city rats: insight from rat spatial behavior into human cognition in urban environments
TRANSCRIPT

Anim Cogn (2011) 14:655–663
DOI 10.1007/s10071-011-0400-yORIGINAL PAPER
City rats: insight from rat spatial behavior into human cognition in urban environments
Osnat Yaski · Juval Portugali · David Eilam
Received: 8 November 2010 / Revised: 4 April 2011 / Accepted: 5 April 2011 / Published online: 20 April 2011© Springer-Verlag 2011
Abstract The structure and shape of the urban environ-ment inXuence our ability to Wnd our way about in the city.Understanding how the physical properties of the environ-ment aVect spatial behavior and cognition is therefore anecessity. However, there are inherent diYculties in empir-ically studying complex and large-scale urban environ-ments. These include the need to isolate the impact ofspeciWc urban features and to acquire data on the physicalactivity of individuals. In the present study, we attempted toovercome the above obstacles and examine the relationbetween urban environments and spatial cognition by test-ing the spatial behavior of rats. This idea originated fromthe resemblance in the operative brain functions and in themechanisms and strategies employed by humans and otheranimals when acquiring spatial information and establish-ing an internal representation, as revealed in past studies.Accordingly, we tested rats in arenas that simulated a gridurban layout (e.g. Manhattan streets) and an irregular urbanlayout (e.g. Jerusalem streets). We found that in the gridlayout, rat movement was more structured and extendedover a greater area compared with their restricted move-ment in the irregular layout. These movement patternsrecall those of humans in respective urban environments,
illustrating that the structure and shape of the environmentaVect spatial behavior similarly in humans and rats. Over-all, testing rats in environments that simulate facets ofurban environments can provide new insights into humanspatial cognition in urban environments.
Keywords Cognitive map · Spatial representation · Spatial behavior · Orientation · Exploration · Urban environment
Introduction
Humans and other animals explore, learn, and Wnd theirway around the environment by using some form of inter-nal representation (Golledge 1999; Jacobs 2003; O’Keefeand Nadel 1978; Tolman 1948; Wang and Spelke 2002).This notion of an internal representation of the externalenvironment was termed a ‘cognitive map’ by Edward Tol-man, a behavioral psychologist (Tolman 1948). Demon-strating the remarkable navigational abilities of rats inspatial problem-solving tasks, Tolman suggested that rats,and by analogy humans, construct a map in their mind/brain and use it for orientation and wayWnding (Tolman1948). A similar notion was introduced by Kevin Lynch, anurban planner, in his seminal book ‘The Image of the City’(Lynch 1960). Based on a Wve-year empirical study in threedisparate US cities (Boston, Jersey City, and Los Angeles),Lynch contended that city dwellers navigate by means of amental image of their cities. He suggested that the legibilityof the urban environment, which reXects the ease withwhich people understand and move about in a city, rests onthe distinctiveness and coherence of Wve urban elements asfollows: landmarks, paths, nodes, districts, and edges. Legi-ble environments enable one to construct a mental image of
David Eilam is a Visiting Professor at the Department of OTANES, University of South Africa.
O. Yaski · D. Eilam (&)Department of Zoology, Faculty of Life Sciences, Tel-Aviv University, 69978 Ramat-Aviv, Israele-mail: [email protected]
J. PortugaliDepartment of Geography and the Human Environment, Faculty of Humanities, Tel-Aviv University, 69978 Tel-Aviv, Israel
123

656 Anim Cogn (2011) 14:655–663
the physical environment and thereby facilitate orientationand wayWnding (Lynch 1960). Lynch ignited the search fortheories and methods aimed at unraveling the relationbetween the spatial cognition and the urban environment,which led to the concept of designing easy to orient urbanenvironments (Darken and Peterson 2001; Haken and Por-tugali 2003). A signiWcant notion on urban spatial cogni-tion, in this respect, is ‘space syntax’ (Hillier 1996; Hillierand Hanson 1984), which scrutinizes the role of urban roadsystems in shaping human spatial behavior. Space syntaxoVers an analytic tool that represents space morphology asa network, providing quantitative measurements of parame-ters such as access or visibility. However, space syntaxfocuses on the centrality of segments (roads) in the urbannetwork and their impact on collective human movement,leaving other perspectives unexplained. Furthermore, spacesyntax has been criticized for providing an explanation for‘how’ people move as a collective without answering thequestion of ‘why’ individuals move (Penn 2003). In orderto answer the latter question, it is necessary to take intoaccount human spatial cognition, and one way to do this isby incorporating empirical research and animal models inorder to assess the impact of the structure on movement.
Following Tolman and Lynch, numerous studies haveprovided evidence that both humans and other animalsestablish a mental representation of the environment, andboth share mechanisms and strategies for acquiring spatialinformation and Wnding their way (Burgess and O’Keefe2003; Golledge 1999; Hills 2006; O’Keefe and Nadel 1978;O’Keefe and Burgess 1996; Thinus-Blanc 1996; Wang andSpelke 2000, 2002). For example, path integration is a nav-igational mechanism that, in the absence of visual informa-tion, enables the traveler (human or non-human, even acreature as ‘simple’ as an ant) to continuously track its cur-rent position in reference to a key point such as home ornest (Etienne et al. 1996; Loomis et al. 1993; Mittelstaedtand Mittelstaedt 1980; Müller and Wehner 1988). Simi-larly, both humans and other animals utilize geometricproperties of the environment for orientation (Cheng 1986;Cheng and Newcombe 2005; see however Cheng 2008;Twyman and Newcombe 2010; Gouteux et al. 2001).
The idea of paralleling spatial behavior of humans andrats is not new. It follows the original and seminal notion ofTolman, who suggested viewing the performances of rats inmazes in relation to the “God-given maze which is ourhuman world” (Tolman 1948). Since then, some of the keybuilding blocks of allocentric spatial representation (i.e.spatial neurons) have been discovered, tested, and appear tobe broadly similar in rats and humans. These include placecells (O’Keefe 1976), head direction cells (Taube et al.1990), grid cells (Hafting et al. 2005), and boundary vectorcells (Lever et al. 2009) in rats. Place cells and head direc-tion cells have also been found in primates (Matsumura
et al. 1999; Robertson et al. 1999), while fMRI evidence ofa grid-cell population signal in humans has also been found(Doeller et al. 2010). All in all, Tolman’s notion on the sim-ilarity of spatial behavior and cognition in humans and ratshas been extended into operative brain functions. Theabove commonalities and the absence of integration of Tol-man’s and Lynch’s concepts (Portugali 2011; Haken andPortugali 2003) led us to the idea that testing rats in settingsthat simulate urban landmarks, paths, and other ‘Lynchianelements’ could provide new insights into human spatialcognition.
Past studies on the relation between the built environmentand the spatial behavior in humans have focused almostexclusively on indoor environments (Arthur and Passini1990; Hölscher et al. 2006; Montello and Sas 2006; Peponiset al. 1990; Weisman 1981). This could be due to the inher-ent diYculties in empirically studying spatial behavior inlarge-scale urban environments. One way to circumvent theproblem of large scales is by testing subjects (either humansor other animals) in a dark environment. In the dark, the sub-ject cannot perceive the entire environment and mustexplore it sector by sector, much as a sighted subject woulddo in a large environment (Ben-Yehoshua et al. 2010; Yaskiet al. 2009). Moreover, in the absence of visual cues, bothhumans and rodents that explore small-scale experimentallayouts were found to use common strategies to acquire spa-tial information (Avni et al. 2008; Avni et al. 2006; Cooperet al. 2001; Golledge et al. 1996; Loomis et al. 1993; Ungar2000; Yaski et al. 2009; Zadicario et al. 2005). We thereforesought in the present study to examine the potential of rats’spatial behavior to serve as an in vivo tool for assessing thelegibility of urban forms. SpeciWcally, we examined whetherrats can provide new insights into human spatial cognition inurban environments.
Materials and Methods
Animals
Male Wistar rats (n = 18; age 3 months; weight 250–300 g)were housed in a temperature-controlled (24°C) room with12/12 h light/dark cycle (lights on at 20:00). Rats were heldin standard rodent cages (40 £ 25 £ 20 cm; 2 rats per cage)and were provided with free access to water and standardrodent chow. At all times, rats were maintained and treatedaccording to the institutional guidelines for animal care anduse in research (permit L-10-013).
Apparatus
A round arena (200 cm diameter) enclosed with a 50-cm-high tin wall was located in a dark, light-proofed room and
123

Anim Cogn (2011) 14:655–663 657
was illuminated with an infrared light source, with an 830-nm Wlter that emits light not visible to the rats (Tracksys,UK). A video camera (Ikegami B/W ICD-47E, Japan) wasplaced above the arena center to provide a top view of theentire arena. Sixteen identical objects (12 £ 12 £ 6 cmcement bricks) were placed in the arena in one of the twofollowing arrangements: (1) a grid layout (Fig. 1a, left), inwhich objects were equally spaced at 25 cm distance inorthogonal rows and columns to form a grid-like array; and(2) an irregular layout (Fig. 1a, right) in which objects werespaced irregularly in the arena with at least 25-cm spacebetween them and from the arena wall. In both layouts, the
average distance between objects was the same. The 25-cmspacing between objects was set according to the average20-cm body length of rats. Thus, a rat could not touch at thesame time two objects or object and arena wall. We previ-ously tried to replicate an urban grid in a ‘grid maze’(Miller and Eilam 2011). However, that maze forces therodents to travel only along speciWc routes, and this is notcomparable with free traveling in urban grids. Therefore,we switched in the present study to testing freely movingrats in an open Weld with objects spaced to simulate anurban grid layout (e.g. the streets of Manhattan) and anirregular urban layout (e.g. the streets of Jerusalem).
Fig. 1 Grid and irregular city layouts, the respective test envi-ronments, and the rats’ paths of movement. a The two simulated urban layouts (top) and their respective test models (bottom): Manhattan (left), as an example of a grid layout; and Jerusalem (right), as an irregular layout. The arrow at the top of the sim-ulated arena represents the start point at which the rats were placed at the beginning of explo-ration. b Paths of progression of four rats in each of the layouts, all in the upper median scores for percentage of area covered during the 20 min of testing. In the grid layout, the rats’ move-ment was dispersed throughout the arena, spanning the objects and the perimeter. In the irregu-lar layout, the rats’ movement was in relation to the start point and the nearby arena wall, cov-ering only a portion of the arena area. The dark areas at the loca-tion of the objects in the grid lay-out represent the convergence of paths at the location of the ob-jects, thus reXecting the layout of the objects (note that Fig. 1b describes only the trajectories, whereas the objects are not plot-ted here but only in Fig. 1a). In contrast, in the irregular layout, convergence of paths at objects is conWned to only a few objects near the start point, thus the lay-out of objects is not revealed by the paths
123

658 Anim Cogn (2011) 14:655–663
Procedure
Rats were randomly assigned to one of the two test groups(n = 9 each), and each rat was tested in only one of the lay-outs. Testing was conducted during the night phase of therats, when they are more active. The test began when a ratwas placed at a Wxed start point near the arena wall (markedby an arrow in Fig. 1a), facing the center of the arena. Itsbehavior was then video-recorded for 20 min. After eachtest, the arena was paper-wiped with detergent.
Data acquisition and analysis
Video Wles were analyzed using ‘Ethovision’ (NoldusInformation Technologies, NL), which tracks the progres-sion of the rat in the arena, providing the time and locationof the center of mass of the rat Wve times per second. Foranalysis, the arena was divided into the following areas:
1. Perimeter area a 15-cm strip along the arena wall(13737 cm2). The perimeter strip was further dividedinto eight identical perimeter areas.
2. Start area one of the eight perimeter areas at whicheach rat was gently placed at the beginning of theexperiment. Accordingly, the center of the start areawas termed the Start point (marked by an arrow inFig. 1a).
3. Object area a circle of 25 cm diameter around eachobject.
4. Arena areas to calculate the covered area, the entirearena was also divided into 84 grid zones (each about373.81 cm2), regardless of object and wall location.
The following parameters were extracted from ‘Ethovision’and exported to ‘Microsoft Excel’ for further analyses:
1. Traveled distance the overall metric distance(m) traveled during the 20 min of observation.
2. Turn angle the mean angular change (degrees) in thedirection of movement, as calculated between each twotime points (Wve time points per second).
3. Meander the mean rate of change in the direction ofprogression relative to the distance traveled, as calcu-lated by dividing the turn angle by the distancebetween each two successive time points. Accordingly,lower meander values characterize locomotion alongrelatively straight trajectories, and higher meander val-ues describe circular or winding trajectories.
4. Duration at area the time spent at each of the perime-ter, start or object areas.
5. Visits to area the incidence of entering each of theperimeter, start or object areas.
6. Objects explored the number of diVerent objects thatwere visited at least once.
7. Area covered utilizing the 84 grid zones, we calculatedthe percentage of the zones that were visited(100% = visiting all 84 zones).
8. Roundtrips to home base the incidence of travel seg-ments that started and ended at the start area.
9. Length of roundtrips to home base the mean metric dis-tance of a roundtrip back to the start area.
10. Object-to-objects trips the incidence of travelingbetween objects without visiting the perimeter area.
11. Object-to-perimeter trips the incidence of travelingbetween an object and the perimeter area.
The data from the two groups were compared by means ofStudent’s t test for independent samples. All t-values aretwo-tailed, and alpha level was set to 0.05.
Results
The impact of object layout on the paths of movement
The impact of the two experimental layouts on the pathsof progression is illustrated in Fig. 1b for four represen-tative rats at each layout. As shown, the ‘grid rats’ trav-eled a greater area and reached more objects comparedwith the rats tested in the irregular layout, where travel-ing was conWned to the vicinity of the start point. Thesechanges in the dispersal of movement were furtherinvestigated by measurable parameters, as detailed else-where.
Rats in the two layouts did not diVer in their overall activity and path shape
In both layouts, the rats did not diVer in their overall trav-eled distance (t16 = 1.33; P = 0.202, Fig. 2a). There wasalso no diVerence in the mean turn angle (t16 = 0.41;P = 0.688) and the mean meander (t16 = 1.64; P = 0.121).Therefore, the diVerent layout of objects did not induce adiVerential level of activity or path shape.
‘Grid rats’ covered a greater area and explored more objects
The ‘grid rats’ covered a greater area (t16 = 2.90; P = 0.011,Fig. 2b) and explored more objects (t16 = 3.33; P = 0.004,Fig. 2c) compared with the rats tested in the irregular lay-out. Indeed, the average number of visits per object washigher in the irregular layout (t16 = 2.72; P = 0.015), but theaverage duration per object did not diVer between the twolayouts (t16 = 1.56; P = 0.138). Thus, despite traveling non-diVerent distances (Fig. 2a), the spatial expansion of thesedistances greatly varied in the two layouts.
123
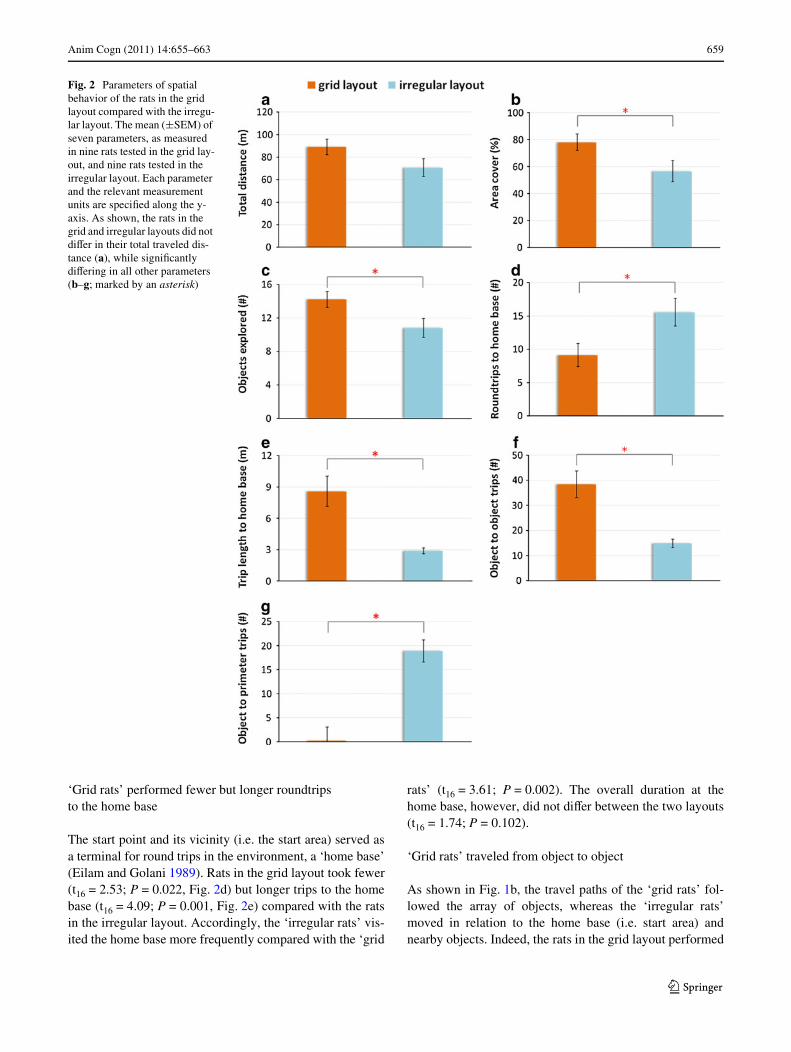
Anim Cogn (2011) 14:655–663 659
‘Grid rats’ performed fewer but longer roundtrips to the home base
The start point and its vicinity (i.e. the start area) served asa terminal for round trips in the environment, a ‘home base’(Eilam and Golani 1989). Rats in the grid layout took fewer(t16 = 2.53; P = 0.022, Fig. 2d) but longer trips to the homebase (t16 = 4.09; P = 0.001, Fig. 2e) compared with the ratsin the irregular layout. Accordingly, the ‘irregular rats’ vis-ited the home base more frequently compared with the ‘grid
rats’ (t16 = 3.61; P = 0.002). The overall duration at thehome base, however, did not diVer between the two layouts(t16 = 1.74; P = 0.102).
‘Grid rats’ traveled from object to object
As shown in Fig. 1b, the travel paths of the ‘grid rats’ fol-lowed the array of objects, whereas the ‘irregular rats’moved in relation to the home base (i.e. start area) andnearby objects. Indeed, the rats in the grid layout performed
Fig. 2 Parameters of spatial behavior of the rats in the grid layout compared with the irregu-lar layout. The mean (§SEM) of seven parameters, as measured in nine rats tested in the grid lay-out, and nine rats tested in the irregular layout. Each parameter and the relevant measurement units are speciWed along the y-axis. As shown, the rats in the grid and irregular layouts did not diVer in their total traveled dis-tance (a), while signiWcantly diVering in all other parameters (b–g; marked by an asterisk)
123

660 Anim Cogn (2011) 14:655–663
more object-to-object trips (t16 = 9.80; P < 0.001, Fig. 2f),while the rats in the irregular layout performed moreobject-to-perimeter trips (t16 = 8.67; P < 0.001, Fig. 2g).
Discussion
Humans, like other animals, are able to Wnd their wayaround in unfamiliar environments by gathering informa-tion from the external surrounding and forming an internalrepresentation (Golledge 1999; Jacobs 2003; O’Keefe andNadel 1978; Tolman 1948; Wang and Spelke 2002). In thepresent study, we tested rats in an unfamiliar experimentalenvironment that simulated a grid urban layout (e.g. Man-hattan streets) and an irregular urban layout (e.g. Jerusalemstreets). We found that movement patterns of rats in theurban-like grid layout were more structured and extendedover a greater area compared with the limited span and lessstructured movement of rats in the irregular urban-like lay-out (Fig. 1b). In the following discussion, we suggest thatspatial behavior of rats and humans is similarly aVected bythe physical properties of the environment. Moreover, dueto the inherent diYculties in empirically studying humanspatial behavior in complex and large-scale urban environ-ments, this study in rats can provide an experimental modelfor acquiring additional insights into some of the cognitiverules that underline the spatial behavior of humans in urbanenvironments.
Relying on the neurophysiological evidence and on thesimilarities in the mechanisms and strategies for spatial ori-entation of humans and animals (see “Introduction”), wetested spatial behavior of freely moving rats in environ-ments that simulated a grid urban layout and an irregularurban layout. We found that in the grid layout, the rats cov-ered more ground, explored more objects, and typicallytraveled from object to object. In contrast, the rats in theirregular layout traveled in only a restricted sector of theenvironment and typically traveled from the start area,where they were introduced into the arena, to nearbyobjects. Since both the ‘grid rats’ and the ‘irregular rats’ didnot diVer in the total distance that they traveled, it is con-cluded that the diVerence in their spatial behavior wasinduced by the diVerent layout of the environmental struc-tures (Fig. 2).
While in geometric grid cities the urban order is appar-ent, in other cities it may look random, but it is not:“Beneath the apparent chaos and diversity of physicalform, there is strong order and a pattern that emerges fromthe myriad of decisions and processes required for a city todevelop and expand physically” (Batty 2008). Indeed, forcenturies, the urban grid was considered as the ideal citylayout (Rose-Redwood 2008), but today the commonlyheld view is that irregular cities, (also termed “bottom-up
organic cities”; Alexander 1965; Hillier 1999) are in manyways better. To date, this assumption is not supported byexperimental studies about the eVect of urban patterns oncognition, or behavior and navigation in cities. Apparently,this is due to the diYculty in conducting experiments withhumans on these issues. In the context of the latter intri-cacy, we suggest that experiments on rats’ spatial behaviorcan shed light on humans’ spatial behavior in respectiveurban environments, as reXected in the following details onthe resemblance between the present Wndings and past Wnd-ings regarding humans’ spatial behavior.
Grid urban layout may increase pedestrian movement byreducing the distance between origin and destination, oVer-ing shortcuts through previously untraveled routes (Frankand Stone 2000; Frank and Engelke 2001; Southworth andOwens 1993). In contrast, irregular urban layout is charac-terized by increased trip length and a decreased choice ofroutes (Frank and Stone 2000; Frank and Engelke 2001;Southworth and Owens 1993). A key comparison of thegrid and irregular urban layouts is that of ‘space syntax’,which is a tool that analyzes the collective movement ofhumans (Hillier 1996; Hillier and Hanson 1984). Accordingto such analysis, urban grids oVer a higher connectivity andthereby higher distribution of pedestrian movementthroughout the city. Irregular urban environments, in con-trast, oVer low connectivity, and movement is thusrestricted to local areas of the city (Hillier 2002; Hillieret al. 1993). Moreover, spatial orientation along orthogonalstreets (e.g. grid urban layout) is easier than along obliquestreets (Montello 1991). This is probably because remem-bering straight lines simpliWes spatial representation.Accordingly, people tend to remember slightly curved linesas straight lines, thus the perceived representation of theenvironment is somewhat distorted in order to facilitate ori-entation (Tversky 1992). The present Wndings in rats seemto reXect the conventional view on grid and irregular urbanenvironments, as well as Hillier’s analysis of pedestrianmovement, demonstrating that the structure of the environ-ment similarly aVects movement patterns in humans andrats. Nevertheless, the cognitive rules that account for theabove similarity in spatial behavior have yet to be uncov-ered.
In an unfamiliar environment, rodents organize theirtraveling in relation to a key location, the ‘home base’(Eilam and Golani 1989). The home base is usually estab-lished near salient landmarks or at the start point of explo-ration, serves as a terminal for roundtrips in the arena, andis characterized by numerous visits (Clark et al. 2006; Draiet al. 2001; Eilam and Golani 1989; Hines and Whishaw2005; Nemati and Whishaw 2007; Tchernichovski et al.1996). Similarly, the rats in the present study established ahome base at the location where they were introduced intothe arena (Fig. 1b). In the irregular layout, the rats primarily
123

Anim Cogn (2011) 14:655–663 661
anchored their roundtrips to the home base and conWnedtheir traveling to nearby objects, as if the irregular layoutmade orientation confusing and therefore required theretracing of steps each time back to the home base (Eilamet al. 2003). In contrast, the rats in the grid layout traveledfurther away from the home base, with longer round tripsthat extended over more objects compared with therestricted trips of the ‘irregular rats’ (Fig. 2). Bearing inmind that the two test layouts diVered mainly in the geo-metric arrangement of the same objects, we suggest that inthe present study, the ordered structure of the grid with itsregularly structured geometry provided a reliable source ofspatial information. This simpliWed the perceived represen-tation of the environment and thereby facilitated orientationand wayWnding. In this sense, the geometric arrangement ofobjects in the grid layout can be viewed like Manhattanstreets—a grid system that is intuitively perceived as a legi-ble environment. In contrast, the irregular object layout thatlacked clear geometry can be viewed as a complex urbanenvironment, like that formed by the winding streets ofJerusalem, or those along the banks of the Charles River inBoston. Altogether, regular geometric forms seem to sim-plify the perceived representation of the environment andfacilitate orientation in both humans and animals (Hillier2002; O’Keefe and Nadel 1978).
DiVerent geometric forms, such as grid and irregular lay-outs, provide diVerent quantity and quality of informationthat aVects spatial behavior. Shannonian information refersto the quantity of information regardless of its meaning andis measured by the reduction in uncertainty (Shannon1948). The amount of uncertainty (i.e. the expected amountof information) is called the ‘entropy’, and the diVerencesin information entropy are commonly measured by bits. InShannonian terms, a grid layout has less entropy because ofits repetitive structure. Thus, a subject that explores a gridlayout compared with an irregular layout has potentiallyless spatial information to gain. This latter point takes us tothe domain of semantic information that is contextual andrefers to the quality (e.g. meaning) of the information(Haken 2006; Haken and Portugali 2003). In terms ofsemantic information, we suggest that a grid layout has aunique quality, since its repetitive structure enables predic-tion of the meaning or quality of the entire structure byexploring only a sector of it. This interaction betweenShannonian and semantic information is an interesting facetof the present study and supports the notion that quantita-tive (Shannonian) and qualitative (semantic) informationare interrelated (Haken 2006; Haken and Portugali 2003).Another interesting facet of the present study are parallelsto the theory of associative learning of temporal intervalsbetween events that reduces uncertainty and elicits antici-patory behavior (Balsam et al. 2010; Balsam and Gallistel2009). Altogether, compared with the irregular layout, the
grid layout of the same objects seems to convey a type ofinformation that makes the grid more legible for spatialmovement.
As noted elsewhere, the empirical study of spatialbehavior in large-scale urban environments has inherentdiYculties. The most obvious diYculties lie in isolatingspeciWc urban features and collecting measurable data onthe physical activity of individuals (Willis et al. 2004). Forthis purpose, a widely used approach is to test humans invirtual city environment (VE) systems (Darken and Sibert1996; Hartley et al. 2003; Jiang 1999; Latini-Corazziniet al. 2010; Omer et al. 2005; Omer and Goldblatt 2007).There are, however, various limitations in VE systems. Forexample, the impact of diVerent viewing perspectives sub-stantially modiWes the perceived image (e.g. bird’s eyeview compared with a walk-through view; Omer et al.2005). This inXuences the subjects’ performance andcauses discrepancies in the results (Hartley et al. 2003;Omer et al. 2005; Omer and Goldblatt 2007). In contrast totesting by means of VEs, the novel approach that we intro-duce in the present study oVers an experimental substratefor uncovering cognitive eVects of speciWc facets of urbandesigns. Consequently, we suggest that spatial behavior ofrats can provide a valid in vivo tool for studying ‘how’ and‘why’ facets of urban environments aVect human spatialcognition and behavior.
Acknowledgments We thank Ms. Naomi Paz for editing the manu-script. This study was supported by the Israel Science Foundation grant177/09 to DE.
References
Alexander C (1965) A city is not a tree. Architectural Forum 122:58–62
Arthur P, Passini R (1990) WayWnding: people, signs, and architecture.McGraw-Hill Ryerson, Toronto
Avni R, Zadicario P, Eilam D (2006) Exploration in a dark open Weld:a shift from directional to positional progression and a proposedmodel of acquiring spatial information. Behav Brain Res171:313–323
Avni R, Tzvaigrach Y, Eilam D (2008) Exploration and navigation inthe blind mole rat (Spalax ehrenbergi): global calibration as aprimer of spatial representation. J Exp Biol 211:2817–2826
Balsam P, Gallistel C (2009) Temporal maps and informativeness inassociative learning. Trends Neurosci 32:73–78
Balsam P, Drew M, Gallistel C (2010) Time and associative learning.Comp Cogn Behav Rev 5:1–22
Batty M (2008) The size, scale, and shape of cities. Science 319:769–771
Ben-Yehoshua D, Yaski O, Eilam D (2010) Spatial behavior: theimpact of global and local geometry. Anim Cogn Publishedonline 24 December 2010
Burgess N, O’Keefe J (2003) Neural representations in human spatialmemory. Trends Cogn Sci 7:517–519
Cheng K (1986) A purely geometric module in the rat’s spatial repre-sentation. Cognition 23:149–178
123

662 Anim Cogn (2011) 14:655–663
Cheng K (2008) Whither geometry? Troubles of the geometric mod-ule. Trends Cogn Sci 12:355–361
Cheng K, Newcombe N (2005) Is there a geometric module for spatialorientation? Squaring theory and evidence. Psychon Bull Rev12:1–23
Clark B, Hamilton D, Whishaw I (2006) Motor activity (exploration)and formation of home bases in mice (C57BL/6) inXuenced by vi-sual and tactile cues: modiWcation of movement distribution, dis-tance, location, and speed. Physiol Behav 87:805–816
Cooper B, Manka T, Mizumori S (2001) Finding your way in the dark:the retrosplenial cortex contributes to spatial memory and naviga-tion without visual cues. Behav Neurosci 115:1012–1028
Darken R, Peterson B (2001) Spatial orientation, wayWnding, and rep-resentation. In: Stanney K (ed) Handbook of virtual environmenttechnology. Lawrence Erlbaum Associates, New Jersey, pp 1–22
Darken R, Sibert J (1996) WayWnding strategies and behaviors in largevirtual worlds. In: Conference on human factors in computingsystems (CHI), Vancouver 1996. ACM, pp 142–149
Doeller C, Barry C, Burgess N (2010) Evidence for grid cells in ahuman memory network. Nature 463:657–663
Drai D, KafkaW N, Benjamini Y, Elmer G, Golani I (2001) Rats andmice share common ethologically relevant parameters of explor-atory behavior. Behav Brain Res 125:133–140
Eilam D, Golani I (1989) Home base behavior of rats (Rattus norvegicus)exploring a novel environment. Behav Brain Res 34:199–211
Eilam D, Dank M, Maurer R (2003) Voles scale locomotion to the sizeof the open-Weld by adjusting the distance between stops: a possi-ble link to path integration. Behav Brain Res 141:73–81
Etienne A, Maurer R, Séguinot V (1996) Path integration in mammalsand its interaction with visual landmarks. J Exp Biol 199:201–209
Frank L, Engelke P (2001) The built environment and human activitypatterns: exploring the impacts of urban form on public health.J Plann Lit 16:202–218
Frank L, Stone B (2000) Linking land use with household vehicleemissions in the central Puget Sound: methodological frameworkand Wndings. Transport Res D- Tr E 5:173–196
Golledge R (1999) Human wayWnding and cognitive maps. In: Goll-edge R (ed) WayWnding behavior: cognitive mapping and otherspatial processes. Johns Hopkins Press, Baltimore, pp 5–45
Golledge R, Klatzky R, Loomis J (1996) Cognitive mapping and way-Wnding by adults without vision. In: Portugali J (ed) The construc-tion of cognitive maps. Kluwer Academic Publishers, Dordrecht,pp 215–246
Gouteux S, Thinus-Blanc C, Vauclair J (2001) Rhesus monkeys usegeometric and non geometric during a reorientation task. J ExpPsychol Gen 130:505–519
Hafting T, Fyhn M, Molden S, Moser M, Moser E (2005) Microstruc-ture of a spatial map in the entorhinal cortex. Nature 436:801–806
Haken H (2006) Information and self-organization: a macroscopic ap-proach to complex systems, 3rd edn. Springer, Berlin
Haken H, Portugali J (2003) The face of the city is its information.J Environ Psychol 23:385–408
Hartley T, Maguire E, Spiers H, Burgess N (2003) The well-worn routeand the path less traveled: distinct neural bases of route followingand wayWnding in humans. Neuron 37:877–888
Hillier B (1996) Space is the machine. Cambridge University Press,Cambridge
Hillier B (1999) The hidden geometry of deformed grids: or, whyspace syntax works, when it looks as though it shouldn’t. EnvironPlann B Plann Des 26:169–191
Hillier B (2002) A theory of the city as object: or, how spatial lawsmediate the social construction of urban space. Urban Des Int7:153–179
Hillier B, Hanson J (1984) The social logic of space. CambridgeUniversity Press, Cambridge
Hillier B, Penn A, Hanson J, Grajewski T, Xu J (1993) Natural move-ment: or, conWguration and attraction in urban pedestrian move-ment. Environ Plann B Plann Des 20:29–66
Hills T (2006) Animal foraging and the evolution of goal-directed cog-nition. Cogn Sci 30:3–41
Hines D, Whishaw I (2005) Home bases formed to visual cues but notto self-movement (dead reckoning) cues in exploring hippocam-pectomized rats. Eur J Neurosci 22:2363–2375
Hölscher C, Meilinger T, Vrachliotis G, Brösamle M, KnauV M (2006)Up the down staircase: wayWnding strategies in multi-level build-ings. J Environ Psychol 26:284–299
Jacobs L (2003) The evolution of the cognitive map. Brain Behav Evol62:128–139
Jiang B (1999) SimPed: simulating pedestrian Xows in a virtual urbanenvironment. J Geogr Inform Decis Anal 3:21–30
Latini-Corazzini L, Nesa M, Ceccaldi M, Guedj E, Thinus-Blanc C,Cauda F, Dagata F, Péruch P (2010) Route and survey processingof topographical memory during navigation. Psychol Res 74:545–559
Lever C, Burton S, Jeewajee A, O’Keefe J, Burgess N (2009) Bound-ary vector cells in the subiculum of the hippocampal formation.J Neurosci 29:9771–9777
Loomis J, Klatzky R, Golledge R, Cicinelli J, Pellegrino J, Fry P(1993) Nonvisual navigation by blind and sighted: assessment ofpath integration ability. J Exp Psychol Gen 122:73–91
Lynch K (1960) The image of the city. MIT Press, CambridgeMatsumura N, Nishijo H, Tamura R, Eifuku S, Endo S, Ono T (1999)
Spatial-and task-dependent neuronal responses during real andvirtual translocation in the monkey hippocampal formation.J Neurosci 19:2381–2393
Miller M, Eilam D (2011) Decision making at a crossroad: why to gostraight ahead, retrace a path, or turn sideways? Anim Cogn14:11–20
Mittelstaedt M, Mittelstaedt H (1980) Homing by path integration in amammal. Naturwissenschaften 67:566–567
Montello D (1991) Spatial orientation and the angularity of urbanroutes: a Weld study. Environ Behav 23:47–69
Montello D, Sas C (2006) Human factors of wayWnding in navigation.In: Karwowski W (ed) International encyclopedia of ergonomicsand human factors, 2nd edn, vol 3. CRC Press/Taylor & Francis,London, pp 2003–2008
Müller M, Wehner R (1988) Path integration in desert ants, Catagly-phis fortis. Proc Natl Acad Sci USA 85:5287–5290
Nemati F, Whishaw I (2007) The point of entry contributes to the orga-nization of exploratory behavior of rats on an open Weld: an exam-ple of spontaneous episodic memory. Behav Brain Res 182:119–128
O’Keefe J (1976) Place units in the hippocampus of the freely movingrat. Exp Neurol 51:78–109
O’Keefe J, Burgess N (1996) Geometric determinants of the placeWelds of hippocampal neurons. Nature 381:425–428
O’Keefe J, Nadel L (1978) The hippocampus as a cognitive map. Ox-ford University Press, Oxford
Omer I, Goldblatt R (2007) The implications of inter-visibility be-tween landmarks on wayWnding performance: an investigationusing a virtual urban environment. Comput Environ Urban Syst31:520–534
Omer I, Goldblatt R, Or U (2005) Virtual city design based on urbanimage theory. Cartogr J 42:15–26
Penn A (2003) Space syntax and spatial cognition: or why the axialline? Environ Behav 35:30–65
Peponis J, Zimring C, Choi Y (1990) Finding the building in wayWnd-ing. Environ Behav 22:555–590
Portugali J (2011) Complexity, cognition and the city. Springer,Heidelberg
123

Anim Cogn (2011) 14:655–663 663
Robertson R, Rolls E, Georges-Francois P, Panzeri S (1999) Headdirection cells in the primate pre-subiculum. Hippocampus9:206–219
Rose-Redwood R (2008) Genealogies of the grid: revisiting Stanislaw-ski’s search for the origin of the grid-pattern town. Geogr Rev98:42–58
Shannon C (1948) A mathematical theory of communication. Bell SysTech J 27:379–423, 623–656
Southworth M, Owens P (1993) The evolving metropolis: studies ofcommunity, neighborhood, and street form at the urban edge.J Am Plann Assoc 59:271–287
Taube J, Muller R, Ranck J Jr (1990) Head-direction cells recordedfrom the postsubiculum in freely moving rats. I. Description andquantitative analysis. Journal Neurosci 10:420–435
Tchernichovski O, Benjamini Y, Golani I (1996) Constraints and theemergence of ‘free’ exploratory behavior in rat ontogeny. Behav-iour 133:519–539
Thinus-Blanc C (1996) Animal spatial cognition: behavioural and neu-ral approach. World ScientiWc, London
Tolman E (1948) Cognitive maps in rats and men. Psychol Rev55:189–208
Tversky B (1992) Distortions in cognitive maps. Geoforum 23:131–138
Twyman A, Newcombe N (2010) Five reasons to doubt the existenceof a geometric module. Cogn Sci 34:1315–1356
Ungar S (2000) Cognitive mapping without visual experience. In: Kit-chin R, Freundschuh S (eds) Cognitive mapping: past, present andfuture. Routledge, London, pp 221–248
Wang R, Spelke E (2000) Updating egocentric representations inhuman navigation. Cognition 77:215–250
Wang R, Spelke E (2002) Human spatial representation: insights fromanimals. Trends Cogn Sci 6:376–382
Weisman J (1981) Evaluating architectural legibility: wayWnding inthe built environment. Environ Behav 13:189–204
Willis A, Gjersoe N, Havard C, Kerridge J, Kukla R (2004) Humanmovement behaviour in urban spaces: implications for the designand modelling of eVective pedestrian environments. EnvironPlann B Plann Des 31:805–828
Yaski O, Portugali J, Eilam D (2009) The dynamic process of cognitivemapping in the absence of visual cues: human data compared withanimal studies. J Exp Biol 212:2619–2626
Zadicario P, Avni R, Zadicario E, Eilam D (2005) ‘Looping’ - anexploration mechanism in a dark open Weld. Behav Brain Res159:27–36
123