ch f06-enzimi di fase ii - giorgio sartor · 4 gs © 2001-2018 ver 3.6.3 enzimi di fase ii...
TRANSCRIPT

1
Enzimi di Fase II
Prof. Giorgio Sartor
Copyright © 2001-2018 by Giorgio Sartor.
All rights reserved. F06 - Versione 3.6.3 – May-18
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 2 -
Enzimi di Fase II
Lipofilo Idrofilo
Xen
obio
tico Metabolita
|OH
Metabolita|
ORFase I
(ossidazione)
Fase II(sintesi)
• Variazione di polarità• Variazione di funzionalità
• Variazione di dimensioni• Variazione di carica• Variazione di solubilità
• Diminuzione dell’attività biologica• Aumentata escrezione
Escrezione

2
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 3 -
Enzimi di fase II
• Catalizzano reazioni di:–Coniugazione con
• Acido glucuronico• Glutatione • Solfato• Acetile• Aminoacidi
–Metilazione
Metallotioneine
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 4 -
Coniugazione con Acido Glucuronico
• Glucuronazione:– Coniugazione di xenobiotici con un H labile con acido
glucuronico da acido uridindifosfoglicerico (UDPGA)
UDPGA
NH
OO–
OH
OH OH
OO P
O
O–
O PO
O–
O
O
OH
O
OH
O
N

3
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 5 -
Formazione di UDPGA
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 6 -
Formazione di UDPGA

4
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 7 -
Glucosio-1-fosfato
fosfoglucomutasiEC 5.4.2.2
G6P
G1P
G16BP
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 8 -
FosfoglucomutasiEC 5.4.2.2 (1C47)
G16BP

5
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 9 -
FosfoglucomutasiEC 5.4.2.2 (1C47)
G16BP
SER-19
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 10 -
Formazione di UDPG
UTP-glucoso-1-fosfato uridililtransferasiEC 2.7.7.9
UTP
UDP

6
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 11 -
Formazione di UDPGA
H2O
UDP-glucoso 6-deidrogenasiEC 1.1.1.22
UDPGNAD+
UDPGA NADH
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 12 -
UDP-glucoso-6-deidrogenasi
NAD+

7
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 13 -
UDP-glucoso-6-deidrogenasi
NAD+
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 14 -
β-UDP-glucuroniltransferasi
• L’enzima β-UDP-glucuroniltransferasi forma O-, N-, S-, C- glucuronati; – Sei forme nel fegato umano– Il cofattore è UDP-glucuronato– Induttori: fenobarbital, indoli, 3-metilcolantrene,
(fumo di sigarette).– Substrati: destrofano, metadone, morfine, p-
nitrofenolo, acido valproico, ormoni steroidei,bilirubina.

8
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 15 -
Coniugazione con acido glucuronico
ROH
UDPglucuronato β-D-glucuronosiltransferasiEC 2.4.1.17
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 16 -
Metabolismo degli androgeni ed estrogeni

9
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 17 -
Metabolismo delle porfirine
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 18 -
• Sindrome di Crigler-Nijar (grave): enzima inattivato; grave iperbilirubinemia; gli induttori non hanno effetto.
• Sindrome di Gilbert (lieve): ridotta attività enzimatica; lieve iperbilirubinemia; il fenobarbital porta la glucuronazione della bilirubina alla normalità
• I pazienti possono glucuronidare p-nitrofenolo, morfine, cloroamfenicolo.
Patologie e polimorfismo genetico

10
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 19 -
Glucuronazione e glucuronidasi
• I coniugati vengono escreti con la bile o con le urine.
• La glucuronidasi della microflora intestinale taglia il legame con l’acido glucuronico.
• La parte agliconica può essere riassorbita ed andare incontro ad un riciclo enteroepatico.
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 20 -
Glucuronazione e glucuronidasi
• Attivazione metabolica di 2,6-dinitrotoluene da parte della glucuronidasi.– La glucuronidasi rimuove l’acido glucuronico da N-
glucuronide;– Il nitro gruppo viene ridotto da N-reduttasi
microbiche;– Il risultante metabolita epatocarcinogeno è
riassorbito.

11
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 21 -
Gli enzimi di fase II catalizzano reazioni di:
• Coniugazione con– Acido glucuronico– Solfato– Glutatione– Acetile– Aminoacidi
• Metilazione Metallotioneine
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 22 -
Coniugazione con il gruppo solfato
• Coniugazione di:– xenobiotici con un OH libero con solfato da
fosfoadeninafosfosolfonato (PAPS)
N N
SO
OO–
O PO
O–O
PO O–
O–
O
NH2
NO
OH
N
12
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 23 -
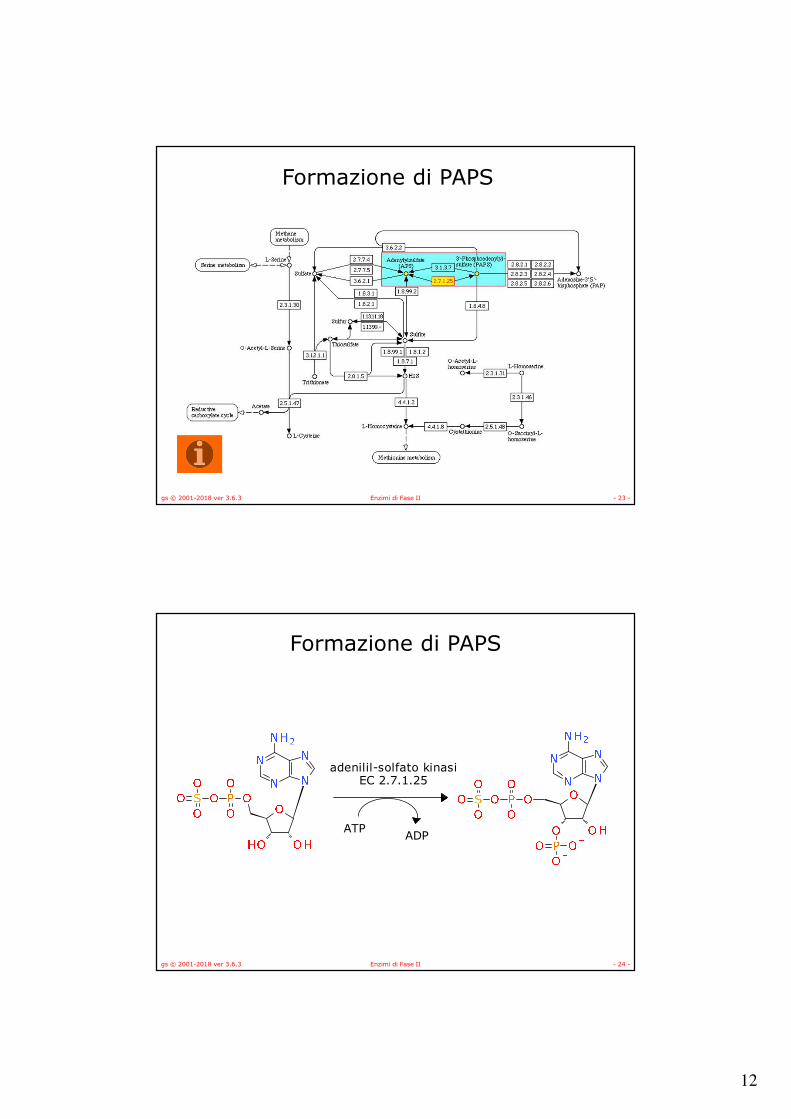
Formazione di PAPS
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 24 -
Formazione di PAPS
ATPADP
adenilil-solfato kinasiEC 2.7.1.25

13
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 25 -
• Le solfotransferasi sono enzimi ubiquitari• Cofattore è 3’-fosfoadenosina-5’-fosfosolfato
(PAPS)• Produce esteri solfati altamente idrosolubili• Eliminati con urine e bile• Xenobiotici e composti endogeni sono solfonati
(fenoli, catecoli, amine, idrossilamine)
Formazione di solfati
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 26 -
Meccanismo della formazione di solfati
PAPS
PAP
steroide solfotransferasiEC 2.8.2.15
H
H
HSOO
O–
O
OCH3
OCH3N N
PO
O–O– O
PO
O–O– O
NH2
NO
OH
N
CH3
H
H
H
OCH3
OH
O
OP
NH2
N
NO
OH
N
N
SO
OO–
O PO
O–O
O O–
O–
+
+

14
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 27 -
Metabolismo degli androgeni ed estrogeni
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 28 -
• La formazione di solfati è una via metabolica ad alta affinità e bassa capacità.– La glucuronazione ha bassa affinità ed alta capacità.– La capacità è limitata dai bassi livelli di PAPS.– A bassi dosaggi prevale la formazione di solfati.– Ad alti dosaggi prevale la glucuronazione.
Formazione di solfati

15
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 29 -
Formazione di solfati
• Esistono quattro solfotransferasi nel citosol degli epatociti umani
• Le arilsolfatasi nella flora intestinale rimuovono il solfato promuovendo il riciclo enteroepatico
• Possono attivare xenobiotici a carcinogenici se il composto coniugato è chimicamente instabile– I solfonati delle idrossilammine sono instabili.
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 30 -
Gli enzimi di fase II catalizzano reazioni di:
• Coniugazione con– Acido glucuronico– Solfato– Glutatione – Acetile– Aminoacidi
• Metilazione Metallotioneine

16
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 31 -
Coniugazione con glutatione
• Coniugazione di xenobiotici (numerosissimi substrati) con una regione elettrofila (epossido) con glutatione ridotto (GSH) – glutatione-S-transferasi
GSH
GSSG
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 32 -
• Il glutatione è un tripeptide formato da glicina, cisteina, acido glutamico– Formato da γ-glutamilcisteina sintetasi (EC 6.3.2.3)
GSH
GLU CYS GLY

17
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 33 -
Sintesi del GSH
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 34 -
Sintesi del GSH

18
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 35 -
Sintesi del GSH
ATPADP
ATPADP
glutamato-cisteina ligasiEC 6.3.2.2
CYS
GLY
GLU
glutatione sintasiEC 6.3.2.3
1M0W 2HGS
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 36 -
Glutatione sintasi (1M0W)
γ-glutamil cisteina
omologo di ATP
Mg++

19
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 37 -
Glutatione sintasi (1M0W)
γ-glutamil cisteina
omologo di ATP
Mg++
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 38 -
Glutatione sintasi (1M0W)
γ-glutamil cisteina
Mg++
omologo di ATP
In rosso: aminoacidi basici
In verde: la superficie della proteina

20
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 39 -
Glutatione sintasi (2HGS)
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 40 -
Vie metaboliche e radicali dell’ossigeno
XENOBIOTICI
METABOLITIREATTIVI
COMPOSTIREDOX
METABOLITIRADICALICI
H2O2
O2
HO.
O2-
O2
H2O + O2
H2O
GSH
GSSG
FAD
FADH2
NADPH + H+
NADP+
O2
NADPH + H+
NADP+
Rotturadel DNA
Inattivazionedi enzimi
Perossidazionelipidica
Addottial DNA
Addottidi proteine Prodotti di
coniugazione
Addottial DNA
Addottidi proteine
Metabolismo
FASE I
FASE II
1
2
3
4
5
Fe++, Cu++
7
6
8
9GR GPX
Ciclo RedoxEnzimi
a Flavina
FASE I

21
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 41 -
Vie metaboliche e radicali dell’ossigeno
XENOBIOTICI
METABOLITIREATTIVI
COMPOSTIREDOX
METABOLITIRADICALICI
H2O2
O2
HO.
O2-
O2
H2O + O2
H2O
GSH
GSSG
FAD
FADH2
NADPH + H+
NADP+
O2
NADPH + H+
NADP+
Rotturadel DNA
Inattivazionedi enzimi
Perossidazionelipidica
Addottial DNA
Addottidi proteine Prodotti di
coniugazione
Addottial DNA
Addottidi proteine
Metabolismo
FASE I
FASE II
1
2
3
4
5
Fe++, Cu++
7
6
8
9GR GPX
Ciclo RedoxEnzimi
a Flavina
FASE I
5. In presenza di ferro l’anione superossido forma il radicale idrossido (Haber-Weiss).
6. L’anione superossido può dismutare a H2O2 ad opera della superossidodismutasi (SOD, EC 1.15.1.1).
7. In presenza di ferro l’ H2O2 forma il radicale idrossido e ossigeno (Fenton).
8. L’ H2O2 può essere degradata ad acqua ad opera della catalasi (CAT, EC 1.11.1.6) o della glutatione perossidasi (GPX, EC 1.11.1.9).
9. La glutatione reduttasi (GR, EC 1.8.1.7) rigenera il glutatione ridotto (GSH) da quello ossidato (GSSG).
GPX GR
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 42 -
• O-demetilazione di organofosfati• Attivazione della trinitroglicerina
– I prodotti sono glutatione ossidato (GSSG), dinitroglicerina, NO (vasodilatore)
• Riduzione di idroperossidi– Metabolismo delle prostaglandine
Coniugazione con GSH

22
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 43 -
Metabolismo delle prostaglandine
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 44 -
Metabolismo delle prostaglandine
H2O
leucotriene-C4 sintasiEC 2.5.1.37

23
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 45 -
• Due tipi di reazione con il GSH– Spiazzamento di alogeni, solfati, solfonati, fosfati,
nitrogruppi– Il glutatione viene addizionato a doppi legami attivati
(epossidi) come nella sintesi delle prostaglandine.
• Substrati:– Idrofobici che contengono un elettrofilo– Possono reagire non enzimaticamente
Coniugazione con il GSH
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 46 -
Coniugazione
RX
HX
glutatione S-transferasiEC 2.5.1.18
24
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 47 -
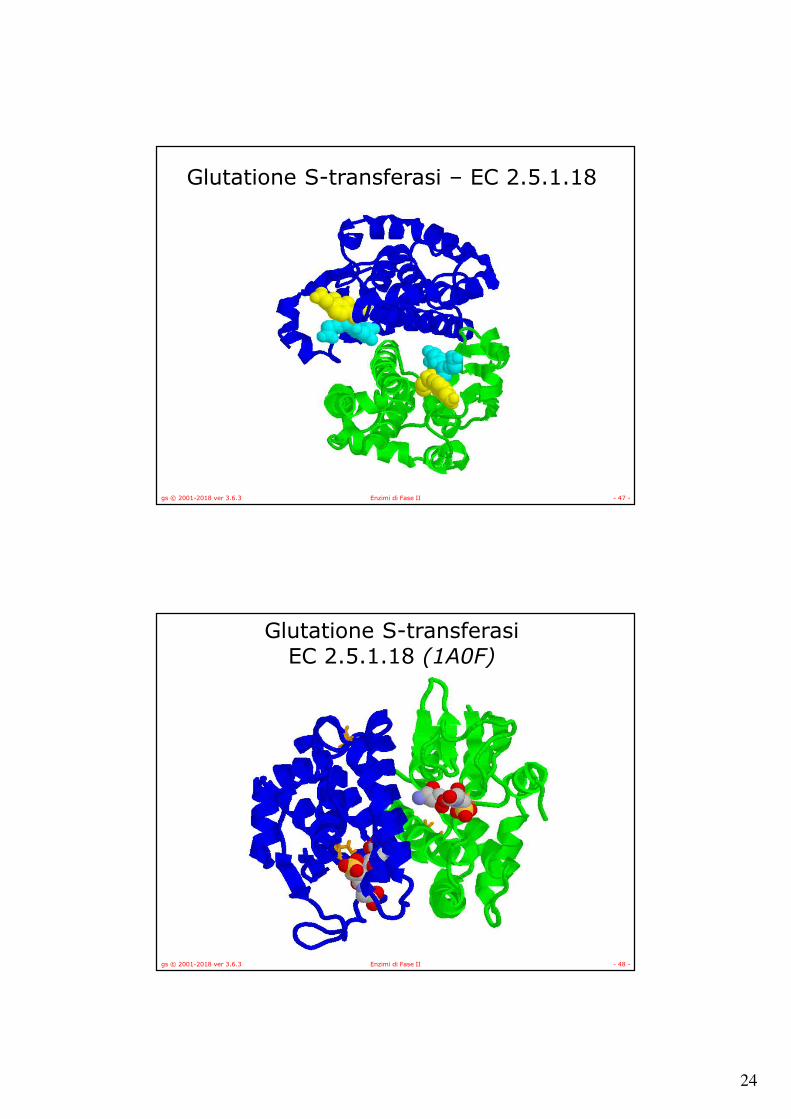
Glutatione S-transferasi – EC 2.5.1.18
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 48 -
Glutatione S-transferasi EC 2.5.1.18 (1A0F)

25
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 49 -
• Glutatione-S-transferasi microsomiale e citosolica
• Almeno quattro classi di glutatione-S-transferasi (α, β, γ, δ), diversi isoenzimi inducibili.– Induttori (3-metilcolantrene, fenobarbital,
corticosteroidi, anti-ossidanti)– La sovraespressione porta a resistenza (insetti
DDT resistenti, piante resistenti all’atrazina, cellule tumorali resistenti alla chemioterapia)
Glutatione S-transferasi
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 50 -
• Escrezione dei glutatione-coniugati:– Intatti nella bile– Convertiti in acido mercapturico nei reni ed escreti
nelle urine • γ-glutamiltransferasi, aminopeptidasi M
Eliminazione dei composti coniugati con il GSH

26
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 51 -
Metabolismo del naftalene
naftalenenaftalene
monoossigenasi
GSH
S-(1,2-diidro-2-idrossi-1-naftil)-
S-(1,2-diidro-2-idrossi-1-naftil)-
S-(1,2-diidro-2-idrossi-1-naftil)-S-(1,2-diidro-2-idrossi-1-naftil)-N-Acido 1-
FASE IFASE II
OH
O
O
OH
CH3
ONH
OH
O
S
CH3
ONH
OHH
H
O
SOH
OHH
H
NH2
O
SOH
H
H
NH
NH2
O
SOH
H
H
NH
NH2
ONH
O
SOHO
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 52 -
γ-glutamiltransferasi
H2O
-glutamiltransferasiEC 2.3.2.2
γ
R-S-alanilglicina Glutamato

27
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 53 -
Gli enzimi di fase II catalizzano reazioni di:
• Coniugazione con– Acido glucuronico– Solfato– Glutatione– Acetile– Aminoacidi
• Metilazione Metallotioneine
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 54 -
Coniugazione con il gruppo acetile
• Coniugazione di:– xenobiotici con un OH libero con gruppo acetile da
acetil-CoA

28
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 55 -
Acetilazione
• Maggiore via di biotrasformazione di amine aromatiche e idrazine
• Diminuisce la solubilità• N-acetiltransferasi (NAT)
– Il cofattore è acetil-CoA
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 56 -
Meccanismo della acetilazioneAcetilCoA
CoA
arilamina N-acetiltransferasiEC 2.3.1.5
O
C H3NH
NN
SHNH
O
NH
O
OHC H3
C H3
OPO
O–
O
PO
O–
P
O
OH
O–O
O
NH2
N
OH
O
N
NH2
NN
O
CH3SNH
O
NH
O
OHCH3
CH3
OPO
O–
O
PO
O–
P
O
O–
OH
O
O
NH2
N
OH
O N
+
+

29
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 57 -
Gli enzimi di fase II catalizzano reazioni di:
• Coniugazione con– Acido glucuronico– Glutatione – Solfato– Acetile– Aminoacidi
• Metilazione Metallotioneine
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 58 -
Coniugazione con aminoacidi
• Alternativa alla glucuronazione• Due vie principali
– Gruppo -COOH del substrato coniugato con -NH2 di glicine, serina, glutamina, richiede attivazione con CoA
• Coniugazione dell’acido benzoico con glicina per dare l’acido ippurico
– Gruppi -NH2 o -NHOH aromatici coniugati con gruppi COOH di serina, prolina, richiedono attivazione con ATP

30
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 59 -
• Specie specificità nell’aminoacido accettore– mammiferi: glicina– uccelli: ornitina– cani e gatti: taurina– primati non umani: glutamina
• Attivazione metabolica– Gli N-esteri di serina o prolina delle idrossilamine sono
instabili e degradano a specie elettrofile reattive.
Coniugazione con aminoacidi
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 60 -
Meccanismo della coniugazione con aminoacidi
glicina N-benzoiltransferasiEC 2.3.1.71
O
HH
O
O–NH
N
N
NH2
N
N
P
O
OHO
–
O OH
PO
O–
PO
O–
C H3
C H3
OH
O O
SHNH
NH
O
O
O
O
O
O–
HH
NH2
N
N
NH2
N
N
P
O
OH
O–
O OH
PO
O–
PO
O–
C H3
C H3
OH
O O
O
SNH
NH
O
O
OO
+
+

31
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 61 -
Gli enzimi di fase II catalizzano reazioni di:
• Coniugazione con– Acido glucuronico– Glutatione– Solfato– Acetile– Aminoacidi
• Metilazione Metallotioneine
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 62 -
• Via metabolica che diminuisce la solubilità• Metiltransferasi
– Trasferisce un gruppo -CH3 a O, N, S, C– Cofattore: S-adenosilmetionina (SAM)
• Substrati sono: fenoli, catecoli, amine, metalli (Hg, As, Se)
Metilazione

32
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 63 -
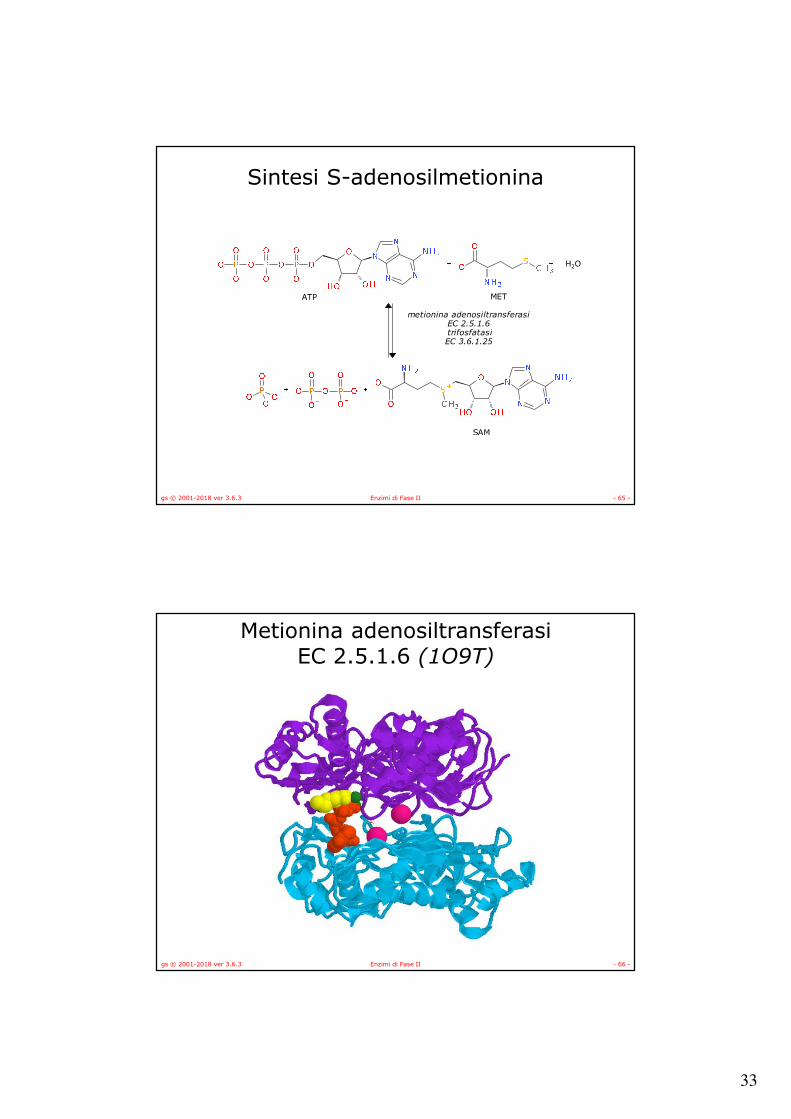
Sintesi di S-adenosilmetionina
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 64 -
Sintesi di S-adenosilmetionina
33
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 65 -
Sintesi S-adenosilmetionina
H2O
metionina adenosiltransferasiEC 2.5.1.6
trifosfatasiEC 3.6.1.25
ATP MET
SAM
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 66 -
Metionina adenosiltransferasiEC 2.5.1.6 (1O9T)

34
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 67 -
Metiltransferasi
• Varie metiltransferasi– fenolo O-metiltransferasi, – catecolo O-metiltransferasi, – N-metiltransferasi, – S-metiltransferasi
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 68 -
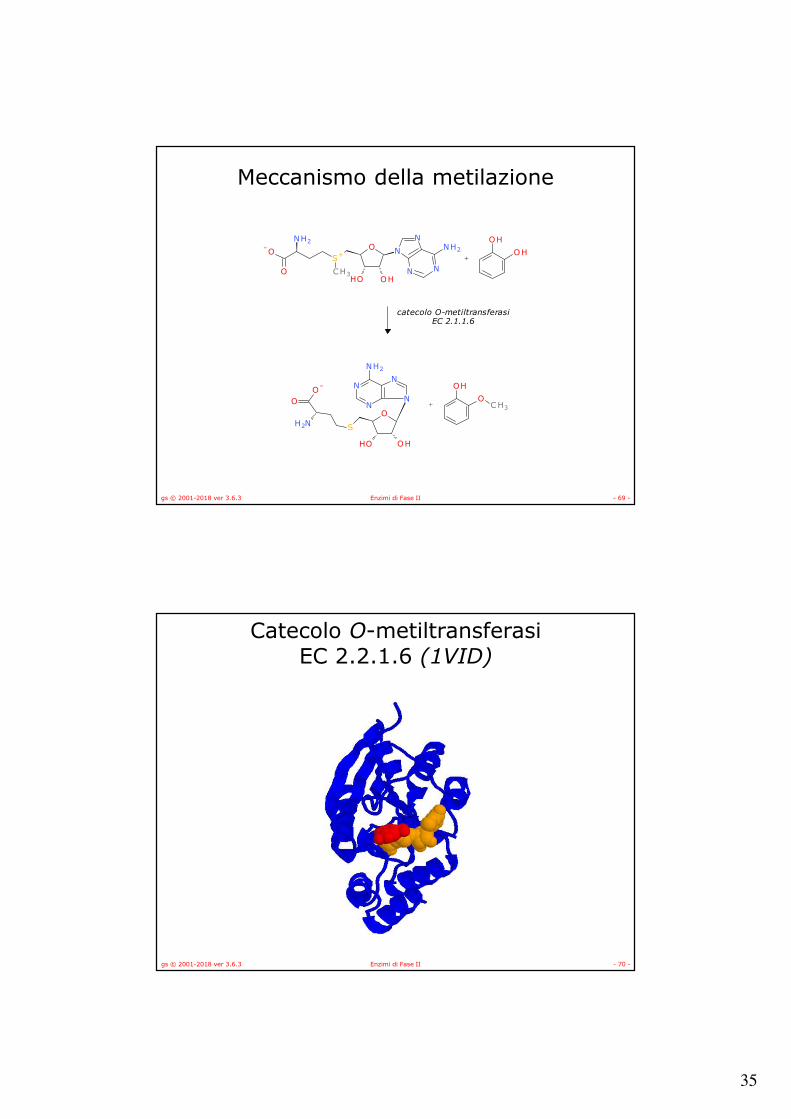
Meccanismo della metilazione
amino N-metiltransferasiEC 2.1.1.49
35
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 69 -
Meccanismo della metilazione
catecolo O-metiltransferasiEC 2.1.1.6
OC H3
OHNN
OO–
NH2 S
OH
NH2
OH
ON
N
OH
OH
N
N
O
O–NH2
S+
CH3OH
NH2
OH
O
N
N+
+
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 70 -
Catecolo O-metiltransferasiEC 2.2.1.6 (1VID)

36
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 71 -
Catecolo O-metiltransferasiEC 2.2.1.6 (1VID)
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 72 -
Mercurio
mercurio(II) reduttasiEC 1.16.1.1 RH H+
alchilmercurio liasiEC 4.99.1.2
NO
O
N
N
OH
PO–
O–O
O
N
NH2
POO–
OPOO–
OO
OH
N+
OH
NH2
O
RHg+
H HN N
O
NH2
N
OH OH
O O PO
O–O P
O
O–O
NH2
PO O–
O–
N
OOH
ON
Hg2+
Hg2+
H+Hg + +
+
++
AND Enantiomer

37
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 73 -
Gli enzimi di fase II catalizzano reazioni di:
• Coniugazione con– Acido glucuronico– Solfato– Glutatione– Acetile– Aminoacidi
• Metilazione Metallotioneine
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 74 -
Metallotioneine
• Le metallotioneine (MT) sono peptidi eproteine ubiquitarie a basso peso molecolaread alto contenuto in aminoacidi solforati emetalli.
• Si ipotizza che giochino un ruolo:– nella fissazione dei metalli in tracce (Zn++, Cu++),– nel controllare la concentrazione di questi ioni,– nella regolazione dei flussi degli ioni ai distretti
cellulari,– nella neutralizzazione dei metalli tossici (Cd++, Hg++)
e nella protezione dallo stress indotto dai metalli.

38
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 75 -
Distribuzione
• Le metallotioneine sono presenti in tutti gli organismi:animali, vegetali e microrganismi.
• Negli animali queste proteine posseggono polimorfismogenetico e sono abbondanti nei tessuti parenchimali(fegato, rene, pancreas e intestino).
• La loro concentrazione dipende da specie, tessuto, età,sesso ed altri fattori non ancora completamenteidentificati
• Nonostante che le metallotioneine siano proteinecitoplasmatiche si sono trovate accumulate nei lisosomie nel nucleo.
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 76 -
Trasporto di metallotioneine
• Schema ipotetico del trasporto di metalli pesanti attraverso l’epitelio branchiale di molluschi bivalvi.– M: metalli in traccia;– MT: metallotioneine.

39
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 77 -
Turnover delle metallotioneine
• Schema del turnover di MT in cellule di molluschi bivalvi (Isani et al., 2000).
– M: Metalli; – MTF: Fattori di
Trascrizione; – MTI: Inibitori di
Trascrizione.
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 78 -
Classificazione
• Il nome deriva dal fatto che hannoun alto contenuto di zolfo e metalli.Tale contenuto varia a secondo delmetallo (fino ad oltre il 20% in peso)
• Nei mammiferi sono caratterizzateda un peso molecolare di 6000-7000Da, contengono da 60 ad 68 aminoacidi di cui 20 Cys che legano 7equivalenti di ione metallicobivalente. Mancano di aminoacidiaromatici. Tutte le Cys sono in formaridotta e sono coordinate con ionimetallici.

40
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 79 -
Classificazione – Le metallotioneine
• La superfamiglia delle the metallotioneine è definitacome quella che comprende i peptidi che assomiglianoalla metallotioneina renale equina che ha le seguenticaratteristiche:– Basso peso molecolare– Composizione:
• Alto contenuto in Cys, basso contenuto in aromatici.• Sequenza caratteristica.
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 80 -
Classificazione – Le famiglie
• Una famiglia di metallotioneine è caratterizzata da unaparticolare sequenza ed è legata ad una o più specie.– I membri di una determinata famiglia appartengono solo a
quella e si pensa siano correlati da un punto di vistaevoluzionistico
– Ogni famiglia è identificata da un numero e dalla specie.– Per esempio: Famiglia 1: vertebrati.

41
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 81 -
Classificazione
• Le sottofamiglie– Si definiscono sottofamiglie di metallotioneine quegli
insiemi di proteine che oltre i caratteri propri delle famigliecondividono un insieme di caratteri più stringenti.
– Per esempio: m1, m2…
• I sottogruppi– Un sottogruppo rappresenta un insieme di sequenza
correlate filogeneticamente. In un albero filogeneticorappresentano un ramo.
– Per esempio: m2U2 = MT-2 di ungulati, sottogruppo dellasottofamiglia m2.
• Le isoforme– Sono i membri di sottogruppi, sottofamiglie e famiglie.– Per esempio MT-1E umana.
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 82 -
Classificazione – I clan
• Un clan è un insieme di proteine che dividono dellecaratteristiche non già definite:– Struttura,– Proprietà termodinamiche,– Affinità per i metalli– Proprietà funzionali– …

42
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 83 -
Sequenza Famiglia Caratteristiche Sottofamiglie
K-x(1,2)-C-C-x-C-C-P-x(2)-C1
vertebrati
Da 60 a 68 AA; 20 Cys (21 in un caso), 19 totalmenteconservate; due domini strutturali, contenenti 9 e 11 Cysche legano 3 e 4 ioni bivalenti rispettivamente. Il gene ècomposto di 3 esoni, 2 introni
m1, m2, m3, m4, m, a, a1, a2, b, ba, t
C-x-C-x(3)-C-T-G-x(3)-C-x-C-x(3)-C-x-C-K
2molluschi
Da 64 a 75 AA; da 18 a 23 Cys, minimo 13 totalmenteconservate; due domini
mo1, mo2, mog, mo
P-[GD]-P-C-C-x(3,4)-C-x-C3
crostacei
da 58 a 60 AA; esistono varianti con e senza Met N-terminale; 18 Cys totalmente conservate; due domini,ognuno con 9 Cys che legano 3 ioni bivalenti
c1, c2, c
P-D-x-K-C-[V,F]-C-C-x(5)-C-x-C-x(4)-C-C-x(4)-C-C-x(4,6)-C-C
4echinodermi
Da 64 a 67 AA; 20 Cys ; due domini strutturali,contenenti 9 e 11 Cys che legano 3 e 4 ioni bivalentirispettivamente
e1, e2
C-G-x(2)-C-x-C-x(2)-Q-x(5)-C-x-C-x(2)D-C-x-C
5ditteri
Da 40 a 43 AA; 10 Cys conservate d1, d2
K-C-C-x(3)-C-C6
nematodi 62 e 74 AAs; 18 Cys, contiene una Tyr n1, n2
una sequenza7
ciliati105 AA, 31 Cys, multiplo pattern CCC, una Tyr ci
C-G-C-S-x(4)-C-x-C-x(3,4)-C-x-C-S-x-C
8funghi I Da 25 a 33 AA; 7 Cys f1
C-X-K-C-x-C-x(2)-C-K-C 11funghi IV
Da 55 a 56 AA; 9 Cys; un pattern CCC; contiene His ePhe f4
K-C-A-C-x(2)-C-L-C 14procarioti
Da 53 a 56 AAs; 9 Cys; una Tyr, una His; contine residuinon comuni
p
[YFH]-x(5,25)-C-[SKD]-C-[GA]-[SDPAT]-x(0,1)-C-x-[CYF]
15piante
Da 45 a 84 AAs; due regioni ric he di Cys (dominio 1 edominio 3) separate da una regione con poche Cys(dominio 2)
p1, p2, p2v, p3, pec, p21
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 84 -
Struttura
• Nonostante che le sequenze aminoacidichesiano diverse hanno caratteristiche strutturalisimili:– Forma a manubrio,– Due domini,– Diverse unità tetraedriche Me(II)-Cys,– Tutte le Cys coinvolte nel legame con lo ione
metallico– Pressoché assente la struttura secondaria.

43
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 85 -
Cu-Metallotioneina da Saccharomyces Cerevisiae (1AQR)
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 86 -
Cu-Metallotioneina da Saccharomyces Cerevisiae (1AQR)

44
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 87 -
Cd-Metallotioneina da Riccio di mareSubunità α (1QJH)
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 88 -
Cd-Metallotioneina da Riccio di mareSubunità α (1QJH)
45
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 89 -
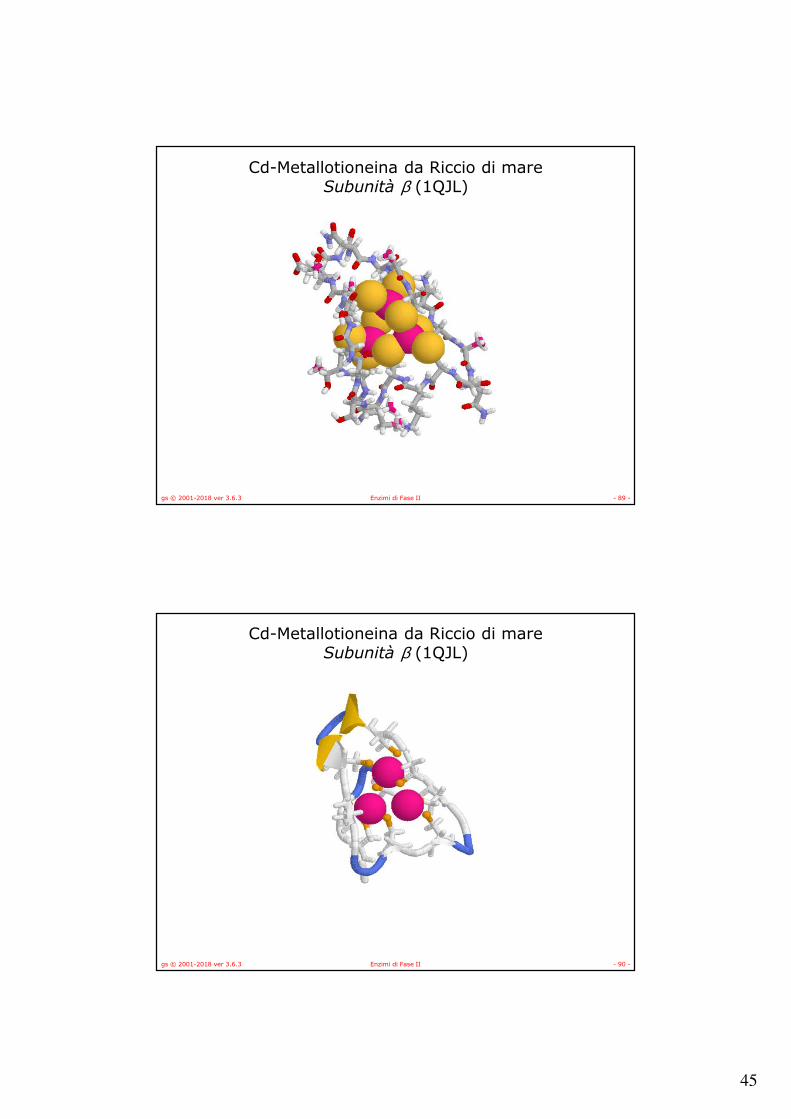
Cd-Metallotioneina da Riccio di mareSubunità β (1QJL)
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 90 -
Cd-Metallotioneina da Riccio di mareSubunità β (1QJL)

46
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 91 -
Cd-Metallotioneina umana (1MHU)
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 92 -
Cd-Metallotioneina umana (1MHU)

47
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 93 -
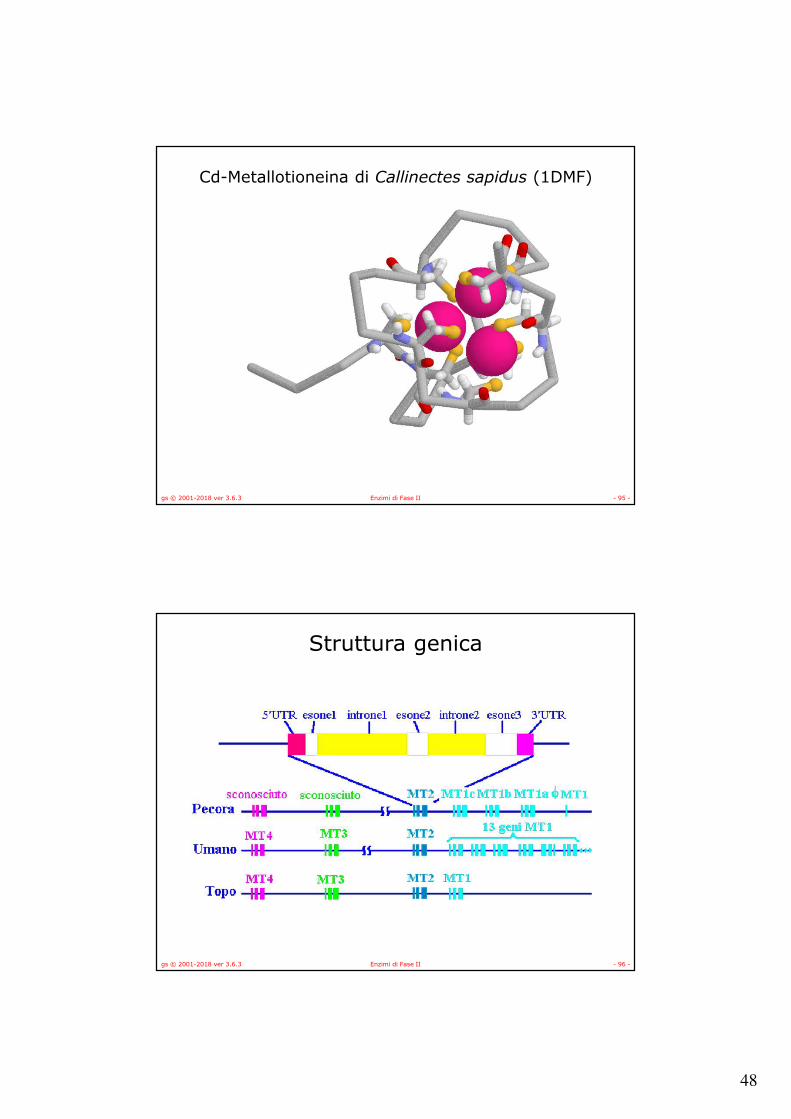
Cd-Metallotioneina di Callinectes sapidus (1DMF)
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 94 -
Cd-Metallotioneina di Callinectes sapidus (1DMF)
48
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 95 -
Cd-Metallotioneina di Callinectes sapidus (1DMF)
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 96 -
Struttura genica

49
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 97 -
Aspetti funzionali
• La più importante delle funzionalità delle metallotioneine è la loro inducibilità da una serie di agenti e condizioni:– Ioni metallici d10 (Cu++, Zn++, Cd++, Tl+, Pb++…)– Ormoni– Citochine– Fattori di crescita– Promotori tumorali– Stress
• È dimostrato il loro aumento fisiologico durante la proliferazione cellulare:
MT-Zn++ + Apo-Zinc-fingers → MT + Zn++- Zinc-fingers
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 98 -
Determinazione
• Le MT sono proteine ricche in cisteina con affinità elevata per i metalli pesanti
• La concentrazione di MT è quantificata valutando il contenuto in Cys
• Reazione di ELLMAN (standard GSH)

50
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 99 -
• Il glutatione è un tripeptide formato da glicina, cisteina, acido glutamico.
GSH
NH
NH
O
SHO
O
O
NH2
O
O
GLU CYS GLY
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 100 -
SS
NN
OO
OO
OO
S N
OO
OSR
SHN
OO
O
+RS +
ditiobisnitrobenzoato, DTNB
Reazione di ELLMAN
Assorbimento a 412 nm

51
gs © 2001-2018 ver 3.6.3 Enzimi di Fase II - 101 -
Referenze sul WEB
• Vie metaboliche – KEGG: http://www.genome.ad.jp/kegg/
• Degradazione degli xenobiotici: http://www.genome.ad.jp/kegg/pathway/map/map01196.html
• Struttura delle proteine: – Protein data bank (Brookhaven): http://www.rcsb.org/pdb/– Hexpasy
• Expert Protein Analysis System: http://us.expasy.org/sprot/• Prosite (protein families and domains): http://www.expasy.org/prosite/• Enzyme (Enzyme nomenclature database): http://www.expasy.org/enzyme/
– Scop (famiglie strutturali): http://scop.berkeley.edu/• Enzimi:
– Nomenclatura - IUBMB: http://www.chem.qmw.ac.uk/iubmb/– Proprietà - Brenda: http://www.brenda.uni-koeln.de/– Expasy (Enzyme nomenclature database): http://www.expasy.org/enzyme/
• Database di biocatalisi e biodegradazione: http://umbbd.ahc.umn.edu/• Citocromo P450: http://www.icgeb.org/~p450srv/• Metallotioneine: http://www.unizh.ch/~mtpage/MT.html• Tossicità degli xenobiotici: Agency for Toxic Substances and Disease Registry
http://www.atsdr.cdc.gov
Crediti e autorizzazioni all’utilizzo• Questo materiale è stato assemblato da informazioni raccolte dai seguenti testi di Biochimica:
– CHAMPE Pamela , HARVEY Richard , FERRIER Denise R. LE BASI DELLA BIOCHIMICA [ISBN 978-8808-17030-9] – Zanichelli
– NELSON David L. , COX Michael M. I PRINCIPI DI BIOCHIMICA DI LEHNINGER - Zanichelli – GARRETT Reginald H., GRISHAM Charles M. BIOCHIMICA con aspetti molecolari della Biologia
cellulare - Zanichelli– VOET Donald , VOET Judith G , PRATT Charlotte W FONDAMENTI DI BIOCHIMICA [ISBN 978-
8808-06879-8] - Zanichelli
• E dalla consultazione di svariate risorse in rete, tra le quali:– Kegg: Kyoto Encyclopedia of Genes and Genomes http://www.genome.ad.jp/kegg/– Brenda: http://www.brenda.uni-koeln.de/– Protein Data Bank: http://www.rcsb.org/pdb/– Rensselaer Polytechnic Institute:
http://www.rpi.edu/dept/bcbp/molbiochem/MBWeb/mb1/MB1index.html
• Il materiale è stato inoltre rivisto e corretto dalla Prof. Giancarla Orlandini dell’Università di Parma alla quale va il mio sentito ringraziamento.
Questo ed altro materiale può essere reperito a partire da: http://www. gsartor.org/pro
• Il materiale di questa presentazione è di libero uso per didattica e ricerca e può essere usato senza limitazione, purché venga riconosciuto l’autore usando questa frase:
Materiale ottenuto dal Prof. Giorgio Sartor
Università di Bologna
Giorgio SartorUfficiale: [email protected]: [email protected]
Aggiornato il 12/05/2018 00:43:46