changes in vascular extracellular matrix composition ... · 1 changes in vascular extracellular...
TRANSCRIPT

1
Changes in vascular extracellular matrix composition during decidual spiral arteriole
remodeling in early human pregnancy.
Samantha D Smith, PhD*1,2, Ruhul H. Choudhury, BSc*1,2, Patricia Matos, BSc1,2, James A
Horn BSc1,2, Stephen J Lye, PhD3, Caroline E Dunk, PhD3, John D Aplin, PhD1,2, Rebecca L 5
Jones, PhD1,2, Lynda K Harris, PhD1,2,4,5
1,2 Maternal and Fetal Health Research Centre (MFHRC), 1Institute of Human Development,
University of Manchester, 2St. Mary's Hospital, Central Manchester University Hospitals NHS
Foundation Trust, Manchester Academic Health Science Centre, Manchester, UK. 10
3Departments of Physiology and Obstetrics and Gynecology, Women’s and Infant’s Health,
Samuel Lunenfeld Research Institute, Mount Sinai Hospital, University of Toronto, Toronto,
Ontario, Canada.
4Manchester Pharmacy School, Stopford Building, Manchester, UK
5To whom all correspondence should be addressed. Maternal and Fetal Health Research Centre, 15
5th Floor (Research), St. Mary’s Hospital, Oxford Road, Manchester, M13 9WL, UK
Tel: 44 (0)161-276-6962
Fax: 44 (0)161-701-6971
20
*SDS and RHC contributed equally to this publication.
Pages: 33 Tables: 2 Figures: 7

2
Running Title: ECM in remodeling spiral arterioles
Keywords (up to 5): extracellular matrix, extravillous trophoblast, placenta, spiral arteriole, 25
vascular remodeling
Author contributions: SDS, SJL, CED, JDA, RLJ and LKH designed the experiments; SDS,
RHC, PM and JAH performed the research; SDS, RHC, PM, JAH and LKH analyzed the data;
SDS and LKH wrote the manuscript. All authors discussed the results and commented on the 30
manuscript.

3
Abstract (250 words)
Uterine spiral arteriole (SA) remodeling in early pregnancy involves a coordinated series of
events including decidual immune cell recruitment, vascular cell disruption and loss, and 35
colonization by placental-derived extravillous trophoblast (EVT). During this process, decidual
SA are converted from narrow, muscular vessels into dilated channels lacking vasomotor
control. We hypothesized that this extensive alteration in SA architecture must require
significant reorganization and/or breakdown of the vascular extracellular matrix (ECM). First
trimester decidua basalis (30 specimens) was immunostained to identify spiral arterioles 40
undergoing trophoblast-independent and -dependent phases of remodeling. Serial sections were
then immunostained for a panel of ECM markers, to examine changes in vascular ECM during
the remodeling process. The initial stages of SA remodeling were characterized by loss of
laminin, elastin, fibrillin, collagen types III, IV and VI from the basement membrane, vascular
media and/or adventitia, and surrounding decidual stromal cells. Loss of ECM correlated with 45
disruption and disorganization of vascular smooth muscle cells, and the majority of changes
occurred prior to extensive colonization of the vessel wall by EVT. The final stages of SA
remodeling, characterized by the arrival of EVT, were associated with the increased mural
deposition of fibronectin and fibrinoid. This study provides the first detailed analysis of the
spatial and temporal loss of ECM from the walls of remodeling decidual SA in early 50
pregnancy.
55

4
Introduction
Remodeling of uterine decidual spiral arterioles (SA) is essential for a successful pregnancy.
During this process, SA are transformed from narrow, muscularized vessels into dilated
channels, that ensures delivery of sufficient blood to the materno-fetal interface to meet the
needs of the developing fetus (Pijnenborg et al., 2006b; Burton et al., 2009). Impaired 60
remodeling is associated with an increased risk of pre-eclampsia, fetal growth restriction, pre-
term labor and stillbirth (Kim et al., 2003; Ball et al., 2006; Brosens et al., 2011). The process
of remodeling is complex, but ultimately involves loss of vascular smooth muscle cells
(VSMC), which are replaced by a fibrinoid matrix containing non-contractile, invading
extravillous trophoblast cells (EVT) derived from the developing placenta (Ashton et al., 2005; 65
Harris et al., 2006). Although complete physiological transformation of the SA is dependent on
the actions of EVT, changes in vessel architecture are observed prior to invasion (Craven et al.,
1998; Smith et al., 2009); these have been termed trophoblast-independent remodeling events
and are maternally regulated (Crocker et al., 2005; Pijnenborg et al., 2006b).
70
Our previous work has shown that decidual SA remodeling can be separated into four distinct
stages (Smith et al., 2009), and that some of these events can be reproduced using a decidual
and placental co-culture model (Hazan et al., 2010). Unremodeled SA, classified as stage I, are
characterized by an intact endothelium, tightly organized layers of circumferential VSMC and
no evidence of immune cell or EVT infiltration. Stage II SA exhibit endothelial cell 75
hypertrophy and loss, coupled with extensive disruption and partial loss of VSMC; these
changes are coincident with infiltration of the vessel wall by uterine natural killer cells (uNK)
and decidual macrophages. Stage III SA contain fewer decidual leukocytes and the majority of

5
vascular cells have been lost. Endovascular EVT are evident and some have adhered to the
arterial wall. Stage IV SA exhibit extensive or complete loss of VSMC and endothelium, and 80
are often lined with a continuous layer of EVT, although some endothelium may be retained.
Arrival of endovascular EVT in the SA is associated with the deposition of fibrinoid, a fibrin-
related, acellular, extracellular material, within the vascular wall. Fibrinoid is believed to confer
structural integrity to the remodeled vessels and provide a scaffold for EVT colonisation (Aplin
and Foden, 1982). Fibrinoid is a recognized hallmark of remodeled SA (Wells et al., 1984); 85
however, the timing of its deposition in relation to EVT invasion is unclear.
To ensure a permanent state of vasodilatation is achieved, the ECM within the decidual SA
must be reorganized in parallel with the disruption and loss of vascular cells. This likely
mediated by local proteases; we and others have shown that invading uNK, macrophages, 90
VSMC and EVT are a potent source of matrix metalloproteinases (MMP) (Smith et al., 2009;
Harris et al., 2010; Anacker et al., 2011; Robson et al., 2012). MMP-2, -7, -9 and -12 have
been localized to EVT in and around remodeling vessels in vivo (Jones et al., 2006; Plaisier et
al., 2008; Smith et al., 2009; Harris et al., 2010), and infiltrating uNK and decidual
macrophages secrete MMP-7 and -9 (Plaisier et al., 2008; Naruse et al., 2009; Smith et al., 95
2009).
Little is known about the specific ECM composition of decidual SA; however, healthy arterial
walls contain collagens, laminin and elastin, and proteoglycans such as decorin, versican and
perlecan (Jacob, 2003). Collagen types I, III and V are observed in large banded fibrils 100
throughout the intima, media and adventitia, whereas collagen type IV is a principal component

6
of the basal lamina to which endothelial cells and VSMC adhere (Shekhonin et al., 1985).
Laminin is also a major protein in basal lamina in the decidua and decidual SA (Charpin et al.,
1985; Wewer et al., 1985; Church et al., 1996), forming a cross-linked structure to which other
membrane and ECM molecules (such as collagen type IV) can bind. Other constituents of the 105
decidual vascular ECM in humans and rodents include collagen type VI, heparan sulphate
proteoglycan, fibrillin and fibronectin (Aplin et al., 1988; Earl et al., 1990; Mulholland et al.,
1992; Dziadek et al., 1995); many of these components can be synthesized by resident vascular
cells (Harris and Aplin, 2007). Unlike the myometrial spiral arteries, which exhibit a well
formed elastic lamina together with concentric mural layers of elastin (Khong et al., 2003), the 110
decidual SA are not believed to contain elastin in the form of a true elastic lamina.
We hypothesized that decidual leukocytes act in conjunction with invading EVT to orchestrate
ECM remodeling within the decidual SA, facilitating the process of vascular transformation. In
the current study, we sought to characterize the ECM composition of intact decidual SA and 115
document changes in ECM architecture in relation to VSMC loss and EVT infiltration.

7
Materials and methods
Tissue collection
Placental and decidual tissue (n = 30) were obtained from women undergoing elective surgical 120
termination of pregnancy at St. Mary’s Hospital, Manchester for social reasons in the first
trimester (6-12 weeks gestation). Written informed consent was obtained from all patients and
ethical approval was obtained from Central Manchester Research Ethics Committee
(03/CM/031) and the North West Research Ethics Committee (08/H1010/28;
08/H1010/55(+5)). 125
Tissue processing and fixation
Decidua and placenta were separated following gross morphological examination and were
randomly sampled. Multiple biopsies of decidua basalis (10-20mm3) were washed in Tris-
buffered saline (TBS) and fixed in 10% neutral buffered formalin at 4°C for 24h, prior to 130
routine histological processing and embedding in paraffin wax.
Immunohistochemistry for vascular and ECM markers
Serial sections of wax-embedded decidua basalis (5µm) were deparaffinized in Histoclear and
alcohol, rehydrated and washed in dH2O. To facilitate antigen unmasking, slides were 135
submerged in sodium citrate buffer (275ml; 0.01M containing 0.05% (v/v) Tween 20, pH 6.0;
room temperature), the container was covered with plastic film to prevent evaporation and
microwaved for 2 X 5 min. Slides were left to cool for 20 min, then washed in cold dH2O for 5
min. Sections to be immunostained for collagen type III were treated differently: slides were
incubated with 1% (w/v) trypsin (Type II-S from porcine pancreas; 1000-2000 units/mg; Sigma 140

8
Aldrich) in 10mM calcium chloride for 20 min at 37°C, then washed in cold dH2O for 5 min.
The staining pattern of every antibody was tested with and without antigen retrieval; antigen
retrieval was only used where necessary, or if it did not affect the staining pattern and was
required for another antibody on a serial section. Endogenous peroxide activity was blocked by
incubating tissue sections in 3% (v/v) hydrogen peroxide for 10 min. Tissue sections were 145
washed three times in 0.05M Tris buffered saline (TBS) and incubated with non-immune block
(10% goat serum (Sigma Aldrich, UK), 2% human serum (in house) and 0.1% Tween-20 in
TBS) to inhibit non-specific antibody binding. Primary antibodies, diluted to the required
concentration in TBS (detailed in Table 1), were applied to the tissue sections, which were
incubated overnight at 4oC in a humidity chamber. Antibody concentrations were optimized 150
prior to characterization of the tissue samples. Negative controls (non-immune or isotype
control IgG, as appropriate) were also included; concentrations were matched to the
corresponding primary antibodies. Slides were washed (3 X 5 min; TBS) and the secondary
antibodies, diluted in TBS (Table 1), were applied for 30 min at room temperature. Slides were
washed (3 X 5 min; TBS) and incubated with avidin peroxidase (5µg/ml in TBS; Sigma 155
Aldrich) for 30 min at room temperature, then further washed in TBS and incubated for 1-5
min with 0.05% (w/v) DAB and 0.015% (v/v) hydrogen peroxide (Sigma Aldrich). All tissue
sections stained with the same antibody were incubated with DAB for the same length of time,
so that comparisons could be made between individual samples. Finally, slides were washed in
dH2O, counterstained with Harris’ hematoxylin (Sigma Aldrich), dehydrated in alcohol and 160
Histoclear, and mounted in DPX (Sigma Aldrich).

9
Periodic Acid Schiff reaction
The Periodic Acid Schiff (PAS) reaction was used to examine fibrinoid deposition in the
decidua; the reaction was performed using a commercially available kit (Sigma Aldrich). 165
Sections of decidua basalis were dewaxed in Histoclear, rehydrated and washed in dH2O.
Periodic acid (1g/dl) was applied to sections for 5 min, which were then rinsed with dH2O, and
incubated with Schiffs reagent for 10 min. After further dH2O washes, sections were
counterstained with Harris’ hematoxylin, dehydrated in alcohol and Histoclear and mounted in
DPX. 170
Microscopy and image analysis
A Leitz Dialux 22 microscope was used in conjunction with a QI Cam Fast 1394 camera and
Image ProPlus 6.0 imaging software; exposure times were matched at image capture. To select
representative spiral arteriole sections for display in each figure, stained slides and captured 175
images were reviewed and discussed by a minimum of 4 authors. Semi-quantitative assessment
of DAB staining intensity was performed by two independent observers: immunostained
samples were assigned a score of − (no staining), + mild staining, ++ moderate staining or +++
(intense staining). Slides were stained in large batches to minimize inter-experimental
variation; comparisons were only made between tissues stained in the same run. 180

10
Results
Changes in decidual vascular ECM during SA remodeling
Initial immunohistochemical analysis of archived decidual tissue was performed using 185
antibodies to smooth muscle actin and HLA-G to identify samples containing remodeling spiral
arterioles. More detailed analysis identified samples of decidua basalis from 30 pregnancies
that each contained examples of unremodeled vessels, vessels colonised by CD45-positive
leukocytes and vessels colonised by HLA-G-positive endovascular trophoblast, as previously
described (Smith et al., 2009). Individual arterioles did not exist in disparate stages of 190
remodeling; the majority of individual tissue sections contained multiple profiles of individual
SA as they spiraled through the tissue, where the proximal end was remodeled and the distal
end was not. Thus, within a single tissue section it was possible to observe profiles of
individual, dilated, trophoblast-lined SA in the proximal decidua (identified by remnants of
placental villi attached to the decidual surface), which could be traced into the distal decidua. 195
The profiles of these distal SA segments exhibited VSMC disruption in the absence of EVT,
and deeper still exhibited intact VSMC layers; thus all stages of remodeling could be observed
within profiles of a single SA. Tracing individual SA through the placental bed as progressive
remodeling occurred ensured that any remodeling vessel that was imaged and analyzed was a
distal portion of an intact SA rather than a vein or a gland; veins were excluded by their 200
morphology (thin layers of VSMCs, intact flattened endothelium, dilated lumens). Every
patient sample contained a minimum of two arterioles in all stages of remodeling. Using these
carefully characterized samples, immunostaining for collagen types III, IV, VI, laminin,
fibronectin, fibrillin and elastin was performed on serial sections, together with markers of
EVT and VSMC, to examine temporal changes in vascular ECM during the remodeling 205

11
process. The reported changes in ECM composition in remodeling SA were consistent across
all samples and gestational ages examined. There was no correlation between the gestational
age of the sample and the proportion of arterioles in each stage of remodeling, as previously
observed (Smith et al., 2009).
210
Collagen type III
Intense pericellular staining of collagen type III was observed in the decidual stroma (Fig 1A;
a). In unremodeled stage I SA (Fig 1C), collagen type III was detected in a thick band (b)
within the adventitia, but was absent from medial VSMC, endothelial cells and the endothelial
basement membrane. In stage II and III remodeling arterioles (Fig 1D, E), which display 215
disruption, disorganization and loss of VSMC, adventitial expression was reduced and fibers
appeared disrupted (c). Pericellular staining of adjacent decidual stromal cells was also reduced
in some areas. In stage IV remodeling arterioles (Fig 1F), which had been heavily colonized by
endovascular EVT, collagen type III immunoreactivity was markedly diminished or absent
from the vessel wall, but was observed in the basement membrane of SA (d) and in the decidual 220
stroma. Decidual stromal cell expression was also reduced in areas of interstitial EVT
infiltration (Fig 1E; e).
Collagen type IV
Intense pericellular staining for collagen type IV was detected around the periphery of decidual 225
stromal cells (Fig 2A; a). In unremodeled stage I SA (Fig 2C), collagen fibers were detected in
tightly bound, organized layers (b). Intense staining was detected in the basement membrane
(BM) underlying the vascular endothelium, and transverse and longitudinal layers of collagen

12
type IV were evident surrounding medial VSMC (Fig 2C). In stage II and III remodeling
arterioles (Fig 2D, E), collagen type IV expression within the vessel wall was weak or absent, 230
especially in areas of VSMC loss. Collagen type IV was still present in the basement membrane
in stage II, but in reduced quantities and gaps (c) were evident (Fig 2D). In stage IV
remodeling arterioles (Fig 2F), collagen type IV expression in the arterial wall was minimal.
Weak staining extending into the stroma was observed; however, pericellular decidual staining
was retained. 235
Collagen type VI
Areas of intense pericellular staining for collagen type VI were observed around the periphery
of a proportion of decidual stromal cells (Fig 3A; a). Although collagen type VI was evident in
the adventitia and outermost VSMC of unremodeled SA (b), and some areas of basement 240
membrane (BM) were weakly immunopositive, it was absent from the inner layers of medial
VSMC (Fig 3C, D; c). Collagen type VI was not observed in VSMC of remodeling arterioles;
however, some endovascular EVT within stage VI SA were immunopositive (Fig 3F; d).
Laminin 245
Pericellular laminin staining of varying intensity was detected in decidual stromal cells (Fig
4A, B; a). Within stage I unremodeled SA (Fig 4D), laminin was detected in the basement
membrane (BM) and weak staining was noted between layers of VSMC (b). In stages II- IV
(Fig 4E-G), laminin staining was very weak or absent in the vessel wall. Laminin was also
absent from endovascular EVT (Fig 4G). 250

13
Fibronectin
Pericellular fibronectin staining was observed in the decidual stroma (Fig 5A; a). In stage I
unremodeled SA (Fig 5C), fibronectin staining was observed in the outmost layers of medial
VSMC (b). In stage II and III remodeling SA (Fig 5D, E) weak, diffuse fibronectin 255
immunoreactivity was observed in some areas of the arterial wall. Interstitial (c) and
endovascular (d) EVT were immunopositive for fibronectin (Fig 5E) and in stage III and IV
remodeled arterioles (Fig 5F), the majority of luminal EVT were also positive for fibronectin
(e).
260
Fibrillin
In stage I unremodeled SA (Fig 6A), fibrillin was expressed in the adventitia and medial
VSMC (a), although discrete layers were not observed. Moderate levels of fibrillin staining
were observed throughout the decidual stroma (Fig 6A, D; b). Fibrillin expression or
localization was not significantly changed in the walls of stage II, III (not shown) or IV 265
remodeling SA (Fig 6B); however, interstitial and endovascular EVT exhibited pericellular
fibrillin expression (Fig 6B, c).
Elastin
Whilst organized, concentric layers of elastin were absent from unremodeled SA, discrete areas 270
of elastin immunoreactivity were observed within the inner layers of VSMC and areas
corresponding to the internal elastic lamina (Fig 6E; d). Decidual cells were also weakly
immunopositive for elastin (e). Following the onset of remodeling, the amount of elastin within
the vascular wall was reduced. In stages II, III and IV, weak elastin immunoreactivity was

14
observed within VSMC, EVT and decidual stromal cells and in some parts of the endothelial 275
basement membrane (Fig 6G; BM), but distinct fibers were no longer seen.
Fibrinoid
Fibrinoid deposition within the decidua was examined using PAS staining. PAS detects
glycogen, elastic fibril-associated glycoprotein and other highly glycosylated ECM as intense 280
pink staining of acellular material. The basement membrane (BM) of stage I SA, which is rich
in glycosylated molecules, was detected as organized bands of PAS-positive material which
encompassed the inner layers of medial VSMC (Fig 7A). PAS also labeled the decidual stromal
cell basement membrane (Fig 7A, B; a). Stage II and III remodeling SA, which exhibit
disruption and loss of VSMC layers (Fig 7B’), stained weakly with PAS (Fig 7B; c). PAS-285
positive fibrinoid present around transformed vessels is thicker and lacks a fibrillar appearance
(c); stage IV SA, which are characterized by extensive endovascular EVT infiltration (Fig
7C’’), were surrounded by areas of diffuse pink staining, indicative of fibrinoid deposition (Fig
7C). Weakly positive intraluminal and intravascular EVT were observed within these vessels
(Fig 7C; d). 290

15
Discussion
This study aimed to characterize the changes in vascular ECM composition during remodeling
of the decidual SA in early pregnancy. Here we confirm previous reports that collagen type IV 295
and laminin are major components of unremodeled vessels (Charpin et al., 1985; Wewer et al.,
1985; Aplin et al., 1988), and demonstrate that they are localized to VSMC and endothelial
basement membranes. We show that collagen types III and VI are present within the adventitia
of intact decidual SA, fibrillin is expressed by medial VSMC, and elastin and fibronectin are
observed in the medial ECM. 300
It has long been recognized that the initial stages of decidual SA remodeling involve severe
disruption and disorganization of the VSMC layers, which is accompanied by endothelial
swelling and basophilia (Craven et al., 1998; Kam et al., 1999); we have previously shown that
these changes are coincident with infiltration of decidual leukocytes (Smith et al., 2009). Here 305
we demonstrate that these alterations in vascular cell morphology are accompanied by
significant reorganization and/or degradation of the vascular ECM. VSMC within SA are
encapsulated within a basement membrane containing collagen type IV and laminin, as seen in
unremodeled stage I vessels. Fibronectin, fibrillin and small elastic fibrils are also evident as
pericellular deposits in some areas of VSMC at stage I. In normally functioning arterioles, cells 310
and ECM act as a unit for the transduction of mechanical forces (Martinez-Lemus, 2012). As a
major component of the decidual vascular ECM, collagen type IV degradation is likely to be
required to facilitate disruption of medial VSMC layers. Indeed, in the early stages of
remodeling where vascular cell layers are disrupted, collagen type IV fibers become
disorganized but remain abundant. We speculate that this initial disruption and ‘loosening’ of 315

16
the ECM, probably induced by local release of proteases and cytokines (Robson et al., 2012),
facilitates colonization by decidual leukocytes (Smith et al., 2009), which may have the
capacity to effect further ECM degradation within the vascular wall (Robson et al., 2012).
Collagen type VI is one of the first ECM components to be lost from stage II remodeling SA. 320
In earlier work we suggested that perivascular collagen type VI plays a role in integrating blood
vessels into the surrounding decidual stroma (Mulholland et al., 1992; Mylona et al., 1995). As
it is localized to the adventitia and outermost layers of VSMC, the first point of contact for
invading decidual leukocytes and EVT, it is the first layer of ECM that must be breached to
allow access to the underlying vascular cells. Similarly, expression of collagen type III and 325
laminin were greatly reduced in stage II, concomitant with evidence of VSMC disruption.
In stage III arterioles, which exhibit major disruption and loss of VSMC and endothelium,
collagen type IV distribution appeared much weaker, and in some areas was visible as
cytoplasmic reactivity in migrating, disrupted VSMC, suggesting degradation of the previously 330
highly organized fibers. In stage IV arterioles, expression of collagen type IV was in the
vascular wall was very low. The fate of the collagen type IV fibers between stages II and IV
remains unknown; dispersed fibers within the stroma may be phagocytosed by macrophages,
which are still present in and around the SA wall in the later stages of remodeling (Smith et al.,
2009). Alternatively, fragments could be further catabolized into smaller non-immunoreactive 335
peptides and may be cleared by the circulation or become incorporated into fibrinoid. Indeed,
the majority of vascular ECM in stage IV SA consisted of PAS-positive material; however,
varying levels of EVT-associated fibronectin, fibrillin, collagen type III and collagen type IV

17
were also evident. A qualitative summary of the progressive loss of ECM components from the
vascular wall during SA remodeling is given in Table 2. 340
Placental fibrinoid has been classified into two broad types: matrix-type fibrinoid and fibrin-
type fibrinoid, although both types are usually deposited together (Kaufmann et al., 1996).
Matrix-type fibrinoid, in which endovascular EVT are embedded, is secreted by endovascular
EVT and reportedly contains fragments of laminin, collagen type IV, heparan sulfate, 345
vitronectin and oncofetal fibronectin. In this study, we used PAS staining to identify the
presence of both forms of fibrinoid, and we have not differentiated between the two. As
observed in primates (Pijnenborg et al., 1996), we demonstrate that fibrinoid is only observed
in the walls of human SA in stage IV when EVT are present. This is consistent both with data
from our in vitro human tissue co-cultures (Hazan et al., 2010), where fibrinoid was not 350
detected (unpublished observations), and the work of others who have shown fibrinoid
deposition to be a late feature of remodeling (Brosens et al., 1967), and is associated with the
presence of interstitial EVT (Kam et al., 1999). Alternatively, the presence of plasma
components in interstitial fluid may be required for fibrin-type fibrinoid deposition. The exact
role of fibrinoid within remodeled SA is unproven but several functions have been proposed. It 355
supports the adhesion of trophoblasts (Aplin and Foden, 1982), and it is likely that during
destruction of the vascular wall and loss of the stability and support provided by the vascular
ECM, it is deposited to maintain structural integrity of the resulting channels. Secretions from
EVT embedded in the vascular wall probably contribute to fibrinoid, and its secretion has also
been detected in the path of interstitial EVT invasion (Pijnenborg et al., 2006a). Differences in 360
the composition of endovascular EVT- and interstitial EVT-derived fibrinoid have previously

18
been noted (Pijnenborg et al., 2006b); further work is still required to define its full range of
functions.
We have previously observed that localized disruption of the vessel wall corresponds with foci 365
of apoptotic vascular cells in remodeling SA (Smith et al., 2009; Hazan et al., 2010). Disruption
of ECM-adhesion molecule interactions that maintain VSMC in situ may also result in
cessation of vital cell survival signals, such as integrin-mediated signaling (Frisch and Francis,
1994). Loss of attachment of adherent cells to the ECM triggers a form of apoptotic cell death,
known as anoikis, which may represent an alternative mechanism for vascular cell loss during 370
active SA remodeling. In addition, healthy VSMC have been observed at a considerable
distance away from actively remodeling SA, suggesting that some VSMC may migrate into the
decidua following their release from the ECM. This could lead to dedifferentiation of VSMC
into myofibroblasts or pericytes and facilitate VSMC dispersal, as previously hypothesized
(Robson et al., 2009). 375
The majority of ECM constituents underwent significant catabolism in the early stages of SA
remodeling, prior to the arrival of endovascular EVT; this is the time when leukocytes infiltrate
the vascular wall (Smith et al., 2009). An increasing body of evidence suggests that decidual
uNK and macrophages actively participate in SA remodeling, but the specific effector functions 380
of these cells are still unclear. Soluble factors produced by uNK include angiopoietin-1,
angiopoietin-2, interferon-gamma and VEGF-C, (Lash et al., 2006; Lash et al., 2011); these
factors have been shown to induce VSMC disorganization and ECM breakdown in isolated
segments of human myometrial SA (Robson et al., 2012). Leukocytes are also a source of

19
MMP, which are likely mediators of ECM degradation in decidua and the arterial wall. Indeed, 385
inhibition of MMP action in the mouse uterus causes major disruption of decidualization
(Alexander et al., 1996). In other vascular beds, and indeed in cycling non-pregnant
endometrium, leukocytes are believed to be major effectors of tissue remodeling, through
production of MMP-3, -7 and -9 (Salamonsen and Lathbury, 2000; Kaitu'u et al., 2005), and
inflammatory mediators such as TGF-β and chemokines (Salamonsen and Lathbury, 2000). 390
EVT are also believed to participate in the degradation of vascular ECM, by producing MMP
capable of cleaving ECM components in the vascular wall (Isaka et al., 2003; Jones et al.,
2006). EVT are known to express a diverse range of MMP in the first trimester of pregnancy
(Anacker et al., 2011), and MMP-2 and -9 are differentially expressed by interstitial and
endovascular EVT (Jones et al., 2006), suggesting they have different functions that may relate 395
to their distinct environments. VSMC resident within remodeling SA have also been identified
as a source of MMP and may contribute to local ECM remodeling (Harris et al., 2010).
In summary, this study has provided a detailed analysis of the temporal loss of ECM from the
walls of remodeling decidual SA in human pregnancy and highlighted that the majority of 400
ECM breakdown occurs as an early event in the remodeling timeline. Whilst this observational
study cannot directly correlate leukocyte infiltration into stage II remodeling SA with ECM
breakdown, our findings suggest that a significant proportion of ECM reorganization occurs
during the trophoblast-independent phase of SA remodeling. We also speculate that the
impaired endovascular EVT invasion and incomplete SA remodeling observed in pre-eclampsia 405
could be ascribed to a reduction in ECM turnover, perhaps due to insufficient number or
activity of decidual uNK, macrophages or EVT. Alternatively, an inherent resistance of

20
maternal vascular ECM to degradation may be the cause, in addition to other defects in
trophoblast invasion; however, more detailed investigations will be required to further elucidate
this complex series of remodeling events. 410

21
Acknowledgements
We thank the research midwives at St. Mary’s Hospital, Manchester for their invaluable help
with subject recruitment and tissue collection. This work was funded by Tommy’s the Baby 415
Charity, The MRC and The Canadian Institute of Health Research. The MFHRC is supported
by Tommy’s, an Action Research Endowment Fund, the Manchester BRC and the Greater
Manchester CLRN. LKH is a BBSRC David Phillips Research Fellow.
420

22
References
Alexander C.M., Hansell E.J., Behrendtsen O., Flannery M.L., Kishnani N.S., Hawkes S.P. and
Werb Z. (1996). Expression and function of matrix metalloproteinases and their inhibitors at
the maternal-embryonic boundary during mouse embryo implantation. Development. 122,
1723-1736. 425
Anacker J., Segerer S.E., Hagemann C., Feix S., Kapp M., Bausch R. and Kammerer U. (2011).
Human decidua and invasive trophoblasts are rich sources of nearly all human matrix
metalloproteinases. Mol. Hum. Reprod. 17, 637-652.
Aplin J.D. and Foden L.J. (1982). A cell spreading factor, abundant in human placenta,
contains fibronectin and fibrinogen. J. Cell Sci. 58, 287-302. 430
Aplin J.D., Charlton A.K. and Ayad S. (1988). An immunohistochemical study of human
endometrial extracellular matrix during the menstrual cycle and first trimester of pregnancy.
Cell Tissue Res. 253, 231-240.
Ashton S.V., Whitley G.S., Dash P.R., Wareing M., Crocker I.P., Baker P.N. and Cartwright
J.E. (2005). Uterine spiral artery remodeling involves endothelial apoptosis induced by 435
extravillous trophoblasts through Fas/FasL interactions. Arterioscler. Thromb. Vasc. Biol. 25,
102-108.
Ball E., Bulmer J.N., Ayis S., Lyall F. and Robson S.C. (2006). Late sporadic miscarriage is
associated with abnormalities in spiral artery transformation and trophoblast invasion. J. Pathol.
208, 535-542. 440
Brosens I., Robertson W.B. and Dixon H.G. (1967). The physiological response of the vessels
of the placental bed to normal pregnancy. J. Pathol. Bacteriol. 93, 569-579.

23
Brosens I., Pijnenborg R., Vercruysse L. and Romero R. (2011). The "Great Obstetrical
Syndromes" are associated with disorders of deep placentation. Am. J. Obstet. Gynecol. 204,
193-201. 445
Burton G.J., Woods A.W., Jauniaux E. and Kingdom J.C. (2009). Rheological and
physiological consequences of conversion of the maternal spiral arteries for uteroplacental
blood flow during human pregnancy. Placenta. 30, 473-482.
Charpin C., Kopp F., Pourreau-Schneider N., Lissitzky J.C., Lavaut M.N., Martin P.M. and
Toga M. (1985). Laminin distribution in human decidua and immature placenta. An 450
immunoelectron microscopic study (avidin-biotin-peroxidase complex method). Am. J. Obstet.
Gynecol. 151, 822-826.
Church H.J., Vicovac L.M., Williams J.D., Hey N.A. and Aplin J.D. (1996). Laminins 2 and 4
are expressed by human decidual cells. Lab. Invest. 74, 21-32.
Craven C.M., Morgan T. and Ward K. (1998). Decidual spiral artery remodelling begins before 455
cellular interaction with cytotrophoblasts. Placenta. 19, 241-252.
Crocker I.P., Wareing M., Ferris G.R., Jones C.J., Cartwright J.E., Baker P.N. and Aplin J.D.
(2005). The effect of vascular origin, oxygen, and tumour necrosis factor alpha on trophoblast
invasion of maternal arteries in vitro. J. Pathol. 206, 476-485.
Dziadek M., Darling P., Zhang R.Z., Pan T.C., Tillet E., Timpl R. and Chu M.L. (1995). 460
Expression of collagen alpha 1(VI), alpha 2(VI), and alpha 3(VI) chains in the pregnant mouse
uterus. Biol. Reprod. 52, 885-894.
Earl U., Estlin C. and Bulmer J.N. (1990). Fibronectin and laminin in the early human placenta.
Placenta. 11, 223-231.

24
Frisch S.M. and Francis H. (1994). Disruption of epithelial cell-matrix interactions induces 465
apoptosis. J. Cell Biol. 124, 619-626.
Harris L.K., Keogh R.J., Wareing M., Baker P.N., Cartwright J.E., Aplin J.D. and Whitley G.S.
(2006). Invasive trophoblasts stimulate vascular smooth muscle cell apoptosis by a fas ligand-
dependent mechanism. Am. J. Pathol. 169, 1863-1874.
Harris L.K. and Aplin J.D. (2007). Vascular remodeling and extracellular matrix breakdown in 470
the uterine spiral arteries during pregnancy. Reprod. Sci. 14, 28-34.
Harris L.K., Smith S.D., Keogh R.J., Jones R.L., Baker P.N., Knofler M., Cartwright J.E.,
Whitley G.S. and Aplin J.D. (2010). Trophoblast- and Vascular Smooth Muscle Cell-Derived
MMP-12 Mediates Elastolysis during Uterine Spiral Artery Remodeling. Am. J. Pathol. 177,
2103-2115. 475
Hazan A.D., Smith S.D., Jones R.L., Whittle W., Lye S.J. and Dunk C.E. (2010). Vascular-
leukocyte interactions: mechanisms of human decidual spiral artery remodeling in vitro. Am. J.
Pathol. 177, 1017-1030.
Isaka K., Usuda S., Ito H., Sagawa Y., Nakamura H., Nishi H., Suzuki Y., Li Y.F. and
Takayama M. (2003). Expression and activity of matrix metalloproteinase 2 and 9 in human 480
trophoblasts. Placenta. 24, 53-64.
Jacob M.P. (2003). Extracellular matrix remodeling and matrix metalloproteinases in the
vascular wall during aging and in pathological conditions. Biomed. Pharmacother. 57, 195-202.
Jones R.L., Findlay J.K., Farnworth P.G., Robertson D.M., Wallace E. and Salamonsen L.A.
(2006). Activin A and inhibin A differentially regulate human uterine matrix 485
metalloproteinases: potential interactions during decidualization and trophoblast invasion.
Endocrinology. 147, 724-732.

25
Kaitu'u T.J., Shen J., Zhang J., Morison N.B. and Salamonsen L.A. (2005). Matrix
metalloproteinases in endometrial breakdown and repair: functional significance in a mouse
model. Biol. Reprod. 73, 672-680. 490
Kam E.P., Gardner L., Loke Y.W. and King A. (1999). The role of trophoblast in the
physiological change in decidual spiral arteries. Hum. Reprod. 14, 2131-2138.
Kaufmann P., Huppertz B. and Frank H.G. (1996). The fibrinoids of the human placenta:
origin, composition and functional relevance. Ann. Anat. 178, 485-501.
Khong T.Y., Adema E.D. and Erwich J.J. (2003). On an anatomical basis for the increase in 495
birth weight in second and subsequent born children. Placenta. 24, 348-353.
Kim Y.M., Bujold E., Chaiworapongsa T., Gomez R., Yoon B.H., Thaler H.T., Rotmensch S.
and Romero R. (2003). Failure of physiologic transformation of the spiral arteries in patients
with preterm labor and intact membranes. Am. J. Obstet. Gynecol. 189, 1063-1069.
Lash G.E., Schiessl B., Kirkley M., Innes B.A., Cooper A., Searle R.F., Robson S.C. and 500
Bulmer J.N. (2006). Expression of angiogenic growth factors by uterine natural killer cells
during early pregnancy. J. Leukoc. Biol. 80, 572-580.
Lash G.E., Naruse K., Robson A., Innes B.A., Searle R.F., Robson S.C. and Bulmer J.N.
(2011). Interaction between uterine natural killer cells and extravillous trophoblast cells: effect
on cytokine and angiogenic growth factor production. Hum. Reprod. 26, 2289-2295. 505
Martinez-Lemus L.A. (2012). The dynamic structure of arterioles. Basic Clin. Pharmacol.
Toxicol. 110, 5-11.
Mulholland J., Aplin J.D., Ayad S., Hong L. and Glasser S.R. (1992). Loss of collagen type VI
from rat endometrial stroma during decidualization. Biol. Reprod. 46, 1136-1143.

26
Mylona P., Kielty C.M., Hoyland J.A. and Aplin J.D. (1995). Expression of type VI collagen 510
mRNAs in human endometrium during the menstrual cycle and first trimester of pregnancy. J.
Reprod. Fertil. 103, 159-167.
Naruse K., Lash G.E., Innes B.A., Otun H.A., Searle R.F., Robson S.C. and Bulmer J.N.
(2009). Localization of matrix metalloproteinase (MMP)-2, MMP-9 and tissue inhibitors for
MMPs (TIMPs) in uterine natural killer cells in early human pregnancy. Hum. Reprod. 24, 553-515
561.
Pijnenborg R., D'Hooghe T., Vercruysse L. and Bambra C. (1996). Evaluation of trophoblast
invasion in placental bed biopsies of the baboon, with immunohistochemical localisation of
cytokeratin, fibronectin, and laminin. J. Med. Primatol. 25, 272-281.
Pijnenborg R., Ball E., Bulmer J.N., Hanssens M., Robson S.C. and Vercruysse L. (2006a). In 520
vivo analysis of trophoblast cell invasion in the human. Methods Mol. Med. 122, 11-44.
Pijnenborg R., Vercruysse L. and Hanssens M. (2006b). The uterine spiral arteries in human
pregnancy: facts and controversies. Placenta. 27, 939-958.
Plaisier M., Koolwijk P., Willems F., Helmerhorst F.M. and van Hinsbergh V.W. (2008).
Pericellular-acting proteases in human first trimester decidua. Mol. Hum. Reprod. 14, 41-51. 525
Robson A., Lash G.E., Innes B.A., Robson S.C. and Bulmer J.N. (2009). Potential Role of
VEGF-C, Ang2 and IFN-gamma in Spiral Artery Remodelling. Reproductive Sciences. 16,
110A-111A (146).
Robson A., Harris L.K., Innes B.A., Lash G.E., Aljunaidy M.M., Aplin J.D., Baker P.N.,
Robson S.C. and Bulmer J.N. (2012). Uterine natural killer cells initiate spiral artery 530
remodeling in human pregnancy. FASEB J. 26, 4876-4885.

27
Salamonsen L.A. and Lathbury L.J. (2000). Endometrial leukocytes and menstruation. Hum.
Reprod. Update. 6, 16-27.
Shekhonin B.V., Domogatsky S.P., Muzykantov V.R., Idelson G.L. and Rukosuev V.S. (1985).
Distribution of type I, III, IV and V collagen in normal and atherosclerotic human arterial wall: 535
immunomorphological characteristics. Coll. Relat. Res. 5, 355-368.
Smith S.D., Dunk C.E., Aplin J.D., Harris L.K. and Jones R.L. (2009). Evidence for immune
cell involvement in decidual spiral arteriole remodeling in early human pregnancy. Am. J.
Pathol. 174, 1959-1971.
Wells M., Hsi B.L., Yeh C.J. and Faulk W.P. (1984). Spiral (uteroplacental) arteries of the 540
human placental bed show the presence of amniotic basement membrane antigens. Am. J.
Obstet. Gynecol. 150, 973-977.
Wewer U.M., Faber M., Liotta L.A. and Albrechtsen R. (1985). Immunochemical and
ultrastructural assessment of the nature of the pericellular basement membrane of human
decidual cells. Lab. Invest. 53, 624-633. 545

28
Figure legends
Figure 1. Serial sections showing immunolocalization of collagen type III in the decidua 550
basalis and spiral arterioles during remodeling in vivo. (A) Collagen (col) type III in the
decidual stroma. (B) Control IgG. (C) Col III expression in unremodeled (stage I) SA; (C’) α-
smooth muscle actin (α-SMA; VSMC); (C’’) HLA-G (EVT). (D) Col III expression in
remodeling (stage II) SA; (D’) α-SMA; (D’’) HLA-G. (E) Col III expression in remodeling
(stage III) SA; (E’) α-SMA; (E’’) HLA-G. (F) Col III expression in remodeled (stage IV) SA; 555
(F’) α-SMA; (F’’) HLA-G. Scale bar = 50µm. Inset = negative control. L = lumen; a =
pericellular col III staining; b = bands of col III; c = disrupted col III fibers; d =
immunoreactivity in the basement membrane; e = reduced col III expression in areas colonized
by interstitial EVT. Serial sections from n=20 individual decidual samples (6-12 weeks
gestation) and a minimum of n=24 vessel sections in each remodeling stage were assessed. 560
Representative images are shown.
Figure 2. Serial sections showing immunolocalization of collagen type IV in the decidua
basalis and spiral arterioles during remodeling in vivo. (A) Collagen (col) type IV in the
decidual stroma. (B) Control IgG. (C) Collagen (col) type IV expression in unremodeled (stage 565
I) SA; (C’) α-SMA (VSMC); (C’’) HLA-G (EVT). BM = basement membrane. (D) Col IV
expression in remodeling (stage II) SA; (D’) α-SMA; (D’’) HLA-G. (E) Col IV expression in
remodeling (stage III) SA; (E’) α-SMA; (E’’) HLA-G. (F) Col IV expression in remodeled
(stage IV) SA; (F’) α-SMA; (F’’) HLA-G. Scale bar = 50µm. Inset = negative control. L =
lumen; a = pericellular col IV staining; b = layers of col IV; BM = basement membrane; c = 570
gaps in the basement membrane. Serial sections from n=24 individual decidual samples (6-12

29
weeks gestation) and a minimum of n=32 vessel sections in each remodeling stage were
assessed. Representative images are shown.
Figure 3. Serial sections showing immunolocalization of collagen type VI in the decidua 575
basalis and spiral arterioles during remodeling in vivo. (A) Collagen (col) type VI in the
decidual stroma. (B) Control IgG. (C) Col VI expression in unremodeled (stage I) SA; (C’) α-
SMA (VSMC); (C’’) HLA-G (EVT). (D) Col VI expression in remodeling (stage II) SA; (D’)
α-SMA (VSMC); (D’’) HLA-G (EVT). (E) Col VI expression in remodeling (stage III) SA;
(E’) α-SMA (VSMC); (E’’) HLA-G (EVT). (F) Col VI expression in remodeled (stage IV) SA; 580
(F’) α-SMA; (F’’) HLA-G. Scale bar = 50µm. Inset = negative control. L = lumen; BM =
basement membrane; a =. pericellular col VI staining; b = adventitia/outer VSMC layers; c =
inner VSMC layers; d = endovascular EVT. Serial sections from n=24 individual decidual
samples (6-12 weeks gestation) and a minimum of n=32 vessel sections in each remodeling
stage were assessed. Representative images are shown. 585
Figure 4. Serial sections showing immunolocalization of laminin in the decidua basalis
and spiral arterioles during remodeling in vivo. (A, B) Laminin in the decidual stroma. (C)
Control IgG. (D) Laminin expression in unremodeled (stage I) SA; (D’) α-SMA (VSMC); (D’’)
HLA-G (EVT). (E) Laminin expression in remodeling (stage II) SA; (E’) α-SMA; (E’’) HLA-590
G. (F) Laminin expression in remodeling (stage III) SA; (F’) α-SMA; (F’’) HLA-G. (G)
Laminin expression in remodeled (stage IV) SA; (G’) α-SMA; (G’’) HLA-G. Scale bar =
50µm. Inset = negative control. L = lumen; BM = basement membrane; a = pericellular laminin
staining; b = layers of laminin. Serial sections from n=24 individual decidual samples (6-12

30
weeks gestation) and a minimum of n=32 vessel sections in each remodeling stage were 595
assessed. Representative images are shown.
Figure 5. Serial sections showing immunolocalization of fibronectin in the decidua basalis
and spiral arterioles during remodeling in vivo.
(A) Fibronectin in the decidual stroma. (B) Control IgG. (C) Fibronectin expression in 600
unremodeled (stage I) SA; (C’) α-SMA (VSMC); (C’’) HLA-G (EVT). (D) Fibronectin
expression in remodeling (stage II) SA; (D’) α-SMA; (D’’) HLA-G. (E) Fibronectin expression
in remodeling (stage III) SA; (E’) α-SMA; (E’’) HLA-G. (F) Fibronectin expression in
remodeled (stage IV) SA; (F’) α-SMA; (F’’) HLA-G. Scale bar = 50µm. Inset = negative
control. L = lumen; a = pericellular fibronectin staining; b = immunopositive VSMC; c = 605
interstitial EVT; d = endovascular EVT; e = luminal EVT. Serial sections from n=24 individual
decidual samples (6-12 weeks gestation) and a minimum of n=32 vessel sections in each
remodeling stage were assessed. Representative images are shown.
Figure 6. Serial sections showing immunolocalization of fibrillin and elastin in the decidua 610
basalis and spiral arterioles during remodeling in vivo. (A) Fibrillin expression in
unremodeled (stage I) SA identified by (A’) α-SMA immunostaining. (B) Fibrillin expression
in remodeled (stage IV) SA identified by (B’) HLA-G immunostaining. (C) Control IgG. (D)
Fibrillin in the decidual stroma. (E) Elastin expression in unremodeled (stage I) SA identified
by (E’) α-SMA immunostaining. (F) Control IgG. (G) Elastin expression in remodeling (stage 615
III/IV) SA identified by (G’) α-SMA and (G’’) HLA-G immunostaining. Scale bar = 50µm. L =
lumen; a = VSMC; b = decidual stroma; c = pericellular fibrillin expression; d = internal elastic

31
lamina; e = immunopositive decidual cell; BM = basement membrane. Serial sections from
n=20 individual decidual samples (6-12 weeks gestation) and a minimum of n=24 vessel
sections in each remodeling stage were assessed. Representative images are shown. 620
Figure 7. Serial sections showing PAS staining in the decidua basalis and spiral arterioles
during remodeling in vivo. (A) PAS staining in unremodeled (stage I) SA; (A’) α-SMA
(VSMC); (A’’) HLA-G (EVT). (B) PAS staining in remodeling (stage II/III) SA; (B’) α-SMA;
(B’’) HLA-G. (C) PAS staining in in remodeled (stage IV) SA; (C’) α-SMA; (C’’) HLA-G. 625
Note endovascular EVT embedded within the fibrinoid (arrowheads). Scale bar = 50µm. L =
lumen; BM = basement membrane; a = pericellular PAS staining; b = reduced staining in areas
of VSMC loss; c = PAS-positive fibrinoid; d = EVT. Serial sections from n=20 individual
decidual samples (6-12 weeks gestation) and a minimum of n=24 vessel sections in each
remodeling stage were assessed. Representative images are shown. 630

32
Table 1. Details of antibodies used for immunohistochemistry
Antibody Source Working
concentration
Dilution
factor
Specificity
Human Leukocyte
Antigen–G (mouse)
Abcam, UK 2µg/ml 1:100 Extravillous
trophoblast
Anti-α-SMA (mouse) Dako, UK 0.18µg/ml 1:800 Vascular
smooth muscle
Anti-collagen type III
(mouse)
Abcam, UK 4.8µg/ml 1:500 Collagen type
III
Anti-collagen type IV
(mouse)
Sigma
Aldrich, UK
1µg/ml 1:200 Collagen type
IV
Anti-collagen type VI
(rabbit)
Sigma
Aldrich, UK
1µg/ml 1:500 Collagen type
VI
Anti-laminin (gamma
1; mouse)
Sigma
Aldrich, UK
4µg/ml 1:50 Laminin
Anti-fibronectin
(mouse)
Abcam, UK 1.8µg/ml 1:100 Fibronectin
Anti-elastin (rabbit) Sigma
Aldrich, UK
4µg/ml 1:250 Elastin
Anti-fibrillin (rabbit) Dr C Kielty,
University of
Manchester,
UK
4µg/ml 1:500 Fibrillin
Isotype control
mouse IgG
Sigma
Aldrich, UK
Matched to
each primary
antibody
- Negative
control
Non immune rabbit
IgG
Sigma
Aldrich, UK
Matched to
each primary
antibody
- Negative
control
Goat anti-mouse IgG Dako, UK 7µg/ml 1:200 Mouse IgG
Swine anti-rabbit IgG Dako, UK 4µg/ml 1:400 Goat IgG
635

33
Table 2. A semi-quantitative summary of changes in vascular extracellular matrix composition
observed during remodeling of decidual SA, as agreed by 2 independent assessors
Stage
Collagen
type III
Collagen
type IV
Collagen
type VI
Laminin Fibronectin Fibrillin Elastin Fibrinoid Intramural
trophoblast
Vascular
smooth
muscle
I +++ +++ + ++ - ++ ++ - - +++
II ++ + + - - ++ + - - ++
III ++ + + - ++ ++ + - ++ +
IV ++ + + - ++ ++ + ++ +++ -
640
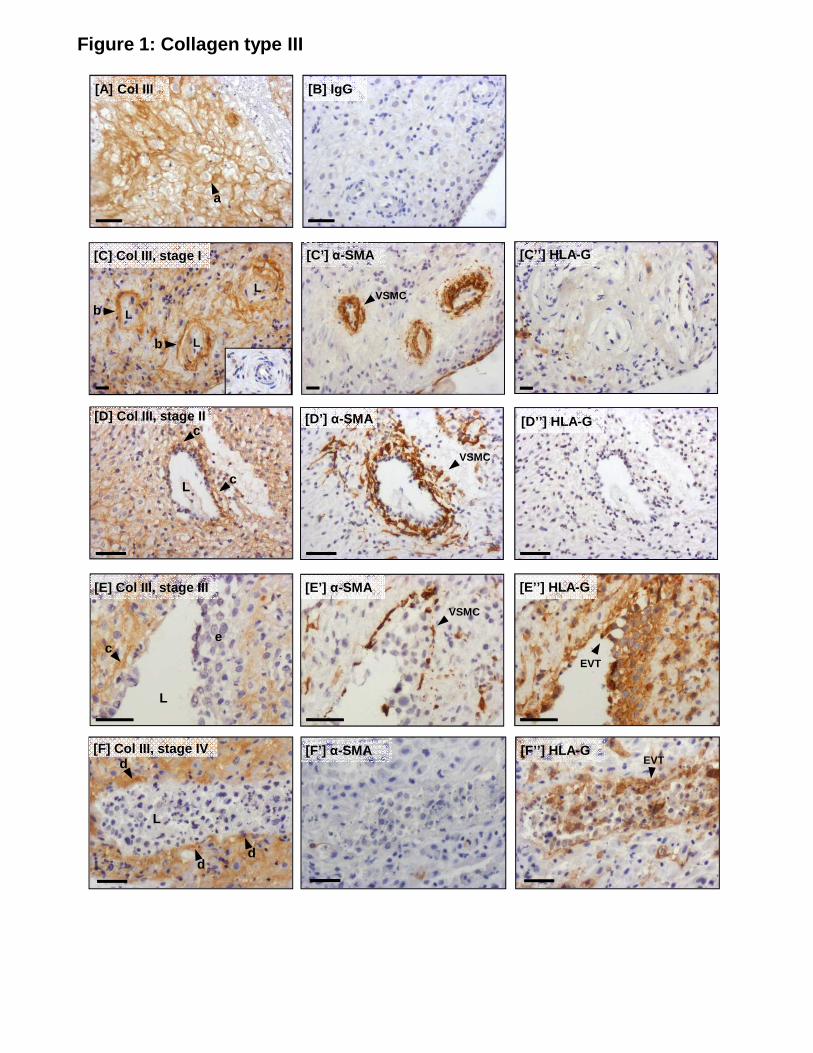
Figure 1: Collagen type III
[C’] α-SMA[C] Col III, stage I [C’’] HLA-G
[D’] α-SMA[D] Col III, stage II [D’’] HLA-G
[F’] α-SMA[F] Col III, stage IV [F’’] HLA-G
[B] IgG[A] Col III
VSMC
EVT
VSMC
[E’] α-SMA[E] Col III, stage III [E’’] HLA-G
VSMC
EVT
L
L
L
L
L
L
b
b
c
c
c
d
dd
a
e
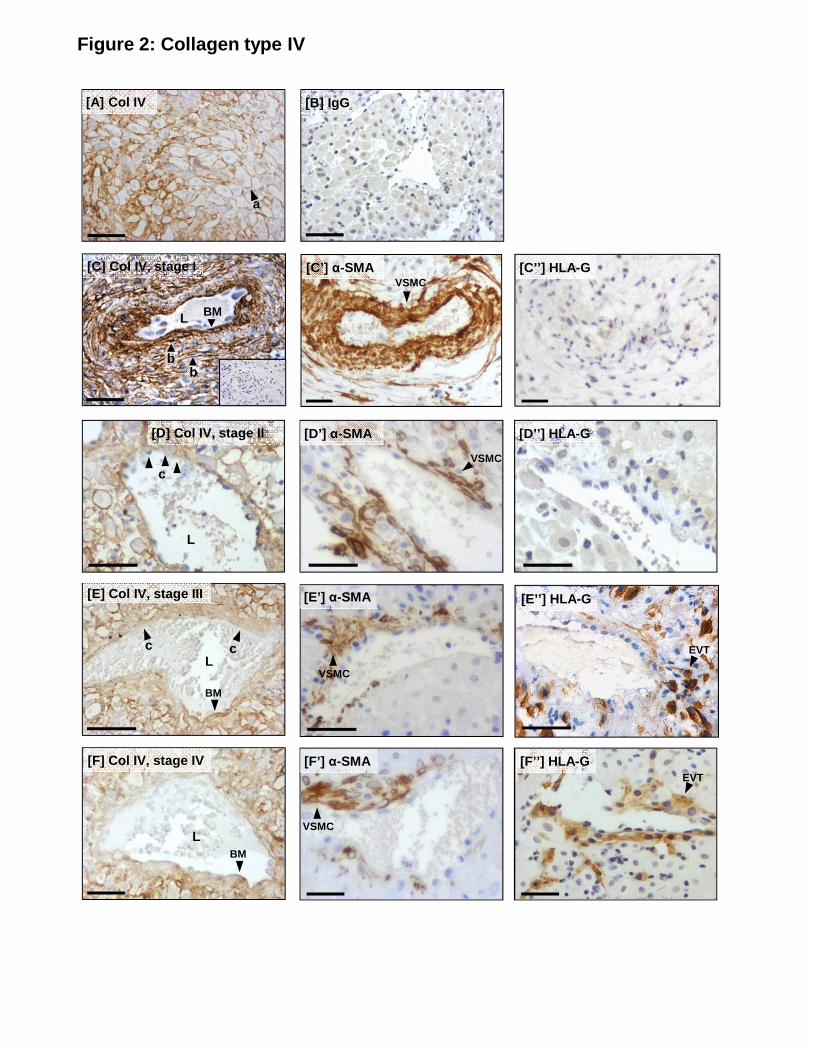
Figure 2: Collagen type IV
[B] IgG
α-SMA
[C’’] HLA-G[C’] α-SMA
[A] Col IV
[D’’] HLA-G[D’] α-SMA[D] Col IV, stage II
[F’’] HLA-G[F’] α-SMA[F] Col IV, stage IVEVT
VSMC
VSMC
VSMC[C] Col IV, stage I
BM
[E’’] HLA-G[E’] α-SMA[E] Col IV, stage III
VSMC
L
L
L
L
c
bb
c
BM
c
BM
a
EVT

Figure 3: Collagen type VI
[A] Col VI
[F’] α-SMA[F] Col VI, stage IV [F’’] HLA-G
[B] IgG
VSMC
EVT
EVT
L
[D’] α-SMA[D] Col VI, stage II [D’’] HLA-G
[C’] α-SMA[C] Col VI, stage I [C’’] HLA-G
VSMC
EVT
L
L
L
[E’] α-SMA[E] Col VI, stage III [E’’] HLA-G
EVT
VSMC
VSMC
a
b
bc
cb
b
d
BM
BM

Figure 4: Laminin
[D’] α-SMA[D] Laminin, stage I [D’’] HLA-G
[E’] α-SMA [E’’] HLA-G
[F] Laminin, stage III
[E] Laminin, stage II
[A] Laminin [C] IgG[B] Laminin
VSMC
VSMC
[G’] α-SMA [G’’] HLA-GEVT
[F’] α-SMA [F’’] HLA-GVSMC
EVT
[G] Laminin, stage IV
LL
L
L
L
BMb BM
b
VSMC
a
a
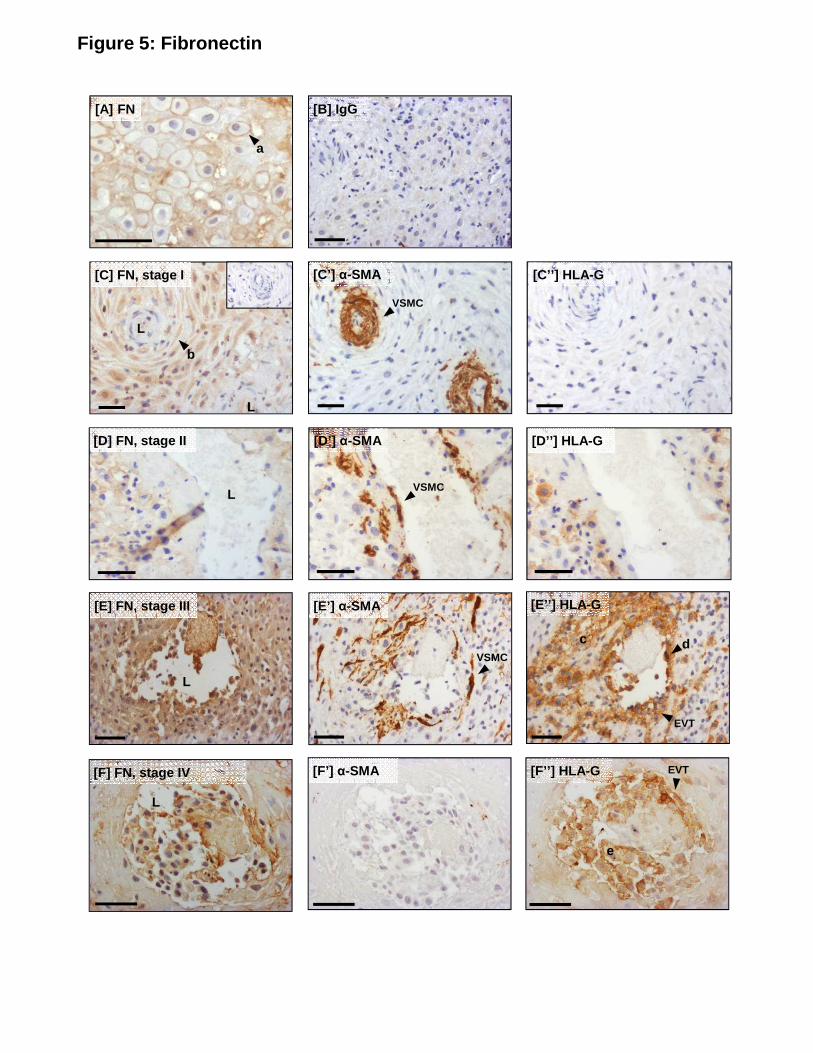
Figure 5: Fibronectin
[C’] α-SMA[C] FN, stage I [C’’] HLA-G
[E] FN, stage III
[F] FN, stage IV
[A] FN [B] IgG
VSMC
[E’] α-SMA [E’’] HLA-G
VSMC
EVT
[F’] α-SMA [F’’] HLA-G EVT
[D] FN, stage II [D’] α-SMA [D’’] HLA-G
VSMC
L
L
L
L
L
a
b
e
dc

Figure 6: Fibrillin/Elastin
Fibrillin
[A’] α-SMA[A] Fibrillin, stage I [C] IgG
[B’] HLA-G[B] Fibrillin, stage IV [D] Fibrillin
[E’] α-SMA[E] Elastin, stage I [F] l IgG
[G’] α-SMA[G] Elastin, stage III/IV [G’’] HLA-G
EVT
VSMC
VSMC
VSMC
EVT
L
L
L
L
L
b
a
c
c
b
e
d
dd
d
BM

Figure 7: Fibrinoid
[A’] α-SMA
VSMC
[B’] α-SMA
VSMC
[C’’] HLA-G
EVT
[A] PAS, stage I
L
[B] PAS, stage II/III
L
[C] PAS, stage IV
d
L
[B’’] HLA-G
[A’’] HLA-G
[C’] α-SMA
BM a
a
b
d
dd d
cVSMC
d