chloromethane, methyl donor in veratryl alcohol ... is a methyl donor in p. chrysosporium...
TRANSCRIPT

Vol. 56, No. 11APPLIED AND ENVIRONMENTAL MICROBIOLOGY, Nov. 1990, p. 3450-34570099-2240/90/113450-08$02.00/0Copyright © 1990, American Society for Microbiology
Chloromethane, Methyl Donor in Veratryl Alcohol Biosynthesis inPhanerochaete chrysosporium and Other Lignin-Degrading Fungi
DAVID B. HARPER,l12* JOHN A. BUSWELL,3t JAMES T. KENNEDY,2 AND JOHN T. G. HAMILTON2
Department ofFood and Agricultural Chemistry, The Queen's University of Belfast,l* and Food and AgriculturalChemistry Research Division, Department of Agriculture for Northern Ireland,2 Newforge Lane,
Belfast B79 5PX, Northern Ireland, and Department of Biology, Paisley Collegeof Technology, Paisley PA] 2BE, Scotland,3 United Kingdom
Received 13 June 1990/Accepted 27 August 1990
Chloromethane, a gaseous natural product implicated in methylation processes in PheUinus pomaceus, hasbeen shown to act as methyl donor in veratryl alcohol biosynthesis in the lignin-degrading fungi Phanerochaetechrysosporium, Phlebia radiata, and Coriolus versicolor, none of which released detectable amounts of CH3Clduring growth. When P. chrysosporium was grown in a medium containing C2H3Cl, levels of C2H3incorporation into the 3- and 4-0-methyl groups of veratryl alcohol were very high and initially similar to thoseobserved when the medium was supplemented with L-[methyl-2H3]methionine. When C2H3Cl was added tocultures actively synthesizing veratryl alcohol, incorporation of C2H3 was very rapid, with 81% of veratrylalcohol labeled after 12 h. By contrast, incorporation of C2H3 from L-[methyl-2H3]methionine was compara-tively slow, attaining 10% after 12 h. It is proposed that these lignin-degrading fungi possess a tightly channeledmultienzyme system in which CH3C1 biosynthesis is closely coupled to CH3Cl utilization for methylation ofveratryl alcohol precursors.
Many species of the family Hymenochaetaceae, a widelydistributed group of wood-rotting bracket fungi, release largequantities of gaseous CH3Cl during secondary metabolism(5, 14, 17, 18). Although CH3C1 has been shown to bederived from methionine (15, 35), no clear physiological rolehad been defined for it until Harper et al. (16) demonstratedrecently that it can act as the methyl donor for biosynthesisof methyl benzoate and methyl furoate during primarymetabolism of Phellinus pomaceus. The broad-specificitymethylating system can esterify a wide range of aromatic andaliphatic acids. Initially CH3Cl biosynthesis is closely cou-pled to utilization, but the system becomes less tightlychanneled in late trophophase, at which stage the release ofgaseous CH3Cl by the fungus begins. Hence, althoughCH3Cl ostensibly appears to be a characteristic secondarymetabolite, its liberation in the idiophase would seem merelyto reflect the breakdown of the strict coordination of biosyn-thesis and utilization of the compound which exists in theprimary metabolic phase.A CH3Cl-utilizing system capable of methylating many
phenols and thiophenol was also found in Phellinus poma-ceus (16), although the natural substrate for the system wasnot identified. The observation that C2H3Cl was incorpo-rated into methyl benzoate by a non-CH3Cl-releasing fun-gus, Fomitopsis pinicola (a member of the family Polypora-ceae), led Harper et al. (16) to speculate that CH3Clbiosynthesis and utilization as the methyl donor might not beconfined to members of the Hymenochaetaceae but couldexist more generally in nature.
Several white rot fungi (20, 22, 26) are known to producethe secondary metabolite veratryl alcohol (3,4-dimethoxy-benzyl alcohol), which is believed to be an important com-ponent of the lignin-degrading system. Veratryl alcohol
* Corresponding author.t Present address: Department of Biology, Chinese University of
Hong Kong, Shatin, New Territories, Hong Kong.
induces elements of the ligninolytic system (8, 9, 25), pre-vents inactivation of lignin peroxidase (13, 32, 33), and isreported to enhance the oxidation of lignin by lignin perox-idase in vitro (12). It may also function as a natural one-electron redox mediator in the depolymerization process,possibly promoting oxidation by creation of activated oxy-gen species (19, 28). The key roles assigned to veratrylalcohol in lignin biodegradation render the biosyntheticroute of the compound and its regulation of critical impor-tance. Biosynthesis of veratryl alcohol in Phanerochaetechrysosporium proceeds via phenylalanine, 3,4-dimethoxy-cinnamyl alcohol, and veratryl glycerol; trapping experi-ments indicate that the 0-methyl groups are derived frommethionine (29). 4-0-Methylation of lignin degradation prod-ucts such as vanillic acid by ligninolytic cultures of thisfungus with methionine as the methyl donor has also beenreported (4).
In the present study the labeling of veratryl alcoholproduced during culture of P. chrysosporium and two otherlignin-degrading fungi was investigated in the presence ofC2H3Cl and, in some instances, L-[methyl-2H3]methionine todetermine whether CH3Cl served as an intermediate in 3-and 4-0-methylation during the synthesis of veratryl alcohol.
MATERIALS AND METHODSOrganisms and maintenance media. P. chrysosporium
Burds INA-12 (CNCM 1-398) was a strain used in investiga-tions by Buswell et al. (3). Coriolus versicolor Quel. Ps 4awas acquired from S. Kawai, Wood Research Institute,Kyoto University, Uji, Kyoto, Japan. Phlebia radiata Fr. 79(ATCC 64658) was also used in some experiments. Fungiwere maintained on 4% (wt/vol) malt extract agar (OxoidLtd., London, United Kingdom).
Chemicals. C2H3Cl (99.9 atom% 2H), C2H31 (99.5 atom%2H), and L-[methyl-2H3]methionine (99.4 atom% 2H) werepurchased from MSD Isotopes, Montreal, Quebec, Canada.C2H302H (99.5 atom% 2H), (C2H3)2SO4 (99 atom% 2H),veratryl alcohol, 3,4,5-trimethoxybenzyl alcohol, and other
3450
on March 30, 2019 by guest
http://aem.asm
.org/D
ownloaded from

CHLOROMETHANE IS A METHYL DONOR IN P. CHRYSOSPORIUM
substituted benzyl alcohols and benzaldehydes were ob-tained from Aldrich Chemical Co., Gillingham, Dorset,United Kingdom. Caffeic, ferulic, isoferulic, and 3,4-dimeth-oxycinnamic acids were also acquired from Aldrich. N-Methyl-N-trimethylsilyltrifluoroacetamide reagent (MSTFA)was obtained from Pierce Chemical Co., Rockford, Ill.
[4-methoxy-2H3]veratryl alcohol (bp 166°C at 11 mm Hg[1,467 Pa]) and [3-methoxy-2H3]veratryl alcohol (bp 164 to165°C at 14 mm Hg [1,867 Pa] were prepared by methylationof vanillin and isovanillin in aqueous NaOH with (C2H3)2SO4by standard procedures followed by reduction of the result-ing aldehydes in methanol with hydrogen in the presence ofRaney nickel at room temperature and pressure. [3,4-dimethoxy-2H6]veratryl alcohol (bp 165°C at 11 mm Hg[1,467 Pa]) was prepared by methylation of 3,4-dihydroxy-benzaldehyde in dimethyl sulfoxide with (C2H3)2SO4 (11)and reduction of the aldehyde with hydrogen in the presenceof Raney nickel.
Culture media and culture conditions. P. chrysosporiumwas grown without agitation at 37°C in 100-ml conical flasksin 10 ml of medium which contained the following (in gramsper liter): KH2PO4, 0.2; MgSO4 .7H20, 0.05; NaCl, 0.59;CaCl2 .2H20, 0.013; thiamine, 0.0025; yeast extract (Difco),0.1; glycerol, 10; L-asparagine monohydrate, 1.0; NH4NO3,0.5; and 2,2-dimethylsuccinic acid, 1.46. It also contained 1ml of trace elements solution per liter (1). The medium wasadjusted to pH 5.0 with 2 M KOH, filter sterilized, andinoculated with approximately 5 x 105 conidia per literprepared as described by Eriksson and Johnsrud (7). Flaskswere flushed with 100% 02 for 2 min before being stopperedwith a polytetrafluoroethylene (PTFE)-coated rubber stop-per. In some experiments the growth medium was supple-mented with 1 mM L-[methyl-2H3]methionine (i.e., 10 ,umolflask-1). When the growth medium required supplementa-tion with C2H3Cl, the following technique was used. Fromthe PTFE-coated rubber stopper was suspended an alumi-num clamp in which a Durham tube was fitted. Immediatelybefore sealing the flasks, 30 mM aqueous C2H3Cl (1.67 ml)was pipetted into the tube to give a concentration of 0.77 mMC2H3Cl in the culture medium after equilibration of gaseousand aqueous phases (i.e., 50 iLmol of C2H3Cl flask-') (17).When the incorporation of precursors added to cultures
after 72 h of growth was investigated, a different procedurewas adopted. The PTFE-coated rubber stopper of each flaskwas fitted with two stoppered glass injection ports, one ofwhich consisted of a capillary tube extending to within 2 mmof the base of the flask. After 72 h of growth the medium wassupplemented with either 1mM L-[methyl-2H3]methionine or0.77mM C2H3Cl by injection through the capillary injectionport of 10 mM methionine (1.1 ml) or 30 mM C2H3Cl (1.67ml), respectively. This technique was also used for supple-mentation of the culture medium with additional methionine(0.5 mM) after 96 h of incubation.
Phlebia radiata was grown under similar conditions to P.chrysosporium, except that a temperature of 29°C was usedand the medium contained the following (in grams per liter):KH2PO4, 0.2; MgSO4 7H2O, 0.5; CaCl2 2H20, 0.1;NH4NO3, 0.039; L-asparagine, 0.069; glucose, 10; and 2,2-dimethylsuccinic acid, 1.46. It also contained 1 ml of traceelements solution per liter (21) and 1 ml of vitamin solutionper liter (23). The medium was adjusted to pH 4.5 with 2 MKOH, filter sterilized, and inoculated with a mycelial sus-pension (1 ml) prepared by manually shaking 0.25 g ofmycelium with 10 ml of sterile water and 5 g of glass beads(diameter, 6 mm) in a 40-ml vial. Flasks were flushed with100% 02 for 2 min before being stoppered with a PTFE-
coated rubber stopper suspended from which, as describedabove, was a Durham tube containing a volume of aqueousC2H3CI such as to give a concentration of 0.77 mM C2H3Clin the medium. After 7 days, cultures were harvested,veratryl alcohol in the supernatant was assayed, and thepercent C2H3 incorporation into the compound was deter-mined.
C. versicolor was grown at 29°C under 02 as outlined forP. chrysosporium, except that the culture medium containedthe following (in grams per liter): KH2PO4, 0.2; MgSO47H20, 0.05; CaCI2, 0.01; NH4NO3, 0.052; L-asparagine,0.086; glucose, 10; and polyacrylic acid (0.01 M in carboxyl),0.72. It also contained 1 ml of mineral solution per liter (23)and 0.5 ml of vitamin solution per liter (23). The medium wasadjusted to pH 4.5, filter sterilized, and inoculated with amycelial suspension (1 ml) prepared as described for Phlebiaradiata. Flasks were flushed with 100% 02 and stopperedwith a PTFE-coated rubber stopper equipped with a C2H3Cl-containing Durham tube giving a concentration of 0.77 mMC2H3C1 in the medium. After 7 days, cultures were har-vested, veratryl alcohol in the supernatant was assayed, andthe percent C2H3 incorporation into the compound wasdetermined.
Lignin peroxidase assay. The activity of lignin peroxidasein culture supernatants was determined by measuring theincrease in A310 due to the oxidation of veratryl alcohol toveratraldehyde in the presence of H202 (31). In the assay,supernatant (1 ml) was incubated with 4 mM veratryl alcoholand 0.27 mM H202 in 60 mM sodium tartrate buffer pH 3.0at 37°C. Activities are expressed as nanomoles of veratral-dehyde formed per minute per milliliter of culture superna-tant.
Assay of veratryl alcohol and determination of percent C21H3incorporation. Culture supernatant (10 ml), after addition of3,4,5-trimethoxbenzyl alcohol (0.2 ml of a 500-,ug/ml solutionin acetone) as an internal standard, was extracted threetimes with chloroform (15 ml per extraction), and the bulkedextract was washed with saturated NaCl solution (10 ml).The extract was dried over anhydrous MgSO4 and evapo-rated to 0.5 ml under reduced pressure. Finally, the solutionwas made up to 5 ml with diethyl ether.
Veratryl alcohol in the extract was determined by gaschromatography-mass spectrometry (GC-MS) on a Hewlett-Packard 5890 gas chromatograph linked to an HP5970 mass-selective detector controlled by an HP300 series computer.The gas chromatograph was equipped with an Ultra 1fused-silica wall-coated open tubular capillary column (25 mby 0.2 mm) with 100% dimethylpolysiloxane as the bondedphase. Helium was used as the carrier gas at a flow rate of1.5 ml min-1, and a split ratio of 20:1 was used. Afterinjection the oven temperature was held at 100°C for 1 minand then programmed at 10°C min-1 up to 160°C followed by30°C min-1 up to 300°C. To quantify veratryl alcohol, ioncurrents at mle 168, 171, and 174 were monitored and theirsum at the retention time of veratryl alcohol was comparedwith that given by an authentic sample of the compound. Theincorporation of C2H3 into the 3- and 4-0-methyl groups ofveratryl alcohol was measured as the relative proportions ofthe three ions after correction for a small proportion of (M -3) ion (approximate intensity, 5%) present in spectra of bothlabeled and unlabeled compounds. As it was not possible onthe basis of the MS fragmentation pattern to distinguishbetween veratryl alcohol monosubstituted with C2H3 in the3-position from that monosubstituted in the 4-position, ver-atryl alcohol with a single C2H3 substitutent is referred tosimply as monolabeled. Thus, the compound giving an ion of
VOL. 56, 1990 3451
on March 30, 2019 by guest
http://aem.asm
.org/D
ownloaded from

APPL. ENVIRON. MICROBIOL.
mle 168 (substitution OCH3, OCH3) is referred to henceforthas unlabeled compound, that giving an ion of mle 171(substitution OCH3, OC2H3) is referred to as monolabeled,and that giving an ion of mle 174 (substitution OC2H3,OC2H3) is referred to as dilabeled.
In experiments on utilization of C2H3Cl and L-[methyl-2H3]methionine during growth of P. chrysosporium, valuesgiven are the mean of two replicates. In experiments involv-ing the addition of labeled precursor to actively growingcultures of P. chrysosporium, the percent incorporations areexpressed as the mean + standard deviation of three to fivereplicates. Results given for Phlebia radiata are the mean offive replicates. Veratryl alcohol in C. versicolor was deter-mined in the bulked supernatants of five replicate cultures.
Determination of nonenzymatic methylation. The extent ofpossible nonenzymatic methylation of phenolic precursorsof veratryl alcohol was investigated by the following proce-dure. Phenolic substrate (0.5 mM) was incubated in sterileculture medium under conditions identical to those of fungalculture at 37°C in the presence of either 1 mM C2H3Cl or 0.25mM C2H31. After a 7-day incubation a solution of thepossible methylated product in unlabeled form was added toeach flask to give a final concentration of 5 puM. The culturemedium was then acidified to pH 3.0 and extracted withchloroform, and the extract was reduced to a small volumeas described above.
For aldehydes and alcohols the solution was made up to 1ml in ether and examined by GC-MS under the conditionsdescribed above for the veratryl alcohol assay. Ion currentsat mle values corresponding to the molecular ions (M) ofpossible methylated products, the M + 3 ions, and in somecases the M + 6 ions were monitored. The ratios of the lasttwo ions to the molecular ion at the retention time of themethylated product were a measure of the nonbiologicalincorporation of C2H3 from C2H3Cl into the monomethy-lated and dimethylated products, respectively.For the substituted cinnamic acids the solution was taken
to dryness and the residue was silylated with 1 ml ofMSTFA. The solution was examined by using GC-MS as
above, except that the gas chromatograph was equippedwith an Ultra 2 fused-silica wall-coated open tubular column(12 m by 0.2 mm) with 95% dimethylpolysiloxane-5% diphe-nyl as the bonded phase. Helium was used as the carrier gas
at a flow rate of 1 ml min-', and a split ratio of 20:1 was
employed. After injection the oven temperature was held at100°C for 1 min and then programmed at 20°C min-' up to300°C and held at this temperature for 5 min. Ion currents atmle values corresponding to the molecular ion (M) and the M+ 3 and M + 6 ions were monitored on a Hewlett-Packard5970 mass-selective detector, and the ratios of the last twoions relative to the molecular ion were again used as a
measure of nonenzymatic methylation.The following conversions were investigated by this tech-
nique: caffeic acid to ferulic, isoferulic, and 3,4-dimethoxy-cinnamic acids; ferulic acid and isoferulic acid to 3,4-dimethoxycinnamic acid; 3,4-dihydroxybenzaldehyde tovanillin, isovanillin, and veratraldehyde; vanillin and isovan-illin to veratraldehyde; and 3-hydroxy-4-methoxybenzyl al-cohol and 3-methoxy-4-hydroxybenzyl alcohol to veratrylalcohol.NMR. 'H nuclear magnetic resonance (NMR) spectra of
C2H3-labeled veratryl alcohols were recorded in C2HC13 at
20°C with tetramethylsilane as the internal standard on a GEOmega 500-MHz NMR spectrometer, with 16,384 datumpoints and a sweep width of 5000 Hz.
Assessment of growth. Mycelial growth was measured as
the weight of mycelia harvested by filtration at variousstages of incubation and dried at 100°C.CH3C1 assay. CH3Cl was determined by GC, using the
headspace technique described by Harper and Kennedy(17). A water/air partition coefficient (wt/vol per wt/vol) of2.21 for CH3C1 at 37°C was used in calculations. Forconfirmation of the presence of CH3Cl in the headspace offungal cultures at concentrations down to 0.2 ng ml-', theGC-MS technique described by Harper et al. (18) was used.
Methionine analysis. Culture medium was filtered througha 0.22-,um membrane filter (Millipore Corp.), and filtrate (8ml) was evaporated to dryness on a Hetovac VRI vacuumconcentrator fitted with a CT 60 cooling trap. The residuewas dissolved in 5 ml of lithium citrate buffer (pH 2.2)containing 2 mM norleucine as internal standard. The sample(15 [lI) was then chromatographed on an LKB model 4400amino acid analyzer by using a lithium citrate buffer elutionprogram (2). The concentration of methionine was calculatedfrom the peak areas by using an internal standard procedure,a calibration standard being chromatographed after everyfive samples.
RESULTS AND DISCUSSION
C2H3 incorporation into veratryl alcohol from L-[methyl-2H3]methionine in growth medium. Figure 1A shows theincorporation of C2H3 into veratryl alcohol at various stagesduring growth of P. chrysosporium on a medium containing1 mM L-[methyl-2H3]methionine; Fig. 2 records the veratrylalcohol concentration in the culture medium and mycelialdry weight during this period. In the early stages of growththe 0-methyl groups in the 3 and 4 positions of veratrylalcohol were almost wholly labeled with C2H3, 83% of thecompound present being dilabeled and 17% monolabeled.This finding confirms the conclusion of Shimada et al. (29)that the 0-methyl groups of veratryl alcohol originate frommethionine. Between 96 and 120 h the level of labeling fellsharply, presumably reflecting exhaustion of labeled methi-onine in the culture medium and the utilization of endoge-nous unlabeled methionine for methylation. The rapidity ofthis decline implies a high rate of veratryl alcohol turnover inthe culture medium as previously observed (24). This isconsistent with the key metabolic role proposed for thecompound. That the decrease in labeling is attributable tothe depletion of labeled methionine in the culture medium isclearly demonstrated by an experiment in which the culturemedium of the fungus grown as in Fig. IA was supplementedwith additional L-[methyl-2H3]methionine (0.5 mM) after 96h. Under these conditions the level of labeling in veratrylalcohol quickly recovered to its original high value, whichwas maintained until the experiment was terminated at 209 h(Fig. 1B).C2H3 incorporation into veratryl alcohol from C2H3Cl in
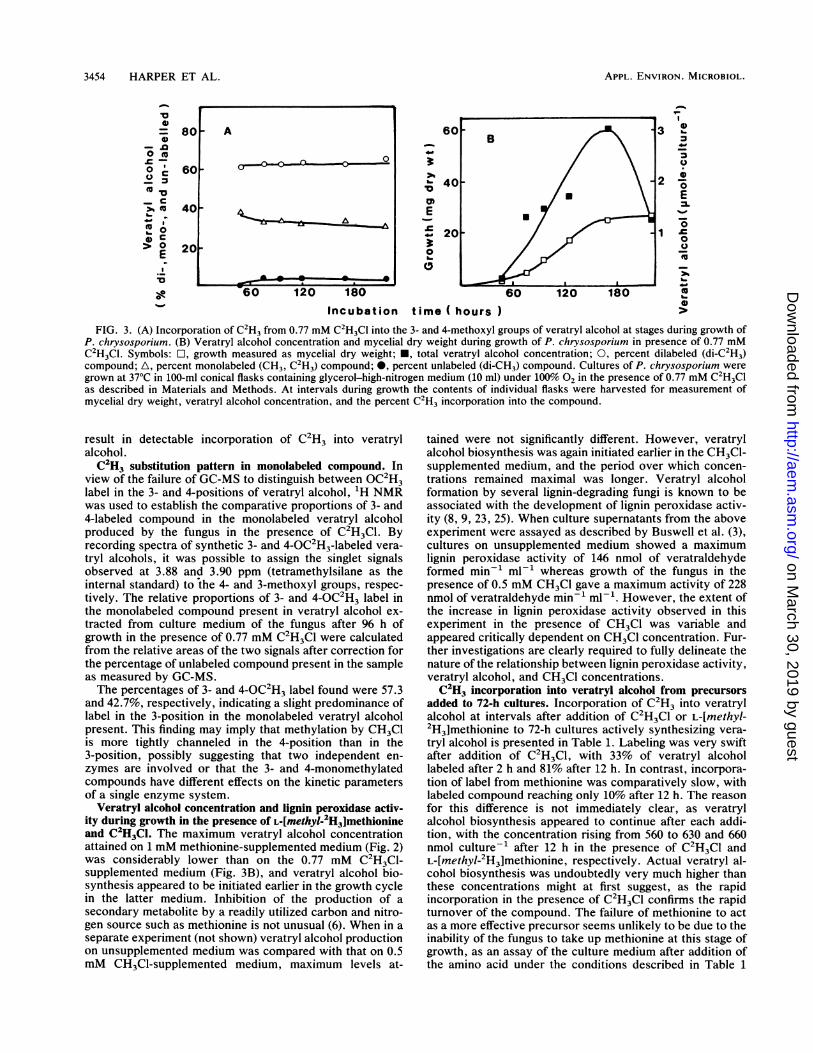
growth medium. Figure 3A illustrates the labeling patternfound in veratryl alcohol present at various stages duringgrowth of the fungus in medium containing 0.77 mM C2H3Cl,whereas Fig. 3B shows the veratryl alcohol concentration inthe culture medium and mycelial dry weight during theexperiment. High levels of incorporation of C2H3 into vera-tryl alcohol were observed throughout the whole incubationperiod. CH3Cl is clearly as effective a precursor as methio-nine for 3- and 4-0-methylation. Maintenance of high levelsof labeling throughout incubation can be attributed to theequilibrium existing between C2H3Cl in the flask headspace(initially 41 ,umol) and that in the aqueous phase (initially 9,umol), which ensures that C2H3C1 in the medium utilized in
3452 HARPER ET AL.
on March 30, 2019 by guest
http://aem.asm
.org/D
ownloaded from

CHLOROMETHANE IS A METHYL DONOR IN P. CHRYSOSPORIUM
.0,
- w
0.
0
80 B 4,
60
40
20 -
. I180 60 120 180
Incubation time (hours )FIG. 1. Incorporation of C2H3 from L-[methyl-2H3]methionine into the 3- and 4-methoxyl groups of veratryl alcohol during growth of P.
chrysosporium. (A) Culture medium contained an initial concentration of 1 mM L-[methyl-2H3]methionine; (B) conditions were similar topanel A, except that a further supplement of L-[methyl-2H3]methionine (0.5 mM) was added at 96 h. Symbols: 0, percent dilabeled (di-C2H3)veratryl alcohol; A, percent monolabeled (CH3, C2H3) veratryl alcohol; 0, percent unlabeled (di-CH3) veratryl alcohol. Cultures of P.chrysosporium were grown at 37°C in 100-ml conical flasks containing glycerol-high-nitrogen medium (10 ml) under 100% 02 in the presenceof 1 mM L-[methyl-2H3]methionine as described in Materials and Methods. In panel B, flasks were supplemented with additionalL-[methyl-2H3]methionine at 96 h (arrowed). At intervals during growth, the contents of individual flasks were harvested for measurement ofmycelial dry weight, veratryl alcohol concentration, and percent C2H3 incorporation into the compound.
metabolism is largely replaced from the gaseous phase.Therefore, at the termination of the experiment at 216 h, theC2H3Cl concentration in the medium was still 0.16 mM.
Unequivocal confirmation that label was incorporated intothe methoxyl groups of veratryl alcohol was obtained bychemical synthesis of the various possible methoxyl-labeledveratryl alcohols and comparison of the mass spectrum of anappropriate mixture of these compounds with that of theveratryl alcohol synthesized by the fungus in the presence ofC2H3Cl (Fig. 4). The mass spectra of unlabeled and dilabeledcompounds (Fig. 4A and B, respectively) are quite different,and both are distinct from the two monolabeled compounds(Fig. 4C and D). However, the latter compounds givepractically identical mass spectra and therefore cannot bedistinguished by the technique. Figure 4E is the massspectrum of veratryl alcohol extracted from culture mediumafter growth of P. chrysosporium for 168 h in the presence of0.77 mM C2H3Cl. On the basis of the relative intensity of themolecular ions at mle 168, 171, and 174 (after correction for
0la
0
0
Incubation time (hours )
.0
-
2
0
0
E
0
0
b-
FIG. 2. Veratryl alcohol concentration during growth of P. chry-sosporium in the presence of 1 mM L-[methyl-2H3]methionine.Symbols: O, growth measured as mycelial dry weight; *, totalveratryl alcohol concentration. Cultures of P. chrysosporium weregrown as described for Fig. 1A.
the M - 3 ion as described in Materials and Methods), it wasconcluded that the ratio of dilabeled to monolabeled tounlabeled compound was 69:27:4. Figure 4F is the massspectrum of a mixture of compounds from Fig. 4B, C, and Ain the above proportion and is virtually indistinguishablefrom the fungal product, demonstrating that C2H3 is incor-porated wholly into the methoxyl groups of veratryl alcoholand validating the ion-monitoring technique used to measurethe proportions of the various labeled species.That methylation was not due to a nonenzymatic reaction
of C2H3CI with a veratryl alcohol precursor produced by thefungus was demonstrated by the isotopic dilution techniquedescribed in Materials and Methods. None of the possibleproducts of methylation were found at concentrations abovethe detection limit of 0.25 ,uM when 0.5 mM caffeic andferulic acids (the precursors postulated by Shimada et al.[29]) and isoferulic acid were incubated for 7 days in sterileculture medium at 37°C with 1 mM C2H3Cl. Additionally,when 0.5 mM 3,4-dihydroxybenzaldehyde, vanillin, isovan-illin, 3-methoxy-4-hydroxybenzyl alcohol, and 3-hydroxy-4-methoxybenzyl alcohol were incubated under similar condi-tions, no evidence of formation of methylated product wasfound at concentrations above the detection limit of 0.1 ,uM.Even when the more potent chemical methylating agentiodomethane was incubated at a concentration of 0.25 mMunder similar conditions with 0.5 mM solutions of the abovesubstrates, no methylated products were observed at con-centrations above the detection limit of 0.25 ,uM. Thesefindings are not unexpected in view of the very weaklynucleophilic nature of unionized phenols. As veratryl alco-hol was typically present at 0.2 mM in the culture medium ofthe fungus, it is clear that the substantial labeling obtained inthe veratryl alcohol biosynthesized in the presence ofC2H3Cl is not explicable in terms of nonenzymatic methyl-ation.The possibility that methylation proceeds by hydrolytic
dehalogenation of CH3Cl to methanol followed by utilizationof methanol as the methyl donor, a route which was elimi-nated in investigations with Phellinus pomaceus (16), is alsorendered highly improbable in P. chrysosporium since addi-tion of 0.5 mM C2H3OH to the growth medium failed to
VOL. 56, 1990 3453
on March 30, 2019 by guest
http://aem.asm
.org/D
ownloaded from
APPL. ENVIRON. MICROBIOL.
80 A
0
60 0
40 4
20
"60 120 180Incubation
I 40
E
.0 20
0
time ( hours )
FIG. 3. (A) Incorporation of C2H3 from 0.77 mM C2H3C1 into the 3- and 4-methoxyl groups of veratryl alcohol at stages during growth ofP. chrysosporium. (B) Veratryl alcohol concentration and mycelial dry weight during growth of P. chrysosporium in presence of 0.77 mMC2H3Cl. Symbols: O, growth measured as mycelial dry weight; *, total veratryl alcohol concentration; 0, percent dilabeled (di-C2H3)compound; A, percent monolabeled (CH3, C2H3) compound; *, percent unlabeled (di-CH3) compound. Cultures of P. chrysosporium weregrown at 37°C in 100-ml conical flasks containing glycerol-high-nitrogen medium (10 ml) under 100% 02 in the presence of 0.77 mM C2H3Clas described in Materials and Methods. At intervals during growth the contents of individual flasks were harvested for measurement ofmycelial dry weight, veratryl alcohol concentration, and the percent C2H3 incorporation into the compound.
result in detectable incorporation of C2H3 into veratrylalcohol.C2H3 substitution pattern in monolabeled compound. In
view of the failure of GC-MS to distinguish between OC2H3label in the 3- and 4-positions of veratryl alcohol, 'H NMRwas used to establish the comparative proportions of 3- and4-labeled compound in the monolabeled veratryl alcoholproduced by the fungus in the presence of C2H3Cl. Byrecording spectra of synthetic 3- and 4-OC2H3-labeled vera-
tryl alcohols, it was possible to assign the singlet signalsobserved at 3.88 and 3.90 ppm (tetramethylsilane as theinternal standard) to the 4- and 3-methoxyl groups, respec-tively. The relative proportions of 3- and 4-OC2H3 label inthe monolabeled compound present in veratryl alcohol ex-tracted from culture medium of the fungus after 96 h ofgrowth in the presence of 0.77 mM C2H3Cl were calculatedfrom the relative areas of the two signals after correction forthe percentage of unlabeled compound present in the sampleas measured by GC-MS.The percentages of 3- and 4-OC2H3 label found were 57.3
and 42.7%, respectively, indicating a slight predominance oflabel in the 3-position in the monolabeled veratryl alcoholpresent. This finding may imply that methylation by CH3Clis more tightly channeled in the 4-position than in the3-position, possibly suggesting that two independent en-zymes are involved or that the 3- and 4-monomethylatedcompounds have different effects on the kinetic parametersof a single enzyme system.
Veratryl alcohol concentration and lignin peroxidase activ-ity during growth in the presence of L-[methyl-2H3]methionineand C2H3Cl. The maximum veratryl alcohol concentrationattained on 1 mM methionine-supplemented medium (Fig. 2)was considerably lower than on the 0.77 mM C2H3Cl-supplemented medium (Fig. 3B), and veratryl alcohol bio-synthesis appeared to be initiated earlier in the growth cyclein the latter medium. Inhibition of the production of a
secondary metabolite by a readily utilized carbon and nitro-gen source such as methionine is not unusual (6). When in aseparate experiment (not shown) veratryl alcohol productionon unsupplemented medium was compared with that on 0.5mM CH3Cl-supplemented medium, maximum levels at-
tained were not significantly different. However, veratrylalcohol biosynthesis was again initiated earlier in the CH3Cl-supplemented medium, and the period over which concen-trations remained maximal was longer. Veratryl alcoholformation by several lignin-degrading fungi is known to beassociated with the development of lignin peroxidase activ-ity (8, 9, 23, 25). When culture supernatants from the aboveexperiment were assayed as described by Buswell et al. (3),cultures on unsupplemented medium showed a maximumlignin peroxidase activity of 146 nmol of veratraldehydeformed min-' ml-' whereas growth of the fungus in thepresence of 0.5 mM CH3Cl gave a maximum activity of 228nmol of veratraldehyde min-' ml-'. However, the extent ofthe increase in lignin peroxidase activity observed in thisexperiment in the presence of CH3Cl was variable andappeared critically dependent on CH3Cl concentration. Fur-ther investigations are clearly required to fully delineate thenature of the relationship between lignin peroxidase activity,veratryl alcohol, and CH3Cl concentrations.C2H3 incorporation into veratryl alcohol from precursors
added to 72-h cultures. Incorporation of C2H3 into veratrylalcohol at intervals after addition of C2H3Cl or L-[methyl-2H3]methionine to 72-h cultures actively synthesizing vera-tryl alcohol is presented in Table 1. Labeling was very swiftafter addition of C2H3Cl, with 33% of veratryl alcohollabeled after 2 h and 81% after 12 h. In contrast, incorpora-tion of label from methionine was comparatively slow, withlabeled compound reaching only 10% after 12 h. The reasonfor this difference is not immediately clear, as veratrylalcohol biosynthesis appeared to continue after each addi-tion, with the concentration rising from 560 to 630 and 660nmol culture-' after 12 h in the presence of C2H3Cl andL-[methyl-2H3]methionine, respectively. Actual veratryl al-cohol biosynthesis was undoubtedly very much higher thanthese concentrations might at first suggest, as the rapidincorporation in the presence of C2H3Cl confirms the rapidturnover of the compound. The failure of methionine to actas a more effective precursor seems unlikely to be due to theinability of the fungus to take up methionine at this stage ofgrowth, as an assay of the culture medium after addition ofthe amino acid under the conditions described in Table 1
0
0- .00 X-
-c_ c
."M
e> 0
E
:6._e
w-
I
0
aU
00E
'a.
-50
0
3454 HARPER ET AL.
on March 30, 2019 by guest
http://aem.asm
.org/D
ownloaded from

CHLOROMETHANE IS A METHYL DONOR IN P. CHRYSOSPORIUM
A
Ill..11
v,'..n3
60t, i6 1,1,, .11.1.....111 .101..1.
CH20H
JOC2H3OCH3
.1 . .-I,. ,1,1,.1,,,..11..1,,.,.1.11 ...
60
CH20H
L O C H3
I I
ISO
OC'H3
Ill.,. x,I Ill,,I IllIll, ...140JII. ,.1 1 'II,
E
--Lllli * 11111- t^f^E| iE l*iXaek kElal llisEfi _4la11-11 . ^0svl-Ill .1111 lffil slr
60 100 140 180
.-III.,.,ll,1ll.,1l.,.,1,lll---I l .,,1 A1ll,,1|1i,
D
60 10o 140
Mass/Charge180
FIG. 4. MS comparison of veratryl alcohol extracted from culture medium of P. chrysosporium grown in the presence of C2H3Cl withauthentic C2H3-labeled and unlabeled standards. (A to D) Mass spectra of various labeled and unlabeled standards of veratryl alcohol. (E)Mass spectrum of veratryl alcohol extracted from fungal culture medium after growth of the fungus for 168 h in presence of 0.77 mM C2H3Cl.(F) Mass spectrum of a mixture of B, C, and A in the proportion 69:27:4.
showed a 29% fall in concentration within 12 h. Therefore,the most feasible explanation of the comparatively lowincorporation of methionine observed in Table 1 is thatexogenous amino acid taken up at this stage of growth isrouted predominantly into competing biochemical pathways,
in particular, protein biosynthesis. These results, whenconsidered in association with those displayed in Fig. 3,provide compelling evidence that CH3Cl must participate as
an intermediate in the methylation of veratryl alcohol pre-cursors.
CH20H
OCH3
100-
60-
20-
100-
60-
20-
.11180100
I ,060
CH20H
1JOC2H3r% 2|4_
B
'Ii''
100-
60-
20-
100-
60-
20-
a0)c0
._
0
0
C F
60
100-
60-
20-
-Ai180
100
60
20
VOL. 56, 1990 3455
140
6!0 100 140 180-
||100 140
100 1i0
on March 30, 2019 by guest
http://aem.asm
.org/D
ownloaded from

APPL. ENVIRON. MICROBIOL.
TABLE 1. Incorporation of C2H3 into veratryl alcohol at intervals after addition of C2H3Cl or L-[methyl-2H3]methionineto 72-h cultures' of P. chrysosporium
Time (h) after % C2H3 substitutionb in 3- and 4-positions of veratryl alcohol ± SDaddition of C2H3CI (0.77 mM) L-[methyl-2H3Jmethionine (1 mM)labeled _____________________________________________________
precursor Di-CH3 CH3, C2H3 Di-C2H3 Di-CH3 CH3, C2H3 Di-C2H3
0 100 0 0 100 0 02 67 ± 14 16 7 17 ± 6 97 5 1 2 1 ± 24 47 ± 8 20 3 33 ± 5 96 6 2 3 2 ± 38 26 ± 11 20 2 54 ± 9 93 10 3 4 4 ± 6
12 19 ± 9 20 4 61 ± 7 89 5 3 2 7 ± 3a Cultures were grown as described in Materials and Methods for 72 h on unsupplemented glycerol-high-nitrogen medium.b Di-CH3, unlabeled compound; CH3, C2H3, monolabeled compound; Di-C2H3, dilabeled compound.
C213 incorporation into veratryl alcohol from C2H3CI inother white rot fungi. Two other white rot fungi, Phlebiaradiata and Coriolus versicolor, both widely used in inves-tigations of lignin degradation and reported to produceveratryl alcohol (20, 22), were also examined for the pres-ence of a CH3Cl-utilizing methylation system. Veratrylalcohol produced at a mean concentration of 50 nmolculture-1 after 7 days of growth of Phlebia radiata in thepresence of 0.77 mM C2H3Cl as described in Materials andMethods gave a mean ratio of dilabeled to monolabeled tounlabeled compound of 82:17:1. Growth of C. versicolor for7 days in the presence of 0.77 mM C2H3Cl as described inMaterials and Methods resulted in a veratryl alcohol con-centration of 4 nmol culture-1 with a ratio of dilabeled tomonolabeled to unlabeled compound of 45:41:14. Thesefindings demonstrate that a CH3Cl-utilizing methylation sys-tem is also operative in both these fungi, albeit rather moretightly channeled in C. versicolor than in P. chrysosporiumor Phlebia radiata.CH3C1 release by fungal cultures. Despite this evidence for
CH3Cl as a metabolic intermediate in all three speciesinvestigated, the release of CH3Cl as a natural product intothe flask headspace could not be detected during the growthof any of the fungi at concentrations above 0.2 ng ml-',which represents the limit of detection of the GC-MS tech-nique used. Similarly, screening of the three species forCH3Cl release on three different solid media, containingglucose, malt extract, and cellulose, respectively, as themain carbon source by the methods of Harper et al. (18)failed to demonstrate the presence of free CH3Cl in theculture headspace. This suggests that, as with the methyla-tion of benzoic acid by CH3Cl in Fomitopsis pinicola (15),biosynthesis of CH3Cl in these species is closely coupled toits utilization by a high-affinity methylation system. Suchchanneling could involve direct transfer of CH3Cl from thebiosynthetic enzyme to the methylation system or, alterna-tively, could entail restriction of CH3Cl to an unmixed layerwithin the cell (10). In either case, the compound is effec-tively confined to a multienzyme complex and never attainsthe status of a freely diffusible intermediate. Nevertheless,the fact that exogenous CH3Cl can act as the methyl donorindicates that access to the methylating enzyme is notrestricted solely to endogenous CH3Cl.An alternative explanation for the failure to detect CH3Cl
as a natural product of these fungi is that rather than being agenuine metabolic iptermediate, the compound is involved inexchange reactions with the true substrate of the methyl-transferase enzyme, which could be either S-adenosylme-thionine or an enzyme-bound activated methyl group. How-ever, to achieve the high labeling levels exhibited in Fig. 3and the speed of incorporation apparent in Table 1, exchange
would have to be very rapid, and this seems exceedinglyunlikely on thermodynamic grounds.
Significance of CH3Cl in fungal metabolism. Participationof CH3Cl in the biosynthesis of a compound of such centralimportance in lignin degradation affords strong support forthe suggestion by Harper et al. (16) that CH3Cl may have awidespread, if previously unsuspected, role in methylationprocesses in many fungi and even perhaps higher plants. Thepart played by the compound as methyl donor may haveescaped recognition hitherto because CH3Cl is a relativelychemically unreactive gas that is readily overlooked bynormal analytical procedures and is usually synthesized andutilized in tightly channeled multienzyme complexes fromwhich the release of free CH3Cl is seldom or never observed.Although this report considers only the role of CH3C1 in
methylation of veratryl alcohol, the presence of CH3Cl-utilizing systems in lignin-degrading fungi may have a muchwider significance. Although ligninases are able to degradelignin in vitro to a limited extent (30), Haemmerli et al. (12)have shown that repolymerization outweighs degradation,probably owing to the phenoloxidizing activity associatedwith the enzyme. Accordingly, they and others postulatethat, inter alia, systems must exist in vivo to preventpolymerization (12, 34). It is conceivable that these includean extracellular methylation system that uses CH3C1, whichcould provide the rapid methylating capability necessary tohalt repolymerization of phenolic cleavage products of lig-ninase action. A sufficiently high-affinity enzyme might evenbe able to use natural CH3Cl normally present at a concen-tration of about 1 ppb (vol/vol) in the atmosphere (27).
ACKNOWLEDGMENTS
We thank Don Annette for methionine analyses and Paul Steven-son, Chemistry Department, The Queen's University of Belfast, forNMR analyses.
LITERATURE CITED1. Ander, P., and K.-E. Eriksson. 1976. The importance of phenol
oxidase activity in lignin degradation by the white rot fungusSporotrichum pulverulentum. Arch. Microbiol. 109:1-8.
2. Andrews, R. P. 1980. Analysis of human serum and plasmausing the LKB 4400 amino acid analyser. Technical LeafletPCN12. LKB Instruments Ltd., Croydon, United Kingdom.
3. Buswell, J. A., B. Molletr and E. Odier. 1984. Ligninolyticenzyme production by Phanerochaete chrysosporium underconditions of nitrogen sufficiency. FEMS Microbiol. Lett. 25:295-299.
4. Chen, C. L., H. M. Chan, and T. K. Kirk. 1982. Aromatic acidsproduced during degradation of lignin in spruce wood by Pha-nerochaete chrysosporium. Holzforschung 36:3-9.
5. Cowan, M. I., T. A. Glen, S. A. Hutchinson, M. E. MacCartney,J. M. Mackintosh, and A. M. Moss. 1973. Production of volatile
3456 HARPER ET AL.
on March 30, 2019 by guest
http://aem.asm
.org/D
ownloaded from

CHLOROMETHANE IS A METHYL DONOR IN P. CHRYSOSPORIUM
metabolites by species of Fomes. Trans. Br. Mycol. Soc.60:347-351.
6. Drew, S. W., and A. L. Demain. 1977. Effect of primarymetabolites on secondary metabolism. Annu. Rev. Microbiol.31:343-356.
7. Eriksson, K.-E., and S. C. Johnsrud. 1983. Mutants of thewhite-rot fungus Sporotrichum pulverulentum with increasedcellulase and P-D-glucosidase production. Enzyme Microb.Technol. 5:425-429.
8. Faison, B. D., and T. K. Kirk. 1985. Factors involved in theregulation of ligninase activity in Phanerochaete chryso-sporium. Appl. Environ. Microbiol. 49:299-304.
9. Faison, B. D., T. K. Kirk, and R. L. Farrell. 1986. Role ofveratryl alcohol in regulating ligninase activity in Phanerocha-ete chrysosporium. Appl. Environ. Microbiol. 52:251-254.
10. Gaertner, F. H. 1978. Unique catalytic properties of enzyme
clusters. Trends Biochem. Sci. 3:64-65.11. Gillis, R. G. 1968. Trideuteromethylation in dimethyl sulphox-
ide. Tetrahedron Lett. 1968:1413-1414.12. Haemmerli, S. D., M. S. A. Leisola, and A. Fiechter. 1986.
Polymerisation of lignins by ligninases. FEMS Microbiol. Lett.35:33-36.
13. Haemmerli, S. D., M. S. A. Leisola, D. Sanglard, and A.Feichter. 1986. Oxidation of benzo (a) pyrene by extracellularligninases of Phanerochaete chrysosporium. J. Biol. Chem.261:6900-6903.
14. Harper, D. B. 1985. Halomethane from halide ion-a highlyefficient fungal conversion of environmental significance. Na-ture (London) 315:55-57.
15. Harper, D. B., and J. T. G. Hamilton. 1988. Biosynthesis ofchloromethane in Phellinus pomaceus. J. Gen. Microbiol. 134:2831-2839.
16. Harper, D. B., J. T. G. Hamilton, J. T. Kennedy, and K. J.McNally. 1989. Chloromethane, a novel methyl donor for bio-synthesis of esters and anisoles in Phellinus pomaceus. Appl.Environ. Microbiol. 55:1981-1989.
17. Harper, D. B., and J. T. Kennedy. 1986. Effect of growthconditions on halomethane production by Phellinus species:biological and environmental implications. J. Gen. Microbiol.132:1231-1246.
18. Harper, D. B., J. T. Kennedy, and J. T. G. Hamilton. 1988.Chloromethane biosynthesis in poroid fungi. Phytochemistry27:3147-3153.
19. Harvey, P. J., H. E. Schoemaker, and J. M. Palmer. 1986.Veratryl alcohol as a mediator and the role of radical cations inlignin biodegradation by Phanerochaete chrysosporium. FEBSLett. 195:242-246.
20. Hatakka, A., A. Kantelinen, A. Tervila-Wilo, and L. Viikari.1987. Production of ligninases by Phlebia radiata in agitatedcultures p. 185-189. In E. Odier (ed.), Lignin enzymic and
microbial degradation. International Symposium, Paris (April23-24), INRA Publications, Paris.
21. Hatakka, A. I., and A. K. Uusi Ravva. 1983. Degradation of14C-labelled poplar wood lignins by selected white-rot fungi.Eur. J. Appl. Microbiol. Biotechnol. 17:235-242.
22. Kawai, S., T. Umezawa, and T. Higuchi. 1986. De novo synthe-sis of veratryl alcohol by Coriolus versicolor. Wood Res.73:18-21.
23. Kirk, T. K., E. Schultz, W. J. Connors, L. F. Lorenz, and J. G.Zeikus. 1978. Influence of cultural parameters on lignin metab-olism by Phanerochaete chrysosporium. Arch. Microbiol. 117:277-285.
24. Leisola, M. S. A., B. Schmidt, U. Thanei-Wyss, and A. Fiechter.1985. Aromatic ring cleavage of veratryl alcohol by Phanero-chaete chrysosporium. FEBS Lett. 189:267-270.
25. Leisola, M. S. A., D. C. Ulmer, R. Waldner, and A. Fiechter.1984. Role of veratryl alcohol in lignin degradation by Phanero-chaete chrysosporium. J. Biotechnol. 1:331-339.
26. Lundquist, K., and T. K. Kirk. 1978. De novo synthesis anddecomposition of veratryl alcohol by a lignin degrading basid-iomycete. Phytochemistry 17:1676.
27. Rasmussen, R. A., L. E. Rasmussen, M. A. K. Khalil, and R. W.Dalluge. 1980. Concentration distribution of methyl chloride inthe atmosphere. J. Geophys. Res. 85:7350-7356.
28. Schoemaker, H. E., P. J. Harvey, R. M. Bowen, and J. M.Palmer. 1985. On the mechanism of enzymatic lignin break-down. FEBS Lett. 183:7-12.
29. Shimada, M., F. Nakatsubo, T. K. Kirk, and T. Higuchi. 1981.Biosynthesis of the secondary metabolite veratryl alcohol inrelation to lignin degradation in Phanerochaete chrysosporium.Arch. Microbiol. 129:321-324.
30. Tien, M., and T. K. Kirk. 1983. Lignin-degrading enzyme fromthe hymenomycete Phanerochaete chrysosporium Burds. Sci-ence 221:661-663.
31. Tien, M., and T. K. Kirk. 1984. Lignin degrading enzyme fromPhanerochaete chrysosporium. Purification, characterization,and catalytic properties of a unique H202-requiring oxygenase.Proc. Natl. Acad. Sci. USA 81:2280-2284.
32. Tonon, F., and E. Odier. 1988. Influence of veratryl alcohol andhydrogen peroxide on ligninase activity and ligninase produc-tion by Phanerochaete chrysosporium. Appi. Environ. Micro-biol. 54:466-472.
33. Wariishi, H., and M. H. Gold. 1989. Lignin peroxidase com-pound III formation, inactivation and conversion to the nativeenzyme. FEBS Lett. 243:165-168.
34. Westermark, U., and K.-E. Eriksson. 1974. Carbohydrate-de-pendent enzymic quinone reduction during lignin degradation.Acta Chem. Scand. B28:204-208.
35. White, R. H. 1982. Biosynthesis of methyl chloride in the fungusPhellinus pomaceus. Arch. Microbiol. 132:100-102.
VOL. 56, 1990 3457
on March 30, 2019 by guest
http://aem.asm
.org/D
ownloaded from