comparison of chelated zinc and zinc sulfate as zinc ... · comparison of chelated zinc and zinc...
TRANSCRIPT

Aquaculture 406–407 (2013) 79–84
Contents lists available at SciVerse ScienceDirect
Aquaculture
j ourna l homepage: www.e lsev ie r .com/ locate /aqua-on l ine
Comparison of chelated zinc and zinc sulfate as zinc sources for growthand immune response of shrimp (Litopenaeus vannamei)
Shimei Lin ⁎, Xin Lin, Yang Yang, Fajian Li, Li Luoa Key Laboratory of Freshwater Fish Reproduction and Development (Ministry of Education), Southwest University, Chongqing 400716, PR Chinab College of Animal Science and Technology, Southwest University, Chongqing 400716, PR China
⁎ Corresponding author at: College of Animal SciencUniversity, Chongqing 400716, PR China. Tel./fax: +8
E-mail address: [email protected] (S. Lin).
0044-8486 © 2013 The Authors. Published by Elsevierhttp://dx.doi.org/10.1016/j.aquaculture.2013.04.026
a b s t r a c t
a r t i c l e i n f oArticle history:Received 14 January 2013Received in revised form 22 April 2013Accepted 22 April 2013Available online 29 April 2013
Keywords:ZincGrowthNon-specific immunityLitopenaeus vannamei
Zinc methionine (ZnMet), zinc lysine (ZnLys), zinc glycine (ZnGly) and zinc sulfate (ZnSO4 · H2O) were eval-uated as dietary zinc sources for Litopenaeus vannamei. Three Zn–amino acid complexes with a molar aminoacid to Zn ratio of 2:1 were compared to Zn sulfate using a casein-based purified diet. Five groups with fourreplicates of shrimps (mean weight 0.72 ± 0.02 g) were given a basal diet either unsupplemented (control)or supplemented with 30 mg Zn kg−1 sulfate (ZnSO4 · H2O) or the organic sources respectively, for12 weeks. Results showed that the source of Zn affects shrimp growth, survival and immune parameters.Shrimp fed diets with organic zinc supplementation produced significantly higher growth, survival and im-mune parameters than ZnSO4 treatment. Shrimp supplemented with ZnMet had the highest weight gainand immune parameters. However, there were no significant differences in weight gain, survival, total hemo-cyte counts, phagocytotic activity, PO, AKP and SOD between the ZnLys and ZnGly groups. Results suggestthat Zn from ZnMet was a better source than the other zinc forms.
© 2013 The Authors. Published by Elsevier B.V. Open access under CC BY-NC-ND license.
1. Introduction
Zinc (Zn) is an essential nutrient that is required in humans and an-imals for many physiological functions including growth, development,reproduction and immune function (Watanabe et al., 1997). Dependingon the doses and the chemical forms of Zn, it can act as nutrients, anti-oxidants, or even toxicants (Lemire et al., 2008). Zinc functions as acofactor in several enzyme systems and is a component of a large num-ber of metalloenzymes, which include carbonic anhydrase, alkalinephosphatase and DNA polymerases (NRC, 2011). However, deficientor excessive zinc has been reported to affect the fish morphology, bio-chemical processes and growth (Watanabe et al., 1997). Like otherheavy metals, deficient or excessive zinc could also exert inhibitory ef-fects on immune responses and increase the severity of infections inhumans and animals (Shankar and Prasad, 1998). The dietary Zn supplyto fish often largely exceeds their actual requirements. Poorly absorbed,Zn is highly concentrated in nature and may cause environmental pol-lution in areas of intensive aquaculture production (NRC, 2011). Ab-sorption of trace elements often limits their utilization. One of thefactors that affect mineral absorption and utilization is their chemical
e and Technology, Southwest6 23 68251196.
B.V. Open access under CC BY-NC-ND l
form. Hence, mineral sources with higher bioavailability should be con-sidered in feed formulation.
Due to higher bioavailability compared to inorganic salts, chelatedminerals as animal feed supplements have attracted considerable atten-tion of the feed manufacturers and the animal producers as a means ofimproving animal performance (Ashmead, 1992; Wang and Lovell,1997). Nowadays, chelated minerals are widely used in the livestockand poultry industries (Puchala et al., 1999; Swiatkiewicz et al., 2001;Wedekind et al., 1992). Some studies regarding chelated mineralshave already been conducted in either fishes (Apines-Amar et al.,2004; Paripatananont and Lovell, 1995; Satoh et al., 2001; Wang andLovell, 1997) or abalone (Tan and Mai, 2001), and beneficial effectson growth performance or immunity have been reported. As forLitopenaeus vannamei, there is little known about the effectiveness ofany chelated minerals including organic Zn sources at present.
Thewhite shrimp (L. vannamei) is one of themost commercially cul-tured shrimp species in South China (FAO, 2010). The shrimp culture in-dustry has often suffered economic losses attributed to outbreaks ofinfectious viral and bacterial diseases. The establishment of healthman-agement regimes and the selection of shrimp that are more resistant todiseases are facilitated through the characterization of effectors of theimmune system. Our understanding on the role of dietary nutrientson shrimphealth is largely based on nutrients such asminerals. Howev-er, potential benefits of organic Zn complexes on the immune functionof shrimp have not been critically evaluated. Thus, the current studywas designed to evaluate the application of different zinc sources, in-cluding inorganic Zn (zinc sulfate) and organic Zn (ZnMet, ZnLys andZnGly), as feed additives in diets for L. vannamei.
icense.

80 S. Lin et al. / Aquaculture 406–407 (2013) 79–84
2. Materials and methods
2.1. Experimental diets
The basal diet formulation is given in Table 1. It was formulatedwith purified ingredients to provide 41% crude protein from caseinand gelatin and 6.5% crude lipid from soybean oil and menhadenfish oil (1:2), which were sufficient to support optimal growth (Royet al., 2006). The composition of mineral premix was modifiedaccording to Kureshy and Davis (2002) without zinc supplementa-tion. The vitamin mixture was similar to that used by Kureshy andDavis (2002). Zinc methionine (ZnMet), zinc lysine (ZnLys), zinc gly-cine (ZnGly) and zinc sulfate (ZnSO4 · H2O) were used as dietary zincsources. The organic source of Zn was developed by Calcialiment(Changsha Xingjia Biotechnology Share Co., Ltd. Hunan, China) andis a Zn–amino acid complex with a molar amino acid to Zn ratio of2:1. Zinc from either sulfate (feed-grade; ZnSO4 · H2O) or the organicsource was added into the basal diet at levels of 30 mg Zn kg−1 dietaccording to NRC (2011), respectively, to prepare the four experimentaldiets. The control groups were basal diet only. The Zn concentrations ofthe basal diets were analyzed by flame photometry after wet decompo-sition according to AOAC (1995) and found to be 6.5 mg Zn/kg diet.
The ingredients were ground in a Hammer mill until they passedthrough a 60-mesh screen. Experimental diets were prepared by thor-oughly mixing the dry ingredients with oil and then adding cold wateruntil a stiff dough resulted. This was then passed through a mincerwith die and the resulting “spaghetti-like” strings were dried using anelectrical fan at 40 °C. After drying, the diets were broken up and sievedinto convenient pellet sizes and stored at −20 °C until being used.
2.2. Experimental animals
Healthy white shrimp, L. vannamei, were obtained from a com-mercial farm in Zhanjiang, Guangdong, China, and acclimated in are-circulated seawater system for 2 weeks prior to the feeding trial.Two thousand shrimps (initial mean weight 0.72 ± 0.02 g) were ran-domly distributed to five treatments and each treatment had fourreplicates. Each 400-l cylindrical fiberglass tank with 100 shrimpswas used as a replicate. The shrimps were fed to apparent satiationfour times a day at 06:00, 12:00, 16:00 and 20:00. For each time,feed remains and feces of shrimp can be removed by the watersystem. During the 12-week feeding trial, water temperature wasmaintained at 28 °C–30 °C, salinity 30–32 ppt, and pH 7.8–8.2. Thezinc concentration in the seawater flowing into the rearing systemwas 4.5 μg/l determined by ICP-OES (n = 3). At the termination of
Table 1Ingredients and proximate composition of the basal diet.
Ingredients % (dry weight)
Vitamin-free casein (Sigma, St. Louis, MD, USA) 40.0Gelatin (Sigma) 6.0Dextrin (Shanghai Chemical, Shanghai, China) 30.0Carboxymethylcellulose (Shanghai Chemical) 5SO/MFO (Food Grade)a 6.0Cholesterol (Shanghai Chemical) 0.5Vitamin premixb 2.0Zinc-free mineral premixc 0.5a-Cellulose (Sigma) 10
a Soybean oil: menhaden fish oil = 1:2.b Vitamin premix (g/kg premix): thiamin HCL 0.5, riboflavin 3.0, pyridoxine HCL 1.0,
DL Ca-pantothenate 5.0, nicotinic acid 5.0, biotin 0.05, folic acid 0.18, vitamin B12 0.002,choline chloride 100.0, inositol 5.0, menadione 2.0, vitamin A acetate (20,000 IU/g) 5.0,vitamin D (400,000 IU/g) 0.002, DL-alpha-tocopheryl acetate (250 IU/g) 8.0, L-ascorbyl-2-monophosphate (35% active C) 25.0, and alpha-cellulose 840.266.
c Zn-free mineral premix (g/100 g premix): cobalt chloride 0.004, cupric sulfatepentahydrate 0.250, ferrous sulfate 4.0, magnesium sulfate heptahydrate 28.398,manganous sulfate monohydrate 0.650, potassium iodide 0.067, sodium selenite 0.010,and zeolite 66.621.
the experiment, the shrimps were fasted for 24 h before harvest.The shrimp were weighed and counted.
2.3. Sample collection
After being fasted for 24 h, hemolymph (100 μl) waswithdrawn fromthe ventral sinus of each shrimp into a 1-ml sterile syringe containing200 μl anticoagulant solution (30 mM trisodium citrate, 10 mM EDTA,0.34 mM sodium chloride 0.12 mM glucose, adjust pH to 7.55 and os-motic pressure to 780 mOsm/kg). The hemolymph from six shrimpsin one tank was pooled. A 1-ml anticoagulant-hemolymph samplewas centrifuged at 700 ×g at 4 °C for 10 min, and supernatant wasused to measure phenoloxidase (PO) activity, superoxide dismutase(SOD) activity and alkaline phosphatase (AKP) activities. About 500 μlanticoagulant-hemolymph was used to measure total hemocyte count(THC) and phagocytic activity of hemocytes.
2.4. Survival and growth performance
At the termination of the experiment, the shrimps were fasted for24 h before harvest. Total number was counted and mean bodyweight of shrimp was measured. Based on recording the weight ofshrimp and counting the number of shrimps, weight gain, feed con-version ratio (FCR) and survival were calculated using the followingequations:
Weight gain %ð Þ ¼ 100� final weight−initial weightð Þ� initial weight
SGR ¼ 100� ln final weight−ln initial weight½ �� total duration of the experiment
FCR ¼ feed given dry weightð Þ �weight gain wet gainð Þ
Survival %ð Þ ¼ final number of shrimps� initial number of shrimpsð Þ� 100:
2.5. Immunological assays
2.5.1. Total hemocyte count (THC)A drop of the anticoagulant-hemolymph was placed on a Buker
hemocytometer to measure total hemocyte count (THC) under opticalmicroscope (XPS-BM-2GA, Shanghai BMOptical InstitutionManufactureCO. LTD.). The hemocytes were counted manually in all 25 squares(0.1 mm3).
2.5.2. Phenoloxidase (PO) activityPhenoloxidase (PO) activity was estimated spectrophotometrical-
ly by recording the formation of dopachrome using L-3,4 dihydroxy-phenylalanine (L-DOPA; Sigma, USA) as substrate according toHernández-López et al. (1996). Briefly, 50 μl hemolymph superna-tant was incubated with 50 μl trypsin (0.1% in cacodylate buffer:0.45 M sodium chloride, 0.10 M trisodium citrate, 0.01 M sodiumcacodylate, pH 7.0) in 96 well microplate at 25 °C for 10 min, andthen 50 μl L-DOPA (0.3% in cacodylate buffer) was added. The absor-bance value was read every 2 min in microplate reader (ModelMultiskan spectrum, Thermo, MA, Waltham, USA) at 490 nm for20 min. Enzyme activity was expressed as the change in absorbanceper minute per milliliter hemolymph supernatant.
2.5.3. Phagocytic activityPhagocytotic activity (PA) was determined according to Itami et
al. (1994). Collected shrimp hemocytes were rinsed with shrimpsaline and the viable cell number adjusted to 1 × 106 cells/ml. The200 μl cell suspension of shrimp hemocytes was inoculated into a
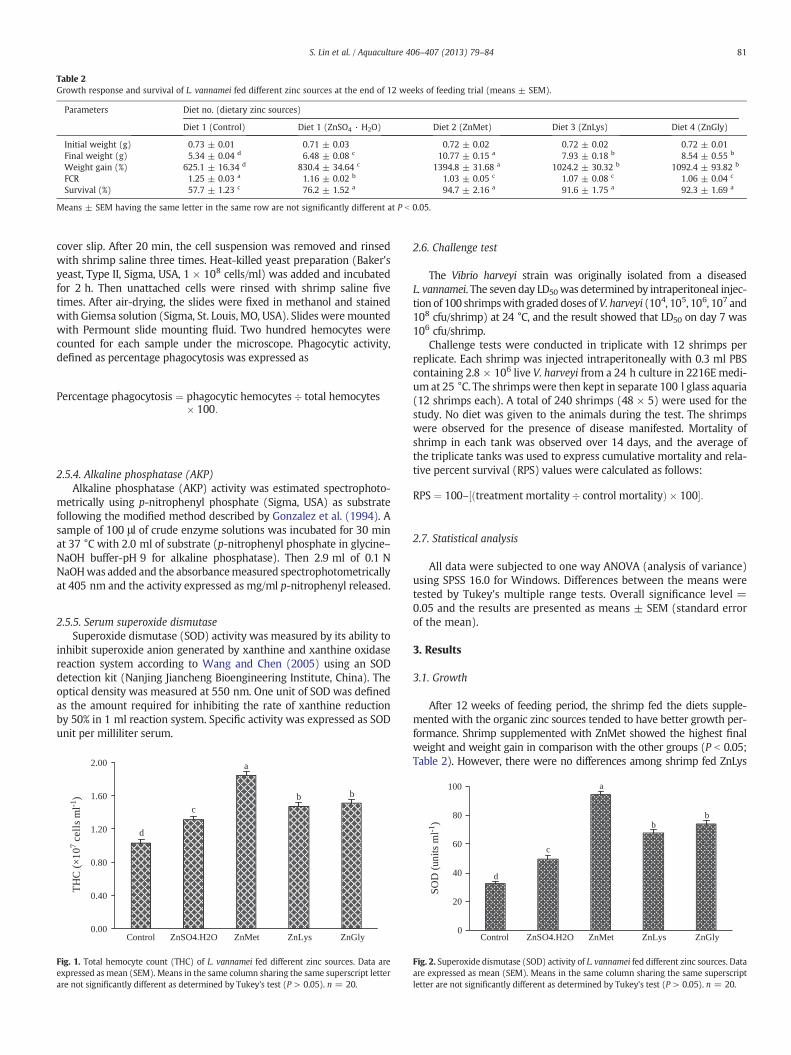
Table 2Growth response and survival of L. vannamei fed different zinc sources at the end of 12 weeks of feeding trial (means ± SEM).
Parameters Diet no. (dietary zinc sources)
Diet 1 (Control) Diet 1 (ZnSO4 · H2O) Diet 2 (ZnMet) Diet 3 (ZnLys) Diet 4 (ZnGly)
Initial weight (g) 0.73 ± 0.01 0.71 ± 0.03 0.72 ± 0.02 0.72 ± 0.02 0.72 ± 0.01Final weight (g) 5.34 ± 0.04 d 6.48 ± 0.08 c 10.77 ± 0.15 a 7.93 ± 0.18 b 8.54 ± 0.55 b
Weight gain (%) 625.1 ± 16.34 d 830.4 ± 34.64 c 1394.8 ± 31.68 a 1024.2 ± 30.32 b 1092.4 ± 93.82 b
FCR 1.25 ± 0.03 a 1.16 ± 0.02 b 1.03 ± 0.05 c 1.07 ± 0.08 c 1.06 ± 0.04 c
Survival (%) 57.7 ± 1.23 c 76.2 ± 1.52 a 94.7 ± 2.16 a 91.6 ± 1.75 a 92.3 ± 1.69 a
Means ± SEM having the same letter in the same row are not significantly different at P b 0.05.
81S. Lin et al. / Aquaculture 406–407 (2013) 79–84
cover slip. After 20 min, the cell suspension was removed and rinsedwith shrimp saline three times. Heat-killed yeast preparation (Baker'syeast, Type II, Sigma, USA, 1 × 108 cells/ml) was added and incubatedfor 2 h. Then unattached cells were rinsed with shrimp saline fivetimes. After air-drying, the slides were fixed in methanol and stainedwith Giemsa solution (Sigma, St. Louis, MO, USA). Slides weremountedwith Permount slide mounting fluid. Two hundred hemocytes werecounted for each sample under the microscope. Phagocytic activity,defined as percentage phagocytosis was expressed as
Percentage phagocytosis ¼ phagocytic hemocytes� total hemocytes� 100:
2.5.4. Alkaline phosphatase (AKP)Alkaline phosphatase (AKP) activity was estimated spectrophoto-
metrically using p-nitrophenyl phosphate (Sigma, USA) as substratefollowing the modified method described by Gonzalez et al. (1994). Asample of 100 μl of crude enzyme solutions was incubated for 30 minat 37 °C with 2.0 ml of substrate (p-nitrophenyl phosphate in glycine–NaOH buffer-pH 9 for alkaline phosphatase). Then 2.9 ml of 0.1 NNaOHwas added and the absorbancemeasured spectrophotometricallyat 405 nm and the activity expressed as mg/ml p-nitrophenyl released.
2.5.5. Serum superoxide dismutaseSuperoxide dismutase (SOD) activity was measured by its ability to
inhibit superoxide anion generated by xanthine and xanthine oxidasereaction system according to Wang and Chen (2005) using an SODdetection kit (Nanjing Jiancheng Bioengineering Institute, China). Theoptical density was measured at 550 nm. One unit of SOD was definedas the amount required for inhibiting the rate of xanthine reductionby 50% in 1 ml reaction system. Specific activity was expressed as SODunit per milliliter serum.
bb
a
c
d
0.00
0.40
0.80
1.20
1.60
2.00
TH
C (
×107 c
ells
ml-1
)
Control ZnSO4.H2O ZnMet ZnLys ZnGly
Fig. 1. Total hemocyte count (THC) of L. vannamei fed different zinc sources. Data areexpressed asmean (SEM). Means in the same column sharing the same superscript letterare not significantly different as determined by Tukey's test (P > 0.05). n = 20.
2.6. Challenge test
The Vibrio harveyi strain was originally isolated from a diseasedL. vannamei. The seven day LD50was determined by intraperitoneal injec-tion of 100 shrimpswith graded doses of V. harveyi (104, 105, 106, 107 and108 cfu/shrimp) at 24 °C, and the result showed that LD50 on day 7 was106 cfu/shrimp.
Challenge tests were conducted in triplicate with 12 shrimps perreplicate. Each shrimp was injected intraperitoneally with 0.3 ml PBScontaining 2.8 × 106 live V. harveyi from a 24 h culture in 2216E medi-um at 25 °C. The shrimpswere then kept in separate 100 l glass aquaria(12 shrimps each). A total of 240 shrimps (48 × 5) were used for thestudy. No diet was given to the animals during the test. The shrimpswere observed for the presence of disease manifested. Mortality ofshrimp in each tank was observed over 14 days, and the average ofthe triplicate tanks was used to express cumulative mortality and rela-tive percent survival (RPS) values were calculated as follows:
RPS ¼ 100– treatment mortality� control mortalityð Þ � 100½ �:
2.7. Statistical analysis
All data were subjected to one way ANOVA (analysis of variance)using SPSS 16.0 for Windows. Differences between the means weretested by Tukey's multiple range tests. Overall significance level =0.05 and the results are presented as means ± SEM (standard errorof the mean).
3. Results
3.1. Growth
After 12 weeks of feeding period, the shrimp fed the diets supple-mented with the organic zinc sources tended to have better growth per-formance. Shrimp supplemented with ZnMet showed the highest finalweight and weight gain in comparison with the other groups (P b 0.05;Table 2). However, there were no differences among shrimp fed ZnLys
d
c
a
bb
0
20
40
60
80
100
Control ZnSO4.H2O ZnMet ZnLys ZnGly
SOD
(un
its m
l-1)
Fig. 2. Superoxide dismutase (SOD) activity of L. vannamei fed different zinc sources. Dataare expressed as mean (SEM). Means in the same column sharing the same superscriptletter are not significantly different as determined by Tukey's test (P > 0.05). n = 20.
bb
a
c
d
0.0
2.0
4.0
6.0
8.0A
KP
(uni
ts m
l-1)
Control ZnSO4.H2O ZnMet ZnLys ZnGly
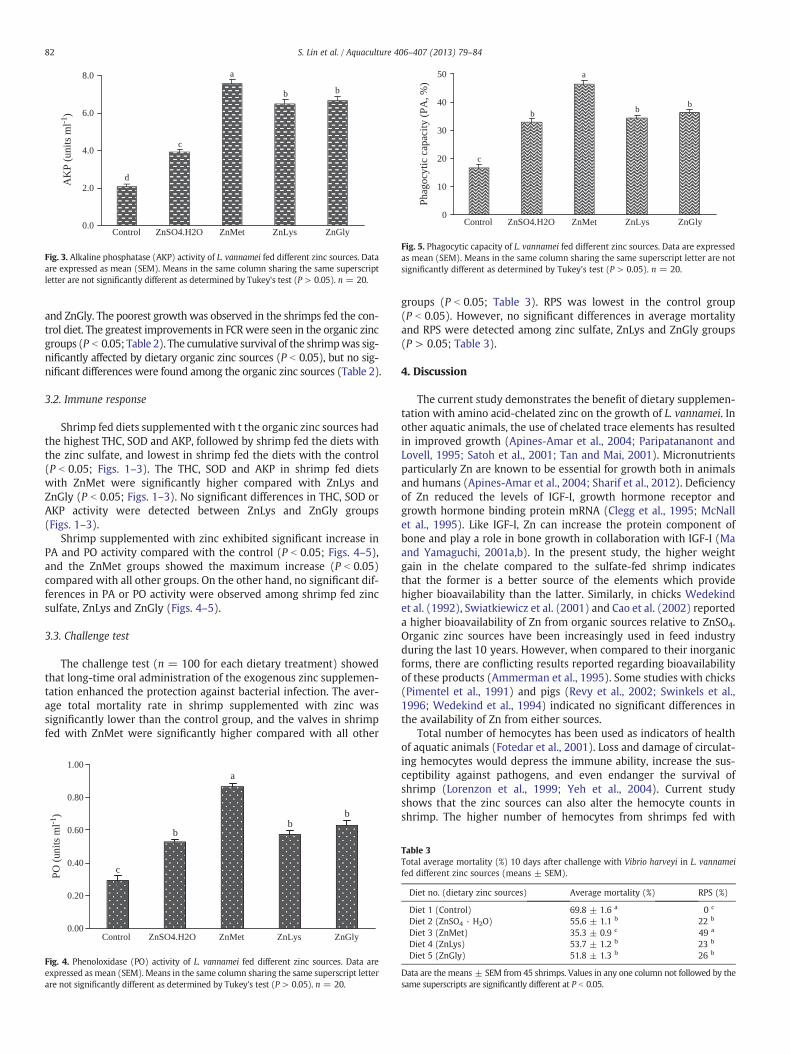
Fig. 3. Alkaline phosphatase (AKP) activity of L. vannamei fed different zinc sources. Dataare expressed as mean (SEM). Means in the same column sharing the same superscriptletter are not significantly different as determined by Tukey's test (P > 0.05). n = 20.
c
b
a
bb
0
10
20
30
40
50
Phag
ocyt
ic c
apac
ity (
PA, %
)
Control ZnSO4.H2O ZnMet ZnLys ZnGly
Fig. 5. Phagocytic capacity of L. vannamei fed different zinc sources. Data are expressedas mean (SEM). Means in the same column sharing the same superscript letter are notsignificantly different as determined by Tukey's test (P > 0.05). n = 20.
82 S. Lin et al. / Aquaculture 406–407 (2013) 79–84
and ZnGly. The poorest growth was observed in the shrimps fed the con-trol diet. The greatest improvements in FCRwere seen in the organic zincgroups (P b 0.05; Table 2). The cumulative survival of the shrimpwas sig-nificantly affected by dietary organic zinc sources (P b 0.05), but no sig-nificant differences were found among the organic zinc sources (Table 2).
3.2. Immune response
Shrimp fed diets supplemented with t the organic zinc sources hadthe highest THC, SOD and AKP, followed by shrimp fed the diets withthe zinc sulfate, and lowest in shrimp fed the diets with the control(P b 0.05; Figs. 1–3). The THC, SOD and AKP in shrimp fed dietswith ZnMet were significantly higher compared with ZnLys andZnGly (P b 0.05; Figs. 1–3). No significant differences in THC, SOD orAKP activity were detected between ZnLys and ZnGly groups(Figs. 1–3).
Shrimp supplemented with zinc exhibited significant increase inPA and PO activity compared with the control (P b 0.05; Figs. 4–5),and the ZnMet groups showed the maximum increase (P b 0.05)compared with all other groups. On the other hand, no significant dif-ferences in PA or PO activity were observed among shrimp fed zincsulfate, ZnLys and ZnGly (Figs. 4–5).
3.3. Challenge test
The challenge test (n = 100 for each dietary treatment) showedthat long-time oral administration of the exogenous zinc supplemen-tation enhanced the protection against bacterial infection. The aver-age total mortality rate in shrimp supplemented with zinc wassignificantly lower than the control group, and the valves in shrimpfed with ZnMet were significantly higher compared with all other
bb
a
b
c
0.00
0.20
0.40
0.60
0.80
1.00
PO (
units
ml-1
)
Control ZnSO4.H2O ZnMet ZnLys ZnGly
Fig. 4. Phenoloxidase (PO) activity of L. vannamei fed different zinc sources. Data areexpressed asmean (SEM). Means in the same column sharing the same superscript letterare not significantly different as determined by Tukey's test (P > 0.05). n = 20.
groups (P b 0.05; Table 3). RPS was lowest in the control group(P b 0.05). However, no significant differences in average mortalityand RPS were detected among zinc sulfate, ZnLys and ZnGly groups(P > 0.05; Table 3).
4. Discussion
The current study demonstrates the benefit of dietary supplemen-tation with amino acid-chelated zinc on the growth of L. vannamei. Inother aquatic animals, the use of chelated trace elements has resultedin improved growth (Apines-Amar et al., 2004; Paripatananont andLovell, 1995; Satoh et al., 2001; Tan and Mai, 2001). Micronutrientsparticularly Zn are known to be essential for growth both in animalsand humans (Apines-Amar et al., 2004; Sharif et al., 2012). Deficiencyof Zn reduced the levels of IGF-I, growth hormone receptor andgrowth hormone binding protein mRNA (Clegg et al., 1995; McNallet al., 1995). Like IGF-I, Zn can increase the protein component ofbone and play a role in bone growth in collaboration with IGF-I (Maand Yamaguchi, 2001a,b). In the present study, the higher weightgain in the chelate compared to the sulfate-fed shrimp indicatesthat the former is a better source of the elements which providehigher bioavailability than the latter. Similarly, in chicks Wedekindet al. (1992), Swiatkiewicz et al. (2001) and Cao et al. (2002) reporteda higher bioavailability of Zn from organic sources relative to ZnSO4.Organic zinc sources have been increasingly used in feed industryduring the last 10 years. However, when compared to their inorganicforms, there are conflicting results reported regarding bioavailabilityof these products (Ammerman et al., 1995). Some studies with chicks(Pimentel et al., 1991) and pigs (Revy et al., 2002; Swinkels et al.,1996; Wedekind et al., 1994) indicated no significant differences inthe availability of Zn from either sources.
Total number of hemocytes has been used as indicators of healthof aquatic animals (Fotedar et al., 2001). Loss and damage of circulat-ing hemocytes would depress the immune ability, increase the sus-ceptibility against pathogens, and even endanger the survival ofshrimp (Lorenzon et al., 1999; Yeh et al., 2004). Current studyshows that the zinc sources can also alter the hemocyte counts inshrimp. The higher number of hemocytes from shrimps fed with
Table 3Total average mortality (%) 10 days after challenge with Vibrio harveyi in L. vannameifed different zinc sources (means ± SEM).
Diet no. (dietary zinc sources) Average mortality (%) RPS (%)
Diet 1 (Control) 69.8 ± 1.6 a 0 c
Diet 2 (ZnSO4 · H2O) 55.6 ± 1.1 b 22 b
Diet 3 (ZnMet) 35.3 ± 0.9 c 49 a
Diet 4 (ZnLys) 53.7 ± 1.2 b 23 b
Diet 5 (ZnGly) 51.8 ± 1.3 b 26 b
Data are the means ± SEM from 45 shrimps. Values in any one column not followed by thesame superscripts are significantly different at P b 0.05.

83S. Lin et al. / Aquaculture 406–407 (2013) 79–84
ZnMet supplemented diets compared to the other diets resulted instrengthening the non-specific defense system of shrimp to bacteriaand pathogens. Smith et al. (2003) demonstrated that the decrease ofimmune parameters may be due to the consumption of the hemocytesby degranulation. In this study, zinc sulfate reduced the hemocytequantity, and decreased the values of some immune parameters tested.This could result in lower survival rate of the shrimps fed zinc sulfate.Moreover, circulating hemocytes are affected by extrinsic factors liketemperature, salinity, pH, and heavy metals (Le Moullac and Haffner,2000). Therefore, different responses in hemocyte count are considereddue to the different zinc sources tested. To our knowledge, the mecha-nism by which organic zinc sources increase numbers of hemocytes isstill not elucidated to date.
Zinc is well-known to play a central role in the immune system (Rinkand Kirchner, 2000). In the present experiment, AKP, SOD, PA and POexhibited a higher activity in the zinc-supplemented group, as demon-strated in other reports (Apines et al., 2003; Apines-Amar et al., 2004;Tan andMai, 2001). Furthermore, ZnMet showed better immune activitycompared to the other zinc sources. This implied that ZnMetwas themosteffective one in enhancing innate immune responses of shrimps. Besidesits involvement in genetic stability andgene expression through the activ-ity of transcription factors and RNA and DNA polymerases, Zn also plays arole inDNA repair and apoptosis (Dreosti, 2001; Falchuk, 1998). Apines etal. (2003) and Apines-Amar et al. (2004) reported that the expression ofDNApolymerase and CuZnSODwas higher in fish fed the chelate than theinorganic source, which could have enhanced the rate of protein synthe-sis. This may help explain the enhanced growth of the shrimp in thisstudy.
In addition, levels of zinc also affect animal immunity. Studiesdemonstrated that the adequate dietary Zn has immunostimulatoryactivities, and keeps the homeostasis of oxidation/reduction, but ex-cessive dietary Zn induces a high oxidative stress in P. monodon(Shiau and Jiang, 2006) and in abalone (Wu et al., 2011). Similarly,the supplemental Zn also resulted in a general disturbance of the im-mune capacities of hemocytes in in vitro experiments (Mottin et al.,2010). Among the zinc sources studied, organic zinc sources showedbetter immune activity compared to ZnSO4. Interestingly, zinc sup-plementation from the chelate at the same levels (36 mg/kg of diet)as the inorganic source significantly increased the level of enzymetranscript, indicating that dietary mineral uptake from the chelatesources improved cellular mineral-dependent functions. Likewise,this corresponds to our previous result (Lin et al., 2011) wherein im-mune activity of shrimp from the chelate was markedly higher com-pared to the sulfate even at one-half supplementation level of theinorganic salt. The higher immune parameters in the chelate, in thisstudy compared to the sulfate-fed shrimp indicated a higher utiliza-tion of Zn. Combined with these findings, it is implied that dietaryZn could induce expression of antioxidant molecules (e.g., superoxidedismutase and metallothionein) and then maintain a better antioxi-dant status against oxidative stress in human and animals (Fang etal., 2002).
The present study showed that amino acid-chelated zinc signifi-cantly reduced cumulative mortality of L. vannamei after being chal-lenged by V. harveyi. The improved resistance of L. vannamei afterchallengemay be partly attributable to the increased AKP and SOD ac-tivity of shrimp compared to zinc sulfate. Therefore, the findings indi-cated that the increased resistance to V. harveyi of L. vannamei wasrelated to the enhanced immune status. More studies are needed tofind the reasons for it. To the best of our knowledge, the exact mech-anisms for these findings were not clear yet.
In summary, the requirement for Zn depends on its chemical form,as suggested by improved growth and hemocyte functions with thesupplementation of amino acid-chelated zinc. Results of this study in-dicate that the level of Zn recommended by NRC (2011) is sufficientfor optimal growth performance and immune responses, and thatZn from organic sources improves these measurements when
supplemented to diets containing 30 ppm of supplemental Zn fromZnMet. Thus, based on weight gain, survival and immunity indices ex-amined in this study, amino acid-chelates are a better source of traceelements for L. vannamei. Potential benefits of organic zinc sources onhemocyte functions need to be further evaluated.
Acknowledgements
The authors thank Yao CF and Yan SK for taking care of the shrimp.Special thanks to Zhang L and Wu YG for helping with the chemicalanalysis.
References
Ammerman, C.B., Baker, D.H., Lewis, A.J., 1995. Bioavailability of Nutrients for Animals:Amino Acids, Minerals, and Vitamins. Academic Press, San Diego, CA.
AOAC, 1995. Official Methods of Analysis, 16th ed. Association of Official AnalyticalChemists, Arlington, VA, USA.
Apines, M.J., Satoh, S., Kiron, V., Watanabe, T., Aoki, T., 2003. Availability of supplemen-tal amino acidchelated trace elements in diets containing tricalcium phosphateand phytate to rainbow trout, Oncorhynchus mykiss. Aquaculture 225, 431–444.
Apines-Amar, M.J.S., Satoh, S., Caipang, C.M.A., Kiron, V.,Watanabe, T., Aoki, T., 2004. Aminoacid-chelate: a better source of Zn,Mn, and Cu for rainbow trout, Oncorhynchus mykiss.Aquaculture 240, 345–358.
Ashmead, H.D., 1992. The Roles of Amino Acid Chelates in Animal Nutrition. NoyesPublications, Park Ridge, NJ 479.
Cao, J., Henry, P.R., Davis, C.D., Cousins, R.J., Miles, R.D., Littell, R.C., Ammerman, C.B.,2002. Relative bioavailability of organic zinc sources based on tissue andmetallothionein in chicks fed conventional dietary zinc concentrations. AnimalFeed Science and Technology 161–170.
Clegg, M.S., Keen, C.L., Donovan, S.M., 1995. Zinc deficiency-induced anorexia influ-ences the distribution of serum insulin-like growth factor-binding proteins in therat. Metabolism 44, 1495–1501.
Dreosti, I.E., 2001. Zinc and the gene. Mutation Research 475, 161–167.Falchuk, K., 1998. The molecular basis for the role of zinc in developmental biology.
Molecular and Cellular Biochemistry 188, 41–48.Fang, Y.Z., Yang, S., Wu, G.Y., 2002. Free radicals, antioxidants, and nutrition. Nutrition
18, 872–879.FAO, 2010. The State of World Fisheries and Aquaculture. 1020-5500978-92-5-
306675-9. Fisheries and Aquaculture Dept., Rome, Italy.Fotedar, S., Tsvetnenko, E., Evans, L., 2001. Effect of air exposure on the immune system
of the rock lobster Panulirus cygnus. Marine and Freshwater Research 52,1351–1355.
Gonzalez, F., Esther Ferez-Vidal, M., Arias, J.M., Mantoya, E., 1994. Partial purificationand biochemical properties of acid and alkaline phosphatases from Myxococcuscoralloides D. Journal of Applied Bacteriology. Journal of Applied Microbiology 77,567–573.
Hernández-López, J., Gollas-Galván, T.S., Vargas-Albores, F., 1996. Activation of theprophenoloxidase system of the brown shrimp (Penaeus californiensis Holmes).Comparative Biochemistry and Physiology. C 113, 61–66.
Itami, T., Takahashi, Y., Tsuchihira, E., Igasu, H., Kondo, M., 1994. Enhancement of diseaseresistance of kuruma prawn Penaeus japonicus and increase in phagocytic activityof prawn hemocytes after oral administration of b 1,3-glucan (Schiz-ophyllan).Proceeding of the Third Asian fisheries Forum, Singapore, 26–30 October 1992. TheAsian Fisheries Society Manila, Philippines, pp. 375–378.
Kureshy, N., Davis, D.A., 2002. Protein requirement for maintenance and maximumweight gain for the Pacific white shrimp, Litopenaeus vannamei. Aquaculture 204,125–143.
Le Moullac, G., Haffner, P., 2000. Environmental factors affecting immune responses inCrustacea. Aquaculture 191, 121–131.
Lemire, J., Mailloux, R., Appanna, V.D., 2008. Zinc toxicity alters mitochondrial metab-olism and leads to decreased ATP production in hepatocytes. Journal of AppliedToxicology 28, 175–182.
Lin, S.M., Mao, S.H., Guan, Y., Lin, X., 2011. Effects of different Zn Mn Cu sources ongrowth performance and non-specific immunity of Litopenaeus vannamei. Fresh-water Fisheries 41 (6), 64–69 (in China, with English abstract).
Lorenzon, S., De Guarrini, S., Smith, V.J., Ferrero, E.A., 1999. Effects of LPS injection oncirculating haemocytes in crustaceans in vivo. Fish & Shellfish Immunology 9,31–50.
Ma, Z.J., Yamaguchi, M., 2001a. Role of endogenous zinc in the enhancement of boneprotein synthesis associated with bone growth of newborn rats. Journal of Boneand Mineral Metabolism 19, 38–44.
Ma, Z.J., Yamaguchi, M., 2001b. Stimulatory effect of zinc and growth factor on boneprotein component in newborn rats: enhancement with zinc and insulin-likegrowth factor-I. International Journal of Molecular Medicine 7, 73–78.
McNall, A.D., Etherton, T.D., Fosmire, G.J., 1995. The impaired growth induced by zincdeficiency in rats is associated with decreased expression of the hepatic insulin-like growth factor I and growth hormone receptor genes. Journal of Nutrition125, 874–879.
Mottin, E., Caplat, C., Mahaut, M.L., Costil, K., Barillier, D., Lebel, J.M., Serpentini, A.,2010. Effect of in vitro exposure to zinc on immunological parameters of

84 S. Lin et al. / Aquaculture 406–407 (2013) 79–84
haemocytes from the marine gastropod Haliotis tuberculata. Fish & Shellfish Immu-nology 29, 846–853.
NRC, 2011. Nutrient Requirements of Fish and Shrimp. National Academies Press,Washington D.C.
Paripatananont, T., Lovell, R.T., 1995. Responses of channel catfish fed organic and inor-ganic sources of zinc to Edwardsiella ictaluri challenge. Journal of Aquatic AnimalHealth 7, 147–154.
Pimentel, J.L., Cook, M.E., Greger, J.L., 1991. Research note: bioavailability of zinc–methioninefor chicks. Poultry Science 70, 1637–1639.
Puchala, R., Sahlu, T., Davis, J.J., 1999. Effects of zinc–methionine on performance ofAngora goats. Small Ruminant Research 33, 1–8.
Revy, P.S., Jondreville, C., Dourmad, J.Y., Guinotte, F., Nys, Y., 2002. Bioavailability of twosources of zinc in weanling pigs. Animal Research 51, 315–326.
Rink, L., Kirchner, H., 2000. Zinc-altered immune function and cytokine production.Journal of Nutrition 130, 1407S–1411S.
Roy, L.A., Davis, D.A., Saoud, I.P., 2006. Effects of lecithin and cholesterol supplementa-tion to practical diets for Litopenaeus vannamei reared in low salinity waters. Aqua-culture 257, 446–452.
Satoh, S., Apines, M.J., Tsukioka, T., Kiron, V., Watanabe, T., Fujita, S., 2001. Bioavailabil-ity of amino acids chelated and glass embedded manganese to rainbow trout,Oncorhynchus mykiss, fingerlings. Aquaculture Research 32S, 18–25.
Shankar, A.H., Prasad, A.S., 1998. Zinc and immune function: the biological basis of alteredresistance to infection. The American Journal of Clinical Nutrition 68, 447S–463S(Suppl.).
Sharif, R., Thomas, P., Zalewski, P., Fenech, M., 2012. The role of zinc in genomic stability.Mutation Research 733, 111–121.
Shiau, S.Y., Jiang, L.C., 2006. Dietary zinc requirements of grass shrimp, Penaeusmonodon, and effects on immune responses. Aquaculture 254, 476–482.
Smith, V.J., Brown, J.H., Hauton, C., 2003. Immunostimulation in crustaceans: does itreally protect against infection? Fish & Shellfish Immunology 15, 71–90.
Swiatkiewicz, S., Koreleski, J., Zhong, D.Q., 2001. The bioavailability of zinc frominorganic and organic sources in broiler chickens as affected by addition of phytase.Journal of Animal and Feed Sciences 10, 317–328.
Swinkels, J.W.G.M., Kornegay, E.T., Zhou, W., Lindemann, M.D., Webb Jr., K.E.,Verstegen, M.W.A., 1996. Effectiveness of a zinc amino acid chelate and zinc sulfatein restoring serumand soft tissue zinc concentrations when fed to zinc-depletedpigs. Journal of Animal Science 74, 2420–2430.
Tan, B.P., Mai, K.S., 2001. Zinc methionine and zinc sulfate as sources of dietary zinc forjuvenile abalone, Haliotis discus hannai Ino. Aquaculture 192, 67–84.
Wang, S.H., Chen, J.C., 2005. The protective effect of chitin and chitosan against Vibrioalginolyticus in white shrimp, Litopenaeus vannamei. Fish & Shellfish Immunology19, 191–204.
Wang, C., Lovell, R.T., 1997. Organic selenium sources, selenomethionine andselenoyeast, have higher bio-availability than an inorganic selenium source, sodi-um selenite, in diets for channel catfish (Ictalurus punc-tatus). Aquaculture 152,223–234.
Watanabe, T., Kiron, V., Satoh, S., 1997. Trace minerals in fish nutrition. Aquaculture151, 185–207.
Wedekind, K.J., Hortin, A.E., Baker, D.H., 1992. Methodology for assessing zinc bioavailability:efficacy estimates for zinc-methionine, zinc sulfate, and zinc oxide. Journal of AnimalScience 70, 178–187.
Wedekind, K.J., Lewis, A.J., Giesemann, M.A., Miller, P.S., 1994. Bioavailability of zincfrom inorganic and organic sources for pigs fed corn-soybean meal diets. Journalof Animal Science 72, 2681–2689.
Wu, C.L., Zhang, W.B., Mai, K.S., Xu, W., Zhong, X.L., 2011. Effects of dietary zinc on geneexpression of antioxidant enzymes and heat shock proteins in hepatopancreas ofabalone Haliotis discus hannai. Comparative Biochemistry and Physiology. C 154,1–6.
Yeh, S.T., Liu, C.H., Chen, J.C., 2004. Effect of copper sulfate on the immune response andsusceptibility to Vibrio alginolyticus in the white shrimp Litopenaeus vannamei. Fish& Shellfish Immunology 175, 437–446.