clinic for horses - unit for reproductive medicine of clinics · clinic for horses - unit for...
TRANSCRIPT

Clinic for Horses - Unit for Reproductive Medicine of Clinics
University of Veterinary Medicine, Hannover
___________________________________________________________________
Investigations on genital blood flow and embryo recovery
after superovulation with eFSH® and on laparoscopic techniques for flushing
the oviduct in the mare.
THESIS
Submitted in partial fulfilment of the requirements for the degree
DOCTOR OF PHILOSOPHY (PhD)
awarded by the University of Veterinary Medicine Hannover
by
Melanie Carola Witt
from Braunschweig
Hannover 2013

Supervisor: Prof. Dr. H. Sieme
Supervision Group: Prof. Dr. H. Sieme
Prof. Dr. S. Meinecke-Tillmann
Prof. Dr. W. Kanitz
1st Evaluation: Prof. Dr. H. Sieme
Clinic for Horses-Unit for Reproductive
Medicine of Clinics
University of Veterinary Medicine Hannover
Prof. Dr. S. Meinecke-Tillmann
Department of Reproductive Biology
University of Veterinary Medicine Hannover
Prof. Dr. W. Kanitz
FBN Leibniz-Institut für Nutztierbiologie
Dummerstorf, Germany
2nd Evaluation: Prof. Dr. Heiner Niemann
FLI Institut für Nutztiergenetik
Mariensee, Germany
Date of final exam: 28.10.2013

Für
Lasse, Lena und Max
und in Erinnerung an
Prof. Erich Klug

„Am Anfang jeder Forschung steht das Staunen.
Plötzlich fällt einem etwas auf.“
Wolfgang Wickler (*1931)

Parts of this thesis have already been published: I Journal articles Witt, M., H. Bollwein, J. Probst, C. Baackmann, E.L. Squires and H. Sieme (2012):
Doppler sonography of the uterine and ovarian arteries during a superovulatory program in horses, Theriogenology 77, 1406–1414
Köllmann, M., A. Rötting, A. Heberling and H. Sieme (2011):
Laparoscopic techniques for investigating the equine oviduct. Equine Vet. J. 43 (1), 106-111
Köllmann, M., J. Probst, C. Baackmann, J. Klewitz, E.S. Squires u. H. Sieme (2008):
Embryogewinnungsrate nach Superovulation mit equinem Hypophysenextrakt (eFSH®) bei der Stute. Pferdeheilkunde 24 (3), 397– 405
II Abstracts and posters Köllmann, M., A. Rötting, A. Heberling u. H. Sieme (2010):
Laparoskopische Technik zur Untersuchung des equinen Eileiters. 21. Arbeitstagung der DVG- Fachgruppe Pferdekrankheiten, 12.-13.03, Hannover ISBN 978-3-939902-62-1
Köllmann, M., A. Rötting, A. Heberling and H. Sieme (2010): Laparoscopic techniques to investigate the equine oviduct. 6th International Conference on Equine Reproduction - What´s New in Equine Reproduction? - Proceedings 5. Leipziger Tierärztekongresss, Leipzig, 21.-23.01.2010, S. 275-277 ISBN 978-3-86583-441-6
Köllmann, M., C. Baackmann, J. Probst, E.L. Squires and H. Sieme (2009):
Luteal and genital blood flow in mares in a superovulation program. 3th Annual Conference of the European Society for Domestic Animal Reproduction – ESDAR, 10.-12.09. 2009, Ghent, Belgium, In: Reproduction in Domestic Animals 44, Suppl.3 ISSN 0936-6768
Köllmann, M., J. Probst, C. Baackmann, J. Klewitz, E.L. Squires and H. Sieme (2008): Embryo recovery rate in mares after treatment with equine follicle stimulating hormone (eFSH®). 41. Jahrestagung Physiologie und Pathologie der Fortpflanzung, 28. - 29.2. 2008, Gießen Reprod. Dom. Anim. 43 (Suppl.), 21
Köllmann, M., J. Probst, C. Baackmann, J. Klewitz, E.L. Squires, u. H. Sieme (2008):

Embryogewinnungsrate nach Superovulation mit equinem Hypophysenextrakt (eFSH®) bei der Stute. 20. Arbeitstagung der DVG- Fachgruppe Pferdekrankheiten, 29.02.-01.03. Hannover Proc. S. 15, ISBN 978-3-939902-62-1
Köllmann, M., J. Probst, C. Baackmann, E.L. Squires and H. Sieme (2008): Embryo recovery after AI with cooled semen 12 and 36 hours after hCG administration in spontaneously ovulating mares and superovulating mares treated with equine pituitary extract (eFSH). 7th Intn. Equine Embryo Transfer Symposium, 09.-11.07.2008, Cambridge, UK, Havemeyer Foundation Abstract book pp.100
Köllmann, M., J. Probst, C. Baackmann, J. Klewitz, E.L. Squires, u. H. Sieme (2008):
Embryogewinnungsrate nach Superovulation mit equinem Hypophysenextrakt (eFSH®) bei der Stute. 35. Jahrestagung der Arbeitsgemeinschaft Embryotransfer Deutschland AET-de, 19./20.06. Dipperz / Friesenhausen
III. Theses Heberling, A. (2010):
Untersuchungen zur Etablierung eines minimal-invasiven chirurgischen Zugangs zum Eileiter der Stute. Hannover, Tierärztl. Hochsch. Diss.
Probst, J. (2009): Untersuchungen zur Superovulation bei der Stute: Einfluss von eFSH® auf Follikelentwicklung und –durchblutung. Hannover, Tierärztl. Hochsch. Diss.
Baackmann, C. (2008): Untersuchungen zur Superovulation bei der Stute: Einfluss von equinem FSH (eFSH®) auf die genitale Durchblutung unter besonderer Berücksichtigung der lutealen Durchblutung. Hannover, Tierärztl. Hochsch. Diss.
This project was supported by the Mehl-Mülhens-Stiftung, Germany

Contents I
Contents
Chapter
1 Introduction……………………………………………………………………… 1
1.1 Study background- embryo transfer in the mare…………… 2
1.2 Introduction in the theme complex of superovulation……… 4
1.2.1 Reproductive cycle in the mare ……………………………… 4
1.2.2 Endocrine regulation of estrus ……………...………….…….. 4
1.2.3 Follicular development..…………………………………….…. 6
1.2.4 Ovulation………………………………………………………… 9
1.2.5 Double ovulations……………………………………………… 10
1.3 Superovulation in the mare……………………………………. 11
1.4 Genital blood flow in the mare………………..…………….. 15
1.4.1 Doppler sonography …………………………………………… 15
1.4.2 Genital blood supply in the mare……………………………… 17
1.4.3 Uterine blood flow during estrus cycle in the mare………… 18
1.4.4 Ovarian blood flow during estrus cycle in the mare……….....18
1.5 The equine oviduct……………………………………………… 19
1.5.1 Fallopian tubes………………………………………………… 19
1.5.2 Equine embryo development in the oviduct………………….. 20
1.5.3 Methods of oviductal flushing or embryo recovery
from the oviduct………………………………………………… 21
1.5.4 Laparoscopic evaluation of the Fallopian tubes in
women…………………………………………….……………… 23
1.6 Aims of the study……………………………………………… 25

II Contents
Chapter
2 Embryo recovery rate following superovulation with equine pituitary
extract (eFSH®) in mares……………………………………………………… 28
3 Doppler sonography of the uterine and ovarian arteries during a
superovulatory program in horses …………………………………………… 32
4 Laparoscopic techniques for investigating the equine oviduct ..……..…. 36
5 General Discussion………………………………………………………….…. 39
5.1 Superovulation………………………………………………………… 40
5.2 Doppler Ultrasonography……………………………………………… 42
5.3 Laparoscopy of the equine oviduct…………………………………… 44
6 Summary……………………………………………………………….……….. 48
7 Zusammenfassung…………………………………………………………….. 51
8 References………………………………………………………………….…… 54
9 Appendix………………………………………………………………………….83
9.1 Material and methods study I and II………………………………….. 83
9.1.1 Animals…………………………………………………………………. 83
9.1.2 Mare management……………………………………………………. 83
9.1.3 Study design…………………………………………………………… 83
9.1.4 Superovulation treatment…………………………………………….. 84
9.1.5 Embryo collection………………………………………………………. 85
9.1.6 Transrectal Doppler sonography……………………………………… 86
9.1.7 Blood collection, progesterone and estrogen analysis……………. 88

Contents III
9.1.8 Statistical analysis……………………………………………………… 88
9.2 Material and methods study III……………………………………….. 89
9.2.1 Animals………………………………………………………………… 89
9.2.2 Preoperative management…………………………………………… 89
9.2.3 Instruments used in experiment I……………………………………. 90
9.2.4 Surgery experiment I…………………………………………………… 91
9.2.5 Preoperative management experiment II…………………………….. 92
9.2.6 Instruments used in experiment II……………………………………. 92
9.2.7 Surgery experiment II………………………………………………….. 94
9.2.8 Postoperative care in both experiments…………………………….. 101

IV Abbreviations
Abbreviations A./ Aa. Arteria/ Arteriae
a.m. ante meridiem
B-mode brightness modulation
BFV blood flow volume
bwt body weight
c cycle
CF continuous-wave doppler
Ch. Charrière
CL corpus luteum
cm centimeter
CO2 carbon dioxide
°C degree Celsius
D day
eCG equine chorionic gonadotropin
EDTA ethylenediaminetetraacetic acid
eFSH equine follicle stimulating hormone
IGF insulin-like growth factor
EIA enzyme immunoassay
EPE equine pituitary extract
et al. et alii
ET embryo transfer
E estradiol
Etot total estrogens
FDP follicular development phase
Fig. figure
FSH follicle stimulating hormone
g gram
g acceleration of gravity
GIFT gamete intra-fallopian tube transfer
GnRH gonadotropin-releasing hormone

Abbreviations V
h hour(s)
hCG human chorionic gonadotropin
hMG human menopausal gonadotropin
ICSI intra-cytoplasmic sperm injection
IGF insulin-like growth factor
IU international units
im intramuscular
iv intravenous
IVF in vitro fertilization
Kg kilogram
L liter
LH luteinizing hormone
mg milligram
MHz megahertz
min minute
mL milliliter
mmHg mm mercury
μL microliter
μm micrometer
Ø mean value
OV ovulation
ov ovarian
ovBFV ovarian blood flow volume
ovPI ovarian pulsatility index
P level of significance
P4 progesterone
pFSH porcine follicle stimulating hormone
PGE2 prostaglandin E2
PGF2α prostaglandin F2α
PI pulsatility index
p.m. post peridiem

VI Abbreviations
POP preovulatory phase
PW pulsed-wave doppler
R./ Rr. ramus/ rami
S.E.M. standard error of mean
SC stimulated cycles
sid semel in die; single a day
TAMV time averaged maximum velocity
USC unstimulated cycles
USP United States Pharmacopeia System
ut uterine
utBFV uterine blood flow volume
utPI uterine pulsatility index
UTJ utero-tubal junction
V./ Vv. vena/ venae

List of figures and tables VII
List of figures and tables
Fig. 1-1 Concentrations of FSH and estradiol in peripheral circulation and size of the largest follicle and concentrations of LH, progesterone and prostaglandinF2α in the peripheral circulation throughout the equine cycle (AURICH 2011)…………… 5
Fig. 1-2 Illustration of the hormonal aspects of deviation using a-two follicle model (GINTHER et al. 2001)…………………………. 8
Fig. 1-3 Follicular development during oestrus cycle of the mare; size of largest follicle, concentrations of hormones in peripheral circulation and occurrence within the largest follicle (AURICH, 2011)……………………………………….. 9
Fig. 1-4 Pulsatile arterial flow over one cardiac cycle………………………. 16
Fig. 1-5 Lateral view of arterial blood supply of the mare´s genital tract (GINTHER 2007)……………………………………………………….. 18
Fig. 1-6 Drawing of lateral view of ovary and associated structures of the mare……………………………………………………………… 20
Fig. 1-7 Scanning electron micrographs of the mare oviduct at the estrus phase……………………………………………………….. 21
Fig. 9-1 eFSH® treatment protocol in cycle 2 and 4………………………….. 85
Fig. 9-2 Ultrasound investigation of the left A. uterina of a mare one day before ovulation……………………………………………………. 87
Fig. 9-3 Ultrasound investigation of the right A. ovarica of a mare two days before ovulation…………………………………………… 87
Fig. 9-4 Trocar set used for experiment I……………………………………… 90
Fig. 9-5 Picture of the flushing catheter……………………………………… 93

VIII List of figures and tables
Fig. 9-6 Picture of the 1) plastic guide sleeve and 2) metal guide sleeve for guidance of the catheter …………………………………………… 94
Fig. 9-7 Drawing of the left flanc region and portal sites…………………….. 95
Fig. 9-8 Laparoscopic view of the right ovary (star), infundibulum (large arrow), oviduct (arrowheads) and tip of the uterine horn (circle) before oviductal flushing……………………………… 96
Fig. 9-9 Picture of the left flanc laparoscopy surgery site for oviductal
flushing in the standing sedated mare………………………………. 97
Fig. 9-10 Laparoscopic view of the left part of the infundibulum „opened up“ by a Babcock forceps; the balloon catheter is directed into the abdominal ostium (large arrow)…………………… 98
Fig. 9-11 The catheter (large arrow) is introduced approximately 2 cm
into the ampulla and the balloon (arrowhead) is insufflated with 2 ml of air, the Babcock forceps is positioned around the catheter and the abdominal ostium to prevent back-flow, methylene blue fluid in the proximal ampulla (asterisks)…… ….. 99
Fig. 9-12 Picture showing the oviductal ampulla filled with methylene
blue solution (arrowheads) following the oviductal flushing, the isthmus (stars) is not visibly filled or distended………………… 100
Tab. 9-1 Overview of the five investigated estrus cycles and the
according treatments (eFSH®-treatment, induction of ovulation with hCG, insemination and embryo collection 6.5 days post OV respectively)…….…………………………… 84
Tab. 9-2 Pain score system…………………………………………………… 101
Tab. 9-3 Clinical details of 10 mares of experiment I and II……………….. 102

Introduction 1
Chapter 1
General Introduction

2 Introduction
1. Introduction
1.1 Study background- embryo transfer in the mare
Embryo transfer (ET) has been part of cattle breeding for more than 35 years
(SCHERZER et al. 2008) and has also gained remarkable interest from the equine
industry after several breeds allowed registration of more than one foal per year.
However, success rates after superovulation and cryopreservation of embryos in
horses are still lagging behind those of cattle (SCHERZER et al. 2008). During the
last 20 years the number of equine ETs performed annually worldwide has grown
enormously. This is illustrated by the International Embryo Transfer Society’s (IETS)
annual statistics for equine ET which document 475, 11672, 27594 and 28661
commercial equine ETs worldwide in 1999, 2004, 2010 and 2011, respectively
(THIBIER 2000, 2005; STROUD 2011, 2012).
The most important breakthroughs were the development of techniques for non-
surgical transfer of embryos that yielded pregnancy rates >80%, not only for freshly
transferred embryos (VOGELSANG et al. 1985; RIERA and MCDONOUGH 1993;
MCKINNON et al. 1998), but also for embryos transported at 5°C for up to 24 h
(CARNEVALE et al. 1987; CARNEY et al. 1991).
The majority of embryos collected from donor mares are from spontaneously
ovulating mares with single ovulations. They are generally recovered 7 or 8 days
after ovulation (SQUIRES et al. 2003). Following fertilization at the ampulla-isthmus
junction, transport through the oviductal isthmus occurs rapidly and the late morula or
early blastocyst enters the uterus through the utero-tubal junction 144 - 156 h after
ovulation (OGURI and TSUTSUMI 1972; WEBER et al. 1996; BATTUT et al. 1997).
Unfertilized eggs on the other hand are retained in the oviduct (VAN NIEKERK and
GERNEKE 1966).
The day of recovery, number of ovulations, age of donor mare and the quality of
semen are factors that affect embryo recovery (SQUIRES 1996). Mean embryo
recovery per cycle from spontaneously ovulating mares with single ovulations is
approximately 50%. An ability to consistently induce multiple follicles > 30 mm and
multiple ovulations in mares would enhance embryo recovery from donor mares,

Introduction 3
provide multiple follicles for collection of oocytes, and improve pregnancy rates from
subfertile mares (SQUIRES 2006).
At present, however, the vast majority (>95%) of horse embryos are transferred fresh
or after cooled storage for up to 24 h, whereas cryopreservation or vitrification is
rarely employed in clinical use (STOUT 2012). A major impediment to the
implementation of embryo cryopreservation in the field is that acceptable pregnancy
rates (>55%) are, at present, achievable only with embryos recovered at an early
developmental stage (day 6–6.5; morula to early blastocyst) when they are <300 µm
in diameter (CZLONKOWSKA et al. 1985; SLADE et al. 1985; ELDRIDGE-
PANUSKA et al. 2005). The influence of size appears to be even more absolute for
vitrification, because embryos >300 µm show a reduced ability to re-expand during
post-warming incubation (HOCHI et al. 1995) and very rarely result in pregnancy
after vitrification and warming (ELDRIDGE-PANUSKA et al. 2005; CARNEVALE
2006; SCHERZER et al. 2011), whereas larger embryos cryopreserved by slow-
freezing do yield normal pregnancies, albeit at a lower rate (<20%) than for small
embryos (SLADE et al. 1985; BARFIELD et al. 2009). The embryonic capsule and
the amount of blastocoel fluid in embryos of larger diameter were hypothesized as
reasons for the low success of vitrification or freezing (MACLELLAN et al. 2002;
BASS et al. 2004; BARFIELD et al. 2009). But nevertheless, in latest works from
laboratories using micromanipulation capabilities, first positive attempts in vitrification
of expanded blastocytes were made (CHOI et al. 2010). It was found that collapsed
(by embryo biopsy) equine blastocysts (initial diameters 407-565 µm) could be
efficiently vitrified and resulted in pregnancy rates of 46% (6/13) (CHOI et al. 2011;
HINRICHS and CHOI 2012).
Moreover, the exact time of the passage into the uterus and rate of embryo
development appear to vary, depending for example on the time of year, type of
semen used (fresh vs. frozen) and age of the donor mare (STOUT 2006). The scale
of the variation in developmental rate was demonstrated by COLCHEN et al. (2000)
who recorded ranges in diameter and cell number, respectively, of 159–365 µm and
272–2217 cells for embryos collected at 168 ± 0.5 h, and 162–245 µm and 117–417
cells for embryos collected at 156 ± 0.5 h after ovulation (COLCHEN et al. 2000).

4 Introduction
A number of inter-related factors have contributed to the slow development and
implementation of equine embryo cryopreservation, and these include the following:
- the absence of commercially available products for reliably stimulating
superovulation
- very poor pregnancy rates following cryopreservation of embryos >300 µm in
diameter
- difficulty in recovering embryos at early developmental stages amenable to
cryopreservation (BATTUT et al. 1997; STOUT 2012)
In the following section an introduction into the fields of superovulation and follicle
development in the mare will be given, and the oviduct as the early storage of
embryos will be introduced in detail.
1.2 Introduction in the theme complex of superovulation 1.2.1 Reproductive cycle in the mare
Mares are a seasonally polyestrous species with ovulatory activity being related to
long days and light. They will experience reproductive activity during the spring and
summer month, between May and October. During the breeding season, average
estrus cycle length is about 21 to 22 days in length. An estrus during the follicular
phase lasts 5–7 days characterized of behavioural signs like increased interest in
stallions and proceptive behaviour in response to the sexual attractivity of a stallion
(CROWELL-DAVIS 2007). This is followed by the luteal phase, or diestrus, and lasts
14 to 16 days (NIE 2007; AURICH 2011). The cycle length is also affected by time of
breeding season or reproductive stage (HEIDLER et al. 2004).
1.2.2 Endocrine regulation of estrus
The endocrinological control of the estrus cycle is governed by the hypothalamic-
pituitary-gonad axis. In the mare, the gonadotropins LH and FSH are considered to
be under the control of GnRH alone. So far there is no evidence that a specific FSH-
releasing factor exists in the horse (AURICH 2011). Hypothalamic GnRH release is
modulated by steroid feedback mechanisms (IRVINE and ALEXANDER 1993). An
early periovulatory rise in peripheral concentrations of LH is accompanied by a

Introduction 5
modest increase in FSH subsequently declining to its nadir concentration while LH is
reaching its maximum (BERGFELT et al. 1991). In the mid-luteal phase, a second
and robust FSH rise occurs with no concomitant increase in LH. This second FSH
surge occurs on different days of the cycle among individual mares (GINTHER et al.
2005). In contrast to other domestic animal species which exhibit a short and
pronounced preovulatory LH surge, no distinct periovulatory LH peak exists in the
mare. However, during estrus the period of elevated concentrations of LH lasts for
several days. A simplified overview of concentrations of FSH, estradiol, LH and
progesterone in relation to the largest follicle is shown in Fig.1-1.
Fig. 1-1: Concentrations of FSH and estradiol in peripheral circulation and size of the largest follicle and concentrations of LH, progesterone and prostaglandinF2α in the peripheral circulation throughout the equine cycle (AURICH 2011)
Estradiol
Estrus
Estrus
Day of estrus cycle

6 Introduction
1.2.3 Follicular development
Mares are a monovulatory species: typically one follicle becomes dominant and
several subordinate follicles regress during the primary follicular wave of the estrus
cycle before ovulation (GINTHER and BERGFELT 1992). Compared to other
domestic animal species, the mares´ ovary has a unique structure characterized by a
large size and weight (35–120 cm3 in volume; 40–80 g in weight) (KIMURA et al.
2005), the presence of an ovulation fossa and an inverted location of its cortex and
medulla (KAINER 1993).
Although scientific interest in equine follicles existed already since the 1920s
(SEABORNE 1925), detailed studies on equine follicle dynamics did start by the
pioneering lead of OJ Ginther at the University of Wisconsin (reviewed in GINTHER
1979, DONADEU and PEDERSEN 2008).
As in other farm animal species and humans, the development of antral follicles in
the horse is characterized by the periodic growth of cohorts of follicles (SIROIS et al.
1989; BERGFELT and GINTHER 1993). Follicular waves are generally divided into
primary and secondary waves during the estrus cycle of the mare. Only the primary
wave appears to produce the dominant follicle that goes on to ovulate during estrus
(GINTHER et al. 2004). The primary wave emerges during midluteal diestrus from
the stimulation of a FSH surge, the dominant follicle becomes preovulatory and
results in ovulation at the end of estrus. The secondary wave emerges during late
estrus of the previous estrus cycle or early diestrus where the dominant follicle is
either anovulatory and regresses, forming an anovulatory hemorrhagic follicle, or
results in secondary ovulation during mid-diestrus (BEG and GINTHER 2006).
In the mare the primary follicular wave emergence is characterized by a follicle
diameter of 6 mm in the largest follicle (GASTAL et al. 1997). A mean number of 7 -
11 follicles emerge over several days and enter a common growth phase of about 3
mm per day (GASTAL et al. 2004; GINTHER et al. 2004). The emergence of each
follicular wave is temporally associated with an FSH surge. FSH reaches a plateau
when the largest follicle reaches a size of about 13 mm in diameter (GASTAL et al.
1997; DONADEU and GINTHER 2001). Subsequently, FSH declines to a
concentration that does not support pronounced further growth of subordinate

Introduction 7
follicles but is sufficient for continuing growth of the dominant follicle. The inhibition
of growth of the smaller follicles does not depend on follicle-to-follicle inhibitory
mechanisms, but follicle deviation involves important changes in the largest follicle
(AURICH 2011). These are characterized by an increased sensitivity to circulating
concentrations of FSH. Dramatic changes in the insulin-like growth factor (IGF)
system (IGF-I and -II, IGF binding protein, IGF binding protein proteases) in the
largest follicle before the beginning of size deviation play a crucial role (BEG and
GINTHER 2006). Simultaneously, the future dominant follicle suppresses circulating
concentrations of FSH, most probably due to follicular synthesis and release of
estrogens (GASTAL et al. 1999; DONADEU and GINTHER 2001) and Inhibin
(WATSON and AL-ZI'ABI 2002).
The low FSH concentration, yet, does not restrict the growth of the dominant follicle
which by that time has acquired the ability to more efficiently use circulating
gonadotropins for growth (GINTHER et al. 2003) and to produce high levels of inhibin
and estradiol. These declining FSH concentrations continue to support growth of the
follicles of the wave until the appearance of the two largest follicles at the time of
deviation: generally 22.5 and 19 mm in diameter, 16 days postovulation or about 8
days before ovulation (GINTHER et al. 2004; JACOB et al. 2009).
Follicular deviation is an abrupt event recognized by the sudden decrease in the
growth rate of the second largest follicle by about 2 days after the beginning of
deviation (ROSER and MEYERS-BROWN 2012). This is thought to be a key
component of the follicular selection process in monovulatoy species such that
usually one follicle becomes the preovulatory follicle, whereas the others regress
owing to an interplay between circulating gonadotropins and follicular factors within
the ovary (BEG and GINTHER 2006; GINTHER 2012). During these 2 days,
depending on the size difference of the two follicles and the subordinate follicles,
treatment with exogenous gonadotropins could enhance the potential for multiple
dominant follicles by rescuing those follicles that start to regress. Mechanisms of
follicle development and deviation are shown in Fig. 1-2 and 1-3.
The critical role of low FSH levels in the deviation mechanism in mares is illustrated
by the disruption of the deviation mechanism after administration of FSH (SQUIRES

8 Introduction
2006) or immunization against inhibin (MCCUE et al. 1992) leading to the
development of multiple ovulatory follicles.
Fig. 1-2: Illustration of the hormonal aspects of deviation using a two- follicle model (GINTHER et al. 2001).
0= dominant and subordinate follicle When the follicles reach about 13 mm, they both secrete increasing concentrations of inhibin during the common-growth phase (before deviation). About a day before deviation, increased estradiol is secreted by the largest follicle under the influence of increased concentrations of LH. Apparently, the increasing estradiol acts in conjunction with Inhibin to continue the reduction in FSH concentrations after deviation. The elevated LH continues to stimulate the production of estradiol by the developing dominant follicle and has a positive diameter effect on the dominant follicle within 2 days after the beginning of deviation.
Fo
llic
le

Introduction 9
Fig. 1-3 Follicular development during the estrus cycle of the mare; size of follicles, hormone changes in peripheral circulation and occurrence within the largest follicle; =follicle = ovulation (AURICH 2011)
1.2.4 Ovulation
The preovulatory follicular development and ovulation in horses differ from other
animal species. The preovulatory follicle is much bigger in size and ovulates at the
ovulation fossa - a specific region of the mare´s ovary (AURICH 2011). The
preovulatory follicle grows at an average rate of 3 mm per day and reaches a
diameter of approximately 35 mm four days before ovulation. Continued growth
occurs up to 2 days before ovulation when follicular size reaches a plateau of
Follicle size
(primary wave)

10 Introduction
approximately 40 mm (GINTHER et al. 2008b). During ovulation, the oocyte enters
the oviduct, while most of the follicular fluid passes into the peritoneal cavity
(TOWNSON and GINTHER 1989) and only a small volume of follicular fluid appears
to accompany the oocyte/cumulus complex into the oviduct (TOWNSON and
GINTHER 1989). Hormones from this fluid are rapidly absorbed into the circulation
leading to a pronounced increase in concentrations of inhibin on the day of ovulation
(BERGFELT et al. 1991). The ovulatory process of the equine follicle involves a
specific and unique pattern of gene regulation in theca and mural granulosa cells.
This includes differences in the expression of a variety of factors among them
prostaglandins and prostaglandin metabolizing enzymes (SAYASITH et al. 2009).
1.2.5 Double ovulations
Spontaneous double ovulations may occur in horses. The double ovulation rate is
affected by various factors such as breed, reproductive status, age and
pharmacological manipulation of the estrus cycle (STABENFELDT et al. 1972;
GINTHER et al. 1982; SIEME and KLUG 1996). The incidence of spontaneous
double ovulation varies between approximately 2% in ponies and 25% in
thoroughbreds, respectively. When two dominant follicles (two follicles >28 mm)
develop in the same follicular wave, double ovulations occur in about 40% of mares
(GINTHER et al. 2008a). These may occur synchronously (within 12 h), but intervals
up to two days and more have been reported between ovulations (GINTHER et al.
2008a). During the 2.5 immediately days before ovulation, the rate of dominant
follicle growth in double ovulating mares is less pronounced than in single ovulating
mares resulting in a lower preovulatory follicle diameter in twin ovulating mares
(GINTHER and BERGFELT 1992). The reduced follicular growth is related to lower
FSH concentrations, most probably due to higher estradiol concentrations from the
two preovulatory follicles (GINTHER et al. 2008a). The peak of FSH levels that
occurs 3 days before deviation is responsible for the development of the dominant
follicle(s), and the decline of FSH causes the regression of subordinate follicles
during the primary follicular wave. But the role of LH, inhibin, and estradiol still needs
to be elucidated in the further development and maturation of the preovulatory follicle

Introduction 11
to ovulation (ROSER and MEYERS-BROWN 2012). Taken together, it is difficult to
discern the role of FSH, LH, and estradiol in inducing one ovulation or double
ovulations during the pre- and periovulatory period. It is conceivable that the effects
of systemic hormones on the intrafollicular factors and their receptors in dominant
and subordinate follicles during the primary follicular wave and the periovulatory
period play a major role in determining whether mares are multiple ovulators
(ROSER and MEYERS-BROWN 2012). Mares that tend to have multiple ovulations
continue to do so in a superovulatory regimen (LOGAN et al. 2007).
1.3 Superovulation in the mare
The percentage of double ovulations in mares is low. The success of advanced
reproductive technologies in the mare would be enhanced by effective superovulation
to provide multiple oocytes and multiple embryos for such techniques as embryo
transfer, gamete intra-fallopian tube transfer (GIFT) and intra-cytoplasmic sperm
injection (ICSI). Superovulation can increase pregnancy rates in normal and
subfertile mares as well as when using semen from subfertile stallions (SQUIRES
2006).
The basis of superovulation is manipulation of the hormones that control the
dominant follicle and inhibit the regression of subordinate follicles (SQUIRES and
MCCUE 2007). Superovulation has been attempted in the cycling mare during the
past 35 years beginning with studies by DOUGLAS et al. (1974). LAPIN and
GINTHER (1977) reported induction of ovulation and multiple ovulations in
seasonally anovulatory and ovulatory mares with an equine pituitary extract (EPE)
preparation. Since then, many other investigators have used various hormone
regimens to induce superovulation in the cycling mare (for reviews see: MCCUE
1996; SQUIRES and MCCUE 2007; SQUIRES and MCCUE 2011). Attempts to
superovulate cyclic mares using preparations of equine chorionic gonadotropin
(DINGER et al. 1982), GnRH (BECKER and JOHNSON 1992; DIPPERT et al. 1992),
porcine FSH (FORTUNE and KIMMICH 1993; CULLINGFORD et al. 2010; RAZ et al.
2010) and active immunization against inhibin (MCCUE et al. 1992; NAMBO et al.

12 Introduction
1998; DERAR et al. 2004) have demonstrated great variability in the results in most
cases.
EPE lead to an increase in the number of smaller follicles. Some of the earlier studies
in pony mares showed an increase in the number of ovulations during anestrus
(DOUGLAS 1979). During the natural breeding season, treatment before a 25- mm
follicle was present resulted in increased ovulations, whereas treatment of mares
with a follicle over 25 mm did not change ovulation rates or increase the number of
ovulations and embryos recovered (DIPPERT et al. 1992). These data suggest that
treatment initiated after the dominant follicle is established, usually around day 15,
may not be effective in rescuing subordinate follicles and increasing ovulation rate. It
was suggested that the reason for this finding was that because of the FSH within the
preparation follicles were rescued from atresia. The variability in the response in
these studies may be due to the variability in the size of the cohort of follicles present
at the time of initial administration of EPE, as the standard time of initial treatment
was 5-6 days postovulation and not based on the size of the follicles present.
Therefore, administration of EPE, before the dominance is established, was found to
be the treatment of choice (PIERSON 1990). Of 170 mares treated with EPE at
Colorado State University, an average of 3.2 ovulations was detected and 1.96
embryos were recovered per mare compared to 0.65 embryo recovered from
untreated control mares (SQUIRES and MCCUE 2007).
In addition, purity of EPE is a problem, as the ratio of FSH to LH does not remain
constant between preparations (ROSER and MEYERS-BROWN 2012).
Equine FSH
In the past decade, a semipurified EPE (eFSH; Bioniche Animal Health, Bogart, GA)
became commercially available. Based on radioimmunoassay, this preparation
contained 110 mg of FSH/mg and 10 mg of LH/mg, an FSH to LH ratio of 10:1
compared with an EPE preparation that had a 5:1 ratio measured by
radioimmunoassay (WELCH et al. 2006). Although eFSH was commercially available,
there was still variability of responses between mares (ALLEN 2005). Factors that
affect the response of mares include day of initial treatment, size of follicles at
initiation, and frequency of treatment injection (ALLEN 2005). To design an optimal

Introduction 13
treatment regimen using eFSH for the present study the following aspects of earlier
studies were considered:
Dose
NISWENDER et al. (2003) first investigated the use of 12 mg (twice-daily
intramuscular injections- total 25 mg/day) or 25 mg of eFSH given in twice-daily
intramuscular injections (total 50 mg/day) to mares during the ovulatory season.
Treatment was initiated 5-6 days postovulation to ensure stimulation to occur during
the active growth phase of follicular waves. For both treatment groups luteolysis was
induced on the second day of treatment and was used to remove the effect of
progesterone. When a majority of follicles measured 35 mm in diameter, ovulation
was induced with either deslorelin or human chorionic gonadotropin. Treatment with
twice daily 12 mg of eFSH increased the number of follicles >35 mm. Ovulations
were also increased to 3.6 versus 1.0 in control animals. Embryos retrieved
increased from 0.5 to 1.9 in mares given the 12-mg-dose twice a day. Treatment with
25 mg of eFSH twice daily resulted in an increased number of follicles but not
ovulation rates. Treatment with 12 mg (twice-daily intramuscular injections- total 25
mg/day) of eFSH was determined as an optimal dose (NISWENDER et al. 2003).
Treatment start
MCCUE et al. (2006, 2007) evaluated different times for treatment start with eFSH
and reached the best results when treatment start was 5–7 days after ovulation when
a cohort of follicles 20–25mm in diameter was present.
Pretreatment
Different protocols for pretreatments before eFSH application to increase embryo
recovery rates have also been reported. The basis for these studies was to induce a
follicular wave with progesterone and estradiol, simulating the mare’s physiological
follicular waves and timing of follicular development and deviation so as to more
accurately time treatment with eFSH. In a study of RAZ et al. (2005) there was no
advantage with a progesterone and estradiol treatment, LOGAN et al. (2007)
reported that pretreatment with progesterone and estradiol-17ß plus 12.5 mg of
eFSH, decreased the number of ovulations compared with administration of eFSH
alone. The number of embryos recovered was 0.7 and 1.5 embryos in the

14 Introduction
progesterone- and estradiol-17b-treated group compared with 2.6 embryos in the
eFSH-only group.
“Coasting”
“Coasting” can be defined as a certain time period of stopping the eFSH treatment
before induction of ovulation. In a study conducted by WELCH et al. (2006) the
authors found a higher embryo recovery rate by stopping the twice-daily treatment of
eFSH at the time of a 32-mm follicle for 42-50 hours before hCG then giving hCG
right after eFSH treatment when a follicle reached 35 mm in diameter. This idea was
adapted from studies in women where results of a continous stimulation program
also showed an ovarian hyperstimulation (FLUKER et al. 1999). An ovarian
hyperstimulation could be seen in studies in cattle (SIRARD et al. 1999) after a
continous stimulation treatment. According to SQUIRES and MCCUE (2011), the
benefits of coasting are to prevent hyperstimulation, which would result in a reduced
receptor response, limit the occurrence of anovulatory follicles, and shorten the
treatment regimen, thereby decreasing the cost of eFSH (SQUIRES and MCCUE
2011).
Recombinant FSH and LH
Given the problems in using EPE and eFSH, in part due to the variability of the ratio
of FSH:LH, it was hypothesized that development of recombinant equine
gonadotropins would provide pure and large quantities of eFSH from the laboratory
using molecular biology and cloning techniques (ROSER and MEYERS-BROWN
2012). Recombinant human FSH was reported to increase follicular activity in
humans, primates, rodents, and cattle (THARASANIT et al. 2006). When tested in
mares, there was no increase in ovulation rate or embryo recovery (ROSER and
MEYERS-BROWN 2012). This may have been due to the fact that the equine FSH
receptors show differences in their DNA sequence and structure compared with other
species (THARASANIT et al. 2006). But the development and efficacy of
recombinant equine gonadotropins (reFSH and reLH) have recently been reported
(JABLONKA-SHARIFF et al. 2007; JENNINGS et al. 2009; MEYERS-BROWN et al.
2010).

Introduction 15
1.4 Genital blood flow in the mare
In human medicine, color Doppler sonography has been used for more than two
decades to predict the outcome of assisted reproduction technologies (BROUSSIN
2007; LAMAZOU et al. 2009). For example, in women undergoing hormonal
treatment, transvaginal color Doppler sonography has been successfully used to
study ovarian blood flow during IVF cycles, and ovarian blood flow was found to be
related to ovarian response to stimulation (WEINER et al. 1993; ZAIDI et al. 1996).
Correlations between genital blood flow and ovarian response to hormonal treatment
have also been verified in cows (HONNENS et al. 2008; 2009). Furthermore, ovarian
blood flow has already been investigated in the mare by BOLLWEIN et al. (2002b).
Using transrectal color Doppler sonography, these authors found characteristic
changes in ovarian blood supply during the estrus cycle in mares, which were related
to alterations of sexual steroid hormone levels (BOLLWEIN et al. 2002b). HONNENS
et al. (2011) investigated the relationships between uterine blood flow, peripheral sex
steroids, expression of endometrial estrogen receptors and nitric oxide synthases
during the estrous cycle in the mare and concluded that the nitric oxide synthase
system plays a major role in regulation of uterine perfusion during the estrous cycle
in the mare. Currently there is no information about ovarian blood flow during
hormonal stimulation of superovulation in the mare.
1.4.1 Doppler sonography
In 1980, Palmer and Driancourt published the first report on the use of transrectal
ultrasound in equine gynaecology, which was rapidly followed by a widespread
utilization of ultrasound scanners for use in this area (GINTHER 1986). This
technology is used for both color-flow and power-flow imaging, and for spectrally
displaying on a viewing screen the blood velocity at a target point in a vessel
(GINTHER et al. 2007). The assessment of ovarian blood flow and ovarian structures
– topics of great interest to equine veterinarians – has received much research
interest in recent years (BOLLWEIN et al. 2002a; GASTAL et al. 2006; MIRO et al.
2010). Doppler ultrasound technology is based on Dopplershift, wherein the
ultrasound frequency of echoes from moving red blood cells is increased or
16 Introduction
decreased as the cells move toward or away from the transducer. In spectral mode,
the blood flow in a specific vessel can be assessed by placing a sample-gate cursor
on the image of the lumen of the vessel (GINTHER 2004). Arterial blood flow to the
reproductive tract is pulsatile in response to the heartbeats or pulsations of the left
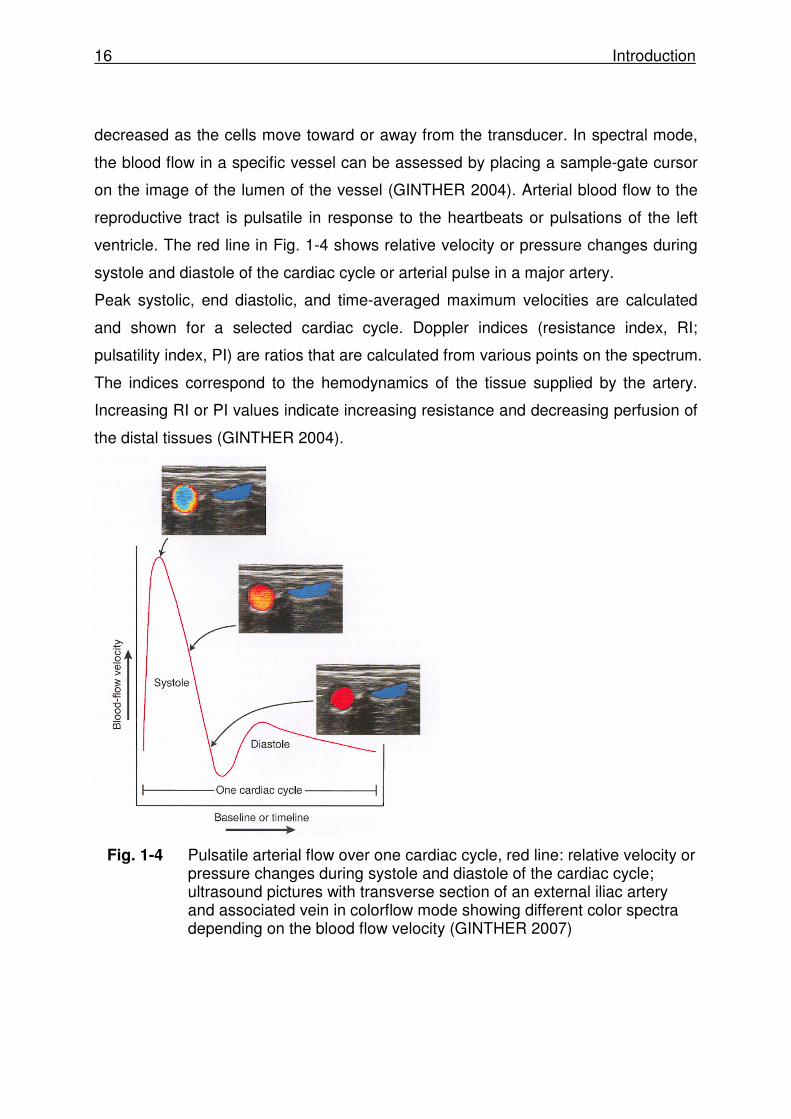
ventricle. The red line in Fig. 1-4 shows relative velocity or pressure changes during
systole and diastole of the cardiac cycle or arterial pulse in a major artery.
Peak systolic, end diastolic, and time-averaged maximum velocities are calculated
and shown for a selected cardiac cycle. Doppler indices (resistance index, RI;
pulsatility index, PI) are ratios that are calculated from various points on the spectrum.
The indices correspond to the hemodynamics of the tissue supplied by the artery.
Increasing RI or PI values indicate increasing resistance and decreasing perfusion of
the distal tissues (GINTHER 2004).
Fig. 1-4 Pulsatile arterial flow over one cardiac cycle, red line: relative velocity or pressure changes during systole and diastole of the cardiac cycle; ultrasound pictures with transverse section of an external iliac artery and associated vein in colorflow mode showing different color spectra depending on the blood flow velocity (GINTHER 2007)

Introduction 17
1.4.2 Genital blood supply in the mare
Uterine artery
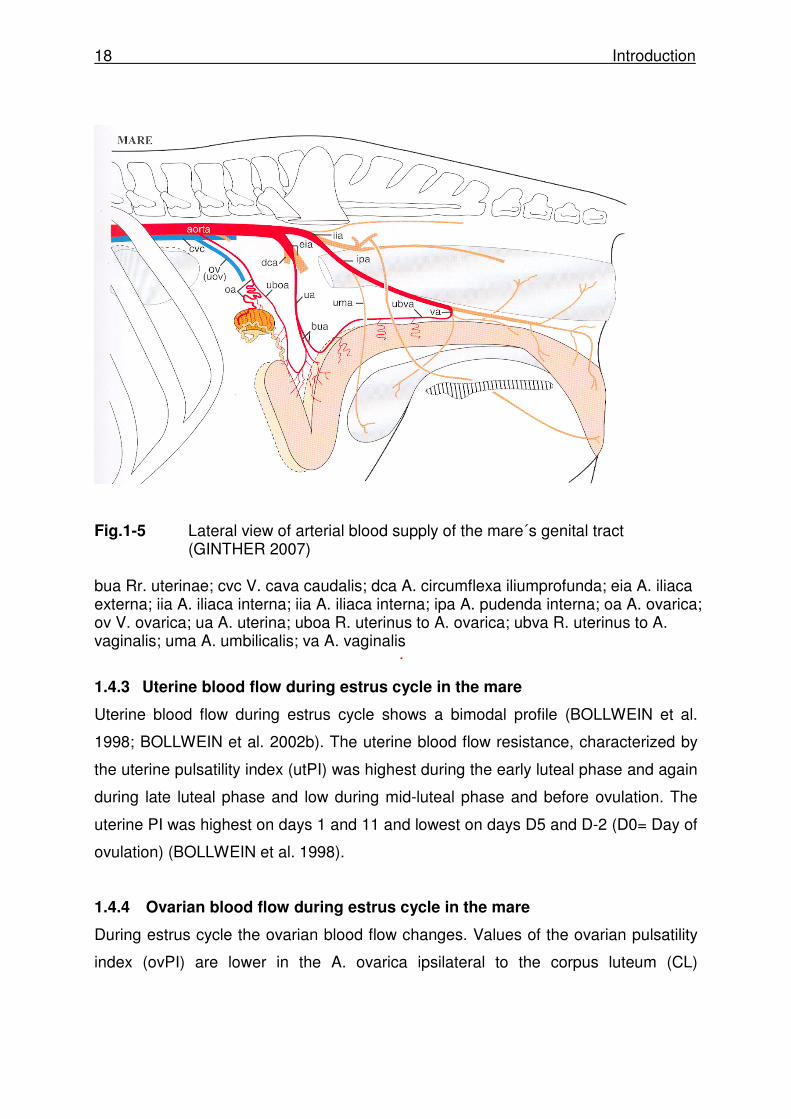
The uterus receives blood from the uterine branch of the ovarian artery, a main
supply from the uterine artery, and the uterine branch of the vaginal artery (Fig. 1-5).
The uterine artery originates from the external iliac artery in the mare. The aorta
continues as a common trunk of a few centimetres between the origins of the
external and internal iliac arteries. Following the mesometrium the uterine artery
forms a cranial and caudal branch (GINTHER 2007).
Ovarian artery
The ovarian artery leaves the aorta, as shown in Fig 1-5, runs dorsally along the
abdominal wall, and enters the mesovarium. The right artery crosses along the vena
cava ventrally. The ovarian artery passes along the cranial aspect of the
mesovarium. In mares the ovarian artery is relatively straight and located a few
centimetres caudal to the uteroovarian vein. The uterine branch of the ovarian artery
or uteroovarian anastomosis is highly variable among individuals and sides
(GINTHER 2007).
The detailed description of location of the uterine and ovarian arteries using Doppler
ultrasound are reviewed by GINTHER (2007).
18 Introduction
Fig.1-5 Lateral view of arterial blood supply of the mare´s genital tract
(GINTHER 2007) bua Rr. uterinae; cvc V. cava caudalis; dca A. circumflexa iliumprofunda; eia A. iliaca externa; iia A. iliaca interna; iia A. iliaca interna; ipa A. pudenda interna; oa A. ovarica; ov V. ovarica; ua A. uterina; uboa R. uterinus to A. ovarica; ubva R. uterinus to A. vaginalis; uma A. umbilicalis; va A. vaginalis
1.4.3 Uterine blood flow during estrus cycle in the mare
Uterine blood flow during estrus cycle shows a bimodal profile (BOLLWEIN et al.
1998; BOLLWEIN et al. 2002b). The uterine blood flow resistance, characterized by
the uterine pulsatility index (utPI) was highest during the early luteal phase and again
during late luteal phase and low during mid-luteal phase and before ovulation. The
uterine PI was highest on days 1 and 11 and lowest on days D5 and D-2 (D0= Day of
ovulation) (BOLLWEIN et al. 1998).
1.4.4 Ovarian blood flow during estrus cycle in the mare
During estrus cycle the ovarian blood flow changes. Values of the ovarian pulsatility
index (ovPI) are lower in the A. ovarica ipsilateral to the corpus luteum (CL)

Introduction 19
compared to the A. ovarica contralateral to the CL during diestrus (D0 - D15). During
D0 to D2 ovPI was highest in the A. ovarica ipsilateral to the CL, decreased until D6
and continuously increased until D15 (BOLLWEIN et al. 2002a). The PI was low on
the expected days of high progesterone concentrations and was attributable to
evaluated blood flow in the CL. During estrus (D-6 to D-1) there was a negative
correlation between the diameters of the largest follicle and the ovPI of the ipsilateral
A. ovarica (BOLLWEIN et al. 2002a).
Currently there is only limited information (PROBST 2009) about ovarian blood flow
during hormonal stimulation of superovulation in the mare.
1.5 The equine oviduct
1.5.1 Fallopian tubes
The first anatomical description of a mammalian oviduct was published by
FALLOPIO in 1561 (cited in BECK and BOOTS 1974). Each uterine tube consists of
an expansive infundibulum covering the ovary´s ovulation fossa, a highly tortuous
ampulla about 6 mm in diameter, and a less tortuous isthmus (about 3 mm in
diameter). The whole uterine tubes are 20-30 cm in length. The isthmus terminates in
a small uterine ostium on a papilla within the cranial end of a uterine horn. The
uterine ostium is about 2 - 3 mm in diameter. The inner circular muscle of the
oviductal musculature increases to form a sphincter at the utero-tubal junction. The
abdominal ostium in the centre of the infundibulum is about 6 mm in diameter. The
distal one third of the oviduct is extremely convoluted and has a well developed
Lamina muscularis (MENEZO and GUERIN 1997). The ovarian bursa of the mare is
a peritoneal pouch extending from the ovulation fossa caudal to the cranial aspect of
the uterine horn. Laterally it is bounded by the uterine tube and mesosalpinx. A fold
of broad ligament containing the proper ligament of the ovary forms the medial part of
the ovarian bursa (KAINER 1993).

20 Introduction
Fig. 1-6: Drawing of lateral view of the right ovary and associated
structures of the mare (GINTHER 1986)
amp=ampulla; inf=infundibulum; ist=isthmus; luh=left uterine horn; mo= mesovarium; ms=mesosalpinx; rl=round ligament; tm=tubal membrane; tuj=tubo-uterine junction left picture=lateral view of the right ovary and associated structures; right picture= lateral view of the right ovary and associated structures with lifted infundibulum and mesosalpinx and view of the mucosal side of the tip of the uterine horn with the tubo- uterine junction (uterine papilla) 1.5.2 Equine embryo development in the oviduct
The oviduct of the mare is the smallest component of the tubular genital tract but is
also the site of significant reproductive events - gamete transport and fertilisation. It is
considered as a reproductive organ having both transport and secretory functions
that are essential for early reproductive events. The equine embryo, in contrast with
embryos of most other domestic species, remains in the oviduct longer (FREEMAN
et al. 1991) and embryo development at the time of uterine entry is relatively
advanced in the horse versus the pig, cow or sheep (FREEMAN et al. 1991). Equine
embryos that enter the uterus are compact morulae to early blastocysts.
Following ovulation, the oocyte arrives in the ampulla of the oviduct still surrounded
by its protective coating of cumulus cells. At the ampullary-isthmic junction it lodges
Introduction 21
and, assuming mating/insemination has already taken place, the oocyte is fertilised
by one of the spermatozoa present (BOYLE et al. 1987; HUNTER 2005). The
developing embryo remains there during its subsequent cleavage divisions
(BETTERIDGE et al. 1982; WEBER et al. 1996). The developing conceptus, still
located at the ampullary-isthmic junction, contains approximately 4 blastomeres, and
the embryonic genome is activated (BETTERIDGE et al. 1982). Embryonic
development continues within the oviduct for another 4 days until the compact morula
begins to secrete PGE2 (WEBER et al. 1991) which induces relaxation of the
ampullary-isthmic ‘sphincter’ and enables the embryo to pass rapidly through the
isthmus and uterotubal junction to enter the uterine lumen at around day 6-6.5 after
ovulation (FREEMAN et al. 1991; BATTUT et al. 1997). At the time of uterine entry,
the embryo is at the late morula or early blastocyst stage of development
(BETTERIDGE et al. 1982; FREEMAN et al. 1991; BATTUT et al. 1997; RAMBAGS
et al. 2005). In contrary to embryos unfertilized eggs are retained in the oviduct (VAN
NIEKERK and GERNEKE 1966).
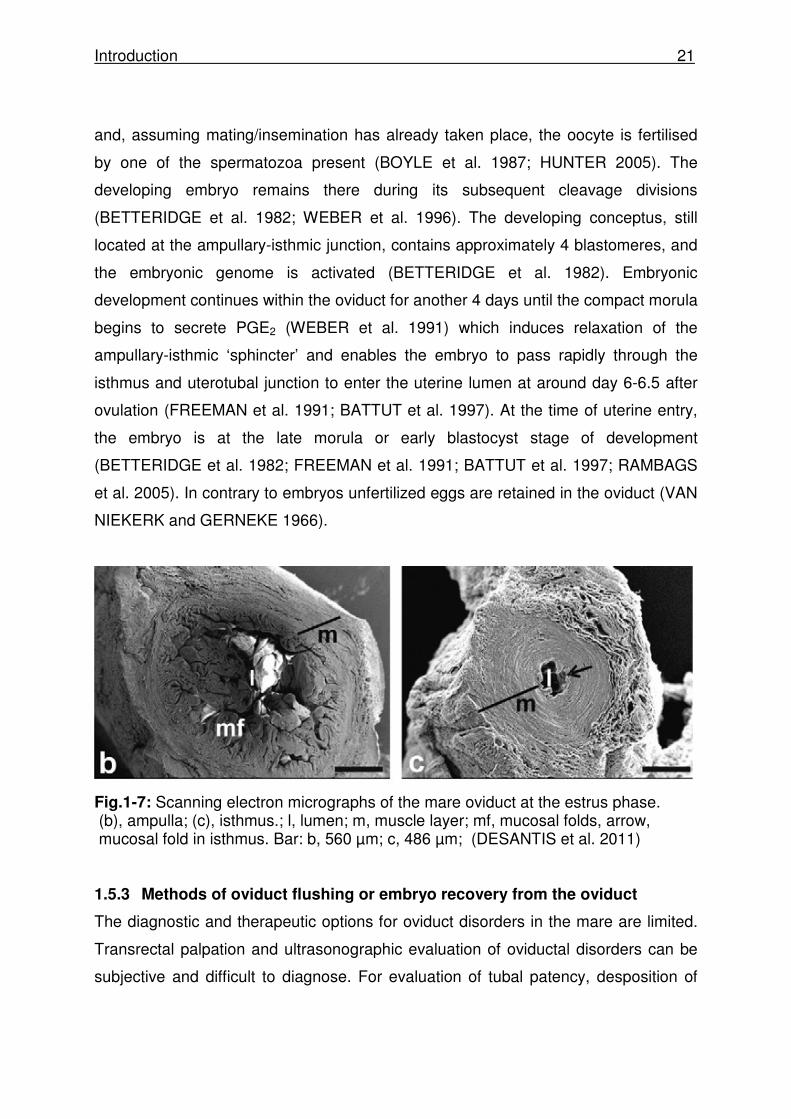
Fig.1-7: Scanning electron micrographs of the mare oviduct at the estrus phase. (b), ampulla; (c), isthmus.; l, lumen; m, muscle layer; mf, mucosal folds, arrow, mucosal fold in isthmus. Bar: b, 560 µm; c, 486 µm; (DESANTIS et al. 2011) 1.5.3 Methods of oviduct flushing or embryo recovery from the oviduct
The diagnostic and therapeutic options for oviduct disorders in the mare are limited.
Transrectal palpation and ultrasonographic evaluation of oviductal disorders can be
subjective and difficult to diagnose. For evaluation of tubal patency, desposition of

22 Introduction
fluorescent microspheres (LEY et al. 1998) and starch granules (ALLEN 1979) on the
surface of the ovary and fimbria have been described but neither test has received
wide acceptance (NEAL 2011).
A major advance in understanding oviducal function in the mare was achieved when
it was demonstrated that Day 5 equine embryos secrete significant quantities of
PGE2 (WEBER et al. 1991). This hormone binds to the oviductal musculature
(WEBER et al. 1992), and continuous infusion of small quantities of PGE2 onto the
surface of the ipsilateral oviduct in inseminated mares via a minipump surgically
implanted into the mesovarium hastens embryonic transport through the oviduct
(WEBER et al. 1991). WEBER et al. (1992) demonstrated marked inhibition by PGE2
of histamine-induced contractility of equine isthmic circular smooth muscle in vitro.
TROEDSSON et al. (2005) observed how PGE2 can also cause contraction of the
longitudinal smooth muscle of the oviduct in rabbits (BLAIR and BECK 1977) and
pigs (RODRIGUEZ-MARTINEZ et al. 1985). These important research findings on
the roles of PGE2 in oviducal transport were supported by the report that application
of a few drops of a PGE2-laced cervical gel onto the surface of the ipsilateral oviduct
of inseminated mares on Day 4 after ovulation hastened entry of the compact
morula-stage embryo into the uterus by 24 h (ROBINSON et al. 2000).
Catheterisation of the equine oviduct through the UTJ is an extremely difficult
procedure, unlike other mammalian species, since the distal third of the duct is
extremely convoluted and has a well-developed Lamina muscularis. This acts as a
sphincter (MENEZO and GUERIN 1997), making mechanical entry from the uterus
exceedingly difficult (KAINER 1993; BENNETT 2007).
Laparoscopy, via a lateral flank approach or a ventral abdominal approach under
general anaesthesia are other possibilities, allowing the upper tract to be viewed in
situ (BENNETT 2007). Zent et al. (1993) successfully restored fertility in 3 of 5
Thoroughbred mares with well documented histories of unexplained conception
failure by flushing saline through their oviducts (from infundibulum to uterine horn)
during surgical laparotomy performed under general anaesthesia.
Another method to collect tubal stage embryos was described by BESENFELDER
and BREM (1998) in the cow. They introduced a transvaginal approach to the oviduct

Introduction 23
and described the collection of tubal stage embryos via a laparoscopic guided
transvaginal oviductal flushing (BESENFELDER et al. 2001).
Currently, the best technique for diagnosis and treatment of oviductal disorders is
laparotomy and exploratory surgery under general anaesthesia and the
catheterisation of the infundibulum (BENNETT 2007), which is relatively invasive. A
simple technique which allows the accurate evaluation of oviducal function, in respect
of gamete transport, would be highly desirable in the investigation of the infertile
mare. Although such a technique remains elusive, considerable progress has been
made with the application of laparoscopic techniques for diagnosis and treatment
(NEAL 2011).
With the development of biotechnologies in the mare, the catheterisation of the
infundibulum was performed in context of oocyte transfer (OT) or gamete
intrafallopian transfer (GIFT) (CARNEVALE 2004; CARNEVALE et al. 1993;
HINRICHS et al. 2000, 2002; SCOTT 2001). Oocytes recovered from a valuable
donor mare are, after being matured in vitro, injected into the fimbria of the oviduct
which is ipsilateral to the ovary containing a maturing follicle in a recipient mare. This
recipient may have been mated or inseminated artificially with semen from the
desired sire immediately before intrafallopian transfer of the donor oocytes, or a low
number (0.5– 5 · 106) of washed spermatozoa may be injected into the oviduct
simultaneously with the M-II stage oocytes (ALLEN 2005). In both techniques the
intrafallopian transfer is performed via an invasive lateral incision and the manual
extorsion of the oviduct without using laparoscopy.
Until now, however, laparoscopic evaluation of the oviduct in the standing sedated
mare has allowed visualisation but not catheterisation and therefore had only limited
diagnostic and therapeutic potential (BENNETT 2007).
1.5.4 Laparoscopic evaluation of the Fallopian tubes in women
Laparoscopy is widely accepted as the ‘‘gold standard’’ method for evaluating tubal
patency. At present, it is considered the most accurate diagnostic test available for
evaluating tubal-related subfertility (SAUNDERS et al. 2011). Its advantages include

24 Introduction
an ability to simultaneously evaluate the abdominal cavity and other pelvic structures
for an enhanced diagnostic evaluation of other etiologies of subfertility. The
procedure also allows for therapeutic excision of endometriotic lesions and, usually,
restoration of abnormal pelvic findings. Laparoscopy incurs, however, operative risks,
costs, and a period of postoperative recovery. As an invasive and expensive
procedure, it is not an ideal first-line screening test for subfertility when suitable
alternative office procedures are available. When clinical history, laboratory, or these
office procedures suggest tubal-related pathology, laparoscopy may disclose a
definitive diagnosis and offer a treatment option (SAUNDERS et al. 2011).

Introduction 25
1.6 Aims of the study
Reasons for the study
Beside the fact that during the last 20 years the number of equine ETs performed
annually worldwide has grown enormously, success rates after superovulation and
cryopreservation of embryos in horses are still lagging behind those of cattle
(SCHERZER et al. 2008).
An ability to consistently induce multiple follicles and ovulations in estrus cycling
mares would enhance embryo recovery from donor mares, provide multiple follicles
for collection of oocytes, and improve pregnancy rates from subfertile mares
(SQUIRES 2006). The basis of superovulation is manipulation of the hormones that
control the dominant follicle and inhibit the regression of subordinate follicles
(SQUIRES and MCCUE 2007).
The most effective drug to induce multiple ovulations so far is eFSH or recombinant
equine FSH. Although results are encouraging, between-mare variability is
considerable (SQUIRES 2006). Being able to identify donor mares that respond
favourably to eFSH based on follicular development, ovulation, and embryo recovery
would be a great advantage. In human medicine, color Doppler sonography has been
used for more than two decades to predict the outcome of assisted reproduction
technologies (BROUSSIN 2007; LAMAZOU et al. 2009). Correlations between
genital blood flow and ovarian response to hormonal treatment have also been found
in cows (HONNENS et al. 2008; HONNENS et al. 2009). Although ovarian blood flow
has already been investigated in the mare by BOLLWEIN et al. (2002) using
transrectal color Doppler sonography, currently there is no information about ovarian
blood flow during hormonal stimulation of superovulation in the mare.
At present, the vast majority (>95%) of horse embryos are transferred fresh or after
chilled storage for up to 24 h, whereas cryopreservation is rarely employed (STOUT
2012). The collection of embryos from the oviducts would be a great advantage, as in
concern of freezing or vitrification acceptable pregnancy rates (>55%) are achievable
only when embryos recovered at an early developmental stage (day 6 to 6.5; morula
to early blastocyst <300 µm in diameter) are transferred (CZLONKOWSKA et al.

26 Introduction
1985; SLADE et al. 1985). Although in recent studies it was also possible to vitrify
expanded blastocysts (CHOI et al. 2011; HINRICHS and CHOI 2012) using an
embryo biopsy technique for blastocoel fluid aspiration in order to shrinken (collapse)
the embryos, this technique requires micromanipulation capabilities and is, at the
moment, no technique for the widespread clinical use.
Thus, a laparoscopic minimally invasive technique for the catheterisation of the
equine oviduct might offer the opportunity for oviductal flushing and thereby the
collection of early stage embryos.
Hypothesis
It was hypothesised that superovulation with eFSH increases ovulation and embryo
recovery rate in the mare, and affects genital blood flow as well as steroid hormone
levels. Changes in genital blood flow might serve as useful parameters in order to
predict mares that respond favourably to a superovulation treatment.
To further optimize efficiency of equine embryo transfer, we hypothesize that entering
the infundibulum and subsequent orthograde flushing of the oviduct is possible by
surgical minimal-invasive laparoscopic techniques in the standing sedated mare.
Aims of the study
Therefore the aim of the present study was to induce superovulation in mares using
eFSH® and to study the effects of stimulation on genital blood flow using color
Doppler ultrasonography and to develop a minimal invasive laparoscopic method for
flushing the oviduct in the standing sedated mare.
In the first part of the study we compared follicle development and ovulation rates in
mares (6 mares in 5 cycles) after spontaneous ovulation and superovulation with
equine pituitary extract (eFSH®) when treatment start was restricted to follicle
diameter, and compared the embryo recovery rate when AI was performed 12h and
36h after hCG application with cooled-stored semen of a fertile stallion (Chapter 2).
Next, it was assessed if uterine and ovarian blood flow in mares during this
superovulation program differs from untreated controls using transrectal Doppler

Introduction 27
sonography. We further investigated if there were relationships between genital blood
flow, steroid hormone levels, and ovarian response (Chapter 3).
In Chapter 4 the development of a minimal invasive laparoscopic technique will be
described in two experiments: The first involved a transvaginal laparoscopic
approach (n=8), the second a laparoscopic flank approach (n=12). Passage of fluid
into the uterus was visualized by post operative hysteroscopy.

Embryo recovery rate following superovulation in mares 28
Chapter 2:
Embryo recovery rate following superovulation with equine pituitary extract
(eFSH®) in mares
Embryogewinnungsrate nach Superovulation mit equinem Hypophysenextrakt (eFSH®) bei der Stute
Melanie Köllmann, Jeanette Probst, Christine Baackmann, Jutta Klewitz,
Edward S. Squires1, Harald Sieme
Klinik für Pferde und Reproduktionsmedizinische Einheit der Kliniken der Stiftung
Tierärztliche Hochschule Hannover, 1Animal Reproduction and Biotechnology
Laboratory, Colorado State University, Fort Collins, USA
Pferdeheilkunde 24 (2008) 3 (Mai/Juni) 397-405
http://www.hippiatrika.com/download.htm?id=20080310

Embryo recovery rate following superovulation in mares 29
The extent of Melanie Witt´s (formerly M. Köllmann) contribution to the article is
evaluated according to the following scale:
A. has contributed to collaboration (0-33%)
B. has contributed significantly (34-66%)
C. has essentially performed this study independently (67-100%)
1. Design of the project including design of individual experiments: C
2. Performing of the experimental part of the study: B
3. Analysis of the experiments: C
4. Presentation and discussion of the study in article form: C

30 Embryo recovery rate following superovulation in mares
2.1 Abstract
Embryo recovery rate following superovulation with equine pituitary extract
(eFSH®) in mares.
Embryo recovery from single ovulating mares is approximately 50% per estrus cycle,
leading to a non-economical state of embryo transfer in the mare. An ability to
consistently induce multiple follicles and ovulations in estrus cycling mares would
enhance embryo recovery from donor mares, provide multiple follicles for collection
of oocytes, and improve pregnancy rates from subfertile mares. There have been
numerous approaches to superovulation of the mare. Injections of porcine FSH,
inhibin vaccines, equine chorionic gonadotropin (eCG) and GnRH have been of
limited success in stimulating multiple ovulations in the mare. Numerous studies have
shown that injection of equine pituitary extract (EPE) will result in three to four
ovulations per estrus cycle and two embryos. Recently, a commercial purified equine
pituitary extract product (eFSH®) has been available. In the present study six
normally cycling mares were investigated over five cycles and ovulation rate and
embryo recovery rate were compared between control cycles and stimulated cycles.
Cycle one and three were designed as control cycles without stimulation and
insemination. In cycles 2 and 4 mares were treated with 12.5 mg eFSH®
intramuscularly twice daily beginning when the diameter of the largest follicle was 20
to 25 mm. Prostaglandin was administered on the second day of eFSH® therapy.
Treatment with eFSH® was continued until follicle(s) were 32-35 mm in diameter. The
mares were subsequently allowed to ‘coast’ for 36 h, after which 2500 IU human
chorionic gonadotropin were administered to induce ovulation. Mares were
inseminated with 750 Mio. progressive motile sperms of a fertile warmblood stallion.
Embryo recovery was performed 6.5 days following ovulation. In the last cycle (5)
mares were treated and inseminated in the same way as in cycle 2 and 4, but without
eFSH® stimulation. Ovulation rate in control cycles was lower (1.3 ovulations) than in
eFSH® treated cycles (4.4 ovulations). The number of days of eFSH® treatment
required for reaching a follicle size of 32-35 mm was on average 4.0 days. Embryo
recovery rate in control mares was 1.2 per cycle, whereas in eFSH® treated mares ø

Embryo recovery rate following superovulation in mares 31
2.9 embryos could be flushed. The eFSH® protocol used in this study was efficient to
induce multiple ovulations and increase embryo recovery rate in mares. Albeit the
number of embryos obtained is quite encouraging, individual mare variation is
considerable. Being able to identify donor mares that respond favourably to eFSH
based on follicular development, ovulation and embryo recovery would be a great
advantage. In current studies a possible influence of follicle development and genital
blood flow on embryo recovery rate is investigated to identify „good donor mares“ in a
superovulation program.
Keywords: mare, reproduction, embryo transfer, superovulation, eFSH

32 Doppler sonography of the uterine and ovarian arteries during superovulation
Chapter 3
Doppler sonography of the uterine and ovarian arteries
during a superovulatory program in horses
M. C. Witt, H. Bollweina, J. Probst, C. Baackmann, E.L. Squiresb, H. Sieme
Clinic for Horses and Unit for Reproductive Medicine, aClinic for Cattle, University of
Veterinary Medicine Hanover Foundation, Buenteweg 9, 30559 Hanover, Germany;
bAnimal Reproduction and Biotechnology Laboratory, Colorado State University, Fort
Collins, Colorado, USA
Theriogenology 77 (2012) 1406–1414
doi:10.1016/j.theriogenology.2011.11.005

Doppler sonography of the uterine and ovarian arteries during superovulation 33

34 Doppler sonography of the uterine and ovarian arteries during superovulation
The extent of Melanie Witt´s contribution to the article is evaluated according to the
following scale:
A. has contributed to collaboration (0-33%)
B. has contributed significantly (34-66%)
C. has essentially performed this study independently (67-100%)
1. Design of the project including design of individual experiments: C
2. Performing of the experimental part of the study: B
3. Analysis of the experiments: B
4. Presentation and discussion of the study in article form: C

Doppler sonography of the uterine and ovarian arteries during superovulation 35
3.1 Abstract
Doppler sonography of the uterine and ovarian arteries during a
superovulatory program in horses
The aim of the present study was to investigate the effects of a gonadotropin
treatment to induce superovulation on ovarian and uterine blood flow and its
relationship with steroid hormone levels and ovarian response in mares, using color
Doppler sonography. Mares were examined sonographically in five consecutive
cycles for three days (t1 to t3) each during the follicular development phase (FDP)
beginning at a follicle size of ≥22 mm, and for four days (D-4 to D-1; D0 = Ovulation)
in the preovulatory phase (POP). After each examination, total estrogens (Etot) and
progesterone (P4) levels were determined in peripheral plasma. Cycles 1, 3, and 5
(c1, c3, c5) were unstimulated cycles (USC); in c2 and c4, the mares were stimulated
(SC) with eFSH and inseminated when in estrus at 12 and 24 h after hCG
administration. Embryo recovery was performed 6.5 days post ovulation. Cycle 5 (c5)
was an unstimulated cycle with hCG treatment, insemination, and embryo recovery.
Ovarian and uterine blood flow was quantified by the blood flow volume (BFV) and
the pulsatility index (PI) in ovarian and uterine arteries. The mean number of
ovulations and developing CL was 1.3 + 0.4 in USC and 4.4 + 3.1 in SC with no
difference (p≥0.05) between the ovaries within mares. No difference (p>0.05) was
observed in utBFV and utPI during FDP between USC and SC, but during POP, utPI
was lower (p<0.05) and utBFV higher (p<0.001) in SC than USC. The ovBFV was
higher (p<0.01) and ovPI lower (p<0.05) in SC compared to USC. All uterine and
ovarian blood flow parameters were related to the number of developing follicles in
SC. Parameters utPI (r=-0.67;p<0.001) and ovPI (r=-0.53; p<0.001) were negatively
correlated with the number of ovulations on t3, and with the number of collected
embryos on t3 (utPI:r=-0.81; p<0.001), D-4 (utPI:r=-0.64; p<0.0001), and D-1
(ovPI:r=-0.41; p<0.01). P4 levels were not positively correlated with utBFV (p>0.05),
but Etot concentrations (D-4: r=0.790; D-3: r=0.639; p<0.001; D-1: r=0.48; p<0.001)
and ovBFV from D-4 to D-1 (r= 0.64; p<0.001) in SC were. The results of the present
study show that in mares treatment with gonadotropins to induce superovulation is

36 Doppler sonography of the uterine and ovarian arteries during superovulation
associated with a marked increase in uterine and ovarian perfusion, concurrent with
the development of multiple follicles and an increase in Etot levels. The increased
blood flow seems to be related to the effectiveness of ovarian response to stimulation.

37 Laparoscopic techniques for investigating the equine oviduct
Chapter 4
Laparoscopic techniques for investigating the equine oviduct
M. Köllmann, A. Rötting, A. Heberling, H. Sieme
Clinic for Horses and Unit for Reproductive Medicine,
University of Veterinary Medicine Hannover Foundation
Equine Vet. J.
(2011) 43 (1) 106-111
doi: 10.1111/j.2042-3306.2010.00143.x

Laparoscopic techniques for investigating the equine oviduct 38
The extent of Melanie Witt´s (formerly M. Köllmann) contribution to the article is
evaluated according to the following scale:
A. has contributed to collaboration (0-33%)
B. has contributed significantly (34-66%)
C. has essentially performed this study independently (67-100%)
1. Design of the project including design of individual experiments: C
2. Performing of the experimental part of the study: C
3. Analysis of the experiments: C
4. Presentation and discussion of the study in article form: C

39 Laparoscopic techniques for investigating the equine oviduct
4.1 Abstract
Laparoscopic techniques for investigating the equine oviduct
Reasons for performing study: The diagnostic and therapeutic options for oviduct
disorders in the mare are limited. The current best techniques require exploratory
surgery under general anaesthesia or flank laparotomy.
Hypothesis: The orthograde flushing of the oviduct for diagnostic or therapeutic
options is possible using laparoscopic techniques in the standing sedated mare.
Methods: Development of a laparoscopic technique for catheterization of the
infundibulum and flushing of the oviduct (sterile methylene blue solution) in the
standing sedated mare was examined in two experiments. The first involved a
transvaginal laparoscopic approach, the second a laparoscopic flank approach.
Passage of fluid into the uterus was assessed by postoperative hysteroscopy.
Results: In experiment I, visualisation of the infundibulum was possible (left side 7/8
cases, right side in 6/8 cases). The beginning of the oviductal ampulla could be seen
in 3 of 8 cases on the left side. An adequate opening of the infundibulum and
visualisation or catheterisation of the abdominal ostium were not possible. In
experiment II, catheterisation of the ampulla was successful in 7 of 11 cases, and in
5 of these 7 cases the injected fluid could be identified in the uterus by postoperative
hysteroscopy.
Conclusion: A transvaginal laparoscopic approach to the oviduct is not appropriate
for oviductal flushing in the mare. However, a laparoscopic flank-approach permits
investigation and flushing of the oviduct.
Potential relevance: Laparoscopic flushing could become a practical method for
diagnosis and therapy of oviduct disorders and a minimally invasive technique for
collection of young embryos or the transfer of gametes (GIFT).

Discussion 40
Chapter 5
Discussion

41 Discussion
5 Discussion
During the last 20 years the number of equine ET´s performed annually worldwide
has grown enormously (International Embryo Transfer Society, IETS). Mean embryo
recovery per cycle from single ovulation mares is approximately 50%. An ability to
consistently induce multiple follicles and ovulations in estrus cycling mares would
enhance embryo recovery from donor mares (SQUIRES 2006). However, success
rates after superovulation and cryopreservation of embryos in horses are still lagging
behind those of cattle (SCHERZER et al. 2008).
At present, the vast majority (>95%) of horse embryos are transferred fresh or after
chilled storage for up to 24 h, whereas cryopreservation is rarely employed. Freezing
and vitrification of embryos <300 µm has been effective (SLADE et al. 1985;
ELDRIDGE-PANUSKA et al. 2005), but at the moment it requires recovery of
embryos on Day 6 after ovulation in clinical praxis.
Therefore the aim of the present study was to induce superovulation in mares using
eFSH® and to study the effects of stimulation on genital blood flow using color
Doppler ultrasonography and to develop a minimal invasive laparoscopic method for
entering the infundibulum and subsequent orthograde flushing of the oviduct in the
standing sedated mare.
5.1 Superovulation
The eFSH® protocol used in this study was efficient to induce multiple ovulations in
mares. The mean number of ovulations and developing CL was 1.3 + 0.4 in
unstimulated cycles (USC) and 4.4 + 3.1 in stimulated cycles (SC) with no
differences (p ≥ 0.05) between the left and right ovaries of both sides. The protocol
was further efficient to increase embryo recovery rate in mares similar to the studys
of SQUIRES and MCUE (2007), LOGAN et al. (2007) and RAZ et al. (2009a). The
total embryo recovery rate was higher (p<0.05) in stimulated cycles (2.9 ± 1.7)
compared to control cycles (1.2 + 0.4).

Discussion 42
But embryo recovery rates per ovulation were lower in SC compared to c5
(unstimulated cycle with hCG treatment, insemination, and embryo recovery) cycles
(66% vs. 87.5%). Several earlier studies noted that with increased number of
ovulations, embryo recovery per ovulation remained the same or decreased,
following repeated injections of either EPE or eFSH (ROSAS et al. 1998;
ALVARENGA et al. 2001; SCOGGIN et al. 2002; SQUIRES 2006; WELCH et al.
2006; LOGAN et al. 2007; ARAUJO et al. 2009; RAZ et al. 2009b). Several factors
can affect the embryo/ovulation ratio, such as oocyte quality (GEARY et al. 1989;
WOODS 1998; CARNEVALE et al. 1999) and the capability of the oocyte to enter the
oviduct via the ovulation fossa (CARMO et al. 2006). According to CARMO et al.
(2006), there was no significant difference in the number of oocytes that appeared in
the oviduct from superovulated mares compared to control mares supporting the
concept that oocyte development and maturation may be a critical factor affecting the
embryo per ovulation ratio in superovulated mares (MEYERS-BROWN et al. 2011).
The findings that an increased number of anovulatory follicles are present after
superovulation regimens (ALVARENGA et al. 2001; SQUIRES 2006; WELCH et al.
2006; RAZ et al. 2009b) can also support the concept that a less than optimum
embryo/ovulation ratio may be a function of oocyte dysfunction.
The between-mare variability in the present study was considerable, as it was
observed in earlier eFSH studies (SQUIRES 2006). This suggests that no one
treatment will work in a similar fashion in all the mares. To further work on the
problems of superovulation in the mare, ROSER et al. (2012) considered if follicular
waves in individual mares need to be evaluated before treatment. They further
assumed that mares then can be subjected to different protocols. Furthermore, they
considered if endogenous levels of FSH, LH, estradiol, inhibin, activin, IGF-I, and
perhaps other hormones or paracrine/autocrine factors in individual mares need to be
measured during the cycle before treatment so as to control the response to
treatment at any one time.
However, understanding the physiology and endocrinology of each reproductive
stage of the mare, will be the key in determining the optimum treatment regimen for

43 Discussion
mares in general. Each mare will have to be approached with the idea in mind that
she may need a different combination of drugs to initiate the desired response.
Beside these considerations, the development and efficacy of recombinant equine
gonadotropins (reFSH and reLH) have recently been reported (JABLONKA-SHARIFF
et al. 2007; JENNINGS et al. 2009; MEYERS-BROWN et al. 2010). The single chain
recombinant products are not contaminated with other hormones or pathogens and
are biologically active in stimulating follicular development and ovulation in the mare.
So more conststant superovulation results may be expected with recombinant
products in future.
But nevertheless, being able to identify donor mares that respond favorably to a
superovulation treatment before the hormone therapy, like demonstrated in the cow
(HONNENS et al. 2008, 2009) or in women, (WEINER et al. 1993; ZAIDI et al. 1996)
would be a great advantage.
In human medicine, color Doppler sonography has been used for more than two
decades to predict the outcome of assisted reproduction technologies (BROUSSIN
2007; LAMAZOU et al. 2009). For example, in women undergoing hormonal
treatment, transvaginal color Doppler sonography has been successfully used to
study ovarian blood flow during IVF cycles, and it was found to be related to ovarian
response to stimulation (WEINER et al. 1993; ZAIDI et al. 1996). Correlations
between genital blood flow and ovarian response to hormonal treatment have also
been found in cows (HONNENS et al. 2008, 2009). Therefore, the aim of this study
was to investigate and compare uterine and ovarian blood flow in mares during a
superovulation program with untreated controls using transrectal Doppler
sonography, and to investigate if there are relationships between genital blood flow,
steroid hormone levels, and ovarian response.
5.2 Doppler Ultrasonography
The hormonal superovulatory treatment with eFSH led to an increase in uterine and
ovarian blood flow and estradiol levels concurrent with the development of multiple
follicles.

Discussion 44
The results of the present study show that mares treatment with gonadotropins to
induce superovulation is associated with a marked increase in uterine and ovarian
perfusion, concurrent with the development of multiple follicles and an increase in Etot
levels. The increased blood flow seems to be related to the effectiveness of ovarian
response to stimulation. The marked increase in ovarian blood flow volume and the
decrease in ovarian PI in SC in the present study were also reported in cows
undergoing hormonal treatment to induce superovulation (HONNENS et al. 2009).
In the present study, we did not detect any differences in uterine or ovarian blood
flow in the presence of one or two follicles, but with increasing numbers of follicles,
the uterine/ovarian BFV increased and uterine/ovarian PI decreased in eFSH cycles.
These findings correspond to a study in human medicine where the uterine PI was
correlated with the number of follicles and collected oocytes in IVF patients
(CACCIATORE et al. 1996).
In the present study, correlations were observed between the uterine and ovarian
BFV and the number of further ovulations in SC. Both parameters were correlated in
the preovulatory phase but also in the follicle developing phase (t2, t3) so that the
BFV might be a parameter that is useful for predicting ovarian response to
superovulatory program in mares.
In contrary to a study in cows, where no significant correlations between the ovBFV,
ovPI, and the number of follicles that developed in response to an eCG hormonal
treatment were detected (HONNENS et al. 2009) we found relationships between the
ovarian BFV (t3, D-4 to D-1), ovPI (t3, D-1) as well as the uterine BFV and utPI and
the number of developing follicles.
Several studies have shown that the superovulatory response in cattle is positively
related to the pools of primordial and growing follicles in the bovine ovary
(MONNIAUX et al. 1983; SINGH et al. 2004). The superstimulatory response can be
predicted by the number of follicles ≥2 mm at wave emergence. The number of
follicles ≥5 mm after ovarian superstimulation can be expected to be approximately
71% of the number of follicles ≥2 mm at the time of wave. A study with color Doppler
ultrasonography revealed that small follicles with blood flow become larger around
the follicle selection period compared with small follicles without blood flow (ACOSTA

45 Discussion
et al. 2005). Since blood supply to individual follicles will provide gonadotropins and
nutrients that support follicular growth, follicle cohorts which include many follicles
with detectable blood flow will show a good response to superovulatory treatment.
Therefore, examination of the number of small follicles which have detectable blood
flow at the start of gonadotrophin treatment may be a useful index as a predictor of
superovulatory response.
In the present study, there were no correlations between the ovarian blood flow
parameters during t1 to t3 and D-4 to D-1 and the embryo recovery rate, which was
found before in studies in cows (HONNENS et al. 2009). But the uterine PI was
correlated with the number of collected embryos on t3 and D-4. Therefore the color
Doppler sonography of the uterine arteries might be useful in predicting the success
of a superovulatory regime in the mares as it has been used in women in IVF
treatments (CACCIATORE et al. 1996; OZTURK et al. 2004). Nevertheless this
correlation should be verified in further studies.
5.3 Laparoscopy of the equine oviduct
Currently, the best techniques for diagnosis and therapy of tubal disorders are
exploratory surgery under general anaesthesia or lateral flank laparotomy with
flushing of the oviduct (BENNETT 2007).
In the present study a minimal invasive laparoscopic technique for oviductal flushing
was successfully established. With the lateral flank approach, an excellent
visualisation of the ovary, infundibulum and the whole oviduct from the side was
possible in all cases. After “opening” the infundibulum, the abdominal ostium could
be seen in 8 cases (72.7%). Catheterisation of the ampulla was successful in 7 of 11
cases (63.6%), and in 5 of these 7 cases the injected fluid could be identified in the
uterine lumen by postoperative hysteroscopy. The laparoscopic flushing of the
oviduct is possible with the lateral flank approach and might be used for the collection
of tubal stage embryos in future.
Laparoscopic techniques for a variety of procedures in horses have been described
in recent years (COLBERN and REAGAN 1987; HANSON and GALUPPO 1999;

Discussion 46
WALMSLEY 1999). Standing female urogenital endoscopic surgery is facilitated by
the more dorsal location of the organs of the female reproduction tract. The most
common reason for laparoscopic surgery on the female urogenital system is
ovariectomy; however, the technique has been used to diagnose periparturient or
reproductive diseases and to perform surgical embryo transfer. Standing surgical
approaches avoid the risk and expense of general anesthesia, but these techniques
are limited by the temperament and size of the patient and the availability of facilities
for restraint (DECHANT and HENDRICKSON 2000). The reported advantages
include increased intraoperative visualisation, the use of smaller incisions, less
extensive manipulation of the abdominal viscera, reduced post operative pain and
reduced periods of convalescence compared with corresponding procedures
performed using conventional methods (HANSON and GALUPPO 1999;
WALMSLEY 1999). The disadvantages of laparoscopy might be the financial
investment associated with the equipment and consumables and the relative
technical difficulty of certain procedures. We assume that the procedure has no
negative influence on fertility because all mares were used for further research and
ET programmes and continued to have good fertility. Negative effects might be
inflammation or adhesions of the fimbria or adhesions in the oviduct which might lead
to an obstruction.
Earlier, the laparoscopically guided administration of PGE2-laced triacetin gel onto
the surface of the oviducts was used to hasten embryonic transport (ROBINSON et
al. 2000) and as a therapeutic option in mares that were presumed to have tubal
obstructions (ALLEN et al. 2006).
The transvaginal approach used in Experiment I was unlike the situation in cattle
(BESENFELDER et al. 2001) inadequate for an investigation or catheterisation of the
oviduct. The visualisation of the medial side of both ovaries was possible and, after
turning the ovary to the mid side, visualisation of the infundibulum was possible in 7
of 8 cases (87.5%) on the left side and in 6 of 8 cases (75.0%) on the right side. The
beginning of the oviductal ampulla could be seen in only 3 of 8 cases (37.5%) on the
left side. An adequate opening of the infundibulum and thereby a visualisation or
catheterisation of the abdominal ostium was not possible with the transvaginal

47 Discussion
laparoscopic approach. Due to the anatomical characteristics of the equine ovary and
infundibulum, catheterisation of the abdominal ostium is only possible following the
180° turn of the ovary in cranio-medial direction and a lifting of the infundibulum
which was not possible using this approach. Nevertheless, the technique used in
Experiment I could be of benefit for the application of a PGE-gel onto the outer
surface of the ovary and ampulla because both sites could be reached in one session.
The techniques of natural orifice transluminal endoscopic surgery (NOTES) using
laparoscopic and endoscopic instrumentation transvaginally into the mare’s abdomen
is seemingly well tolerated and safe and provides adequate observation of most
structures within the dorsal caudal region of the abdomen on the side of endoscope
or laparoscope insertion (ALFORD and HANSON 2010). The transvaginal approach,
to the equine abdomen is not novel. Transvaginal ovariectomy, by colpotomy, is well
established in mares (WALKER and VAUGHAN 1980; COLBERN and REAGAN
1987; EMBERTSON 2006).
The transvaginal laparoscopic approach to the oviduct is not appropriate for oviductal
flushing in the mare. However, a laparoscopic flank-approach permits investigation
and flushing of the oviduct. Laparoscopic flushing could become a practical method
for the diagnosis and therapy of oviduct disorders.
Recently modification of vitrification methods allowed the establishment of a
vitrification technique that supports normal pregnancy rates for expanded blastocysts
up to 650 µm in diameter. This technique uses blastocyst collapse, vitrification
medium containing ethylene glycol and galactose, in combination with warming
medium containing sucrose, and a low-volume vitrification device (HINRICHS and
CHOI 2012). Vitrification of expanded equine blastocysts might be one method to
make the embryo transfer management of equine reproduction more efficient. On the
other hand the laparoscopic technique used in this study has the further potential to
function as a minimally invasive technique for collection of young embryos <300 µm,
which are then available for cryopreservation. This would give potential for simplifying
recipient mare management and facilitating long-term storage and international
transport of embryos.

Discussion 48
The results of the present study show that in mares treatment with gonadotropins to
induce superovulation is associated with a marked increase in uterine and ovarian
perfusion, concurrent with the development of multiple follicles and an increase in Etot
levels. The uterine PI was correlated with the number of collected embryos on t3 and
D-4. The transrectal color Doppler sonography of the uterine arteries might be useful
in predicting the success of a superovulatory regime in the mares if this correlation
can be verified in further studies. The laparoscopic flank-approach permits
investigation and flushing of the oviduct and could become a practical method for
diagnosis and therapy of oviduct disorders and a minimally invasive technique for
collection of young embryos or the transfer of gametes (GIFT).

Summary 49
Chapter 6
Summary

50 Summary
6 Summary Melanie Witt - Investigations on genital blood flow and embryo recovery after
superovulation with eFSH® and on laparoscopic techniques for flushing the oviduct in
the mare.
The aim of the present study was to investigate the effects of a gonadotropin
treatment to induce superovulation on ovarian and uterine blood flow and its
relationship with steroid hormone levels and ovarian response in mares, using color
Doppler sonography and to develop a minimal invasive laparoscopic method for
flushing the oviduct in the standing sedated mare.
Methods: In the present study six mares were investigated over five cycles and
ovulation rate and embryo recovery rate were compared between unstimulated
cycles (USC= c1, c3, c5) and stimulated cycles (SC= c2, c4). Cycle 5 (c5) was an
unstimulated cycle with hCG treatment, insemination, and embryo recovery. The
mares were examined by transrectal color Dopler sonography for three days (t1 to t3)
during the follicular development phase (FDP) beginning at a follicle size of ≥22 mm,
and for four days (D-4 to D-1; D0 = Ovulation) in the preovulatory phase (POP).
Ovarian and uterine blood flow was quantified by the blood flow volume (BFV) and
the pulsatility index (PI) in ovarian and uterine arteries. Total estrogens (Etot) and
progesterone (P4) levels were determined in peripheral plasma each day. In c2 and
c4, the mares were stimulated with 12.5 mg eFSH® (Bioniche-Animal Health, Athens,
GA, USA) im twice daily beginning when the diameter of follicles was 20 to 25 mm.
Prostaglandin F2α was administered on the second day of eFSH® therapy. The eFSH
treatment was continued until follicle(s) were 32-35 mm in diameter. Following a 36 h
lasting coasting period 2500 IU hCG (Ovogest®, Intervet, Unterschleißheim) was
administered iv to induce ovulation. Mares were inseminated with cooled-stored
semen (750 Mill. progressively motile spermatozoa) 12 and 36 h post hCG. Embryo
recovery was performed 6.5 days after ovulation (D0).
The development of a laparoscopic technique for catheterisation of the infundibulum
and flushing of the oviduct in the standing sedated mare was examined using 10
mares in two different experiments. The first involved a transvaginal laparoscopic

Summary 51
approach (n=8), the second a laparoscopic flank approach (n=12). Passage of fluid
into the uterus was assessed by post operative hysteroscopy.
Results: The mean number of ovulations and developing CL was 1.3 + 0.4 in USC
and 4.4 + 3.1 in SC. The mean number of collected embryos was 1.2 vs 2.9. No
difference (p>0.05) was observed in utBFV and utPI during FDP between USC and
SC, but during POP, utPI was lower (p<0.05) and utBFV higher (p<0.001) in SC than
USC. The ovBFV was higher (p<0.01) and ovPI lower (p<0.05) in SC compared to
USC. All uterine and ovarian blood flow parameters were related to the number of
developing follicles in SC. Parameters utPI (r=-0.67;p<0.001) and ovPI (r=-0.53;
p<0.001) were negatively correlated with the number of ovulations on t3, and with the
number of collected embryos on t3 (utPI:r=-0.81; p<0.001), D-4 (utPI:r=-0.64;
p<0.0001), and D-1 (ovPI:r=-0.41; p<0.01).
By transvaginal approach (Experiment I) visualisation of the infundibulum was
possible (left side 7/8 cases, right side in 6/8 cases). Access to the oviductal ampulla
could be achieved in 3 of 8 cases on the left side. Stretching of the infundibulum and
visualisation or catheterisation of the abdominal ostium was not possible. The
laparoscopic approach (Experiment II) enables catheterisation of the ampulla (7/11),
and in 5/7 the injected fluid could be visualized in the uterus by postoperative
hysteroscopy.
Potential relevance: The results of the present study show that in mares treatment
with gonadotropins to induce superovulation is associated with a marked increase in
uterine and ovarian perfusion, concurrent with the development of multiple follicles
and an increase in Etot levels. The uterine PI on t3 and D-4 was correlated with the
number of collected embryos. Transrectal color Doppler sonography of the uterine
arteries might be useful in predicting the success of a superovulatory regime in the
mare, however, this correlation needs to be verified by further studies. The
laparoscopic flank-approach permits visualization, entering and orthograde flushing
of the oviduct. This procedure bears the potential to aid in diagnosis and therapy of
oviductal disorders and represents a minimal invasive technique in the standing
sedated mare for collection of early stage embryos as well as intraoviductal transfer
of gametes (GIFT).

52 Zusammenfassung
Chapter 7
Zusammenfassung

Zusammenfassung 53
7 Zusammenfassung
Melanie Witt - Investigations on genital blood flow and embryo recovery after
superovulation with eFSH® and on laparoscopic techniques for flushing the oviduct in
the mare.
Ziel der Arbeit war es Zusammenhänge zwischen der ovariellen Stimulationsantwort
nach einer Superovulationsbehandlung bei Stuten und dem mittels transrektaler
Dopplersonographie gemessenen genitalen Blutfluss sowie den
Steroidhormonwerten im Blut zu untersuchen. Des Weiteren war es Ziel dieser
Studie eine minimal-invasive laparoskopische Technik zur Katheterisierung des
Infundibulums und Spülung des Eileiters an der stehenden, sedierten Stute zu
entwickeln.
Methode: Sechs Stuten wurden über 5 Zyklen untersucht und die Anzahl der
Ovulationen und gewonnenen Embryonen aus den stimulierten Zyklen mit denen der
Kontrollzyklen verglichen. Zyklus 1 und 3 wurden als Kontrollzyklen ohne Stimulation
und Besamung definiert. Im zweiten und vierten Zyklus wurden die Stuten jeweils ab
einer Follikelgröße von 22-25 mm mit einer zweimal täglichen Gabe von 12,5 mg
eFSH® (Fa. Bioniche-Animalhealth, Athens, GA, USA) bis zu einer durchschnittlichen
Follikelgröße von 32-35 mm stimuliert. Am zweiten Tag der eFSH® Behandlung
wurden 250 μg Cloprostenol (Estrumate®) intramuskulär injiziert. Nach einer 36
Stunden dauernden coasting“-Periode wurde 2500 IE humanes Choriogonadotropin
(Ovogest® Intervet, Unterschleißheim) i.v. appliziert und die Stuten 12 und 36
Stunden später mit gekühltem Frischsperma (750 Mio. progressiv motile Spermien)
inseminiert. Die Embryonengewinnung erfolgte an Tag 6,5 nach Ovulation durch
Uterusspülung. Im fünften Zyklus wurde eine Embryonengewinnung nach
Ovulationsinduktion und zweimaliger Besamung wie in den Zyklen 2 und 4, jedoch
ohne vorherige Stimulation durchgeführt. Zusätzlich wurden der uterine und ovarielle
Blutfluss täglich ab einer Follikelgröße von 22 mm bis zur Ovulation hin mittels
transrektaler Dopplersonographie dokumentiert und mithilfe der Parameter Pulsatility
Index (PI) und Blutflussvolumen (BFV) definiert. Die Entwicklung einer
laparoskopischen Technik zur Katheterisierung des Infundibulums und der Spülung

54 Zusammenfassung
des Eileiters erfolgte an 10 Stuten in zwei Experimenten. Im Ersten wurde ein
transvaginaler Zugang (n=8), im Zweiten ein lateraler Flankenzugang (n=12) gewählt.
Die Flüssigkeitspassage in den Uterus wurde mittels Hysteroskopie nachgewiesen.
Ergebnisse: Im Rahmen der in dieser Studie durchgeführten
Superovulationsbehandlung mit eFSH war die durchschnittlichen Ovulationen in den
Kontrollzyklen und stimulierten Zyklen (1,3 versus 4,4 OV), die
Embryonengewinnungsrate 1,2 vs 2,9 Embryonen. Mittels Dopplersonographie
konnte gezeigt werden, dass es im Rahmen einer Superovulationsbehandlung zu
einem gesteigerten uterinen und ovariellen Blutfluss kommt, der mit der Entwicklung
multipler Follikel und einer Erhöhung des Östradiolwertes im Blut einhergeht. Die
utPI (r=-0,67; p<0,001) und ovPI (r=-0,53; p<0,001) waren an t3 negativ mit der
Anzahl an OV korreliert sowie mit der Anzahl an gewonnenen Embryonen an t3
(utPI:r=-0,81; p<0,001), D-4 (utPI:r=-0,64; p<0,0001), and D-1 (ovPI:r=-0,41; p<0,01).
Der transvaginale Zugang ermöglichte eine Visualisierung der medialen Seite beider
Ovarien und nach Drehung des Ovars nach median konnte der Beginn der Ampulla
in 3/8 (37.5%) Fällen am linken Ovar gesehen werden. Eine Öffnung des
Infundibulums und Katheterisierung der Ampulla war mit dem transvaginalen Zugang
nicht möglich. Der laterale Flankenzugang machte eine Visualisierung des Ovars,
Infundibulums und des gesamten Oviducts in allen Fällen möglich. Nach der
Auffaltung des Infundibulums war das Ostium abdominale in 8 (72.7%) Fällen
sichtbar und eine Katheterisierung der Ampulla gelang in 7/11 (63.6%) Fällen. In 5
der 7 Spülungen konnte die Spülflüssigkeit intrauterine mittels Hysteroskopie
nachgewiesen warden.
Ausblick: In der vorliegenden Arbeit korrelierte der uterine PI mit der Anzahl
gewonnener Embryonen an t3 und D-4, so dass die transrektale Dopplersonographie
der uterinen Arterien hilfreich sein könnte den Erfolg einer
Superovulationsbehandlung vorherzusagen. Jedoch sollten in weiteren Studien diese
Ergebnisse bestätigt werden. Die laparoskopische Eileiterspülung ist mittels eines
lateralen Flankenzugangs möglich und könnte für die Gewinnung tubaler
Embryonalstadien und den intratubalen Transfer von Gameten (GIFT) zur Verfügung
stehen.

References 55
Chapter 8
References

56 References
ACOSTA, T. J., K. G. HAYASHI, M. MATSUI and A. MIYAMOTO (2005):
Changes in follicular vascularity during the first follicular wave in lactating cows.
J Reprod Dev 51, 273-280
ALFORD, C. and R. HANSON (2010):
Evaluation of a transvaginal laparoscopic natural orifice transluminal endoscopic
surgery approach to the abdomen of mares.
Vet. Surg. 39, 873-878
ALLEN, W., B. M. KESSY and D. E. NOAKES (1979):
Evaluation of uterine tube function in pony mares.
Vet. Rec. 105, 364-366
ALLEN, W. (2005):
The development and application of the modern reproductive technologies to horse
breeding.
Reprod. Domest. Anim. 40, 310-329
ALLEN, W. R., S. WILSHER, L. MORRIS, J. CROWHURST, M. HILLYER and H.
NEAL (2006):
Laparoscopic application of PGE2 to re-establish oviducal patency and fertility in
infertile mares: a preliminary study.
Equine Vet. J. 38, 454-459
ALVARENGA, M. A., P. M. MCCUE, J. BRUEMMER, J. R. NEVES NETO and E. L.
SQUIRES (2001):
Ovarian superstimulatory response and embryo production in mares treated with
equine pituitary extract twice daily.
Theriogenology 56, 879-887

References 57
ARAUJO, G. H., A. N. ROCHA FILHO, E. P. LOPES, C. F. MOYA and M. A.
ALVARENGA (2009):
Use of a low dose of equine purified FSH to induce multiple ovulations in mares.
Reprod. Domest. Anim. 44, 380-383
ARTHUR, G. (1958):
An analysis of the reproductive function of mares based on postmortem examination.
Vet. Rec. 70, 682-686
AURICH, C. (2011):
Reproductive cycles of horses.
Anim. Reprod. Sci. 124, 220-228
BARFIELD, J. P., P. M. MCCUE, E. L. SQUIRES and G. E. SEIDEL, JR. (2009):
Effect of dehydration prior to cryopreservation of large equine embryos.
Cryobiology 59, 36-41
BASS, L. D., D. J. DENNISTON, L. J. MACLELLAN, P. M. MCCUE, G. E. SEIDEL,
JR. and E. L. SQUIRES (2004):
Methanol as a cryoprotectant for equine embryos.
Theriogenology 62, 1153-1159
BATTUT, I., S. COLCHEN, F. FIENI, D. TAINTURIER and J. F. BRUYAS (1997):
Success rates when attempting to nonsurgically collect equine embryos at 144, 156
or 168 hours after ovulation.
Equine Vet. J. (Suppl) 25, 60-62

58 References
BECK, L. and L. BOOTS (1974):
The comparative anatomy, histology and morphology of the mammalian oviduct.
In: The Oviduct and its Functions. Proceedings of a Symposium University of
Georgia October 1972).
Ed.: JOHNSON, A. D., C. W. FOLEY
Academic-Press, New York and London, pp. 1-51
BECKER, S. E. and A. L. JOHNSON (1992):
Effects of gonadotropin-releasing hormone infused in a pulsatile or continuous
fashion on serum gonadotropin concentrations and ovulation in the mare.
J. Anim. Sci. 70, 1208-1215
BEG, M. A. and O. J. GINTHER (2006):
Follicle selection in cattle and horses: role of intrafollicular factors.
Reproduction 132, 365-377
BENNETT, S. (2007):
Diagnosis of oviductal disorders and diagnostic techniques.
In: Current Therapy in Equine Reproduction
SAMPER, J. C., PYCOCK, J. F., MCKINNON, A. O. (Eds.)
Saunders Elsevier, St. Louis, Missouri, USA, pp. 78-82
BERGFELT, D. and O. J. GINTHER (1993):
Relationships between FSH surges and follicular waves during the estrus cycle in
mares.
Theriogenology 39, 781-796
BERGFELT, D., B. G. MANN, N. B. SCHWARTZ and O. J. GINTHER (1991):
Circulating concentrations of immunoreactive inhibin and FSH during the estrus
cycle of mares.
J. Equine Vet. Sci. 11, 319-322

References 59
BESENFELDER, U., G. BREM, 1998:
Tubal transfer of bovine embryos: a simple endoscopic method reducing long term
exposure of in vitro produced embryos.
Theriogenology 50, 739–745
BESENFELDER, U., V. HAVLICEK, G. MOSSLACHER and G. BREM (2001):
Collection of tubal stage bovine embryos by means of endoscopy. A technique
report.
Theriogenology 55, 837-845
BETTERIDGE, K. J., M. D. EAGLESOME, D. MITCHELL, P. F. FLOOD and R.
BERIAULT (1982):
Development of horse embryos up to twenty two days after ovulation: observations
on fresh specimens.
J. Anat. 135, 191-209
BLAIR, W. D. and L. R. BECK (1977):
In vivo effects of prostaglandin F2 alpha and E2 on contractility and diameter of the
rabbit oviduct using intraluminal transducers.
Biol. Reprod. 16, 122-127
BOLLWEIN, H., J. MAIERL, R. MAYER and R. STOLLA (1998):
Transrectal color Doppler sonography of the A. uterina in cyclic mares.
Theriogenology 49, 1483-1488
BOLLWEIN, H., R. MAYER, F. WEBER and R. STOLLA (2002a):
Luteal blood flow during the estrus cycle in mares.
Theriogenology 57, 2043-2051

60 References
BOLLWEIN, H., F. WEBER, B. KOLBERG and R. STOLLA (2002b):
Uterine and ovarian blood flow during the estrus cycle in mares.
Theriogenology 65, 2129-2138
BOYLE, M. S., D. G. CRAN, W. R. ALLEN and R. H. HUNTER (1987):
Distribution of spermatozoa in the mare's oviduct.
J. Reprod. Fertil. (Suppl.) 35, 79-86
BROUSSIN, B. (2007):
The clinical value of ultrasound for endometrial receptivity assessment in Assisted
Reproductive Techniques (ART) procedures.
Gynecol. Obstet. Fertil. 35, 570-575
CACCIATORE, B., N. SIMBERG, P. FUSARO and A. TIITINEN (1996):
Transvaginal Doppler study of uterine artery blood flow in in vitro fertilization-embryo
transfer cycles.
Fertil Steril 66, 130-134
CARMO, M., M. ALVERANGA, L. LOSINNO and J. AQUILAR (2006):
Oocyte transport to the oviduct of superovulated mares.
Anim. Reprod. Sci. 94, 337-339
CARNEVALE E. (2004):
Oocyte transfer and gamete intrafallopian transfer in the mare.
Proc. 15th Congr. Anim. Reprod. Research. and Practice III 82–83, 617–632
CARNEVALE, E. (2006):
Vitrification of equine embryos.
Vet. Clin. North. Am. Equine Pract. 22, 831-841

References 61
CARNEVALE, E., E. SQUIRES and A. MCKINNON (1987):
Comparison of Ham’s F10 with CO2 or Hepes buffer for storage of equine embryos
at 5 C for 24 h.
J. Anim. Sci. 65, 1775–1781
CARNEVALE E., O. J. GINTHER (1993):
Use of a linear ultrasonic transducer for a transvaginal aspiration and transfer of
oocytes in the mare.
J. Eq. Vet. Sci. 13, 331–333
CARNEVALE, E., M. USON, J. BOZZOLA, S. KING, S. SCHMITT and H. GATES
(1999):
Comparison of oocytes from young and old mares with light and electron microscopy.
Theriogenolgy 5, 299
CARNEY, N. J., E. L. SQUIRES, V. M. COOK, G. E. SEIDEL, JR. and D. J. JASKO
(1991):
Comparison of pregnancy rates from transfer of fresh versus cooled, transported
equine embryos.
Theriogenology 36, 23-32
CHOI, Y. H., D. L. HARTMAN, S. B. BLISS, S. S. HAYDEN, T. L. BLANCHARD and
K. HINRICHS (2010):
High pregnancy rates after transfer of large equine blastsocysts collapsed via
micromanipulation before vitrification.
Reprod. Fertil. Dev. 22, 203
CHOI, Y. H., I. C. VELEZ, F. L. RIERA, J. E. ROLDAN, D. L. HARTMAN, S. B.
BLISS, T. L. BLANCHARD, S. S. HAYDEN and K. HINRICHS (2011):
Successful cryopreservation of expanded equine blastocysts.
Theriogenology 76, 143-152

62 References
COLBERN, G. and W. REAGAN (1987):
Ovariectomy by colpotomy in mares.
Contin. Educ. Pract. Vet. 9, 1035-1038
COLCHEN, S., I. BATTUT, F. FIENI, D. TAINTURIER, B. SILIART and J. F.
BRUYAS (2000):
Quantitative histological analysis of equine embryos at exactly 156 and 168 h after
ovulation.
J. Reprod. Fertil. (Suppl.) 56, 527-537
CROWELL-DAVIS, S. (2007):
Sexual behavior of mares.
Horm. Behav. 52, 12-17
CULLINGFORD, E., E. SQUIRES, P. MCCUE and S. G. JR. (2010):
Attempts at superovulation of mares with porcine follicle stimulating hormone and
recombinant equine follicle stimulating hormone.
J. Equine Vet. Sci. 30, 305-309
CZLONKOWSKA, M., M. BOYLE and W. ALLEN (1985):
Deep freezing of horse embryos.
J. Reprod. Fertil. 75, 485–490
DECHANT, J. and D. HENDRICKSON (2000):
Standing female equine urogenital endoscopic surgery.
Vet. Clin. North. Am. Equine Pract. 16, 301-315

References 63
DERAR, R. I., Y. MAEDA, S. M. HOQUE, T. OSAWA, G. WATANABE, K. TAYA and
Y. MIYAKE (2004):
Effect of active immunization of pony mares against recombinant porcine inhibin
alpha subunit on ovarian follicular development and plasma steroids and
gonadotropins.
J. Vet. Med. Sci. 66, 31-35
DESANTIS, S., S. ZIZZA, G. ACCOGLI, F. ACONE, R. ROSSI and L. RESTA (2011):
Morphometric and ultrastructural features of the mare oviduct epithelium during
oestrus.
Theriogenology 75, 671-678
DINGER, J. E., E. E. NOILES and M. J. BATES (1982):
Ovarian activity in progesterone treated mares following administration of pregnant
mare serum gonadotropin.
Theriogenology 17, 305-311
DIPPERT, K. D., S. HOFFERER, E. PALMER, D. J. JASKO and E. L. SQUIRES
(1992):
Initiation of superovulation in mares 5 or 12 days after ovulation using equine
pituitary extract with or without GnRH analogue.
Theriogenology 38, 695-710
DONADEU, F. X. and O. J. GINTHER (2001):
Effect of number and diameter of follicles on plasma concentrations of inhibin and
FSH in mares.
Reproduction 121, 897-903
DONADEU, F. X. and H. G. PEDERSEN (2008):
Follicle development in mares.
Reprod. Domest. Anim. (Suppl.) 43, 224-231

64 References
DOUGLAS, R. H., L. Nuti, O. J. GINTHER (1974):
Induction of ovulation and multiple ovulation in seasonally-anovulatory mares with
equine pituitary fraction.
Theriogenology 2, 133–143
DOUGLAS, R. H. (1979):
Review of induction of superovulation and embryo transfer in the equine.
Theriogenology 11, 33-46
ELDRIDGE-PANUSKA, W. D., V. C. DI BRIENZA, G. E. SEIDEL, JR., E. L.
SQUIRES and E. M. CARNEVALE (2005):
Establishment of pregnancies after serial dilution or direct transfer by vitrified equine
embryos.
Theriogenology 63, 1308-1319
EMBERTSON, R. (2006):
Ovaries and uterus
In: Equine Surgery
Saunders,W.B., St. Louis, MO, third Edition, pp. 855-864
FALLOPPIO, G. (1561):
Observationes anatomicae.
Ulma, M A. and G. Perchachino, Venice
apud Iacobum Keruer, 1562
cit. from BECK and BOOTS (1974)
FLUKER, M. R., W. M. HOOPER and A. A. YUZPE (1999):
Withholding gonadotropins ("coasting") to minimize the risk of ovarian
hyperstimulation during superovulation and in vitro fertilization-embryo transfer
cycles.
Fertil. Steril. 71, 294-301

References 65
FORTUNE, J. and T. KIMMICH (1993):
Purified pig FSH increases the rate of double ovulation in mares.
Equine Vet. J. (Suppl.) 25, 95-98
FREEMAN, D. A., J. A. WEBER, R. T. GEARY and G. L. WOODS (1991):
Time of embryo transport through the mare oviduct.
Theriogenology 36, 823-830
GASTAL, E. L., M. O. GASTAL, M. A. BEG and O. J. GINTHER (2004):
Interrelationships among follicles during the common-growth phase of a follicular
wave and capacity of individual follicles for dominance in mares.
Reproduction 128, 417-422
GASTAL, E. L., M. O. GASTAL, D. R. BERGFELT and O. J. GINTHER (1997):
Role of diameter differences among follicles in selection of a future dominant follicle
in mares.
Biol. Reprod. 57, 1320-1327
GASTAL, E. L., M. O. GASTAL and O. J. GINTHER (2006):
Relationships of changes in B-mode echotexture and color-Doppler signals in the
wall of the preovulatory follicle to changes in systemic oestradiol concentrations and
the effects of human chorionic gonadotrophin in mares.
Reproduction 131, 699-709
GASTAL, E. L., M. O. GASTAL, M. C. WILTBANK and O. J. GINTHER (1999):
Follicle deviation and intrafollicular and systemic estradiol concentrations in mares.
Biol. Reprod. 61, 31-39
GEARY, R., J. WEBER and G. WOODS (1989):
Hastened transport of equine embryos through the oviduct of the mare.
Theriogenology 31, 973-978

66 References
GINTHER, O. J. (1979):
Reproductive Biology of the Mare: Basic and Applied Aspects.
Equiservices Plublishing, Cross Plains, WI, USA
GINTHER, O. J. (1986):
Ultrasonic Imaging and Reproduction: Horses.
Equiservices Publishing, Cross Plains, WI, USA
GINTHER, O. J. (2004):
Doppler ultrasound in equine reproduction: principles, techniques and potential.
J. Equine Vet. Sci. 24, 516-526
GINTHER, O. J. (2007):
Ultrasonic Imaging and Animal Reproduction: Color Doppler Ultrasonography.Book 4
Equiservices Publishing, Cross Plains, WI, USA
GINTHER, O. J. (2012):
The mare: a 1000-pound guinea pig for study of the ovulatory follicular wave in
women.
Theriogenology 77, 818-828
GINTHER, O. J., M. A. BEG, D. R. BERGFELT, F. X. DONADEU and K. KOT (2001):
Follicle selection in monovular species.
Biol Reprod 65, 638-647
GINTHER, O. J., M. A. BEG, K. KOT, C. MEIRA and D. R. BERGFELT (2003):
Associated and independent comparisons between the two largest follicles preceding
follicle deviation in cattle.
Biol Reprod 68, 524-529

References 67
GINTHER, O. J. and D. R. BERGFELT (1990):
Effect of GnRH treatment during the anovulatory season on multiple ovulation rate
and on follicular development during the ensuring pregnancy in mares.
J Reprod Fertil 88, 119-126
GINTHER, O. J. and D. R. BERGFELT (1992):
Associations between FSH concentrations and major and minor follicular waves in
pregnant mares.
Theriogenology 38, 807-821
GINTHER, O. J., R. H. DOUGLAS and J. R. LAWRENCE (1982):
Twinning in mares: A survey of veterinarians and analyses of theriogenology records.
Theriogenology 18, 333-347
GINTHER, O. J., E. J. GASTAL, M. O. GASTAL and M. A. BEG (2005):
Regulation of circulating gonadotropins by the negative effects of ovarian hormones
in mares.
Biol. Reprod. 73, 315-323
GINTHER, O. J., E. L. GASTAL, M. O. GASTAL and M. A. BEG (2007):
Effect of prostaglandin F2alpha on ovarian, adrenal, and pituitary hormones and on
luteal blood flow in mares.
Domest. Anim. Endocrinol. 32, 315-328
GINTHER, O. J., E. L. GASTAL, M. O. GASTAL and M. A. BEG (2008a):
Dynamics of the equine preovulatory follicle and periovulatory hormones: what’s
new?
J. Equine Vet. Sci. 28, 454–460

68 References
GINTHER, O. J., E. L. GASTAL, M. O. GASTAL, D. R. BERGFELT, A. R.
BAERWALD and R. A. PIERSON (2004):
Comparative study of the dynamics of follicular waves in mares and women.
Biol. Reprod. 71, 1195-1201
GINTHER, O. J., M. O. GASTAL, E. L. GASTAL, J. C. JACOB, M. A. SIDDIQUI and
M. A. BEG (2008b):
Effects of age on follicle and hormone dynamics during the oestrus cycle in mares.
Reprod. Fertil. Dev. 20, 955-963
GUILLO, F. and Y. COMBARNOUS (1983):
Purification of equine gonadotropins and comparative study of their acid-dissociation
and receptor binding specifity.
Biochim. Biophys. Acta 755, 229-236
HANSON, C. A. and L. D. GALUPPO (1999):
Bilateral laparoscopic ovariectomy in standing mares: 22 cases.
Vet. Surg. 28, 106-112
HEIDLER, B., J. E. AURICH, W. POHL and C. AURICH (2004):
Body weight of mares and foals, estrus cycles and plasma glucose concentration in
lactating and non-lactating Lipizzaner mares.
Theriogenology 61, 883-893
HINRICHS K., R. W.BETSCHART, P. M. MCCUE, E. L. SQUIRES (2000):
Effect of timing of follicle aspiration on pregnancy rate after oocyte transfer in mares.
J. Reprod. Fertil. 56 (Suppl.), 493–498

References 69
HINRICHS K., C. C. LOVE, S. P. BRINSKO, Y. H. CHOI, D. D. VARNER (2002):
In vitro fertilization of in vitro-matured equine oocytes: effect of maturation medium,
duration of maturation, and sperm calcium ionophore treatment, and comparison with
rates of fertilization in vivo after oviducal transfer.
Biol. Reprod. 67, 256–262
HINRICHS, K. and Y.-H. CHOI (2012):
Equine Embryo Biopsy, Genetic Testing, and Cryopreservation.
J. Equine Vet. Sci. 23, 390-397
HOCHI, S., T. FUJIMOTO and N. OGURI (1995):
Large equine blastocysts are damaged by vitrification procedures.
Reprod. Fertil. Dev. 7, 113-117
HONNENS, A., H. NIEMANN, K. HERZOG, V. PAUL, H. H. MEYER and H.
BOLLWEIN (2009):
Relationships between ovarian blood flow and ovarian response to eCG-treatment of
dairy cows.
Anim. Reprod. Sci. 113, 1-10
HONNENS, A., H. NIEMANN, V. PAUL, H. H. MEYER and H. BOLLWEIN (2008):
Doppler sonography of the uterine arteries during a superovulatory regime in cattle.
Uterine blood flow in superovulated cattle.
Theriogenology 70, 859-867
HONNENS, A., S. WEISSER, H. WELTER, R. EINSPANIER and H. BOLLWEIN
(2011):
Relationships between uterine blood flow, peripheral sex steroids, expression of
endometrial estrogen receptors and nitric oxide synthases during the estrous cycle in
the mare.
J. Reprod. Dev. 57, 43-48

70 References
HUNTER, R. H. (2002):
Vital aspects of Fallopian tube physiology in pigs.
Reprod. Domest. Anim. 37, 186-190
HUNTER, R. H. (2005):
The Fallopian tubes in domestic mammals: how vital is their physiological activity?
Reprod. Nutr. Dev. 45, 281-290
IMEL, K. J., E. L. SQUIRES, R. P. ELSDEN (1981):
Collection and transfer of equine embryos.
J. Am. Vet. Med. Assoc. 179, 987-991
IRVINE, C. H., S. L. GASTAL (1993):
GnRH
In: Equine Reproduction
MCKINNON, A.O., J. L. VOSS (Eds.)
Williams and Wilkins, Media, USA, pp. 37-44
JABLONKA-SHARIFF, A., J. F. ROSER, G. R. BOUSFIELD, M. W. WOLFE, L. E.
SIBLEY, M. COLGIN and I. BOIME (2007):
Expression and bioactivity of a single chain recombinant equine luteinizing hormone
(reLH).
Theriogenology 67, 311-320
JACOB, J. C., E. L. GASTAL, M. O. GASTAL, G. R. CARVALHO, M. A. BEG and O.
J. GINTHER (2009):
Temporal relationships and repeatability of follicle diameters and hormone
concentrations within individuals in mares.
Reprod. Domest. Anim. 44, 92-99

References 71
JENNINGS, M., I. BOIME, D. DAPHNA-IKEN, A. JABLONKA-SHARIFF, A. CONLEY
and M. COLGIN (2009):
The efficacy of recombinant equine follicle stimulating hormone (reFSH) to promote
follicular growth in mares using a follicular suppression model.
Anim. Reprod. Sci. 116, 291-307
KAINER, R. A. (1993):
Reproductive Organs of the mare.
In: Equine Reproduction
MCKINNON, A.O., J. L. VOSS (Eds.)
Williams and Wilkins, Media, USA, pp. 10-12
KIMURA, J., Y. HIRANO, S. TAKEMOTO, Y. NAMBO, T. ISHINAZAKA, R. HIMENO,
T. MISHIMA, S. TSUMAGARI and H. YOKOTA (2005):
Three-dimensional reconstruction of the equine ovary.
Anat. Histol. Embryol. 34, 48-51
LAMAZOU, F., V. LETOUZEY, E. ARBO, M. GRYNBERG, J. M. LEVAILLANT, R.
FRYDMAN and R. FANCHIN (2009):
The role of ultrasound examination to evaluate ovarian reserve of infertile patients.
Gynecol. Obstet. Fertil. 37, 425-431
LAPIN, D. R. and O. J. GINTHER (1977):
Induction of ovulation and multiple ovulations in seasonally anovulatory and ovulatory
mares with an equine pituitary extract.
J. Anim. Sci. 44, 834-842

72 References
LEY, W., B. J. BOWEN, P. DASCANIO, N. PARKER, T. BAILEY and W. DIGRASSI
(1998):
Modified technique to evaluate uterine tubal pantency in the mare.
In: Proceedings of the 44th Annual Convention of the American Assocoation of
Equine Practitioners pp.56-59
LOGAN, N. L., P. M. MCCUE, M. A. ALONSO and E. L. SQUIRES (2007):
Evaluation of three equine FSH superovulation protocols in mares.
Anim Reprod Sci 102, 48-55
MACLELLAN, L. J., E. M. CARNEVALE, M. A. COUTINHO DA SILVA, P. M.
MCCUE, L. G. J. SEIDE and S. EL (2002):
Cryopreservation of small and large equine embryos pre-treated with cytochalasin-B
and /or trypsin.
Theriogenolgy 58, 717-720
MCCUE, P. M. (1996):
Superovulation.
Vet. Clin. North. Am. Equine Pract. 12, 1-11
MCCUE, P. M., N. J. CARNEY, J. P. HUGHES, J. RIVIER, W. VALE and B. L.
LASLEY (1992):
Ovulation and embryo recovery rates following immunization of mares against an
inhibin alpha-subunit fragment.
Theriogenology 38, 823-831
MCCUE, P. M., M. M. LEBLANC and E. L. SQUIRES (2007):
eFSH in clinical equine practice.
Theriogenology 68, 429-433

References 73
MCCUE, P. M., N. LOGAN, M. ALONSO and E. L. SQUIRES (2006):
Superovulation of mares: Effects of eFSH treatment during consecutive or alternate
cycles and as short-term therapy before follicular divergence.
Anim. Reprod. Sci. 94 404-407
MCKINNON, A. O., E. L. SQUIRES, J. L. VOSS and V. M. COOK (1998):
Equine embryo transfer: a review.
Compend. Contin. Educ. Pract. Vet. 10, 343-355
MCKINNON, A. O., J. L. VOSS, E. L. SQUIRES and E. M. CARNEVALE (1993):
Diagnostic ultrasonography.
In: Equine Reproduction
MCKINNON, A.O., J. L. VOSS (Eds.)
Wiliams and Wilkins, Media, PA, USA, pp. 266-302
MENEZO, Y. and P. GUERIN (1997):
The mammalian oviduct: biochemistry and physiology.
Eur. J. Obstet. Gynecol. Reprod. Biol. 73, 99-104
MEYERS-BROWN, G., P. MCCUE, K. NISWENDER, E. SQUIRES, C. DELUCA and
L. BIDSTRUP (2010):
Superovulation in mares using recombinant equine follicle stimulating hormone
(reFSH): ovulation rates, embryo retrieval and hormone profiles.
J. Equine Vet. Sci. 30, 560-568
MIRO, L., O. ANGLADA, A. CRISCI, E. TABERNER and N. FABREGAS (2010):
Color Doppler ultrasonographic evaluation of luteolysis in donkeys.
Anim. Reprod. Sci. 45, 94

74 References
MONNIAUX, D., D. CHUPIN and J. SAUMANDE (1983):
Superovulatory responses of cattle.
Theriogenology 19, 55-81
NAMBO, Y., H. KANEKO, S. NAGATA, M. OIKAWA, T. YOSHIHARA, N.
NAGAMINE, G. WATANABE and K. TAYA (1998):
Effect of passive immunization against inhibin on FSH secretion, folliculogenesis and
ovulation rate during the follicular phase of the estrus cycle in mares.
Theriogenology 50, 545-557
NEAL, H. N. (2011):
Laparoscopic evaluation of oviducal patency.
Proceedings of the 50th British Equine Veterinary Association Congress BEVA
Sep. 7 – 10, 2011 Liverpool, United Kingdom pp.140-141
NIE, G. (2007):
Estrus suppression.
In: Current Therapy in Equine Reproduction
Saunders Elsevier, St. Louis, Mi, USA, pp. 26-31
NISWENDER, K., M. ALVARENGA, P. MCCUE, Q. HARDY and E. SQUIRES
(2003):
Superovulation in cycling mares using equine follicle stimulating hormone (eFSH).
J. Equine Vet. Sci. 23, 497-500
OGURI, N. and Y. TSUTSUMI (1972):
Non-surgical recovery of equine eggs, and an attempt at non-surgical egg transfer in
horses.
J. Reprod. Fertil. 31, 187-195

References 75
OZTURK, O., S. BHATTACHARYA, E. SARIDOGAN, E. JAUNIAUX and A.
TEMPLETON (2004):
Role of utero-ovarian vascular impedance: predictor of ongoing pregnancy in an IVF-
embryo transfer programme.
Reprod Biomed Online 9, 299-305
PALMER, E. and M. DRIANCOURT (1980):
Use of ultrasonic echography in equine gynecology.
Theriogenolgy 13, 203-216
PIERSON, R. A. and O. J. GINTHER (1990):
Ovarian folliclular response of mares to an equine pituitary extract after suppression
of follicular development.
Anim Reprod Sci 22, 131-144
PROBST, J. (2009):
Untersuchungen zur Superovulation bei der Stute: Einfluss von eFSH® auf
Follikelentwicklung und –durchblutung.
Hannover, tierärztl. Hochsch. Diss.
RAMBAGS, B. P., P. J. KRIJTENBURG, H. F. DRIE, G. LAZZARI, C. GALLI, P. L.
PEARSON, B. COLENBRANDER and T. A. STOUT (2005):
Numerical chromosomal abnormalities in equine embryos produced in vivo and in
vitro.
Mol. Reprod. Dev. 72, 77-87
RAZ, T., M. D. AMORIM, B. C. STOVER and C. E. CARD (2010):
Ovulation, pregnancy rate and early embryonic development in vernal transitional
mares treated with equine- or porcine-FSH.
Reprod. Domest. Anim. 45, 287-294

76 References
RAZ, T., S. CARLEY and C. CARD (2009a):
Comparison of the effects of eFSH and deslorelin treatment regimes on ovarian
stimulation and embryo production of donor mares in early vernal transition.
Theriogenology 71, 1358-1366
RAZ, T., A. GRAY, B. HUNTER and C. CARD (2009b):
Early effects of equine FSH (eFSH) treatment on hormonal and reproductive
parameters in mares intended to carry their own pregnancy.
Anim. Reprod. Sci. 115, 76-87
RAZ, T., S. CARLEY, J. GREEN, M. CORRIGAN and C. CARD (2005):
Effect of prostaglandin in early diestrus or progesterone and estradiol administration
on equine FSH-treated donor mare embryo recovery and recipient pregnancy rate.
In: Proceedings of the 51st Annual Convention of the American Association of
Equine Practitioners, December 3-7, 2005 Seattle, WA, USA, pp. 236-241
RIERA, F. and J. MCDONOUGH (1993):
Commercial embryo transfer in polo ponies in Argentina.
Equine Vet. J. (Suppl.) 15, 116-119
ROBINSON, S. J., H. NEAL and W. ALLEN (2000):
Modulation of oviductal transport in mares by local application of prostaglandin E2.
J. Reprod. Fert. 56, 587-592
RODRIGUEZ-MARTINEZ, H., H. EKWALL and L. PLOEN (1985):
Ultrastructure and innervation of smooth muscle in the porcine oviduct.
Anat. Histol. Embryol. 14, 33-46

References 77
ROSAS, C. A., R. H. ALBERIO, J. L. BARANAO, A. AGUERO and M. G. CHAVES
(1998):
Evaluation of two treatments in superovulation of mares.
Theriogenology 49, 1257-1264
ROSER, J. and G. MEYERS-BROWN (2012):
Superovulation in the mare: A work in progress.
J. Equine Vet. Sci. 32, 376-386
SAUNDERS, R. D., J. M. SHWAYDER and S. T. NAKAJIMA (2011):
Current methods of tubal patency assessment.
Fertil. Steril. 95, 2171-2179
SAYASITH, K., N. BOUCHARD, M. DORE and J. SIROIS (2009):
Gonadotropin-dependent regulation of the prostaglandin E2 receptor in equine
preovulatory follicles during the ovulatory process in mares.
Mol. Reprod. Dev. 76, 191-201
SCHERZER, J., C. DAVIS and D. HURLEY (2011):
Laserassisted vitrification of large equine embryos.
Reprod. Domest. Anim. 46, 1104-1106
SCHERZER, J., R. A. FAYRER-HOSKEN, L. RAY, D. J. HURLEY and G. L.
HEUSNER (2008):
Advancements in large animal embryo transfer and related biotechnologies.
Reprod. Domest. Anim. 43, 371-376
SCOGGIN, C. F., C. MEIRA, P. M. MCCUE, E. M. CARNEVALE, T. M. NETT and E.
L. SQUIRES (2002):
Strategies to improve the ovarian response to equine pituitary extract in cyclic mares.
Theriogenology 58, 151-164

78 References
SCOTT T. J., E. M. CARNEVALE, L. J. MACLELLAN, C. F. SCOGGIN CF, E. L.
SQUIRES (2001):
Embryo development rates after transfer of oocytes matured in vivo, in vitro, or within
oviducts of mares.
Theriogenology 55, 705–715
SEABORNE, E. (1925):
The oestrus cycle of the mare and some associated phenomena.
Anat. Rec. 30, 277-286
SIEME, H. and E. KLUG (1996):
Special experiences with direct ovulation control in mares for insemination.
Prakt. Tierarzt 77, 76-78
SINGH, J., M. DOMINGUEZ, R. JAISWAL and G. P. ADAMS (2004):
A simple ultrasound test to predict the superstimulatory response in cattle.
Theriogenology 62, 227-243
SIRARD, M. A., L. PICARD, M. DERY, K. COENEN and P. BLONDIN (1999):
The time interval between FSH administration and ovarian aspiration influences the
development of cattle oocytes.
Theriogenology 51, 699-708
SIROIS, J., B. BALL and J. FORTUNE (1989):
Patterns of growth and regression of ovarian follicles during the oestrus cycle and
after hemiovariectomy in mares.
Equine Vet. J. (Suppl.) 8, 43-48
SLADE, N. P., T. TAKEDA, E. L. SQUIRES, R. P. ELSDEN and G. E. SEIDEL, JR.
(1985):
A new procedure for the cryopreservation of equine embryos.
Theriogenology 24, 45-58

References 79
SQUIRES, E. L. (1996):
Maturation and fertilization of equine oocytes.
Vet. Clin. North Am. Equine Pract. 12, 31-45
SQUIRES, E. L. (2006):
Superovulation in mares.
Vet. Clin. North Am. Equine Pract. 22, 819-830
SQUIRES, E. L., E. M. CARNEVALE, P. M. MCCUE and J. E. BRUEMMER (2003):
Embryo technologies in the horse.
Theriogenology 59, 151-170
SQUIRES, E. L., R. H. GARCIA, O. J. GINTHER, J. L. VOSS and G. E. SEIDEL, JR.
(1986):
Comparison of equine pituitary extract and follicle stimulating hormone for
superovulating mares.
Theriogenology 26, 661-670
SQUIRES, E. L. and P. MCCUE (2011):
Superovulation.
In: Equine Reproduction. Second Edition.
MCKINNON, A. O., E. L. SQUIRES, W. E. VAALA, D. D. VARNER (Eds.)
Wiley-Blackwell, Ames, Iowa, pp. 2277-2290
SQUIRES, E. L. and P. M. MCCUE (2007):
Superovulation in mares.
Anim. Reprod. Sci. 99, 1-8

80 References
SQUIRES, E. L., A. O. MCKINNON, E. M. CARNEVALE, R. MORRIS and T. M.
NETT (1987):
Reproductive characteristics of spontaneous single and double ovulating mares and
superovulated mares.
J Reprod Fertil Suppl 35, 399-403
STABENFELDT, G. H., J. P. HUGHES and J. W. EVANS (1972):
Ovarian activity during the estrus cycle of the mare.
Endocrinology 90, 1379-1384
STOUT, T. (2006):
Equine embryo transfer:review of developing potential.
Equine Vet. J. 38, 467-478
STOUT, T. (2012):
Cryopreservation of equine embryos: current state-of-the-art.
Reprod. Domest. Anim. (Suppl.) 47, 84-89
STROUD B. (2011):
The year 2010 worldwide statistics of embryo transfer in domestic farm animals.
Statistics and Data Retrieval Committee Report.
WHEELER, M. B. (Ed.)
IETS Embryo Transfer Newsletter 29, 12–23
ISSN 1083-4699
STROUD B. (2012):
The year 2011 worldwide statistics of embryo transfer in domestic farm animals.
Statistics and Data Retrieval Committee Report.
WHEELER, M. B. (Ed.)
IETS Embryo Transfer Newsletter 30, 16-26
ISSN 1083-4699

References 81
THARASANIT, T., B. COLENBRANDER, M.M. BEVERS, T.A. STOUT (2006):
Effects of recombinant human follicle stimulating hormone on follicle development
and ovulation in the mare.
Theriogenology 65, 1071-81
THIBIER, M. (2000):
The IETS statistics of embryo transfers in livestock in the world for the year 1999: a
new record for bovine in vivo-derived embryos transferred.
IETS Newsletter 18, 24-28
THIBIER, M. (2005):
Significant increases in transfers of both in vivo derived and in vitro produced
embryos in cattle and contrasted trends in other species in 2004.
IETS Newsletter 23, 11-17
TOWNSON, D. H. and O. J. GINTHER (1989):
Ultrasonic characterization of follicular evacuation during ovulation and fate of the
discharged follicular fluid in mares.
Anim. Reprod. Sci. 20, 131-141
VAN NIEKERK, C. and W. GERNEKE (1966):
Persistence and parthenogenetic cleavage of tubal ova in the mare.
Onderstepoort. J. Vet. Res. 33, 195-232
VOGELSANG, S., K. BONDIOLI and J. MASSEY (1985):
Commercial application of equine embryo transfer.
Equine Vet. J. (Suppl.) 3, 89-91

82 References
WALKER, D. and J. VAUGHAN (1980):
Bovine and Equine Urogenital Surgery.
In: Urogenital Surgery
Lea & Febiger, Philadelphia, PA, pp. 265-301
WALMSLEY, J. P. (1999):
Review of equine laparoscopy and an analysis of 158 laparoscopies in the horse.
Equine Vet. J. 31, 456-464
WATSON, E. D. and M. O. AL-ZI'ABI (2002):
Characterization of morphology and angiogenesis in follicles of mares during spring
transition and the breeding season.
Reproduction 124, 227-234
WEBER, J. A., D. FREEMAN, D. VANDERWALL and G. WOODS (1991):
Prostaglandin E2 secretion by oviductal transport-stage equine embryos.
Biol. Reprod. 45, 540-543
WEBER, J. A., G. WOODS, D. FREEMAN and D. VANDERWALL (1992):
Prostaglandin E2-specific binding to the equine oviduct.
Prostaglandins 43, 61-65
WEBER, J. A., G. L. WOODS and J. J. AGUILAR (1996):
Location of equine oviductal embryos on day 5 post ovulation and oviductal transport
time of day 5 embryos autotransferred to the contralateral oviduct.
Theriogenology 46, 1477-1483

References 83
WEINER, Z., I. THALER, J. LEVRON, N. LEWIT and J. ITSKOVITZ-ELDOR (1993):
Assessment of ovarian and uterine blood flow by transvaginal color Doppler in
ovarian-stimulated women: correlation with the number of follicles and steroid
hormone levels.
Fertil. Steril. 59, 743-749
WELCH, S., D. DENNISTON, J. HUDSON, J. BRUEMMER, P. MCCUE and E.
SQUIRES (2006):
Exogenous eFSH, follicle coasting, and hCG as a novel superovulation regimen in
mares.
J. Equine Vet. Sci. 26, 262-270
WOODS, G. (1998):
Pregnancy loss: a major cause of subfertility in the mare.
Equine Pract. 11, 29-32
ZAIDI, J., R. PITTROF, A. SHAKER, A. KYEI-MENSAH, S. CAMPBELL and S. L.
TAN (1996):
Assessment of uterine artery blood flow on the day of human chorionic gonadotropin
administration by transvaginal color Doppler ultrasound in an in vitro fertilization
program.
Fertil. Steril. 65, 377-381
ZENT, W. W., L. LIU and M. SPIRITO (1993):
Oviduct flushing as a treatment for infertility in the mare.
Equine Vet. J. (Suppl.) 15, 47-48

84 Appendix
9 Appendix
9.1 Material and methods study I and II
9.1.1 Animals
Six clinically healthy and normally cycling mares (3 Trotter, 2 Warmblood, 1
Thoroughbred) between 4 to 14 years of age were examined at the Unit for
Reproductive Medicine of the University of Veterinary Medicine Hannover, Germany,
during five estrus cycles in the breeding season 2007. Four mares were nulli-, one
uni-, and one pluripar. They were kept in barns and on pasture and were fed with hay
and oats. This animal study was approved by the Hannover Regional Administration
(reference number: 33.12-42502-04-07/1235) and conducted in compliance with the
German Protection of Animals Act (TierschG).
9.1.2 Mare management
Uterine swabs were collected transcervically from all mares at the start of the study
for bacteriological and cytological assessments. Mares were teased daily using a
stallion and were examined daily using B-mode sonography. Estrus was defined as
the time period during which mares showed behavioral signs of estrus and
endometrial edema on sonographic B-mode images. The day of ovulation (Day 0)
was defined as the day of disappearance of the preovulatory follicle or, if there were
multiple follicles, as the day of disappearance of the first follicle.
9.1.3 Study design
The mares were examined during five estrus cycles (Table 9-1). Each mare received
no treatment in the first cycle (c1); treatment with equine FSH (eFSH®, Bioniche-
Animal Health, Athens, GA, USA), insemination, and embryo flushing in the second
cycle (c2); no treatment in the third cycle (c3); and treatment with eFSH®,
insemination, and embryo flushing in the fourth cycle (c4). In the fifth cycle (c5),
mares were also inseminated and embryo flushing was performed, but without any
stimulation of multiple ovulations. Mares were treated with hCG to induce ovulation.

Appendix 85
Following embryo recovery after stimulated cycles (SC), the mares received
prostaglandin on D 6.5 (250 μg of cloprostenol, Estrumate®, Essex-Tierarznei,
Munich, Germany) to induce luteolysis. The short cycles following luteolysis were not
used for examination, and after the next ovulation c3 and c5 were continued.
Color Doppler assessments of uterine and ovarian blood flow were performed on
every cycle day starting at a follicular size of 22 mm until ovulation. Blood samples
were taken after each examination. For better comparison of data, results were
presented for the first three days of the follicular development phase (FDP) t1 to t3
(t1=starting with a follicle diameter of 22 mm, and after the first eFSH treatment in
c2/c4, which were on different days of the cycle in each mare). For comparison of
genital blood flow in the preovulatory phase (POP), data of cycle days -4 to -1 (D-4 to
D-1) were compared.
Tab. 9-1: Overview of the five investigated estrus cycles and the according treatments (eFSH®-treatment, induction of ovulation with hCG, insemination and embryo collection 6.5 days post OV respectively)
cycle eFSH®
treatment
Induction of
ovulation with hCG
insemination embryo collection 6.5
days post OV
1 - - - -
2 + + + +
3 - - - -
4 + + + +
5 - + + +
9.1.4 Superovulation treatment
In stimulation cycles, mares were treated with 12.5 mg of eFSH® im (lateral neck
musculature) twice daily (7 a.m. and 7 p.m.), starting when follicular diameter was
>22 mm. On the second day of eFSH® therapy (following the third eFSH injection),
prostaglandin (250 μg of cloprostenol, Estrumate®, Essex-Tierarznei, Munich,
Germany) was administered im (lateral neck musculature). Treatment with eFSH®
was continued until follicle(s) were 32-35 mm in diameter (duration between 2.5 - 8.5
days; Ø 4.0 ± 1.6 days of FSH® treatment; Ø 100 ± 40 mg eFSH® in total ). The
mares were subsequently allowed to “coast” for 36 h, and were then injected with

86 Appendix
2500 IU of iv hCG (Ovogest®, Intervet, Unterschleissheim, Germany) to induce
ovulation. Mares were inseminated with cooled-stored semen (750 x 106
progressively motile spermatozoa, 20 ml in skin-milk extender INRA 82 collected
from one fertile warmblood stallion on the same day) 12 and 36 h after hCG injection
(Fig. 9-1). Insemination was performed using a sterile lubricated gloved arm guiding
the insemination pipette (Besamungspipette Universal für Pferde, Minitüb GmbH,
Tiefenbach, Germany) transcervically into the uterine body. The plunger of the
syringe was slowly depressed, introducing the semen.
D4-5 D6.5
OV foll.-Ø until foll.-Ø 36h “coasting“ OV
≥22-25mm ≥32-35mm ↑ ↑ ↑ ↑ ↑ ↑ ↑ ↑ ↑ ↑ ↑ eFSH
® (2xdaily 12.5mg) 2500IU 1.ins. 2.ins. embryo
PGF2α at 2. treatment day hCG iv +12h +36h collection
Fig. 9-1: eFSH® treatment protocol in cycle 2 and 4 (D= day, foll= follicle, OV=
ovulation, ins.= insemination)
9.1.5 Embryo collection
Transcervical embryo collection in cycles 2, 4 and 5 was performed by two persons
(JP, JK) 6.5 days after ovulation using a standard non-surgical procedure (IMEL et al.
1981). A balloon catheter (Ø 32 Ch., Embryospülkatheter, Minitüb, Tiefenbach,
Germany) was introduced into the uterus and the uterus was flushed four times with
1 L of 37 °C warm Dulbecco´s phosphate buffered saline (PBS) supplemented with
0.1 g/L bovine serum albumin fraction V (Sigma-Aldrich, Taufkirchen). The solution
was collected in a Petri dish (Miniflush Schale®, Minitüb, Tiefenbach, Germany) after
being filtered through a filter (Miniflush Filter®, pore size 73 µm Minitüb, Tiefenbach,
Germany). The collected fluid was inspected under a transmitted-light microscope
(Wild Hamburg, Germany) with 20 to 115 fold magnification. If not all expectable
embryos were found in the flush a second flush with four times of 1 l Dulbecco´s PBS
solution was performed directly following injection of 20 IU oxytocin iv (Oxytocin® Fa.
WDT, Garbsen, Germany). Oocytes were not included in the research.

Appendix 87
9.1.6 Transrectal Doppler sonography
In all cycles, examinations were carried out daily (starting at a follicle diameter of
22 mm) at the same time of day (09.00 to 11.00 a.m.). Doppler measurements were
always performed by the same operator (JP) using an ultrasound device (Logic P5,
GE Healthcare, Solingen, Germany) equipped with a 4 to 10 MHz linear probe (I 739,
GE Healthcare, Solingen, Germany). Collected data were digitized on DVDs
(Verbatim, 4.7 GB, Eschborn, Germany) and saved with e-Film Workstation 1.5.3 (E-
Film Medical Inc. Toronto, Canada) using the software program Pixelflux Scientific
(Chameleon-Software, Leipzig, Germany).
To measure uterine blood flow by transrectal ultrasonography, uterine arteries were
located in B-mode (brightness modulation). At the division from the aorta the A. iliaca
externa was followed latero-ventrally until the A. circumflexa iliumprofunda and the A.
uterina separated from the A. iliaca externa. The A. uterina was measured in
diameter shortly after the origin of the A. uterina.
The Aa. ovaricae were found following the ovarian vessel convolute in the
mesovarium. The A. ovarica was followed retrograde, and proximal of the separation
of the Ramus uterinus of the A. ovarica the diameter of the A. uterina was measured.
Following the measurement in B-mode, the CF-mode (continuous-wave Doppler)
was activated and the probe was positioned that the maximum blood flow was
visible. Using the PW-mode (pulsed-wave Doppler) several Doppler waves were
documented. For analysis of uterine and ovarian blood flow, the values of two similar
uniform consecutive pulse waves were averaged using the software program
PixelFlux Scientific (Chameleon-Software, Leipzig, Germany). Ovarian and uterine
blood flow were characterized by parameters pulsatility index (PI) and blood flow
volume (BFV), which were described in detail in earlier studies for ovarian
(HONNENS et al. 2009; BOLLWEIN et al. 2004) and uterine blood flow (HONNENS
et al. 2009; BOLLWEIN et al. 1998, 2000). The program PixelFlux Scientific
calculates the peak systolic velocity, the end diastolic velocity and the time averaged
maximum velocity over the time of a cardiac cycle (TAMV). From these data the
pulsatility index (PI) is calculated. Using the value of TAMV and the vessels diameter
the blood flow volume (BFV) can be calculated.
88 Appendix
Fig. 9-2: Ultrasound investigation of the left A. uterina of a mare one day before ovulation 1: A. uterina identification in B-mode, 2: measurement of the arteries diameter, 2: uterina blood flow using CF-mode, 4: measurement in PW-mode
Fig. 9-3: Ultrasound investigation of the right A. ovarica of a mare two days before ovulation 1: ovarian vessels convolute in the mesovarium in CF-mode, 2: measurement of the arteries diameter, 2: ovarian blood flow using CF-mode, 4: measurement in PW-mode
vessels convolute A. ovarica

Appendix 89
9.1.7 Blood collection, progesterone and estrogen analysis
EDTA-blood samples were obtained from the jugular vein using a vacutainer system
(Becton Dickinson Vacutainer System containing EDTA, Belliver Industrial Estate,
Plymouth, UK) after each scan. Plasma was separated after 2 h (centrifugation for
10 min at 1800 x g) and frozen in Sarstedt-tubes (Sarstedt AG & Co/ Nümbrecht) at -
20°C until analysis of plasma progesterone (P4) and total estrogen (Etot) levels.
Samples were assayed in the endocrinologic laboratory of the Clinic for Cattle,
School of Veterinary Medicine Hannover, Germany. They were analysed in duplicate,
and a separate assay was performed for each trial.
P4 was estimated by enzyme immunoassay (EIA) as published earlier (PRAKASH et
al. 2000). The intra- and inter-assay coefficients of variation were 2.7% and 3.8%,
respectively.
Plasma levels of Etot were estimated following hydrolysis and extraction by EIA as
described previously (MEYER et al. 2000). The intra- and inter-assay coefficients of
variation were 5.4% and 9.6%, respectively.
9.1.8 Statistical analysis
Statistical analysis was carried out using SPSS for Windows, version 15.0 (SPSS
Inc., USA). The Shapiro-Wilk test was used to test for normal distribution of data. All
data were presented as mean ± S.E.M. for better comparison. Student`s t-test
(normally distributed data) and the Mann-Whitney U-test (not normally distributed
data) were used to determine differences in measurements between groups and
days.
Correlations between BFV, PI values and number of follicles, ovulations, embryos
and plasma hormone levels were determined by the Pearson (normally distributed
data) or Spearman (not normally distributed data) correlation coefficient (r).
Differences and correlations with p<0.05 were considered to be statistically
significant.

90 Appendix
9.2 Material and methods study III
The study was performed in the Clinic for Horses and Unit for Reproductive
Medicine, University of Veterinary Medicine Hannover, Germany. Experiment I was
conducted between July 2007 and September 2007 and experiment II between
November 2008 and June 2009. This research was conducted in accordance with
Animal Welfare (No. 33.12-42502-04-07/1235).
9.2.1 Animals
Ten mares (3-16 years old) were examined and randomized by replicate into two
groups for experiment I (8 surgeries) and II (12 surgeries). Two mares underwent
both experiments (Table 9-3). For both procedures feed was withheld for 24 hours
preoperatively. Animals had full access to water. The mares were judged to be in
good physical health on the basis of an initial physical examination. Gynaecologic
examinations were performed before surgery to assess the day of the oestrus cycle.
The time interval between surgeries in the same mare was between 3 and 102 days
(Ø 45 days).
9.2.2 Preoperative management
For both experiments, mares received a central venous catheter (Vygonüle T®,
Vygon, Aachen, Germany) in the jugular vein for the duration of the preoperative
preparations and surgery time (experiment I: 81.25 min, experiment II Ø 26.8 min).
Antimicrobial therapy was provided with sodium benzylpenicillin (Penicillin
„Grünenthal“ 10 Mega®, Grünenthal, Aachen, Germany, 22.000 IU/kg iv) and
gentamicin (Genta 100®, cp-pharma, Burgdorf, Germany, 6.6 mg/kg sid, iv)
administered as a single dose immediately preoperatively. Flunixin meglumine
(Flunidol®, cp-pharma, Burgdorf, Germany, 1.1 mg/kg bwt, iv) was given to provide
perioperative analgesia. Post operative analgesia was maintained with flunixin
meglumine (Flunidolgel®, cp-pharma, Burgdorf, Germany, 1.1 mg/kg bwt, per os) for
48 hours or longer as required.

Appendix 91
The mares were restrained in stocks and sedated with a combination of detomidine
hydrochloride (Domosedan®, Pfizer, Karlsruhe, Germany, 10 μg/kg) and butorphanol
tartrate (Torbugesic®, Fort Dodge, Würselen, Germany, 10 μg/kg) administered
intravenously. Detomidine and butorphanol (0.5–1 μg/kg, iv) were administered
repeatedly as needed during surgery.
9.2.3 Instruments used in experiment I
1) Universal tube (Ø 12.5 mm, length 52.0 cm STORZ, Tuttlingen, 66016 AB)
2) mandrin with truncated tip (Ø 10.0 mm, length 52.0 cm, STORZ, Tuttlingen)
3) sharp mandrin (Ø 10.0 mm, length 52.0 cm, STORZ, Tuttlingen)
4) tubing system with two channels for catheter and laparoscope (STORZ, Tuttlingen)
5) laparoscope (Ø 10.0 mm, length 57.0 cm, 30° forward Hopkins optic, STORZ,
Tuttlingen, 6015 BV) All instruments are shown in Fig. 9-4.
Fig. 9-4: Trocar set used for experiment I, 1) Universal tube, mandrin with truncated tip, 3) sharp mandarin, 4) tubing system with two channels, 5) laparoscope
Furthermore were used a
KARL STORZ Endovision TELECAM, color system PAL (STORZ) containing of:
- TELECAM pal camera head (STORZ 20210030)
- TELECAM pal camera-control unit (STORZ 20210020)
- Digivideo picture processing system (STORZ 20202020)
ø 12,5 mm 52 cm length
30 °
1)
2)
3)
4)
5)

92 Appendix
- cold light source Xeno n 300 (STORZ 20133020)
- glass fibre-light cable (STORZ 495NE, Ø 3.5 mm, lenght3 m)
- 36 cm color monitor (Sony, Trinitr on color video Monitor, PVM-2053MD)
- Video-recorder (Panasonic, Viddeo Cassette Recorder NVFS88 HQ)
9.2.4 Surgery experiment I
For experiment I (n=8), mares were fixed in a stock, the tail of the mares were fixed
and secured with a bandage and the stock was covered with a sterile sheet (Buster®-
OP-Cover, 90 cm x 120 cm). Following manual evacuation of the rectum, the outer
genital were prepared antiseptically using a soap (Lifosan soft®, B. Braun, Melsungen,
Germany) and iodine (Braunol®, Braun, Melsungen, Germany). The vestibulum and
vagina were flushed with 500 ml of an iodine solution (NACL 0.9%, Ecotainer® ,
Braun, Melsungen, Germany) and (0.5 % iodine solution (Braunol® , B. Braun,
Melsungen, Germany) by inserting the left hand with a sterile glove and a flushing
tube into the vagina.
In the first two surgeries, the mares received an epidural anaesthesia with
lidocainhydrochloride 2% (Lidocain®, belapharm, Vechta, Germany).
The trocar set (Storz, Tuttlingen, Germany) consisting of a universal tube (12.5 mm x
52 cm) and a mandrin with a truncated tip were hold in the operators right hand,
covered with a sterile glove an were inserted and positioned middorsally in the fornix
of the vagina. The trocar was fixed by the operator’s left hand and the right hand was
inserted into the rectum. Following a transrectal manual control of the correct position
of the trocar and that no intestine was in front of the tip of the trocar, the atraumatic
mandrin was replaced by a sharp mandarin. The trocar set was introduced through
the vaginal wall into the peritoneal cavity, which was confirmed by passive influx of
air. The mandrin was replaced by a tubing system, containing the laparoscope
(Storz, Tuttlingen, Germany, Ø 10 mm, 30° forward Hopkins optic) and a 6 Ch.
flushing catheter (ERU®, Willy RÜSCH GmbH, Kernen i. R., Germany) in the working
channel. The ovaries were fixed by manipulation per rectum, and were turned 180° in
cranio-medial direction so that the infundibulum and the adjacent part of the ampulla
could be clearly identified. An attempt was then made to introduce the catheter into

Appendix 93
the abdominal ostium. The instruments were removed and the mare was fastened in
the stable for 2 hours to prevent aspiration due to the sedation.
9.2.5 Preoperative management experiment II
For experiment II (n=12) the paralumbar fossa was clipped and prepared aseptically
(Hibiscrub®, Chlorhexidine gluconate, Regent Medical, irlam, Manchester, UK). After
the infiltration of skin and muscle at the surgery side with 80 - 100 ml lidocaine
(Lidocain®, belapharm, Vechta, Germany) the mare and stock were covered with
sterile sheets (Buster®-OP-Cover, 90 cm x 120 cm).
9.2.6 Instruments used in experiment II
The following instruments were needed or developed for experiment two:
Endoscope-set (KARL STORZ, Tuttlingen, Germany) containing:
- 4 trocar sleeves, one with automatic valve for CO2–insufflation (Ø12.5 mm,
3 x lengths 20 cm, 1x length 15 cm, STORZ, 62103H3)
- trocar with truncated tip (Ø 10.0 mm, length 28 cm, STORZ, 62103 AL)
- laparoscope with a 30° viewing angle (Ø 10 mm, length 57 cm, Hopkins® optic,
62032 BP, STORZ)
KARL STORZ Endovision TELECAM, color system PAL (STORZ) containing:
- TELECAM pal camera head (STORZ, 20210030)
- TELECAM pal camera-control unit (STORZ, 20210020)
- Digivideo picture processing system (STORZ, 20202020)
- cold light source Xeno n 300 (STORZ, 20133020)
- glass fibre-light cable (STORZ, 495NE, Ø 3.5 mm, length 3 m)
- 36 cm color monitor (Sony, Trinitron color video Monitor, PVM-2053MD)
- video-recorder (Panasonic, Video Cassette Recorder NVFS88 HQ)
- CO2-insufflator with insufflation-tube (DVM-medizintechnik GmbH Ludwigsfelde
95C8032)
- sterile filter (mtp medical technical promotion GmbH, Neuhausen, RGF 031122-02)
- 2 Babcock forceps (atraumatic, Ø 10 mm, length 43 cm, Click’line ®, STORZ)

94 Appendix
- 2 scalpel handles with blades (size 22 and 10)
- injection filter (Sterifit 0.2 Ym, Braun, Melsung, Germany)
- suture material (Difalon 1 USP, Braun, Tuttlingen, Germany)
- surgical instrument set (needle holder, clamps, scissors)
- methylene blue solution:
(2 g of methylene blue powder (Microscopy Certistain Methylenblau, MERCK
KGaG, Darmstadt, Germany) were given in 1 L 0.9% NaCl solution (NaCl 0.9%
solution Ecotainer 1 L, Braun, Melsungen, Germany)
40 ml of the solution were filtered through an injection filter (Sterifit 0.2 Ym®, Braun,
Melsung, Germany) and aspirated into a 10 ml syringe.
- balloon catheter (Fig. 9-5):
ureter catheter with balloon after Steffens and Vahlensiek, Ch. 7, 75 cm length,
PVC material (26 30 00, Willy RÜSCH GmbH, Kernen i. R., Germany) with a two
channel system (one channel for balloon insufflation of 5 ml air and one for fluid
application, “flute-tip” with three “eyes”, the one in front of the balloon was closed by
a 2 cm x 15 mm piece of PVC tape (Coronaplast®, Conmetall, Celle, Germany)
Fig. 9-5: Picture of the flushing catheter a: catheter with three openings, one closed by a PVC tape, b: 5 ml syringe, c: balloon insufflated with 3 ml of air
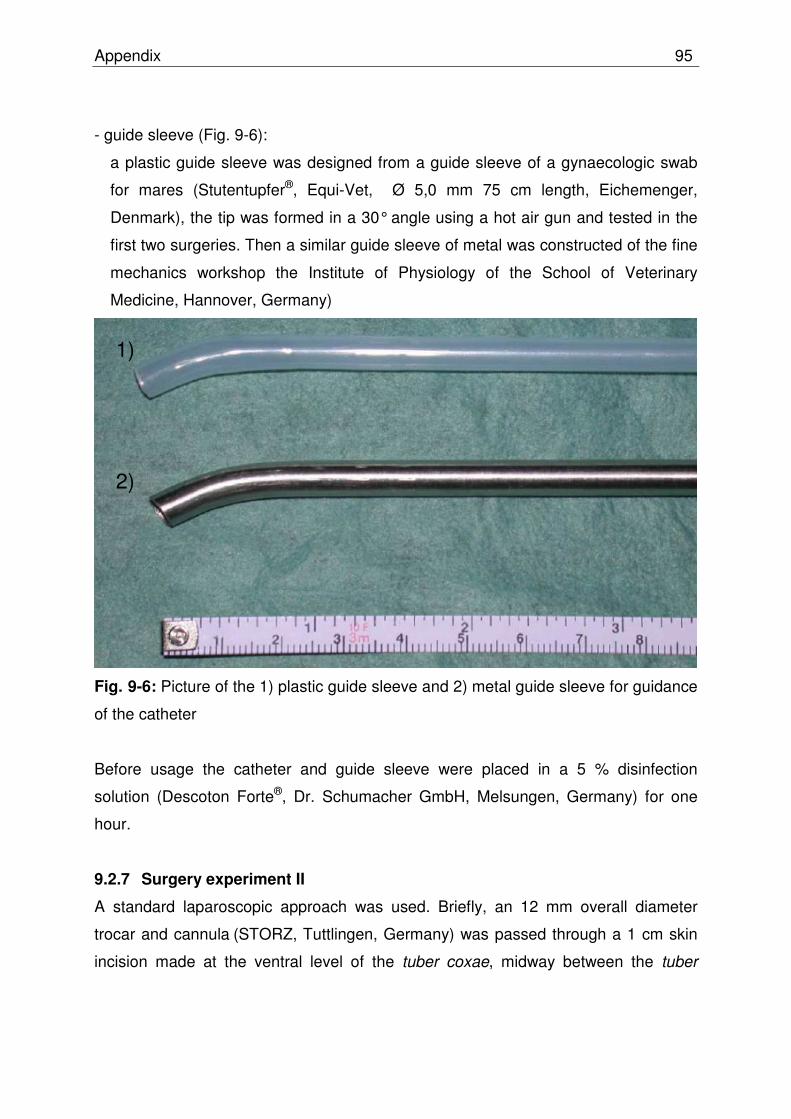
Appendix 95
- guide sleeve (Fig. 9-6):
a plastic guide sleeve was designed from a guide sleeve of a gynaecologic swab
for mares (Stutentupfer®, Equi-Vet, Ø 5,0 mm 75 cm length, Eichemenger,
Denmark), the tip was formed in a 30° angle using a hot air gun and tested in the
first two surgeries. Then a similar guide sleeve of metal was constructed of the fine
mechanics workshop the Institute of Physiology of the School of Veterinary
Medicine, Hannover, Germany)
Fig. 9-6: Picture of the 1) plastic guide sleeve and 2) metal guide sleeve for guidance
of the catheter
Before usage the catheter and guide sleeve were placed in a 5 % disinfection
solution (Descoton Forte®, Dr. Schumacher GmbH, Melsungen, Germany) for one
hour.
9.2.7 Surgery experiment II
A standard laparoscopic approach was used. Briefly, an 12 mm overall diameter
trocar and cannula (STORZ, Tuttlingen, Germany) was passed through a 1 cm skin
incision made at the ventral level of the tuber coxae, midway between the tuber
1)
2)
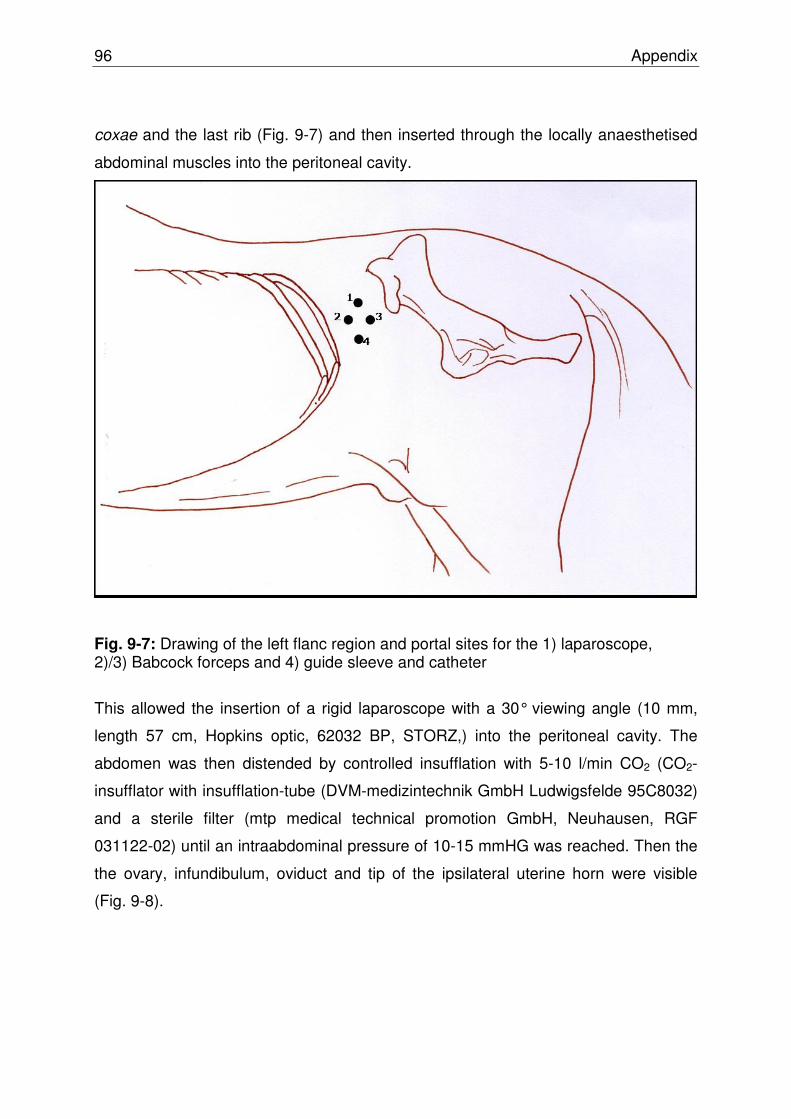
96 Appendix
coxae and the last rib (Fig. 9-7) and then inserted through the locally anaesthetised
abdominal muscles into the peritoneal cavity.
Fig. 9-7: Drawing of the left flanc region and portal sites for the 1) laparoscope, 2)/3) Babcock forceps and 4) guide sleeve and catheter
This allowed the insertion of a rigid laparoscope with a 30° viewing angle (10 mm,
length 57 cm, Hopkins optic, 62032 BP, STORZ,) into the peritoneal cavity. The
abdomen was then distended by controlled insufflation with 5-10 l/min CO2 (CO2-
insufflator with insufflation-tube (DVM-medizintechnik GmbH Ludwigsfelde 95C8032)
and a sterile filter (mtp medical technical promotion GmbH, Neuhausen, RGF
031122-02) until an intraabdominal pressure of 10-15 mmHG was reached. Then the
the ovary, infundibulum, oviduct and tip of the ipsilateral uterine horn were visible
(Fig. 9-8).
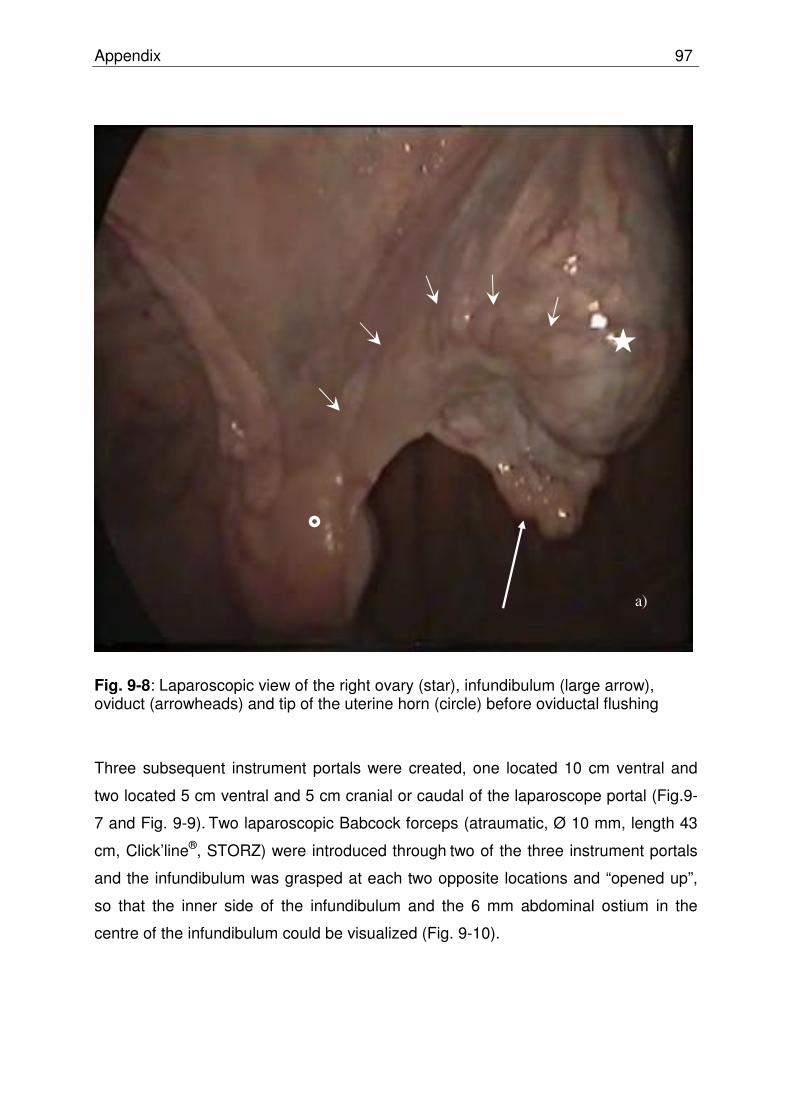
Appendix 97
Fig. 9-8: Laparoscopic view of the right ovary (star), infundibulum (large arrow), oviduct (arrowheads) and tip of the uterine horn (circle) before oviductal flushing
Three subsequent instrument portals were created, one located 10 cm ventral and
two located 5 cm ventral and 5 cm cranial or caudal of the laparoscope portal (Fig.9-
7 and Fig. 9-9). Two laparoscopic Babcock forceps (atraumatic, Ø 10 mm, length 43
cm, Click’line®, STORZ) were introduced through two of the three instrument portals
and the infundibulum was grasped at each two opposite locations and “opened up”,
so that the inner side of the infundibulum and the 6 mm abdominal ostium in the
centre of the infundibulum could be visualized (Fig. 9-10).
a)
�
�
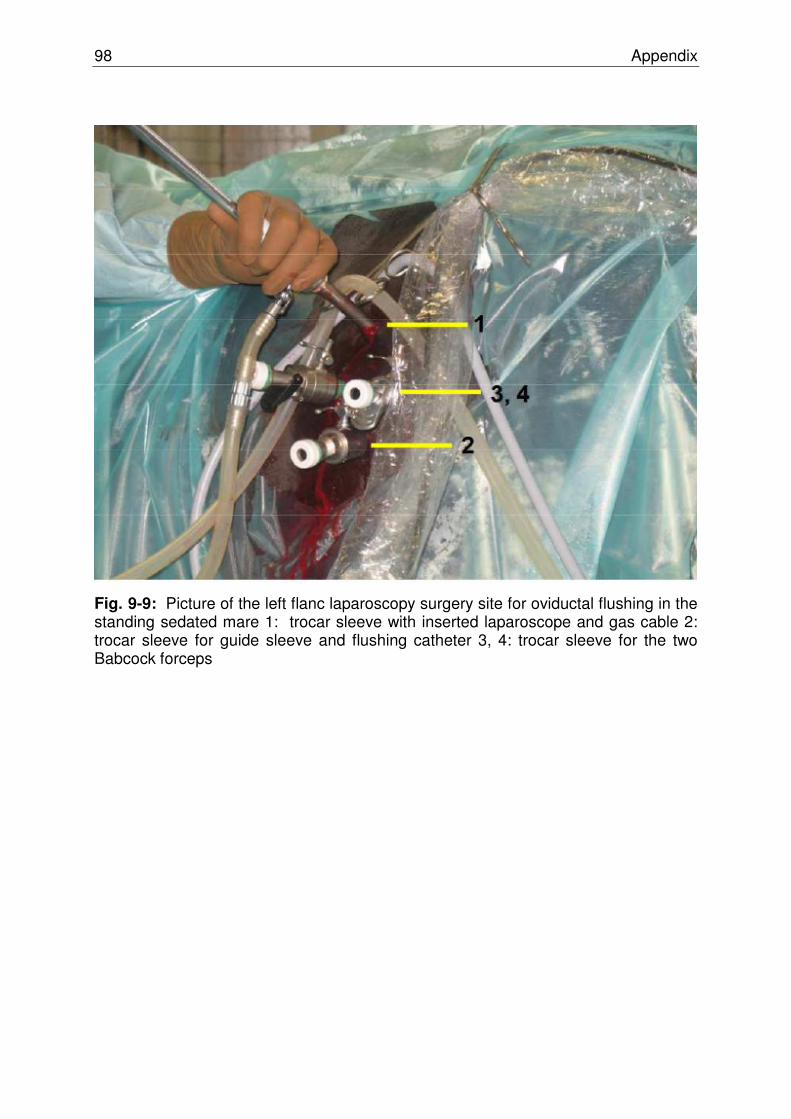
98 Appendix
Fig. 9-9: Picture of the left flanc laparoscopy surgery site for oviductal flushing in the standing sedated mare 1: trocar sleeve with inserted laparoscope and gas cable 2: trocar sleeve for guide sleeve and flushing catheter 3, 4: trocar sleeve for the two Babcock forceps

Appendix 99
Fig. 9-10: Laparoscopic view of the left part of the infundibulum „opened up“ by a Babcock forceps; the balloon catheter is directed into the abdominal ostium (large arrow)
A 7 Ch. balloon catheter (ureter catheter with balloon after Steffens and Vahlensiek,
Ch. 7, Art. Nr. 26 30 00, Willy RÜSCH GmbH, Kernen i. R., Germany) inserted in the
guide sleeve which was curved at the tip was positioned into the lower instrument
portal and directed into the abdominal ostium of the oviduct and approximately 2 cm
into the ampulla. The balloon was inflated with 2 ml of air. One of the Babcock
forceps holding the infundibulum was removed and positioned around the abdominal
ostium and the catheter to secure the catheter in place and to avoid a back pressure
of the catheter (Fig. 9-11).
a)
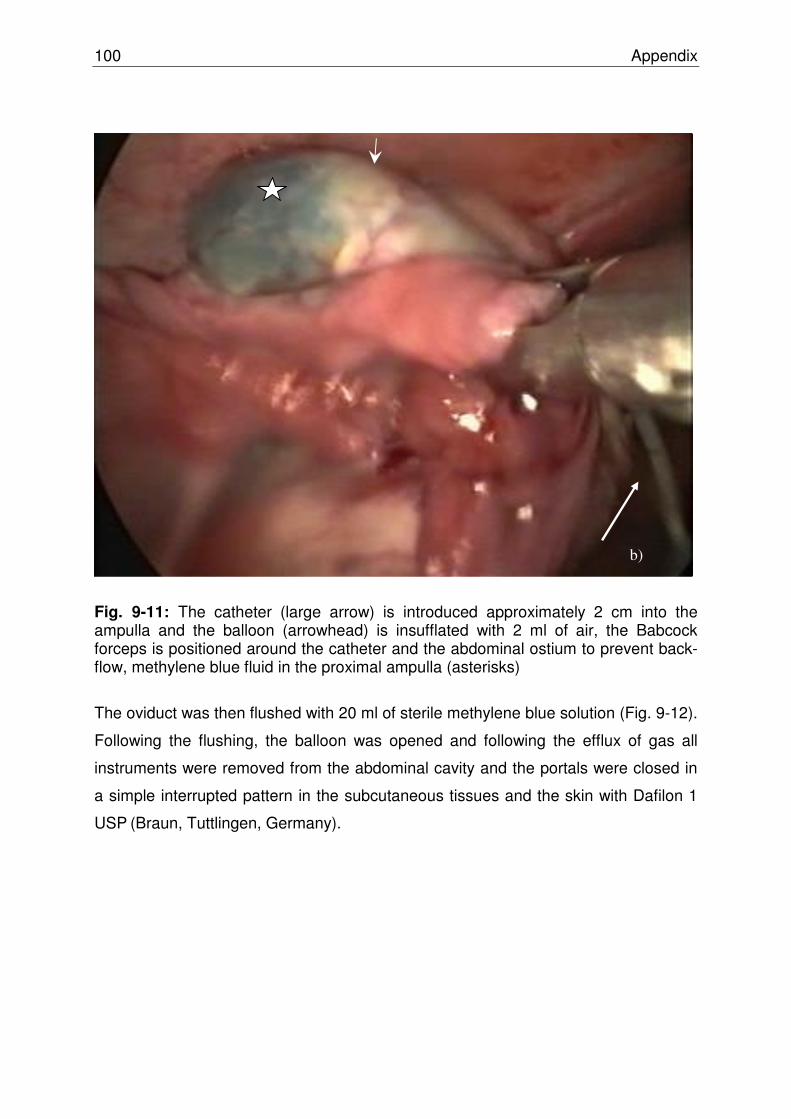
100 Appendix
Fig. 9-11: The catheter (large arrow) is introduced approximately 2 cm into the ampulla and the balloon (arrowhead) is insufflated with 2 ml of air, the Babcock forceps is positioned around the catheter and the abdominal ostium to prevent back-flow, methylene blue fluid in the proximal ampulla (asterisks)
The oviduct was then flushed with 20 ml of sterile methylene blue solution (Fig. 9-12).
Following the flushing, the balloon was opened and following the efflux of gas all
instruments were removed from the abdominal cavity and the portals were closed in
a simple interrupted pattern in the subcutaneous tissues and the skin with Dafilon 1
USP (Braun, Tuttlingen, Germany).
b)
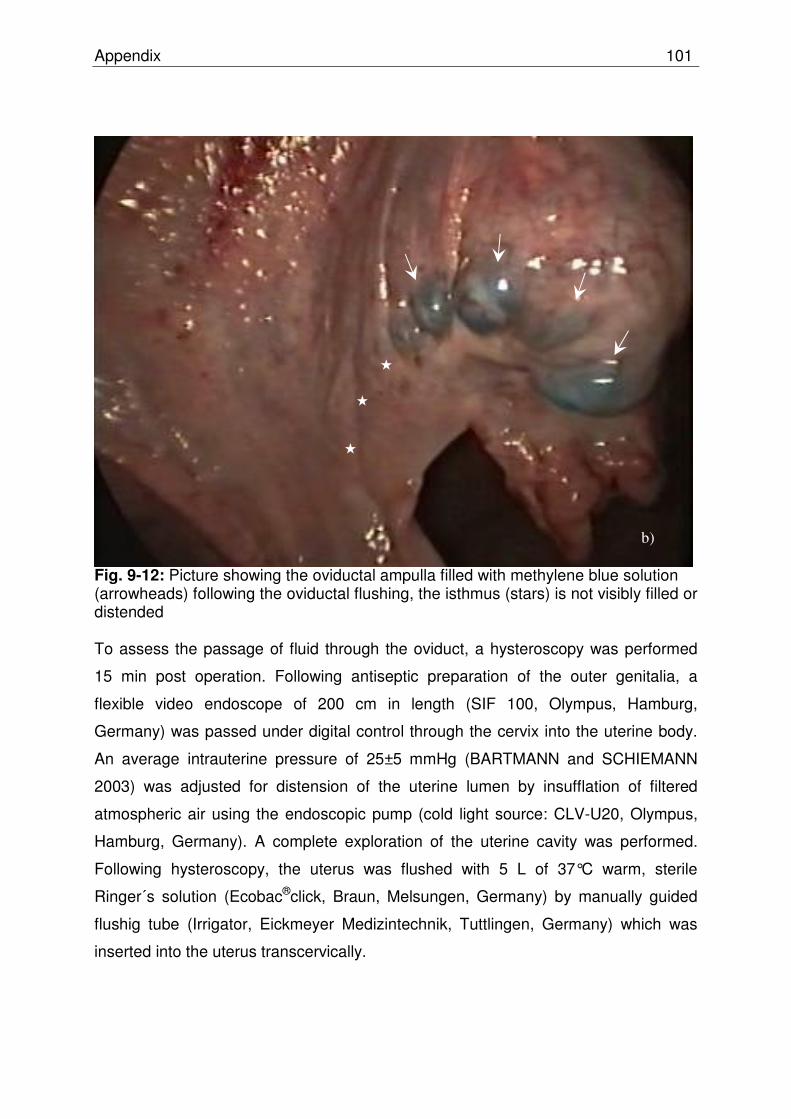
Appendix 101
Fig. 9-12: Picture showing the oviductal ampulla filled with methylene blue solution (arrowheads) following the oviductal flushing, the isthmus (stars) is not visibly filled or distended To assess the passage of fluid through the oviduct, a hysteroscopy was performed
15 min post operation. Following antiseptic preparation of the outer genitalia, a
flexible video endoscope of 200 cm in length (SIF 100, Olympus, Hamburg,
Germany) was passed under digital control through the cervix into the uterine body.
An average intrauterine pressure of 25±5 mmHg (BARTMANN and SCHIEMANN
2003) was adjusted for distension of the uterine lumen by insufflation of filtered
atmospheric air using the endoscopic pump (cold light source: CLV-U20, Olympus,
Hamburg, Germany). A complete exploration of the uterine cavity was performed.
Following hysteroscopy, the uterus was flushed with 5 L of 37°C warm, sterile
Ringer´s solution (Ecobac®click, Braun, Melsungen, Germany) by manually guided
flushig tube (Irrigator, Eickmeyer Medizintechnik, Tuttlingen, Germany) which was
inserted into the uterus transcervically.
b)
�
�
�

102 Appendix
9.2.8 Postoperative care in both experiments
The central venous catheter was removed and mares were examined in their stable
6, 12 and 18 hours post surgery (day 0) for signs of postoperative pain. Using a pain-
score system (see Table 9-2) the sum of scores were used. A total score of 0-4 was
defined as not painful, 5-8 as slight painful, 9-17 as moderate painful and 18-24 as
highly painful. Additionally, a clinical examination containing behaviour, appetite,
pulse (beats/min), respiration (breaths/min), rectal temperature, gut sounds, capillary
refill time, mucous membranes was performed daily from days 0-5. At days 1, 3 and
5 evaluations of an EDTA-blood sample was performed. The values in brackets were
defined as normal. Haematocrit (25-45%), total protein (55-75 g/l), leucocytes (5-10
G/l). Administration of flunixin meglumine (Flunidolgel 1.1 mg/kg bwt, per os) was
continued for 48 hours.
Tab. 9-2 Pain score system parameter/points 0 1 2 3
behaviour alert calm dull colic, apathy
appetite good not eating
defecation existing not existing,
diarrhoea pulse (beats/min)
28-40 >40 >52 >60
respiration (breaths/min)
<16 >16 >32 >40
abdominal wall tension
relaxed slightly tense moderately tense
highly tense
pressure pain of wound
insensible slightly painful moderately painful
highly painful
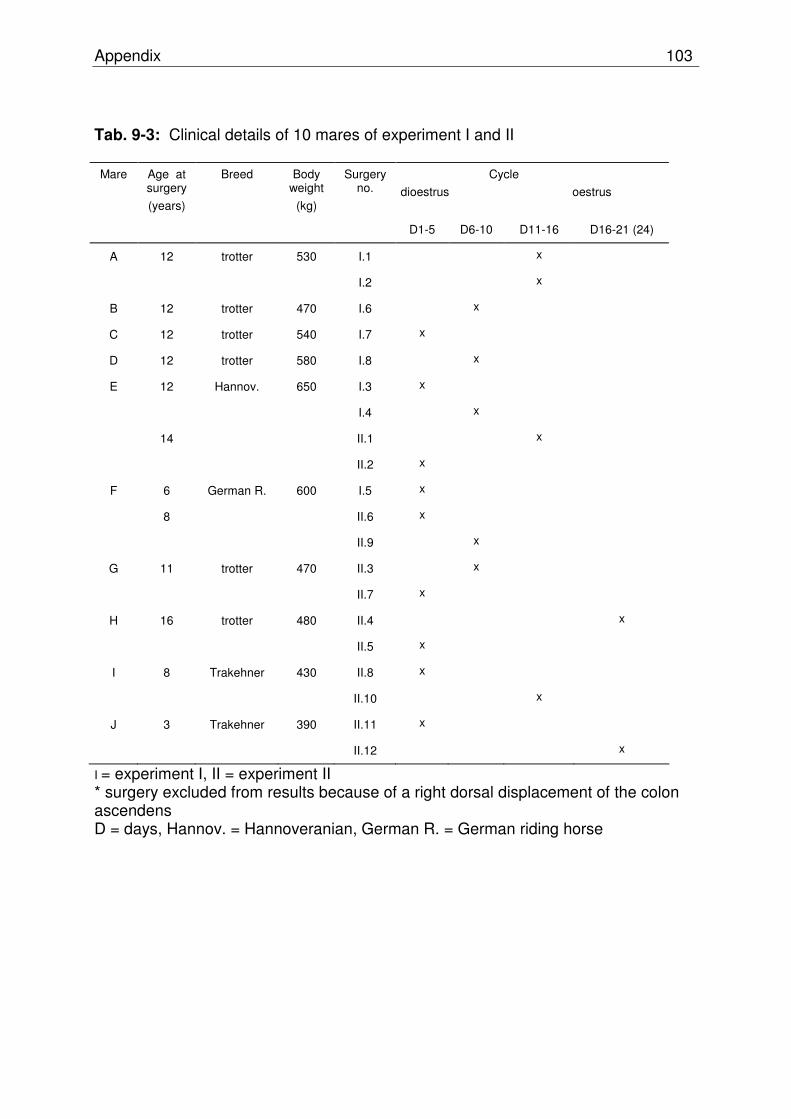
Appendix 103
Tab. 9-3: Clinical details of 10 mares of experiment I and II Mare Age at
surgery
(years)
Breed Body weight
(kg)
Surgery no.
Cycle
dioestrus oestrus
D1-5 D6-10 D11-16 D16-21 (24)
A 12 trotter 530 I.1 x
I.2 x
B 12 trotter 470 I.6 x
C 12 trotter 540 I.7 x
D 12 trotter 580 I.8 x
E 12 Hannov. 650 I.3 x
I.4 x
14 II.1 x
II.2 x
F 6 German R. 600 I.5 x
8 II.6 x
II.9 x
G 11 trotter 470 II.3 x
II.7 x
H 16 trotter 480 II.4 x
II.5 x
I 8 Trakehner 430 II.8 x
II.10 x
J 3 Trakehner 390 II.11 x
II.12 x
I = experiment I, II = experiment II * surgery excluded from results because of a right dorsal displacement of the colon ascendens D = days, Hannov. = Hannoveranian, German R. = German riding horse

104 Appendix
Composition of skin-milk extender INRA 82 (page 85) 25 g glucose 1.5 g lactose 1.5 g raffinose 0.4 g potassium-citrate 0.3 g tri-sodium-citrate-dihydrat 4.76 g HEPES 0.5 g penicillin 0.5 g gentamicin ad 0.5 l aqua bidest. ad 1000 ml skin milk (0.3%)
Composition of phosphate buffered saline, PBS (page 85) Solution 1: 64 g NaCl 1.6 g KCl 0.8 g MgCl26H2O 0.8 g CaCl2 in aqua bidest ad 1000 ml Solution 2: 1.2 g NaH2PO4H2O 10 g Na2HPO42H2O 1.6 g KH2PO4 9.8 g glucoseH2O 0.288 g pyruvat 0.8 g bovine sermalbumine fraktion V (Sigma Aldrich,Taufkirchen, Germany) in auqa bidest ad 7000 ml Mixture of solution 1 and 2 (8 L) , 37-38°C warm

Appendix 105
Erklärung
Hiermit erkläre ich, dass ich die Dissertation “Investigations on genital blood flow and
embryo recovery after superovulation with eFSH® and on laparoscopic techniques for
flushing the oviduct in the mare” selbständig verfasst habe.
Ich habe keine entgeltliche Hilfe von Vermittlungs- bzw. Beratungsdiensten
(Promotionsberater oder anderer Personen) in Anspruch genommen. Niemand hat
von mir unmittelbar oder mittelbar entgeltliche Leistungen für Arbeiten erhalten, die
im Zusammenhang mit dem Inhalt der vorgelegten Dissertation stehen.
Ich habe die Dissertation an folgender Institution angefertigt:
Klinik für Pferde und Reproduktionsmedizinische Einheit der Kliniken der Stiftung
Tierärztliche Hochschule Hannover.
Die Dissertation wurde bisher nicht für eine Prüfung oder Promotion oder für einen
ähnlichen Zweck zur Beurteilung eingereicht.
Ich versichere, dass ich die vorstehenden Angaben nach bestem Wissen vollständig
und der Wahrheit entsprechend gemacht habe.
Hannover, den Melanie Witt

106 Appendix
Acknowledgements
I express my gratitude to my supervisor Prof. Dr. Harald Sieme for entrusting me with
this interesting work and for his continuous academic guidance, confidence,
motivation, patience and good humor.
I thank Prof. Dr. Sabine Meinecke-Tillmann and Prof. Dr. Wilhelm Kanitz, from my
tutorial group, for thoughtful discussions, comments and suggestions.
I also thank the Mehl-Mülhens-Stiftung, Germany, who supported this project.
Many thanks to Dr. Anna Rötting, for the interesting discussions and qualified help in
many issues concerning surgery and laparoscopy and for the fact that she never lost
the belief those laparoscopies would be successful…
I thank Antje Heberling, Christine Baackmann and Jeannette Probst for working
together on the interesting projects.
I also thank to the National Stud Celle for semen collection any day of the week.
Thanks to the laboratory of the Clinic for Cattle, University of Veterinary Medicine
Hannover and JProf. Marion Piechotta for the help in hormone analysis.
Special thanks to Joachim König and Juliane Marx and all the Veterinarian
Technicians of the Clinic for horses for technical assistance and patience during
laparoscopies.
Thanks to Katharina Höffmann for demonstrating the technique of transvaginal tubal
flushing in the cow.
I thank all colleagues from the Clinic for horses and the Reproductive Unit of Clinics,
University of Veterinary Medicine Hannover for the good times we have had together.
I thank Astrid Bienert-Zeit, Jessica Cavalleri, Florian Geburek, Rolf Wagels, Anna
Rötting, Miriam v. Borstel, Jutta Klewitz and Frauke Uhlendorf for the great time in
the Clinic for horses, for their general help and fun and friendship!
Special thanks to my family, Lars, Lasse, Lena and Max, I love you!