the primary neurotransmitter, and thus is known as
the glutamatergic through-pathway. Signal processing occurs within the retina prior
to the information reaching the lateral geniculate nucleus, both along the through-
pathways and by lateral transmission from amacrine and horizontal cells.
One way that the retina processes images is by dividing its neural signals
between parallel circuits (2). The circuit that is sensitive to light increments, that is
light with intensity above the mean ambient light intensity, is known as the On
pathway. On the other hand, the circuit that is sensitive to light decrements, or light
with intensity below the mean ambient light intensity, is known as the Off pathway.
This division into parallel circuits has important

2
consequences for vision. Separate channels are used to encode distinct parts of an
image. The specialization of one channel exclusively for light intensities above a
mean and another channel exclusively for light intensities below a mean results in
robust signal transfer for both light increments and light decrements; this is because
RGCs respond to both light increments and light decrements with high frequency
action potentials that are sent along the optic nerve to the lateral geniculate nucleus
(1). In addition, the division into parallel On and Off pathways is critical because it
results in the doubling of the retina’s dynamic range, or the range of light intensities
that can be effectively encoded into neural signals. The effects of halving the retinal
dynamic range are dramatic (3). In the rhesus monkey, the On channel was
reversibly blocked by intraocular injection of mGluR6 agonist APB. With only active
Off channels, the monkeys were unable to detect light increments. Moreover, the
monkeys experienced a severe loss in contrast sensitivity. Therefore, the parallel On
and Off pathways are critical for normal contrast sensitivity, which in turn is crucial
for resolving detail in everyday visual scenes. Since these parallel pathways are so
important for normal vision, it is vital that both the functional mechanisms of each
pathway and the differences between them are deeply understood. This thesis
focuses on the Off pathways and Off response modulation for two important
reasons. First, Off pathways form one half of the parallel circuits that are so
important for contrast sensitivity, and therefore should be studied in their own
right. Second, Off pathways have a relatively simple glutamatergic through-pathway
unlike On pathways that have an inverted synapse between the cones and bipolar

3
cells (4). Thus, for practical experimental reasons, we focus on modulation of Off
responses.
In this thesis, I examine the modulation of Off responses in the light-
adapted mouse retina. APB, a potent agonist of mGluR6 receptors has been shown to
increase Off response amplitudes under dark adaptation by disrupting crossover
inhibition from On pathways to Off pathways (5, 6, 7, 8). Therefore, I investigated
the effects of APB on Off response amplitudes under light adaptation and attempted
to determine the mechanism of APB’s effects, as detailed in Chapter 1. Investigators
have found that CB1Rs are localized to numerous sites in the mouse retina, but the
effects of this endocannabinoid (eCB) system on light responses are not understood
(9). I used CB1R agonists and antagonists to study the role the retinal eCB system
plays in modulating Off responses, as discussed in Chapter 2. Finally, previous
research has shown that GABAC receptors are localized to many bipolar cell axon
terminals, and these receptors modulate On response kinetics (10). I used GABAC
receptor-specific antagonist TPMPA to study the role of GABAC receptors in shaping
Off responses.
1. Retina structure and organization
The retina lines the posterior portion of the eye and is around 200 microns
thick in both humans and mice (11). Three main stages of processing occur within
the retina before the visual signals are sent to the lateral geniculate nucleus of the
thalamus (1). First, the photoreceptors transform the optical image made up of
photons into a neural signal in a process known as transduction. Second, the neural

4
signal is transmitted to interneurons including bipolar cells. Third, the neural signal
is transmitted to the RGCs that fire action potentials that travel to the thalamus.
Other retinal neurons include the horizontal cells with somas in the outer plexiform
layer (OPL) and amacrine cells with somas in the inner plexiform layer (IPL).
Photoreceptors are localized to the most posterior layer of the retina known
as the photoreceptor layer (12). In the absence of photons, photoreceptors are
depolarized due to a constant influx of sodium ions known as the dark current.
When photons are present, however, photoreceptors hyperpolarize. Rods are the
photoreceptor subtype that are active in dark adaptation and as such are involved in
night vision. They are extremely sensitive and can transduce single photons (13,
14). Cones, on the other hand, are active in light adaptation and are commonly
associated with visual acuity and color vision. They are not as sensitive as rods and
need the relatively abundant amount of photons present in daylight to function.
Both rods and cones release glutamate from their axon terminals.
Bipolar cells and horizontal cells receive direct glutamatergic input from the
photoreceptors (1). Their cell bodies are localized to the inner nuclear layer (INL)
and their dendrites stratify in the OPL. Rod bipolar cells receive inputs from rods,
cone bipolar cells receive inputs from cones, and one type of mouse bipolar cell
receives inputs from both rods and cones. The bipolar cell axon terminals stratify in
the IPL and release glutamate. Horizontal cells mediate lateral transmission in the
retina (15). They release GABA from their axon terminals that are localized to the
OPL. Depending on subtype, horizontal cells may feed back onto photoreceptors

5
and/or feed forward onto bipolar cells. Functionally, the horizontal cells average
photoreceptor signals (1).
The amacrine cells receive direct glutamatergic inputs from bipolar cells (16,
17). Most amacrine cell bodies are localized to the INL, but so-called displaced
amacrine cell bodies can be found in the ganglion cell layer (GCL). There are
approximately 40 subtypes of amacrine cell. The AII amacrine cell mediates forward
transmission and is crucial under dark adaptation. It receives inputs exclusively
from rod bipolar cells and connects to either On-cone bipolar cells via gap junctions
or Off-cone bipolar cells via glycinergic synapses. Other amacrine cell subtypes
mediate lateral transmission. Depending on subtype, amacrine cells can release
neurotransmitters including but not limited to glycine, GABA, dopamine, or
acetylcholine (18).
RGCs are the final output neurons of the retina. Their cell bodies are localized
to the GCL, the innermost layer of the retina, and they receive direct inputs from
bipolar cells and amacrine cells (1). Ganglion cells have diverse morphology and
physiology and can be subdivided into at least 20 subtypes. The On subtype has
dendrites that stratify in sublamina b, the innermost part of the IPL whereas the Off
subtype has dendrites that stratify in sublamina a, the outermost part of the IPL (19,
20). On RGCs respond with action potentials when there are light increments in the
center of their receptive field and Off RGCs respond with action potentials when
there are light decrements in the center of their receptive field.

6
2. On and Off pathways
The retina divides its neural signals into parallel circuits for light increments
and light decrements (1). In the light-adapted retina, also known as photopic
conditions, there is one circuit for light increments known as the On-cone signaling
pathway and one circuit for light decrements known as the Off-cone signaling
pathway. The On-cone signaling pathway proceeds as follows: 1) cones
hyperpolarize in response to photons, 2) glutamate release from cones onto On-
cone bipolar cells ceases, 3) mGluR6 receptors localized to On-cone bipolar cells are
inactive resulting in depolarization of the On-cone bipolar cells, 4) glutamate
released from On-cone bipolar cells depolarizes On ganglion cells. For light
decrements, the Off-cone signaling pathway proceeds as follows: 1) cones
depolarize in response to a decrease of photons, 2) cones release glutamate onto
Off-cone bipolar cells, 3) ionotropic glutamate receptors localized to Off-cone
bipolar cells are activated resulting in depolarization of the Off-cone bipolar cells, 4)
glutamate released from Off-cone bipolar cells depolarizes Off ganglion cells.
Under scotopic conditions of the dark-adapted retina, there are two circuits
for light increments (21). The main pathway for light increments in scotopic
conditions is known as the primary rod On pathway, and it proceeds as follows: 1)
rods hyperpolarize in response to photons, 2) glutamate release from rods onto rod
bipolar cells ceases, 3) mGluR6 receptors localized to rod bipolar cells are inactive
resulting in depolarization of the rod bipolar cells, 4) glutamate released from rod
bipolar cells depolarizes AII amacrine cell, 5) On-cone bipolar cells also depolarize
because of their gap junctions with AII amacrine cells, 6) glutamate released from

7
On-cone bipolar cells depolarizes On ganglion cells. The other pathway for light
increments in scotopic conditions is known as the secondary rod On pathway. In
this pathway, the rods form gap junctions with cones so when the rods
hyperpolarize the cones do as well. The neural signal then travels down the On-cone
signaling pathway just as it would under light adaptation.
There are three circuits for light decrements under scotopic conditions (22).
The primary rod Off pathway is as follows: 1) rods depolarize in response to
absence of photons, 2) rods release glutamate onto rod bipolar cells, 3) rod bipolar
cells expressing mGluR6 receptors hyperpolarize in response to glutamate, 4)
glutamate release from rod bipolar cells to AII amacrine cells ceases, 5) AII amacrine
cells hyperpolarize and cease release of glycine onto Off-cone bipolar cells, 6) Off-
cone bipolar cells depolarize and release glutamate onto Off ganglion cells. In the
secondary rod Off pathway, rods make glutamatergic synapses directly onto Off-
cone bipolar cells. Therefore, when the rods depolarize, they release glutamate onto
Off-cone bipolar cells that release glutamate onto Off RGCs. Finally, in the tertiary
rod Off pathway, rods form gap junctions with cones (23). Thus, when the rods
depolarize, the cones do as well resulting in a release of glutamate from cones that
depolarizes Off-cone bipolar cells, and ultimately depolarizes Off ganglion cells. For
both light increments and light decrements, the pathways are ranked (i.e. primary,
secondary, etc.) in order from most to least sensitive. The On and Off signaling
pathways are shown in Figure 1.

8
Figure 1. Diagram of rod and cone signaling pathways. A: One cone pathway and
two rod pathways for transmitting On responses. On cone signaling pathway: cones
> On-cone bipolar cells > On RGCs. Primary rod On pathway: rods > rod bipolar cells
> AII amacrine cells > gap junction > On-cone bipolar cells > On RGCs. Secondary rod
On pathway: rods > gap junction > cones > On-cone bipolar cells > On RGCs. B: One
cone pathway and three rod pathways for transmitting Off responses. Off cone
signaling pathway: cones > Off-cone bipolar cells > Off RGCs. Primary rod Off
pathway: rods > rod bipolar cells > AII amacrine cells > Off-cone bipolar cells > Off
RGCs. Secondary rod Off pathway: rods > Off-cone bipolar cells > Off RGCs. Tertiary
rod Off pathway: rods > gap junction > cones > Off-cone bipolar cells > Off RGCs. R=
rod, C= cone, RB= rod bipolar cell, On CB= On-cone bipolar cell, Off CB= Off-cone
bipolar cell, AII= AII amacrine cell, On RGC= On retinal ganglion cell, Off RGC= Off
retinal ganglion cell. Dotted lines represent gap junctions. Bolded cell types express
mGluR6 receptors at the dendrites.
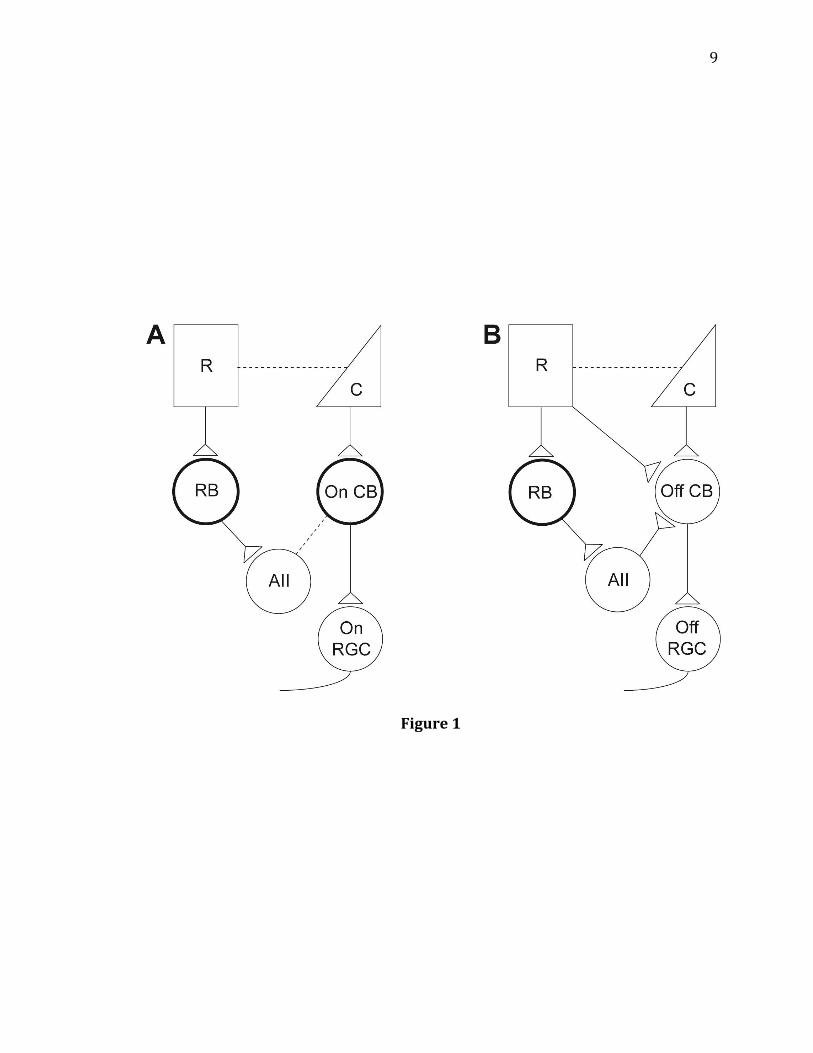
9
Figure 1

10
In the lab, we are capable of experimentally determining whether a ganglion
cell receives inputs from On pathways, Off pathways, or both On and Off pathways.
RGCs are voltage-clamped in the whole-cell mode and a spot of light, centered on the
ganglion cell, is shined onto the retina. On ganglion cells respond with an inward
current at light onset whereas Off ganglion cells respond with an inward current at
light offset. On-Off ganglion cells, with bistratified dendrites that collect inputs from
both On and Off pathways, respond with an inward current both at light onset and at
light offset. These inward currents, provided they are large enough to reach
threshold, would trigger action potentials in vivo that would be transmitted via the
optic nerve (24).
3. mGluR6 receptors in the retina
In the retina, mGluR6 receptors have been localized to rod bipolar cell and
On-cone bipolar cell dendrites (25, 26). These receptors are G-protein coupled and
close a cation channel when bound to glutamate. Research has shown that this
cation channel may be TRPM1 (27). As a result of the cation channel closure in the
rod bipolar cell or On-cone bipolar cell dendrites, the bipolar cell hyperpolarizes.
However, Off-cone bipolar cells do not express mGluR6 receptors (28, 29, 30).
Instead, they express a combination of AMPA and Kainate receptors that depolarize
the Off-cone bipolar cell when bound to glutamate. It is this difference in glutamate
receptor expression at the bipolar cell that creates the distinct On and Off pathways.
While the On and Off pathways carry distinct neural signals representing
different parts of an optical scene, these pathways also crosstalk. One way that On

11
and Off pathways crosstalk is through crossover inhibition (31, 32, 33, 34).
Activation of On pathways inhibits Off pathways and vice versa via glycinergic
amacrine cells that are involved in lateral transmission (35, 36, 37). DL-2-Amino-4-
phosphonobutyric acid (APB) is a potent mGluR6 agonist (38). When it binds to
mGluR6 receptors on On-cone bipolar cell dendrites, the cell hyperpolarizes and
glutamate release from the On-cone bipolar cell axon terminal ceases. Since Off-cone
bipolar cells do not express mGluR6 receptors, APB does not directly affect them.
Thus, APB has been used experimentally to specifically block On pathways.
Previous studies have shown that in the dark-adapted retina, APB blocks On
responses but increases the amplitude of Off responses (5, 6, 7, 8). In this scenario,
APB inactivates the On pathway preventing crossover inhibition of Off pathways
and resulting in potentiation of Off responses. Since these previous studies have
been performed under dark adaptation, the rod pathways were active. During
application of APB, rod bipolar cells and On-cone bipolar cells are hyperpolarized
resulting in the total block of On pathways, and partial block of Off pathways. As the
primary rod Off pathway is blocked by APB, Off responses are carried by the
secondary and possibly by the rare mouse tertiary rod Off pathways. The effects of
APB on Off responses in the light-adapted retina are unknown, however.
4. CB1 receptors in the retina
Previous investigations of the effects of Cannabis use on vision provided the
first evidence of a retinal eCB system (9). Anecdotal evidence was provided by
Jamaican fishermen who claimed that smoking marijuana improved their night

12
vision and helped them catch fish at night (39). Cannabis sativa or
tetrahydrocannabinol administration was found to improve night vision in a field
study in Morocco (40). The field researcher hypothesized that these effects were
mediated at the retinal level. Marijuana smoking has also been shown to have other
visual effects such as a decrease in visual acuity during testing with a Snellen chart
and an increase in photosensitivity (41). Only after the eCB 2-AG was isolated in the
retina was the existence of a retinal eCB system confirmed (42).
The expression of CB1Rs in the retina has been investigated in human,
mouse, rat, chick, tiger salamander, goldfish, and rhesus monkey (9, 42, 43, 44). In
one study, CB1Rs were found to be expressed in rod synaptic terminals, cone
synaptic terminals, amacrine cells, RGCs, and sporadically in the IPL (42). Distinct
bands of expression were observed in the IPL near the INL and GCL. In another
study and using a different antibody, CB1R expression was detected in rat rod
bipolar cells, PKC-immunoreactive GABAergic amacrine cells, the outer plexiform
layer, and the IPL but not the GCL (43). In goldfish, CB1R immunoreactivity was
detected in cone pedicles, Müller’s cells, bipolar cell bodies, bipolar cell axon
terminals, and in the IPL, but CB1Rs were not detected in rod spherules or in
horizontal cells (44). Within the IPL, CB1R expression was strongest in the On
sublamina.
Various experiments have been performed to determine the functions of the
retinal eCB system. The majority of these studies have focused on the role this
system plays in modulating ion channel currents. Brain researchers have shown that
CB1Rs are activated by endogenous retrograde neurotransmitters (45, 46). In the

13
retina, KCl or mGluR1 agonist DHPG puff onto bipolar cell bodies elicited release of a
retrograde neurotransmitter that reversibly inhibited cone IK(V) currents by
approximately 25% (47). This effect was blocked by CB1R antagonist SR141716A
indicating that the retrograde transmitter was an eCB acting at CB1 receptors. Bath
application of CB1R agonist modulates several voltage-dependent ion channel
currents including HVA Ca2+ currents, ICa currents, and IK(V) currents in
photoreceptors, bipolar cells, and cultured RGCs (42, 44, 47, 48, 49, 50, 51, 52).
CB1R agonist concentrations as high as 10M were used in several studies because
biphasic effects emerge at lower concentrations like 1M, and because researchers
wanted to ensure that the lipophilic drug would penetrate all retinal synaptic layers
(48, 53, 54, 55, 56).
Several studies have investigated the role of the retinal eCB system in
regulating neurotransmitter release. A variety of CB1R agonists have been shown to
decrease generalized norepinephrine, dopamine, and glutamate release in the retina
(57, 58, 59). This is consistent with findings in other studies demonstrating that
CB1Rs activate Gi/o to reduce neurotransmitter release (60). CB1R agonists also
affect miniature post-synaptic current (mini) frequency. In cultured chick amacrine
cells with a low initial mini frequency, CB1R agonists increased the mini frequency
via a CB1R-mediated mechanism (61). CB1R agonists also decreased spontaneous
excitatory post-synaptic current (EPSC) and inhibitory post-synaptic current (IPSC)
frequencies recorded from mouse RGCs (62). Only one study has investigated CB1R
effects on light-evoked responses (63). In that study, researchers found that cone
responses to light offset returned to baseline faster following CB1R agonist

14
application. CB1R antagonist SR141716A did not block this phenomenon, however,
possibly indicating a non-CB1R effect.
5. GABAC receptors in the retina
GABAC receptors, like GABAA receptors, are ionotropic GABA receptors that
flux chloride (64). TPMPA is an antagonist that specifically blocks GABAC receptors,
but GABAA receptors can be blocked specifically by bicuculline and picrotoxin (PTX)
blocks both GABAA and GABAC receptors (65, 66). While GABAA receptor expression
is predominant in other parts of the central nervous system, GABAC receptors are
highly expressed in the retina. In mammals, GABAC receptors are localized to rod
and cone bipolar cell axon terminals (67). Bipolar cell axon terminals receive GABA
inputs from GABAergic amacrine cells (1). Light GABAA expression has also been
observed in bipolar cell axon terminals, but the majority of GABAA receptors are
expressed in cones, bipolar cell dendrites, and ganglion cells (68). In ferret, the ratio
of GABAC to GABAA (GABAC: GABAA) expression is higher in On bipolar cells than Off
bipolar cells indicating differences in GABA receptor expression between On and Off
pathways (69). Further, GABAC: GABAA expression is higher in rod bipolar cells than
cone bipolar cells. Following with the morphological data, GABAC receptors mediate
the majority of chloride current induced by GABA puff onto bipolar cell axon
terminals (70). In the study, the ratio of GABAC to GABAA-mediated current differed
between bipolar cells in tiger salamander retina, but overall, 80% of the current
induced by GABA puff was mediated by GABAC receptors.

15
GABAC receptors modulate bipolar cell excitability and help shape light-
evoked responses. It has been shown that GABAC-mediated currents in bipolar cells
exhibit a longer latency and time course than GABAA-mediated currents (71, 72, 73).
Light-evoked inhibitory currents recorded from bipolar cells were blocked by PTX,
the GABAA and GABAC antagonist, but not by the GABAA-specific antagonist
bicuculline (74). This research indicates that GABAC receptors mediate inhibition
onto bipolar cells, modulate excitability of bipolar cells, and ultimately modulate
neurotransmission from bipolar cells to RGCs. Indeed, under one level of light
adaptation, block of GABAC receptors with 50M TPMPA increased total charge
transfer and decay time of On EPSCs recorded from mouse RGCs (10). In contrast,
TPMPA did not affect Off responses recorded from Off RGCs. GABAC receptor
knockout decreased On RGC but not Off RGC dynamic range, or the range of light
intensities that elicit between 5% and 95% of the maximum response, compared to
wild type.
6. Hypothesis
The retina uses parallel circuits to effectively transmit neural signals for both
light increments and light decrements (1). Off pathways form one half of these
parallel circuits and are crucial for proper contrast sensitivity and therefore, vision.
Previous investigations, including receptor localization studies, indicate that
mGluR6 receptors, CB1 receptors, and GABAC receptors may modulate Off
responses. I performed whole-cell voltage clamping of mouse RGCs and record light-
evoked Off responses primarily under light adaptation. To determine the role of

16
mGluR6 receptors in shaping Off responses under light adaptation, I applied
mGluR6-specific agonist APB. To examine the role of CB1 receptors in shaping Off
responses, I used CB1R agonists 2-arachidonoyl glycerol (2-AG) and WIN 55,212-2
(WIN) and various CB1R antagonists. To study how GABAC receptors modulate Off
responses under different levels of light adaptation, I applied the GABAC- specific
antagonist TPMPA. In these studies, I tested the hypotheses that: APB increases Off
response amplitudes under light adaptation as it does under dark adaptation by
reducing crossover inhibition from On pathways to Off pathways; CB1R agonists
reduce glutamate release along the Off pathway resulting in a decrease in Off response
amplitudes; and TPMPA increases the decay time of Off responses as has been
previously shown in On responses.
7. Statement of specific aims
Specific Aim 1: investigated whether mGluR6 agonist APB increases RGC
light-evoked Off amplitudes by reducing crossover inhibition of Off pathways under
light adaptation. Retinal On pathways exhibit crossover inhibition of Off pathways
and vice versa via glycinergic amacrine cells (31, 32, 33, 34). The mGluR6 agonist
APB is commonly used to block On pathways and has been shown to prevent
crossover inhibition of Off pathways under dark adaptation resulting in an increase
in Off response amplitudes (5, 6, 7, 8). We hypothesized that APB would also
potentiate Off responses under light adaptation by reducing crossover inhibition of
Off pathways. For experiments in this specific aim, I performed whole-cell voltage
clamp on mouse RGCs under different adaptation levels and used APB to determine

17
if Off responses were uniformly potentiated. In addition, I applied different mGluR6
agonists to ensure that APB effects were mediated by mGluR6 receptors. Further, I
applied dopamine receptor antagonists to determine if the APB effects were
dopamine transmission-dependent. The experiments for this specific aim are
explained in detail in Chapter 1.
Specific Aim 2: investigated whether cannabinoids acting at CB1Rs
attenuate RGC light-evoked Off responses under light adaptation. Classically,
presynaptic CB1Rs are activated by eCBs released by postsynaptic cells to decrease
neurotransmitter release from the presynapse (75, 76, 77, 78, 79). In the retina,
CB1Rs are localized at presynaptic sites at glutamatergic synapses along the Off
pathway (9, 42, 43, 44). Therefore, we hypothesized that bath application of CB1R
agonists would reduce glutamatergic transmission and decrease Off response
amplitudes. For experiments in this specific aim, I performed whole-cell patch
clamping of mouse RGCs under light adaptation. I applied either CB1R antagonists,
such as O-2050 or AM251, or CB1R agonists, such as 2-AG and WIN, and recorded
light-evoked responses to determine the role of CB1 receptors in modulating Off
responses. Also, I co-perfused APB during CB1R agonist treatment to eliminate
crossover inhibition. Further, I applied TPMPA both prior to and during CB1R
agonist perfusion to determine if CB1R agonists increased Off amplitudes in some
RGCs via disinhibition. The experiments for this specific aim are explained in detail
in Chapter 2.
Specific Aim 3: investigated whether GABAC receptor antagonist TPMPA
modulates Off responses. GABAC receptors have been localized to the axon terminals

18
of bipolar cells (67). Under one level of light adaptation, block of GABAC receptors
with TPMPA has been shown to selectively potentiate On EPSCs recorded from On
RGCs (10). TPMPA releases bipolar cells from inhibition leading to greater release of
glutamate from bipolar cells onto RGC dendrites. For experiments in this specific
aim, I performed whole-cell patch clamping of mouse On-Off RGCs under different
levels of light adaptation. I applied TPMPA to determine if both On and Off response
decay times increase with GABAC receptors blocked. Additionally, I applied APB
both prior to and during application of TPMPA to investigate whether TPMPA
increases Off responses via a decrease in crossover inhibition. Further, experiments
were performed at different ambient light intensities to determine if GABAC-
mediated modulation of Off responses is consistent or dynamic. The experiments for
this specific aim are explained in detail in Chapter 3.

19
CHAPTER 1
Dopamine modulates the Off pathway in light-adapted mouse retina
Introduction
In the retina, light increment information is transmitted by On pathways and
light decrement information is transmitted by Off pathways. Regardless of
adaptation level, On responses require transmission by On-cone bipolar cells (1).
The On-cone bipolar cells express mGluR6 receptors at their dendrites. These
mGluR6 receptors differ from the ionotropic glutamate receptors expressed by Off-
cone bipolar cells in two crucial ways. First, mGluR6 receptors are metabotropic G-
protein coupled receptors. Second, mGluR6 receptor activation closes a cation
channel resulting in the hyperpolarization of the On-cone bipolar cell (27, 80). Thus,
mGluR6 agonist APB can be applied to hyperpolarize On-cone bipolar cells and
block On pathways.
Off-bipolar cells do not express mGluR6 receptors and therefore cannot be
directly affected by APB. Nevertheless, under dark adaptation, Off response
amplitudes have been shown to increase following application of APB (5, 6, 7, 8). It
has been shown that APB hyperpolarizes On-cone bipolar cells resulting in a
disruption of glutamate release onto glycinergic amacrine cells (34, 35, 36, 37). The
amacrine cells cease

20
releasing glycine onto Off-cone bipolar cells resulting in disinhibition and larger Off
responses. These studies have revealed that On and Off pathways are not completely
discrete. Instead, the pathways communicate through crossover inhibition.
However, the studies have not demonstrated that the effects of APB are consistent
across adaptation levels.
In the current study, patch clamp recordings were made from RGCs in mouse
retinas under different adaptation levels. We found that APB decreases Off response
amplitudes under light adaptation by inhibiting dopaminergic amacrine cells that
synapse onto Off-cone bipolar cells resulting, ultimately, in inhibition of Off-cone
bipolar cell hyperpolarization-activated cyclic nucleotide-gated (HCN) channels. The
results presented in this study suggest that crosstalk between On and Off pathways
occurs under light adaptation via dopaminergic amacrine cells.

21
Materials and Methods
The basic methods used in this study were similar to those used previously
(81, 82, 83, 84). All procedures were in compliance with National Institutes of
Health guidelines and were approved by the campus animal use committees of
Tulane University. Animals were dark-adapted overnight prior to the experiments.
All procedures, including animal surgery, dissection of retinas, and recordings from
cells, were performed in complete darkness. Infrared goggles were used to visualize
the tissue on the dissecting and recording microscopes and to maneuver in the
recording room. LEDs (850nm) were used to provide light to the dissecting
microscope while the illumination from the recording microscope was passed
through a 850-nm cutoff filter.
Retinal preparation. Retinas were obtained from 3-4-month-old mice (C57BL/6
from Charles River). After a lethal dose of barbiturate (Beuthanasia-D; 360 mg/kg
i.p.), the eyes were removed and placed in oxygenated L15 (Sigma, St. Louis, MO;
L1518) at 37C for 12 min. The retinas were then carefully peeled from the eyecup
and stored at room temperature in minimum essential medium Eagle (MEME;
Mediatech, Herndon, VA; catalog No. 51-010-PC), and continuously bubbled with
95% O2+ and 5% CO2. There was no cesium in the bath solution. A small piece of
retina was placed GCL upward in the recording chamber and stabilized with an
overlying piece of filter paper. A 2-mm hole in the filter paper provided access for
the recording electrode. Cells were visualized through a 40X objective mounted on
an upright epifluorescence microscope (Nikon).

22
During recordings, the retina was perfused continuously with MEME (1.5
ml/min) through a gravity-fed line, heated with a dual channel temperature
controller (Warner Instruments, Hamden, CT), and continuously bubbled with 95%
O2 and 5% CO2. A calibrated thermocouple monitored the temperature in the
recording chamber, which was maintained at 35C. Recordings from each individual
cell usually lasted 30-120 min., and retinal segments from which recordings were
made typically remained viable for 8-12 hr. Patch electrodes were filled with a
solution containing (in mM) cesium methanesulfonate 118, CsCl 12, CaCl2 0.5, MgCl2
0.5, HEPES 10, EGTA 5, 0.5% Lucifer yellow, QX-314 3, pH 7.4, osmolarity 290
mOsm. The chloride equilibrium potential (ECl) with this internal solution was
approximately -58.0 mV. QX-314 (3 mM) was included in the electrode solution to
eliminate sodium currents in the recorded cell. All recordings were made with the
whole-cell configuration. By the end of the experiment, the soma and the dendritic
arborizations of the recorded cell were usually completely filled with Lucifer yellow.
Once complete filling had been achieved, the retina was removed and fixed in 4%
paraformaldehyde for 6-8 hr at 4C.
Subsequently, by using a Leica TCS SP2 confocal microscope (Leica
Microsystems, Heidelberg, Germany), high-resolution and three-dimensional images
were made of each cell. Scans were taken at 0.25-0.7-m intervals along the z-axis
depending on the objective used. The dendritic stratifications of RGCs in the IPL
were determined by rotating the confocal stack image 90. DAPI was used to label
the nuclei of the GCL and the INL. The depth of the IPL was defined as the area
between the GCL and the inner border of the INL. The outer two-fifths of the IPL is

23
sublamina a, the Off sublamina; the inner three-fifths of the IPL is sublamina b, the
On sublamina. Ganglion cells with their dendrites ramify in sublamina a, sublamina
b, or both are Off, On, or On-Off RGCs, respectively. Only morphologically identified
Off and On-Off RGCs were included in this study. Because we focused mainly on the
effects of APB on the Off responses under dark and light adaptation, and for
statistical analyses, we grouped Off and On-Off cells together. Images of
morphological Off RGCs and On-Off RGCs are similar to those from our previous
studies (83, 84, 85).
Electrophysiology. Whole-cell patch clamp recordings were made from RGCs in dark-
and light-adapted retinas. Patch pipettes with a tip resistance between 3 and 7 M
were pulled from thick-walled 1.5-mm-ODS borosilicate glass on a Sutter
Instruments (Novato, CA) puller (P-97). Whole-cell patch clamp recordings were
made with a Multiclamp 700B patch clamp amplifier (Axon instruments,
Burlingame, CA). The data were low-pass filtered at rates between 1 and 2 kHz and
digitized at rates of 5 kHz before storage on a computer for subsequent offline
analysis. To attain whole-cell access, the vitreous and the limiting membrane
overlying the recording area were removed by gently brushing the retinal surface
with the tip of a glass pipette. Recordings were made from ganglion cells located
around a region about midway between the geometric center and the peripheral rim
of the retina and obtained by patching onto cells with clear, nongranular cytoplasm.
High-resistance seals were obtained by moving the patch electrode onto the cell
membrane and applying gentle suction. After formation of a high-resistance seal

24
between the electrode and the cell membrane, transient currents caused by pipette
capacitance were electronically compensated by the circuit of the MultiClamp 700B
patch clamp amplifier. Recordings from cells with a seal resistance 1 G were
discarded. After forming the whole-cell configuration by gentle suction, the series
resistance was measured, and it was 7-16 M. Recordings were terminated
whenever significant increases (20%) in series resistance occurred. Light-evoked
EPSCs were recorded at the membrane potential of -58 mV, the chloride reversal
potential of the RGCs. For light-evoked EPSCs, the averaged amplitudes of each cell
before and after drug application were obtained from five trials. The results were
expressed as mean SE.
Light stimulus. Light-evoked responses were obtained by delivering square-wave
spots of light to the retina from a 1-in.-diameter computer monitor, with a green
(P43, 545 nm light) phosphor (Lucivid MR1-103 MicroBrightField, Colchester, VT),
through the camera port of the microscope (86). Light responses to a 600 m spot of
light were recorded for each cell. Then, the size of the spot of light was adjusted to
evoke the maximal current recorded under voltage clamp before the effects of APB
were tested. For each cell, different-sized spots, from 200 to 600 m were used. The
spots of light were always centered on the soma. The stimuli were programmed in
Matlab (Mathworks, Natick, MA), using the Psychophysics Toolbox extensions (87,
88). The intensity of the spot of light was calibrated with a spectroradiometer/
photometer (UDT instruments, S350/268R) and expressed in terms of the time-
averaged rate of photoisomerizations per rod per second (Rh/rod/sec), the only

25
unit that the retina understands (89). The instrument was calibrated relative to
standards of the National Institute of Standards and Technology.
The method of recording from light-adapted retinas is essentially the same as
in our previous studies (83, 84). Initially, the whole-cell patch clamp recordings
were made from a ganglion cell under the dark-adapted condition; then, a
background light was delivered to the retina to induce light adaptation, and the
recording was continued from the same cell. For these studies, a background light of
constant brightness at 150, 1500, or 15000 Rh/rod/sec was provided full-field by
the computer-controlled 1-in.-diameter monitor (Lucivid) for 10 min to allow the
transition from dark adaptation to adaptation under the background light. The
background light with intensity at 150 Rh/rod/sec is in the mesopic range (21, 90).
With this background light, both rods and cones can be activated. In contrast, the
background light with intensity at 1500 or 15000 Rh/rod/sec is in the photopic
range. Rods are completely inactivated by 1500 or 15000 Rh/rod/sec background
light (83, 84).
Light stimuli with intensity of 350 Rh/rod/sec and light stimuli with
intensities greater than that of the background light were used in the dark- and
light-adapted retina, respectively to evoke light responses from ganglion cells. For
each cell, different intensities were used to evoke light responses under light
adaptation. Then, the lowest intensity required to evoke an optimal response was
determined and used in the experiments to test the effects of APB on the ganglion
cell. An optimal response was characterized as the largest synaptic current
amplitude (in voltage clamp mode) of the light-evoked responses of each cell. Under

26
dark or different background light conditions, light stimuli were delivered once
every 20 sec to limit alteration of the adaptation level.
Drug application. APB (20 M; Sigma), L-2-amino-4-phosphonobutyric acid (L-AP4;
4 M; Sigma), (S)-3,4-dicarboxyphenylglycine (DCPG; 3 M; Tocris, Ellisville, MO),
strychnine (STR; 2 M; Sigma), picrotoxin (PTX; 100 M; Sigma), (1,2,5,6-
tetrahydropyridin-4-yl)methylphosphinic acid (TPMPA; 50 M; Sigma), SCH 23390
hydrochloride (20 or 10 M; Tocris), spiperone hydrochloride (20 or 10 M;
Tocris), and ZD 7288 (100 or 30 M; Tocris) were freshly dissolved in MEME on the
day of the experiment and administered through a gravity-fed line. The pH was
adjusted to 7.4. All drugs were prepared and stored in accordance with the
manufacturer’s recommendations. The solutions were heated with a dual-channel
temperature controller (Warner Instruments) and continuously bubbled with 95%
O2 and 5% CO2. A six-position rotary valve (Western Analytical Products, Wildomar,
CA) was used to switch between bath and drug solutions.

27
Results
Effects of APB and L-AP4 on the light-evoked Off responses in Off and On-Off ganglion
cells under dark adaptation and different ambient light conditions
To determine the effects of APB on Off responses, morphologically identified
Off and On-Off mouse RGCs were voltage-clamped and light-evoked Off responses
were recorded before and after bath application of APB. In addition, these cells were
either dark-adapted or adapted to another ambient light intensity. Researchers have
identified three Off pathways that function under dark adaptation: 1) the primary
pathway: rod > rod bipolar cell > AII amacrine cell > glycinergic synapse > Off-cone
bipolar cell > Off ganglion cell; 2) the secondary pathway: rod > gap junction > cone
> Off-cone bipolar cell > Off ganglion cell; and 3) the tertiary pathway: rod > Off-
cone bipolar cell > Off ganglion cell (2, 4, 21, 91). APB blocks the primary pathway
by hyperpolarizing rod bipolar cells and the tertiary pathway is rarely seen in the
mouse retina (23). Thus, following APB application in the dark-adapted retina, Off
responses are mainly mediated by the secondary Off pathway. However, the light
stimulus intensity of 350 Rh/rod/sec that was used to elicit responses in dark-
adapted cells can also activate M cones (92). Therefore, some of the Off responses
under dark adaptation were likely mediated by the Off cone pathway.
Bath applied APB (20 M) completely blocked On responses in On-Off RGCs
and under all tested ambient light intensities (Figure 2). The effects of APB on Off
responses varied depending on adaptation, however. Under dark adaption, APB
significantly increased the amplitude of Off responses recorded from Off and On-Off
RGCs (Figure 2A,B, n= 15). After washout, the potentiated Off amplitudes returned

28
to baseline. At a background light intensity of 150 Rh/rod/sec, however, APB
effects were inconsistent and no significant increase or decrease in Off amplitude
was observed (Figure 2C,D, n= 7). The effects of APB were also tested under a
background light intensity of 1500 Rh/rod/sec (Figure 2E,F, n=20) or 15000
Rh/rod/sec (Figure 2G,H, n= 8). In both conditions, APB significantly reduced the
amplitude of Off responses. These attenuated Off responses recovered following a
washout.
In order to confirm that the observed APB effects are mediated by mGluR6
receptors, we repeated the above experiment with the more specific mGluR6
agonist, L-AP4 (Figure 3A-D, n= 8). Effects of L-AP4 were similar to those of APB.
Following bath application of L-AP4, On responses were blocked in all recorded On-
Off RGCs. In addition, L-AP4 increased the amplitude of Off responses under dark
adaptation (Figure 3A,B, n= 8). Further, L-AP4 decreased the amplitude of Off
responses under a background of 1500 Rh/rod/sec (Figure 3C,D, n=8). These
results confirm that the APB effects under dark and light adaptation are due to
activation of mGluR6 receptors.
Previous researchers have shown that L-AP4 can activate mGluR8 (93, 94).
To ensure that L-AP4 effects are not in fact mediated by mGluR8 receptors, we
repeated the above experiment with mGluR8 agonist DCPG (Figure 3E-G, n= 7). The
effects of DCPG were not similar to those of APB and L-AP4. Under a background of
1500 Rh/rod/sec, DCPG increased On response amplitude (Figure 3F, n= 7) but did
not consistently affect Off response amplitudes (Figure 3G, n= 7). These data suggest

29
that APB and L-AP4 are not acting at mGluR8 receptors to affect Off responses under
light adaptation.
Effects of strychnine, picrotoxin, and TPMPA on the APB-induced reduction of Off
responses in the light-adapted mouse retina
Previous research has shown that crossover inhibition between On and Off
pathways occurs via glycinergic neurotransmission. To determine if inhibitory
mechanisms are involved when APB reduces Off responses under light adaptation,
glycine receptor antagonist STR, GABAA/C receptor antagonist PTX, and GABAC
receptor antagonist TPMPA were bath applied before and during application of APB
(Figure 4). When STR was applied to block glycine receptors, APB still reduced Off
response amplitudes under light adaptation (Figure 4A,B, n= 16). Similar results
were seen when both GABAA and GABAC receptors were blocked by PTX (Figure
4C,D, n= 16) and also when only GABAC receptors were blocked by TPMPA (Figure
4E,F, n= 16). Further, simultaneous application of STR, PTX, and TPMPA did not
prevent the reduction of Off response amplitudes by APB (Figure 4G,H, n= 7). These
results indicate that the underlying mechanism of APB-induced reduction of Off
responses under light adaptation does not involve glycinergic or GABAergic
transmission.
Effects of dopamine receptor antagonists on the APB-induced reduction of Off
responses of RGCs

30
As the retina transitions from dark adaptation to light adaptation, dopamine
begins to be released by dopaminergic amacrine cells (95, 96, 97). Dopamine has
been shown to increase the amplitude of light-evoked responses in RGCs (98). Thus,
it is possible that a disruption in dopamine transmission may reduce Off response
amplitudes. Indeed previous studies have shown that APB inhibits dopaminergic
amacrine cells (99, 100). To determine if APB reduces Off responses in the light-
adapted mouse retina by disrupting dopamine receptor activation, D1 and D2
antagonists (SCH 23390 and spiperone, respectively; 20 M) were bath applied
before and during APB application (Figure 5). Our results show that Off responses
decreased with D1 receptors blocked (Figure 5A,B, n= 10) and Off responses
increased with D2 receptors blocked (Figure 5C,D, n= 8). When APB was applied
after D1 block, Off amplitudes no longer decreased. Rather, APB increased Off
amplitudes following D1 block. However, when APB was applied after D2 block, Off
amplitudes still decreased. These results indicate that dopaminergic transmission
involving D1 receptors is necessary for the APB-induced reduction of Off responses
under light adaptation. Similar results were seen when experiments were repeated
with L-AP4 (Figure 5G,H, n= 5).
To further study the role of D1 and D2 receptors in APB-induced reduction in
Off amplitudes, both SCH 23390 and spiperone were co applied (Figure 5E,F, n= 9).
Off amplitudes increased with both D1 and D2 receptors blocked. APB did not
significantly change the Off amplitudes when D1 and D2 receptors were blocked.
Moreover APB did not significantly change Off amplitudes under light adaptation
when SCH 23390, STR, PTX, and TPMPA were applied (Figure 6A,B, n= 7). Under a

31
background light of 150 Rh/rod/sec, APB did decrease Off amplitudes when STR,
PTX, and TPMPA were applied (Figure 6C,D, n= 7). These results indicate that under
mesopic conditions, blocking inhibitory neurotransmission allows APB to reduce Off
amplitudes as was also seen under light adaptation.
HCN channel blocker (ZD 7288) prevented the APB-induced reduction of Off responses
in RGCs in the light-adapted mouse retina
Dopamine has been shown to modulate HCN channels in the retina (101,
102). Additionally, APB inhibits a subgroup of dopaminergic amacrine cells (99,
100). Thus, APB may affect HCN function resulting in the observed reduction in Off
responses under light adaptation. To determine if HCN channels are involved in
APB-induced reduction of Off responses under light adaptation, HCN antagonist ZD
7288 (100 M) was applied before and during bath application of APB (Figure 7).
Under dark adaptation, ZD 7288 did not significantly change On responses (Figure
7A,B, n= 12), but it did significantly decrease Off responses (Figure 7A,C, n= 15). On
the contrary, ZD 7288 decreased both On (Figure 7D,E, n= 11) and Off (Figure 7D,F,
n= 15) responses under light adaptation. The modulatory effects of ZD 7288 were
more pronounced under light adaptation. These data follow a previous study
showing that HCN channels are activated in bright light conditions (103).
When ZD 7288 blocked HCN channels under dark adaptation, APB increased
Off amplitudes similarly to how Off amplitudes increased without HCN channel
block (Figure 7C, n= 15). In contrast, under light adaptation, APB no longer
decreased Off amplitudes when HCN channels were blocked by ZD 7288 (Figure 7F,

32
n= 15). These results indicate that HCN channels are indeed involved in the
mechanism underlying the APB-induced reduction of Off responses under light
adaptation.
APB-induced reduction of Off responses did not depend on the membrane potentials of
RGCs in the light-adapted mouse retina
HCN channels are expressed by Off-cone bipolar cells, but also by RGCs (103,
104). Thus, it is possible that ZD 7288 blocked RGC-expressed HCN channels to
prevent the APB-induced reduction of Off responses. HCN channels are active when
the neuron is hyperpolarized but inactive in depolarized conditions like -40 mV
(102, 105). Therefore, if RGC-localized HCN channels were the major site involved in
the APB-induced reduction of Off responses, then the APB-induced reductions of Off
responses should be eliminated by RGC depolarization. To test that possibility, RGCs
were voltage-clamped at depolarized, resting, and hyperpolarized membrane
potentials under light adaptation and APB was applied (Figure 8). In all three
conditions, APB still decreased Off amplitudes indicating that HCN channels
localized to RGCs do not underlie the APB-induced reduction of Off responses
(Figure 8B, n= 10).

33
Discussion
Previous studies have shown that On and Off pathways cross talk via
crossover inhibition mediated by glycinergic amacrine cells (34, 35, 36, 37). Other
researchers have shown that under dark adaptation, APB activates mGluR6
receptors causing a disruption in crossover inhibition and ultimately increasing Off
response amplitudes (5, 6, 7, 8). The results of the present study demonstrate that
APB does indeed increase Off response amplitudes under dark adaptation, but
effects on Off and On-Off RGCs differ under different adaptations. In the light-
adapted retina, APB significantly decreased Off response amplitudes. When D1
receptors or HCN channels were blocked, APB-induced reduction of Off responses
under light adaptation was prevented. These findings provide evidence that under
light adaptation, On and Off pathways cross talk via dopaminergic amacrine cells
that modulate HCN channel activity at the Off-cone bipolar cell. Moreover, these
findings demonstrate that mGluR6 receptor activity modulates this dopaminergic
mechanism of crosstalk. Taking together the findings in this study and previous
studies, I identified that mGluR6 receptors play an important role in modulating Off
response amplitudes.
APB differentially modulated Off responses of RGCs in dark- and light-adapted mouse
retinas
APB is an mGluR6 agonist that is commonly used in retinal research to block
On pathways. Off responses are also affected by APB, however. Under dark
adaptation, it has been shown that APB blocks APB-sensitive Off responses in the

34
primary rod pathway (81, 106). The remaining Off responses, mainly from the
secondary Off pathway, increase following APB application. Most likely, APB reduces
crossover inhibition via glycinergic amacrine cells under dark adaptation to
increase the Off amplitudes. However, the effects of APB at different adaptation
levels are unclear. The current study used voltage clamp recordings to show that
APB differentially affects Off responses of mouse RGCs depending on adaptation
level. Similar to previous studies, APB increased Off amplitudes under dark
adaptation. Under light adaptation, however, APB decreased Off amplitudes.
We simulated mesopic conditions by adapting the retina to a background
light intensity of 150 Rh/rod/sec. Under these conditions, with both rod and cone
pathways active, APB neither increased nor decreased Off response amplitudes
(107). Therefore, the effect of APB to increase Off amplitudes under dark adaptation
and the effect of APB to decrease Off amplitudes under light adaptation may have
cancelled out. Indeed, when crossover inhibition was blocked with GABA and
glycine antagonists under mesopic conditions, APB decreased Off amplitudes. Taken
together, these results indicate that under mesopic conditions, Off potentiation
requiring crossover inhibition and Off attenuation requiring dopamine occlude each
other.
Glycinergic and/ or GABAergic inhibition and other members of group III mGluRs were
not involved in the APB-induced reduction of Off responses of RGCs in the light-adapted
mouse retina

35
Block of inhibitory neurotransmission with STR, PTX, TPMPA or a
combination of these drugs did not prevent APB-induced reduction of Off responses.
These results indicate that while inhibitory neurotransmission is involved in the
APB effects under dark adaptation, it is not involved in the mechanism under light
adaptation.
In previous studies, APB and L-AP4 have been shown to activate group III
mGluRs including mGluR4, mGluR6, mGluR7, and mGluR8 (108, 109, 110, 111). The
effects of activation of group III mGluRs other than mGluR6 on Off responses in the
mouse retina is currently unclear. In the present study, we applied the mGluR8
agonist DCPG and found that it did not significantly change Off amplitudes. The
results indicate that APB and L-AP4 effects on Off responses occur primarily due to
activation of mGluR6 receptors.
Dopamine receptors were involved in the APB-induced reduction of Off responses of
RGCs in the light-adapted mouse retina
Dopamine release in the retina is stimulated by light adaptation (95, 96, 97).
APB has been previously shown to inhibit dopaminergic amacrine cells, resulting in
a decrease in dopamine release (99, 100). The results from the present study show
that the D1 antagonist SCH 23390 prevents APB from decreasing Off responses
under light adaptation. This indicates that dopaminergic transmission is involved in
the APB-induced reduction of Off responses.
D1 receptor activation has been shown to activate HCN channels (102). When
HCN channels were blocked in the present study, Off response amplitudes

36
decreased. Therefore, it is possible that under light adaptation, APB disrupts D1
activation resulting in HCN inhibition and Off response reduction. When D1
receptors were blocked with SCH 23390, APB actually increased the Off response
amplitudes. These results suggest that under light adaptation, APB has competing
effects. The first effect is the one described above that results in Off response
amplitude reduction. The second is that APB increases Off responses by removing
crossover inhibition, as occurs under dark adaptation. Thus, the mechanism
underlying APB-induced increases of Off responses observed under dark adaptation
is still active under light adaptation. However, under light adaptation, the
mechanism involving disruption of dopamine transmission has more robust effects,
resulting in a net decrease in Off response amplitudes. These findings are supported
by our results showing that APB did not significantly change Off amplitudes when
both the glycinergic and dopaminergic mechanisms were blocked.
While blocking D1 receptors prevented APB-induced reduction of Off
responses under light adaptation, blocking D2 receptors did not. D1 receptor block
decreased Off responses and D2 receptor block increased Off responses. Thus, D1
and D2 receptors may play opposite roles in the retina to modulate light-evoked
responses under light adaptation. It remains unclear why D2 receptor block
significantly increased Off response amplitudes. Future studies should identify the
functional role of D2 receptors in shaping light responses in the retina.
HCN channels were involved in the mechanisms that underlie the APB-induced
reduction of Off responses of RGCs in the light-adapted mouse retina

37
HCN channel block by ZD 7288 prevented the APB-induced reduction of Off
responses. It has been shown that dopamine activates HCN channels, so both
dopamine and HCN may be involved in the APB-induced reduction of Off responses
under light adaptation (102). In the present study, we demonstrated that HCN
channels upstream of RGCs, and not localized to RGCs themselves, are involved in
the APB effects. Previous studies showed that HCN channels are also localized to Off-
cone bipolar cells, cones, and amacrine cells and it is activation of these channels
that is likely modulated by APB application (101, 103, 112, 113, 114).
The results presented here, along with previous studies, indicate that under
light adaptation in the mouse retina, APB inhibits dopaminergic amacrine cells
which inhibits HCN channels resulting in a decrease in Off response amplitudes. The
hypothesized functional circuit illustrated in Figure 9 is as follows: On-cone bipolar
cells activate dopaminergic amacrine cells, which then activate D1 receptors on Off-
cone bipolar cells; after that, HCN channels are activated and glutamate release from
the Off-cone bipolar cell is elevated. APB disrupts this circuit by inhibiting the On-
cone bipolar cells, preventing the activation of dopaminergic amacrine cells. This
circuit demonstrates another way in which On and Off pathways are capable of
crosstalk.

38
Figure 2. The effects of APB on RGC Off responses under different ambient light
conditions. A: Recordings from an On-Off RGC under dark adaptation. APB increased
the amplitude of light-evoked Off responses of this cell, indicated by arrows, and the
response recovered after wash-out. The stimulus light intensity was 350 Rh*/rod/s.
The light onset and offset are indicated above the recording traces. B: Bath
application of APB significantly increased the remaining Off responses of recorded
RGCs under dark adaptation (n= 15, 4 Off cells, and 11 On-Off cells; ** p= 0.001,
paired t-test). C, D: Under a background light of 150 Rh*/rod/s, APB had little effect
on Off responses of RGCs (n= 7, 1 Off cell, 6 On-Off cells; N.S. p= 0.111, paired t-test).
The stimulus light intensity was 6000 Rh*/rod/s. E, F: Under a background light of
1500 Rh*/rod/s, APB significantly decreased the averaged amplitude of Off
responses of the recorded cells (n= 20, 6 Off cells, 14 On-Off cells, ** p= 0.007, paired
t-test). The stimulus light intensity was 15000 Rh*/rod/s. G, H: Under a background
light of 15000 Rh*/rod/s, APB significantly decreased the averaged amplitude of Off
responses of the recorded cells (n= 8, 2 Off cells, 6 On-Off cells; ** p= 0.004, paired t-
test). The stimulus light intensity was 25000 Rh*/rod/s.

39
Figure 2

40
Figure 3. The effects of L-AP4 and DCPG on RGC Off responses under different
ambient light conditions. A, B: Under dark adaptation, mGluR6 agonist L-AP4
significantly increased the amplitudes of Off responses of recorded RGCs (n= 8, 2 Off
cells, 6 On-Off cells; ** p= 0.001, paired t-test). The stimulus light intensity was 350
Rh*/rod/s. C, D: Under the background light of 1500 Rh*/rod/s, L-AP4 significantly
decreased the amplitudes of Off responses of recorded RGCs (n= 8, 1 Off cell, 7 On-
Off cells; ** p= 0.009, paired t-test). The stimulus light intensity was 15000
Rh*/rod/s. E, F, and G: Under a background light of 1500 Rh*/rod/s and a stimulus
light intensity of 15000 Rh*/rod/s, mGluR8 agonist DCPG significantly increased the
averaged amplitude of On responses of recorded RGCs (n= 7, 7 On-Off cells; * p=
0.040, paired t-test). However, DCPG had little effect on the averaged amplitude of
Off responses of recorded RGCs (n= 7, 7 On-Off cells; N.S. p= 0.263, paired t-test).

41
Figure 3

42
Figure 4. Antagonists of glycine, GABAA, and GABAC receptors did not eliminate the
APB-induced reductions of Off responses of RGCs in light-adapted mouse retinas.
Recordings were made under the background light of 1500 Rh*/rod/s from three
On-Off RGCs as shown in A, C, and E, respectively. A, C, and E: APB still decreased
the Off responses of the three cells following perfusion with STR, PTX, and TPMPA,
respectively. B, D, and F: The APB-induced reductions of Off responses were not
eliminated by either STR, or PTX, or TPMPA, respectively. G and H: STR, PTX and
TPMPA together did not prevent the APB-induced reductions of Off responses of
RGCs in light-adapted mouse retinas (** p= 0.006, *p= 0.014, ** p=0.005, and ** p=
0.001 in B, D, F and H, respectively, paired t-test).

43
Figure 4

44
Figure 5. Effects of dopamine receptor antagonists on the APB-induced reductions
of Off responses of RGCs in light-adapted mouse retinas. All recordings were made
from RGCs with a background light of 1500 Rh*/rod/s and a stimulus light intensity
of 15000 Rh*/rod/s. A, B: A D1 blocker, SCH 23390, significantly decreased the light-
evoked Off responses of the recorded cells, but it prevented and reversed the APB-
induced reduction of the Off responses in these cells (n= 10, 2 Off cells, 8 On-Off
cells; * p= 0.027 for SCH 23390 alone and ** p= 0.007 for APB with SCH 23390,
paired t-test). C, D: A D2 blocker, spiperone, significantly increased the light-evoked
Off responses of the recorded cells, but did not prevent the APB-induced reduction
of the Off responses in these cells (n= 8, 3 Off cells, 5 On-Off cells; ** p= 0.008 for
spiperone alone and * p= 0.015 for APB with spiperone, paired t-test). E, F: Bath
application of D1 and D2 blockers together significantly increased the light-evoked
Off responses in the recorded cells. APB, when applied with SCH 23390 and
Spiperone, had little effect on Off responses of RGCs (n= 9, ** p=0.006, N.S. p= 0.966,
paired t-test). G, H: SCH 23390 significantly decreased the light-evoked Off
responses of the recorded cells, but it prevented and reversed the L-AP4-induced
reduction of the Off responses in these cells (n= 5, 1 Off cell, 4 On-Off cells; * p=
0.022 for SCH 23390 alone and * p= 0.013 for L-AP4 with SCH 23390, paired t-test).
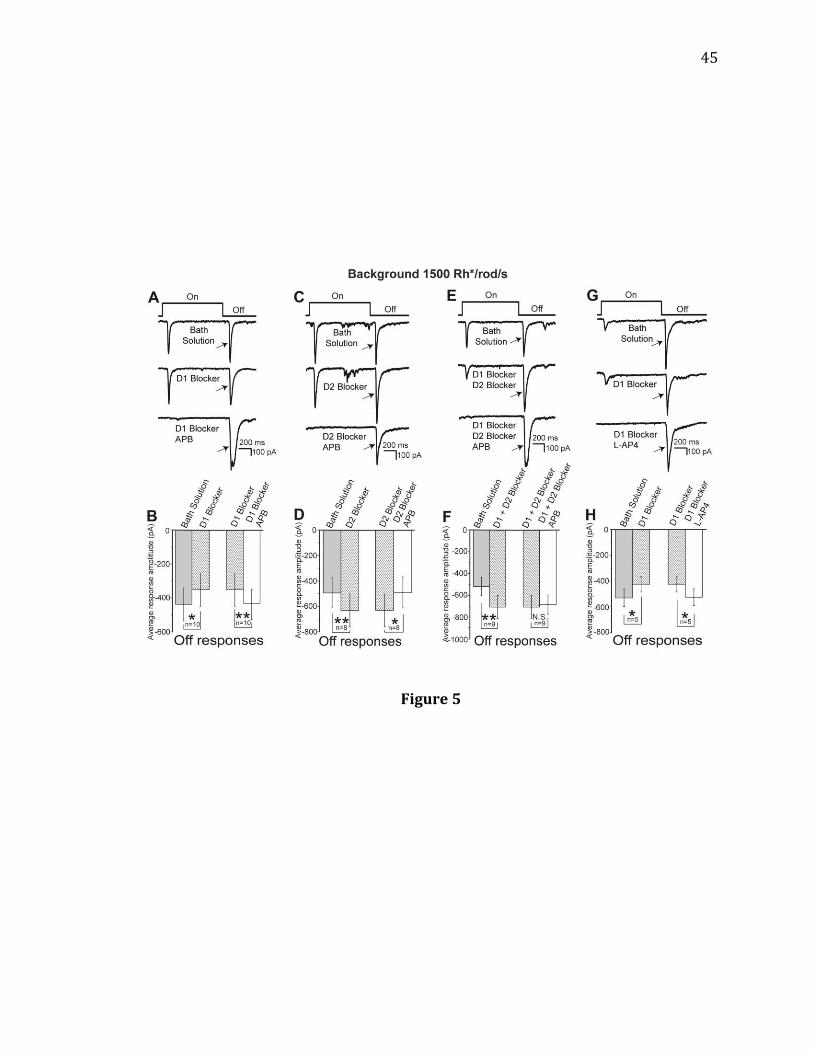
45
Figure 5

46
Figure 6. Effects of APB on light-evoked Off responses when D1 receptors and
inhibitory receptors are blocked. A, B: Under a background light of 1500 Rh*/rod/s,
APB had little effect on the averaged amplitude of Off responses of the recorded cells
after D1, GABA, and glycine receptors blocked together (n= 7, N.S. p= 0.101, paired t-
test). The stimulus light intensity was 15000 Rh*/rod/s. C, D: Under a background
light of 150 Rh*/rod/s, following GABA and glycine receptor block by PTX, TPMPA,
and STR, bath application of APB decreased the averaged amplitude of Off responses
of the recorded cells (n= 7, * p= 0.026, paired t-test).
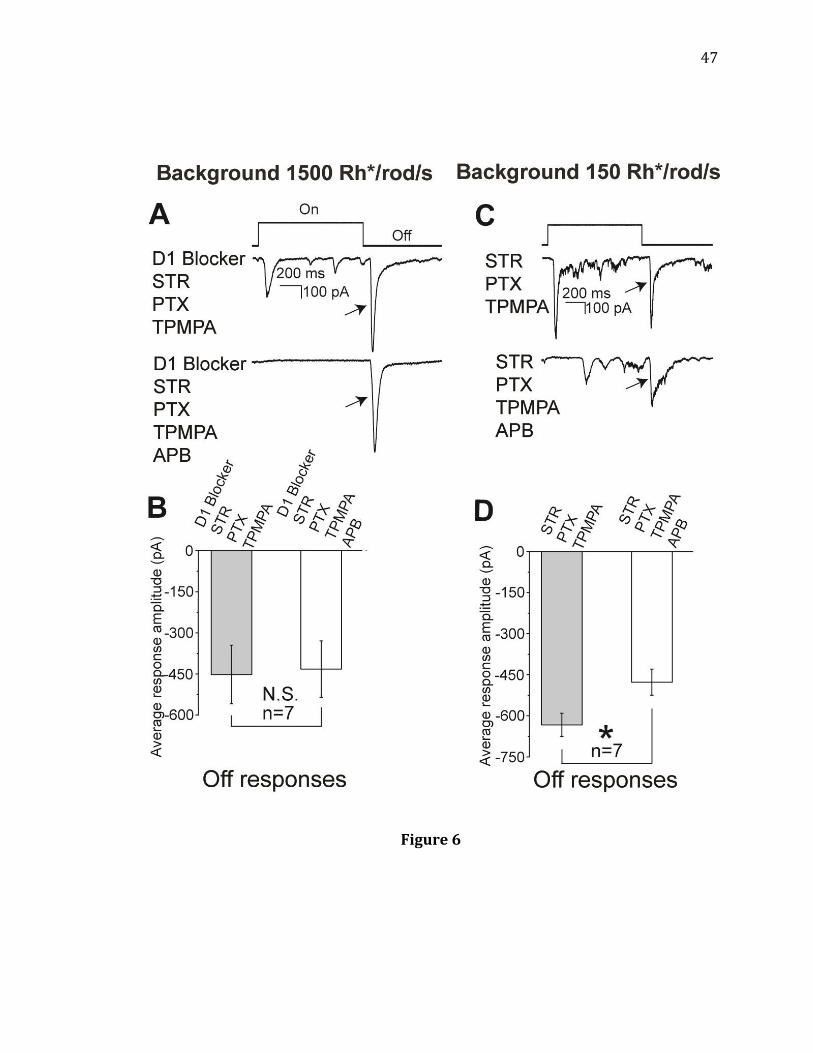
47
Figure 6

48
Figure 7. HCN channel antagonist ZD 7288 prevented the APB-induced reduction in
Off responses under light adaptation. A: Recordings from an On-Off RGC in a dark-
adapted retina. The stimulus light intensity was 350 Rh*/rod/s. B: ZD 7288 did not
significantly decrease the light-evoked On responses of the recorded On-Off cells
under dark adaptation (n= 12, N.S. p= 0.170, paired t-test). C: ZD 7288 significantly
decreased the amplitudes of light-evoked Off responses of the recorded cells under
dark adaptation (n= 15, 3 Of cells, 12 On-Off cells, * p= 0.041, paired t-test). The
average percent reduction was (9.2 ± 1.3) %. After blocking HCN channels with ZD
7288, APB still significantly increased the amplitude of light-evoked Off responses of
the recorded cells under dark adaptation (n= 15, ** p= 0.001, paired t-test). D:
Recordings were made from an On-Off ganglion cell under a background light of
1500 Rh*/rod/s. The stimulus light intensity was 15000 Rh*/rod/s. E: ZD 7288
significantly decreased the light-evoked On responses of the recorded On-Off cells
under the background light of 1500 Rh*/rod/s (n= 11, * p= 0.047, paired t-test). F:
ZD 7288 significantly decreased the amplitudes of light-evoked Off responses of the
recorded cells under a background light of 1500 Rh*/rod/s (n= 15, 4 Off cells, 11
On-Off cells, * p= 0.015, paired t-test). The average percent reduction was (47.6 ±
7.5) %. After blocking the HCN channels with ZD 7288, APB did not further decrease
the averaged amplitude of light-evoked Off responses in the same recorded cells (n=
15, N.S. p= 0.131, paired t-test).
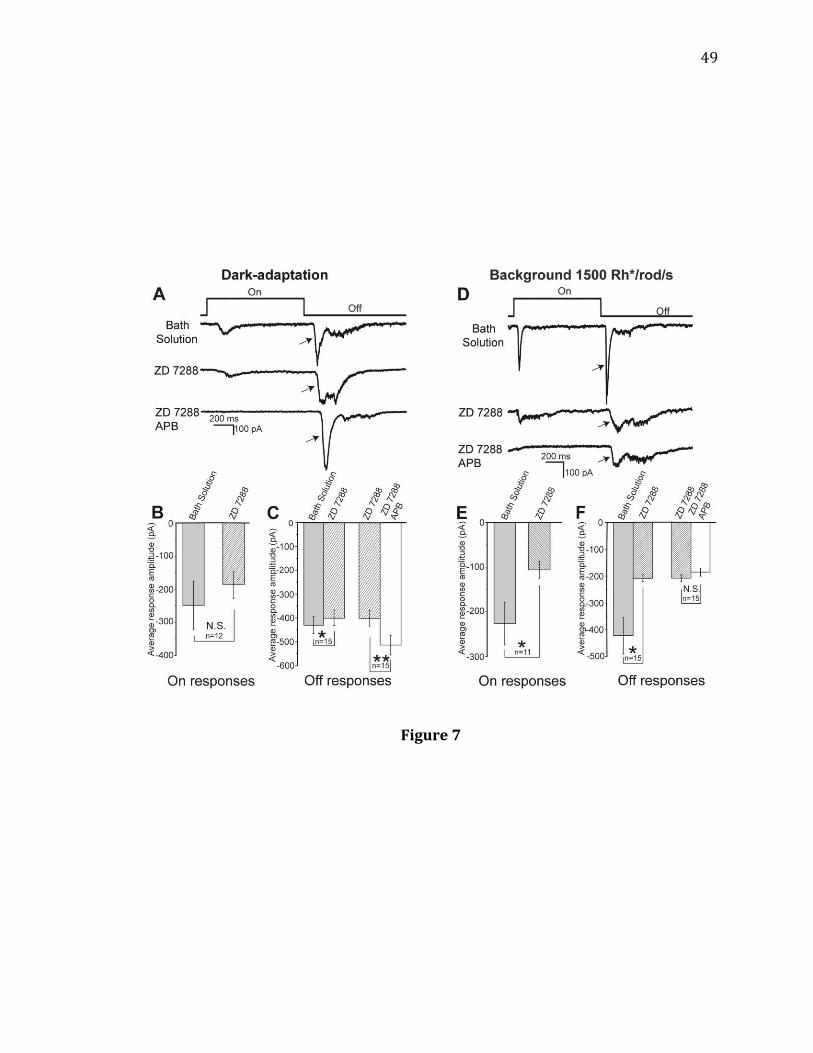
49
Figure 7

50
Figure 8. A depolarized holding potential of -40 mV, which inactivates HCN
channels in the recorded RGCs, did not prevent the APB-induced reduction of Off
responses under light adaptation. A: Under a background light of 1500 Rh*/rod/s,
recordings were made from an On-Off ganglion cell with holding potentials at -40
mV (left panel), -60 mV (middle panel), and -90 mV (right panel). The stimulus light
intensity was 15000 Rh*/rod/s. Bath application of APB decreased the Off
responses of these cells at all three holding potentials. B: Under the background
light of 1500 Rh*/rod/s, the averaged Off response amplitudes of the recorded cells
were significantly reduced by APB at the holding potentials of -40 mV, -60 mV, and -
90 mV (n= 10, 2 Off cells; 8 On-Off cells, * p= 0.032 at -40 mV, * p= 0.015 at -60 mV,
and ** p= 0.004 at -90 mV, paired t-test).
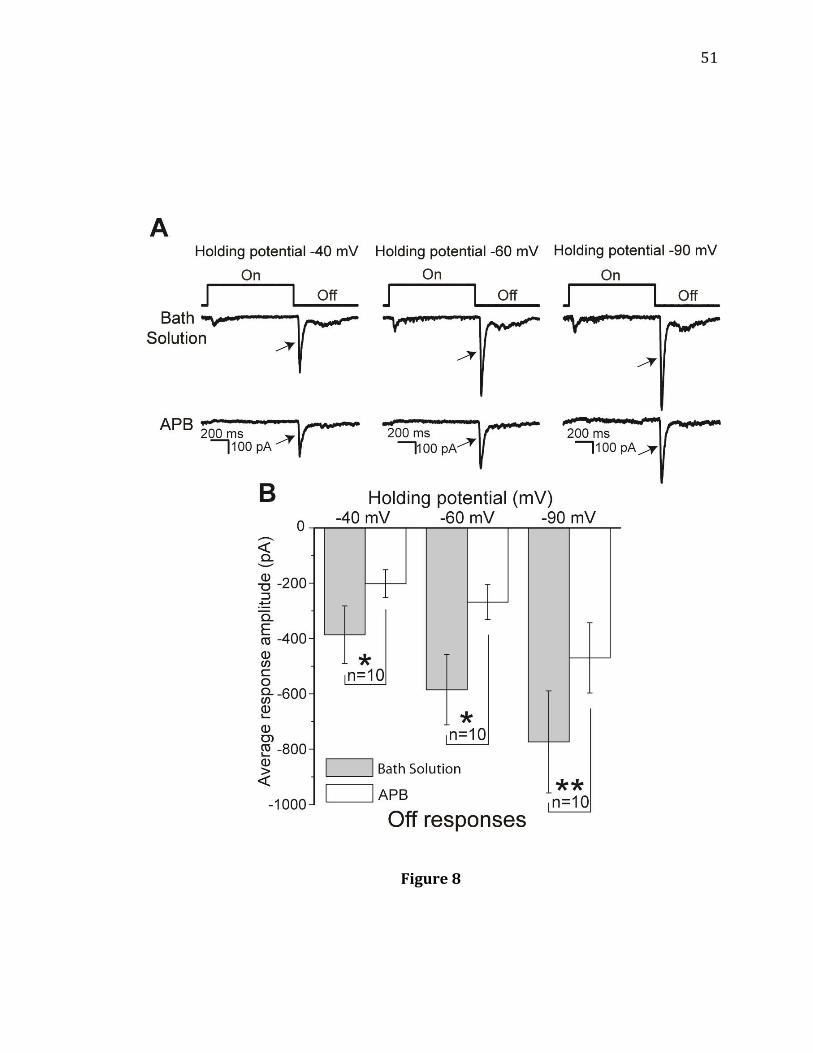
51
Figure 8

52
Figure 9. A diagram showing a functional circuit from On pathways to Off pathways
under light adaptation: On-cone bipolar cells activate dopaminergic amacrine cells
that release dopamine, which in turn to activates D1 receptors on Off-cone bipolar
cells. Then, D1 receptors activate HCN channels on Off-cone bipolar cells. C: cone; ON
CB: On-cone bipolar cell; OFF CB: Off-cone bipolar cell; ON RGC: On retinal ganglion
cell; OFF RGC: Off retinal ganglion cell; DA: dopaminergic amacrine cell; D1:
dopaminergic type 1 receptor; HCN: hyperpolarization-activated cyclic nucleotide-
gated channel.
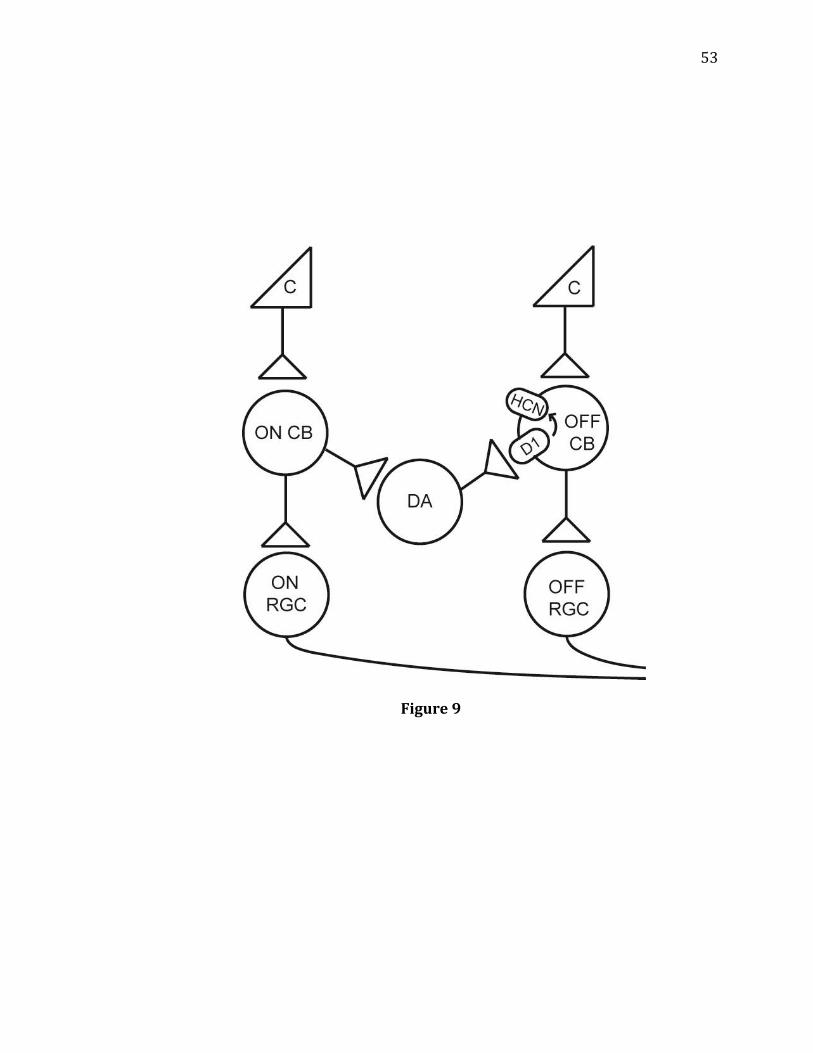
53
Figure 9

54
CHAPTER 2
CB1Rs modulate Off responses in light-adapted mouse retina Introduction
CB1Rs are metabotropic G-protein coupled receptors that are involved in
regulating neurotransmitter release (115). When post-synaptic neurons are excited
and depolarize, they release eCBs that travel retrograde to the presynaptic neuron
and bind to CB1Rs (79). Following signal transduction, neurotransmitter release
from the presynapse is reduced (116). Activity-dependent eCB suppression of
excitatory neurotransmitter release is known as depolarization-induced
suppression of excitation (DSE), and activity-dependent eCB suppression of
inhibitory neurotransmitter release is known as depolarization-induced
suppression of inhibition (DSI) (75, 117, 118). Thus far, the majority of CB1R
research has focused on functions in the brain, but CB1Rs have also been discovered
in the retina.
CB1Rs are expressed widely in the retina of various species including mouse.
One study has shown that CB1Rs are localized to rod synaptic terminals, cone
synaptic terminals, amacrine cells, RGCs, and sporadically throughout the IPL (43).
Another study used a different antibody and found CB1R expression in rod bipolar
cells, PKC-immunoreactive GABAergic amacrine cells, in the OPL, in the IPL, but not
in the GCL (42). While the localization studies do not completely agree, it is clear

55
that CB1Rs are positioned to affect neurotransmission at glutamatergic and possibly
GABAergic synapses.
Previous studies investigating the functions of CB1Rs have mainly focused on
shaping ion channel currents such as the HVA Ca2+ currents in photoreceptors,
bipolar cells, and cultured RGCs (9). Application of CB1R agonists has been shown to
decrease the release of neurotransmitters like dopamine and glutamate, which
follows with previous studies in the brain showing that CB1R activation prevents
neurotransmitter release (57, 59). Only one study investigated CB1R and its role in
shaping light-evoked responses (63). The researchers found that the cone response
to light offset returned to baseline faster following application of CB1R agonist.
However, previous studies have not demonstrated the role of the eCB system in
shaping light responses in RGCs, the final output neurons of the retina.
In the current study, patch clamp recordings were made from RGCs in mouse
retinas under light adaptation. I found that a phenomenon similar to DSE occurs in
some RGCs, but it is unclear if CB1Rs are involved in the underlying mechanism.
Further, I found that there are two functional populations of RGCs: one where CB1R
agonists increase Off responses, and one where CB1R agonists decrease Off
responses. The results presented in this study suggest that CB1R activation
increases Off response amplitudes by disinhibiting Off-cone bipolar cells.

56
Materials and Methods
The basic methods used in this study were similar to those used previously in
this laboratory (81, 82, 83, 84). The materials and methods used for the experiments
outlined in this Chapter are also similar to those described in Chapter 1. All
procedures were performed under light adaptation with a background light
intensity of 2.9x104 photons/m2/s.
Retinal preparation. Retinal preparations performed in these experiments are
similar to those described in Chapter 1. For current-clamp experiments, patch
electrodes were filled with a solution containing (in mM) potassium gluconate 120,
NaCl 10, MgCl2 0.5, CaCl2 0.5, HEPES 10, EGTA 5, 0.5% Lucifer yellow, pH 7.4,
osmolarity 280 mOsm. The ECl with this internal solution was approximately -58
mV.
Electrophysiology. The basic electrophysiology methods were similar to those
described in Chapter 1, except that all experiments were performed under light
adaptation. For DSE experiments, RGCs were first maintained at -58 mV, the
chloride reversal potential. Three light responses at least 10 seconds apart were
recorded (pre-depolarization) and then the membrane potential was depolarized to
0 mV for 10 seconds before returning to -58 mV for the rest of the recording.
Immediately following membrane depolarization, light responses were recorded
every ten seconds for 3 minutes. The first three light responses after depolarization
are henceforth referred to as “post-depolarization” and the first three light

57
responses at least two minutes after depolarization are referred to as “recovery.”
For current clamp experiments, current was applied so that the baseline membrane
voltage was approximately -58 mV.
Light stimulus. The light stimuli were presented to the light-adapted retina using the
Lucivid, as described in Chapter 1. For these studies, a background light of constant
brightness, 2.9x104 photons/m2/s, was provided full-field by lights in the
recording room. At this background light intensity, rods are completely inactivated
and only cone-mediated responses are recorded (83).
The method of recording from light-adapted retinas is similar to those in our
previous studies. Throughout the procedure, including during retinal preparation,
dissection, and recordings, the retina was light-adapted. Light stimuli with intensity
greater than 2.9x104 Rh/rod/sec were used to evoke light responses from ganglion
cells.
Drug application. 2-arachidonoyl glycerol (2-AG; 5 M; Tocris), AM251 (2 M;
Tocris), APB (20 M; Sigma), O-2050 (2-10 M; Tocris), (1,2,5,6-tetrahydropyridin-
4-yl)methylphosphinic acid (TPMPA; 50 M; Sigma), WIN 55,212-2 (WIN; 10 M;
Sigma) were freshly dissolved in MEME on the day of the experiment and
administered through a gravity-fed line.

58
Results
DSE in RGC light-evoked Off responses
In certain preparations, when postsynaptic neurons are transiently
depolarized, glutamate release from presynaptic neurons is reduced resulting in
smaller EPSCs in the postsynaptic cell (118). It is believed that the depolarized
postsynaptic neuron releases eCBs that travel retrogradely to activate
presynaptically localized CB1Rs that reduce neurotransmitter release. To determine
if DSE occurs in the retina, specifically at the synapses between Off-cone bipolar
cells and RGCs, morphologically identified Off and On-Off mouse RGCs were voltage-
clamped and light-evoked Off responses were recorded before and after transient
membrane depolarization of RGCs (Figure 10).
Light responses were recorded both before and immediately after a 10
second RGC-membrane depolarization to 0 mV that was designed to evoke DSE.
Responses were again recorded 2 minutes after membrane depolarization to
determine if response amplitudes recovered from DSE. Experiments were first
carried out in MEME without drugs applied to the bath solution (Figure 10A-C, n=
12). The averaged response amplitude after depolarization did not significantly
decrease in the 12 total recorded RGCs (Figure 10B, n= 12). However, in one
population of RGCs, Off response amplitudes significantly decreased following
depolarization, which is consistent with DSE (Figure 10A,C, n= 7). The decrease in
amplitude following depolarization in these RGCs was 23%, similar to EPSC
amplitude reductions reported in other studies (45, 79). In these cells where DSE
occurred, Off responses returned to pre-depolarization amplitudes by 2 minutes

59
post-depolarization, similar to the recovery time reported in the other studies.
These results suggest that DSE occurs at some, but not all synapses between Off-
cone bipolar cells and RGCs.
The protocol used to evoke DSE in control conditions was repeated following
bath application of 2M CB1R antagonist O-2050 (Figure 10D-F, n= 10). Again, the
averaged response amplitude after depolarization did not significantly decrease in
the 10 total recorded RGCs (Figure 10E, n= 10). However, in the same 7 RGCs where
DSE occurred in control conditions, DSE also occurred following bath application of
O-2050 (Figure 10F, n= 7). The amplitude reduction due to depolarization in the
presence of O-2050 was 11%. These results indicate that blocking CB1Rs may
reduce, but not totally prevent DSE in this preparation. Moreover, the observed DSE
may be mediated by mechanisms that do not involve CB1Rs. Previous researchers
using other preparations have also found that CB1R antagonists could not block DSE
(118, 119).
Effects of CB1R antagonist O-2050 on light-evoked Off responses of RGCs
CB1Rs are localized to glutamatergic synapses in the retina and CB1R
antagonists have been shown to increase neurotransmitter release in other
preparations (120). Therefore we hypothesized that bath application of CB1R
antagonist O-2050 would increase glutamate release in Off pathways and increase
Off response amplitudes. To further our understanding of eCB functions in the retina
and determine if CB1R antagonists increase light-evoked Off responses in RGCs, Off
responses were recorded before and after bath application of 2M CB1R antagonist

60
O-2050 (Figure 11). Consistent with our hypothesis, O-2050 significantly increased
Off response amplitudes in one population of Off and On-Off RGCs (Figure 11A-C, n=
8). However in another population of Off and On-Off RGCs, O-2050 significantly
decreased Off response amplitudes (Figure 11D-F, n= 28). Experiments were
repeated with CB1R antagonist AM251 (2 M) and similar results were found (data
not shown). These data suggest that there are two functional populations of RGCs:
blocking CB1Rs in one population of RGCs increases Off responses, and blocking
CB1Rs in another population of RGCs decreases Off responses.
Effects of CB1R agonist WIN on light-evoked Off responses of RGCs
We originally operated under the hypothesis that activating CB1Rs with WIN
would reduce glutamate release and reduce Off responses (9). To determine if CB1R
agonists decrease Off response amplitudes, Off responses were recorded before and
after bath application of 10 M CB1R agonist WIN (Figure 12). In one population of
Off and On-Off RGCs, WIN significantly decreased Off response amplitudes, as we
hypothesized (Figure 12D-F, n= 9). However, in another population of Off and On-
Off RGCs, WIN significantly increased Off response amplitudes (Figure 12A-C, n= 7).
These results are consistent with our previous findings of two functional
populations of RGCs after O-2050 application. Experiments were repeated with the
eCB and CB1R agonist 2-AG (5 M) and similar results were found (data not shown).
Washout of WIN or 2-AG for between 20 and 30 minutes did not reliably return Off
amplitudes to control levels.

61
Effects of CB1R agonist 2-AG when crossover inhibition is blocked with mGluR6 agonist
APB
On pathways inhibit Off pathways via crossover inhibition (31, 32, 33, 34).
The circuit for crossover inhibition is as follows: cone > On-cone bipolar cell >
glycinergic amacrine cell > Off-cone bipolar cell > RGC. Applying mGluR6 agonist
APB blocks crossover inhibition. Activation of mGluR6 receptors, which are
localized to On-cone bipolar cell dendrites, results in hyperpolarization of On-cone
bipolar cells. This prevents glycinergic amacrine cells from being activated,
disinhibits Off-cone bipolar cells, and increases Off responses (5, 6, 7, 8). As
explained previously, in one population of Off and On-Off RGCs, application of CB1R
agonists increased the amplitude of Off responses, contrary to our original
hypothesis. To determine if CB1R agonists increase Off responses by reducing
crossover inhibition of Off pathways, we applied APB (20 M) to block crossover
inhibition and then recorded Off responses before and after bath application of 5 M
CB1R agonist 2-AG (Figure 13). With crossover inhibition blocked by APB, 2-AG still
increased the amplitude of Off responses in one population of Off and On-Off RGCs
(Figure 13A,B, n= 15) and decreased the amplitude of Off responses in another
population of Off and On-Off RGCs (Figure 13C,D, n= 10). APB eliminated On
responses in every recorded On-Off cell, indicating that On pathways were blocked,
On-cone bipolar cells were hyperpolarized, and crossover inhibition was indeed
blocked. These results indicate that CB1R agonists increase Off response amplitudes
in one population of Off and On-Off RGCs, but the underlying mechanism does not
involve modulation of crossover inhibition.

62
Off response amplitude enhancement by CB1R agonists is prevented when GABAC
receptors are blocked by TPMPA
Much of the CB1R immunoreactivity in the retina is localized to
glutamatergic synapses (9). However, in one study, CB1R immunoreactivity was
discovered at the axon terminals of a subtype of PKC-immunoreactive GABAergic
amacrine cell (42). Previous researchers studying eCBs in the brain have found that
eCBs can reduce GABA release from presynaptic neurons (75, 121). Activation of
CB1Rs localized to GABAergic amacrine cells could prevent GABA release and
disinhibit cells that receive inputs from GABAergic amacrine cells. Off-cone bipolar
cells that preferentially express GABAC receptors, as opposed to GABAA receptors,
receive inputs from GABAergic amacrine cells (73, 74, 122). On-cone bipolar cells
also express GABAC receptors (123). To determine if CB1R agonists increase Off
amplitudes in one population of RGCs by reducing GABAergic inhibition onto Off-
cone bipolar cells, we applied APB (20 M) to block crossover inhibition, we applied
TPMPA (50 M) to block GABAC receptors, and we recorded Off responses before
and after application of 5 M 2-AG (Figure 14). Bath application of CB1R agonist 2-
AG significantly reduced Off response amplitudes in all recorded Off and On-Off
RGCs (Figure 14B, n= 5). Thus, increases in Off response amplitudes by a CB1R
agonist were prevented when GABAC receptors were blocked. These results suggest
that CB1R agonists may disinhibit Off-cone bipolar cells that mostly express GABAC
receptors resulting in greater Off amplitudes in one population of Off and On-Off
RGCs. We suspect that CB1R agonists reduce GABA release from GABAergic
amacrine cells that both express CB1Rs and synapse onto Off-cone bipolar cells.

63
Discussion
Studies of the retinal eCB system have focused primarily on CB1R
localization and CB1R-mediated modulation of isolated channel currents (9). The
available CB1R localization studies do not completely agree (42, 43). However, each
study has reported CB1R expression at the glutamatergic synapses in the OPL and
IPL as well as at amacrine cells. In particular importance to the present study, CB1R
expression has also been reported at the axon terminals of a subtype of PKC-
immunoreactive GABAergic amacrine cell. CB1Rs have been found to modulate
voltage-dependent potassium, chloride, and calcium currents in various retinal
neurons (42, 44, 47, 48, 49, 50, 51, 52). In addition, CB1R agonists decrease
generalized dopamine and glutamate release in the retina, consistent with brain
studies showing that CB1R activation decreases neurotransmitter release (58, 59).
In the only study that investigated the role of CB1Rs in shaping light-evoked
responses, researchers found that CB1R agonist WIN decreased the latency for cone
responses to return to baseline following light offset (63). However, the role of
CB1Rs in shaping light-evoked response output has not yet been investigated. The
results of the present study demonstrate that DSE, a phenomenon involving CB1Rs
that has been studied extensively in the brain, occurs in some but not all RGCs.
There are functionally two populations of Off and On-Off RGCs: 1) in the first
population, CB1R activation increases Off amplitudes; 2) in the second population,
CB1R activation decreases Off amplitudes. Reduction of crossover inhibition is not
the mechanism underlying CB1R agonist-mediated increases in Off response
amplitude. Rather, our results indicate that CB1R activation can reduce GABAergic

64
inhibition onto Off-cone bipolar cells resulting in the observed increase in Off
response amplitudes. Taken together, these findings provide evidence that under
light adaptation, CB1Rs participate in shaping light-evoked Off responses.
DSE occurs in some RGCs but CB1Rs may not be involved
DSE occurs when a postsynaptic neuron depolarizes and subsequently
synthesizes eCBs that activate CB1Rs localized to presynaptic glutamatergic
neurons (118, 124). The activation of CB1Rs leads to a reduction of glutamate
release from the presynaptic neuron and a subsequent reduction in excitation of the
postsynaptic neuron. DSE, along with its counterpart DSI, which occurs at inhibitory
synapses, is thought be a key mechanism of eCB functions in the brain. However,
even though CB1Rs have been discovered in the retina, DSE has not yet been studied
in the retina. The current study used voltage clamp recordings to show that a
phenomenon similar to DSE occurs at the synapse between Off-cone bipolar cells
and RGCs.
Under light adaptation, we recorded light-evoked Off responses in Off and
On-Off RGCs before (pre-depolarization) and after (post-depolarization) a 10 second
RGC membrane depolarization to 0 mV. In previous studies, the excitatory current-
reducing effect of DSE weakened over several minutes until current amplitudes
returned to pre-depolarization levels (118, 124). Therefore, we also recorded Off
responses 2 minutes after depolarization to see if Off amplitudes returned to pre-
depolarization levels (recovery). Off response amplitudes decreased following
depolarization in some but not all recorded RGCs. This may be due to different

65
expression levels of CB1Rs at different synapses. In cells where DSE did occur, CB1R
antagonist O-2050 reduced but did not eliminate the DSE. This data may indicate
that at least some of the DSE we observed in RGCs is not mediated by CB1R
activation. It is possible that the DSE that is not blocked following O-2050
application is mediated by an undiscovered cannabinoid receptor. Indeed, in a
previous study, cannabinoid receptor agonist WIN was able to reduce EPSCs
recorded from hippocampal neurons in a CB1R knockout mouse (119). The
researchers in that study hypothesize that cannabinoids may be acting at
presynaptic glutamatergic neurons expressing non-CB1R cannabinoid receptors.
Taken together, these results indicate that DSE occurs in some RGCs but may only be
partially mediated by CB1Rs.
Effects of manipulating retinal CB1R activity on light-evoked Off responses in RGCs
CB1Rs are expressed at glutamatergic synapses in the retina (9). In a 2008
review, Dr. Yazulla hypothesized that application of CB1R agonists would decrease
glutamate release at these synapses and reduce light-evoked responses. Thus, we
hypothesized that blocking CB1Rs with specific antagonists would increase light-
evoked responses. The current study used voltage clamp recordings to show that O-
2050 does indeed increase Off responses in one population of RGCs but decreases
Off responses in another population of RGCs.
Consistent with our hypothesis, O-2050 increased the amplitude of light-
evoked Off responses in some RGCs. Taking into account CB1R localization studies,
it is likely that O-2050 is blocking CB1Rs and preventing eCBs from reducing

66
glutamate release. Contrary to our hypothesis, however, O-2050 reduced the
amplitude of light-evoked Off responses in other RGCs. To test if O-2050 similarly
modulated RGC action potentials, a major way that the retina communicates with
the brain, we repeated the experiment in current clamp (data not shown). In some
cells, Off action potential frequency increased following O-2050 application as we
hypothesized, but in other cells Off action potential decreased. This data follows
with the voltage clamp data above. In addition, voltage clamp experiments were
repeated with another CB1R antagonist, AM251. Similar results to those seen with
O-2050 were also seen with AM251, indicating that the effect was mediated by
CB1Rs.
We had hypothesized that CB1R agonists would decrease the amplitude of
Off responses. In one population of RGCs, CB1R agonist WIN decreased Off
responses as we hypothesized. Contrary to our hypothesis, however, WIN increased
Off responses in another population of RGCs. Experiments were repeated with the
CB1R agonist 2-AG yielding similar results. Washout of CB1R agonists was carried
out but effect was non-reversible. This was not surprising since difficulty in washing
out the lipophilic CB1R agonists has been reported in the past (50, 116, 125, 126).
These data follow with the O-2050 data indicating that there are functionally two
populations of RGCs in terms of CB1R effects on Off responses. We believe that in
cells where CB1R antagonists increased Off responses, CB1R agonists would have
decreased Off responses. Further, we believe that in cells where CB1R antagonists
decreased Off responses, CB1R agonists would have increased Off responses. Due to

67
difficulties with washouts, however, carrying out the above experiment would have
proved problematic.
Crossover inhibition does not underlie CB1R agonist-mediated increases in Off
response amplitude
Given the expression of CB1Rs at retinal glutamatergic synapses, we
hypothesized that CB1R agonists would reduce glutamate release and decrease Off
responses. In many RGCs, however, CB1R agonists increased Off responses.
Therefore, we began to investigate alternative ways in which CB1R agonists may
increase Off responses. On pathways inhibit Off pathways via crossover inhibition. It
is possible that CB1R agonists increased Off responses in some RGCs by reducing the
amount of crossover inhibition of Off pathways. For instance, if CB1R agonists
decreased glutamate release in On pathways, glycinergic amacrine cells could be
inhibited and release of glycine onto Off pathways could be reduced. This crossover
inhibition can be reliably blocked with mGluR6 agonist APB. In the present study,
we showed that even with crossover inhibition blocked by APB, CB1R agonist still
increased Off response amplitudes in one population of RGCs and decreased Off
response amplitudes in another population of RGCs. These results suggest that a
reduction in crossover inhibition is not the mechanism underlying the CB1R
agonist-mediated increases in Off amplitudes in one population of RGCs.
GABAC-mediated inhibition is necessary for the CB1R agonist-mediated increases in Off
response amplitude

68
Since we found that crossover inhibition did not underlie CB1R agonist-
mediated Off response amplitude increases, we explored another possible
explanation. Studies have shown that CB1Rs are not only localized to glutamatergic
synapses (9). Rather, studies agree that CB1Rs are expressed at the axon terminals
of amacrine cells. In one study, CB1R immunoreactivity was found at the axon
terminals of a subtype of PKC-immunoreactive GABAergic amacrine cell indicating
that CB1Rs may regulate retinal GABA release (43). These GABAergic amacrine cells
often feed forward onto the axon terminals of cone bipolar cells (127). Therefore,
CB1R agonists may reduce GABA release from these amacrine cells and disinhibit
Off-cone bipolar cells upstream of our recorded Off and On-Off RGCs. In the present
study, we found that with crossover inhibition blocked by APB and with GABAC
receptors blocked by TPMPA, 2-AG was no longer able to increase Off response
amplitudes recorded from RGCs. In the retina, GABAC receptors are primarily
expressed by On and Off-cone bipolar cells (128). Therefore, we believe that a
reduction in GABAergic input at the synapse between amacrine cells and Off-cone
bipolar cells underlies the CB1R agonist-mediated increases in Off responses in one
population of RGCs.
Taken together, the results of this study and results from previous studies
indicate that Off responses can be differentially modulated by CB1R agonists and
antagonists. Our hypothesized circuit for the CB1R agonist-mediated increases in Off
response, as illustrated in Figure 15, is as follows: 1) CB1R agonist activates CB1Rs
at GABAergic amacrine cells that provide inputs to Off-cone bipolar cells, 2) GABA
release is reduced and GABAC receptors on Off-cone bipolar cells are not activated

69
resulting in disinhibition, and 3) Off-cone bipolar cells release more glutamate onto
Off and On-Off RGCs. For the population of RGCs where CB1R agonists decreased Off
responses we hypothesize the following circuit: 1) CB1R agonist activates CB1Rs at
cone and Off-cone bipolar cell synaptic terminals, 2) Glutamate release along the Off
pathway is reduced, and 3) RGC Off responses are reduced. We believe that both
circuits may be involved in modulating individual Off responses. However, the
relative strength of one circuit compared to the other due to differences in CB1R
expression, number of synapses, or another variable may result in the two
populations of RGCs we have observed in this study.

70
Figure 10. DSE of light-evoked Off EPSCs recorded from RGCs. Recordings were
made from RGCs with a background light of 1.6x104 photons/m2/s and a stimulus
light intensity of 6.7x106 photons/m2/s. A: Averaged Off responses before RGC
depolarization (black trace), shortly after depolarization (dotted trace), and two
minutes after depolarization (gray trace) in one On-Off RGC perfused with normal
bath solution. In this cell, 10 second RGC membrane depolarization temporarily
reduced the averaged Off response amplitude. The averaged Off response amplitude
recovered two minutes after depolarization. B: RGC depolarization did not
significantly reduce Off response amplitudes in all recorded RGCs (n= 12, 2 Off cells,
10 On-Off cells; N.S. p= 0.158, paired t-test). C: RGC depolarization significantly
reduced Off response amplitudes in one population of recorded RGCs (n= 7, 7 On-Off
cells, * p= 0.041, paired t-test). D: Averaged Off responses before RGC depolarization
(black trace), shortly after depolarization (dotted trace), and two minutes after
depolarization (gray trace) in one On-Off RGC with bath applied CB1R antagonist O-
2050. In this cell, 10 second RGC membrane depolarization temporarily reduced the
averaged Off response amplitude. The averaged Off response amplitude recovered
two minutes after depolarization. E: RGC depolarization did not reduce Off response
amplitudes in all recorded cells with CB1Rs blocked by O-2050 (n= 10, 2 Off cells, 8
On-Off cells, N.S. p= 0.801, paired t-test). F: RGC depolarization significantly reduced
Off response amplitudes in the same cells depicted in panel C even though O-2050
was included in the bath solution (n= 7, 7 On-Off cells, ** p= 0.009, paired t-test).
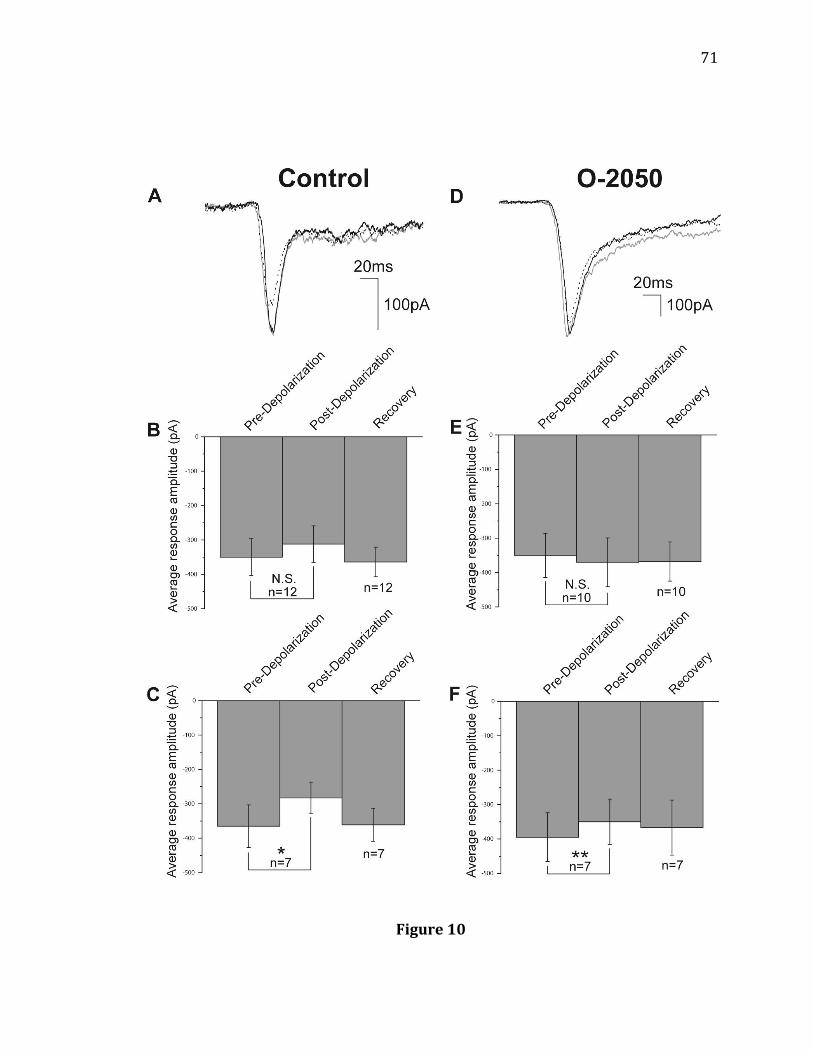
71
Figure 10

72
Figure 11. Effects of CB1R antagonist O-2050 on light-evoked Off responses of
RGCs. O-2050 increased OFF responses in one population of recorded RGCs (A, B,
and C). A: Recording traces from an OFF RGC. O-2050 increased the amplitude of
OFF responses in this cell. B: Recording traces from an ON-OFF RGC. O-2050
increased the amplitude of OFF responses in this cell. C: In one population of RGCs,
O-2050 significantly increased the amplitude of OFF responses (n= 8, 1 Off cell, 7
On-Off cells; ** p= 0.004, paired t-test). O-2050 decreased OFF responses another
population of recorded RGCs (D, E, and F). D: Recording traces from an OFF RGC. O-
2050 decreased the amplitude of Off responses in this cell. E: Recording traces from
an On-Off RGC. O-2050 decreased the amplitude of Off responses in this cell. F: In
one population of RGCs, O-2050 significantly decreased the amplitude of Off
responses (n= 28, 9 Off cells, 19 On-Off cells; *** p0.001, paired t-test).
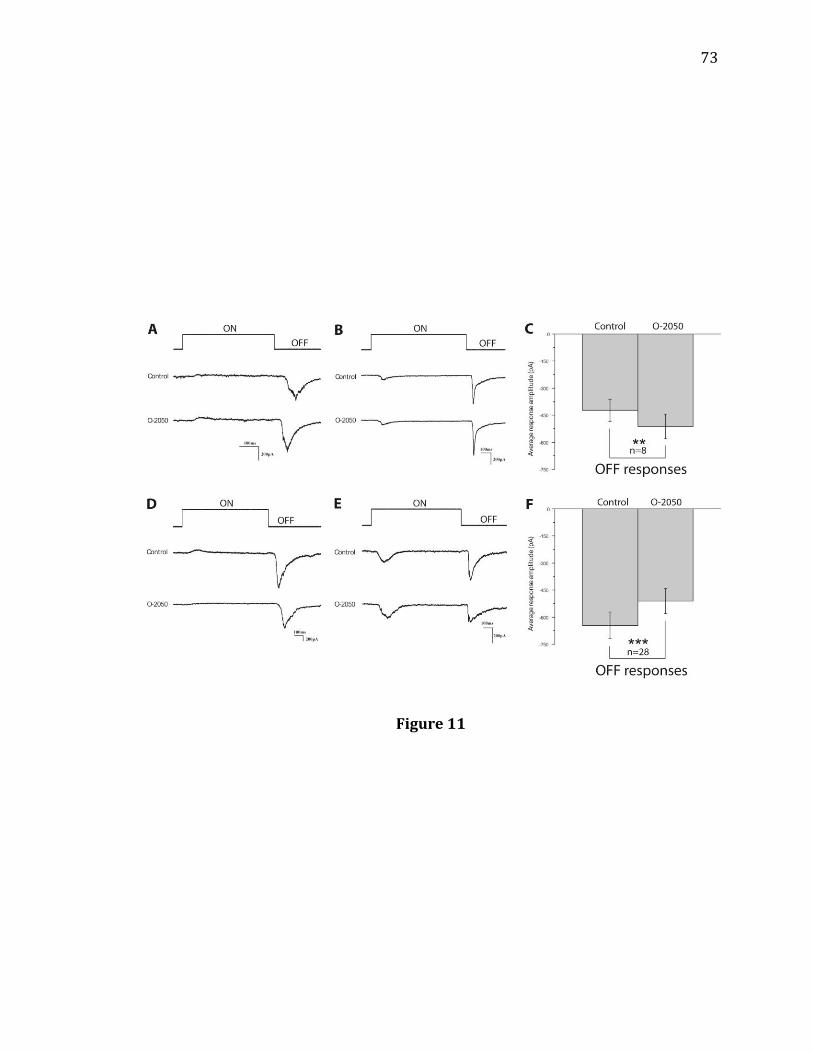
73
Figure 11

74
Figure 12. Effects of CB1R agonist WIN on light-evoked Off responses of RGCs. WIN
increased Off responses in some RGCs (A, B, and C). A: Recording traces from an Off
RGC. WIN increased the amplitude of Off responses in this cell. B: Recording traces
from an On-Off RGC. WIN increased the amplitude of Off responses in this cell. C: In
one population of recorded RGCs, WIN significantly increased the amplitude of Off
responses (n= 7, 2 Off cells, 5 On-Off cells; * p= 0.028, paired t-test). WIN decreased
Off responses in some RGCs (D, E, and F). D: Recording traces from an Off RGC. WIN
decreased the amplitude of Off responses in this cell. E: Recordings from an On-Off
RGC. WIN decreased the amplitude of Off responses in this cell. F: In one population
of recorded RGCs, WIN significantly decreased the amplitude of Off responses (n= 9,
1 Off cell, 8 On-Off cells; * p= 0.013, paired t-test).
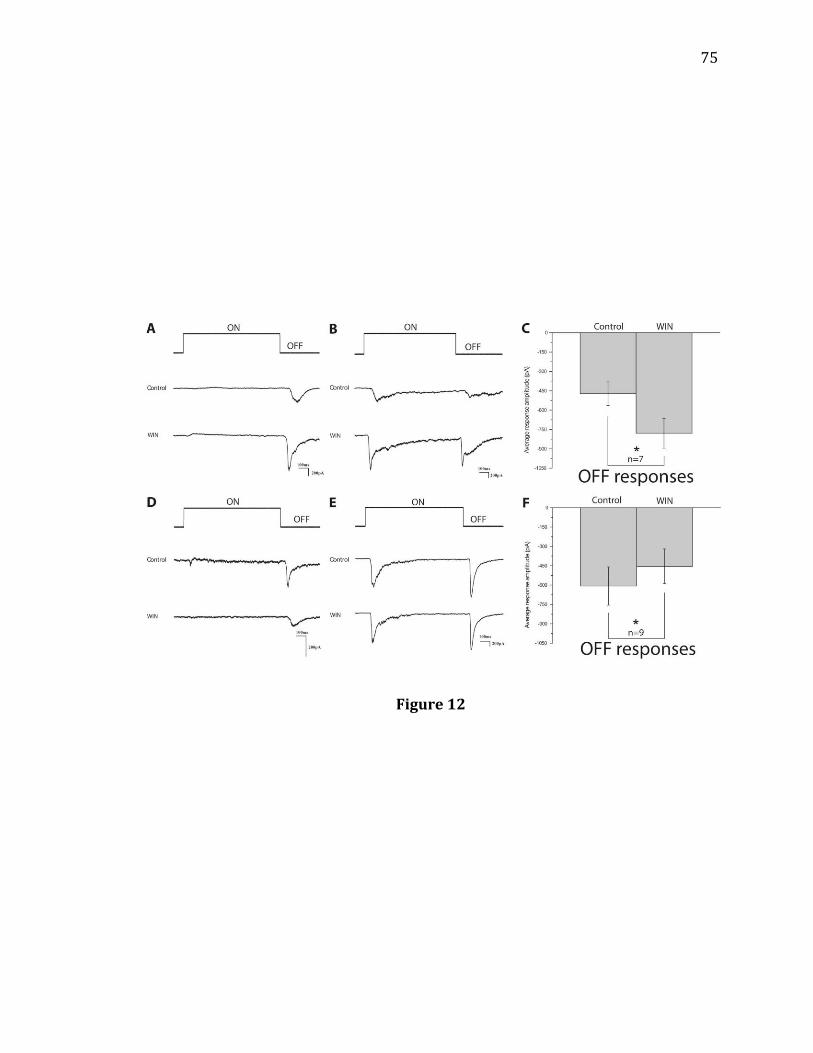
75
Figure 12

76
Figure 13. Effects of CB1R agonist 2-AG when crossover inhibition is blocked with
mGluR6 agonist APB. 2-AG increased Off responses in some RGCs even with
crossover inhibition blocked by APB (A and B). A: Recording traces from an On-Off
RGC. APB blocked the On responses and decreased the Off response amplitudes in
this cell. 2-AG increased the amplitude of Off responses in this cell. B: In one
population of recorded RGCs, 2-AG in the presence of APB increased the amplitude
of Off responses (n= 15, 9 Off cells, 6 On-Off cells; *** p0.001, paired t-test). 2-AG
decreased Off responses in some RGCs even with crossover inhibition blocked by
APB (C and D). C: Recording traces from an On-Off RGC. APB blocked the On
responses and decreased the Off response amplitudes in this cell. 2-AG decreased
the amplitude of OFF responses in this cell. D: In one population of recorded RGCs,
2-AG in the presence of APB decreased the amplitude of Off responses (n= 10, 1 Off
cell, 9 On-Off cells; * p= 0.012, paired t-test).
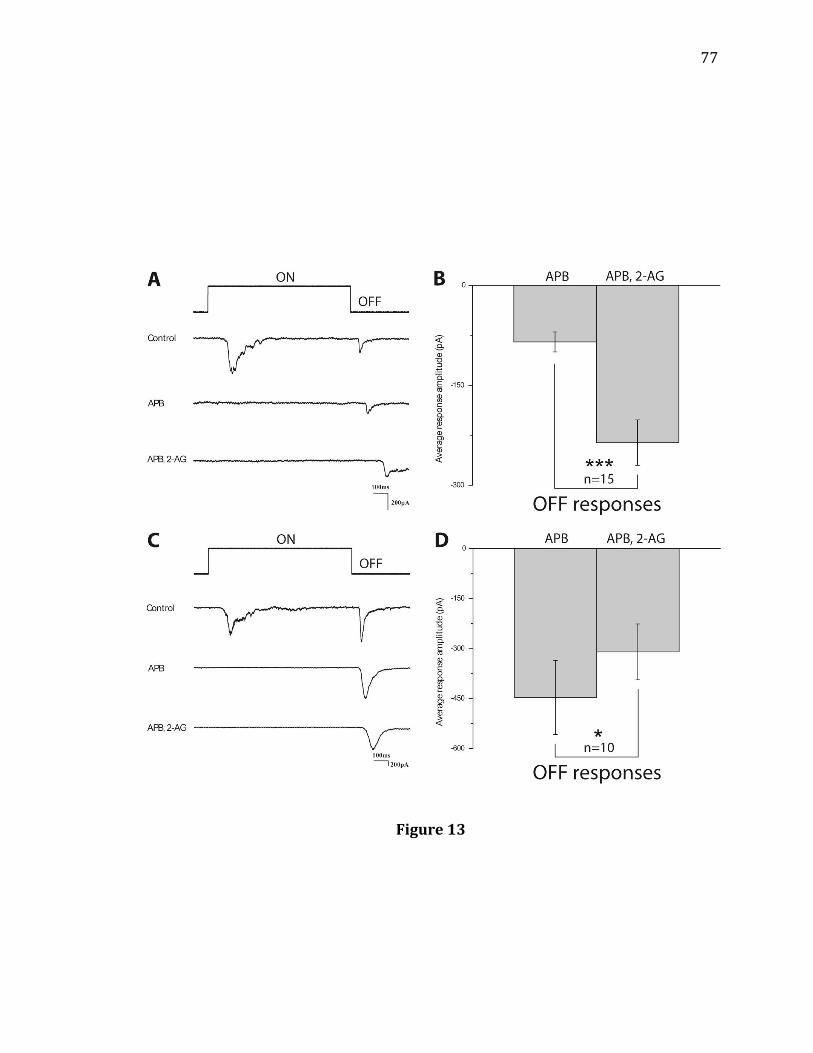
77
Figure 13

78
Figure 14. Increases in Off response amplitudes are prevented when GABAC
receptors are blocked by TPMPA. 2-AG decreased Off responses in all recorded RGCs
with crossover inhibition blocked by APB and GABAC transmission blocked by
TPMPA (A and B). A: Recording traces from an On-Off RGC. APB blocked the On
responses and decreased the Off response amplitudes in this cell. TPMPA, co-
perfused with APB, increased Off response amplitudes indicating GABAC block and
subsequent disinhibition. 2-AG, co-perfused with APB and TPMPA, decreased Off
response amplitudes in this cell. B: After application of APB and TPMPA, 2-AG
significantly decreased the amplitude of Off responses in recorded RGCs (n= 5, 2 Off
cells, 3 On-Off cells, * p= 0.015, paired t-test).
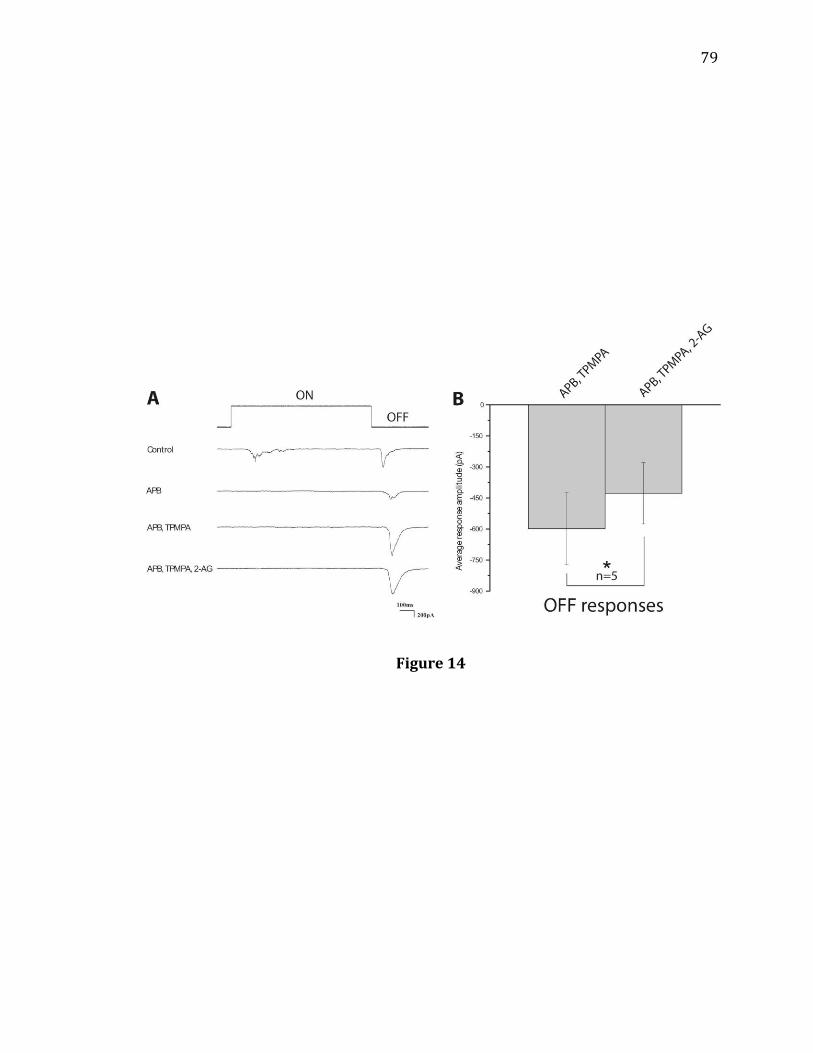
79
Figure 14

80
Figure 15. Diagram showing a functional circuit for CB1R-mediated modulation of
light-evoked Off response amplitudes: CB1R agonists activate CB1Rs at cone
terminals and/or Off-cone bipolar cell terminals to decrease glutamate release and
decrease Off amplitudes. Alternatively, CB1R agonists activate CB1Rs at GABAergic
amacrine cell terminals to decrease GABA release onto Off-cone bipolar cell
terminals that express GABAC receptors resulting in an increase in OFF amplitudes.
C: cone; OFF BC: Off-cone bipolar cell; OFF RGC: Off retinal ganglion cell; A:
GABAergic amacrine cell; GABAC: GABAC receptor.
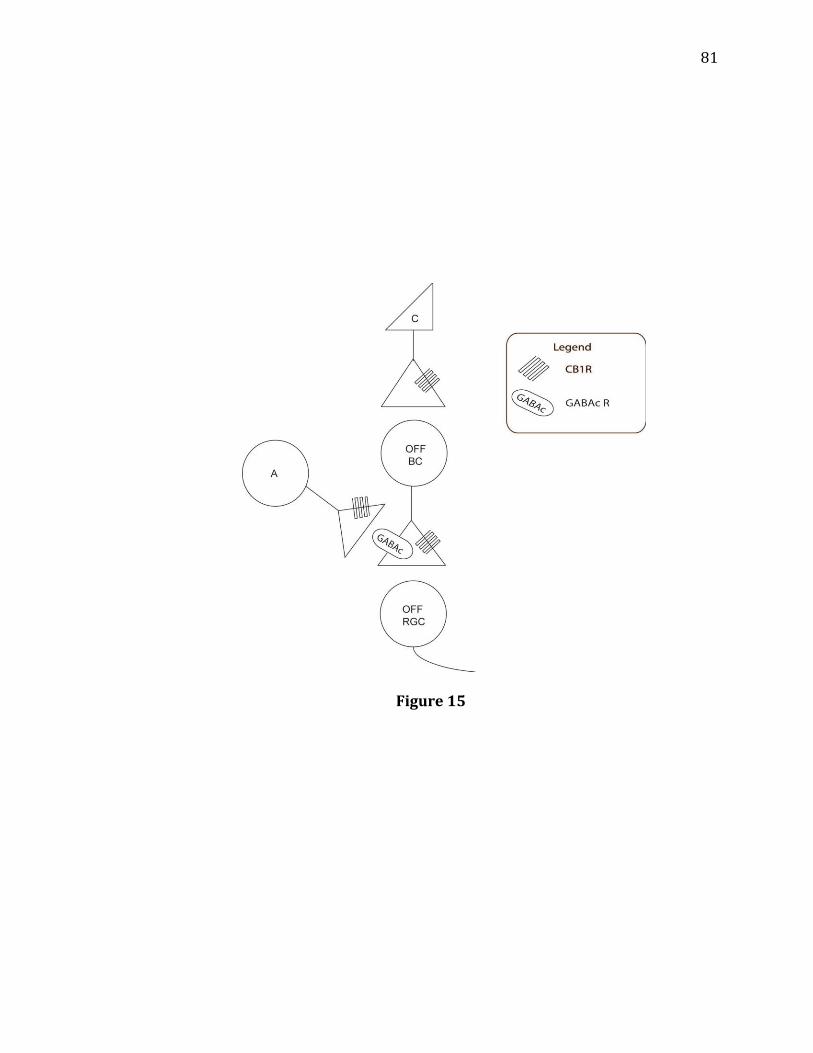
81
Figure 15

82
CHAPTER 3 GABAC receptor antagonist TPMPA modulates light-evoked Off responses in light-adapted mouse retina Introduction One way that Off pathways are modulated is by lateral inhibitory inputs from
amacrine cells (129, 130). Many GABAergic amacrine cells make synapses with the
axon terminals of Off-cone bipolar cells to provide this lateral inhibition. In the
mammalian retina, both On and Off-cone bipolar cells express GABAA and GABAC
receptors, but they do not express the metabotropic GABAB receptors (128, 131).
GABAC receptor expression is much greater than GABAA expression, however, in
both On and Off bipolar cell terminals (67, 132). Indeed, almost all GABA-evoked
current in bipolar cells is mediated by GABAC receptors (69, 122 133). The GABA-
evoked current can be parsed out experimentally using receptor-specific
antagonists. GABAA receptors can be specifically blocked with bicuculline, GABAC
receptors can be specifically blocked with TPMPA, and both GABAA and GABAC
receptors are blocked by PTX (134).
Several experiments have investigated the functional roles of GABAC
receptors. Blocking GABAC receptors with TPMPA alters bipolar cell responses to
GABA puff (135, 136). GABA-induced responses

83
became shorter following GABAC block leading researchers to hypothesize that
GABAC receptors may primarily modulate sustained glutamate release from bipolar
cells. Experimenters have also shown that GABAA versus GABAC expression differs
by bipolar cell type with important functional implications (69). In ferret, rod
bipolar cell responses were primarily mediated by GABAC receptors and were
slowest. On bipolar cell responses were slightly faster and were mediated by both
GABAA and GABAC receptors. Off bipolar cells had the fastest responses and these
responses were mostly mediated by GABAA receptors. These results indicated that
On pathways may be modulated more strongly by GABAC receptors than Off
pathways. Indeed, in the light-adapted GABAC receptor knockout mouse retina, On
responses evoked by electrical stimulation of bipolar cells were significantly
different from wild-type (10). However, Off response kinetics in GABAC-null mice
were not significantly different from wild-type. The researchers in the above study
concluded that GABAC receptors more effectively limit signaling from On than Off-
cone bipolar cells. However, the previous retinal GABAC studies have not
demonstrated that GABAC receptors modulate light-evoked responses consistently
under different ambient light conditions.
In the current study, patch clamp recordings were made from RGCs in mouse
retinas under two ambient light conditions. Under low photopic conditions, I found
that GABAC block by TPMPA increased both On and Off response decay times
recorded from On-Off RGCs. Further, I found that under high photopic conditions,
TPMPA had similar effects to those observed in dim light. Blocking crossover
inhibition with APB did not prevent TPMPA-induced increases in Off decay time.

84
The results presented in this study suggest that GABAC receptors shape both On and
Off responses in the light-adapted mouse retina under both low and high photopic
ambient light conditions.

85
Materials and Methods
The basic methods used in this study were similar to those used previously in
the laboratory (81, 82, 83, 84). The materials and methods used for the experiments
outlined in this Chapter are also similar to those described in Chapters 1 and 2.
Retinal preparation. Retinal preparations performed in these experiments are
similar to those described in Chapters 1 and 2.
Electrophysiology. Whole-cell patch clamp recordings were made from RGCs in light-
adapted retinas, as described in Chapters 1 and 2. Measurements for light-evoked
EPSCs are detailed in Chapter 1. Three light-evoked responses were averaged under
control conditions and also under each different drug condition.
Light stimulus. The light stimuli were presented to the light-adapted retina using the
Lucivid, as described in Chapter 1. For these studies, each RGC was dissected and
prepared with a background light intensity of 3000 photons/m2/s (low photopic).
Control responses were recorded at the above background light intensity. After
control responses were recorded, drugs were bath applied and the retina was
adapted to a background light intensity of 2.9x104 photons/m2/s (high-photopic).
At both of these background light intensities, rods are completely inactivated and
only cone-mediated responses are recorded.
Light stimuli with intensities greater than that of the background light were
used to evoke light responses from RGCs. For each cell, different intensities, ranging
from 3.2x103 photons/m2/s to 1.6x104 photons/m2/s under the low photopic

86
conditions and between 6.25x104 photons/m2/s and 6.7x106 photons/m2/s
under the high photopic conditions were used to evoke light responses.
In this study, we investigated the role of TPMPA in modulating light-evoked
response kinetics. The time for light-evoked EPSCs to decay to 10% of the peak
amplitude was measured with Minianalysis (Synaptosoft, Inc.). EPSC traces were
visualized using Clampfit 9 (Molecular Devices, Inc.) and averaged within drug
conditions using Origin 6.0 (OriginLab Corporation). Averaged traces before and
after TPMPA application were normalized by amplitude and overlaid to illustrate
differences in light response kinetics.
Drug application. APB (20 M; Sigma) and (1,2,5,6-tetrahydropyridin-4-
yl)methylphosphinic acid (TPMPA; 50 M; Sigma) were freshly dissolved in MEME
on the day of the experiment and administered through a gravity-fed line.

87
Results
TPMPA increased the decay time of both On and Off responses under low photopic
background illumination
In a previous study performed under one background light intensity, GABAC
receptor antagonist TPMPA significantly increased On but not Off response decay
time (10). Moreover, On responses in GABAC knockout mice strongly differed from
On responses in wild-type, but Off responses in GABAC knockout mice were no
different from Off responses in wild-type. Responses were stimulated by electrical
tetanus localized to the OPL where bipolar cell dendrites stratify. To determine if
TPMPA selectively increases light-evoked On response decay time under dim light
conditions, we recorded light-evoked responses from On-Off RGCs under a low
photopic ambient light intensity of 3000 photons/m2/s before and after bath
application of 50 M TPMPA (Figure 16). With GABAC receptors blocked by TPMPA,
light-evoked On response average time to decay to 10% of peak amplitude
significantly increased (Figure 16A,B, n= 12). The average On decay time increased
62%. However, Off response decay times also significantly increased (Figure 16C,D,
n= 12). The average Off decay time increased 82%. These results indicate that under
low photopic conditions, TPMPA alters both light-evoked On and light-evoked Off
kinetics.
TPMPA increased the decay time of both On and Off responses under high photopic
background illumination

88
As was discussed in Chapter 1, lateral inhibition of Off pathways changes
depending upon the intensity of background illumination. In the previous study
demonstrating the effects of TPMPA in RGC responses, experiments were only
performed at one background light intensity (10). To determine if TPMPA increases
light-evoked On and Off response decay time consistently at different ambient light
intensities, we increased the background light intensity from 3000 photons/m2/s
to a high photopic intensity of 2.9x104 photons/m2/s and recorded light-evoked
responses before and after bath application of 50 M TPMPA (Figure 17). The same
cells were used for both the low photopic and high photopic conditions. After the
effects of TPMPA were recorded in the low photopic condition, washout of TPMPA
proceeded for 10 minutes. At the same time, the retina was illuminated with and
adapted to the high photopic ambient light intensity. Under high photopic
conditions, GABAC receptor block by TPMPA significantly increased On response
average time to decay to 10% of peak amplitude (Figure 17A,B, n= 11). The On
response decay time increased 67%. Moreover, Off response average time to decay
to 10% of peak amplitude under photopic conditions also significantly increased
(Figure 17C,D, n= 11). TPMPA increased the Off response decay time 85%. These
results indicate that under high photopic conditions, TPMPA alters both On and Off
response kinetics.
Blocking crossover inhibition with mGluR6 agonist APB does not prevent the TPMPA-
induced increase in Off response decay time

89
Researchers found that only On response kinetics were significantly altered
in GABAC knockout mice or after application of TPMPA (10). However, we found that
both On and Off decay times significantly increased following bath application of
TPMPA. In our experiment, we used light to evoked RGC responses. As we
demonstrated in Chapter 1, light-evoked Off responses can be modulated by
crossover inhibition from On pathways. To determine if TPMPA modulates
crossover inhibition to increase Off response decay times, we bath applied APB (20
M) to block crossover inhibition and then recorded light-evoked responses from
On-Off RGCs before and after application of 50 M TPMPA (Figure 18). Even with
crossover inhibition blocked by APB, Off response average time to decay to 10% of
peak amplitude significantly increased in TPMPA. Indeed, Off response decay times
increased 64%. These results indicate that a reduction in crossover inhibition does
not underlie the TPMPA-induced increases in Off response decay time.

90
Discussion
Both On and Off-cone bipolar cells express GABAC receptors at their axon
terminals (137, 138, 139, 140). GABAergic amacrine cells release GABA that
activates the chloride-fluxing GABAC receptors at bipolar cell terminals (128). This
results in inhibition of the bipolar cell and reduces subsequent release of glutamate
from bipolar cells onto RGCs. GABAergic transmission in the IPL, like that described
above, plays an important role in shaping the surround inhibition and temporal
responses in RGCs (135, 141). Thus, GABAC mediated transmission is crucial to
important visual properties like edge detection, motion sensitivity, and direction
sensitivity.
TPMPA can be used to specifically block GABAC receptors (65). When GABAC
receptors are knocked out or blocked by TPMPA, GABAR-mediated chloride
currents become more transient, with a fast rise and decay time, because of the
elimination of a longer-latency chloride current (10). This alters neurotransmitter
release from bipolar cells and alters kinetics of EPSCs recorded from RGCs. While
GABAC receptors are expressed at both On and Off-cone bipolar cell axon terminals,
a previous study using a ferret model demonstrated that GABA-induced currents in
bipolar cells differed depending on cell type (69). GABA-induced currents in ferret
On bipolar cells were mostly mediated by GABAC receptors, whereas GABA-induced
currents in ferret Off bipolar cells were mostly mediated by GABAA receptors and
were transient. Moreover, in GABAC knockout mice, On responses recorded from
RGCs were significantly different from wild-type, but Off responses were not
significantly different from wild-type (10). Thus, the previous studies indicate that

91
GABAC-mediated inhibition plays a larger role in modulating On responses than Off
responses. However, whether these previous findings are consistent under different
background light intensities is not known. Further, it is not yet clear whether the
results from previous studies will be consistent with results of experiments where
TPMPA is used to block GABAC receptors and light is used to evoke responses. The
results of the present study demonstrate that TPMPA increases both On and Off
response decay times under dim ambient light conditions. Similar results were
found when the background light intensity was increased. Crossover inhibition does
not underlie the TPMPA-induced increase in Off decay times. Taken together, these
findings provide evidence that under two background light intensities, GABAC block
by TPMPA alters both On and Off response kinetics.
TPMPA increases both On and Off response decay time under low photopic background illumination Under a low photopic background light intensity of 3000 photons/m2/s,
TPMPA altered both On and Off response amplitude and kinetics. By blocking GABAC
receptors with TPMPA, both On and Off pathways were disinhibited. This facilitated
increased glutamate release from bipolar cells onto the recorded RGCs. In addition,
the temporal properties of both On and Off responses changed. On EPSCs had a
much longer time to decay. Surprisingly, Off EPSC decay times also increased,
contrary to the findings in previous studies (10). Indeed, TPMPA increased Off decay
times to a greater extent than On decay times. In the earlier study, RGC responses
before and after application of TPMPA were evoked with an electrical stimulus. It is

92
possible that the differences between the previous study and the current study are
due to differences in stimulation methods. However, light-evoked responses were
also used in the aforementioned previous study to determine the differences in both
On and Off kinetics between wild-type and GABAC knockout mice. In that study,
GABAC knockout mice had significantly different On but not Off responses compared
to wild-type. Currently it is unclear why Off responses were affected by TPMPA in
the present study, but not in the previous study.
TPMPA increases both On and Off response decay time under high photopic background illumination Given the surprising result of TPMPA-induced differences in Off response
kinetics, we repeated the above experiment at the higher background light intensity
of 2.9x104 photons/m2/s. From our results in Chapter 1, in addition to results from
other studies, we hypothesized that the relative strength of inhibition may change at
different ambient light intensities. Therefore, we adapted the retina to a high
photopic, but still light-adapted, ambient illumination. In the present study, we
showed that under high photopic background illumination, TPMPA still increased
both On and Off response amplitude and decay times. In fact, the factors by which
TPMPA increased both On and Off decay times were consistent across the two
background illumination levels. These results suggest that our surprising finding
that TPMPA altered Off response kinetics was not due to differences in ambient light
intensity compared to the previous study.

93
Blocking crossover inhibition with APB does not prevent TPMPA-induced increases in Off response decay time In the previous study, TPMPA only increased On response decay times (10).
These On responses were evoked electrically with the stimulus localized to the OPL
where bipolar cell dendrites stratify. Given the results from the previous study, we
had not expected that TPMPA would increase Off response decay times. However,
we used light to evoke the stimulus rather than an electric tetanus. It is possible that
differences in the stimulus methods resulted in different circuits being activated.
For instance, light-evoked Off responses are known to be modulated by crossover
inhibition from On pathways. Crossover inhibition of Off pathways could potentially
be inactive when using an electrical stimulus, and this could underlie the differences
found between the previous and the current study. Therefore, we blocked crossover
inhibition with APB and recorded light-evoked responses before and after
application of TPMPA. Even with crossover inhibition blocked by APB, TPMPA still
increased Off response decay times. These results indicate that modulation of
crossover inhibition does not underlie the TPMPA-induced increases in Off response
decay times.
Taken together, the results of this study and those of previous studies
indicate that GABAC receptors play a crucial role in shaping the kinetics of light
responses in RGCs. Studies showing GABAC expression at Off-cone bipolar cell
terminals, as well as our present study, suggest that both On and Off responses are
modulated by GABAC receptors (Figure 19). It is currently unclear why the results of
our study differed from the results in 10. Future experiments should be conducted
to determine the basis for the inconsistency.

94
Figure 16. GABAC antagonist TPMPA increases the decay time of On and Off
responses in On-Off RGCs under low photopic conditions. Recordings were made
from RGCs with a background light of 3000 photons/m2/s and a stimulus light
intensity of 1.6x104 photons/m2/s. TPMPA increased the On decay time in On-Off
RGCs (A and B). A: Overlaid On responses normalized by amplitude from a single
On-Off RGC before (black trace) and after bath application of TPMPA (gray trace). B:
TPMPA significantly increased the average time for the On response to decay to
10% of the peak On amplitude (n= 12, 12 On-Off cells, * p= 0.010, paired t-test).
TPMPA increased the Off decay time in On-Off RGCs (C and D). C: Overlaid Off
responses normalized by amplitude from the same On-Off RGC as A before (black
trace) and after bath application of TPMPA (gray trace). D: TPMPA significantly
increased the average time for the Off response to decay to 10% of the peak Off
amplitude (n= 12, 12 On-Off cells, *** p< .001, paired t-test).
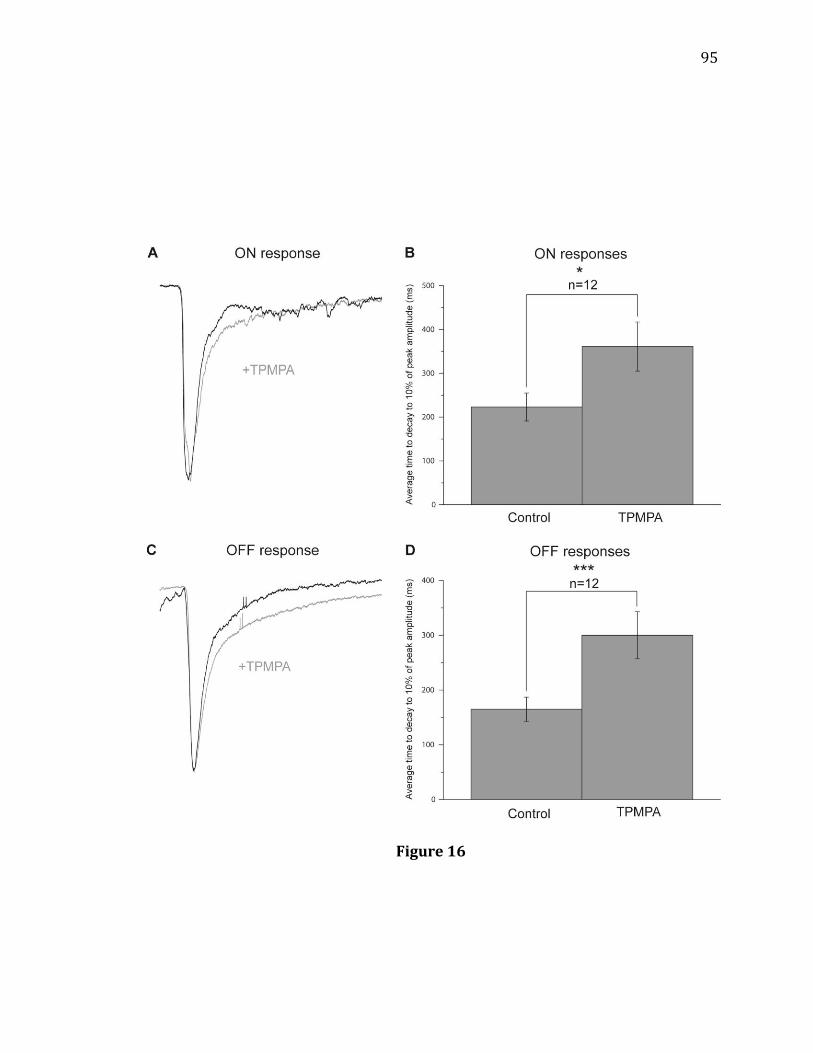
95
Figure 16

96
Figure 17. GABAC antagonist TPMPA increases the decay time of On and Off
responses in On-Off RGCs under high photopic conditions. Recordings were made
from RGCs with a background light of 2.9x104 photons/m2/s and a stimulus light
intensity of 6.7x106 photons/m2/s. TPMPA increased the On decay time in On-Off
RGCs (A and B). A: Overlaid On responses normalized by amplitude from a single
On-Off RGC before (black trace) and after bath application of TPMPA (gray trace). B:
TPMPA significantly increased the average time for the On response to decay to
10% of the peak On amplitude (n= 11, 11 On-Off cells, * p= 0.014, paired t-test).
TPMPA increased the Off decay time in On-Off RGCs (C and D). C: Overlaid Off
responses normalized by amplitude from the same On-Off RGC as A before (black
trace) and after bath application of TPMPA (gray trace). D: TPMPA significantly
increased the average time for the Off response to decay to 10% of the peak Off
amplitude (n= 11, 11 On-Off cells, *** p< .001, paired t-test).

97
Figure 17

98
Figure 18. Blocking crossover inhibition with mGluR6 agonist APB does not prevent
the TPMPA-induced increase in Off response decay time under high photopic
conditions. TPMPA increases Off response decay time recorded from On-Off RGCs
with crossover inhibition blocked by APB (A and B). A: Overlaid Off responses
normalized by amplitude from an On-Off RGC during bath application of APB (black
trace) and bath application of APB plus TPMPA (gray trace). B: TPMPA significantly
increased the average time for the Off response to decay to 10% of peak Off
amplitude even with crossover inhibition blocked by APB (n= 11, 11 On-Off cells, p=
0.015, paired t-test).

99
Figure 18

100
Figure 19. Diagram showing a functional circuit for GABAC receptor-mediated
modulation of Off responses. GABAergic amacrine cells release GABA onto Off-cone
bipolar cells. The Off-cone bipolar cells, which express GABAC receptors at the axon
terminals become inhibited and subsequent glutamate release from the Off-cone
bipolar cell is reduced. Ultimately, Off responses recorded from Off RGCs are
reduced. TPMPA blocks this circuit by blocking GABAC receptors and preventing
GABA from inhibiting the Off-cone bipolar cells. C: cone; OFF BC: Off-cone bipolar
cell; OFF RGC: Off retinal ganglion cell; A: GABAergic amacrine cell; GABAC: GABAC
receptor.

101
Figure 19

102
CONCLUSION
Neural signals in the retina are carried by parallel pathways dedicated to
light increments and light decrements (1). Having parallel circuits doubles the
retina’s dynamic range, increases signal transfer efficiency, and facilitates contrast
sensitivity. Loss of function of either pathway severely impairs vision (3). Therefore,
it is crucial to deeply understand both the circuit for light increments, known as the
On pathway, and the circuit for light decrements, known as the Off pathway. In this
thesis, modulation of Off responses by mGluR6, CB1, and GABAC receptors were
studied using patch clamp electrophysiology of RGCs and pharmacological
manipulations. The experiments I have described in this thesis demonstrate how
several neurotransmitter receptors shape the responses recorded from RGCs, the
final output neurons of the retina.
Even though On and Off pathways are parallel and have opposite functions,
they are not completely isolated from one other. In previous experiments performed
in the dark-adapted retina, it has been shown that On pathways can inhibit Off
pathways via crossover inhibition (31, 32, 33, 34). When On-cone bipolar cells
become excitable they release glutamate that activates glycinergic amacrine cells.
These amacrine cells then release glycine onto Off-cone bipolar cells and inhibit Off
responses. Blocking

103
crossover inhibition with mGluR6 agonist APB disinhibits the Off-cone bipolar cells
and increases the amplitude of RGC Off responses recorded under dark adaptation
(5, 6, 7, 8). The results from the experiments detailed in Chapter 1 confirm that APB
increases Off response amplitudes under dark adaptation. However, we found that
APB decreases Off response amplitudes under light adaptation. This suggests that
the impact of crossover inhibition is dynamic and depends on the ambient light
intensity. APB-induced increases in Off response amplitude under dark adaptation
required glycinergic transmission. We found that the APB-induced decreases in Off
response amplitude under light adaptation, however, did not involve inhibitory
neurotransmission because the effect persisted even with STR, PTX, or TPMPA
included in the bath solution. This indicated that On pathways may modulate Off
pathways using a novel mechanism that does not involve crossover inhibition. When
we blocked D1 receptors, APB-induced decreases in Off responses under light
adaptation were blocked. This suggested that dopaminergic transmission involving
D1 receptors is necessary for the observed effects of APB under light adaptation.
Finally, blocking HCN channels also prevented APB-induced decreases in Off
responses under light adaptation. Taken together these results indicate a novel way
in which On pathways can modulate Off responses. Under dark adaptation, On
pathways inhibit Off pathways via glycinergic amacrine cells. Under light adaptation,
this crossover inhibition pathway is still active but is occluded by a stronger and
opposite crosstalk mechanism: 1) On-cone bipolar cells excite dopaminergic
amacrine cells that are only active during light adaptation, 2) dopamine is released
from these amacrine cells and binds to D1 receptors expressed by Off-cone bipolar

104
cells, 3) activation of the metabotropic D1 receptors results in activation of HCN
channels , and 4) HCN channel activation increases glutamate release from Off-cone
bipolar cells resulting in greater Off EPSCs in RGCs. APB blocks the above
mechanism by hyperpolarizing On-cone bipolar cells resulting in a decrease in Off
response amplitudes. Our findings in dark and light adaptation suggest that mGluR6
receptors, despite being localized to On pathways, are capable of modulating Off
responses. Moreover, mGluR6-mediated modulation of Off responses is dynamic
and the mechanisms involved are dependent on the retinal adaptation.
The retinal eCB system is a relatively new field of study. CB1R localization
and functions in the retina have been investigated, however, the role of CB1Rs in
shaping light-evoked output from the retina is not yet clear (9). The results
presented in Chapter 2 support our hypothesis that the retinal eCB system does
modulate Off response output from the light-adapted retina. We found that DSE, an
extensively studied mechanism of eCB function in the brain, also occurs at some
synapses between Off-cone bipolar cells and RGCs. The DSE may not be completely
mediated by CB1Rs, however. In addition, we found that there are functionally two
populations of RGCs. In the first population, CB1R antagonists increase Off
responses and CB1R agonists decrease Off responses. We suggest that the
antagonists increase glutamate release from cones and bipolar cells resulting in a
larger Off response. The agonists decrease glutamate release at the same cells
resulting in a smaller Off response. In the second population, CB1R antagonists
decrease Off responses and CB1R agonists increase Off responses. Our results
indicate that the antagonists decrease Off responses by increasing GABA release

105
from GABAergic amacrine cells that provide inputs to Off-cone bipolar cells.
Consistent with this, our results suggest that agonists increase Off responses by
decreasing GABA release from GABAergic amacrine cells that provide inputs to Off-
cone bipolar cells. Taken together, the results from this study indicate that the
retinal eCB system does modulate signal output from the retina and likely has an
impact on vision.
As previous studies have shown, including our own in Chapter 1, lateral
inhibition modulates Off pathways (129, 130). Lateral inhibition from GABAergic
amacrine cells to bipolar cells is mainly mediated by GABAC receptors (134).
However, in a previous study, even though both On and Off-cone bipolar cells
express GABAC receptors, knocking out or blocking GABAC receptors only affected
On but not Off responses in RGCs under light adaptation (10). The results from the
experiments detailed in Chapter 3 indicate that both On and Off responses are
affected by TPMPA, a GABAC specific antagonist. Contrary to the results in the
previous study, TPMPA increased both On and Off response decay times as would be
expected if GABAC receptors, with their long-latency and sustained effects, were
blocked. These results were consistent at two levels of ambient light intensity and
with crossover inhibition blocked by APB. It remains unclear why our results differ
from those of the previous study. Taken together, the results presented in this study
indicate that GABAC receptors play an important role in modulating the strength and
temporal properties of Off responses.
In conclusion, the data contained in this thesis indicate that mGluR6, CB1,
and GABAC receptors are all capable of modulating Off response output from the

106
retina. Collectively, the results in this thesis suggest several methods in which Off
pathways can be modulated. First, we showed that crosstalk between On and Off
pathways modulates Off responses under dark and light adaptation. Second, we
demonstrated that retrograde neurotransmitter systems affect Off response
strength. Finally, we illustrated how lateral inhibition shapes Off responses. The
mechanisms described in this thesis illustrate both the complexity and the
importance of the retina with regard to visual signal processing.

107
LIST OF ABBREVIATIONS
2-AG Endocannabinoid and CB1R agonist 2-arachidonoyl glycerol APB mGluR6 receptor agonist DL-2-amino-4-phosphonobutyric acid CB1R Cannabinoid 1 receptor DCPG mGluR8 agonist (S)-3,4-dicarboxyphenylglycine DSE Depolarization-induced suppression of excitation DSI Depolarization-induced suppression of inhibition ECl Chloride equilibrium potential eCB Endocannabinoid, an endogenous cannabinoid EPSC Excitatory post-synaptic current GABA Inhibitory neurotransmitter -aminobutyric acid GCL Ganglion cell layer HCN Hyperpolarization-activated cyclic nucleotide-gated channel INL Inner nuclear layer IPL Inner plexiform layer IPSC Inhibitory post-synaptic current MEME Bath solution used in experiments, minimum essential medium Eagle mGluR6 Metabotropic glutamate receptor 6 ONL Outer nuclear layer OPL Outer plexiform layer

108
PTX GABAA/C receptor antagonist picrotoxin RGC Retinal ganglion cell SE Standard error STR Glycine receptor antagonist strychnine TPMPA GABAC receptor specific antagonist (1,2,5,6-tetrahydropyridin-4-
yl)methylphosphinic acid WIN Synthetic CB1R agonist WIN 55, 212-2

109
LIST OF REFERENCES
1. Sterling P, Demb J (2004) Retina. In: Synaptic Organization of the Brain, 5th Edition (Shepherd G, ed), pp 163-169. New York: Oxford University Press.
2. Sharpe LT, Stockman A (1999) Rod pathways: the importance of seeing nothing.
Trends Neurosci 22:497-504. 3. Schiller PH, Sandell JH, Maunsell JH (1986) Functions of the ON and OFF channels
of the visual system. Nature 322:824-825. 4. Schiller PH (1992) The ON and OFF channels of the visual system. Trends
Neurosci 15:86-92. 5. Massey SC, Redburn DA, Crawford ML (1983) The effects of 2-amino-4-
phosphonobutyric acid (APB) on the ERG and ganglion cell discharge of rabbit retina. Vision Res 23:1607-1613.
6. Jin XT, Brunken WJ (1996) A differential effect of APB on ON- and OFF-center
ganglion cells in the dark adapted rabbit retina. Brain Res 708:191-196. 7. Higgs MH, Romano C, Lukasiewicz PD (2002) Presynaptic effects of group III
metabotropic glutamate receptors on excitatory synaptic transmission in the retina. Neuroscience 115:163-172.
8. Popova E, Mitova L, Vitanova L, Kupenova P (2003) Participation of the
GABAergic system in the action of 2-amino-4-phosphonobutyrate on the OFF responses of frog retinal ganglion cells. Vision Res 43:607-616.
9. Yazulla S (2008) Endocannabinoids in the retina: from marijuana to
neuroprotection. Prog Retin Eye Res 27:501-526. 10. Sagdullaev BT, McCall MA, Lukasiewicz PD (2006) Presynaptic inhibition
modulates spillover, creating distinct dynamic response ranges of sensory output. Neuron 50:923-935.
11. Alamouti B, Funk J (2003) Retinal thickness decreases with age: an OCT study.
Br J Ophthalmol 87:899-901.

110
12. Masland RH (2012) The neuronal organization of the retina. Neuron 76:266-280. 13. Yau KW (1994) Phototransduction mechanism in retinal rods and cones. The
Friedenwald Lecture. Invest Ophthalmol Vis Sci 35:9-32. 14. Bloomfield SA, Dacheux RF (2001) Rod vision: pathways and processing in the
mammalian retina. Prog Retin Eye Res 20:351-384. 15. Weiler R, Pottek M, He S, Vaney DI (2000) Modulation of coupling between
retinal horizontal cells by retinoic acid and endogenous dopamine. Brain Res Brain Res Rev 32:121-129.
16. Kolb H, Nelson R, Mariani A (1981) Amacrine cells, bipolar cells and ganglion
cells of the cat retina: a Golgi study. Vision Res 21:1081-1114. 17. MacNeil MA, Masland RH (1998) Extreme diversity among amacrine cells:
implications for function. Neuron 20:971-982. 18. Cook PB, McReynolds JS (1998b) Modulation of sustained and transient lateral
inhibitory mechanisms in the mudpuppy retina during light adaptation. J Neurophysiol 79:197-204.
19. Famiglietti EV, Jr., Kolb H (1976) Structural basis for ON-and OFF-center
responses in retinal ganglion cells. Science 194:193-195. 20. Nelson R, Famiglietti EV, Jr., Kolb H (1978) Intracellular staining reveals
different levels of stratification for on- and off-center ganglion cells in cat retina. J Neurophysiol 41:472-483.
21. Volgyi B, Deans MR, Paul DL, Bloomfield SA (2004) Convergence and
segregation of the multiple rod pathways in mammalian retina. J Neurosci 24:11182-11192.
22. Purkiss TJ, Hughes A, DeMarco PJ, Jr. (2001) Processing of scotopic increments
and decrements. Vis Neurosci 18:119-125. 23. Protti DA, Flores-Herr N, Li W, Massey SC, Wassle H (2005) Light signaling in
scotopic conditions in the rabbit, mouse and rat retina: a physiological and anatomical study. J Neurophysiol 93:3479-3488.
24. Weinstein GW, Hobson RR, Baker FH (1971) Extracellular recordings from
human retinal ganglion cells. Science 171:1021-1022. 25. Shiells RA, Falk G, Naghshineh S (1981) Action of glutamate and aspartate
analogues on rod horizontal and bipolar cells. Nature 294:592-594.

111
26. Nakanishi S, Nakajima Y, Masu M, Ueda Y, Nakahara K, Watanabe D, Yamaguchi S, Kawabata S, Okada M (1998) Glutamate receptors: brain function and signal transduction. Brain Res Brain Res Rev 26:230-235.
27. Morgans CW, Zhang J, Jeffrey BG, Nelson SM, Burke NS, Duvoisin RM, Brown RL
(2009) TRPM1 is required for the depolarizing light response in retinal ON-bipolar cells. Proc Natl Acad Sci U S A 106:19174-19178.
28. Sasaki T, Kaneko A (1996) L-Glutamate-induced responses in OFF-type bipolar
cells of the cat retina. Vision Res 36:787-795. 29. DeVries SH, Schwartz EA (1999) Kainate receptors mediate synaptic
transmission between cones and 'Off' bipolar cells in a mammalian retina. Nature 397:157-160.
30. DeVries SH (2000) Bipolar cells use kainate and AMPA receptors to filter visual
information into separate channels. In: Neuron, pp 847-856. United States. 31. Menger N, Pow DV, Wassle H (1998) Glycinergic amacrine cells of the rat retina.
J Comp Neurol 401:34-46. 32. O'Brien BJ, Richardson RC, Berson DM (2003) Inhibitory network properties
shaping the light evoked responses of cat alpha retinal ganglion cells. Vis Neurosci 20:351-361.
33. Molnar A, Hsueh HA, Roska B, Werblin FS (2009) Crossover inhibition in the
retina: circuitry that compensates for nonlinear rectifying synaptic transmission. J Comput Neurosci 27:569-590.
34. van Wyk M, Wassle H, Taylor WR (2009) Receptive field properties of ON- and
OFF-ganglion cells in the mouse retina. Vis Neurosci 26:297-308. 35. Molnar A, Werblin F (2007) Inhibitory feedback shapes bipolar cell responses in
the rabbit retina. J Neurophysiol 98:3423-3435. 36. Hsueh HA, Molnar A, Werblin FS (2008) Amacrine-to-amacrine cell inhibition in
the rabbit retina. J Neurophysiol 100:2077-2088. 37. Manookin MB, Beaudoin DL, Ernst ZR, Flagel LJ, Demb JB (2008) Disinhibition
combines with excitation to extend the operating range of the OFF visual pathway in daylight. J Neurosci 28:4136-4150.
38. Slaughter MM, Miller RF (1981) 2-amino-4-phosphonobutyric acid: a new
pharmacological tool for retina research. Science 211:182-185. 39. West ME (1991) Cannabis and night vision. Nature 351:703-704.

112
40. Russo EB, Merzouki A, Mesa JM, Frey KA, Bach PJ (2004) Cannabis improves night vision: a case study of dark adaptometry and scotopic sensitivity in kif smokers of the Rif mountains of northern Morocco. In: J Ethnopharmacol, pp 99-104. Ireland.
41. Dawson WW, Jimenez-Antillon CF, Perez JM, Zeskind JA (1977) Marijuana and
vision--after ten years' use in Costa Rica. Invest Ophthalmol Vis Sci 16:689-699.
42. Straiker A, Stella N, Piomelli D, Mackie K, Karten HJ, Maguire G (1999)
Cannabinoid CB1 receptors and ligands in vertebrate retina: localization and function of an endogenous signaling system. Proc Natl Acad Sci U S A 96:14565-14570.
43. Yazulla S, Studholme KM, McIntosh HH, Deutsch DG (1999)
Immunocytochemical localization of cannabinoid CB1 receptor and fatty acid amide hydrolase in rat retina. J Comp Neurol 415:80-90.
44. Yazulla S, Studholme KM, McIntosh HH, Fan SF (2000) Cannabinoid receptors on
goldfish retinal bipolar cells: electron-microscope immunocytochemistry and whole-cell recordings. Vis Neurosci 17:391-401.
45. Diana MA, Marty A (2004) Endocannabinoid-mediated short-term synaptic
plasticity: depolarization-induced suppression of inhibition (DSI) and depolarization-induced suppression of excitation (DSE). Br J Pharmacol 142:9-19.
46. Ohno-Shosaku T, Hashimotodani Y, Maejima T, Kano M (2005) Calcium
signaling and synaptic modulation: regulation of endocannabinoid-mediated synaptic modulation by calcium. Cell Calcium 38:369-374.
47. Fan SF, Yazulla S (2007) Retrograde endocannabinoid inhibition of goldfish
retinal cones is mediated by 2-arachidonoyl glycerol. Vis Neurosci 24:257-267.
48. Fan SF, Yazulla S (2003) Biphasic modulation of voltage-dependent currents of
retinal cones by cannabinoid CB1 receptor agonist WIN 55212-2. Vis Neurosci 20:177-188.
49. Straiker A, Sullivan JM (2003) Cannabinoid receptor activation differentially
modulates ion channels in photoreceptors of the tiger salamander. In: J Neurophysiol, pp 2647-2654. United States.
50. Fan SF, Yazulla S (2004) Inhibitory interaction of cannabinoid CB1 receptor and
dopamine D2 receptor agonists on voltage-gated currents of goldfish cones. Vis Neurosci 21:69-77.

113
51. Fan SF, Yazulla S (2005) Reciprocal inhibition of voltage-gated potassium currents (I K(V)) by activation of cannabinoid CB1 and dopamine D1 receptors in ON bipolar cells of goldfish retina. Vis Neurosci 22:55-63.
52. Lalonde MR, Jollimore CA, Stevens K, Barnes S, Kelly ME (2006) Cannabinoid
receptor-mediated inhibition of calcium signaling in rat retinal ganglion cells. Mol Vis 12:1160-1166.
53. Glass M FC (1997) Concurrent stimulation of cannabinoid CB1 and dopa... [J
Neurosci. 1997] - PubMed result. In: Journal of Neuroscience. 54. Maneuf YP, Brotchie JM (1997) Paradoxical action of the cannabinoid WIN
55,212-2 in stimulated and basal cyclic AMP accumulation in rat globus pallidus slices. Br J Pharmacol 120:1397-1398.
55. Sulcova E, Mechoulam R, Fride E (1998) Biphasic effects of anandamide.
Pharmacol Biochem Behav 59:347-352. 56. Calandra B, Portier M, Kerneis A, Delpech M, Carillon C, Le Fur G, Ferrara P,
Shire D (1999) Dual intracellular signaling pathways mediated by the human cannabinoid CB1 receptor. Eur J Pharmacol 374:445-455.
57. Schlicker E, Timm J, Gothert M (1996) Cannabinoid receptor-mediated
inhibition of dopamine release in the retina. Naunyn Schmiedebergs Arch Pharmacol 354:791-795.
58. Weber B, Schlicker E (2001) Modulation of dopamine release in the guinea-pig
retina by G(i)- but not by G(s)- or G(q)-protein-coupled receptors. In: Fundam Clin Pharmacol, pp 393-400. France.
59. Opere CA, Zheng WD, Zhao M, Lee JS, Kulkarni KH, Ohia SE (2006) Inhibition of
potassium- and ischemia-evoked [3H] D-aspartate release from isolated bovine retina by cannabinoids. In: Curr Eye Res, pp 645-653. England.
60. Savinainen JR, Laitinen JT (2004) Detection of cannabinoid CB1, adenosine A1,
muscarinic acetylcholine, and GABA(B) receptor-dependent G protein activity in transducin-deactivated membranes and autoradiography sections of rat retina. Cell Mol Neurobiol 24:243-256.
61. Warrier A, Wilson M (2007) Endocannabinoid signaling regulates spontaneous
transmitter release from embryonic retinal amacrine cells. Vis Neurosci 24:25-35.
62. Middleton TP, Protti DA (2011) Cannabinoids modulate spontaneous synaptic
activity in retinal ganglion cells. Vis Neurosci 28:393-402.

114
63. Struik ML, Yazulla S, Kamermans M (2006) Cannabinoid agonist WIN 55212-2 speeds up the cone response to light offset in goldfish retina. Vis Neurosci 23:285-293.
64. Lukasiewicz PD, Shields CR (1998b) A diversity of GABA receptors in the retina.
In: Semin Cell Dev Biol, pp 293-299. England: 1998 Academic Press. 65. Ragozzino D, Woodward RM, Murata Y, Eusebi F, Overman LE, Miledi R (1996)
Design and in vitro pharmacology of a selective gamma-aminobutyric acidC receptor antagonist. Mol Pharmacol 50:1024-1030.
66. Eggers ED, Lukasiewicz PD (2011) Multiple pathways of inhibition shape
bipolar cell responses in the retina. In: Vis Neurosci, pp 95-108. England. 67. Wassle H, Koulen P, Brandstatter JH, Fletcher EL, Becker CM (1998) Glycine and
GABA receptors in the mammalian retina. In: Vision Res, pp 1411-1430. England.
68. Lam DM (1997) Neurotransmitters in the vertebrate retina. Invest Ophthalmol
Vis Sci 38:553-556. 69. Shields CR, Tran MN, Wong RO, Lukasiewicz PD (2000) Distinct ionotropic
GABA receptors mediate presynaptic and postsynaptic inhibition in retinal bipolar cells. J Neurosci 20:2673-2682.
70. Lukasiewicz PD, Shields CR (1998a) Different combinations of GABAA and
GABAC receptors confer distinct temporal properties to retinal synaptic responses. J Neurophysiol 79:3157-3167.
71. Feigenspan A, Wassle H, Bormann J (1993) Pharmacology of GABA receptor Cl-
channels in rat retinal bipolar cells. Nature 361:159-162. 72. Pan ZH, Lipton SA (1995) Multiple GABA receptor subtypes mediate inhibition
of calcium influx at rat retinal bipolar cell terminals. J Neurosci 15:2668-2679.
73. Lukasiewicz PD (1996) GABAC receptors in the vertebrate retina. Mol Neurobiol
12:181-194. 74. Lukasiewicz PD, Werblin FS (1994) A novel GABA receptor modulates synaptic
transmission from bipolar to ganglion and amacrine cells in the tiger salamander retina. J Neurosci 14:1213-1223.
75. Vincent P, Armstrong CM, Marty A (1992) Inhibitory synaptic currents in rat
cerebellar Purkinje cells: modulation by postsynaptic depolarization. J Physiol 456:453-471.

115
76. Di S, Malcher-Lopes R, Halmos K, Tasker J (2003) Nongenomic glucocorticoid inhibition via endocannabinoid release in the hypothalamus: a fast feedback mechanism. J Neurosci 23:4850-4857.
77. Di S, Malcher-Lopes R, Marcheselli VL, Bazan NG, Tasker JG (2005a) Rapid
glucocorticoid-mediated endocannabinoid release and opposing regulation of glutamate and gamma-aminobutyric acid inputs to hypothalamic magnocellular neurons. Endocrinology 146:4292-4301.
78. Di S, Boudaba C, Popescu I, Weng F, Harris C, Marcheselli V, Bazan N, Tasker J
(2005b) Activity-dependent release and actions of endocannabinoids in the rat hypothalamic supraoptic nucleus. J Physiol 569:751-760.
79. Hill MN, Tasker JG (2012) Endocannabinoid signaling, glucocorticoid-mediated
negative feedback, and regulation of the hypothalamic-pituitary-adrenal axis. Neuroscience 204:5-16.
80. Morgans CW, Brown RL, Duvoisin RM (2010) TRPM1: the endpoint of the
mGluR6 signal transduction cascade in retinal ON-bipolar cells. Bioessays 32:609-614.
81. Wang GY (2006) Unique functional properties of the APB sensitive and
insensitive rod pathways signaling light decrements in mouse retinal ganglion cells. Vis Neurosci 23:127-135.
82. Wang GY, van der List DA, Nemargut JP, Coombs JL, Chalupa LM (2007) The
sensitivity of light-evoked responses of retinal ganglion cells is decreased in nitric oxide synthase gene knockout mice. In: J Vis, pp 7 1-13. United States.
83. Bai X, Zhu J, Yang J, Savoie BT, Wang GY (2009) Mechanisms that limit the light
stimulus frequency following through the DL-2-amino-4-phosphonobutyric acid sensitive and insensitive rod Off-pathways. Neuroscience 162:184-194.
84. Nemargut JP, Zhu J, Savoie BT, Wang GY (2009) Differential effects of
charybdotoxin on the activity of retinal ganglion cells in the dark- and light-adapted mouse retina. In: Vision Res, pp 388-397. England.
85. Yang J, Nemargut JP, Wang GY (2011) The roles of ionotropic glutamate
receptors along the On and Off signaling pathways in the light-adapted mouse retina. Brain Res 1390:70-79.
86. Demb JB, Haarsma L, Freed MA, Sterling P (1999) Functional circuitry of the
retinal ganglion cell's nonlinear receptive field. J Neurosci 19:9756-9767. 87. Brainard DH (1997) The Psychophysics Toolbox. Spat Vis 10:433-436.

116
88. Pelli DG (1997) The VideoToolbox software for visual psychophysics: transforming numbers into movies. Spat Vis 10:437-442.
89. Lyubarsky AL, Daniele LL, Pugh EN, Jr. (2004) From candelas to
photoisomerizations in the mouse eye by rhodopsin bleaching in situ and the light-rearing dependence of the major components of the mouse ERG. Vision Res 44:3235-3251.
90. Field GD, Rieke F (2002) Nonlinear signal transfer from mouse rods to bipolar
cells and implications for visual sensitivity. Neuron 34:773-785. 91. Tsukamoto Y, Morigiwa K, Ueda M, Sterling P (2001) Microcircuits for night
vision in mouse retina. J Neurosci 21:8616-8623. 92. Pang JJ, Gao F, Wu SM (2003) Light-evoked excitatory and inhibitory synaptic
inputs to ON and OFF alpha ganglion cells in the mouse retina. J Neurosci 23:6063-6073.
93. Duvoisin RM, Zhang C, Ramonell K (1995) A novel metabotropic glutamate
receptor expressed in the retina and olfactory bulb. J Neurosci 15:3075-3083. 94. Wu S, Wright RA, Rockey PK, Burgett SG, Arnold JS, Rosteck PR, Jr., Johnson BG,
Schoepp DD, Belagaje RM (1998) Group III human metabotropic glutamate receptors 4, 7 and 8: molecular cloning, functional expression, and comparison of pharmacological properties in RGT cells. Brain Res Mol Brain Res 53:88-97.
95. Bauer B, Ehinger B, Aberg L (1980) [3H]-dopamine release from the rabbit
retina. Albrecht Von Graefes Arch Klin Exp Ophthalmol 215:71-78. 96. Godley BF, Wurtman RJ (1988) Release of endogenous dopamine from the
superfused rabbit retina in vitro: effect of light stimulation. Brain Res 452:393-395.
97. Witkovsky P (2004) Dopamine and retinal function. Doc Ophthalmol 108:17-40. 98. Hu EH, Pan F, Volgyi B, Bloomfield SA (2010) Light increases the gap junctional
coupling of retinal ganglion cells. J Physiol 588:4145-4163. 99. Zhang DQ, Zhou TR, McMahon DG (2007) Functional heterogeneity of retinal
dopaminergic neurons underlying their multiple roles in vision. J Neurosci 27:692-699.
100. Contini M, Lin B, Kobayashi K, Okano H, Masland RH, Raviola E (2010) Synaptic
input of ON-bipolar cells onto the dopaminergic neurons of the mouse retina. J Comp Neurol 518:2035-2050.

117
101. Muller F, Scholten A, Ivanova E, Haverkamp S, Kremmer E, Kaupp UB (2003) HCN channels are expressed differentially in retinal bipolar cells and concentrated at synaptic terminals. Eur J Neurosci 17:2084-2096.
102. Chen L, Yang XL (2007) Hyperpolarization-activated cation current is involved
in modulation of the excitability of rat retinal ganglion cells by dopamine. Neuroscience 150:299-308.
103. Knop GC, Seeliger MW, Thiel F, Mataruga A, Kaupp UB, Friedburg C, Tanimoto
N, Muller F (2008) Light responses in the mouse retina are prolonged upon targeted deletion of the HCN1 channel gene. Eur J Neurosci 28:2221-2230.
104. Oi H, Partida GJ, Lee SC, Ishida AT (2008) HCN4-like immunoreactivity in rat
retinal ganglion cells. Vis Neurosci 25:95-102. 105. Akopian A, Witkovsky P (1996) D2 dopamine receptor-mediated inhibition of
a hyperpolarization-activated current in rod photoreceptors. J Neurophysiol 76:1828-1835.
106. Soucy E, Wang Y, Nirenberg S, Nathans J, Meister M (1998) A novel signaling
pathway from rod photoreceptors to ganglion cells in mammalian retina. Neuron 21:481-493.
107. Doly M (1994) Transduction of the light message: from the photon to the optic
nerve. Fundam Clin Pharmacol 8:147-154. 108. Nawy S, Sie A, Copenhagen DR (1989) The glutamate analog 2-amino-4-
phosphonobutyrate antagonizes synaptic transmission from cones to horizontal cells in the goldfish retina. Proc Natl Acad Sci U S A 86:1726-1730.
109. Koulen P, Kuhn R, Wassle H, Brandstatter JH (1999) Modulation of the
intracellular calcium concentration in photoreceptor terminals by a presynaptic metabotropic glutamate receptor. Proc Natl Acad Sci U S A 96:9909-9914.
110. Cartmell J, Schoepp DD (2000) Regulation of neurotransmitter release by
metabotropic glutamate receptors. J Neurochem 75:889-907. 111. Hirasawa H, Shiells R, Yamada M (2002) A metabotropic glutamate receptor
regulates transmitter release from cone presynaptic terminals in carp retinal slices. J Gen Physiol 119:55-68.
112. Kim IB, Lee EJ, Kang TH, Chung JW, Chun MH (2003) Morphological analysis of
the hyperpolarization-activated cyclic nucleotide-gated cation channel 1 (HCN1) immunoreactive bipolar cells in the rabbit retina. J Comp Neurol 467:389-402.

118
113. Koizumi A, Jakobs TC, Masland RH (2004) Inward rectifying currents stabilize the membrane potential in dendrites of mouse amacrine cells: patch-clamp recordings and single-cell RT-PCR. Mol Vis 10:328-340.
114. Fyk-Kolodziej B, Pourcho RG (2007) Differential distribution of
hyperpolarization-activated and cyclic nucleotide-gated channels in cone bipolar cells of the rat retina. J Comp Neurol 501:891-903.
115. van der Stelt M, Di Marzo V (2005) Cannabinoid receptors and their role in
neuroprotection. In: Neuromolecular Med, pp 37-50. United States. 116. Wilson RI, Nicoll RA (2001) Endogenous cannabinoids mediate retrograde
signalling at hippocampal synapses. In: Nature, pp 588-592. England. 117. Kreitzer AC, Regehr WG (2001b) Retrograde inhibition of presynaptic calcium
influx by endogenous cannabinoids at excitatory synapses onto Purkinje cells. In: Neuron, pp 717-727. United States.
118. Ohno-Shosaku T, Maejima T, Kano M (2001) Endogenous cannabinoids
mediate retrograde signals from depolarized postsynaptic neurons to presynaptic terminals. In: Neuron, pp 729-738. United States.
119. Hajos N, Ledent C, Freund TF (2001) Novel cannabinoid-sensitive receptor
mediates inhibition of glutamatergic synaptic transmission in the hippocampus. Neuroscience 106:1-4.
120. Degroot A, Kofalvi A, Wade MR, Davis RJ, Rodrigues RJ, Rebola N, Cunha RA,
Nomikos GG (2006) CB1 receptor antagonism increases hippocampal acetylcholine release: site and mechanism of action. Mol Pharmacol 70:1236-1245.
121. Pitler TA, Alger BE (1992) Postsynaptic spike firing reduces synaptic GABAA
responses in hippocampal pyramidal cells. J Neurosci 12:4122-4132. 122. Lukasiewicz PD, Maple BR, Werblin FS (1994) A novel GABA receptor on
bipolar cell terminals in the tiger salamander retina. J Neurosci 14:1202-1212.
123. Zhang J, Slaughter MM (1995) Preferential suppression of the ON pathway by
GABAC receptors in the amphibian retina. J Neurophysiol 74:1583-1592. 124. Kreitzer AC, Regehr WG (2001a) Cerebellar depolarization-induced
suppression of inhibition is mediated by endogenous cannabinoids. In: J Neurosci, p RC174. United States.

119
125. Wilson RI, Kunos G, Nicoll RA (2001) Presynaptic specificity of endocannabinoid signaling in the hippocampus. In: Neuron, pp 453-462. United States.
126. Bojesen IN, Hansen HS (2003) Binding of anandamide to bovine serum
albumin. J Lipid Res 44:1790-1794. 127. Vaughn JE, Famiglietti EV, Jr., Barber RP, Saito K, Roberts E, Ribak CE (1981)
GABAergic amacrine cells in rat retina: immunocytochemical identification and synaptic connectivity. J Comp Neurol 197:113-127.
128. Lukasiewicz PD, Eggers ED, Sagdullaev BT, McCall MA (2004b) GABAC
receptor-mediated inhibition in the retina. In: Vision Res, pp 3289-3296. England.
129. Cook PB, McReynolds JS (1998a) Lateral inhibition in the inner retina is
important for spatial tuning of ganglion cells. Nat Neurosci 1:714-719. 130. Flores-Herr N, Protti DA, Wassle H (2001) Synaptic currents generating the
inhibitory surround of ganglion cells in the mammalian retina. J Neurosci 21:4852-4863.
131. Slaughter MM, Pan ZH (1992) The physiology of GABAB receptors in the
vertebrate retina. Prog Brain Res 90:47-60. 132. Grunert U (1999) Distribution of GABAA and glycine receptors in the
mammalian retina. Clin Exp Pharmacol Physiol 26:941-944. 133. Euler T, Masland RH (2000) Light-evoked responses of bipolar cells in a
mammalian retina. J Neurophysiol 83:1817-1829. 134. Lukasiewicz P, Eggers E, Sagdullaev B, McCall M (2004a) GABAC receptor-
mediated inhibition in the retina. Vision Res 44:3289-3296. 135. Dong CJ, Werblin FS (1998) Temporal contrast enhancement via GABAC
feedback at bipolar terminals in the tiger salamander retina. J Neurophysiol 79:2171-2180.
136. Freed MA, Smith RG, Sterling P (2003) Timing of quantal release from the
retinal bipolar terminal is regulated by a feedback circuit. Neuron 38:89-101. 137. Fletcher EL, Koulen P, Wassle H (1998) GABAA and GABAC receptors on
mammalian rod bipolar cells. J Comp Neurol 396:351-365.

120
138. Koulen P, Brandstatter JH, Enz R, Bormann J, Wassle H (1998a) Synaptic clustering of GABA(C) receptor rho-subunits in the rat retina. Eur J Neurosci 10:115-127.
139. Koulen P, Malitschek B, Kuhn R, Bettler B, Wassle H, Brandstatter JH (1998b)
Presynaptic and postsynaptic localization of GABA(B) receptors in neurons of the rat retina. Eur J Neurosci 10:1446-1456.
140. Shen W, Slaughter MM (2001) Multireceptor GABAergic regulation of synaptic communication in amphibian retina. In: J Physiol, pp 55-67. England.
141. Caldwell JH, Daw NW (1978) Effects of picrotoxin and strychnine on rabbit
retinal ganglion cells: changes in centre surround receptive fields. J Physiol 276:299-310.

121
BIOGRAPHY
Joshua Nathaniel Pahng was born to Sung-Cue Pahng and Colleen Marvin at
Camp Lejune Naval Hospital in Jacksonville, North Carolina on November 24th, 1985.
In 2000, he and his family moved to Manassas, Virginia where he attended
Stonewall Jackson High School. He earned his Bachelor of Science degree at the
College of William and Mary in Williamsburg, Virginia in 2008. In the fall of 2008,
Joshua entered the Ph.D. Program in Neuroscience at Tulane University in New
Orleans, Louisiana. There, he conducted his research under the mentorship of his
advisor, Dr. Guoyong Wang.