chapitre 1 reproduction sexuée des organismes …ekladata.com/hyseh4vguxoqtczh0uh2tfdnwa4.pdf ·...
TRANSCRIPT

Chapitre 1Reproduction sexuée des
organismes animaux et végétaux
Partie IID - Ontogenèse et reproduction

3. La fusion des gamètes et la formation d’un nouvel individu
2

Les gamètes, cellules haploïdes
3
spermatozoïde motile
gamète femelleapex
acrosome
empli d'enzymes
d'hydrolyse
noyau haploïde
condensé
centriole proximal
centriole distal
microtubules
spirale
mitochondriale
cytosol réduit
col
tête
pièce intermédiaire
/0"1'00'$.'$23$4)$.'$0+51
pièce principale
pièce terminale
!"#$
!"#$%&'()"*+,+-.'
cellule folliculaire
de la couronne radiée
zone pellucide
1er globule
polaire
noyau bloqué en
métaphase II
membrane plasmique
à microvillosités
granules corticaux
espace périvitellin
mitochondrie

Les gamètes en présence
4
Un seul globule polaire est visible ici : l’ovocyte n’a pas achevé sa méiose
Ovocyte de diamètre 80 µmSpermatozoïde delongueur 100 µm
5 µm

La motilité du spermatozoïde
5

Courbure du flagelle
6

Reconnaissance, fixation et réaction acrosomique
7
28
Du rapprochement des gamètes à la fécondation chez les animaux et végétaux
Le gamète femelle (figure 13.20b) est formé par l’ovocyte II entouré de la zone pellucide (enveloppe de glycoprotéines) et d’une couche de cellules folliculaires (la couronne radiée). L’ovocyte n’a pas achevé sa méiose mais reste bloqué en métaphase de 2e division. L’ovocyte de mammifère ne contient que très peu de réserves. Il abrite des vésicules d’exocytose contenant des enzymes appelées granules corticaux.
Reconnaissance spécifique et fixation à la zone pellucideLes spermatozoïdes s’engagent à travers les espaces entre les cellules de la couronne radiée qu’ils traversent en quelques minutes grâce à la hyaluronidase présente sur leur membrane. Quelques dizaines de spermatozoïdes seulement atteignent la zone pellucide.La reconnaissance de sites spécifiques entre le spermatozoïde et la zone pellucide est un barrage à la fécondation entre espèces différentes. La zone pellucide est composée de glycoprotéines fibrillaires les ZP1, ZP2 et ZP3. La fixation du spermatozoïde à la zone pellucide se fait par l’apex. Chez la souris, la ZP3 est riche en chaînes oligosaccharidiques qui se lient à la galac-tosyl-transférase, enzyme membranaire du spermatozoïde démasquée lors de la capacitation (figure 13.21a). Celle-ci permet ainsi l’accrochage du spermatozoïde à la zone pellucide. Le mécanisme débute à l’apex puis s’étend et la tête du spermatozoïde se couche sur la zone pel-lucide. D’autres protéines que la ZP3 semblent également impliquées dans ces mécanismes de fixation et de reconnaissance spécifique.
noyau
enzyme membranaire
(galactosyltrasnférase)
acrosome
Spermatozoïde
zone pellucide
ovocyte
membrane avec
microvillosités
glycoprotéine
ZP3
oligosaccharide
liaison progressive de la membrane
du spermatozoïde à la zone pellucide
(a) (b)
enzymes de
l'acrosome
déversées
fusion de la membrane de l'acrosome
avec la membrane plasmique
dégradation de la zone pellucide
!"#$
FIGURE 13.21 (a) reconnaissance, fixation ; (b) réaction acrosomique du spermatozoïde.
Réaction acrosomique et franchissement de la zone pellucideLa fixation à la zone pellucide provoque l’activation de récepteurs membranaires du sperma-tozoïde et l’ouverture des canaux Ca2+. L’influx calcique déclenche la fusion de la membrane plasmique et de la membrane externe de l’acrosome. Progressivement, les ouvertures de l’acro-some augmentent de diamètre, confluent et l’ensemble formé par la membrane plasmique et la membrane externe de l’acrosome est résorbé. Ces mécanismes, constituent la réaction acro-somique (figure 13.21b). Il en résulte la libération des enzymes contenues dans l’acrosome : hyaluronidase, acrosine, et β-N-acétylglucosaminidase, à l’endroit même où est fixé le sperma-tozoïde. Ces enzymes permettent la dissolution de l’acide hyaluronique de la zone pellucide. La poussée du spermatozoïde par son flagelle est le moteur de l’avancée oblique vers l’ovocyte, aidée par les enzymes de l’acrosome.En quelques minutes, le spermatozoïde passe la zone pellucide, pénètre dans l’espace périvitellin et se couche contre la membrane plasmique de l’ovocyte II qui est hérissée de microvillosités. Il s’immobilise alors (figure 13.22a).
Liaison aux ZP3 => influx calcique => exocytose de l’acrosome

Étapes de la fécondation
8
noyau
granules corticaux
microvillosités de la
membrane de l'ovocyte
Ovocyte
spermatozoïde
a
H+
Na+
Dépolarisation bloquant
la fusion d'un autre spermatozoïde
Fixation et fusion des membranes
b
centriole proximal
exocytose des granules corticaux
c
le contenu des granules corticaux bloque définitivement
la polyspermie en modifiant la zone pellucide
noyau spermatiquedégénérescence des organites
du spermatozoïde réveil métabolique de l'ovocytefin de méiose
FIGURE 13.22 La fécondation.

Des modifications de l’ovocyte II
9
Contact entre membranes du spermatozoïde et ovocyte II
Effet immédiatentrée de Na+ et sortie d’H+
dépolarisation observée chez l’oursinhyperpolarisation chez les mammifères
=> pas de nouvelle fusion mb possible
Effet différépics d’influx de Ca2+
=> exocytose des granules corticaux
reprise du métabolismefin de méiose

Evolution des pronuclei
10
Interdigitation des
enveloppes nucléairesMigration
Réplication ADN
Duplication du centrosome
Condensation de l’ADN
Rupture des enveloppes nucléaires
Alignement des chromosomes sur la plaque
équatoriale d’un même fuseau
mitotique
✄

Résumé des étapes concernant le spz
11

Le pollen apporte les gamètes mâles
12 http://www.jpboseret.eu/index.php?page=gametogenese-vegetale

Les gamètes femelles
13
32
Du rapprochement des gamètes à la fécondation chez les animaux et végétaux
stigmate
style
cavité ovarienne
ovaire
2 téguments
nucelle (parenchyme
de réserve)
sac embryonnairechalaze
funicule
O
V
U
L
E
!""#$%2 synergides
appareil filiforme
oosphère
gamète femelle
2 noyaux de la
cellule centrale
vacuole
3 antipodes
(rôle nutritif)
paroi
'"#$%
FIGURE 13.24 Ovule et sac embryonnaire chez les angiospermes.
Le déroulement de la double fécondationLe tube pollinique traverse l’appareil filiforme d’une des 2 synergides et, au contact du cyto-plasme, l’apex du tube pollinique éclate, permettant ainsi la décharge des 2 gamètes mâles dans l’une des synergides (figure 13.25a et b).Les 2 gamètes mâles migrent. Un gamète mâle (souvent, le plus riche en plastes) fusionne avec l’oosphère ; ils sont à l’origine du zygote principal diploïde (œuf-embryon). L’autre gamète mâle (le plus riche en mitochondries), fusionne avec la cellule centrale ; ils sont à l’origine du zygote accessoire triploïde (œuf-albumen). Comme deux gamètes mâles sont impliqués dans la fécondation et qu’il s’y forme 2 zygotes, celle-ci est appelée double fécondation (figure 13.25c).
FIGURE 13.25 La double fécondation des angiospermes.

La double fécondation
14
32
Du rapprochement des gamètes à la fécondation chez les animaux et végétaux
FIGURE 13.24 Ovule et sac embryonnaire chez les angiospermes.
Le déroulement de la double fécondationLe tube pollinique traverse l’appareil filiforme d’une des 2 synergides et, au contact du cyto-plasme, l’apex du tube pollinique éclate, permettant ainsi la décharge des 2 gamètes mâles dans l’une des synergides (figure 13.25a et b).Les 2 gamètes mâles migrent. Un gamète mâle (souvent, le plus riche en plastes) fusionne avec l’oosphère ; ils sont à l’origine du zygote principal diploïde (œuf-embryon). L’autre gamète mâle (le plus riche en mitochondries), fusionne avec la cellule centrale ; ils sont à l’origine du zygote accessoire triploïde (œuf-albumen). Comme deux gamètes mâles sont impliqués dans la fécondation et qu’il s’y forme 2 zygotes, celle-ci est appelée double fécondation (figure 13.25c).
cellule centrale
du sac embryonnaire
à deux noyaux
tube pollinique noyau végétatif
2 gamètes mâles
oosphère
synergide
dégénérescente
paroi
appareil filiforme
membrane
de l'oosphère
membrane de
la cellule centrale
synergide
dégénérée
(a) (b) (c)
!"#$%
FIGURE 13.25 La double fécondation des angiospermes.

Evolution du sac embryonnaire
15Mémo visuel de biologie - 2e édition: L'essentiel en fiches Par Daniel Richard,Patrick Chevalet,Thierry Soubaya
Le zygote principal donne naissance à l’embryon (radicule + gemmule ou tigelle + 1 ou 2 cotylédons).Le zygote accessoire donne naissance à l’albumen, tissu de réserve transitoire ou permanent.
périsperme
Graine à périsperme Graine albuminée Graine exalbuminée
rare : les réserves de la graine reste dans le nucelle

Les devenirs de la fleur
16
OVULE → GRAINESAC EMBRYONNAIRE → EMBRYON
OVAIRE → FRUIT
Sac embryonnairezygote principal ➝ embryonzygote accessoire ➝ albumen
✭✭
La graine (avec ou sans le fruit) est dispersée.
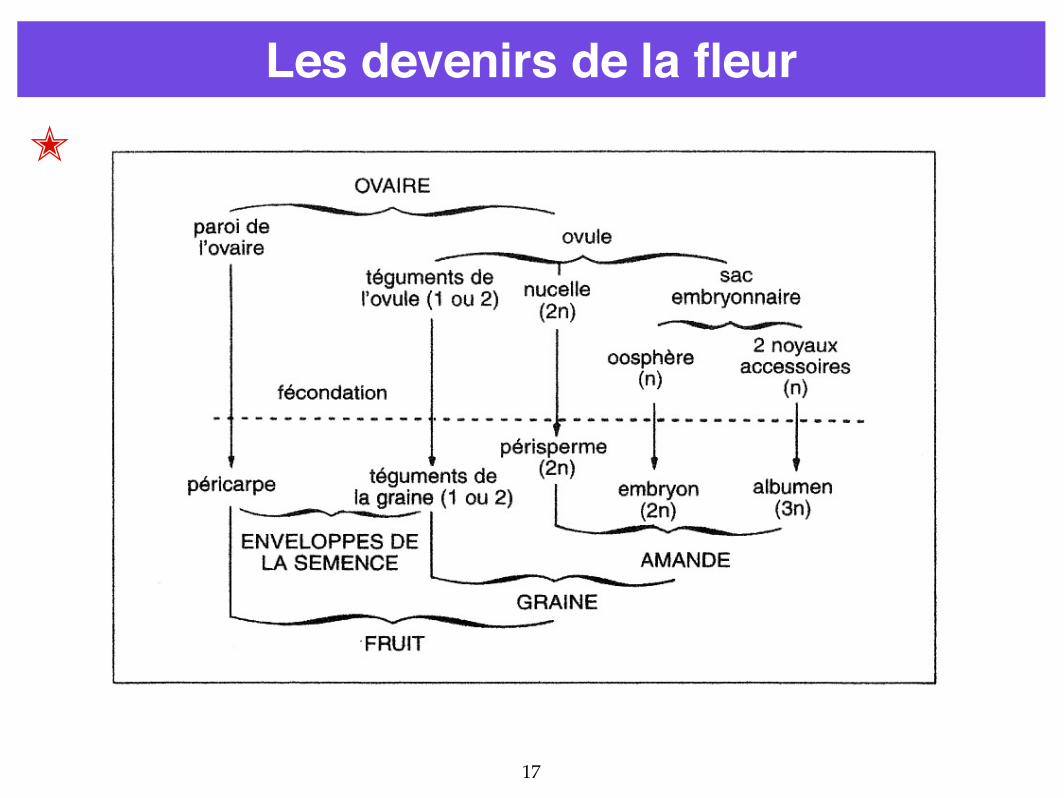
Les devenirs de la fleur
17
✭

4. Des mécanismes favorisant la fusion entre gamètes
de génomes différents
18

Conséquences génétiques de la reproduction
19
Hypothèses sex-ratio 0,5 et 2 descendants par génération
Croissance démographique
Diversificationgénétique

Contrôle de la maturité des prothalles
20
Prothalle à maturité femelle
Archégones mûrsAnthéridiogèneslibération
+
Accélération de la maturité mâle
Anthéridies
spz
1
3
2
4

Incompatibilité sporophytique
21
Locus Sgrande diversité allélique
Lorsqu’un des allèles du parent mâle est commun avec le génotype du parent femelle, le grain de pollen ne germe pas.

Incompatibilité sporophytique
22
Locus S à 3 gènes
- gène scr : le gène est exprimé dans l’anthère qui produit le pollen.Les protéines SCR sont retrouvées sur la paroi d’exine du pollen.
- gène srk : le gène est exprimé dans les cellules de surface du stigmate. Les protéines SRK sont des enzymes membranaires impliquées dans la transduction du signal.
- gène slg : le gène est exprimé dans les cellules de surface du stigmate.Il code pour une glycoprotéine de la paroi, récepteur des SCR. L’allèle slg1 ne reconnaît que la protéine SCR1.

Incompatibilité sporophytique
23Attention, il existe des systèmes complexes
avec des phénomènes de dominance
C. EscuyerSRK4SRK3

Reconnaissance de parentés
24 Ligoud, INRA Prod. Anim., 2006, 19 (2), 119-134
Le CMH, locus très variable, est impliqué dans l’immunité. Il est aussi à l’origine de la sécrétion de protéines odorantes très diverses.

5. La reproduction sexuée dans les cycles de développement
25

Cycle de développement du cerf
26
Cerf mâle spermatozoïdes
Cerf femelle
Méiosephotopériode
(automne)Fécondation
Cellule oeuf
Faon
Parade nuptiale
Développement
intrautérin
Naissance
Croissance
Acquisition de la
maturité sexuelle
automne
fin du printemps
ovocytes
Gamétogenèse
Gamétogenèse
Multiplication
Diversification
Diversification

Cycle de développement du polypode
27
Sporange
Méiose
Spores
Pied feuillé
Prothalle
Anthéridie
Archégone
FécondationOosphère
Spermatozoïde
Zygote
Germination
Germination
Sporophyte
diploïde
(2 n)
Gamétophyte
haploïde
(n)
Gamétogenèse
Sporogenèse
printemps
Multiplication
Diversification
Diversification

Cycle de développement d’une angiosperme
28
Fleur
Pied feuillé
Sac
embryonnaire
Oosphère
2 noyaux polaires
Double
fécondation
Zygote
Pollinisation
Germination
Germination
Sporophyte
diploïde
(2 n)
Gamétophytes
haploïdes
(n)
Etamine
CarpelleMéiose
Pollen
2 spermatozoïdes
Développement
postembryonnaire
Embryon
dans la graine
Développement
embryonnaire
Développement
floral
photopériode
facteurs internes
Gamétogenèse
Sporogenèse
Multiplication
Diversification
Diversification

Caractéristiques d’un cycle de développement
29
Aspect génétiquealternance de phases haploïde / diploïdesexualisationdiversification génétique
Aspect trophique : coût énergétiquedépenses liées au contrôle du cycle (perception des saisons)développement préalable important des organismes matures2 stratégies de reproduction : r et K
Cas particulier des végétaux supérieursalternance de générations : sporophyte et gamétophytes

30
Reproduction asexuée Reproduction sexuée
Rapide, peu coûteux en énergie, simple
Repose sur la mitose et sur la dédifférenciation
Pas de variabilité au niveau du génotype, les individus sont des clones, le génotype parental est conservé sauf mutations
Permet une colonisation rapide d’un habitat homogèneEfficacité dans l’exploitation des ressources
Fragilité vis-à-vis des changements environnementauxFragilité vis-à-vis d’agents pathogènesPropagation de mutations délétères
Lent, coûteux en énergie (cellules spécialisées, parade nuptiale, certains stades fragiles, aléas des rencontres)
Deux évènements clés :la méiose et la fécondation
Variabilité importante des individus :- par la rencontre de gamètes de génotypes éloignés, - par la formation de nouvelles combinaisons d’allèles=> unicité des individus
Moins efficace dans la colonisation rapide, mais des structures de dispersion existent (spores, graines)
Grande aptitude d’adaptation dans un habitat hétérogène, et aux changements environnementauxRésistance aux pathogènesElimination des mutations délétères, capacité de coévolution avec des orga-nismes mutualistes ou pathogènes