changes induced by natural scrapie in the calretinin-immunopositive cells and fibres of the sheep...
TRANSCRIPT

ORIGINAL PAPER
Changes Induced by Natural Scrapiein the Calretinin-Immunopositive Cells and Fibresof the Sheep Cerebellar Cortex
Adolfo Toledano & María-Isabel Álvarez &
Eva Monleón & Adolfo Toledano-Díaz &
Juan-José Badiola & Marta Monzón
Published online: 26 November 2011# Springer Science+Business Media, LLC 2011
Abstract Calretinin (CR)-immunopositive cells and fibresin the cerebellar cortex (vermal archicerebellum and neo-cerebellum) of scrapie-affected, ARQ/ARQ, Rasa Arago-nesa breed sheep were studied in comparison with healthy,young and aged, ARQ/ARQ, Rasa Aragonesa animals andwith Manchega breed sheep. The scrapie-affected sheepshowed signs of both cellular involution and hypertrophic/hyperimmunoreactive responses in all neuronal subtypes;the distribution of the neuronal subtypes in the archi- andneocerebellum, however, did not change compared withcontrols. The results suggest that the different CR expres-sion and/or CR content of cerebellar cortical neurons inscrapie-affected sheep are more related to their specificfunctions than any neuroprotective response. The reductionin the cell density of some CR-immunopositive neuronalsubsets (i.e. unipolar brush cells) is contradictory to thesupposed neuroprotective role of the calcium bindingprotein CR. However, the hyperimmunoreactivity of manyCR-immunopositive neuronal subsets (e.g. the Purkinjecells) suggests the involvement of an over-expression of
CR (transitory or restricted to selected neurons) as anadaptative mechanism to fight against the neurodegenera-tion caused by this prion disease. The changes in thenumber of immunopositive cells and the hypertrophic/hyperimmunoreactive response seen in scrapie-affected andaged sheep suggests that some different and some similarmechanisms are at work in this disease and aging.
Keywords Archicerebellum . Neocerebellum . Calretinin .
Scrapie . Sheep
Introduction
In the cerebellum of mammals, a large number of neuronsof different type/subtype show calretinin (CR) immunopo-sitivity. A high density of CR-immunopositive fibres is alsoseen. The intensity of this immunopositivity, as well as thenumber and location of these cerebellar cells and fibres,seems to depend on species [1–9] and on the cerebellarregion in question [4, 9]. In a previous report, the specialcerebellar pattern of CR-immunoreactivity in sheep wasdescribed in healthy adult animals belonging to twodifferent breeds [10]. A very high density of CR-immunopositive neurons and fibres showing intense immu-noreactivity was observed in the archicerebellum, as well asin a large number of CR-immunopositive neurons andfibres in the neocerebellum. In addition, important differ-ences in the main features of the CR-immunoreactivepattern were recognised between sheep and other mamma-lian species.
Like other members of the large family of elongationfactor-handed calcium-binding proteins (CaBPs) [11, 12],CR is currently understood to be a calcium buffer that
A. Toledano (*) :M.-I. ÁlvarezInstituto Cajal, CSIC,Avda. Dr. Arce 37,28002 Madrid, Spaine-mail: [email protected]
E. Monleón : J.-J. Badiola :M. MonzónCentro de Investigación en Encefalopatías y EnfermedadesTransmisibles Emergentes, Universidad de Zaragoza,Miguel Servet 177,50013 Zaragoza, Spain
A. Toledano-DíazDepartamento de Reproducción Animal, INIA,Avd. Puerta de Hierro s/n,Madrid, Spain
Cerebellum (2012) 11:593–604DOI 10.1007/s12311-011-0335-5

confers general protection against the pathogenic mecha-nisms of different neuron diseases. However, its exact rolein the neuronal processes of the cerebellum is unclear sincestriking differences can exist in the CR content ofmorphofunctionally similar neurons located in differentlobules (especially when they belong to different regionswith very different functions that appeared at different timesover the course of evolution, e.g. the archicerebellum andneocerebellum) and even neurons close to one another inthe same lobule. Moreover, wide variation is seen amongmammals in their CR-immunoreactivity patterns. Neitherthe physiological significance of these CR-enriched neu-rons nor their pathophysiological variations are wellunderstood [2, 13–16]. A reduction in CR can causecerebellar dysfunction resulting in problems of motorcoordination. For instance, in CR-null mice, the functionsof the granule and Purkinje cells are impaired [15, 17, 18],while induced CR expression in granule cells restoresnormal cerebellar functions [19].
High neuronal CR and other CaBP contents are thoughtto underlie the resistance of the cerebellum to the neuro-degeneration caused by Alzheimer’s Disease (AD) [20–22].In fact, the CR-immunopositive neurons in the brain arethought to be generally unaffected in AD [23–27], probablybecause the calcium buffering capacity of CR is involved inthe resistance of these cells to amyloid toxicity [28]. Asimilar situation is seen in experimental transmissiblespongiform encephalopathies (TSEs), in which corticalinhibitory CR-immunopositive neurons appear to be resis-tant to neurotoxic prion deposits [29]. However, thisneuroprotective effect of CR has been contested. Abnormalsubsets of CR-immunopositive cells, or the loss of suchcells, has been observed in AD [30, 31], and an age-relatedloss of CR-immunoreactive hippocampal interneurons hasbeen reported in transgenic models of AD [31, 32].Moreover, the re-evaluation of the roles of the differentCaBPs has suggested that these proteins are rapid modu-lators of intracellular calcium concentration (critical forsynaptic plasticity) rather than general systems for protec-tion against neuronal involution [13, 16, 33].
The present study was designed to investigate thechanges induced by scrapie in the CR-immunopositiveneurons and fibres of the cerebellar cortex and theirrelationship with the pathological involution that occurs inthis neurodegenerative prion disease of a very greatscientific interest given its importance as a model of humanTSEs. In this work, different functional regions of thecerebellum that appeared at different times during sheepevolution (lobule X of the archicerebellum and lobule VIIbof the neocerebellum) were examined because of theirnotable differences in CR immunoreactivity [4, 6, 9, 10].Changes in CR-immunoreactivity were compared withthose seen for calbindin (CB) and related to the glial
response and prion deposition that characterise scrapie. TheCB immunoreactive response has previously been used as acontrol in the examination of the responses of other CaBPsand to control for the folium-to-folium variation ofimmunoreactions [10]; CB immunoreactivity is very stable,therefore, any major change in this variable suggeststechnical errors have occurred. Scrapie-induced neuro-degeneration was also compared with physiological senileinvolution. The results help clarify the role of CR in theinvolutive process in prion diseases and confirm theexistence of neuronal circuits characterised by high CRcontents.
Experimental Procedures
Animal and Sample Processing
The experimental animals used in this study included 17Rasa Aragonesa sheep—six young, healthy, positive con-trol (i.e. scrapie-negative) animals, 4–5 years old (group A);five aged (8–9 years old), healthy positive control animals(group B) and six scrapie-affected animals aged 4–5 yearswith neurological symptoms (in the clinically advancedphase of the disease, typical form) and showing a similarpattern of intense PrPsc deposition in the cerebellum (groupC). All these sheep had an ARQ/ARQ genotype andbelonged to a regularly monitored flock with a highincidence of natural scrapie (positive flock) kept forresearch purposes at the Research Centre for Encephalo-pathies and Transmissible Emerging Diseases (Universidadde Zaragoza, Zaragoza, Spain). Ten further healthy animalsbelonging to the same breed and with the same genotypewere selected from different flocks in which no scrapie hadever been reported (negative flocks); these animals wereused as negative controls (group D). In addition, ten healthySpanish Manchega sheep of different ages (five young [4–5 years old], healthy control animals [group E] and fiveaged [9 years old] but healthy animals [Group F]) wereexamined to test for breed-dependent variations. These lastanimals belonged to the Department of Animal Reproduc-tion at the INIA, Madrid, Spain. All animals werediagnosed by histopathological and immunohistochemicaltechniques proposed by the World Organization for AnimalHealth (OIE Manual of Standards for Diagnostic Tests andVaccines. Office International des Epizooties. Paris, 2008).Rules established by the Spanish National ResearchCouncil’s guide for animal experimentation, based onSpanish and European guidelines, were followed withrespect to animal handling and care. Necropsies and brainprocessing were performed according to the protocols forthe analysis of TSEs, for which fresh (for biochemical,molecular and genetic analysis) and immersion-fixed (for
594 Cerebellum (2012) 11:593–604

morphological and histochemical analysis) samples werecollected. Following an i.v. injection of pentobarbital, allanimals were sacrificed by exsanguination. The brain wasimmediately removed and one hemisphere placed in 4%formaldehyde in phosphate buffered saline (PBS). Tissuesfrom the lymphoreticular system were collected and thepresence/absence of scrapie prion (PrPsc) determined inorder to detect possible pre-clinical cases. Brain sampleswere trimmed, post-fixed and embedded in paraffin waxaccording to standard procedures. Serial transverse sections(5 μm) of the cerebellum were mounted on treated glassslides (Vectabond; Vector, Burlingame, CA, USA). Vermallobules X (archicerebellum) and VIIb (neocerebellum) (asdescribed in [34]) were studied using selected groups of 50serial sections at the appropriate level. Ten sections werestained with haematoxylin and eosin (H/E) followingstandard procedures. Sections were de-waxed and hydratedusing routine methods before immunostaining.
Calretinin Immunohistochemistry
Endogenous peroxidase activity was inhibited by incu-bation of the sections for 15 min in 3% H2O2 inmethanol. Non-specific antigens were blocked by incu-bating for 20 min in diluted Normal SerumTM (Vector,Burlingame, CA, USA). Slides were pre-treated withproteinase K (Dako, Denmark; 2 μg/ml) for 10 min at37°C. Monoclonal mouse anti-human CR antibodies(clone DAK Calret 1; Dako, Denmark) were used at adilution of 1:100 to detect CR in brain sections. Allsections were incubated at room temperature for 1 h. TheEnVision™ system (Dako, Denmark) was used to visual-ise immunoreactions; 0.02% of 3,3′-diaminobenzidine(DAB, Sigma Chem, St. Louis MO, USA) was used as achromogen. The sections were then stained with H/E,dehydrated through a graded alcohol series, cleared inxylene and mounted with Entellan (Merck, KgaA, Darmstadt,Germany). These tissue sections were analysed using an E400Nikon Eclipse microscope (Nikon, Japan). Control sectionswere incubated without anti-CR antibody, or after the anti-body’s pre-incubation with CR; no neurons or fibres becamestained. The DAB nickel variant [35] was used to enhance thecontrast at low magnification in two sections from eachgroup (groups A–F).
Glial Fibrillary Acidic Protein and CBImmunohistochemistry
After de-waxing and rehydration with PBS, the inhibitionof endogenous peroxidase, the blocking of non-specificantigens and the retrieval of antigens by a short (10 min)incubation with proteinase K, sections parallel to those usedin CR detection, were treated to show glial fibrillary acidic
protein (GFAP) immunoreactive glial cells and CB-immunoreactive Purkinje cells. The antibodies employedwere rabbit polyclonal antibodies raised against GFAP(1:500) (Chemicon, Temecula, CA, USA) and calbindin D-28 k (1:1,000) (Chemicon, Temecula, CA, USA). TheEnVision™ system and DAB were used to visualise thereactions. The sections were then stained with H/E,dehydrated through a graded alcohol series, cleared inxylene and mounted with Entellan. Control sectionsincubated without GFAP or anti-CB antibodies showed nostained glial cells, neurons or fibres.
Prion (PrPsc) Immunohistochemistry
PrPsc was detected immunohistochemically using theprotocol described by Monleón et al. (2004) [36]. Briefly,the following pre-treatments were used for antigen retrieval:immersion in 98% formic acid for 15 min, proteinase Ktreatment for 15 min at 37°C (Roche, Switzerland; 4 μg/ml)and hydrated autoclaving. A step with blocking reagent(Dako, Denmark) for 5 min was included before incubationwith the primary antibody (L42, R-Biopharm, Germany;1:500) for 30 min at room temperature. The EnVision™system and DAB were used to visualise the reactions.Control sections incubated without PrP-antibody showedno stained immunodeposits.
Statistics
The density and percentage of immunopositive cells of eachneuronal subtype were calculated as previously described[10]. Five sections with a minimum separation of 50 μmwere randomly selected at the level of lobes X (archicer-ebellum) and VIIb (neocerebellum) in each animal to countthe number of immunopositive cells. In each section, 12areas of 370×240 μm including the Purkinje and granulecell layers (the long axis being parallel to the Purkinje celllayer) were randomly selected and the number of CR-immunopositive Purkinje, Golgi, Lugaro, brush (or unipolarbrush) and granule cell bodies recorded. In another 12randomly selected areas of 370×240 μm including thePurkinje cell and molecular layers, the number of CR-immunopositive basket and stellate cells were counted.When necessary, a parallel section was used to classify theimmunopositive cells. ACT and NIS-D (Nikon, Japan) andIPPlus (Image Pro Plus, Media Cybernetics, Iowa) softwarewas used for morphometric analysis and enumeration. Thepercentage of immunopositive neurons of each cortico-cerebellar subtype was obtained in relation to the totalnumber of each subtype in five H/E-stained sections using apreviously described protocol [10]. The “pale cells” ofAltman and Bayer (1977) [37] observed in H/E-stainedsections were deemed to be brush cells.
Cerebellum (2012) 11:593–604 595

The involution of the cerebellar cortex in scrapie-affected animals was analysed by examining the reductionin the density of Purkinje cells (identified by theirnucleolus) and the reduction of the granule cell layer usingIPPlus (Image Pro Plus, Media Cybernetics, Iowa) softwarein morphometric analysis. The reduction in Purkinje celldensity was calculated by comparing the number ofnucleoli per unit length of the Purkinje cell layer in coronaltissue sections of lobules X and VIIb [38]. Five randomlyselected H/E-stained sections with a minimum separation of25 μm were examined in animals from groups A–F. Thedensity reduction of the granule cell layer was determinedby comparing the cross-sectional area of this layer per unitlength of the Purkinje cell layer in coronal tissue sections.
ANOVA was used to examine the homogeneity of theresults for animals of groups A–F. The Student’s t test wasused to compare means.
Results
CR-Immunopositive Neurons and Fibres in Young Control(Groups A, D and E) and Old Animals (Groups B and F)
All types of neuron and fibre in the cerebellar cortex ofhealthy sheep (groups A, B, D and E) showed very intenseCR-immunopositivity. The qualitative and quantitativeresults obtained for the young control animals (groups A,D, E) were very similar to those reported in our previousstudy performed on intravitally perfused as well asimmersion-fixed (following TSE protocols) cerebella [10].ANOVA showed no significant differences between thepositive and negative control Rasa Aragonesa flocks(groups A and D), nor the Manchega and Rasa Aragonesabreeds (groups A and E).
The healthy Rasa Aragonesa and Manchega sheep aged8–9 years (groups B and F) showed a general pattern of celland fibre CR-immunopositivity similar to that seen in theyoung animals (groups A and E), with lobule-dependentvariations in immunopositive cell density. In the aged,positive control Rasa Aragonesa sheep (group B), the totalnumber of Purkinje cells decreased by 4–9% and thegranule cell layer section area decreased by 6–15%. Thenumber of unipolar brush cells was significantly reduced inall lobules (Fig. 1). Immunoreactivity remained high, but asubset of cells showed lower immunoreactivity. CR-immunopositive Golgi cells increased in average density,but not significantly so. CR-immunopositive Purkinje cellsincreased in average density in lobule VIIb and X. Threesubsets of these immunopositive cells were observed(Fig. 1) with different immunoreactivities: (a) immuno-positive Purkinje cell bodies with homogenously dispersed(although few), fine granular deposits (+ in Fig. 1), (b)
immunopositive somata with homogeneously dispersed,fine granular deposits of medium intensity (the mostcommon Purkinje cell types in young animals) (++) and(c) heterogeneously dispersed (granular like), highly reac-tive or hyperimmunoreactive Purkinje cell bodies (notobserved in young animals [groups A, D and E]) (+++).A small but significant increase in the number of Purkinjecells of the above b subset was seen (p=0.034 in lobule Xand p=0.014 in lobule VIIb). In lobule X, the number ofimmunopositive Lugaro cells was not significantly differentto that seen in the young healthy animals (group A) (p=0.568), although a significant reduction (p<0.001) was seenin lobule VIIb (Fig. 1). The perikarya of granule, basketand stellate cells were only immunoreactive in certainregions of the folia, but with no important differencescompared with young animals (group A) observed. All theimmunopositive fibres (climbing and mossy fibres in thegranule cell layer, and climbing and parallel fibres andbasket and stellate axons in the molecular layer) seen inyoung animals (groups A, D and E) were present in agedanimals (groups B and F), maintaining the variation fromfolium to folium characteristic of the young animals (groupA, D and E). Some of these elements were hypertrophied.The results for the aged (9 years old) Manchega and controlnegative (8–9 years old) Rasa Aragonesa sheep (groups Fand B) were very similar.
CR-Immunopositive Neurons and Fibresin Scrapie-Affected Animals
A. General PatternThe Rasa Aragonesa sheep in a clinically advanced
phase of scrapie (group C) showed a general pattern ofcell and fibre immunoreactivity similar to that seen inhealthy animals (group A) but with more involutedfolia and a large reduction in immunopositive celldensity. The total number of Purkinje cells decreasedby 13.3±5.1% and the granule cell layer section areaby 14.7±7.1%. However, in selected regions, adramatic reduction in the Purkinje cell density(>25%) and/or section area of the granule cell layer(>25%) was observed. Very variable patterns of CR-immunopositive structures were recorded in differentcortical areas. These ranged from an intense reductionin certain types of immunopositive neurons or fibres tostrong CR-immunoreactivity in normal, hypertrophicor atrophic cells of all types (Fig. 2a–k). Only in a fewinstances did regions of lower reactivity correspond tomore involuted folia. Special cerebellar CR-richcircuits already described in control animals [10] wereobserved, the most prominent formed by either bandsor groups of CR-immunopositive mossy fibres and themoss-like endings of climbing fibres in synaptic
596 Cerebellum (2012) 11:593–604

Fig. 1 Rasa Aragonesa positive flock (groups A, B and C). Number of CRunipolar brush, Golgi, Purkinje and Lugaro cells in healthy control (youngand aged animals; groups A and B) and scrapie-affected sheep (group C) inlobules X and VIIb of the cerebellar vermis. Areas of 370×240 μm (88.8×103 μm2) including the Purkinje and granule cell layers were randomlyselected for counting. The percentage of CR-immunonegative (−) and CR-immunoreactive Purkinje cells (+/+++) are represented in the graphs, aswell as the different subsets of immunopositive Purkinje cells: aimmunopositive Purkinje cell bodies of homogeneously dispersed (al-
though few), fine granular deposits (+ in Fig. 1), b immunopositive somatawith homogeneously diffuse, fine granular, deposits of medium intensity(the most common Purkinje cells in young animals) (++), and cheterogeneously distributed (granular-like), highly reactive or hyperimmu-noreactive Purkinje cell bodies (not observed in young animals) (+++).Data are expressed as mean±SEM. **p<0.005; *p<0.05; n (group A;young healthy animals—young a.)=6; n (group B; old healthy animals—aged a.)=5; n (group C; scrapie-affected animals—scrapie a.)=6. Totalcounts/group=5 sections×12 areas×number of animals/group
Cerebellum (2012) 11:593–604 597

contact with the unipolar brush cells (generally inclusters) or close CR-immunopositive Purkinje cells.In scrapie-affected sheep (group C), these CR-richcircuits were mainly present in areas of high CR-immunoreactivity (Fig. 2a–e), but the immunoreactivechanges induced hindered their observation. Vacuolarstructures characteristic of the spongiform encephalo-pathies were observed (Figs. 2e, f, h, k and 3). No clearboundaries were seen between the granule cell layerand the white matter in many areas, or between thePurkinje cell layer and the molecular layer. Differentelements entered the white matter (unipolar brush,Golgi and granule cells; Fig. 2e) and molecular layer(unipolar brush, Golgi and Purkinje cells; Fig. 2g). Ingeneral, all these “ectopic” cells showed intensereactivity. Hyperplasia of glial cells was observed inthe Purkinje and granule cell layers (Fig. 2c, d, f, j).
B. Unipolar Brush CellsThe number of unipolar brush cells was significant-
ly lower in all the lobules of group C animalscompared with the healthy animals of group A (Figs. 1and 2a–c). Almost all these cells were intense CR-immunoreactive, including a significant number (12–29% in folia of the lobule X, 6–17% in lobule VIIb) ofdystrophic-vacuolated unipolar brush cells; only onesubset (<12% in all lobules) showed lower intensity.Unipolar brush cell-like immunonegative cells werenot seen. Moreover, the density of “pale cells” in H/E-stained sections was close to that recorded for unipolarbrush cells. Some unipolar brush cells (14–24% inlobule X, 12–18% in lobule VIIb) were hypertrophicand/or hyperimmunoreactive and showed a morecomplex cell architecture (Fig. 2d, f, i). Especially incertain regions of lobule X, the apical dendritic endingsof these cells showed hypertrophy, a complex shapeand very intense immunoreactivity. A part (16–27% inall the lobules) of the terminal dendrites of thedystrophic unipolar brush cells were in close relation-ship with, or included in (Fig. 2k) vacuolar structures.Axons were, in general, less immunoreactive.
C. Golgi, Lugaro and Granule CellsImmunopositive Golgi cells were fewer in number
in the archicerebellum scrapie-affected sheep (group C)than in the controls (group A), but not significantly so(Figs. 1 and 2h). Immunopositive Lugaro cells werefound at a lower density only in the neocerebellum ofthese affected animals (Fig. 1). The processes of someof these cells showed low immunoreactivity. Immuno-positive granule cell perikarya were few in all thelobules examined (Figs. 2a–c), although an increasednumber of immunopositive granule cells was seen insome areas where hypertrophic/hyperactive Purkinjecells and unipolar brush cells were in the majority
(Fig. 2a–c). In general, all the axons showed lessimmunoreactivity than in the controls, although insome areas of both lobules intense CR-immunopositiveaxons (mainly parallel fibres) were seen.
D. Purkinje CellsThe most variable immunopositive cells were the
Purkinje cells, both in terms of their density andinmunodeposit intensity (Fig. 2d, f and j). Severalmorphological variations were seen in scrapie-affectedanimals (group C). Hypertrophic (Fig. 2j), atrophic(Fig. 2f) and dystrophic Purkinje cells (Fig. 2g) wereseen in high proportion. In the archicerebellum (lobuleX), the average density of the immunopositive Pur-kinje cells was greater than that observed in control
Fig. 2 Calretinin-immunopositive (CR) cells and fibres in scrapie-affected sheep (group C). a Cerebellar lobule VIIb; b–k, lobule X. aArea of intense immunoreactivity with variable immunopositivity inthe Purkinje cells (Pc) and in the granule cells (gc). Some basket cells(arrow) and climbing fibres (black right-pointing triangle) wereimmunopositive in the molecular layer as well as many mossy fibresin the granule cell layer. b Area of intense immunoreactivity withvariable immunopositivity in the Purkinje cells including one“ectopic” Purkinje cell (Pc-e) in the molecular layer and one atrophicPurkinje cell (Pc-a). Some Bergmann-like glial processes are observedin the molecular layer (arrow) together with climbing and parallelfibres. c Higher magnification of the granule cell layer of b. In thegranule cell layer of these areas of intense CR-immunoreactivity,many cellular elements with intense CR-immunoreactivity are seen. Ahigh density of CR-immunopositive mossy fibres, granule cells (gc)and astrocytes (a) exist. Thin, thick and sheet glial-like processes areobserved surrounding CR-immunopositive and CR-immunonegativegranule cells and vacuolar structures (asterisk). d The unipolar brushcells were in lower number than in healthy animals (group A), but theremaining cells maintained a very high immunopositivity (arow).Golgi cells presented very variable immunoreactivity; in the image,one immunopositive Golgi cell with hyperactive dendrites (G+) andtwo CR-immunonegative Golgi cells (G−). e Unipolar brush cells(arrow) in the deepest region of the granule cell layer (gL) and in thewhite matter (wm) were observed. In the white matter, the CR-immunopositive fibres do not form fascicles of fibres running inparallel (similar to that seen in the healthy animals [group A]). f Inareas of not intense CR-immunoreactivity, the immunopositiveelements were the same above-mentioned cells and processes. In theimage, atrophic (Pc-a) and dystrophic (intra/peri-vacuolar) (Pc-v)Purkinje cells, brush (arrow) and granule cells, and glial cell bodiesand processes (black right-pointing triangle), these mainly surround-ing vacuolar structures (asterisk), can be observed. g A dystrophicPurkinje cell, “ectopic” in the molecular layer, with intense CR-immunoreactivity in the body and newly formed dendrites. The thickand thin dendrites of other Purkinje cells show weak immunoreactivity(black right-pointing triangle). h A CR-immunopositive small Golgicell near a vacuolar structure (asterisk) without CR-immunopositivestructures surrounding it. i A unipolar brush cell, of subtype Pdescribed in [10], with a hypertrophied dendrite (arrow). j Hypertro-phic, hyper-reactive Purkinje cells. Only two of the surrounding glialcells show CR-immunoreactivity (black right-pointing triangle). k Avacuolar structure (asterisk) showing part of the terminal dendrite of aunipolar brush cell (arrow). This dystrophic structure maintains highCR-immunoreactivity. Another normal unipolar brush cell, subtype Odescribed in [10], can be observed at the bottom (black right-pointingtriangle). Scale bars—50 (a, b), 13.5 (c, j, k), 20 (d, f), 65 (e), and57.5 μm (g, h, i)
�
598 Cerebellum (2012) 11:593–604

animals (group A), both when considering the positivecells together or as three Purkinje cell subsets of low,medium and high immunoreactivity (the subsets +, ++,+++ in Fig. 1). However, in the neocerebellum (lobuleVIIb), the average density of immunopositive Purkinjecells was reduced compared with the control group(Fig. 1). Wide variation was seen in immunodepositintensity. In these scrapie-affected animals (group C),selected Purkinje cells showed very intense immuno-reactivity (subset +++ in Fig. 1; images in Fig. 2a, b)
not observed in the controls but only a large number ofPurkinje cells showing medium or low immunoreac-tivity (++ and + in Fig. 1) was observed in lobule X.Larger areas with groups of up to 25 Purkinje cells ofvariable immunopositivity were observed. No relation-ships could be established between Purkinje cellimmunoreactivity and cell shape (hypertrophic, dys-trophic or atrophic), the presence of internal micro-vacuolisation, the level of surrounding vacuolisation orthe number of surrounding glial cells. Ectopic Purkinje
Cerebellum (2012) 11:593–604 599
cells located into the molecular layer showed highreactivity (Fig. 2g). Axon and dendritic processes ofsome Purkinje cells were intensely immunostained.
E. Basket and Stellate CellsThe density of CR-immunopositive basket and
stellate cells in scrapie-affected animals (group C)was similar to that observed in healthy animals (groupA). The dendritic processes were more intenselyimmunostained in these immunopositive neurons thanin those from the healthy controls (Fig. 2d, h and j).
F. Afferent FibresImmunopositive climbing and mossy fibres were
observed in the granule cell layer in scrapie-affectedanimals (group C) with many elements hypertrophiedand/or very immunostained (Fig. 2c). The white mattershowed the fascicles of CR-immunopositive fibres(including the efferent ones) running in parallel, butthey were distorted and heterogeneous in shape andCR-immunoreactivity (Fig. 2e). In the molecular layer,areas with intense immunopositive climbing fibres(Fig. 2a–c) together with immunopositive hypertrophicor thin Purkinje cell dendrites and parallel fibres(Fig. 2j) were seen.
G. Glial CellsCR-immunoreactivity was observed in selected
glial-like cells in scrapie-affected animals (group C),mainly in cells near the Purkinje cells (probablyprotoplasmic astroglial and Golgi epithelial cells)(Figs. 2j and 3). Some CR-immunopositiveBergmann-like processes were seen in the molecularlayer (Fig. 2c). In the granule cell layer, protoplasmicastrocyte-like cells, as well as lightly or intenselystained thin or thick processes and immunoreactivesheets (probably glial processes) between the granulecells, near the Purkinje cells or surrounding vacuolarstructures, were observed (Fig. 2a–c). Similar immu-nostained structures were not observed in youngcontrol animals (groups A, D and E). Processes of
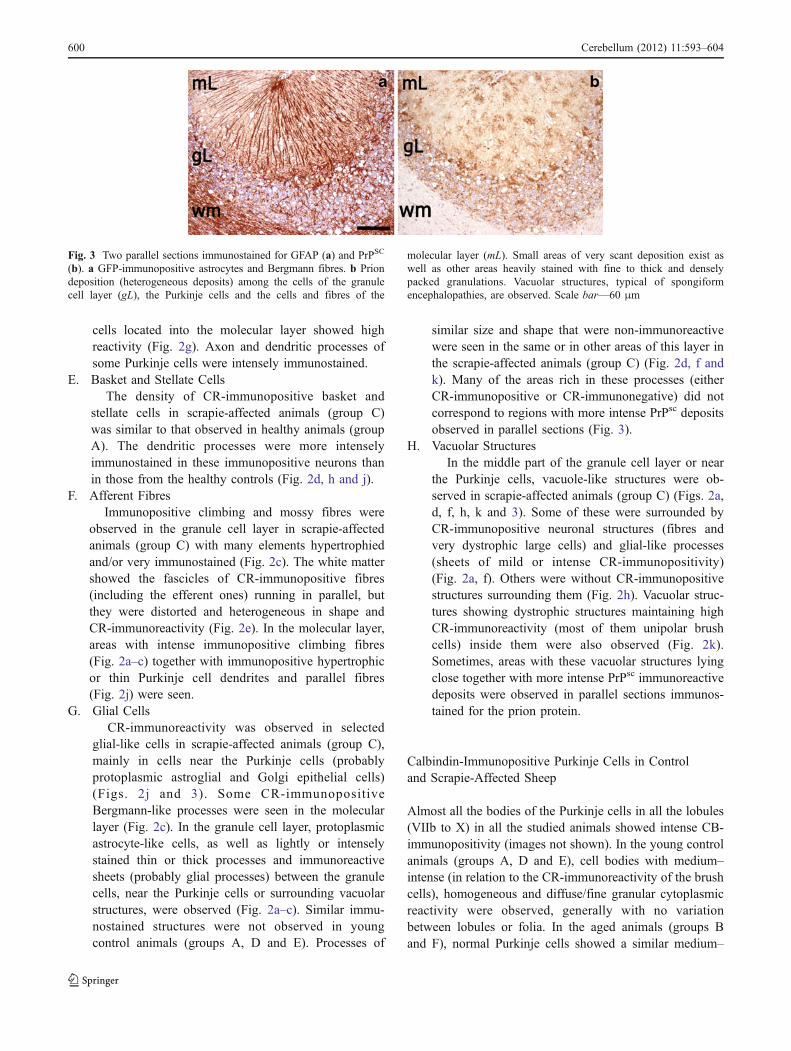
similar size and shape that were non-immunoreactivewere seen in the same or in other areas of this layer inthe scrapie-affected animals (group C) (Fig. 2d, f andk). Many of the areas rich in these processes (eitherCR-immunopositive or CR-immunonegative) did notcorrespond to regions with more intense PrPsc depositsobserved in parallel sections (Fig. 3).
H. Vacuolar StructuresIn the middle part of the granule cell layer or near
the Purkinje cells, vacuole-like structures were ob-served in scrapie-affected animals (group C) (Figs. 2a,d, f, h, k and 3). Some of these were surrounded byCR-immunopositive neuronal structures (fibres andvery dystrophic large cells) and glial-like processes(sheets of mild or intense CR-immunopositivity)(Fig. 2a, f). Others were without CR-immunopositivestructures surrounding them (Fig. 2h). Vacuolar struc-tures showing dystrophic structures maintaining highCR-immunoreactivity (most of them unipolar brushcells) inside them were also observed (Fig. 2k).Sometimes, areas with these vacuolar structures lyingclose together with more intense PrPsc immunoreactivedeposits were observed in parallel sections immunos-tained for the prion protein.
Calbindin-Immunopositive Purkinje Cells in Controland Scrapie-Affected Sheep
Almost all the bodies of the Purkinje cells in all the lobules(VIIb to X) in all the studied animals showed intense CB-immunopositivity (images not shown). In the young controlanimals (groups A, D and E), cell bodies with medium–intense (in relation to the CR-immunoreactivity of the brushcells), homogeneous and diffuse/fine granular cytoplasmicreactivity were observed, generally with no variationbetween lobules or folia. In the aged animals (groups Band F), normal Purkinje cells showed a similar medium–
Fig. 3 Two parallel sections immunostained for GFAP (a) and PrPSC
(b). a GFP-immunopositive astrocytes and Bergmann fibres. b Priondeposition (heterogeneous deposits) among the cells of the granulecell layer (gL), the Purkinje cells and the cells and fibres of the
molecular layer (mL). Small areas of very scant deposition exist aswell as other areas heavily stained with fine to thick and denselypacked granulations. Vacuolar structures, typical of spongiformencephalopathies, are observed. Scale bar—60 μm
600 Cerebellum (2012) 11:593–604

intense immunoreactivity, but hypertrophic somata, den-drites and axons sometimes showed more intense immuno-reactivity. Immunonegative cells were infrequent. In thescrapie-affected animals (group C), a wide variety of CB-immunoreactive Purkinje cell bodies were seen, with moregranular deposits of variable intensity in many cells,including atrophic, dystrophic and hypertrophic elements.Some Purkinje cells with vacuolar structures showedintense immunoreactivity. Negative Purkinje cells were lessthan 5%. In lobule VIIb, but not in lobule X, areas ofgenerally lower immunoreactivity were observed, but stillincluded hypertrophic/hyper-reactive elements.
GFAP-Immunopositive Glial Cells in Controland Scrapie-Affected Sheep
Protoplasmic and fibrous astrocytes were immunostained inscrapie-affected animals (group C) by the two polyclonalantibodies used, as were the Bergmann fibres in themolecular layer (Fig. 3). Only the size and shape of somesubtypes of GFAP-immunostained astrocytes and glialprocesses were studied for comparison with the non-neuronal cellular structures showing CR-immunopositivity.A homogeneous network of GFAP-immunopositive astro-cytes was observed in the granular layer of young healthyanimals (groups A, D and E). The number and intensity ofthe astrocytes was greater in the old controls (groups B andF), but far superior in the scrapie-affected animals (groupC). In this last group, many processes were hypertrophic/hyperimmunoreactive, but no areas of greater accumulationof hypertrophic cells existed. Bergmann fibres wereimmunostained in a very variable manner in all the foliaof all the lobules in all sheep groups. In the aged (groups Band F) and scrapie-affected animals (group C), moreprocesses were stained, their thickness being greater insome folia (Fig. 3). The morphological similarity betweensome GFAP-immunostained astrocytes and the Bergmannfibres, and between CR-immunostained glial-like cells andglial-like processes, was noted (Fig. 2c, f, j). In some areasof the Purkinje cell layer of the scrapie-affected animals(group C; many of which showed intense reductions in thedensity of Purkinje cells), a great number of GFAP-immunonegative glial bodies were observed.
Immunopositive PrPsc Deposits in Scrapie-AffectedAnimals
The scrapie-affected sheep (group C) showed very intense PrPimmunoreactivity in lobules VIIb and X. The pattern ofdeposition was similar in all these animals. The PrPsc
immunoreaction was observed in all cerebellar layers,revealing a non-homogeneous but constant presence of priondeposition. Small areas of variable size and shape but of very
low immunoreactivity (very fine, dispersed granules) wereseen close together with other small areas or “deposits” withgranules in far greater numbers (confluent in many instances)(Fig. 3). The most intense immunoreaction was seen betweenthe granule cells, in the deeper region of the molecular layerand near the Purkinje cells (Fig. 3). The parallel sectionsprocessed for CR or PrPsc immunodetection showed themost heavily stained prion deposits to be usually close toareas of low cell density between the granule cells, but notclose to groups of cells showing CR-immunoreactivity.Purkinje cells and unipolar brush cells of pathologicalappearance were observed close to the small areas both ofvery low and intense prion deposition. The parallel study ofPrPsc - and GFAP-immunostained sections revealed nocorrespondence between heavy prion protein deposits andhyperimmunoreactive glial cells. Only in areas of greaterdensity of GFAP-immunonegative glial bodies in thePurkinje cell layer was such a correlation appreciated.
Discussion
In the scrapie-affected animals (group C), intense variationswere observed in the size, shape and CR-immunopositivityof many elements of the different subsets of neurons andfibres. The density of the different CR-immunopositiveneuronal subsets decreased, except for the Purkinje cellsubset in the archicerebellum, but many elements of these(unipolar brush cells, Purkinje, Golgi and Lugaro cells)were hypertrophic. These cells showed greater complexityin their shape, generally accompanied by more intense CR-immunopositivity. These hypertrophic/hyperimmunoreac-tive neurons were observed in cerebellar areas of highCR-immunoreactivity (lobule X) as well as in areas withsmall numbers of CR-immunopositive elements (morecommon in lobule VIIb). The hyperimmunoreactivityshown by these neurons sometimes involved the finestdendrites and the axons (Purkinje cells), but was not relatedto their morphology (hypertrophy, atrophy, dystrophy),intra- or extracellular spongiform pathology, or surroundingastrogliosis (unipolar brush cells, Purkinje, Golgi andLugaro cells). The number of immunoreactive granule,basket and stellate cell perikarya remained low, but moreregions of the molecular layer showed stained axons arisingfrom these cells. Similarly, extracorticocerebellar afferentmossy and climbing fibres showed increased immunoreac-tivity in some areas. Together, these results confirm that,besides a reduction in the neuronal density induced byscrapie, some cells of the CR-immunopositive subsets ofneurons in the cerebellar cortex respond to this pathologyby becoming hypertrophic and/or hyperimmunoreactive.Moreover, the special cerebellar CR-rich circuits describedin healthy sheep are maintained.
Cerebellum (2012) 11:593–604 601

It is noteworthy that the hypertrophy/hyperimmunoreac-tivity shown by some neurons, as well as the loss of othercells from each subset, was independent of the intensity ofPrPsc immunodeposits in the close vicinity (as observed inparallel sections because the identification of cell types inPrPsc-immunostained serial slides was very difficult).However, many medium–large, complex, vacuole-likestructures in the granule cell layer (typical of scrapie) wereseen close to cells or cellular structures showing very highCR-immunopositivity. Some of these were the large cells ofthis layer (both hypertrophic and dystrophic) or differenttypes of axon, dendrite or glial process. In scrapie-affectedanimals (group C), the intense and sustained accumulationof pathological prions in the form of diffuse depositions ordense deposits in the molecular and granule cell layers,together with homogeneous astrogliosis, suggest that,ultimately, atrophy/cell loss as well as hypertrophy/hyper-immunoreactivity may be PrPsc-induced. However, otherfactors might be operational. For example, the “ectopic”cells (Purkinje, unipolar brush cell and Golgi cells) enteringthe molecular layer via morphological changes induced bythe disease (astrogliosis, cell loss, neuronal dystrophy) arealso CR-hyperimmunoreactive, a fate probably induced bytheir new environment. Analysis of the exact relationshipbetween PrPsc deposition, or the vacuolisation associatedwith scrapie, and the changes in CR expression in differentcell types, will require studies on the colocalisation ofbiomarkers and ultrastructural modifications.
A significant reduction in the number of unipolar brushcells (a mean of approximately 50%) was seen in thescrapie-affected animals (group C). However, this was notexactly the case for the Purkinje cells, in which a significantincrease in CR-immunoreactivity in the archicerebellumand a decrease in immunoreactivity in the neocerebellumwas observed, leading to an overall 13.3% of reduction inthe total cell count. In the neocerebellum, Lugaro cellnumbers and immunoreactivity declined. Golgi and granulecells were unchanged in all lobules. These data suggest thatresistance to involution varies, the neurons with the highestCR-content being the most sensitive to this prion-inducedprocess. CR-immunoreactive neurons have been regardedas showing special resistance to the cellular pathology ofAD [23–26, 28], dementia with Lewy bodies [39] and priondiseases [29]. Only selected CR neuronal subtypes inspecific brain regions show tangles and/or only mildmorphological changes [24, 25, 30]. This neuroprotectiveeffect cannot be generally extended to other CaBPs sinceparvalbumin-immunopositive neurons are highly sensitiveto degeneration in prion diseases [29]. The cerebellum, theCNS region with the highest neuronal content of CaBPs, isthat least affected in AD [38]. However, it can develop allthe neuropathological hallmarks of TSE (spongiformdegeneration, astroglial/microglial proliferation, neuronal
loss and abnormal PrP deposition) [40, 41] and does so inclose relationship with pathological changes at the synapses[42]. Prominent PrPsc deposition in the cerebellum occursin scrapie, even in supposedly genetically scrapie-resistantsheep [43]. The fact that neurons containing CaBPs otherthan CR express PrPc (the cellular form of the prion proteinnecessary for the development of prion diseases [44]) couldbe related to cell pathology.
The changes in the number of immunopositive cells andthe hypertrophic/hyperimmunoreactive response seen inscrapie-affected (group C) and aged sheep (groups B andF) suggests that some similar and some different mecha-nisms are simultaneously at work in each. Unipolar brushcells were the most sensitive to prion problems despite theirvery high CR content, whereas the response of the othercell subtypes varied in the different lobules. Certainly, thePurkinje cells in the archicerebellum appeared more proneto increasing their CR expression in scrapie, the opposite ofthat seen in the neocerebellum. The wide variety of CB-immunoreactive Purkinje cells in the scrapie-affected sheep(group C), together with their greater CR reactivity,suggests the existence of complex involutive/adaptativemechanisms and/or different responses by different Pur-kinje cell subsets in scrapie and aging. CR and calbindinexpression and/or content seems to be related to the specificfunction of different neuronal subtypes (e.g. the Golgi,Purkinje and Lugaro cells) and to be differently regulated indifferent lobules.
The existence of functionally distinct sets and arrange-ments of neurons and fibres, together with the differentCaBPs involved in the physiological control of intracellularcalcium currents (with important roles in synaptic plasticity,neurotransmission and signalling cascade regulation; [13–15, 33]), plus the different responses to scrapie-inducedneurodegeneration, suggests that the major role of theseproteins is not that of endogenous neuroprotection butfunctional modulation. The present results agree well withthis idea since the unipolar brush cells, the most CR-immunopositive neuronal type, were the most involuted inscrapie-affected animals. In addition, CR-immunopositivityincreased in some normal and hypertrophic Purkinje, Golgiand Lugaro cells. This hypertrophy might be an adaptiveresponse. Different studies on the temporal and spatialcorrelations between CR-immunopositivity and calcium-dependent events, such as synaptogenesis, neurite elonga-tion or remodelling, strongly suggest CR is able to inducemicroplasticity [45]. The results obtained in the study of theCR and CB immunoreactivities showed that each wasassociated with specific involutive/adaptative responses toaging and prion-induced neurodegeneration.
Special cerebellar CR-rich circuits seem to exist [10], themost prominent formed by either bands or groups of CR-immunopositive mossy fibres and the moss-like endings of
602 Cerebellum (2012) 11:593–604

climbing fibres in synaptic contact with the unipolar brushcells (generally in clusters) or close CR-immunopositivePurkinje cells. In scrapie-affected sheep (group C), theseCR-rich circuits were present, but the immunoreactivechanges induced hindered their observation. It has beenproposed that afferent fibres connecting with the unipolarbrush cells might represent a rapid cerebellar reflex circuit,bypassing much of the multisynaptic route of the cortex [4].The reduction in unipolar brush cells in scrapie-affectedanimals might be related to the motor symptoms (ataxia,tremor) associated with the disease.
Most reviews report the granule cells to show the greatestCR expression. These cells provide the major excitatory inputinto Purkinje cells through parallel fibres [13–15]. CR-deficient mice show dramatic alterations in motor coordina-tion and Purkinje cell firing [14, 15, 17], but when thecalcium-buffering capacity [14] or CR-expression is restored[19], normal cerebellar function is recovered. In manyspecies, as well as in the sheep, the CR-immunoreaction ingranule cell perikarya is weak, while that of their axons ismuch more intense. Certainly, CR expression in granule cellsseems to be more important than the staining of theperikarya. The mobilisation of CR to specific subcellulardomains in the axons, dendrites or perikarya seems to bevery important in cerebellar physiology.
This is the first time that CR-immunopositive glial-likecells (protoplasmic astrocytes and Golgi epithelial cells[46]) have been described. This agrees well with astroglialcells in the cerebellar cortex showing CR-immunopositivityin human AD cerebella (Toledano et al., unpublished).Upregulated CaBP expression has been described in somecells in certain situations, e.g. calbindin is expressed byastroglial cells in the injured brain [47] or in culture [48],and strong CR expression is frequently seen in many typesof cell, e.g. malignant tumour cell lines [49]. However, itshould be noted that CR gene promoter activity seems to beregulated differently in neurons and cancer cells [50]. Norelationship was observed between the always dispersedCR-immunopositive glial-like cells and areas of prominentPrPsc deposition.
This work shows that the scrapie prion induces changes ofdifferent sorts in the various CR-immunoreactive cells of thesheep cerebellar cortex. This suggests that different patho-physiological mechanisms are at work in scrapie and that eachsubtype of neuron, perhaps even each cell, may respond in aspecific manner based on its own characteristics and thedamage it has suffered. Future work should include thecolocalisation of physiological and pathological markers incells affected to different extents by the disease.
Acknowledgements This study was partly supported by grants fromthe Spanish Ministry of Science (EET 2001-4844-C03-01 and PN-CTQ 2009–09538).
Conflict of Interest The authors have no conflicts of interest; allwork is only in basic research at official scientific institutions.
References
1. Fortin M, Marchand R, Parent A. Calcium-binding proteins inprimate cerebellum. Neurosci Res. 1998;30:155–68.
2. Geurts FJ, Timmermans J, Shigemoto R, De Schutter E.Morphological and neurochemical differentiation of large granularlayer interneurons in the adult rat cerebellum. Neuroscience.2001;104:499–512.
3. Geurts FJ, De Schutter E, Dieudonne S. Unraveling the cerebellarcortex: cytology and cellular physiology of large-sized interneur-ons in the granular layer. Cerebellum. 2003;2:290–9.
4. Yan XX, Garey LJ. Calretinin immunoreactivity in the monkeyand cat cerebellum: cellular localisation and modular distribution.J Hirnforsch. 1996;37:409–19.
5. Yan XX, Garey LJ. Complementary distributions of calbindin,parvalbumin and calretinin in the cerebellar vermis of the adultcat. J Hirnforsch. 1998;39:9–14.
6. Rogers JH. Immunoreactivity for calretinin and other calcium-binding proteins in cerebellum. Neuroscience. 1989;31:711–21.
7. Braak E, Braak H. The new monodendritic neuronal type withinthe adult human cerebellar granule cell layer shows calretinin-immunoreactivity. Neurosci Lett. 1993;154:199–202.
8. Nunzi MG, Shigemoto R, Mugnaini E. Differential expression ofcalretinin and metabotropic glutamate receptor mGluR1alphadefines subsets of unipolar brush cells in mouse cerebellum. JComp Neurol. 2002;451:189–99.
9. Dino MR, Willard FH, Mugnaini E. Distribution of unipolar brushcells and other calretinin immunoreactive components in themammalian cerebellar cortex. J Neurocytol. 1999;28:99–123.
10. Álvarez MI, Lacruz C, Toledano-Díaz A, Monleón E, Monzón M,Badiola JJ, Toledano A. Calretinin-immunopositive cells andfibres in the cerebellar cortex of normal sheep. Cerebellum.2008;7:417–29.
11. Schafer BW, Heizmann CW. The S100 family of EF-handcalcium-binding proteins: functions and pathology. Trends Bio-chem Sci. 1996;21:134–40.
12. Kawasaki H, Nakayama S, Kretsinger RH. Classification andevolution of EF-hand proteins. Biometals. 1998;11:277–95.
13. Bastianelli E. Distribution of calcium-binding proteins in thecerebellum. Cerebellum. 2003;2:242–62.
14. Gall D, Roussel C, Susa I, D’Angelo E, Rossi P, Bearzatto B,Galas MC, Blum D, Schurmans S, Schiffmann SN. Alteredneuronal excitability in cerebellar granule cells of mice lackingcalretinin. J Neurosci. 2003;23:9320–7.
15. Gall D, Roussel C, Nieus T, Cheron G, Servais L, D’Angelo E,Schiffmann SN. Role of calcium binding proteins in the control ofcerebellar granule cell neuronal excitability: experimental andmodeling studies. Prog Brain Res. 2005;148:321–8.
16. Saftenku EE. Effect of calretinin an Ca(2+) signals in cerebellargranule cells: implications of cooperative Ca(2+) binding.Cerebellum. 2011; doi:10.1007/s12311-011-0263-4
17. Schiffmann SN, Cheron G, Lohof A, d’Alcantara P, Meyer M,Parmentier M, Schurmans S. Impaired motor coordination andPurkinje cell excitability in mice lacking calretinin. Proc NatlAcad Sci USA. 1999;96:5257–62.
18. Servais L, Cheron G. Purkinje cell rhythmicity and synchronicityduring modulation of fast cerebellar oscillation. Neuroscience.2005;134:1247–59.
19. Bearzatto B, Servais L, Roussel C, Gall D, Baba-Aissa F,Schurmans S, de Kerchove d’Exaerde A, Cheron G, SchiffmannSN. Targeted calretinin expression in granule cells of calretinin-
Cerebellum (2012) 11:593–604 603

null mice restores normal cerebellar functions. FASEB J.2006;20:380–2.
20. Villa A, Podini P, Panzeri MC, Racchetti G, Meldolesi J.Cytosolic Ca2+ binding proteins during rat brain ageing: loss ofcalbindin and calretinin in the hippocampus, with no change in thecerebellum. Eur J Neurosci. 1994;6:1491–9.
21. Kishimoto J, Tsuchiya T, Cox H, Emson PC, Nakayama Y. Age-related changes of calbindin-D28k, calretinin, and parvalbuminmRNAs in the hamster brain. Neurobiol Aging. 1998;19:77–182.
22. Lemmens MA, Sierksma AS, Rutten BP, Debbusseb F, StinbuschHW, Lucassen PJ, Schmitz C. Age-related changes of neuronnumbers in the frontal cortex of a transgenic mouse model ofAlzheimer’s disease. Brain Struct Funct. 2011; doi:10.1007/s00429-011-0305-1
23. Fonseca M, Soriano E. Calretinin-immunoreactive neurons in thenormal human temporal cortex and in Alzheimer’s disease. BrainRes. 1995;691:83–91.
24. Sampson VL, Morrison JH, Vickers JC. The cellular basis for therelative resistance of parvalbumin and calretinin immunoreactiveneocortical neurons to the pathology of Alzheimer’s disease. ExpNeurol. 1997;145:295–302.
25. Leuba G, Kraftsik R, Saini K. Quantitative distribution ofparvalbumin, calretinin, and calbindin D-28 k immunoreactiveneurons in the visual cortex of normal and Alzheimer cases. ExpNeurol. 1998;152:278–91.
26. Mikkonen M, Alafuzoff I, Tapiola T, Soininen H, Miettinen R.Subfield- and layer-specific changes in parvalbumin, calretininand calbindin-D28K immunoreactivity in the entorhinal cortex inAlzheimer’s disease. Neuroscience. 1999;92:515–32.
27. Takei H, Buckleair LW, Rivera A, Powell SZ. Brain tissuemicroarrays in neurodegenerative diseases: validation of method-ology and immunohistochemical study of growth-associatedprotein-43 and calretinin. Pathol Int. 2007;57:778–83.
28. Pike CJ, Cotman CW. Calretinin-immunoreactive neurons areresistant to beta-amyloid toxicity in vitro. Brain Res.1995;67:293–8.
29. Guentchev M, Groschup MH, Kordek R, Liberski PP, Budka H.Severe, early and selective loss of a subpopulation of GABAergicinhibitory neurons in experimental transmissible spongiformencephalopathies. Brain Pathol. 1998;8:615–23.
30. Brion JP, Résibois A. A subset of calretinin-positive neurons areabnormal in Alzheimer’s disease. Acta Neuropathol. 1994;88:33–43.
31. Takahashi H, Brasnjevic I, Rutten BP, Van Derr Kolk N, Perl DP,Bouras C, Steinbusch HW, Schmitz C, Hof PR, Dickstein DL.Hippocampal interneuron loss in an APP/PS1 double mutantmouse and in Alzheimer’s disease. Brain Struct Funct.2010;214:145–60.
32. Baglieto-Vargas D, Moreno-Gonzalez I, Sánchez-Varo R, JimenezS, Trujillo-Estrada L, Sánchez-Mejias E, Torres M, Romero-Acebal M, Ruano D, Vizuete M, Vitorica J, Guiterrez A.Calretinin interneurons are early targets of extracellular amyloid-beta pathology in PS1/AbetaPP Alzheimer mice hippocampus. JAlzheimer Dis. 2010;21:119–32.
33. Schwaller B, Meyer M, Schiffmann S. “New” functions for “old”proteins: the role of the calcium-binding proteins calbindin D-28 k, calretinin and parvalbumin, in cerebellar physiology. Studieswith knockout mice. Cerebellum. 2000;1:241–58.
34. Saigal RP, Karamanlidis AN, Voogd J, Michaloudi H, ManganaO. Cerebellar afferents from motor nuclei of cranial nerves, the
nucleus of the solitary tract, and nuclei coeruleus and para-brachialis in sheep, demonstrated with retrograde transport ofhorseradish peroxidase. Brain Res. 1980;197:200–6.
35. Shu S, Ju G, Fan L. The glucose oxidase-DAB-nickel method inperoxidase histochemistry of the nervous system. Neurosci Lett.1988;85:169–71.
36. Monleon E, Monzon M, Hortells P, Vargas A, Acin C, Badiola JJ.Detection of PrPsc on lymphoid tissues from naturally affectedscrapie animals: comparison of three visualization systems. JHistochem Cytochem. 2004;52:145–51.
37. Altman J, Bayer SA. Time of origin and distribution of a new celltype in the rat cerebellar cortex. Exp Brain Res. 1977;29:265–74.
38. Toledano A, Alvarez MI, Rivas L, Lacruz C, Martinez-RodriguezR. Amyloid precursor protein in the cerebellar cortex ofAlzheimer’s disease patients devoid of cerebellar beta amyloiddeposits: immunocytochemical study of five cases. J NeuralTransm. 1999;106:1151–69.
39. Gómez-Tortosa E, Sanders JL, Newell K, Hyman BT. Corticalneurons expressing calcium binding proteins are spared indementia with Lewy bodies. Acta Neuropathol. 2001;101:36–42.
40. Wood JL, McGill IS, Done SH, Bradley R. Neuropathology ofscrapie: a study of the distribution patterns of brain lesions in 222cases of natural scrapie en sheep, 1982–1991. Vet Rec.1997;140:167–74.
41. Siso S, Puig B, Varea R, Vidal E, Acin C, Prinz M, Montrasio F,Badiola J, Aguzzi A, Pumarola M, Ferrer I. Abnormal synapticprotein expression and cell death in murine scrapie. Acta Neuro-pathol. 2002;103:615–26.
42. Ferrer I. Synaptic pathology and cell death in the cerebellum inCreutzfeldt-Jakob disease. Cerebellum. 2002;1:213–22.
43. Buschmann A, Luhken G, Schultz J, Erhardt G, Groschup MH.Neuronal accumulation of abnormal prion protein in sheepcarrying a scrapie-resistant genotype (PrPARR/ARR). J GenVirol. 2004;85:2727–33.
44. Moleres FJ, Velayos JL. Expression of PrP(C) in the rat brain andcharacterization of a subset of cortical neurons. Brain Res.2005;1056:10–21.
45. Schierle GS, Gander JC, D’Orlando C, Ceilo MR, Vogt Weisen-horn DM. Calretinin-immunoreactivity during postnatal develop-ment of the rat isocortex: a qualitative and quantitative study.Cereb Cortex. 1997;7:130–42.
46. Ramón y Cajal S. Histologie du systéme nerveux de l’homme etdes vertébrés. 1911;vol II. Paris, Maloine
47. Mattson MP, Cheng B, Baldwin SA, Smith-Swintosky VL, KellerJ, Geddes JW, Scheff SW, Christakos S. Brain injury and tumornecrosis factors induce calbindin D-28 k in astrocytes: evidencefor a cytoprotective response. J Neurosci Res. 1995;42:357–70.
48. Wernyj RP, Mattson MP, Christakos S. Expression of calbindin-D28k in C6 glial cells stabilizes intracellular calcium levels andprotects against apoptosis induced by calcium ionophore andamyloid beta-peptide. Mol Brain Res. 1999;64:69–79.
49. Lugli A, Forster Y, Haas P, Nocito A, Bucher C, Bissig H,Mirlacher M, Storz M, Mihatsch MJ, Sauter G. Calretininexpression in human normal and neoplastic tissues: a tissuemicroarray analysis on 5233 tissue samples. Hum Pathol.2003;34:994–1000.
50. Billing-Marczak K, Zieminska E, Lesniak W, Lazarewicz JW,Kuznicki J. Calretinin gene promoter activity is differentlyregulated in neurons and cancer cells. Role of AP2-like ciselement and zinc ions. Biochim Biophys Acta. 2004;1678:14–21.
604 Cerebellum (2012) 11:593–604