christina ziegler feb 15 th 2010. mechanisms of tolerance induction (1)clonal deletion -negative...
TRANSCRIPT

Christina Ziegler
Feb 15th 2010

Mechanisms of tolerance induction
(1) Clonal deletion- negative selection of thymocytes with high affinity TCR for MHC:self-
antigen (central tolerance)(2) Clonal anergy- auto-reactive T cells encountering their Ag in absence of co-stimulatory
signal become non-responsive to Ag (peripheral tolerance)(3) Clonal ignorance- removal of auto-reactive T cells not encountering their Ag in periphery(4) Anti-idiotypic antibody- Ab against specific idiotypes of other Ab or TCR(5) Regulatory T cells (suppressor cells) - suppressive function via production of TGF-β and IL-10 or cell-cell contact(6) Termination of tolerance- By prolonged absence/exposure to tolerogen, damage of immune system or
immunization with cross-reactive Ag

Development of autoimmune diseases
AUTOIMMUNITY
Breakdown of mechanisms controlling central and/or peripheral tolerance by
(1)Sequestration of antigen- antigen develops late or is only expressed in particular
organ
(2) Escape of autoreactive clones - defective negative selection in thymus
(3) Lack/deficiency of regulatory T cells
(4) Cross-reactive antigens - pathogens antigen may cross-react with self-antigens leading to an autoimmune response like e.g. streptococcal nephritis

Characteristics of regulatory T cells
Natural Tregs (nTregs) developed in thymus with high affinity for self-antigen- CD25+ Foxp3+ CTLA-4+ (5–10% of total CD4+ αβ T cells)
Adaptive Tregs (aTregs) develop from conventional T cells in periphery and can be divided into (a) Th3 cells (CD4+ CD25 - Foxp3-)
-activated by IL-10 which induced its secretion; acts autocrine(b) Tr1 cells (CD4+ CD25 - Foxp3-)
- require IL-10 for maturation, then secrete TGF-β and IL-10- like Th3 cells, Tr1 are abundant in intestine and likely induce
tolerance to food Ag(c) CD8+ Tregs (CD8+ CD25 - Foxp3-)
- shown to suppress CD4+ cells in vitro

Stability of regulatory T cells – STATUS QUO –
• Tregs retain Foxp3 expression under homeostatic conditions after adoptive transfer maybe via positive feedback loop
• During inflammation, Tregs have lower Foxp3 expression • Possible that IL-6 acts in synergy with IL-1 to downregulate
Foxp3• CD4+CD25-Foxp3+ were shown to convert into Th cells
SUM: Peripheral Tregs can become unstable under certain conditions.

Mouse model to analyse stability of Tregs
Adapted from http://commons.wikimedia.org/wiki/File:CreLoxP_experiment.png

Development of Foxp3+ T cells in Foxp3-GFP-Cre x R26-YFP mice
Majority of Foxp3+ cells developed from CD4SP thymocytes (a).
Most Foxp3+ transcription is initiated after maturation of CD4SP thymocytes in the thymus (b ).
Conclusion: Foxp3+ Treg cells develop as ´escape´ mechanism during negative selection process after exposure to self-Ag.

´Ex-Foxp3´ T cells show fading Foxp3 translation in periphery
15% - 20% of YFP cells lack Foxp3 and GFP expression in thymus and peripheral lymphoid organs, respectively (c).
Different peripheral lymphoid organs showed similar proportions of CD4+ T cells expressing Foxp3 at various maturation stages (e).
Conclusion: Certain population of T cells called ´ex-Foxp3´ had ceased translation of Foxp3.

Methylation status of ex-Foxp3+ Treg as indicator for their stability
Differentiation of Tconv, Tregs and ex-Foxp3 Tregs using CD4 vs Foxp3 or GFP vs YFP (A).
Methylation of CpG islands is the principle control mechanism: 90% of CpG motives in TSDR of Foxp3 locus of naive CD4+ Foxp3- Tconv cells are methylated (d).
Tregs were mostly de-methylated (GFP+YFP+), while ex-Foxp3 Tregs (GFP-YFP+) Tregs had random methylation status (d).
Conclusion: Factors controlling the expression of the Foxp3 led to re-methylation of this locus at certain stage in ex-Foxp3 Tregs .

´Ex-Foxp3´ T cells have a non-Treg cell surface phenotype in the periphery
YFP+ ex-Foxp3 T cells were CD25-GITRlowCD127high and thus differ considerably from Foxp3+ Tregs (a).
Loss of ´signature´ Treg markers FR4, CTLA-4 and CD103 on ex-Foxp3 T cells in comparison to Tconv and Foxp3 + Tregs (b).
Conclusion: Ex-Foxp3 T cells do no longer show Treg specific phenotype indicating their instability in homeostatic conditions.
(b) thick line: Tconv cells thin line: Foxp3+ Tregs
filled: ex-Foxp3 T cells

´Ex-Foxp3´ T cells show an effector-memory phenotype
Ex-Foxp3 T cells (GFP-YFP+) showed an activated-memory T cell phenotype (CD62Llow-
highCD44high) (a).
Stimulated YFP+ T cells secreted IFN-γ (b) and IL-17 in GALT (c). Th1 or Th17?
Conclusion: Ex-Foxp3 T cells show an effector-memory T cell phenotype those cytokine profile depends on the microenviron-ment.

Mouse model to study the Foxp3 expression during an autoimmune disease
NOD MOUSE• the non-obese diabetic mouse is a model of autoimmune disease•develops spontaneous autoimmune diabetes similar to T1D in humans incl.
- pancreas-specific autoantibodies- autoreactive CD4+ and CD8+ T cells
•Inflamed pancreatic β islets have lower Treg to Teffector ratio
Theory: Lower Foxp3 expression in the autoimmune disease shifts balance of Tregs to ex-Foxp3 cell phenotype.
Approach: Crossing of Foxp3-GFP-Cre mouse with R26-YFP-NOD mouse
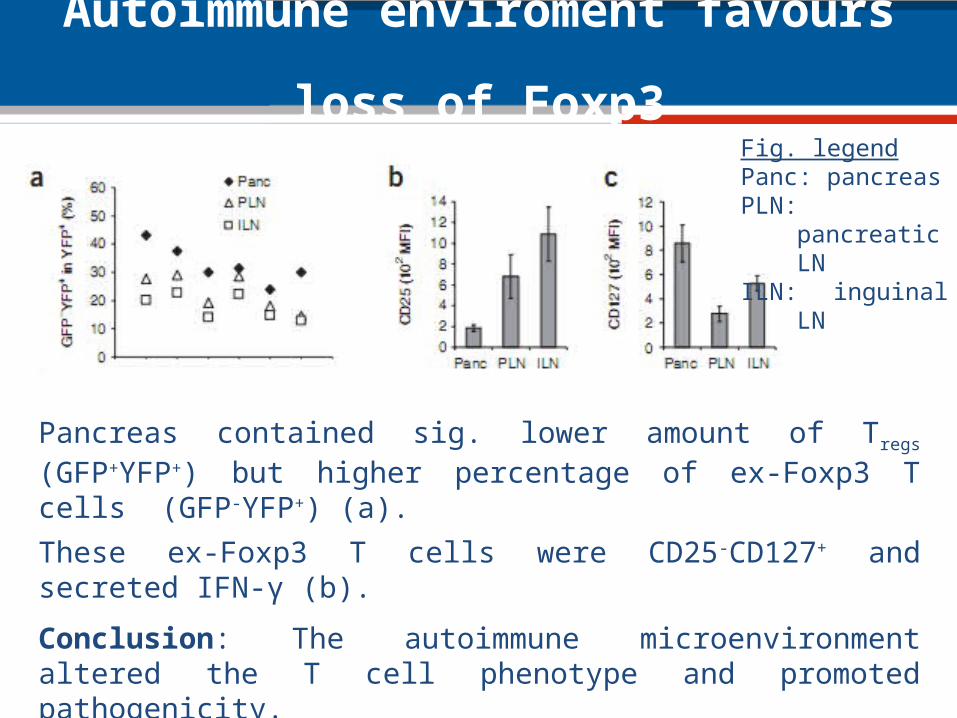
Autoimmune enviroment favours loss of Foxp3
Pancreas contained sig. lower amount of Tregs (GFP+YFP+) but higher percentage of ex-Foxp3 T cells (GFP-YFP+) (a).
These ex-Foxp3 T cells were CD25-CD127+ and secreted IFN-γ (b).
Conclusion: The autoimmune microenvironment altered the T cell phenotype and promoted pathogenicity.
Appearance of ex-Foxp3 T cells was likely consequence of antigen recognition in inflamed area.
Fig. legendPanc: pancreasPLN: pancreatic LNILN: inguinal LN

Mouse model to study if auto-reactive T cells favour development of ex-Foxp3 T cells
BDC2.5 TCR-tg mouseTCR of CD4+ T cells in the BDC2.5 TCR-tg mouse are reactive to a natural pancreatic islet β cell antigen
Theory: Auto-reactive T cells in pancreas changes the percentage of ex-Foxp3 cells and their pathogenic potential.
Approach: Crossing of Foxp3-GFP-Cre x R26-YFP mouse with BDC2.5 TCR-tg mouse.

Autoimmune environment favours loss of Foxp3
Proportions of thymic CD4+Tconv and ex-Foxp3 T cells (GFP-YFP+) similar between non-tg and BCD2.5 mice (d). However, spleen and LN of BCD2.5 mice had more ex-Foxp3 cells (d and e) similar to situation in pancreas of NOD mice.
Conclusion: Strong affinity to self-antigen especially during inflammation promotes generation of ex-Foxp3 T cells.

Mouse model to study if auto-reactive T cells favour development of ex-Foxp3 T cells
NOD Tcra-/- mouseLack αβ T cells and thus are completely protected from autoimmune diabetes.
NOD Rag2-/- mouseHas immunodeficiency and combined cellular and humoral immune defects.
Theory: Tregs are unstable and potentially pathogenic in autoimmune conditions.
Approach: Adoptive transfer of Tregs from Foxp3-GFP-Cre x R26-YFP x BDC2.5 TCR-tg mouse into
a) NOD Tcra-/- mouse and b) NOD Rag2-/- mouse

Ex-Foxp3 cells can be generated from nTregs or aTregs
Adoptively transferred nTregs from BDC2.5 TCR-tg Foxp3-GFP-Cre x R26-YFP mouse into the NOD Tcra-/-
a) had to 1/3 down-regulated Foxp3, b) effector-memory phenotype (a).
After adoptive transfer of Foxp3-
cells into the NOD Rag2-/- mouse, those expressing BDC2.5 TCR were 0.3% YFP+ in the pancreas.
Conclusion: Ex-Foxp3 cells can be generated from instable nTregs or to a lesser extend from abortive aTregs.

Auto-reactive ex-Foxp3 T cells turn into effector cells and then induce T1D
Ex vivo expansion of ex-Foxp3, Tconv and Tregs from BDC2.5 TCR-tg mice for 7-9 d.
20% of GFP+YFP+ and 2% of YFP+ lost Foxp3 expression (b).
Adoptive transfer of three T cell subtypes into the NOD Rag2-/- mouse i)Tregs did not alter the blood glucose levelsii)Tconv and ex-Foxp3 T cells induced diabetes (c and d).
Conclusion: Auto-reactive Ex-Foxp3 T cells turn into effector cells after self-antigen recognition and induce T1D.

Ontogeny of ex-Foxp3 T cells
Unclear if ex-Foxp3 originate from i) aborted Fopx3+ aTreg cells that had converted from Tconv orii) Tconv in the periphery or iii) loss of Foxp3 expression in true CD4+Foxp3+ nTreg cells
Analysis of the CDR3 in various CD4+ T cell subsets from BDC2.5 TCR-tg mice showed that i) all subsets had productive VJ gene rearrangementii) Treg and Tconv cells had dinstinct TCR Vα2 repertoire as only 13% of CDR3 sequence was present in Tconv iii) Ex-Foxp3 cells shared 24% and 36 % sequence CDR3 similarity to Treg
and Tconv, respectively.
Conclusion: Ex-Foxp3 cells have substantial overlap of TCR repertoire with Treg and Tconv and can probably originate from both T cell subtypes.

Summary and conclusions
Substantial fraction of Tregs are unstable in the periphery as a significant percentage
(a)down-regulates Foxp3(b)loses their characteristic Treg phenotype(c)exhibits an activated-memory phenotype and (d)produces pathogenic cytokines(e)loses their suppressive function (f)triggers development of autoimmune disease
•´ex-Foxp3´ T cell levels were elevated in autoimmune conditions •cells share ontogeny with Foxp3+ Tregs and Tconv thus likely originate from nTregs and aTregs
THEORY: Foxp3 instability can lead to the generation of pathogenic effector-memory T cells that promote autoimmunity.

Thank you for your attention!

Possible reasons for the development of autoimmune diseases
• Foxp3 instability can lead to the generation of pathogenic effector-memory T cells that promoter autoimmunity
• Functional deficiency of IL-2 signalling in Treg cells in autoimmunity may disturb the positive feedback loop that controls Foxp3 stability
• Dysfunctional microRNA or Dicer can affect Foxp3 stability• Destabilized Foxp3 possibly involves epigenetic changes in the Foxp3
locus• Early inflammatory cytokines induced by the innate immune system may
disable Tregs and enhance immunity by creating locally pathogenic autoreactive T cell repertoire