characterization of the heparin-binding domain of human extracellular superoxide dismutase
TRANSCRIPT

Ž .Biochimica et Biophysica Acta 1340 1997 21–32
Characterization of the heparin-binding domain of human extracellularsuperoxide dismutase
Lena A.E. Tibell a,), Ingmar Sethson b, Alexei V. Buevich b
a Department of Biochemistry, Umea UniÕersity, S-901 87 Umea, Sweden˚ ˚b Department of Organic Chemistry, Umea UniÕersity, S-901 87 Umea, Sweden˚ ˚
Received 18 November 1996; revised 28 January 1997; accepted 28 January 1997
Abstract
Ž .The C-terminal, heparin-binding domain of human extracellular superoxide dismutase hEC-SOD has been studied as aŽ .fusion to human carbonic anhydrase II HCAII . This technique allows the properties of the EC-SOD domain to be
characterized. At the same time, it allows us to differentiate the contributions from the domain, from those propertiesoriginating from other parts of EC-SOD. The fusion of the 27 C-terminal amino acids of hEC-SOD to the C-terminal of
Ž .HCAII FusCC resulted in the formation of a monomeric protein, which binds to heparin-Sepharose with approximately thesame affinity as the tetrameric hEC-SOD. The structure of the fused C-terminal was characterized by CD and NMRspectroscopy and the data were compatible with the presence of a-helical structures as suggested by secondary structurepredictions. The NMR data show that the C-terminal of FusCC moves independently from the rest of the protein and that itscentral part is involved in conformational exchange. The NOESY spectra demonstrate that the C-terminal in both FusCCand hEC-SOD binds to heparin, and that arginine side chains take part in the binding.
Ž .Keywords: Superoxide dismutase, extracellular; Heparin binding; Human
Abbreviations: FusCC, fusion protein comprising 27 C-termi-nal amino acids of hEC-SOD to the C-terminal of HCAII; GAG,glycosaminoglycan; HSQC, heteronuclear single-quantum coher-ence; PCR, polymerase chain reaction; NMR, nuclear magneticresonance; NOE, nuclear Overhouser enhancement; NOESY, nu-clear Overhouser enhancement spectroscopy; SOD, superoxidedismutase; SOD1, gene encoding intracellular Cu- and Zn-con-taining superoxide dismutase; TOCSY, total correlation spec-troscopy; TPPI, time proportional phase incrementation; CuZn-SOD, intracellular Cu- and Zn-containing superoxide dismutase,EC 1.15.1.1.; EC-SOD, extracellular Cu- and Zn-containing su-peroxide dismutase, EC 1.15.1.1.; rhEC-SOD, recombinant hu-man extracellular superoxide dismutase; HCAII, human carbonicanhydrase II, EC 4.2.1.1.
) Ž . Ž .Corresponding author. Tel. 46 90-16-76-33; Fax: 46 90-16-76-61; e-mail: [email protected]
1. Introduction
Aerobic organisms have developed an array ofdefense mechanisms against oxygen free radicals and
Ž .superoxide dismutases SOD take part in this de-fense. SODs are metalloenzymes that are present inmost aerobic organisms and serve to catalyze thedismutation of superoxide radicals. Mammalian cellsproduce two different types of CuZn-SOD: one intra-
Ž .cellular and one extracellular form EC-SOD .Extra- and intracellular Cu- and Zn-containing
SOD enzymes catalyze the same reaction with aboutthe same efficiency and with nearly identical kinetic
w x Ž .parameters 1 . The central part of the human h EC-
0167-4838r97r$17.00 Copyright q 1997 Elsevier Science B.V. All rights reserved.Ž .PII S0167-4838 97 00024-1

( )L.A.E. Tibell et al.rBiochimica et Biophysica Acta 1340 1997 21–3222
SOD sequence is clearly homologous with the se-quence of the intracellular CuZnSODs, including themetal-binding ligands and SS-forming cysteines thathave been demonstrated in CuZnSOD by X-ray crys-tallography. However, the two enzyme forms alsodiffer in several respects. The human intracellularCuZnSOD contains 153 amino acids per subunit, hasa subunit molecular mass of 16 kDa and is a dimer.On the other hand, hEC-SOD is a tetrameric secretedprotein in which each subunit contains 222 aminoacids and a N-linked oligosaccharide, resulting in a
Žcalculated molecular mass of 26.7 kDa 24.2kDafrom the peptide and about 2.5 kDa from the oligo-
. w xsaccharide 2,3 . The hEC-SOD forms a tetramerthat is secreted. We have earlier demonstrated theN-terminal domain of hEC-SOD to be responsible for
w xthe tetramerization 4 .Heparin sulfate and other glycosaminoglycans
Ž .GAG are present in large amounts in the extracellu-lar matrix of different tissues where they are involved
w xin different cellular activities 5 . One of the charac-teristic properties of hEC-SOD is its binding to sul-fated glycosaminoglycans such as heparin and to
w xheparin-Sepharose 6 . In addition to hEC-SOD, awide range of extracellular proteins have been shownto bind to heparin and heparin sulfate. Many of theseproteins are involved in the blood coagulation path-way and participate in signal transduction, cell regu-
Žlation e.g. those that are connected with neutrophil.activation, growth-factors and cytokines or inflam-
w xmation 7 . Several reports have shown that the C-terminal of EC-SOD is essential for binding to hep-
w xarin 8,9 . However, it is not known whether thebinding is moderated by interactions to other parts of
w xthe protein. Sandstrom et al. 9 and Carlsson et al.¨w x10 concluded that the heparin binding is cooperativeand dependent on the tetramerization, while Willems
w xet al. 11 proposed that heparin binding was depen-dent on the structure of the C-terminal domain.
Our investigation aimed to elucidate whether theC-terminal domain is solely responsible for this hep-arin affinity, to get some structural information aboutthe C-terminal domain, and to elucidate if the te-trameric structure of hEC-SOD affects the heparinaffinity, i.e. whether EC-SOD heparin binding iscooperative.
For this study we constructed a fusion proteincomprising the C-terminal domain of EC-SOD and
Fig. 1. Coomassie blue-stained SDS-PAGE slab gels of FusCCand HCAII. Lane 1: FusCC after affinity purification. Lane 2:Total cell extract of FusCC producing E. coli.
Ž .human carbonic anhydrase II HCAII . This tech-nique has obvious technical advantages for produc-
w xtion and purification 4 . However, the most impor-tant advantage of the technique in this context is thatdifferent domains of EC-SOD can be expressed bothindependently and in combinations, and the func-tional roles of individual domains can be studied
Ž .separately. The fusion protein, FusCC Fig. 1 , con-sists of HCAII, followed by a poly Gly-spacer and
Žthe 27 C-terminal amino acids of hEC-SOD amino.acids 196-222 :
FusCC s HCAII y AlaGlyGlyGlyGlySer y 27
=C-terminal aa of hEC-SOD
The fusion protein was readily produced in Es-cherichia coli, and then purified to homogeneity.
2. Material and methods
2.1. Construction of the gene fusion
Gene fusion was made using the expression plas-w xmid pACA 12 . This plasmid contains two restriction
sites for the restriction enzyme HindIII and, prior toconstruction of the gene fusion, one of these sites wasremoved by in vitro site-directed mutagenesis accord-
w xing to the method of Kunkel 13 . The resultingplasmid, pACA1, contains an unique HindIII site 9nucleotides before the corresponding stop codon. The

( )L.A.E. Tibell et al.rBiochimica et Biophysica Acta 1340 1997 21–32 23
DNA fragment coding for the C-terminal portion ofhEC-SOD was isolated from a human placenta cDNA
Ž .library kindly provided by J.L. Millan by use ofP C R a n d th e o lig o n u c le o t id e s : 5 X-GCGCGCAAAGCTTCCGCTGGAGGCGGTGGAA-G C G G G C C C G G G C T C - 3 X , a n d 5 X-GACTGCAAGCTTCTATCAGGCGGCCTTGCAC-TCGCTCTC-3X. This resulted in PCR synthesis of aDNA fragment with the following composition: 7nucleotides – HindIII site – codons for Ser-Ala-Gly-Gly-Gly-Gly-Ser – 81 nucleotides coding for the 27C-terminal amino acids of hEC-SOD – two stopcodons – HindIII site – 6 nucleotides.
The DNA fragment and the expression plasmidwere both cleaved by HindIII and subsequently puri-fied by using agarose gel electrophoresis. The vectorfragment was treated by alkaline phosphatase fol-lowed by ligation to the DNA fragment and transfec-
Ž . w xtion of E. coli BL21 DE3 14 . Several colonies ofthe transfected E. coli were isolated, and the com-plete coding regions of their corresponding plasmidswere checked by DNA sequencing before a singleclone was used for protein production. All sequenc-ing was performed according to the chain-termination
w xmethod 15 . The plasmid was given the namepFusCC.
2.2. Isolation and purification of the fusion protein
Ž .E. coli BL21 DE3 harboring the constructedexpression plasmid was grown in 2=Luria Broth,0.5 mM ZnSO and 50 mgrml ampicillin at 238C.415N-enriched FusCC was produced by growing thecells in minimal medium containing 0.5 grl 15NH Cl4w x16 . Protein production was initiated by adding IPTGto a final concentration of 0.5 mM at a cell densitycorresponding to OD660 s0.5, and then allowing thecells to grow for 6–12 h. The fusion protein was
w xpurified by affinity chromatography 4,17 .
2.3. Analysis of fusion protein
The CO -hydration activity of HCAII was mea-2w xsured according to Rickli 18 . N-terminal amino acid
sequences were determined using an Applied Biosys-tems model 477A sequencing system. Determinationof the protein extinction coefficient was carried out
w xaccording to the method of Gill and von Hippel 19 ,
which is based on the amino acid composition ofeach protein and the measured absorption values at280 nm for the native protein, and protein denaturedin 6 M guanidinium chloride. SDS-PAGE, Westernblotting and immunodetections were performed as
w xdescribed earlier 4 . Isoelectric focusing was carriedout using a PhastGel IEF 3–9 gradient medium and aPharmacia PhastSystem instrument. The apparentmolecular mass of each of the native fusion proteinwas estimated by size-exclusion chromatography on a
w xSuperose 12 column as described previously 4 . Cir-Ž .cular dichroism CD spectra were recorded on a
ŽCD6 spectrodichrograph Jobin-Yvon Instruments. w xSA, Longjumeau, France as described previously 4 .
2.4. Heparin-Sepharose chromatography
Chromatography of the proteins was carried out asw xpreviously described 20 at room temperature in a
2.5 ml heparin-Sepharose column using an equilibra-tion buffer composed of 15 mM sodium cacodylate,pH 6.5, containing 50 mM NaCl. Bound proteinswere eluted using the same buffer with a gradient of
Ž .NaCl 50 mM–1 M . A Pharmacia FPLC apparatuswas used. The FusCC protein showed that a minorfraction did not bind to the column, some smallerfractions eluted at about 0.3–0.4 M NaCl, and amajor fraction eluted at about 0.55 M NaCl. All thosepeaks contained HCAII activity, and were found toreact with both anti-HCAII IgG and anti-hEC-SODIgG. SDS-PAGE analysis of the different peaksshowed that the earlier fractions had somewhat lowermolecular masses than the fractions that eluted laterin the salt gradient. The most probable explanationfor these results is that minor fractions of the C-terminal of FusCC was partially degraded, resultingin lower affinity for heparin-Sepharose for the de-graded species. Repeated chromatography of the ma-
Ž .jor fraction eluting at 0.55 M NaCl resulted in achromatogram with only one single peak at 0.55 M
Ž .NaCl Fig. 6A .
2.5. NMR measurements
The NMR samples of 0.6–1.0 mM concentrationŽ .were prepared in aqueous solution 10% D O with2
20 mM phosphate buffer at pH 7.0 and 40 mM NaCl.The NMR measurements were done at 308C, 78C and

( )L.A.E. Tibell et al.rBiochimica et Biophysica Acta 1340 1997 21–3224
28C on a AMX2-500 Bruker spectrometer operatingat a proton frequency of 500.13 MHz. The 1H chemi-cal shift of the water signal was used as a reference:4.70 ppm at 308C; 4.98 ppm at 78C; and 5.04 ppm at
w x28C 21 . The nitrogen chemical shifts were calcu-15 1 w xlated using J ratio Nr Hs0.101329118 21 .
w xThe two-dimensional NOESY 22 and TOCSYw x23 experiments were recorded in the phase-sensitive
w xTPPI mode 24 . Water suppression was achieved byw xthe WATERGATE method 25 . 200 ms mixing time
and 70 ms spin lock time were used in NOESY andTOCSY experiments, respectively. 256 complexpoints in the t and 1024 complex points in the t1 2
dimensions were acquired. 16 or 32 scans per eacht -point with 1–2 s delay between scans were used.1
The spectral width was 8333.3 Hz in both dimen-sions. A lorentzian-gaussian window function in t2
and pr2-shifted squared sine function in t dimen-1
sions were applied prior to Fourier transformation.Ž . Ž1 15 .The two-dimensional 2D H- N -HSQC exper-
iments were performed in the sensitivity-enhancedw xgradient selection mode 26 . The number of acquired
Ž15 .points were 128 complex points in t N and 10241Ž1 .complex points in t H . The spectral widths were2
8333.3 Hz and 3598.1 Hz in the t and t dimensions,2 1
respectively. 8, 16 or 32 scans were acquired forevery t -point with 1 s delay between scans. The t1 1
points were linear predicted to 256 complex pointsand thereafter zero-filled to 512. A lorentzian-gaus-sian window function in t and pr2-shifted squared2
sine function in the t dimension were applied prior1
to Fourier transformation.Ž .The three-dimensional 3D NOESY-HSQC and
TOCSY-HSQC experiments were performed in the
States-TPPI mode for the NOESY- and TOCSY-Ž .transfer t and in the gradient selection mode for1
Ž .the HSQC-transfer t . The number of acquired2
complex points were 1024)64)16 or 1024)50)16 inŽ1 . Ž1 . Ž15 .t H , t H and t N dimensions, respec-3 N 2 1
tively. 16 scans for each increment and 1 s delaybetween scans were used. Linear prediction andzero-filling in the indirect dimensions finally gave amatrix of 1024=256=64 complex points. The pro-cessing of the 3D spectra were performed with thesame window functions which were used in the cor-responding 2D experiments.
w Ž .Heparin Lovens was 5000 IUr ml 33 mgrml¨Ž .from Løvens Kemiske Fabrik Ballerup, Denmark .
Recombinant human EC-SOD was a generous giftfrom Symbicom AB.
3. Results and discussion
3.1. FusCC fusion protein
The fusion protein, FusCC, was produced in E.coli, resulting in an approximate yield of 30% of the
Žtotal protein content of the E. coli cultures Fig. 1,. Žlane 2 and purified by affinity chromatography Fig.
.1, lane 1 .A number of analyses were carried out to verify
the identity of the fusion protein. The expressionplasmid was isolated from the clone that was used forproduction of the fusion protein. DNA sequencing ofthis plasmid confirmed the construction and alsoshowed that no mutations had occurred sponta-neously. The produced fusion protein reacted with
Table 1A comparison of physiochemical properties of fusion and native proteins
d eŽ .Protein Apparent molecular mass kD p I Specific CO activity Extinction coefficient2
aNative Subunitb c y1 y1Ž . Ž . Ž . Ž .Observed Predicted AUrmg M cm
FusCC 28 32 33 8.0 4 850 59 800HCAII 30 30 29 7.3 20 000 54 800EC-SOD 150 32 27 4.9 y 42 040
a Estimated from size-exclusion chomatography using Superose 12.b Estimated from SDS-PAGE.c Calculated from amino acid sequences and oligosacharide sequence in the case of EC-SOD.d w xMeasured as described by Rickli et al. 18 .e w xMeasured as described by Gill and von Hippel 19 .
( )L.A.E. Tibell et al.rBiochimica et Biophysica Acta 1340 1997 21–32 25
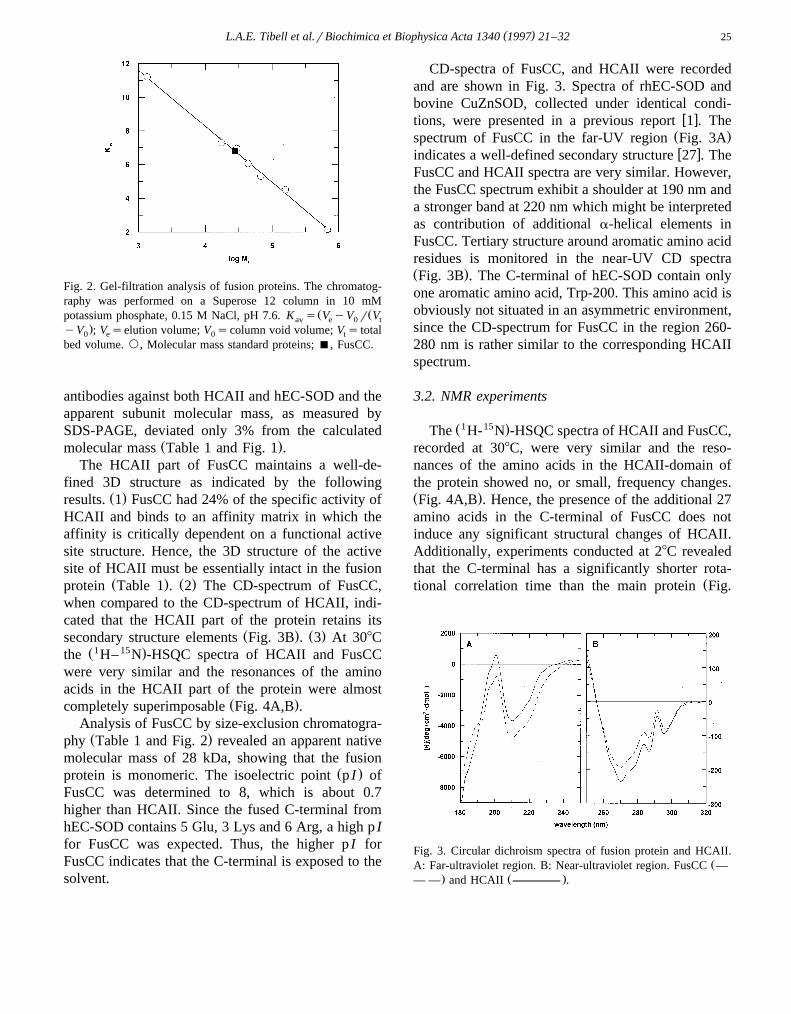
Fig. 2. Gel-filtration analysis of fusion proteins. The chromatog-raphy was performed on a Superose 12 column in 10 mM
Ž Žpotassium phosphate, 0.15 M NaCl, pH 7.6. K s V yV r Vav e 0 t.yV ; V selution volume; V scolumn void volume; V s total0 e 0 t
bed volume. `, Molecular mass standard proteins; B, FusCC.
antibodies against both HCAII and hEC-SOD and theapparent subunit molecular mass, as measured bySDS-PAGE, deviated only 3% from the calculated
Ž .molecular mass Table 1 and Fig. 1 .The HCAII part of FusCC maintains a well-de-
fined 3D structure as indicated by the followingŽ .results. 1 FusCC had 24% of the specific activity of
HCAII and binds to an affinity matrix in which theaffinity is critically dependent on a functional activesite structure. Hence, the 3D structure of the activesite of HCAII must be essentially intact in the fusion
Ž . Ž .protein Table 1 . 2 The CD-spectrum of FusCC,when compared to the CD-spectrum of HCAII, indi-cated that the HCAII part of the protein retains its
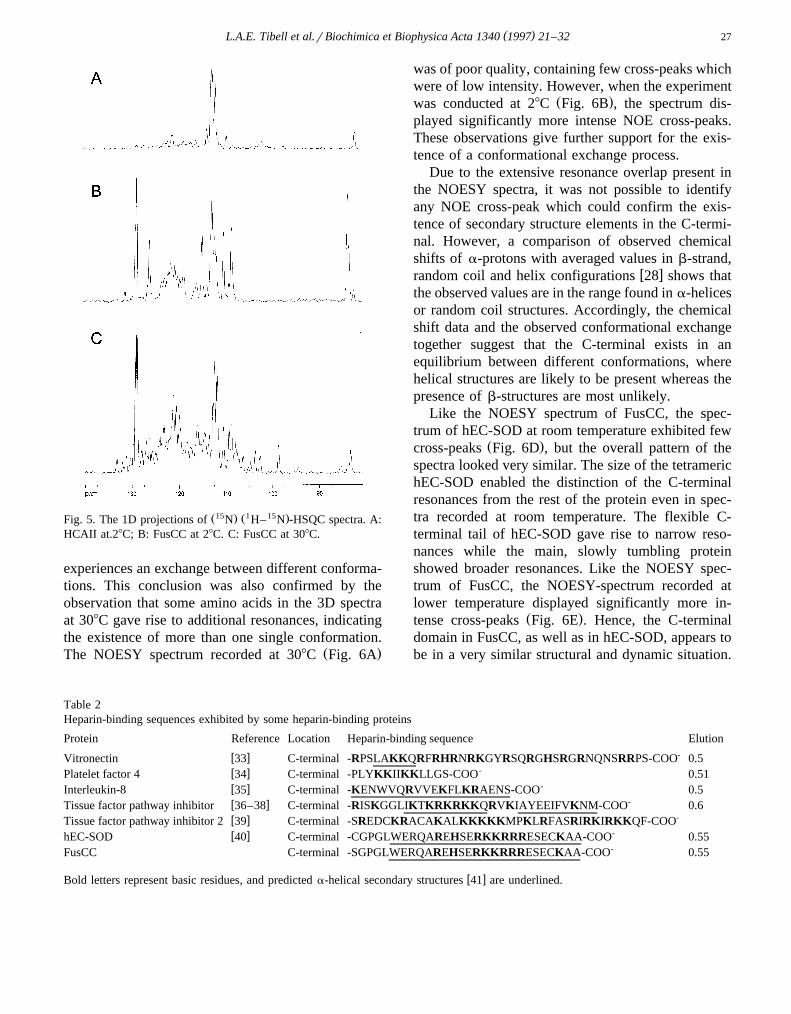
Ž . Ž .secondary structure elements Fig. 3B . 3 At 308CŽ1 15 .the H– N -HSQC spectra of HCAII and FusCC
were very similar and the resonances of the aminoacids in the HCAII part of the protein were almost
Ž .completely superimposable Fig. 4A,B .Analysis of FusCC by size-exclusion chromatogra-Ž .phy Table 1 and Fig. 2 revealed an apparent native
molecular mass of 28 kDa, showing that the fusionŽ .protein is monomeric. The isoelectric point p I of
FusCC was determined to 8, which is about 0.7higher than HCAII. Since the fused C-terminal fromhEC-SOD contains 5 Glu, 3 Lys and 6 Arg, a high p Ifor FusCC was expected. Thus, the higher p I forFusCC indicates that the C-terminal is exposed to thesolvent.
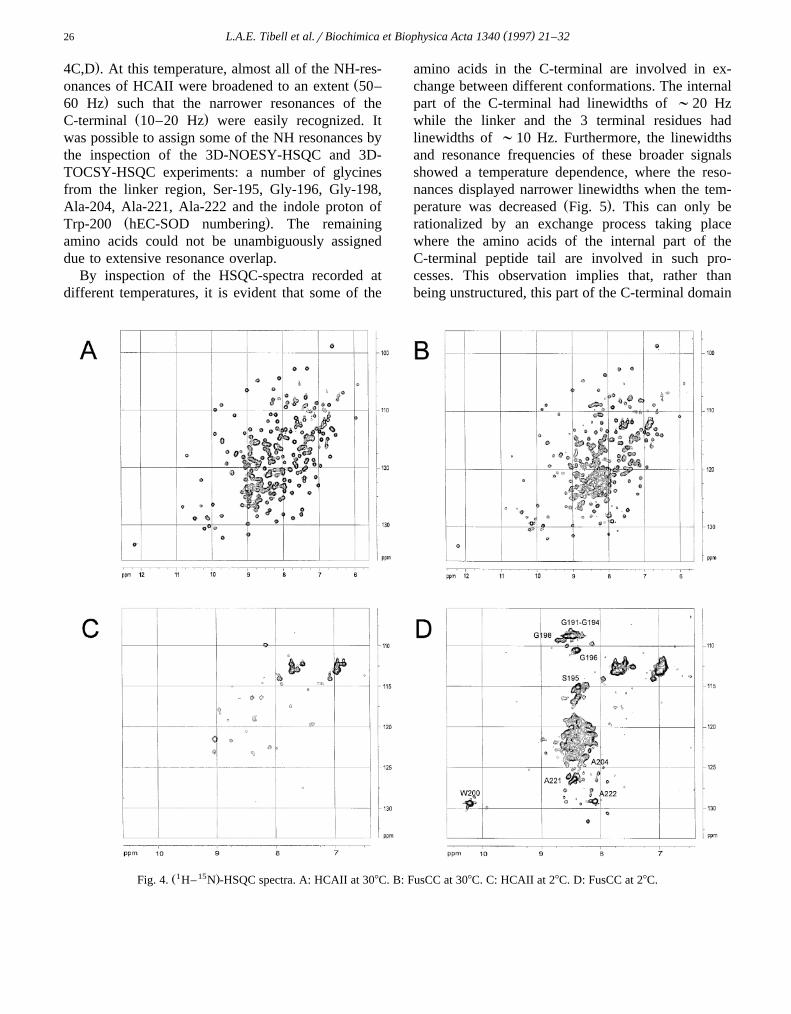
CD-spectra of FusCC, and HCAII were recordedand are shown in Fig. 3. Spectra of rhEC-SOD andbovine CuZnSOD, collected under identical condi-
w xtions, were presented in a previous report 1 . TheŽ .spectrum of FusCC in the far-UV region Fig. 3Aw xindicates a well-defined secondary structure 27 . The
FusCC and HCAII spectra are very similar. However,the FusCC spectrum exhibit a shoulder at 190 nm anda stronger band at 220 nm which might be interpretedas contribution of additional a-helical elements inFusCC. Tertiary structure around aromatic amino acidresidues is monitored in the near-UV CD spectraŽ .Fig. 3B . The C-terminal of hEC-SOD contain onlyone aromatic amino acid, Trp-200. This amino acid isobviously not situated in an asymmetric environment,since the CD-spectrum for FusCC in the region 260-280 nm is rather similar to the corresponding HCAIIspectrum.
3.2. NMR experiments
Ž1 15 .The H- N -HSQC spectra of HCAII and FusCC,recorded at 308C, were very similar and the reso-nances of the amino acids in the HCAII-domain ofthe protein showed no, or small, frequency changes.Ž .Fig. 4A,B . Hence, the presence of the additional 27amino acids in the C-terminal of FusCC does notinduce any significant structural changes of HCAII.Additionally, experiments conducted at 28C revealedthat the C-terminal has a significantly shorter rota-
Žtional correlation time than the main protein Fig.
Fig. 3. Circular dichroism spectra of fusion protein and HCAII.ŽA: Far-ultraviolet region. B: Near-ultraviolet region. FusCC —
. Ž .— — and HCAII .
( )L.A.E. Tibell et al.rBiochimica et Biophysica Acta 1340 1997 21–3226
.4C,D . At this temperature, almost all of the NH-res-Žonances of HCAII were broadened to an extent 50–
.60 Hz such that the narrower resonances of theŽ .C-terminal 10–20 Hz were easily recognized. It
was possible to assign some of the NH resonances bythe inspection of the 3D-NOESY-HSQC and 3D-TOCSY-HSQC experiments: a number of glycinesfrom the linker region, Ser-195, Gly-196, Gly-198,Ala-204, Ala-221, Ala-222 and the indole proton of
Ž .Trp-200 hEC-SOD numbering . The remainingamino acids could not be unambiguously assigneddue to extensive resonance overlap.
By inspection of the HSQC-spectra recorded atdifferent temperatures, it is evident that some of the
amino acids in the C-terminal are involved in ex-change between different conformations. The internalpart of the C-terminal had linewidths of ;20 Hzwhile the linker and the 3 terminal residues hadlinewidths of ;10 Hz. Furthermore, the linewidthsand resonance frequencies of these broader signalsshowed a temperature dependence, where the reso-nances displayed narrower linewidths when the tem-
Ž .perature was decreased Fig. 5 . This can only berationalized by an exchange process taking placewhere the amino acids of the internal part of theC-terminal peptide tail are involved in such pro-cesses. This observation implies that, rather thanbeing unstructured, this part of the C-terminal domain
Ž1 15 .Fig. 4. H– N -HSQC spectra. A: HCAII at 308C. B: FusCC at 308C. C: HCAII at 28C. D: FusCC at 28C.
( )L.A.E. Tibell et al.rBiochimica et Biophysica Acta 1340 1997 21–32 27
Ž15 . Ž1 15 .Fig. 5. The 1D projections of N H– N -HSQC spectra. A:HCAII at.28C; B: FusCC at 28C. C: FusCC at 308C.
experiences an exchange between different conforma-tions. This conclusion was also confirmed by theobservation that some amino acids in the 3D spectraat 308C gave rise to additional resonances, indicatingthe existence of more than one single conformation.
Ž .The NOESY spectrum recorded at 308C Fig. 6A
was of poor quality, containing few cross-peaks whichwere of low intensity. However, when the experiment
Ž .was conducted at 28C Fig. 6B , the spectrum dis-played significantly more intense NOE cross-peaks.These observations give further support for the exis-tence of a conformational exchange process.
Due to the extensive resonance overlap present inthe NOESY spectra, it was not possible to identifyany NOE cross-peak which could confirm the exis-tence of secondary structure elements in the C-termi-nal. However, a comparison of observed chemicalshifts of a-protons with averaged values in b-strand,
w xrandom coil and helix configurations 28 shows thatthe observed values are in the range found in a-helicesor random coil structures. Accordingly, the chemicalshift data and the observed conformational exchangetogether suggest that the C-terminal exists in anequilibrium between different conformations, wherehelical structures are likely to be present whereas thepresence of b-structures are most unlikely.
Like the NOESY spectrum of FusCC, the spec-trum of hEC-SOD at room temperature exhibited few
Ž .cross-peaks Fig. 6D , but the overall pattern of thespectra looked very similar. The size of the tetramerichEC-SOD enabled the distinction of the C-terminalresonances from the rest of the protein even in spec-tra recorded at room temperature. The flexible C-terminal tail of hEC-SOD gave rise to narrow reso-nances while the main, slowly tumbling proteinshowed broader resonances. Like the NOESY spec-trum of FusCC, the NOESY-spectrum recorded atlower temperature displayed significantly more in-
Ž .tense cross-peaks Fig. 6E . Hence, the C-terminaldomain in FusCC, as well as in hEC-SOD, appears tobe in a very similar structural and dynamic situation.
Table 2Heparin-binding sequences exhibited by some heparin-binding proteins
Protein Reference Location Heparin-binding sequence Elution-w xVitronectin 33 C-terminal -RPSLAKKQRFRHRNRKGYRSQRGHSRGRNQNSRRPS-COO 0.5
-w xPlatelet factor 4 34 C-terminal -PLYKKIIKKLLGS-COO 0.51-w xInterleukin-8 35 C-terminal -KENWVQRVVEKFLKRAENS-COO 0.5
-w xTissue factor pathway inhibitor 36–38 C-terminal -RISKGGLIKTKRKRKKQRVKIAYEEIFVKNM-COO 0.6-w xTissue factor pathway inhibitor 2 39 C-terminal -SREDCKRACAKALKKKKKMPKLRFASRIRKIRKKQF-COO
-w xhEC-SOD 40 C-terminal -CGPGLWERQAREHSERKKRRRESECKAA-COO 0.55-FusCC C-terminal -SGPGLWERQAREHSERKKRRRESECKAA-COO 0.55
w xBold letters represent basic residues, and predicted a-helical secondary structures 41 are underlined.

( )L.A.E. Tibell et al.rBiochimica et Biophysica Acta 1340 1997 21–3228
Fig.
6.A
mid
epr
oton
regi
ons
ofN
OE
SYsp
ectr
a.A
:Fu
sCC
at30
8C.
B:
FusC
Cat
28C
.C
:Fu
sCC
at30
8Cw
ithhe
pari
n.D
:hE
C-S
OD
at30
8C.
E:
hEC
-SO
Dat
28C
.F:
hEC
-SO
Dat
308C
with
hepa
rin.
( )L.A.E. Tibell et al.rBiochimica et Biophysica Acta 1340 1997 21–32 29
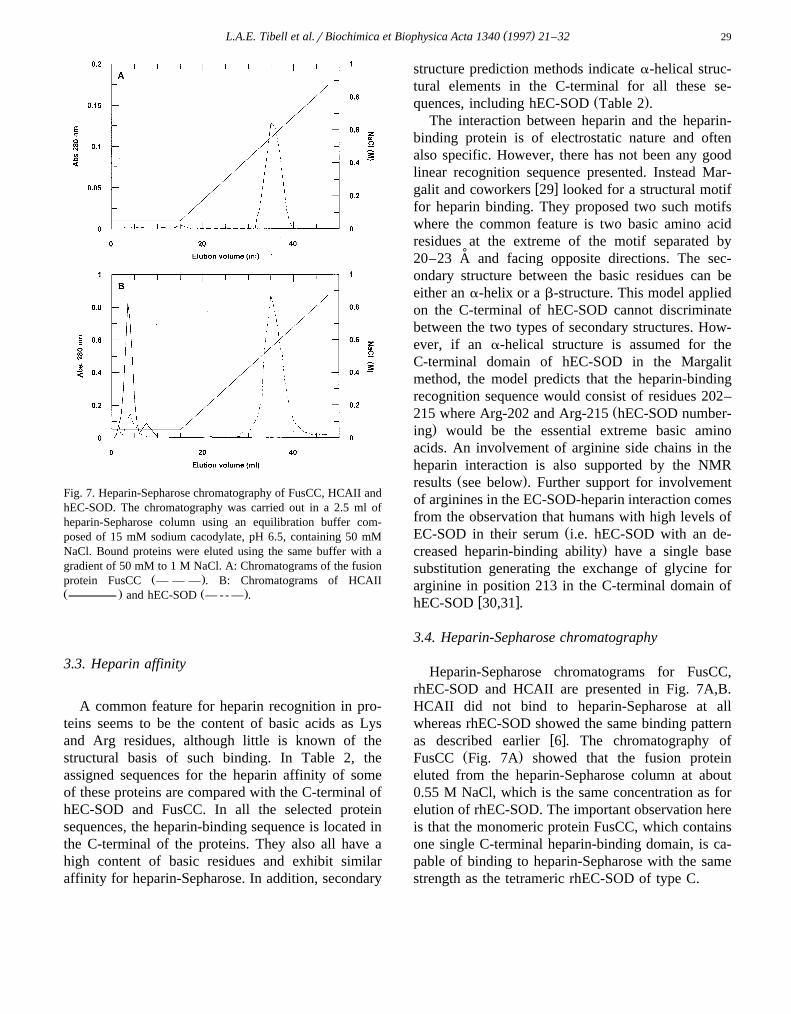
Fig. 7. Heparin-Sepharose chromatography of FusCC, HCAII andhEC-SOD. The chromatography was carried out in a 2.5 ml ofheparin-Sepharose column using an equilibration buffer com-posed of 15 mM sodium cacodylate, pH 6.5, containing 50 mMNaCl. Bound proteins were eluted using the same buffer with agradient of 50 mM to 1 M NaCl. A: Chromatograms of the fusion
Ž .protein FusCC — — — . B: Chromatograms of HCAIIŽ . Ž .and hEC-SOD — - - — .
3.3. Heparin affinity
A common feature for heparin recognition in pro-teins seems to be the content of basic acids as Lysand Arg residues, although little is known of thestructural basis of such binding. In Table 2, theassigned sequences for the heparin affinity of someof these proteins are compared with the C-terminal ofhEC-SOD and FusCC. In all the selected proteinsequences, the heparin-binding sequence is located inthe C-terminal of the proteins. They also all have ahigh content of basic residues and exhibit similaraffinity for heparin-Sepharose. In addition, secondary
structure prediction methods indicate a-helical struc-tural elements in the C-terminal for all these se-
Ž .quences, including hEC-SOD Table 2 .The interaction between heparin and the heparin-
binding protein is of electrostatic nature and oftenalso specific. However, there has not been any goodlinear recognition sequence presented. Instead Mar-
w xgalit and coworkers 29 looked for a structural motiffor heparin binding. They proposed two such motifswhere the common feature is two basic amino acidresidues at the extreme of the motif separated by
˚20–23 A and facing opposite directions. The sec-ondary structure between the basic residues can beeither an a-helix or a b-structure. This model appliedon the C-terminal of hEC-SOD cannot discriminatebetween the two types of secondary structures. How-ever, if an a-helical structure is assumed for theC-terminal domain of hEC-SOD in the Margalitmethod, the model predicts that the heparin-bindingrecognition sequence would consist of residues 202–
Ž215 where Arg-202 and Arg-215 hEC-SOD number-.ing would be the essential extreme basic amino
acids. An involvement of arginine side chains in theheparin interaction is also supported by the NMR
Ž .results see below . Further support for involvementof arginines in the EC-SOD-heparin interaction comesfrom the observation that humans with high levels of
ŽEC-SOD in their serum i.e. hEC-SOD with an de-.creased heparin-binding ability have a single base
substitution generating the exchange of glycine forarginine in position 213 in the C-terminal domain of
w xhEC-SOD 30,31 .
3.4. Heparin-Sepharose chromatography
Heparin-Sepharose chromatograms for FusCC,rhEC-SOD and HCAII are presented in Fig. 7A,B.HCAII did not bind to heparin-Sepharose at allwhereas rhEC-SOD showed the same binding pattern
w xas described earlier 6 . The chromatography ofŽ .FusCC Fig. 7A showed that the fusion protein
eluted from the heparin-Sepharose column at about0.55 M NaCl, which is the same concentration as forelution of rhEC-SOD. The important observation hereis that the monomeric protein FusCC, which containsone single C-terminal heparin-binding domain, is ca-pable of binding to heparin-Sepharose with the samestrength as the tetrameric rhEC-SOD of type C.

( )L.A.E. Tibell et al.rBiochimica et Biophysica Acta 1340 1997 21–3230
3.5. NMR and the interaction with heparin
When FusCC was titrated with heparin the NMRexperiments revealed that significant changes oc-
Ž .curred Fig. 6C . The NOESY spectrum showed muchmore intense NOE cross-peaks, which resembled thepattern observed in the NOESY spectrum of FusCCrecorded at 28C. By inspection of NOESY spectrarecorded on pure heparin samples, it was evident thatthese changes were not caused by interactions withinthe added heparin. Even more conclusive evidencefor heparin binding was found when the changes ofthe arginine guanidinium protons were inspected. Atlow temperature the exchange between the guani-dinium protons of the arginines in the C-terminal andwater was slow enough to allow detection. The fre-quencies of these resonances changed significantly
Ž .upon binding to heparin Fig. 8 . This observation isw xin a good agreement with a recent publication 32 in
which the detailed structure of the interaction be-tween heparin-derived tetra- and hexamers and basicfibroblast growth factor is described. In that study theimportance of the interaction between charged sidechains of basic amino acids and heparin sulfate groupswas emphasized. In our study it was not possible,from the NMR spectra, to deduce if Lys residues areinvolved in the heparin binding.
When heparin was added to hEC-SOD, the inten-sity of the existing cross-peaks in the NOESY spec-trum at room temperature increased significantly and
Ž .a number of new cross-peaks appeared Fig. 6F . It isalso interesting to note that these changes were dif-ferent from those observed in the NOESY spectrumof pure hEC-SOD recorded at low temperature, whichrules out the possibility that the appearing NOEcross-peaks are solely due to viscosity changes. Ac-cordingly, heparin binds to the C-terminal as evidentfrom the observed changes in the NOESY spectrum.
In summary one can conclude that FusCC clearlyis a monomeric protein, which contain one singleC-terminal heparin-binding domain and with a hep-arin affinity of the same strength as that shown byhEC-SOD. The strong binding to heparin-Sepharosecan, most probably, be assigned to the C-terminaldomain alone since HCAII does not bind to heparin-
Ž .Sepharose at all Fig. 7B . In addition, it is notprobable that fusion of the C-terminal to HCAIIcreate a novel combined binding site, because HCAIIdoes not contain any patches with basic residues, butinstead the charged residues are evenly distributed onthe molecule. Further, the observation that the C-terminal constitutes the functional structure in FusCCindicate that the heparin binding of EC-SOD is notmodulated by the rest of the EC-SOD molecule. In
Ž1 15 .Fig. 8. The arginine-guanidinium resonances of H– N -HSQC spectra. A: FusCC at 28C. B: FusCC at 28C with addition of heparin.

( )L.A.E. Tibell et al.rBiochimica et Biophysica Acta 1340 1997 21–32 31
addition, the NMR results indicate that the C-terminaldomain moves independently from the main protein,which supports its role as an independent heparin-binding unit.
Based on results, that show differences inheparin-binding affinity for homo- and heterote-
w xtramers consisting of wild-type and natural 31 orw xconstructed 9,10 mutants, it has been proposed
w x9,10,31 that the heparin binding to EC-SOD iscooperative. However, our results, that FusCC bindsto heparin with the same affinity as the EC-SODvariant having the highest affinity, indicate that thebinding of hEC-SOD to heparin-Sepharose is notnecessarily cooperative in nature.
From our NMR results it has not been possible toelucidate the structure of the C-terminal but, ratherthan being unstructured, it is most likely involved inconformational exchange. The presence of a-helicalconformations are probable, whereas b-structures areunlikely to occur. Finally, the heparin binding to theC-terminal of FusCC and hEC-SOD is evident, andinvolvement of arginine side chains in the interactionare confirmed by the NMR results.
Acknowledgements
The authors wish to thank Dr. Karin Hjalmarssonand Dr. Bengt-Harald Jonsson for valuable discus-sions, Eleonore Skarfstad for technical assistance,¨and Per-Olov Freskgard for performing the CD-spec-˚tra measurements. This work was supported by grantsfrom Magnus Bergvalls Stiftelse and Stiftelsen LarsHiertas Minne.
References
w x Ž .1 Tibell, L., Aasa, R. and Marklund, S. 1993 Arch. Biochem.Biophys. 304, 429–443.
w x2 Tibell, L., Hjalmarsson, K., Edlund, T., Skogman, G., En-˚ Ž .gstrom, A. and Marklund, S.L. 1987 Proc. Natl. Acad. Sci.¨
USA 84, 6634–6638.w x3 Edlund, A., Edlund, T., Hjalmarsson, K., Marklund, S.L.,
Ž .Sandstrom, J., Stromqvist, M. and Tibell, L. 1992 Biochem.¨ ¨J. 288, 451–456.
w x Ž .4 Tibell, L.A.E., Skarfstad, E. and Jonsson, B.H. 1996¨Biochim. Biophys. Acta 1292, 47–52.
w x Ž .5 Kjellen, L. and Lindahl, U. 1991 Ann. Rev. Biochem. 60,443–465.
w x Ž .6 Marklund, S.L. 1982 Proc. Natl. Acad. Sci. USA. 79,7634–7638.
w x Ž .7 Klagsbrun, M. 1990 Curr.Opin. Cell. Biol. 2, 857–863.w x8 Inoue, M., Watanabe, N. Morino, Y., Tanaka,Y., Amachi,
Ž .T. and Sasaki, J. 1990 FEBS Lett. 269, 89–92.w x9 Sandstrom, J., Carlsson, L., Marklund, S.L. and Edlund, T.¨
Ž .1992 J. Biol. Chem. 267, 18205–18209.w x Ž .10 Carlsson, L.M., Marklund, L.S. and Edlund, T. 1996 Proc.
Natl. Acad. Sci. USA. 93, 5219–5222.w x11 Willems, J., Zwijsen. A., Slegers, H., Nicolas. S., Bettada-
Ž .pura, J., Raymackers, J. and Scarcez, T. 1993 J. Biol.Chem. 33, 24614–24621.
w x12 Nair, S.K., Calderone, T.L., Christianson, D.W. and Fierke,Ž .C.A. 1991 J. Biol. Chem. 266, 17320–17325.
w x Ž .13 Kunkel, T.A. 1985 Proc. Natl. Acad. Sci. USA. 82, 488–492.
w x Ž .14 Studier, F.W. and Moffat, B.A. 1986 J. Biol. Chem. 189,113–130.
w x Ž .15 Sanger, F., Nichlen, S. and Coulson, A.R. 1977 Proc. Natl.Acad. Sci. USA 74, 5463–5467.
w x16 Martensson, L.-G., Jonasson, P., Freskgard, P.-O., Svens-˚ ˚Ž .son, M., Carlsson, U. and Jonsson, B.-H. 1995 Biochem-
istry 34, 1011–1021.w x17 Khalifah, R.G., Strader, D.J., Bryant, S.H. and Gibson, S.M.
Ž .1977 Biochemistry 16, 2241–2247.w x18 Rickli, E.E., Ghazanfar, S.A.S., Gibbons, B.H and Edsall,
Ž .J.T. 1964 J. Biol. Chem. 239, 1065–1078.w x Ž .19 Gill, S.C. and von Hippel, P.H. 1989 Anal. Biochem. 182,
319–326.w x Ž .20 Karlsson, K., Lindahl, U. and Marklund, S.L. 1988
Biochem. J. 256, 29–33.w x21 Wishart, D.S., Bigam C.G., Yao, G., Abildgaard, F., Dyson,
Ž .H.J., Oldfield, E., Markley, J.L. and Sykes, B.D. 1995 J.Biomol. NMR 6, 135–140.
w x22 Jeener, J., Meier, B.H., Bachmann, P. and Ernst, R.R.Ž .1979 J. Chem. Phys. 71, 4546–4553.
w x Ž .23 Braunschweiler, L. and Ernst, R.R. 1983 J. Magn. Reson.53, 521–528.
w x Ž .24 Marion, D. and Wutrich, K. 1983 Biochem. Biophys. Res.¨Commun. 113, 967–974.
w x Ž .25 Piotto, M., Saudek, V. and Sklenar, V. 1992 J. Biomol.NMR, 2, 661–664.
w x Ž .26 Kay, L.E., Keifer, P. and Saarinen, T. 1992 J. Am. Chem.Soc. 114, 10663–10665.
w x Ž .27 Johnson, Jr., W.C. 1990 PROTEINS: Struct. Funct. Genet.7, 205–214.
w x Ž .28 Wishart, D.S., Sykes, B.D. and Richards, F.M. 1991 J.Mol. Biol. 222, 311–333.
w x Ž .29 Margalit, H., Fischer, N. and Ben-Sasson, S.A. 1993 J.Mol. Biol. 268, 19228–19231.
w x30 Sandstrom, J., Nilsson, P., Karlsson, K. and Marklund, S.L¨Ž .1994 J. Biol. Chem. 269, 19163–19166.
w x31 Adachi, T., Yamada, H., Yamada, Y., Morihara, N., Ya-mazaki, N., Murakami, T., Futenma, A., Kato, K. and
Ž .Hirano, K. 1996 Biochem. J. 313, 235–239.

( )L.A.E. Tibell et al.rBiochimica et Biophysica Acta 1340 1997 21–3232
w x32 Faham, S., Hileman, R.E., Fromm, J.R., Linhardt, R.J. andŽ .Rees, D.C. 1996 Science 271, 1116–1120.
w x33 Suzuki, S., Pierschbacher, M.D., Hayman, E.G., Nguyen,¨ Ž .K., Ohgren, Y. and Ruoslahti, E. 1984 J. Biol. Chem. 259,
15307–15314.w x34 Zucker, M.B., Katz, I.R., Thorbecke, J., Milton, D.C. and
Ž .Holt, J. 1989 Proc. Natl. Acad. Sci. USA, 86, 7571–7574.w x35 Webb, L.M., Ehrengruber, M.U., Clark-Lewis, I., Baggi-
Ž .olini, M. and Rot, A. 1993 Proc. Natl. Acad. Sci. USA 90,7158–7162.
w x36 Wun, T.C., Kretzmer, K.K., Girard, T.J., Miletich, J.P. andŽ .Broze, G.J. Jr. 1988 J. Biol. Chem. 263, 6001–6004.
w x37 Valentin, S., Larnkjaer, A., Oestergaard, P., Nielsen, J.I. andŽ .Nordfang, O. 1994 Thromb. Res. 75, 173–183.
w x38 Wesselschmidt, R., Likert, K., Girard, T., Wun, T.C. andŽ .Broze, G.J. 1992 Blood 79, 2004–2020.
w x39 Sprecher, C.A., Kisiel, W., Mathewes, S. and Foster, D.C.Ž .1994 Proc. Natl. Acad. Sci. USA 91, 3353–3357.
˚w x40 Hjalmarsson, K., Marklund, S.L., Engstrom, A. and Edlund,¨Ž .T. 1987 Proc. Natl. Acad. Sci. USA 84, 6340–6433.
w x Ž .41 Garnier, J., Osguthorpe, D.J. and Robson, B. 1978 J. Mol.Biol. 120, 97–120.