ceramide and ceramide 1-phosphate are negative … · · 2010-11-11ceramide and ceramide...
TRANSCRIPT

of June 14, 2018.This information is current as
Induced by Lipopolysaccharide ProductionαNegative Regulators of TNF-
Ceramide and Ceramide 1-Phosphate Are
and Andrzej SobotaAlicja Bielawska, Jacek Bielawski, Katarzyna Kwiatkowska Szczepan Józefowski, Maciej Czerkies, Anna Lukasik,
http://www.jimmunol.org/content/185/11/6960doi: 10.4049/jimmunol.0902926November 2010;
2010; 185:6960-6973; Prepublished online 1J Immunol
Referenceshttp://www.jimmunol.org/content/185/11/6960.full#ref-list-1
, 35 of which you can access for free at: cites 75 articlesThis article
average*
4 weeks from acceptance to publicationFast Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
Submit online. ?The JIWhy
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. All rights reserved.1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on June 14, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
by guest on June 14, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

The Journal of Immunology
Ceramide and Ceramide 1-Phosphate Are NegativeRegulators of TNF-a Production Induced byLipopolysaccharide
Szczepan Jozefowski,* Maciej Czerkies,* Anna Łukasik,* Alicja Bielawska,†
Jacek Bielawski,† Katarzyna Kwiatkowska,* and Andrzej Sobota*
LPS is a constituent of cell walls of Gram-negative bacteria that, acting through the CD14/TLR4 receptor complex, causes strong
proinflammatory activation of macrophages. In murine peritoneal macrophages and J774 cells, LPS at 1–2 ng/ml induced maximal
TNF-a and MIP-2 release, and higher LPS concentrations were less effective, which suggested a negative control of LPS action.
While studying the mechanism of this negative regulation, we found that in J774 cells, LPS activated both acid sphingomyelinase
and neutral sphingomyelinase and moderately elevated ceramide, ceramide 1-phosphate, and sphingosine levels. Lowering of the
acid sphingomyelinase and neutral sphingomyelinase activities using inhibitors or gene silencing upregulated TNF-a and MIP-2
production in J774 cells and macrophages. Accordingly, treatment of those cells with exogenous C8-ceramide diminished TNF-a
and MIP-2 production after LPS stimulation. Exposure of J774 cells to bacterial sphingomyelinase or interference with ceramide
hydrolysis using inhibitors of ceramidases also lowered the LPS-induced TNF-a production. The latter result indicates that
ceramide rather than sphingosine suppresses TNF-a and MIP-2 production. Of these two cytokines, only TNF-a was negatively
regulated by ceramide 1-phosphate as was indicated by upregulated TNF-a production after silencing of ceramide kinase gene
expression. None of the above treatments diminished NO or RANTES production induced by LPS. Together the data indicate that
ceramide negatively regulates production of TNF-a and MIP-2 in response to LPS with the former being sensitive to ceramide
1-phosphate as well. We hypothesize that the ceramide-mediated anti-inflammatory pathway may play a role in preventing
endotoxic shock and in limiting inflammation. The Journal of Immunology, 2010, 185: 6960–6973.
Ceramide generated by hydrolysis of membrane sphingo-myelin by neutral sphingomyelinase (NSMase) or acidsphingomyelinase (ASMase) or by de novo synthesis has
been implicated in mediating or regulating numerous cellularprocesses as diverse as proliferation, differentiation and senescence,stress responses, apoptosis, and inflammation. The complexity ofthe ceramide-mediated signaling is augmented by the fact thatceramide may also serve as a precursor of other bioactive sphin-golipids, such as ceramide 1-phosphate, sphingosine, or sphingosine1-phosphate, sometimes exerting biological effects opposing those
of ceramide (1). The exact molecular mechanisms by whichceramide performs its functions are not completely understood.Ceramide has been reported to affect the activity of numerousenzymes involved in intracellular signaling, such as serine/threonine phosphatases PP1 and PP2A, kinase suppressor of Ras,protein kinase Raf-1, and different isoforms of protein kinase C (2–4). Besides acting as a second messenger, ceramide may also in-duce effects secondary to changes it produces in the biophysicalproperties of biological membranes. The conversion of sphingo-myelin to ceramide within artificial or natural membranes has beenreported to induce their fusion/fission, vesiculation, and channelformation, as well as to facilitate the interaction of plasma mem-brane receptors with lipid rafts (5–11).Ceramide has been postulated to modulate cellular responses to
LPS, a major constituent of cell walls of Gram-negative bacteria.LPS induces strong proinflammatory and microbicidal activation ofmacrophages and other cell types, reflected by production ofcytokines, such as TNF-a, chemokines, PGs, and NO, which, ifnot appropriately controlled, may lead to an organism’s deathresulting from septic shock (12). The recognition of enterobacte-rial LPS by macrophages involves cooperation between the GPI-anchored receptor CD14, which binds LPS with high affinity, andthe MD-2/TLR4 heterodimer (13–15). CD14 binds LPS mono-mers extracted from LPS micelles by soluble LPS-binding protein(LBP) and transfers them to MD-2, which leads to aggregation ofMD-2–associated TLR4 and initiation of signal transductionpathways (16). Recent studies have indicated a dichotomy of theTLR4 signaling cascades, which either require or do not requireparticipation of the adapter protein MyD88 (17).The scarce reports on ceramide participation in LPS signaling
pathways are often contradictory. Stimulation with LPS has beenreported to cause activation of ASMase and ASMase-dependent
*Laboratory of Plasma Membrane Receptors, Department of Cell Biology, NenckiInstitute of Experimental Biology, 02-093 Warsaw, Poland; and †Lipidomics SharedResource, Department of Biochemistry and Molecular Biology, Medical Universityof South Carolina, Charleston, SC 29425
Received for publication September 3, 2009. Accepted for publication September 21,2010.
This work was supported by Grants N N301 0147 33 and N N301 0302 34 from theMinistry of Science and Higher Education (Poland).
Address correspondence and reprint requests to Dr. Andrzej Sobota, Department ofCell Biology, Nencki Institute of Experimental Biology, 3 Pasteur Street, 02-093Warsaw, Poland. E-mail address: [email protected]
Abbreviations used in this paper: ASMase, acid sphingomyelinase; B13, (1R,2R)-2-N-myristoylamino-1-(4-nitrophenyl)-1,3-propandiol; bSMase, bacterial sphingomye-linase; C8-Cer-1P, C8-ceramide 1-phosphate; C24:1-Cer-1P, C24:1-ceramide 1-phos-phate; CERK, ceramide kinase; D-e-MAPP, (1S,2R)-D-erythro-2-(N-myristoylamino)-1-phenyl-1-propanol; dhC16, C16-dihydroceramide; dhSph, dihydrosphingosine;GW4869, N,N9-Bis[4-(4,5-dihydro-1H-imidazol-2-yl)phenyl]-3,39-p-phenylene-bis-ac-rylamide dihydrochloride; HBSS, solution composed of 125 mM NaCl, 4 mM KCl,10 mM NaHCO3, 1 mM KH2PO4, 10 mM glucose, 1 mM CaCl2, 1 mMMgCl2, and 20mM Hepes; IS, internal standard; LBP, LPS-binding protein; OxPAPC, oxidized 1-palmitoyl-2-arachidonyl-sn-glycero-3-phosphocholine; NSMase, neutral sphingomyeli-nase; PCERA-1, phospho-ceramide–analogue-1; siRNA, small interfering RNA; Sph,sphingosine; Sph-1P, sphingosine 1-phosphate.
Copyright� 2010 by TheAmericanAssociation of Immunologists, Inc. 0022-1767/10/$16.00
www.jimmunol.org/cgi/doi/10.4049/jimmunol.0902926
by guest on June 14, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

ceramide production in macrophage-like cell lines. However, neitherexogenous ceramide nor ceramide generated from plasmamembranesphingomyelin by bacterial sphingomyelinase (bSMase) were foundto stimulate NF-kB activation or TNF-a and IL-6 release in mac-rophages or macrophage-like cell lines (18–20). The lipid has beenproposed to act as a TLR4 agonist but also as a CD14 ligand thatcan trigger the assembly of multimolecular complexes aroundCD14 (21, 22). In the latter case, the complexes lacked TLR4 andwere, therefore, most likely unable to trigger signaling (22). A fewreports have indicated a positive role of the ASMase-releasedceramide in the macrophage responses to LPS. Direct and indirect(by targeting an upstream phospholipase C) inhibition of ASMasesuppressed the LPS-induced ceramide production, recruitment ofTLR4 to lipid rafts, activation of NF-kB (a transcription factorplaying an essential role in the induction of inflammatory media-tors), and cytokine release from THP-1 cells (23, 24). Quite to thecontrary, C8-ceramide reduced TNF-a and IL-6 but not IL-10 pro-duction in LPS-stimulated macrophages exhibiting simultaneouslyan opposite effect in mast cells (25), whereas C6-ceramide inhibitedLPS-stimulated IL-8 production in human aortic endothelial cells(26). Most importantly, ASMase KO mice display an unimpairedincrease in serum TNF-a in response to LPS (27).To our knowledge, only a few earlier reports addressed the role of
NSMase in regulation of IL-8 release (28) and expression of in-ducible NO-synthase (29, 30) in LPS-stimulated epithelial cells,glioma cells, and macrophages, respectively. The studies werebased on application of then newly synthesized inhibitors ofNSMase and indicated a positive role of the NSMase-generatedceramide in cell reaction to LPS.Because of the importance of the LPS-induced responses in the
immunity against Gram-negative bacteria and in the pathogenesisof septic shock, we sought to study in more detail the role of sphin-golipid mediators in LPS signaling. Our data indicate that cer-amide negatively regulates LPS-stimulated production of TNF-aand MIP-2 but not that of NO or the chemokine RANTES, andceramide 1-phosphate, product of ceramide kinase (CERK) activ-ity, inhibits generation of TNF-a only.
Materials and MethodsReagents
(1S,2R)-D-erythro-2-(N-myristoylamino)-1-phenyl-1-propanol (D-e-MAPP) and(1R,2R)-2-N-myristoylamino-1-(4-nitrophenyl)-1,3-propandiol (B13) werefrom Lipidomics Shared Resource, Medical University of South Carolina.C8-ceramide and C8-ceramide 1-phosphate were purchased from Avanti PolarLipids (Alabaster, AL), C2-dihydroceramide from Biomol (Hamburg, Ger-many), and sphingosine 1-phosphate and N,N-dimethylsphingosine fromCayman Chemical (Ann Arbor, MI). Ultrapure LPS from Escherichia coli0111:B4, containing, according to the manufacturer, no detectable pro-teins, was purchased from List Biological Laboratories (Campbell, CA).N,N9-Bis[4-(4,5-dihydro-1H-imidazol-2-yl)phenyl]-3,39-p-phenylene-bis-acrylamide dihydrochloride (GW4869) imipramine, sphingomyelinasefrom Bacillus cereus (bSMase), D-e-sphingosine, and C2-ceramide werefrom Sigma-Aldrich (Poznan, Poland).
GW4869 was suspended in DMSO at 7.5 mM, aliquoted, and stored at270˚C. Before use it was solubilized by adding 1/20 volume of 5% meth-anesulfonic acid. Stock solutions (20–50 mM) of other drugs and sphin-golipids were prepared in DMSO, except for imipramine, which was dis-solved at 400 mM in dimethylformamide, and N,N-dimethylsphingosine,which was provided as an ethanol solution.
Cell isolation and culture
BALB/c mice were purchased from the Center of Experimental Medicine(Białystok, Poland). Female mice, 9–14 wk old, were euthanized by in-halation of isoflurane (Baxter, Deerfield, IL), followed by cervical dislo-cation. Inflammatory peritoneal cells, elicited with 1 ml aged 3% thio-glycolate (Difco, Lawrence, KS) injected i.p. 4–5 d earlier, were washed outwith PBS, resuspended in RPMI 1640mediumwith 25mMHEPES (Sigma-Aldrich), pH 7.4, supplemented with 10% FCS (Life Technologies, Warsaw,
Poland) and 2 mM L-glutamine (RPMI/10% FCS medium), and plated at1.33 105 cells/well in 96-well tissue culture-treated plates. After overnightincubation, nonadherent cells were removed by washing leaving adherentmacrophages for experiments. The studies were reviewed and approvedby an appropriate local animal ethics committee.
The culture of J774A.1 macrophage-like cells was maintained undernonadherent or loosely adherent conditions in RPMI/10% FCS medium inunmodified polystyrene (bacteriological grade) Petri dishes. One day beforeexperiments, the cells were plated at 13 105/well in 96-well tissue culture-treated plates.
Cell treatment
Adherent macrophages were stimulated in 160 ml medium with LPS for 4 h(TNF-a, MIP-2, or RANTES production) or 24 h (NO release). Whenindicated, cells were preincubated for 80 min with 100 mM imipramine, 14mM GW4869, 25 mM B13, 25 mM D-e-MAPP, or for 30 min with 10 mMceramide kinase inhibitor K1 (Calbiochem, San Diego, CA) or equivalentamounts of solvents (37˚C) before stimulation with LPS in the presence ofthe inhibitors. When the effects of the inhibitors on LPS-stimulated NOproduction were to be assessed, cells were co-incubated with LPS andinhibitors for 3 h only (and for the remaining 21 h in medium alone) be-cause a 24-h co-incubation with imipramine decreased cell viability. Inanother set of experiments, cells were preincubated for 40 min with 10 mMsphingolipids (C2-ceramide, C2-dihydroceramide, C8-ceramide, C8-ceramide1-phosphate, sphingosine, or sphingosine 1-phosphate), and the lipids werealso present during stimulation with LPS. C8-ceramide 1-phosphate stocksolution was warmed up to 50˚C for 1 min before adding to cell cultures.When N,N-dimethylsphingosine was applied, cells were treated with thedrug (15 mM) for 30 min prior to addition of 10 mM C8-ceramide or10 mM sphingosine for a further 40 min incubation, which was followed bystimulation of cells with LPS in the presence of the drug and lipids. Ex-ogenous bSMase (0.5 U/ml) was applied in 160 ml serum-free RPMImedium for 5–60 min (4˚C).
TNF-a, MIP-2, NO, and RANTES assays
TNF-a, MIP-2, and RANTES levels in culture supernatants of cells treatedfor 4 h with LPS were determined with the use of appropriate murineELISA Development Kits (PeproTech, Rocky Hill, NJ), according to themanufacturer’s instructions, using a Sunrise plate reader (Tecan Group,Mannedof, Switzerland). Nitrite concentrations in 100-ml aliquots of cul-ture supernatants were determined 24 h after LPS application according toa modified Griess method, as described previously (31).
Silencing of ASMase, NSMase, and CERK genes
J774 cells were harvested by centrifugation (10 min, 400 3 g) and resus-pended (2 3 106 cells/100 ml) in Nucleofector Solution V (Amaxa, Koeln,Germany) prewarmed to room temperature. Mouse ASMase, NSMase(Smpd2), or CERK small interfering RNA (siRNA) or control scrambledsiRNA (Ambion, Austin, TX) was added to the cells (75 pmol/106 cells),and the cells were submitted to transfection using the T-020 program anda Nucleofector II device (Amaxa) according to the manufacturer’s protocol.Immediately after transfection, 900 ml RPMI/10%FCS medium (preheatedto 37˚C) was added to the cells, and the cells were cultured for 48 h forfurther experiments.
ASMase activity assay
Cells were plated at 1 3 106 in 1 ml RPMI/10% FCS in 12-well tissueculture plates and, when indicated, were stimulated by adding 1 ml LPSstock solutions without changing medium. After chilling on ice andwashing with cold solution composed of 125 mMNaCl, 4 mMKCl, 10 mMNaHCO3, 1 mMKH2PO4, 10 mM glucose, 1 mM CaCl2, 1 mMMgCl2, and20 mM Hepes (pH 7.4) (HBSS), cells were lysed for 30 min at 4˚C in 260ml of a buffer composed of 0.2% Triton X-100, 50 mM NaCl, 2 mM EDTA,20 mM Tris-HCl, pH 7.4. Aliquots of cell lysates (100 ml) were mixed with150 ml ASMase assay buffer containing 135 mM sodium acetate (final pH5), 1.3 mM EDTA, and liposomes of 10 mM NBD-C6-sphingomyelin(Avanti) and 30 mM 1,2-dioleoyl-sn-glycerol-3-phosphocholine (32). Af-ter 1.5 h at 37˚C, the reaction was stopped by adding 0.45 ml 2-propanol,1.5 ml heptane, and 0.2 ml PBS. After phase separation, the fluorescence ofthe upper phase was measured with a Spex spectrofluorimeter (Yobin-Yvon,Edison, NJ) with excitation at 460 nm and emission at 534 nm. The fluo-rescence values were normalized for protein content determined with theBradford ULTRA kit (Novexin, Cambridge, U.K.).
NSMase activity assay
To assay NSMase activity, the release of phosphorylcholine using un-modified sphingomyelin (Avanti) as a substrate was measured according to
The Journal of Immunology 6961
by guest on June 14, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

Ref. 33. Cells (1 3 106 in 1 ml RPMI/10% FCS in 12-well tissue cultureplates) were preincubated for 1 h with 100 mM imipramine to depleteASMase. When indicated, LPS was added in 1-ml aliquots to cell cultures,and at various time points of stimulation, cells were scraped into 130 mllysis buffer composed of 0.2% Triton X-100, 5 mM MgCl2, proteaseinhibitors (2 mM PMSF, 10 mg/ml aprotinin, leupeptin, and pepstatineach), and 50 mM HEPES, pH 7.4. The cells were lysed for 30 min on iceand 100 ml of the lysates was mixed with 100 ml of a solution containing0.4% Triton X-100, 1 mM C16-sphingomyelin, 6 mM 4-hydroxyphenyl-acetic acid, 10 U/ml alkaline phosphatase (Pierce, Rockford, IL), 0.5 U/mlcholine oxidase, 20 U/ml HRP, and 100 mM HEPES, pH 7.4. After 1 h(37˚C), the reaction was terminated by diluting samples with 0.5 ml ice-cold 10 mM EDTA in PBS, and the fluorescence of the product ofhydroxyphenylacetic acid oxidation was measured at 320 nm/400 nm ina Spex spectrofluorimeter.
Determination of total cellular ceramide by radiolabeling
Cells were labeled for 40 h with [3H]serine (Perkin Elmer, Waltham, MA)at 4 mCi/ml per 1 3 106 cells in RPMI 1640 medium supplemented with6% FBS. After the treatment, cells were washed with HBSS solution,scraped from the dishes, and resuspended in the same buffer. After cen-trifugation (3 min, 400 3 g), cells were lysed in 1 ml 0.1% SDS, and 100ml of the resulting lysate was used for determination of protein contentusing the Bradford method. The remaining 900 ml was subjected to lipidextraction and separation by TLC according to Ref. 34. After drying andvisualizing lipids by iodine vapor, the position of ceramide was determinedby comparison with a standard C16-ceramide (Avanti). Designated spotswere scraped off and quantified by liquid scintillation in a BeckmanLS6000TA counter (Vienna, Austria) and normalized against protein amo-unt in the samples.
Assessment of cell surface ceramide level
The ceramide content in the external leaflet of the plasma membrane wasdetermined by flow cytometry in triplicate with the use of mouse anti-ceramide IgM, clone 15B4 (Alexis, Lausen, Switzerland). Suspendedcells were fixed for 15 min with 1% paraformaldehyde in HBSS (4˚C),washed, blocked with 3% goat serum in PBS, and incubated with 2 mg/mlof the anti-ceramide IgM or the same concentration of control, nonimmuneIgM in PBS (30 min). After extensive washing with PBS, cells were in-cubated for 40 min with FITC-conjugated goat anti-mouse IgM Ab(Sigma-Aldrich), washed again, resuspended in 500 ml PBS, and analyzedin a Becton Dickinson FACSCalibur flow cytometer (Warsaw, Poland). Persample, 10,000 cells were analyzed, and the mean of the total fluorescenceintensity was taken as the value reflecting the amount of the cell surfaceceramide. To calculate ceramide-specific signals, the value of a nonspecificfluorescence detected in samples treated with control IgM was subtractedfrom fluorescence of samples exposed to anti-ceramide IgM.
Liquid chromatography-mass spectrometry analysis ofendogenous sphingolipids
Advanced analyses were performed on a Thermo Finnigan TSQ 7000(Ringoes, NJ), triple-stage quadrupole mass spectrometer operating ina multiple reaction monitoring positive ionization mode as described (35).Quantitative analysis of the cellular level of sphingolipids was based on thecalibration curves generated by spiking an artificial matrix with knownamounts of target standards and an equal amount of the internal standard(IS). The target analyte to IS peak area ratios from the samples were simi-larly normalized to their respective IS and compared with the calibrationcurves using a linear regression model. Final results were expressed asthe level of the particular sphingolipids/protein (pmol/mg). Protein con-centration was determined in 0.5% SDS-cell lysates with Micro BCA Pro-tein Assay Kit (Pierce).
Cell viability
Cells were incubated for 15 min with 0.5 mg/ml MTT in 100 ml RPMI/10%FCS medium. Subsequently, the wells were washed once with PBS, and theproduct of MTT reduction was dissolved in 100 ml DMSO and measured at509 nm in a Sunrise plate reader. Treatment with 1 and 100 ng/ml LPS (4–24 h) decreased cell viability by ∼5 and ∼10%, respectively. The sphin-gomyelinase inhibitors and sphingolipids tested did not impair cell via-bility.
Immunoblotting
Proteins of whole lysates of J774 cells were separated by 10% SDS-PAGE,transferred onto nitrocellulose, and probed with rabbit anti-CERK (Santa
Cruz Biotechnology, Santa Cruz, CA) and mouse anti-actin Ab (RocheDiagnostics, Mannheim, Germany) followed by goat anti-rabbit or anti-mouse IgG conjugated with peroxidase (Santa Cruz Biotechnology). Im-munoreactive bands were visualized with SuperSignal West Pico Chemi-luminescent substrate (Pierce) and analyzed by densitometry using TL100image analysis software (NonlinearDynamics,NewcastleUponTyne,U.K.).
Data analysis
After assessing homogeneity of variances with the F test, statistical sig-nificance of differences between groups was evaluated with unpairedStudent t test for single comparison or ANOVA for multiple comparisons,with the assumption that p values ,0.05 indicate statistically significantdifferences (GraphPad Prism software, GraphPad Software, La Jolla, CA).
ResultsTNF-a production in response to LPS is negatively modulatedby sphingomyelinases
To examine the involvement of sphingomyelin turnover in LPSsignaling, we interfered with the ASMase and NSMase activity inJ774 macrophage-like cells and assessed TNF-a production stimu-lated by 1 and 100 ng/ml LPS. First, ASMase activity was dimini-shed with imipramine, which acts by inducing proteolytic degra-dation of the enzyme (36). Treatment of the cells with 100 mMimipramine decreased the ASMase activity by ~90% (Fig. 1A).The drug, however, did not change the level of TNF-a release inresponse to 1 ng/ml LPS. In contrast, in cells stimulated with100 ng/ml LPS, a pretreatment with imipramine enhanced TNF-aproduction by 45% (Fig. 1C). TNF-a generation was affected ina similar manner by GW4869, an inhibitor of neutral sphingo-myelinase, which also increased by ~41% the amount of TNF-aproduced by the cells at 100 ng/ml LPS (Fig. 1C). The reportedselectivity of GW4869 (37) was confirmed also for J774 cells: thedrug (14 mM) had no effect on ASMase activity (Fig. 1A) butinhibited NSMase by 60% (Fig. 1B). Notably, sphingomyelinaseactivity measured at neutral pH was also reduced by imipramine(27% inhibition), which suggested the activity was in part dueto that of ASMase. Accordingly, the sphingomyelinase activitydetected at neutral pH was strongly inhibited (by ~86%) after anexposure of cells to imipramine and GW4869 combined (Fig. 1B).Combination of imipramine and GW4869 had a synergistic effecton the TNF-a production stimulated in J774 cells by 100 ng/mlLPS, enhancing it by 121%. The combined drugs, however, didnot enhance TNF-a production stimulated by 1 ng/ml LPS (Fig.1C). When applied to thioglycolate-elicited macrophages, imip-ramine and GW4869 exerted a positive effect on TNF-a pro-duction in a manner qualitatively similar to that seen in J774 cells.Pretreatment of cells with either drug led to a 2-fold elevation ofTNF-a release induced by 10 ng/ml and 1.6- to 2-fold elevation ofTNF-a at 1000 ng/ml LPS. However, neither imipramine norGW4869 affected the TNF-a generation in cells stimulated with1 ng/ml LPS (Fig. 1E). To examine whether the positive effect ofthe ASMase and NSMase inhibitors was limited to LPS-stimula-ted TNF-a production, we estimated MIP-2 generation in cells pre-treated with those drugs. MIP-2 was chosen for analysis, as thesynthesis of both TNF-a and MIP-2 is controlled by the MyD88-dependent signaling pathway (38, 39). Imipramine and GW4869upregulated MIP-2 generation stimulated by 10 or 1000 ng/ml LPSbut not that stimulated by 1 ng/ml LPS, as they did with respect toTNF-a production (Fig. 1F).To verify that the imipramine and GW4869 effects could be
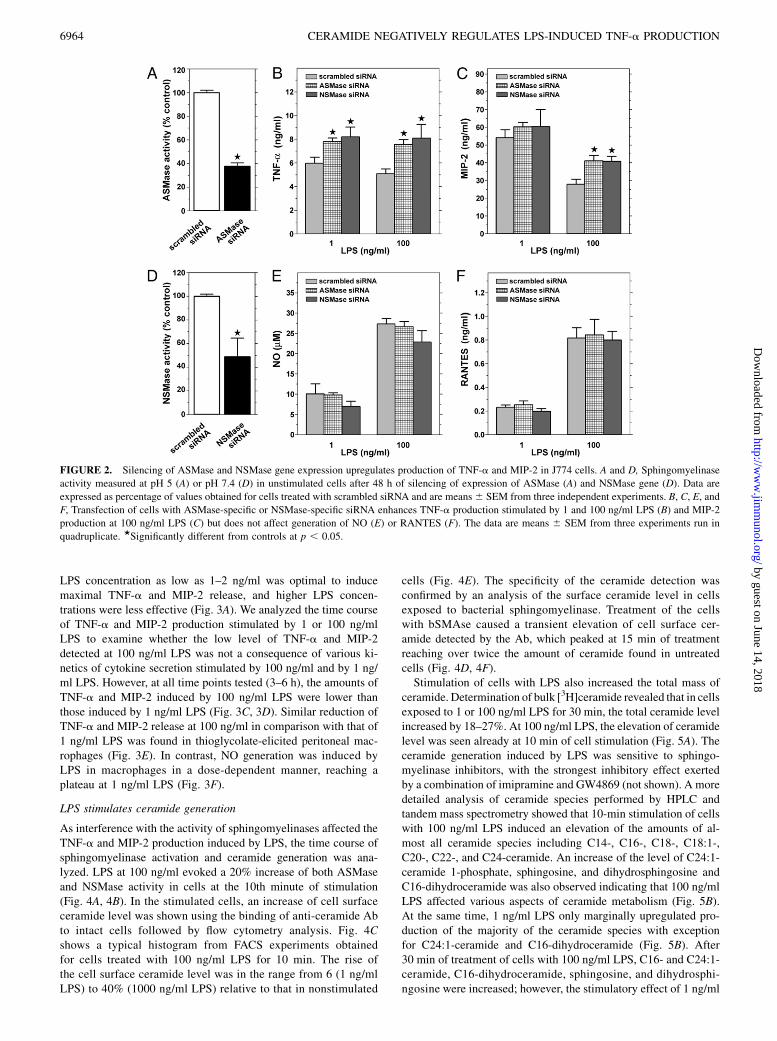
ascribed to their inhibitory action toward ASMase and NSMase,the release of TNF-a was measured in cells after silencing of theexpression of the two sphingomyelinases. An analysis of sphin-gomyelinase activity reveled that transfection of cells with speci-fic siRNA led to a depletion of ASMase and NSMase activity in
6962 CERAMIDE NEGATIVELY REGULATES LPS-INDUCED TNF-a PRODUCTION
by guest on June 14, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
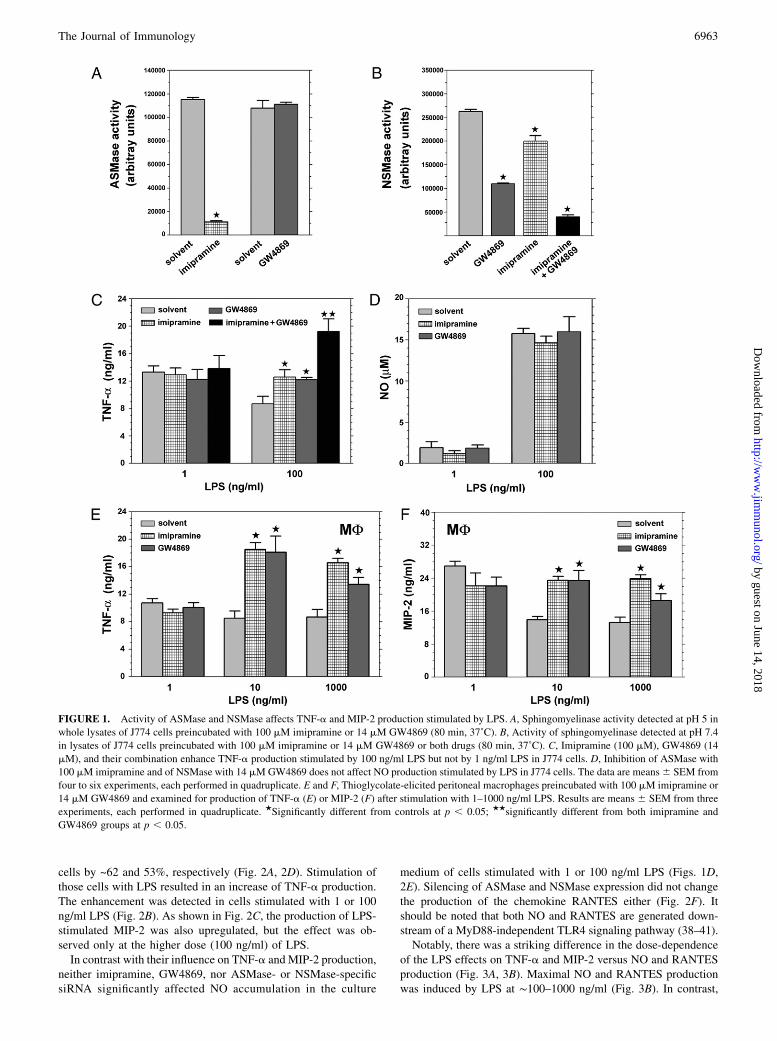
cells by ~62 and 53%, respectively (Fig. 2A, 2D). Stimulation ofthose cells with LPS resulted in an increase of TNF-a production.The enhancement was detected in cells stimulated with 1 or 100ng/ml LPS (Fig. 2B). As shown in Fig. 2C, the production of LPS-stimulated MIP-2 was also upregulated, but the effect was ob-served only at the higher dose (100 ng/ml) of LPS.In contrast with their influence on TNF-a and MIP-2 production,
neither imipramine, GW4869, nor ASMase- or NSMase-specificsiRNA significantly affected NO accumulation in the culture
medium of cells stimulated with 1 or 100 ng/ml LPS (Figs. 1D,2E). Silencing of ASMase and NSMase expression did not changethe production of the chemokine RANTES either (Fig. 2F). Itshould be noted that both NO and RANTES are generated down-stream of a MyD88-independent TLR4 signaling pathway (38–41).Notably, there was a striking difference in the dose-dependence
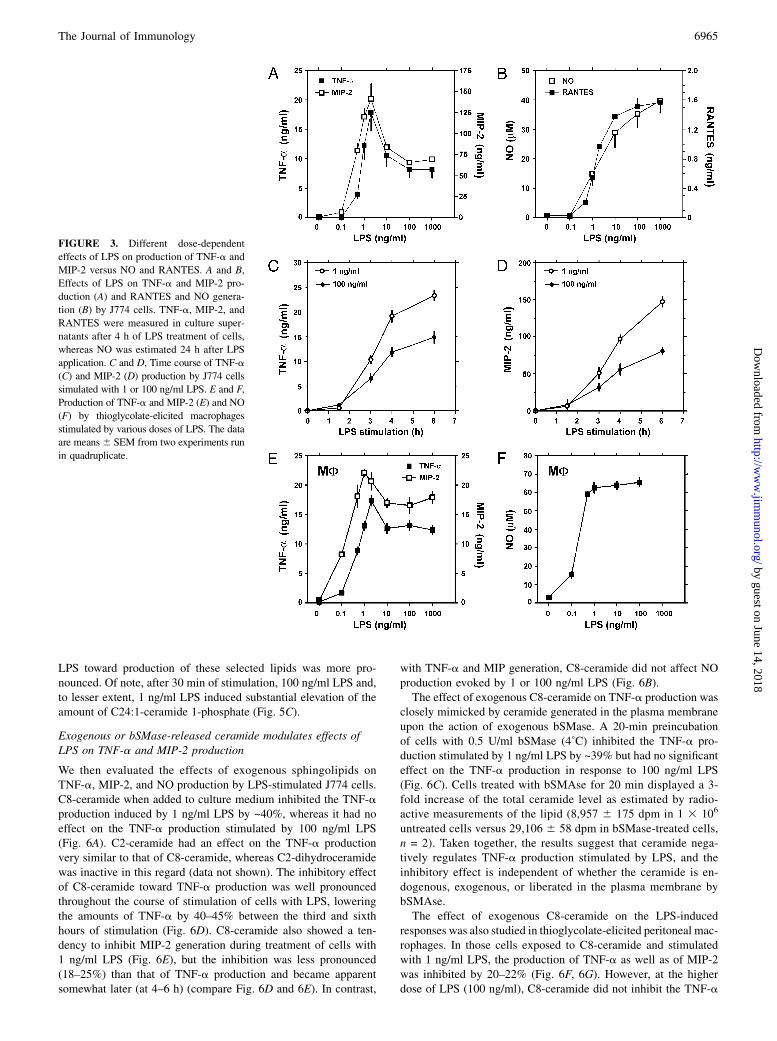
of the LPS effects on TNF-a and MIP-2 versus NO and RANTESproduction (Fig. 3A, 3B). Maximal NO and RANTES productionwas induced by LPS at ∼100–1000 ng/ml (Fig. 3B). In contrast,
FIGURE 1. Activity of ASMase and NSMase affects TNF-a and MIP-2 production stimulated by LPS. A, Sphingomyelinase activity detected at pH 5 in
whole lysates of J774 cells preincubated with 100 mM imipramine or 14 mM GW4869 (80 min, 37˚C). B, Activity of sphingomyelinase detected at pH 7.4
in lysates of J774 cells preincubated with 100 mM imipramine or 14 mM GW4869 or both drugs (80 min, 37˚C). C, Imipramine (100 mM), GW4869 (14
mM), and their combination enhance TNF-a production stimulated by 100 ng/ml LPS but not by 1 ng/ml LPS in J774 cells. D, Inhibition of ASMase with
100 mM imipramine and of NSMase with 14 mMGW4869 does not affect NO production stimulated by LPS in J774 cells. The data are means6 SEM from
four to six experiments, each performed in quadruplicate. E and F, Thioglycolate-elicited peritoneal macrophages preincubated with 100 mM imipramine or
14 mM GW4869 and examined for production of TNF-a (E) or MIP-2 (F) after stimulation with 1–1000 ng/ml LPS. Results are means 6 SEM from three
experiments, each performed in quadruplicate. ★Significantly different from controls at p , 0.05; ★★significantly different from both imipramine and
GW4869 groups at p , 0.05.
The Journal of Immunology 6963
by guest on June 14, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
LPS concentration as low as 1–2 ng/ml was optimal to inducemaximal TNF-a and MIP-2 release, and higher LPS concen-trations were less effective (Fig. 3A). We analyzed the time courseof TNF-a and MIP-2 production stimulated by 1 or 100 ng/mlLPS to examine whether the low level of TNF-a and MIP-2detected at 100 ng/ml LPS was not a consequence of various ki-netics of cytokine secretion stimulated by 100 ng/ml and by 1 ng/ml LPS. However, at all time points tested (3–6 h), the amounts ofTNF-a and MIP-2 induced by 100 ng/ml LPS were lower thanthose induced by 1 ng/ml LPS (Fig. 3C, 3D). Similar reduction ofTNF-a and MIP-2 release at 100 ng/ml in comparison with that of1 ng/ml LPS was found in thioglycolate-elicited peritoneal mac-rophages (Fig. 3E). In contrast, NO generation was induced byLPS in macrophages in a dose-dependent manner, reaching aplateau at 1 ng/ml LPS (Fig. 3F).
LPS stimulates ceramide generation
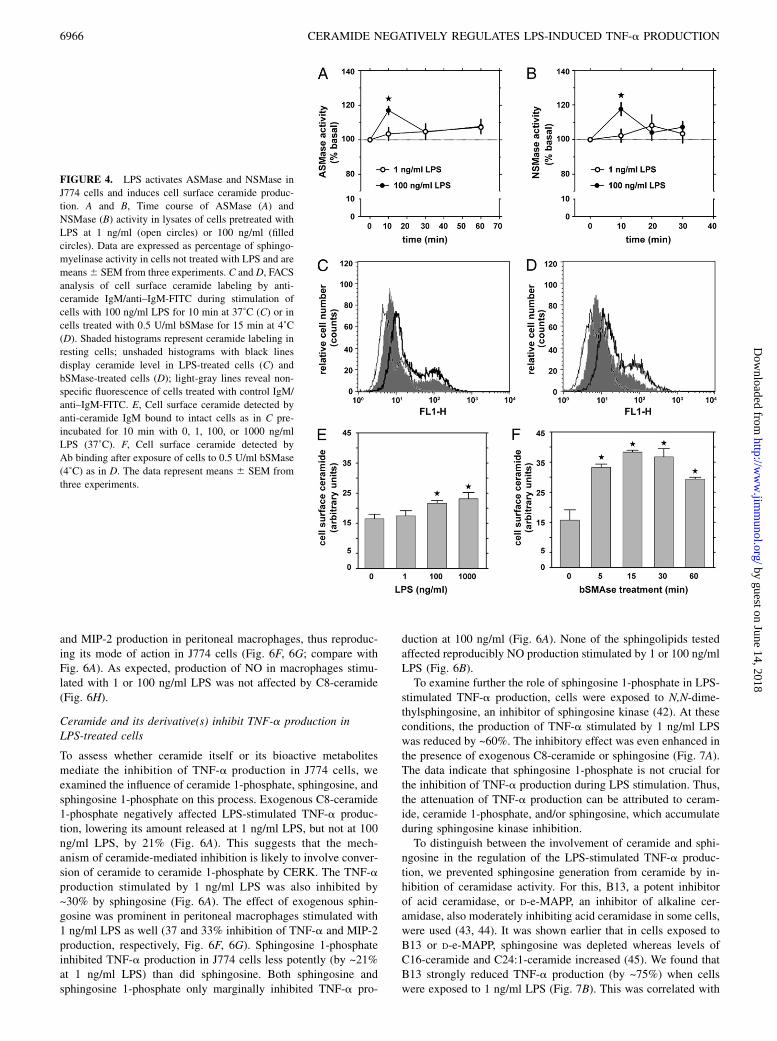
As interference with the activity of sphingomyelinases affected theTNF-a and MIP-2 production induced by LPS, the time course ofsphingomyelinase activation and ceramide generation was ana-lyzed. LPS at 100 ng/ml evoked a 20% increase of both ASMaseand NSMase activity in cells at the 10th minute of stimulation(Fig. 4A, 4B). In the stimulated cells, an increase of cell surfaceceramide level was shown using the binding of anti-ceramide Abto intact cells followed by flow cytometry analysis. Fig. 4Cshows a typical histogram from FACS experiments obtainedfor cells treated with 100 ng/ml LPS for 10 min. The rise ofthe cell surface ceramide level was in the range from 6 (1 ng/mlLPS) to 40% (1000 ng/ml LPS) relative to that in nonstimulated
cells (Fig. 4E). The specificity of the ceramide detection wasconfirmed by an analysis of the surface ceramide level in cellsexposed to bacterial sphingomyelinase. Treatment of the cellswith bSMAse caused a transient elevation of cell surface cer-amide detected by the Ab, which peaked at 15 min of treatmentreaching over twice the amount of ceramide found in untreatedcells (Fig. 4D, 4F).Stimulation of cells with LPS also increased the total mass of
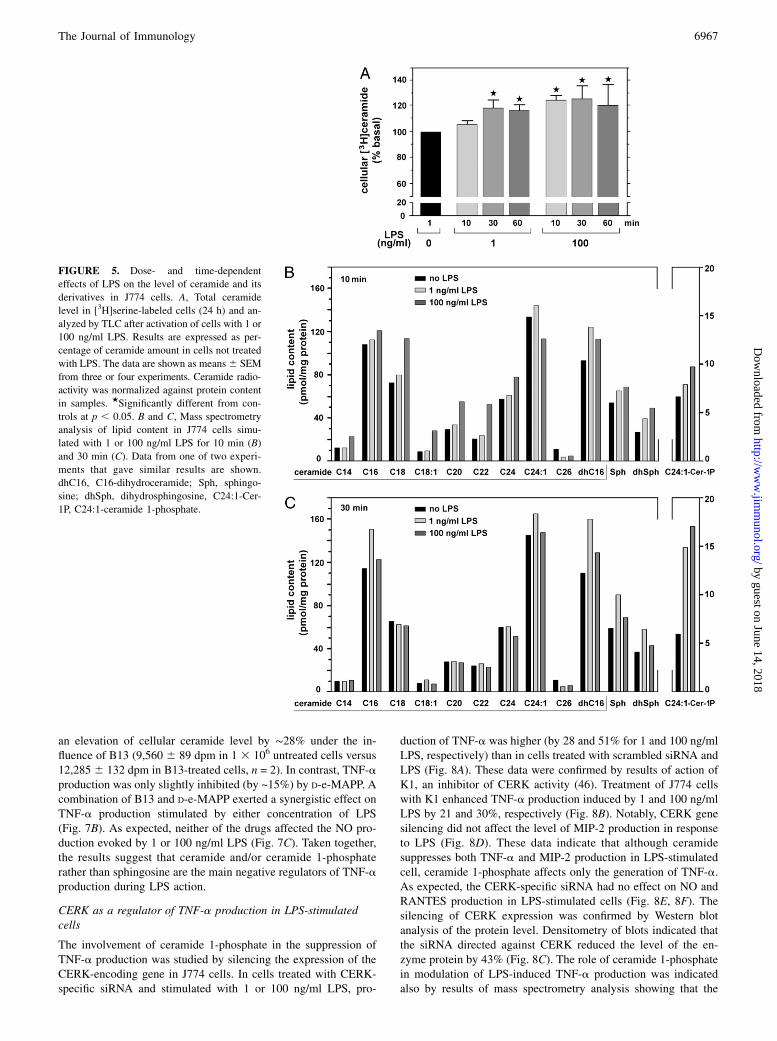
ceramide. Determination of bulk [3H]ceramide revealed that in cellsexposed to 1 or 100 ng/ml LPS for 30 min, the total ceramide levelincreased by 18–27%. At 100 ng/ml LPS, the elevation of ceramidelevel was seen already at 10 min of cell stimulation (Fig. 5A). Theceramide generation induced by LPS was sensitive to sphingo-myelinase inhibitors, with the strongest inhibitory effect exertedby a combination of imipramine and GW4869 (not shown). A moredetailed analysis of ceramide species performed by HPLC andtandem mass spectrometry showed that 10-min stimulation of cellswith 100 ng/ml LPS induced an elevation of the amounts of al-most all ceramide species including C14-, C16-, C18-, C18:1-,C20-, C22-, and C24-ceramide. An increase of the level of C24:1-ceramide 1-phosphate, sphingosine, and dihydrosphingosine andC16-dihydroceramide was also observed indicating that 100 ng/mlLPS affected various aspects of ceramide metabolism (Fig. 5B).At the same time, 1 ng/ml LPS only marginally upregulated pro-duction of the majority of the ceramide species with exceptionfor C24:1-ceramide and C16-dihydroceramide (Fig. 5B). After30 min of treatment of cells with 100 ng/ml LPS, C16- and C24:1-ceramide, C16-dihydroceramide, sphingosine, and dihydrosphi-ngosine were increased; however, the stimulatory effect of 1 ng/ml
FIGURE 2. Silencing of ASMase and NSMase gene expression upregulates production of TNF-a and MIP-2 in J774 cells. A and D, Sphingomyelinase
activity measured at pH 5 (A) or pH 7.4 (D) in unstimulated cells after 48 h of silencing of expression of ASMase (A) and NSMase gene (D). Data are
expressed as percentage of values obtained for cells treated with scrambled siRNA and are means6 SEM from three independent experiments. B, C, E, and
F, Transfection of cells with ASMase-specific or NSMase-specific siRNA enhances TNF-a production stimulated by 1 and 100 ng/ml LPS (B) and MIP-2
production at 100 ng/ml LPS (C) but does not affect generation of NO (E) or RANTES (F). The data are means 6 SEM from three experiments run in
quadruplicate. ★Significantly different from controls at p , 0.05.
6964 CERAMIDE NEGATIVELY REGULATES LPS-INDUCED TNF-a PRODUCTION
by guest on June 14, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
LPS toward production of these selected lipids was more pro-nounced. Of note, after 30 min of stimulation, 100 ng/ml LPS and,to lesser extent, 1 ng/ml LPS induced substantial elevation of theamount of C24:1-ceramide 1-phosphate (Fig. 5C).
Exogenous or bSMase-released ceramide modulates effects ofLPS on TNF-a and MIP-2 production
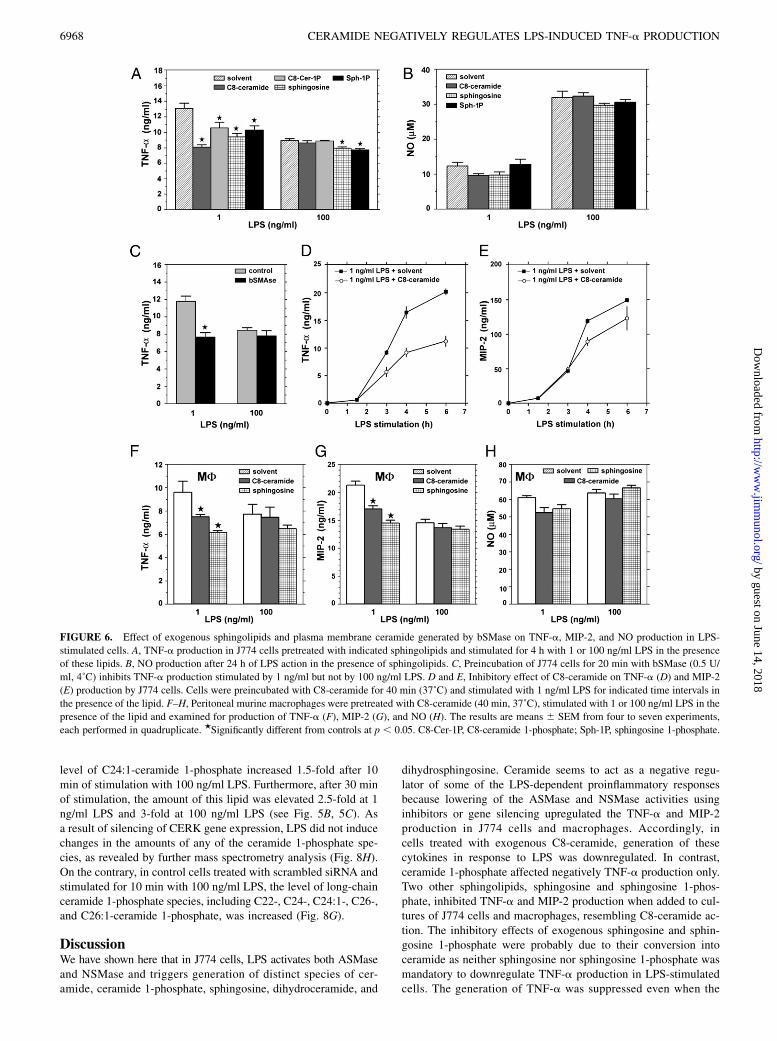
We then evaluated the effects of exogenous sphingolipids onTNF-a, MIP-2, and NO production by LPS-stimulated J774 cells.C8-ceramide when added to culture medium inhibited the TNF-aproduction induced by 1 ng/ml LPS by ~40%, whereas it had noeffect on the TNF-a production stimulated by 100 ng/ml LPS(Fig. 6A). C2-ceramide had an effect on the TNF-a productionvery similar to that of C8-ceramide, whereas C2-dihydroceramidewas inactive in this regard (data not shown). The inhibitory effectof C8-ceramide toward TNF-a production was well pronouncedthroughout the course of stimulation of cells with LPS, loweringthe amounts of TNF-a by 40–45% between the third and sixthhours of stimulation (Fig. 6D). C8-ceramide also showed a ten-dency to inhibit MIP-2 generation during treatment of cells with1 ng/ml LPS (Fig. 6E), but the inhibition was less pronounced(18–25%) than that of TNF-a production and became apparentsomewhat later (at 4–6 h) (compare Fig. 6D and 6E). In contrast,
with TNF-a and MIP generation, C8-ceramide did not affect NOproduction evoked by 1 or 100 ng/ml LPS (Fig. 6B).The effect of exogenous C8-ceramide on TNF-a production was
closely mimicked by ceramide generated in the plasma membraneupon the action of exogenous bSMase. A 20-min preincubationof cells with 0.5 U/ml bSMase (4˚C) inhibited the TNF-a pro-duction stimulated by 1 ng/ml LPS by ~39% but had no significanteffect on the TNF-a production in response to 100 ng/ml LPS(Fig. 6C). Cells treated with bSMAse for 20 min displayed a 3-fold increase of the total ceramide level as estimated by radio-active measurements of the lipid (8,957 6 175 dpm in 1 3 106
untreated cells versus 29,106 6 58 dpm in bSMase-treated cells,n = 2). Taken together, the results suggest that ceramide nega-tively regulates TNF-a production stimulated by LPS, and theinhibitory effect is independent of whether the ceramide is en-dogenous, exogenous, or liberated in the plasma membrane bybSMAse.The effect of exogenous C8-ceramide on the LPS-induced
responses was also studied in thioglycolate-elicited peritoneal mac-rophages. In those cells exposed to C8-ceramide and stimulatedwith 1 ng/ml LPS, the production of TNF-a as well as of MIP-2was inhibited by 20–22% (Fig. 6F, 6G). However, at the higherdose of LPS (100 ng/ml), C8-ceramide did not inhibit the TNF-a
FIGURE 3. Different dose-dependent
effects of LPS on production of TNF-a and
MIP-2 versus NO and RANTES. A and B,
Effects of LPS on TNF-a and MIP-2 pro-
duction (A) and RANTES and NO genera-
tion (B) by J774 cells. TNF-a, MIP-2, and
RANTES were measured in culture super-
natants after 4 h of LPS treatment of cells,
whereas NO was estimated 24 h after LPS
application. C and D, Time course of TNF-a
(C) and MIP-2 (D) production by J774 cells
simulated with 1 or 100 ng/ml LPS. E and F,
Production of TNF-a and MIP-2 (E) and NO
(F) by thioglycolate-elicited macrophages
stimulated by various doses of LPS. The data
are means6 SEM from two experiments run
in quadruplicate.
The Journal of Immunology 6965
by guest on June 14, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
and MIP-2 production in peritoneal macrophages, thus reproduc-ing its mode of action in J774 cells (Fig. 6F, 6G; compare withFig. 6A). As expected, production of NO in macrophages stimu-lated with 1 or 100 ng/ml LPS was not affected by C8-ceramide(Fig. 6H).
Ceramide and its derivative(s) inhibit TNF-a production inLPS-treated cells
To assess whether ceramide itself or its bioactive metabolitesmediate the inhibition of TNF-a production in J774 cells, weexamined the influence of ceramide 1-phosphate, sphingosine, andsphingosine 1-phosphate on this process. Exogenous C8-ceramide1-phosphate negatively affected LPS-stimulated TNF-a produc-tion, lowering its amount released at 1 ng/ml LPS, but not at 100ng/ml LPS, by 21% (Fig. 6A). This suggests that the mech-anism of ceramide-mediated inhibition is likely to involve conver-sion of ceramide to ceramide 1-phosphate by CERK. The TNF-aproduction stimulated by 1 ng/ml LPS was also inhibited by~30% by sphingosine (Fig. 6A). The effect of exogenous sphin-gosine was prominent in peritoneal macrophages stimulated with1 ng/ml LPS as well (37 and 33% inhibition of TNF-a and MIP-2production, respectively, Fig. 6F, 6G). Sphingosine 1-phosphateinhibited TNF-a production in J774 cells less potently (by ~21%at 1 ng/ml LPS) than did sphingosine. Both sphingosine andsphingosine 1-phosphate only marginally inhibited TNF-a pro-
duction at 100 ng/ml (Fig. 6A). None of the sphingolipids testedaffected reproducibly NO production stimulated by 1 or 100 ng/mlLPS (Fig. 6B).To examine further the role of sphingosine 1-phosphate in LPS-
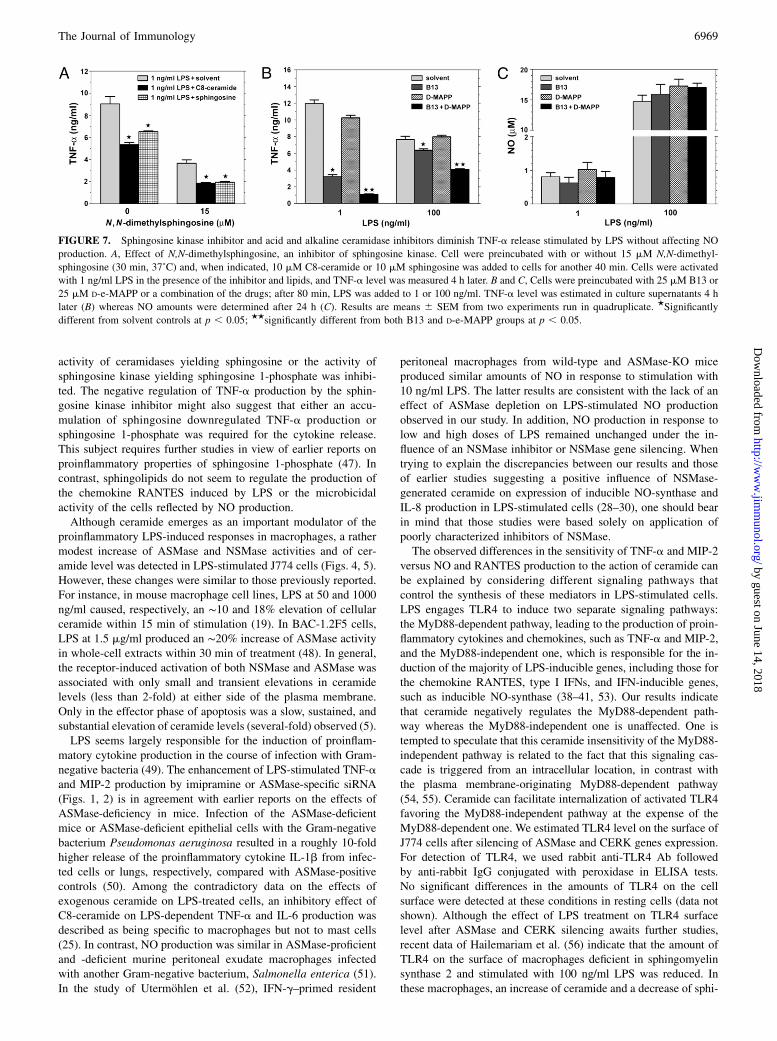
stimulated TNF-a production, cells were exposed to N,N-dime-thylsphingosine, an inhibitor of sphingosine kinase (42). At theseconditions, the production of TNF-a stimulated by 1 ng/ml LPSwas reduced by ~60%. The inhibitory effect was even enhanced inthe presence of exogenous C8-ceramide or sphingosine (Fig. 7A).The data indicate that sphingosine 1-phosphate is not crucial forthe inhibition of TNF-a production during LPS stimulation. Thus,the attenuation of TNF-a production can be attributed to ceram-ide, ceramide 1-phosphate, and/or sphingosine, which accumulateduring sphingosine kinase inhibition.To distinguish between the involvement of ceramide and sphi-
ngosine in the regulation of the LPS-stimulated TNF-a produc-tion, we prevented sphingosine generation from ceramide by in-hibition of ceramidase activity. For this, B13, a potent inhibitorof acid ceramidase, or D-e-MAPP, an inhibitor of alkaline cer-amidase, also moderately inhibiting acid ceramidase in some cells,were used (43, 44). It was shown earlier that in cells exposed toB13 or D-e-MAPP, sphingosine was depleted whereas levels ofC16-ceramide and C24:1-ceramide increased (45). We found thatB13 strongly reduced TNF-a production (by ~75%) when cellswere exposed to 1 ng/ml LPS (Fig. 7B). This was correlated with
FIGURE 4. LPS activates ASMase and NSMase in
J774 cells and induces cell surface ceramide produc-
tion. A and B, Time course of ASMase (A) and
NSMase (B) activity in lysates of cells pretreated with
LPS at 1 ng/ml (open circles) or 100 ng/ml (filled
circles). Data are expressed as percentage of sphingo-
myelinase activity in cells not treated with LPS and are
means6 SEM from three experiments. C and D, FACS
analysis of cell surface ceramide labeling by anti-
ceramide IgM/anti–IgM-FITC during stimulation of
cells with 100 ng/ml LPS for 10 min at 37˚C (C) or in
cells treated with 0.5 U/ml bSMase for 15 min at 4˚C
(D). Shaded histograms represent ceramide labeling in
resting cells; unshaded histograms with black lines
display ceramide level in LPS-treated cells (C) and
bSMase-treated cells (D); light-gray lines reveal non-
specific fluorescence of cells treated with control IgM/
anti–IgM-FITC. E, Cell surface ceramide detected by
anti-ceramide IgM bound to intact cells as in C pre-
incubated for 10 min with 0, 1, 100, or 1000 ng/ml
LPS (37˚C). F, Cell surface ceramide detected by
Ab binding after exposure of cells to 0.5 U/ml bSMase
(4˚C) as in D. The data represent means 6 SEM from
three experiments.
6966 CERAMIDE NEGATIVELY REGULATES LPS-INDUCED TNF-a PRODUCTION
by guest on June 14, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
an elevation of cellular ceramide level by ∼28% under the in-fluence of B13 (9,560 6 89 dpm in 1 3 106 untreated cells versus12,2856 132 dpm in B13-treated cells, n = 2). In contrast, TNF-aproduction was only slightly inhibited (by ~15%) by D-e-MAPP. Acombination of B13 and D-e-MAPP exerted a synergistic effect onTNF-a production stimulated by either concentration of LPS(Fig. 7B). As expected, neither of the drugs affected the NO pro-duction evoked by 1 or 100 ng/ml LPS (Fig. 7C). Taken together,the results suggest that ceramide and/or ceramide 1-phosphaterather than sphingosine are the main negative regulators of TNF-aproduction during LPS action.
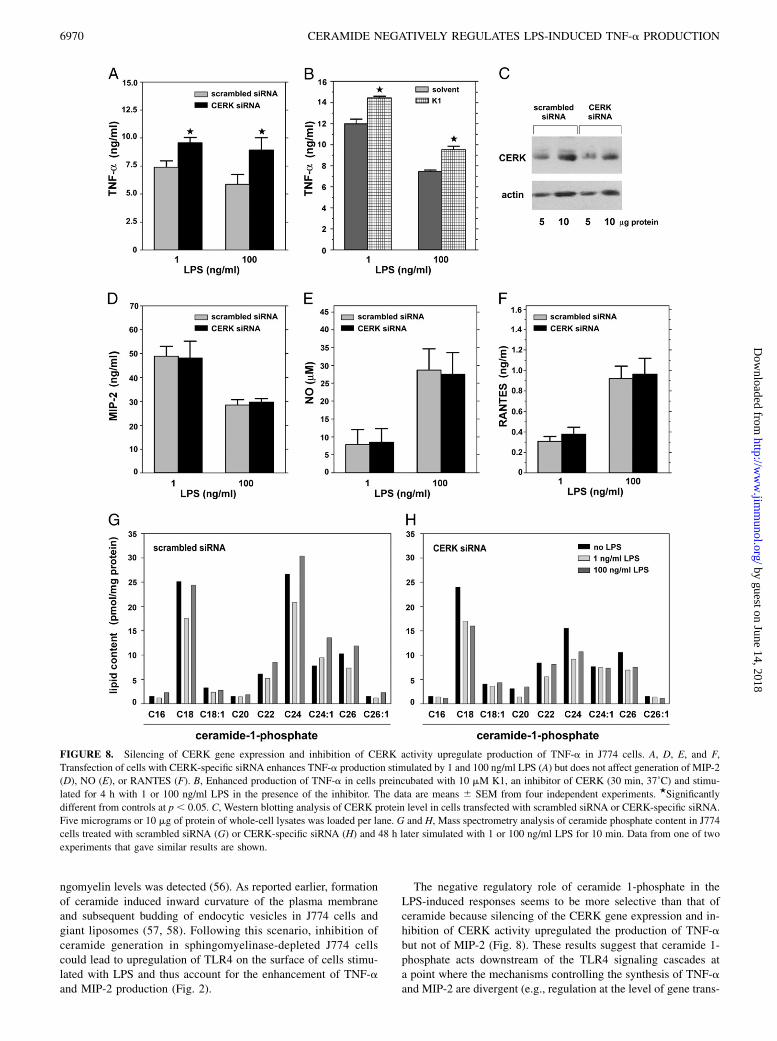
CERK as a regulator of TNF-a production in LPS-stimulatedcells
The involvement of ceramide 1-phosphate in the suppression ofTNF-a production was studied by silencing the expression of theCERK-encoding gene in J774 cells. In cells treated with CERK-specific siRNA and stimulated with 1 or 100 ng/ml LPS, pro-
duction of TNF-a was higher (by 28 and 51% for 1 and 100 ng/mlLPS, respectively) than in cells treated with scrambled siRNA andLPS (Fig. 8A). These data were confirmed by results of action ofK1, an inhibitor of CERK activity (46). Treatment of J774 cellswith K1 enhanced TNF-a production induced by 1 and 100 ng/mlLPS by 21 and 30%, respectively (Fig. 8B). Notably, CERK genesilencing did not affect the level of MIP-2 production in responseto LPS (Fig. 8D). These data indicate that although ceramidesuppresses both TNF-a and MIP-2 production in LPS-stimulatedcell, ceramide 1-phosphate affects only the generation of TNF-a.As expected, the CERK-specific siRNA had no effect on NO andRANTES production in LPS-stimulated cells (Fig. 8E, 8F). Thesilencing of CERK expression was confirmed by Western blotanalysis of the protein level. Densitometry of blots indicated thatthe siRNA directed against CERK reduced the level of the en-zyme protein by 43% (Fig. 8C). The role of ceramide 1-phosphatein modulation of LPS-induced TNF-a production was indicatedalso by results of mass spectrometry analysis showing that the
FIGURE 5. Dose- and time-dependent
effects of LPS on the level of ceramide and its
derivatives in J774 cells. A, Total ceramide
level in [3H]serine-labeled cells (24 h) and an-
alyzed by TLC after activation of cells with 1 or
100 ng/ml LPS. Results are expressed as per-
centage of ceramide amount in cells not treated
with LPS. The data are shown as means6 SEM
from three or four experiments. Ceramide radio-
activity was normalized against protein content
in samples. ★Significantly different from con-
trols at p , 0.05. B and C, Mass spectrometry
analysis of lipid content in J774 cells simu-
lated with 1 or 100 ng/ml LPS for 10 min (B)
and 30 min (C). Data from one of two experi-
ments that gave similar results are shown.
dhC16, C16-dihydroceramide; Sph, sphingo-
sine; dhSph, dihydrosphingosine, C24:1-Cer-
1P, C24:1-ceramide 1-phosphate.
The Journal of Immunology 6967
by guest on June 14, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
level of C24:1-ceramide 1-phosphate increased 1.5-fold after 10min of stimulation with 100 ng/ml LPS. Furthermore, after 30 minof stimulation, the amount of this lipid was elevated 2.5-fold at 1ng/ml LPS and 3-fold at 100 ng/ml LPS (see Fig. 5B, 5C). Asa result of silencing of CERK gene expression, LPS did not inducechanges in the amounts of any of the ceramide 1-phosphate spe-cies, as revealed by further mass spectrometry analysis (Fig. 8H).On the contrary, in control cells treated with scrambled siRNA andstimulated for 10 min with 100 ng/ml LPS, the level of long-chainceramide 1-phosphate species, including C22-, C24-, C24:1-, C26-,and C26:1-ceramide 1-phosphate, was increased (Fig. 8G).
DiscussionWe have shown here that in J774 cells, LPS activates both ASMaseand NSMase and triggers generation of distinct species of cer-amide, ceramide 1-phosphate, sphingosine, dihydroceramide, and
dihydrosphingosine. Ceramide seems to act as a negative regu-lator of some of the LPS-dependent proinflammatory responsesbecause lowering of the ASMase and NSMase activities usinginhibitors or gene silencing upregulated the TNF-a and MIP-2production in J774 cells and macrophages. Accordingly, incells treated with exogenous C8-ceramide, generation of thesecytokines in response to LPS was downregulated. In contrast,ceramide 1-phosphate affected negatively TNF-a production only.Two other sphingolipids, sphingosine and sphingosine 1-phos-phate, inhibited TNF-a and MIP-2 production when added to cul-tures of J774 cells and macrophages, resembling C8-ceramide ac-tion. The inhibitory effects of exogenous sphingosine and sphin-gosine 1-phosphate were probably due to their conversion intoceramide as neither sphingosine nor sphingosine 1-phosphate wasmandatory to downregulate TNF-a production in LPS-stimulatedcells. The generation of TNF-a was suppressed even when the
FIGURE 6. Effect of exogenous sphingolipids and plasma membrane ceramide generated by bSMase on TNF-a, MIP-2, and NO production in LPS-
stimulated cells. A, TNF-a production in J774 cells pretreated with indicated sphingolipids and stimulated for 4 h with 1 or 100 ng/ml LPS in the presence
of these lipids. B, NO production after 24 h of LPS action in the presence of sphingolipids. C, Preincubation of J774 cells for 20 min with bSMase (0.5 U/
ml, 4˚C) inhibits TNF-a production stimulated by 1 ng/ml but not by 100 ng/ml LPS. D and E, Inhibitory effect of C8-ceramide on TNF-a (D) and MIP-2
(E) production by J774 cells. Cells were preincubated with C8-ceramide for 40 min (37˚C) and stimulated with 1 ng/ml LPS for indicated time intervals in
the presence of the lipid. F–H, Peritoneal murine macrophages were pretreated with C8-ceramide (40 min, 37˚C), stimulated with 1 or 100 ng/ml LPS in the
presence of the lipid and examined for production of TNF-a (F), MIP-2 (G), and NO (H). The results are means 6 SEM from four to seven experiments,
each performed in quadruplicate. ★Significantly different from controls at p , 0.05. C8-Cer-1P, C8-ceramide 1-phosphate; Sph-1P, sphingosine 1-phosphate.
6968 CERAMIDE NEGATIVELY REGULATES LPS-INDUCED TNF-a PRODUCTION
by guest on June 14, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
activity of ceramidases yielding sphingosine or the activity ofsphingosine kinase yielding sphingosine 1-phosphate was inhibi-ted. The negative regulation of TNF-a production by the sphin-gosine kinase inhibitor might also suggest that either an accu-mulation of sphingosine downregulated TNF-a production orsphingosine 1-phosphate was required for the cytokine release.This subject requires further studies in view of earlier reports onproinflammatory properties of sphingosine 1-phosphate (47). Incontrast, sphingolipids do not seem to regulate the production ofthe chemokine RANTES induced by LPS or the microbicidalactivity of the cells reflected by NO production.Although ceramide emerges as an important modulator of the
proinflammatory LPS-induced responses in macrophages, a rathermodest increase of ASMase and NSMase activities and of cer-amide level was detected in LPS-stimulated J774 cells (Figs. 4, 5).However, these changes were similar to those previously reported.For instance, in mouse macrophage cell lines, LPS at 50 and 1000ng/ml caused, respectively, an ∼10 and 18% elevation of cellularceramide within 15 min of stimulation (19). In BAC-1.2F5 cells,LPS at 1.5 mg/ml produced an ∼20% increase of ASMase activityin whole-cell extracts within 30 min of treatment (48). In general,the receptor-induced activation of both NSMase and ASMase wasassociated with only small and transient elevations in ceramidelevels (less than 2-fold) at either side of the plasma membrane.Only in the effector phase of apoptosis was a slow, sustained, andsubstantial elevation of ceramide levels (several-fold) observed (5).LPS seems largely responsible for the induction of proinflam-
matory cytokine production in the course of infection with Gram-negative bacteria (49). The enhancement of LPS-stimulated TNF-aand MIP-2 production by imipramine or ASMase-specific siRNA(Figs. 1, 2) is in agreement with earlier reports on the effects ofASMase-deficiency in mice. Infection of the ASMase-deficientmice or ASMase-deficient epithelial cells with the Gram-negativebacterium Pseudomonas aeruginosa resulted in a roughly 10-foldhigher release of the proinflammatory cytokine IL-1b from infec-ted cells or lungs, respectively, compared with ASMase-positivecontrols (50). Among the contradictory data on the effects ofexogenous ceramide on LPS-treated cells, an inhibitory effect ofC8-ceramide on LPS-dependent TNF-a and IL-6 production wasdescribed as being specific to macrophages but not to mast cells(25). In contrast, NO production was similar in ASMase-proficientand -deficient murine peritoneal exudate macrophages infectedwith another Gram-negative bacterium, Salmonella enterica (51).In the study of Utermohlen et al. (52), IFN-g–primed resident
peritoneal macrophages from wild-type and ASMase-KO miceproduced similar amounts of NO in response to stimulation with10 ng/ml LPS. The latter results are consistent with the lack of aneffect of ASMase depletion on LPS-stimulated NO productionobserved in our study. In addition, NO production in response tolow and high doses of LPS remained unchanged under the in-fluence of an NSMase inhibitor or NSMase gene silencing. Whentrying to explain the discrepancies between our results and thoseof earlier studies suggesting a positive influence of NSMase-generated ceramide on expression of inducible NO-synthase andIL-8 production in LPS-stimulated cells (28–30), one should bearin mind that those studies were based solely on application ofpoorly characterized inhibitors of NSMase.The observed differences in the sensitivity of TNF-a and MIP-2
versus NO and RANTES production to the action of ceramide canbe explained by considering different signaling pathways thatcontrol the synthesis of these mediators in LPS-stimulated cells.LPS engages TLR4 to induce two separate signaling pathways:the MyD88-dependent pathway, leading to the production of proin-flammatory cytokines and chemokines, such as TNF-a and MIP-2,and the MyD88-independent one, which is responsible for the in-duction of the majority of LPS-inducible genes, including those forthe chemokine RANTES, type I IFNs, and IFN-inducible genes,such as inducible NO-synthase (38–41, 53). Our results indicatethat ceramide negatively regulates the MyD88-dependent path-way whereas the MyD88-independent one is unaffected. One istempted to speculate that this ceramide insensitivity of the MyD88-independent pathway is related to the fact that this signaling cas-cade is triggered from an intracellular location, in contrast withthe plasma membrane-originating MyD88-dependent pathway(54, 55). Ceramide can facilitate internalization of activated TLR4favoring the MyD88-independent pathway at the expense of theMyD88-dependent one. We estimated TLR4 level on the surface ofJ774 cells after silencing of ASMase and CERK genes expression.For detection of TLR4, we used rabbit anti-TLR4 Ab followedby anti-rabbit IgG conjugated with peroxidase in ELISA tests.No significant differences in the amounts of TLR4 on the cellsurface were detected at these conditions in resting cells (data notshown). Although the effect of LPS treatment on TLR4 surfacelevel after ASMase and CERK silencing awaits further studies,recent data of Hailemariam et al. (56) indicate that the amount ofTLR4 on the surface of macrophages deficient in sphingomyelinsynthase 2 and stimulated with 100 ng/ml LPS was reduced. Inthese macrophages, an increase of ceramide and a decrease of sphi-
FIGURE 7. Sphingosine kinase inhibitor and acid and alkaline ceramidase inhibitors diminish TNF-a release stimulated by LPS without affecting NO
production. A, Effect of N,N-dimethylsphingosine, an inhibitor of sphingosine kinase. Cell were preincubated with or without 15 mM N,N-dimethyl-
sphingosine (30 min, 37˚C) and, when indicated, 10 mM C8-ceramide or 10 mM sphingosine was added to cells for another 40 min. Cells were activated
with 1 ng/ml LPS in the presence of the inhibitor and lipids, and TNF-a level was measured 4 h later. B and C, Cells were preincubated with 25 mM B13 or
25 mM D-e-MAPP or a combination of the drugs; after 80 min, LPS was added to 1 or 100 ng/ml. TNF-a level was estimated in culture supernatants 4 h
later (B) whereas NO amounts were determined after 24 h (C). Results are means 6 SEM from two experiments run in quadruplicate. ★Significantly
different from solvent controls at p , 0.05; ★★significantly different from both B13 and D-e-MAPP groups at p , 0.05.
The Journal of Immunology 6969
by guest on June 14, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
ngomyelin levels was detected (56). As reported earlier, formationof ceramide induced inward curvature of the plasma membraneand subsequent budding of endocytic vesicles in J774 cells andgiant liposomes (57, 58). Following this scenario, inhibition ofceramide generation in sphingomyelinase-depleted J774 cellscould lead to upregulation of TLR4 on the surface of cells stimu-lated with LPS and thus account for the enhancement of TNF-aand MIP-2 production (Fig. 2).
The negative regulatory role of ceramide 1-phosphate in theLPS-induced responses seems to be more selective than that ofceramide because silencing of the CERK gene expression and in-hibition of CERK activity upregulated the production of TNF-abut not of MIP-2 (Fig. 8). These results suggest that ceramide 1-phosphate acts downstream of the TLR4 signaling cascades ata point where the mechanisms controlling the synthesis of TNF-aand MIP-2 are divergent (e.g., regulation at the level of gene trans-
FIGURE 8. Silencing of CERK gene expression and inhibition of CERK activity upregulate production of TNF-a in J774 cells. A, D, E, and F,
Transfection of cells with CERK-specific siRNA enhances TNF-a production stimulated by 1 and 100 ng/ml LPS (A) but does not affect generation of MIP-2
(D), NO (E), or RANTES (F). B, Enhanced production of TNF-a in cells preincubated with 10 mM K1, an inhibitor of CERK (30 min, 37˚C) and stimu-
lated for 4 h with 1 or 100 ng/ml LPS in the presence of the inhibitor. The data are means 6 SEM from four independent experiments. ★Significantly
different from controls at p, 0.05. C, Western blotting analysis of CERK protein level in cells transfected with scrambled siRNA or CERK-specific siRNA.
Five micrograms or 10 mg of protein of whole-cell lysates was loaded per lane. G and H, Mass spectrometry analysis of ceramide phosphate content in J774
cells treated with scrambled siRNA (G) or CERK-specific siRNA (H) and 48 h later simulated with 1 or 100 ng/ml LPS for 10 min. Data from one of two
experiments that gave similar results are shown.
6970 CERAMIDE NEGATIVELY REGULATES LPS-INDUCED TNF-a PRODUCTION
by guest on June 14, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

cription). The TNF-a gene promoter is regulated cooperativelyby several LPS-induced transcription factors with NF-kB as themain activator and CREB as a repressor (59–61). It was shownrecently that exogenous ceramide phospho-ceramide-analogue-1(PCERA-1), induced elevation of cAMP in cells leading, in turn,to a downregulation of TNF-amRNA and protein (61). It is temptingto speculate that endogenous ceramide 1-phosphate generated byCERK suppresses production of TNF-a in LPS-stimulated cells in asimilar manner acting through CREB. In the case of MIP-2, CREBseems to positively regulate gene transcription (62), but its possibleattenuation in CERK-depleted cells does not limit the MIP-2 syn-thesis. At present, this thesis is speculative because it predicts thatendogenous ceramide 1-phosphate affects CREB activity, which isalso regulated by exogenous PCERA-1 after its binding to a Gs
protein-coupled receptor (63). In contrast, there are data indicat-ing that exogenous ceramide 1-phosphate binds to a Gi protein-coupled receptor different from the PCERA-1 receptor (64, 65).Our data based on depletion of CERK activity demonstrate thatceramide 1-phosphate is a potent suppressor of LPS-inducedTNF-a production validating further studies on the mechanismof its action in macrophages.Besides its positive role in host defense, TNF-a is also the
primary mediator of LPS-induced septic shock (12, 27, 66, 67).Thus, a plausible physiological role of the ceramide-mediated (andpossibly also ceramide 1-phosphate–mediated) negative regulationof proinflammatory activation of macrophages could be to preventendotoxic shock during infection with Gram-negative bacteria.Such a role is supported by the observation that ASMase-deficientmice exhibited strongly increased lethality as a result of acute in-fection with P. aeruginosa that was prevented by neutralization ofthe proinflammatory cytokine IL-1b with a specific Ab (50). Ac-tivation of the sphingomyelin-ceramide pathway by LPS releasedfrom destroyed bacteria can curb the severity of the response toinfection. Ceramide could also participate in the termination of theproinflammatory responses to LPS acting downstream of receptorsfor oxidized phospholipids. The inflammatory process is associatedwith gradual accumulation of oxidized phospholipids in tissues(68), and oxidized phospholipids have been demonstrated to exertpotent anti-inflammatory effects in vivo (69). Recently, oxidized 1-palmitoyl-2-arachidonyl-sn-glycero-3-phosphocholine (OxPAPC)and other oxidized phospholipids were found to inhibit LPS-dependent synthesis of TNF-a and IL-8 (26, 69, 70). The in-hibitory action of OxPAPC toward the LPS-induced IL-8 releasewas mediated by ceramide generated by NSMase (26). We alsofound that OxPAPC at 5 mg/ml inhibited the TNF-a productioninduced by 1 ng/ml LPS in J774 cells. The inhibitory effect waspartially reversed by imipramine and fully by GW4869, suggestingactivation of both NSMase and ASMase by OxPAPC (data notshown). Our data support the thesis that the inhibition of TNF-aproduction by oxidized phospholipids may represent a negativefeedback mechanism preventing excessive proinflammatory re-sponses. Oxidized phospholipids can directly bind to LBP/CD14/MD-2 thereby enhancing ceramide production (69, 70). It ispossible that receptors for the oxidized lipids, scavenger receptorsand PGE2 receptor, can also mediate negative regulation of pro-duction of proinflammatory cytokines (26, 71–73).Although ceramide strongly inhibits secretion of proinflam-
matory mediators, such as TNF-a, IL-6, and chemokines (Refs. 25,26, and 74 and this study), it does not seem to impair the NO-dependent microbicidal activity of macrophages (Refs. 51 and 52and this study). These selective anti-inflammatory properties ofceramide could perhaps be exploited therapeutically in treatmentof acute inflammation or chronic inflammatory disorders such asasthma, arthritis, psoriasis, and inflammatory bowel disease. The
feasibility of such an approach is supported by the recently pub-lished study by Sun et al. (74). In a murine in vivo model of LPS-or Staphylococcus aureus-induced corneal inflammation, topicaldelivery of low doses of a liposomal formulation of C6-ceramidestrongly inhibited the inflammation without appreciable cellularapoptosis or reduction in wound-healing responses (74).Note added in proof. During the publication of this article,Rozenova et al. (75) published data showing that ASMase-derivedceramide was a negative regulator of posttranslational processingof TNF-a in macrophages and suppressed TNF-a secretion afterLPS stimulation. Macrophages isolated from ASMase-deficientmice produced severalfold more TNF-a than their wild-typecounterparts in response to LPS. These data correlate with the re-sults presented in our report.
AcknowledgmentsWe thank Lipidomics Shared Resource, Medical University of South Car-
olina for performing sphingolipid analyses and providing ceramidase in-
hibitors.
DisclosuresThe authors have no financial conflicts of interest.
References1. Yang, J., Y. Yu, S. Sun, and P. J. Duerksen-Hughes. 2004. Ceramide and other
sphingolipids in cellular responses. Cell Biochem. Biophys. 40: 323–350.2. van Blitterswijk, W. J. 1998. Hypothesis: ceramide conditionally activates atypical
protein kinases C, Raf-1 and KSR through binding to their cysteine-rich domains.Biochem. J. 331: 679–680.
3. Galadari, S., K. Kishikawa, C. Kamibayashi, M. C. Mumby, and Y. A. Hannun.1998. Purification and characterization of ceramide-activated protein phospha-tases. Biochemistry 37: 11232–11238.
4. Kitatani, K., J. Idkowiak-Baldys, and Y. A. Hannun. 2007. Mechanism of in-hibition of sequestration of protein kinase C a/betaII by ceramide. Roles ofceramide-activated protein phosphatases and phosphorylation/dephosphorylationof protein kinase C a/betaII on threonine 638/641. J. Biol. Chem. 282: 20647–20656.
5. van Blitterswijk, W. J., A. H. van der Luit, R. J. Veldman, M. Verheij, andJ. Borst. 2003. Ceramide: second messenger or modulator of membrane structureand dynamics? Biochem. J. 369: 199–211.
6. Megha, and E. London. 2004. Ceramide selectively displaces cholesterol fromordered lipid domains (rafts): implications for lipid raft structure and function. J.Biol. Chem. 279: 9997210004.
7. Nybond, S., Y. J. Bjorkqvist, B. Ramstedt, and J. P. Slotte. 2005. Acyl chainlength affects ceramide action on sterol/sphingomyelin-rich domains. Biochim.Biophys. Acta 1718: 61–66.
8. Silva, L. C., R. F. M. de Almeida, B. M. Castro, A. Fedorov, and M. Prieto. 2007.Ceramide-domain formation and collapse in lipid rafts: membrane reorgani-zation by an apoptotic lipid. Biophys. J. 92: 502–516.
9. Cremesti, A., F. Paris, H. Grassme, N. Holler, J. Tschopp, Z. Fuks, E. Gulbins,and R. Kolesnick. 2001. Ceramide enables fas to cap and kill. J. Biol. Chem. 276:23954–23961.
10. Grassme, H., V. Jendrossek, J. Bock, A. Riehle, and E. Gulbins. 2002. Ceramide-rich membrane rafts mediate CD40 clustering. J. Immunol. 168: 298–307.
11. Abdel Shakor, A. B., K. Kwiatkowska, and A. Sobota. 2004. Cell surface ceramidegeneration precedes and controls FcgammaRII clustering and phosphorylation inrafts. J. Biol. Chem. 279: 36778–36787.
12. Pfeffer, K., T. Matsuyama, T. M. Kundig, A. Wakeham, K. Kishihara, A. Shahinian,K. Wiegmann, P. S. Ohashi, M. Kronke, and T. W. Mak. 1993. Mice deficient forthe 55 kd tumor necrosis factor receptor are resistant to endotoxic shock, yet suc-cumb to L. monocytogenes infection. Cell 73: 457–467.
13. Perera, P.-Y., S. N. Vogel, G. R. Detore, A. Haziot, and S. M. Goyert. 1997.CD14-dependent and CD14-independent signaling pathways in murine macro-phages from normal and CD14 knockout mice stimulated with lipopolysaccha-ride or taxol. J. Immunol. 158: 4422–4429.
14. Hoshino, K., O. Takeuchi, T. Kawai, H. Sanjo, T. Ogawa, Y. Takeda, K. Takeda,and S. Akira. 1999. Cutting edge: Toll-like receptor 4 (TLR4)-deficient mice arehyporesponsive to lipopolysaccharide: evidence for TLR4 as the Lps geneproduct. J. Immunol. 162: 3749–3752.
15. Moore, K. J., L. P. Andersson, R. R. Ingalls, B. G. Monks, R. Li, M. A. Arnaout,D. T. Golenbock, and M. W. Freeman. 2000. Divergent response to LPS andbacteria in CD14-deficient murine macrophages. J. Immunol. 165: 4272–4280.
16. Jerala, R. 2007. Structural biology of the LPS recognition. Int. J. Med. Micro-biol. 297: 353–363.
17. Lu, Y. C., W. C. Yeh, and P. S. Ohashi. 2008. LPS/TLR4 signal transductionpathway. Cytokine 42: 145–151.
The Journal of Immunology 6971
by guest on June 14, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

18. Medvedev, A. E., J. C. G. Blanco, N. Qureshi, and S. N. Vogel. 1999. Limitedrole of ceramide in lipopolysaccharide-mediated mitogen-activated protein ki-nase activation, transcription factor induction, and cytokine release. J. Biol.Chem. 274: 9342–9350.
19. MacKichan, M. L., and A. L. DeFranco. 1999. Role of ceramide in lipopoly-saccharide (LPS)-induced signaling. LPS increases ceramide rather than actingas a structural homolog. J. Biol. Chem. 274: 1767–1775.
20. Lo, C. J., M. Fu, F. R. Lo, and H. G. Cryer. 1999. Macrophage TNF mRNAexpression induced by LPS is regulated by sphingomyelin metabolites. Shock 11:411–415.
21. Pfeiffer, A., A. Bottcher, E. Orso, M. Kapinsky, P. Nagy, A. Bodnar, I. Spreitzer,G. Liebisch, W. Drobnik, K. Gempel, et al. 2001. Lipopolysaccharide andceramide docking to CD14 provokes ligand-specific receptor clustering in rafts.Eur. J. Immunol. 31: 3153–3164.
22. Fischer, H., P. Ellstrom, K. Ekstrom, L. Gustafsson, M. Gustafsson, andC. Svanborg. 2007. Ceramide as a TLR4 agonist; a putative signalling inter-mediate between sphingolipid receptors for microbial ligands and TLR4. Cell.Microbiol. 9: 1239–1251.
23. Cuschieri, J., E. Bulger, J. Billgrin, I. Garcia, and R. V. Maier. 2007. Acidsphingomyelinase is required for lipid Raft TLR4 complex formation. Surg.Infect. (Larchmt) 8: 91–106.
24. Sakata, A., T. Ochiai, H. Shimeno, S. Hikishima, T. Yokomatsu, S. Shibuya,A. Toda, R. Eyanagi, and S. Soeda. 2007. Acid sphingomyelinase inhibition su-ppresses lipopolysaccharide-mediated release of inflammatory cytokines frommacrophages and protects against disease pathology in dextran sulphate sodium-induced colitis in mice. Immunology 122: 54–64.
25. Chiba, N., A. Masuda, Y. Yoshikai, and T. Matsuguchi. 2007. Ceramide inhibitsLPS-induced production of IL-5, IL-10, and IL-13 from mast cells. J. Cell.Physiol. 213: 126–136.
26. Walton, K. A., B. G. Gugiu, M. Thomas, R. J. Basseri, D. R. Eliav, R. G. Salomon,and J. A. Berliner. 2006. A role for neutral sphingomyelinase activation in theinhibition of LPS action by phospholipid oxidation products. J. Lipid Res. 47:1967–1974.
27. Haimovitz-Friedman, A., C. Cordon-Cardo, S. Bayoumy, M. Garzotto,M. McLoughlin, R. Gallily, C. K. Edwards, III, E. H. Schuchman, Z. Fuks, andR. Kolesnick. 1997. Lipopolysaccharide induces disseminated endothelial apo-ptosis requiring ceramide generation. J. Exp. Med. 186: 1831–1841.
28. Sakata, A., K. Yasuda, T. Ochiai, H. Shimeno, S. Hikishima, T. Yokomatsu,S. Shibuya, and S. Soeda. 2007. Inhibition of lipopolysaccharide-induced releaseof interleukin-8 from intestinal epithelial cells by SMA, a novel inhibitor ofsphingomyelinase and its therapeutic effect on dextran sulphate sodium-inducedcolitis in mice. Cell. Immunol. 245: 24–31.
29. Amtmann, E., W. Baader, and M. Zoller. 2003. Neutral sphingomyelinase in-hibitor C11AG prevents lipopolysaccharide-induced macrophage activation.Drugs Exp. Clin. Res. 29: 5–13.
30. Won, J. S., Y. B. Im, M. Khan, A. K. Singh, and I. Singh. 2004. The role ofneutral sphingomyelinase produced ceramide in lipopolysaccharide-mediatedexpression of inducible nitric oxide synthase. J. Neurochem. 88: 583–593.
31. Jozefowski, S., and L. Kobzik. 2004. Scavenger receptor A mediates H2O2
production and suppression of IL-12 release in murine macrophages. J. Leukoc.Biol. 76: 1066–1074.
32. Hinkovska-Galcheva, V., L. Kjeldsen, P. J. Mansfield, L. A. Boxer, J. A. Shayman,and S. J. Suchard. 1998. Activation of a plasma membrane-associated neu-tral sphingomyelinase and concomitant ceramide accumulation during IgG-dependent phagocytosis in human polymorphonuclear leukocytes. Blood 91:4761–4769.
33. Hojjati, M. R., and X. C. Jiang. 2006. Rapid, specific, and sensitive measure-ments of plasma sphingomyelin and phosphatidylcholine. J. Lipid Res. 47: 673–676.
34. Hanada, K., and M. Nishijima. 2000. Selection of mammalian cell mutants insphingolipid biosynthesis. Methods Enzymol. 312: 304–317.
35. Bielawski, J., Z. M. Szulc, Y. A. Hannun, and A. Bielawska. 2006. Simultaneousquantitative analysis of bioactive sphingolipids by high-performance liquidchromatography-tandem mass spectrometry. Methods 39: 82–91.
36. Hurwitz, R., K. Ferlinz, and K. Sandhoff. 1994. The tricyclic antidepressantdesipramine causes proteolytic degradation of lysosomal sphingomyelinase inhuman fibroblasts. Biol. Chem. Hoppe Seyler 375: 447–450.
37. Luberto, C., D. F. Hassler, P. Signorelli, Y. Okamoto, H. Sawai, E. Boros,D. J. Hazen-Martin, L. M. Obeid, Y. A. Hannun, and G. K. Smith. 2002. In-hibition of tumor necrosis factor-induced cell death in MCF7 by a novel inhibitorof neutral sphingomyelinase. J. Biol. Chem. 277: 41128–41139.
38. Bjorkbacka, H., K. A. Fitzgerald, F. Huet, X. Li, J. A. Gregory, M. A. Lee,C. M. Ordija, N. E. Dowley, D. T. Golenbock, and M. W. Freeman. 2004. Theinduction of macrophage gene expression by LPS predominantly utilizes Myd88-independent signaling cascades. Physiol. Genomics 19: 319–330.
39. Hirotani, T., M. Yamamoto, Y. Kumagai, S. Uematsu, I. Kawase, O. Takeuchi,and S. Akira. 2005. Regulation of lipopolysaccharide-inducible genes by MyD88and Toll/IL-1 domain containing adaptor inducing IFN-b. Biochem. Biophys.Res. Commun. 328: 383–392.
40. Hoebe, K., X. Du, P. Georgel, E. Janssen, K. Tabeta, S. O. Kim, J. Goode, P. Lin,N. Mann, S. Mudd, et al. 2003. Identification of Lps2 as a key transducer ofMyD88-independent TIR signalling. Nature 424: 743–748.
41. Lee, J. Y., C. A. Lowell, D. G. Lemay, H. S. Youn, S. H. Rhee, K. H. Sohn,B. Jang, J. Ye, J. H. Chung, and D. H. Hwang. 2005. The regulation of theexpression of inducible nitric oxide synthase by Src-family tyrosine kinasesmediated through MyD88-independent signaling pathways of Toll-like receptor4. Biochem. Pharmacol. 70: 1231–1240.
42. Edsall, L. C., J. R. Van Brocklyn, O. Cuvillier, B. Kleuser, and S. Spiegel. 1998.N,N-Dimethylsphingosine is a potent competitive inhibitor of sphingosine kinasebut not of protein kinase C: modulation of cellular levels of sphingosine 1-phosphate and ceramide. Biochemistry 37: 12892–12898.
43. Bielawska, A., M. S. Greenberg, D. Perry, S. Jayadev, J. A. Shayman, C. McKay,and Y. A. Hannun. 1996. (1S,2R)-D-erythro-2-(N-myristoylamino)-1-phenyl-1-propanol as an inhibitor of ceramidase. J. Biol. Chem. 271: 12646–12654.
44. Raisova, M., G. Goltz, M. Bektas, A. Bielawska, C. Riebeling, A. M. Hossini,J. Eberle, Y. A. Hannun, C. E. Orfanos, and C. C. Geilen. 2002. Bcl-2 over-expression prevents apoptosis induced by ceramidase inhibitors in malignantmelanoma and HaCaT keratinocytes. FEBS Lett. 516: 47–52.
45. Bielawska, A. J., J. Bielawski, Z. M. Szulc, N. Mayroo, X. Liu, A. Bai,S. Elojeimy, B. Rembiesa, J. Pierce, J. S. Norris, and Y. A. Hannun. 2008. Novelanalogs of D-e-MAPP and B13. Part 2: signature effects on bioactive sphingo-lipids. Bioorg. Med. Chem. 16: 1032–1045.
46. Kumada, H., S. Mitsutake, Y. Inagaki, S. Mitsunaga, H. Tsuchikawa, S. Katsumura,and Y. Igarashi. 2007. Kinetics of the ceramide kinase inhibitor K1, a suppressorof mast-cell activation. Biosci. Biotechnol. Biochem. 71: 2581–2584.
47. Snider, A. J., K. A. Orr Gandy, and L. M. Obeid. 2010. Sphingosine kinase: rolein regulation of bioactive sphingolipid mediators in inflammation. Biochimie 92:707–715.
48. Procyk, K. J., M. R. Rippo, R. Testi, F. Hofmann, P. J. Parker, and M. Baccarini.2000. Lipopolysaccharide induces jun N-terminal kinase activation in macro-phages by a novel Cdc42/Rac-independent pathway involving sequential acti-vation of protein kinase C z and phosphatidylcholine-dependent phospholipaseC. Blood 96: 2592–2598.
49. Hellerud, B. C., J. Stenvik, T. Espevik, J. D. Lambris, T. E. Mollnes, andP. Brandtzaeg. 2008. Stages of meningococcal sepsis simulated in vitro, withemphasis on complement and Toll-like receptor activation. Infect. Immun. 76:4183–4189.
50. Grassme, H., V. Jendrossek, A. Riehle, G. von Kurthy, J. Berger, H. Schwarz,M. Weller, R. Kolesnick, and E. Gulbins. 2003. Host defense against Pseudo-monas aeruginosa requires ceramide-rich membrane rafts. Nat. Med. 9: 322–330.
51. McCollister, B. D., J. T. Myers, J. Jones-Carson, D. R. Voelker, and A. Vazquez-Torres. 2007. Constitutive acid sphingomyelinase enhances early and late mac-rophage killing of Salmonella enterica serovar Typhimurium. Infect. Immun. 75:5346–5352.
52. Utermohlen, O., U. Karow, J. Lohler, and M. Kronke. 2003. Severe impairmentin early host defense against Listeria monocytogenes in mice deficient in acidsphingomyelinase. J. Immunol. 170: 2621–2628.
53. Thomas, K. E., C. L. Galligan, R. D. Newman, E. N. Fish, and S. N. Vogel. 2006.Contribution of interferon-beta to the murine macrophage response to the toll-like receptor 4 agonist, lipopolysaccharide. J. Biol. Chem. 281: 31119–31130.
54. Kagan, J. C., and R. Medzhitov. 2006. Phosphoinositide-mediated adaptor re-cruitment controls Toll-like receptor signaling. Cell 125: 943–955.
55. Kagan, J. C., T. Su, T. Horng, A. Chow, S. Akira, and R. Medzhitov. 2008.TRAM couples endocytosis of Toll-like receptor 4 to the induction of interferon-b. Nat. Immunol. 9: 361–368.
56. Hailemariam, T. K., C. Huan, J. Liu, Z. Li, C. Roman, M. Kalbfeisch, H. H. Bui,D. A. Peake, M. S. Kuo, G. Cao, et al. 2008. Sphingomyelin synthase 2 de-ficiency attenuates NFkappaB activation. Arterioscler. Thromb. Vasc. Biol. 28:1519–1526.
57. Zha, X., L. M. Pierini, P. L. Leopold, P. J. Skiba, I. Tabas, and F. R. Maxfield.1998. Sphingomyelinase treatment induces ATP-independent endocytosis. J.Cell Biol. 140: 39–47.
58. Holopainen, J. M., M. I. Angelova, and P. K. Kinnunen. 2000. Vectorial buddingof vesicles by asymmetrical enzymatic formation of ceramide in giant lip-osomes. Biophys. J. 78: 830–838.
59. Liu, H., P. Sidiropoulos, G. Song, L. J. Pagliari, M. J. Birrer, B. Stein,J. Anrather, and R. M. Pope. 2000. TNF-a gene expression in macrophages:regulation by NF-k B is independent of c-Jun or C/EBP b. J. Immunol. 164:4277–4285.
60. Feng, C., A. G. Mery, E. M. Beller, C. Favot, and J. A. Boyce. 2004. Adeninenucleotides inhibit cytokine generation by human mast cells through a Gs-cou-pled receptor. J. Immunol. 173: 7539–7547.
61. Goldsmith, M., D. Avni, G. Levy-Rimler, R. Mashiach, O. Ernst, M. Levi,B. Webb, M. M. Meijler, N. S. Gray, H. Rosen, and T. Zor. 2009. A ceramide-1-phosphate analogue, PCERA-1, simultaneously suppresses tumour necrosisfactor-alpha and induces interleukin-10 production in activated macrophages.Immunology 127: 103–115.
62. Jaramillo, M., and M. Olivier. 2002. Hydrogen peroxide induces murine mac-rophage chemokine gene transcription via extracellular signal-regulated kinase-and cyclic adenosine 59-monophosphate (cAMP)-dependent pathways: in-volvement of NF-k B, activator protein 1, and cAMP response element bindingprotein. J. Immunol. 169: 7026–7038.
63. Avni, D., A. Philosoph, M. M. Meijler, and T. Zor. 2009. The ceramide-1phosphateanalogue PCERA-1 modulates tumor necrosis factor-a and interleukin-10 pro-duction in macrophages via the cAMP-PKA-CREB pathway in a GTP-dependentmanner. Immunology 129: 375–385.
64. Granado, M. H., P. Gangoiti, A. Ouro, L. Arana, M. Gonzalez, M. Trueba, andA. Gomez-Munoz. 2009. Ceramide 1-phosphate (C1P) promotes cell migrationInvolvement of a specific C1P receptor. Cell. Signal. 21: 405–412.
65. Levi, M., M. M. Meijler, A. Gomez-Munoz, and T. Zor. 2010. Distinct receptor-mediated activities in macrophages for natural ceramide-1-phosphate (C1P) andfor phospho-ceramide analogue-1 (PCERA-1). Mol. Cell. Endocrinol. 314: 248–255.
6972 CERAMIDE NEGATIVELY REGULATES LPS-INDUCED TNF-a PRODUCTION
by guest on June 14, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

66. Beutler, B., I. W. Milsark, and A. C. Cerami. 1985. Passive immunization againstcachectin/tumor necrosis factor protects mice from lethal effect of endotoxin.Science 229: 869–871.
67. Rothe, J., W. Lesslauer, H. Lotscher, Y. Lang, P. Koebel, F. Kontgen, A. Althage,R. Zinkernagel, M. Steinmetz, and H. Bluethmann. 1993. Mice lacking the tu-mour necrosis factor receptor 1 are resistant to TNF-mediated toxicity but highlysusceptible to infection by Listeria monocytogenes. Nature 364: 798–802.
68. Watson, A. D., N. Leitinger, M. Navab, K. F. Faull, S. Horkko, J. L. Witztum,W. Palinski, D. Schwenke, R. G. Salomon, W. Sha, et al. 1997. Structural iden-tification by mass spectrometry of oxidized phospholipids in minimally oxidizedlow density lipoprotein that induce monocyte/endothelial interactions and evi-dence for their presence in vivo. J. Biol. Chem. 272: 13597–13607.
69. Bochkov, V. N., A. Kadl, J. Huber, F. Gruber, B. R. Binder, and N. Leitinger.2002. Protective role of phospholipid oxidation products in endotoxin-inducedtissue damage. Nature 419: 77–81.
70. Erridge, C., S. Kennedy, C. M. Spickett, and D. J. Webb. 2008. Oxidizedphospholipid inhibition of toll-like receptor (TLR) signaling is restricted toTLR2 and TLR4: roles for CD14, LPS-binding protein, and MD2 as targets forspecificity of inhibition. J. Biol. Chem. 283: 24748–24759.
71. Podrez, E. A., E. Poliakov, Z. Shen, R. Zhang, Y. Deng, M. Sun, P. J. Finton,L. Shan, B. Gugiu, P. L. Fox, et al. 2002. Identification of a novel family ofoxidized phospholipids that serve as ligands for the macrophage scavenger re-ceptor CD36. J. Biol. Chem. 277: 38503–38516.
72. Greenberg, M. E., M. Sun, R. Zhang, M. Febbraio, R. Silverstein, andS. L. Hazen. 2006. Oxidized phosphatidylserine-CD36 interactions play an es-sential role in macrophage-dependent phagocytosis of apoptotic cells. J. Exp.Med. 203: 2613–2625.
73. Li, R., K. P. Mouillesseaux, D. Montoya, D. Cruz, N. Gharavi, M. Dun,L. Koroniak, and J. A. Berliner. 2006. Identification of prostaglandin E2 receptorsubtype 2 as a receptor activated by OxPAPC. Circ. Res. 98: 642–650.
74. Sun, Y., T. Fox, G. Adhikary, M. Kester, and E. Pearlman. 2008. Inhibition ofcorneal inflammation by liposomal delivery of short-chain, C-6 ceramide. J.Leukoc. Biol. 83: 1512–1521.
75. Rozenova, K. A., G. M. Deevska, A. A. Karakashian, and M. N. Nikolova-Karakashian. 2010. Studies on the role of acid sphingomyelinase and ceramidein the regulation of tumor necrosis factor alpha (TNFalpha)-converting enzymeactivity and TNFalpha secretion in macrophages. J. Biol. Chem. 285: 21103–21113.
The Journal of Immunology 6973
by guest on June 14, 2018http://w
ww
.jimm
unol.org/D
ownloaded from