comparison of the effects of fk-506, cyclosporin a and rapamycin
TRANSCRIPT

Immunology 1991 73 316-321
Comparison of the effects of FK-506, cyclosporin A and rapamycinon IL-2 production
D. J. HENDERSON, I. NAYA, R. V. BUNDICK, G. M. SMITH & J. A. SCHMIDTDepartment of Biochemistry, Fisons Pharmaceuticals plc, Loughborough, Leicestershire
Acceptedfor publication 11 March 1991
SUMMARY
The immunosuppressive compounds FK-506, cyclosporin A (CsA) and rapamycin inhibit both thehuman and mouse mixed lymphocyte reactions (MLR) with IC50s of 2-5 x 10-10 M for FK-506 andrapamycin and l0- M for CsA. FK-506 and CsA were also potent inhibitors of A23187/PMA-stimulated IL-2 production by Jurkat and HuT-78 cells but had no effect on the response of mouseCTLL cells to IL-2. IC50 values for inhibition of IL-2 production closely matched those for inhibitionof the MLR and both drugs were active only during the first 4-6 hr following stimulation. In contrast,rapamycin was a poor inhibitor of IL-2 production, although it inhibited cellular responses to IL-2.The IC50 values for these two activities indicated that neither alone accounted for rapamycininhibition of the MLR. FK-506 and CsA affected IL-2 gene transcription in Jurkat cells by the samemechanism. Both inhibited the appearance of the transcription factor, NFAT, whereas rapamycindid not. The appearance of another transcription factor, NFK#, was unaffected by all three drugs.The effects of FK-506 and CsA on IL-2 gene expression, therefore, are similar even though the twodrugs act through distinct cytosolic receptors.
INTRODUCTION
Cyclosporin A (CsA) has been in clinical use as an immuno-suppressive drug for a number of years. Despite its success, theexact mechanism of its action is unknown. Recently, two novelimmunosuppressive agents have been described, FK-506 andrapamycin. These compounds have aroused considerable inter-est because both appear to be 10-100-fold more active than CsAin various models.'-7 FK-506 and rapamycin are structurallyrelated and although both, like CsA, are cyclic microbialmetabolites, they have no structural relationship with CsA.
CsA and FK-506 both block antigen- and mitogen-driven T-cell activation.3-7 These effects are selective as the drugsspecifically block induction of 'early' genes but have no effect onthe induction of 'immediate early' or the 'late' genes of T-cellactivation.3' -'0 Early genes include those coding for interleukin-2 (IL-2), IL-3, IL-4, tumour necrosis factor alpha (TNF-ox) andinterferon-gamma (IFN-y).8 Specific non-expression of theseproteins would be expected to result in immunosuppression andthis activity could, therefore, largely explain the immunosup-pressive properties of both drugs. Exactly how the transfer of
Abbreviations: CsA, cyclosporin A; IL-2, interleukin-2; MLR,mixed lymphocyte reaction; PMA, phorbol- 1 2-myristate- 13-acetate;NFAT, nuclear factor of activated T cells; NFKJJ, nuclear factor K/3.
Correspondence: Dr J. A. Schmidt, Dept. of Biochemistry, FisonsPharmaceuticals plc, Bakewell Road, Loughborough, LeicestershireLEI I ORH, U.K.
the mitogenic signal to new gene expression is blocked isunknown. CsA and FK-506 specifically inhibit the Ca2+ arm ofthe T-cell activation signal,3 6 however, they do not simply blockCa2+ influx but affect the signal at some later point in thetransduction pathway. '°0" Recent studies have focused on IL-2gene expression in CsA-treated cells. DNA sequences within 300bp upstream of the human IL-2 gene transcription start sitespecify both tissue-specific expression and inducibility andseveral protein factors are known to bind to this region.'2-'4 Oneof these factors, NFAT,14"15 protects the region of the IL-2 genepromoter from -263 to - 290 bases upstream of the transcrip-tion start site in DNase I footprinting experiments. Another,NFKf,'4"6 protects the region from - 187 to -207. Both ofthese factors are absent in protein extracts of unstimulated cellsbut present in activated cell extracts. CsA selectively inhibits theappearance of the NFAT transcription factor.'7"8
Little is known about the action of rapamycin other thanthat it also possesses immunosuppressive properties both in vitroand in vivo.2"19-2I Recently, it has been suggested that its mode ofaction is different from that of FK-506,2' even though it binds tothe same intracellular receptor (our unpublished data).22'23
We have compared the effects of CsA, FK-506 and rapamy-cin on various systems with a view to understanding further howthey mediate immunosuppression. Here, we show that CsA,FK-506 and rapamycin all inhibit the mixed lymphocytereaction (MLR). However, whilst inhibition of IL-2 productioncan explain the action of FK-506 and CsA on the MLR,rapamycin action is not fully explained either by inhibition of
316
ADONIS 001928059100158C

Immunosuppressive drugs and IL-2 production
IL-2 production or by inhibition of the response of cells to IL-2.Furthermore, FK-506 and CsA, but not rapamycin, inhibit theappearance of the same transcription factor required forexpression of the IL-2 gene.
MATERIALS AND METHODS
Cells and cell linesHuman peripheral blood mononuclear cells (PBMC) wereprepared from blood taken from healthy volunteers in 3%sodium citrate. PBMC were isolated by centrifugation onFicoll-Hypaque (Lymphoprep, Nycomed, Birmingham, U.K.)according to the manufacturers instructions. C57/BL andBALB/c mice were obtained from Olac, Bicester, Oxon, U.K.Spleen cell suspensions were prepared by gently dissociatingaseptically removed spleens and washing cells twice with RPMI-1640 medium. The human T-lymphoblast cell lines Jurkat, J6and Hut-78 (obtained from the Imperial Cancer Research Fund,London, U.K.) were maintained in RPMI-1640 medium con-taining 5% foetal calf serum (FCS), 100 U/ml penicillin, 100 U/ml streptomycin and 2 mm glutamine. Mouse CTLL-2 cells(from the ECACC, Porton Down, Sussex, U.K.) were grown inRPMI-1640 medium containing 5% FCS, antibiotics, gluta-mine, 20 gM 2-mercaptoethanol and 20 U/ml human recombi-nant IL-2 (Boehringer Mannheim, Mannheim, Germany).
Drugs and other chemicalsCsA and FK-506 were kindly provided by the FujisawaPharmaceutical Company, Osaka, Japan, and rapamycin byAyerst Research, Montreal, Canada. Drugs were dissolved inethanol and stored at -20° as 10 mg/ml stock solutions. Foruse, all three drugs were diluted in ethanol to 10 x the requiredconcentration to minimize handling losses on plastic. Phorbol-12-myristate-13-acetate (PMA) and A23187 were purchasedfrom Sigma, Poole, Dorset, U.K., dissolved in DMSO at 1 mg/ml and 10 mm, respectively, and stored at -20°.
Mixed lymphocyte reactions (MLR)Two-way human or mouse MLR were set up using humanPBMC from two individual donors pre-tested to give a strongresponse or spleen cells from C57/BL and BALB/c mice. Drugswere dispensed at varying concentrations in triplicate in flat-bottomed 96-well plates, ethanol allowed to evaporate and cellsadded (50 pl from each donor at 3 x 106/ml in serum-free RPMI-1640 medium containing antibiotics and glutamine). Volumeswere made up to 200 pl with RPMI-1640 containing heat-inactivated pooled human serum for human cells or FCS formouse cells so that the final serum concentration was 10%.Plates were incubated at 370, 5% CO2 for 96 hr and cells pulsedwith 3H-methyl-thymidine ([3H]TdR, 0-5 pCi/well, Amersham,Amersham, Bucks, U.K.) for the last 6 hr before harvesting ontoglass-fibre filters. Incorporated radioactivity was counted byliquid scintillation spectrometry.
Induction of IL-2 in T-cell lines and assay for IL-2 activityDrugs were dispensed in flat-bottomed 96-well plates as for theMLR, ethanol allowed to evaporate and 50 of a mixture ofA23187 (1 pM) and PMA (200 ng/ml) in growth medium were
added to each well as appropriate. Jurkat J6 or Hut-78 cells wereadded in 200 pl ofgrowth medium to give 2 x 105 or 105 cells perwell, respectively. Plates were incubated at 37°, 5% CO2 for 24 hr
and the IL-2 released was assayed by transferring 100 pl of theculture supernatant to a second plate containing 100 pl ofCTLL-2 cells (2 x 104) per well. Plates were incubated for 18 hrand cultures pulsed with [3H]TdR (0-5 pCi per well) for a further4 hr. Cells were harvested onto glass fibre filters and incorpor-ated radioactivity counted as described above.
IL-2 gene promoter binding activity assaysJurkat cells were stimulated for 3 hr with 0 4 pM A23 187 and 20ng/ml PMA at a cell density of 3 x 106/ml. Nuclear proteinextracts were prepared from 109 cells as in ref. 24 and pelletsfinally resuspended in a 1-ml volume giving a typical proteinconcentration of 4 mg/ml. Aliquots of treated and control cellswere incubated for a further 20 hr and supernatants assayed forIL-2 as described above to confirm that IL-2 was induced.
Oligonucleotides for gel shift experiments were labelled byphosphorylation with y32P-ATP (BRL 5' terminus DNA label-ling system) and annealed to their complements in 50mM NaCl,20 mm Tris, pH 8, by slow cooling to room temperature from90°. Protein-binding reactions contained 15 pg nuclear proteinextract, 2 ng labelled annealed oligonucleotide, 1 pg poly dIdC,5% glycerol, 1 mm EDTA, 1 mm DTT, 50 mm NaCl and 10 mMTris-HCl, pH 7-5, in a 30 pl volume. Binding was for 30 min atroom temperature, after which 5 kl of 20% glycerol, 01%bromophenol blue were added, 9 p1 of the reaction loaded on a4% polyacrylamide gel and bound complexes analysed byelectrophoresis in Tris borate buffer at approximately 11 V/cm.25 In oligonucleotide competition experiments, a 20-foldexcess of unlabelled oligonucleotide was added to the proteinbinding reaction.
Six oligonucleotides covering the sequences 5'-AAA-TATGTGAATATGTAAAACATTTTGAC-3' (NFIL2A site,-63 to -93), 5'-ACAAAGAGGGATTTCACCTACATC-CAT-3' (NFKJ5 site, - 186 to -212), 5'-GAG-GAAAAACTGTTTCATACAGAAGGCGTTA-3' (NFAT-1site, -257 to -285) and their complements were synthesized onan ABI PCRmate and purified on 20% polyacrylamide gelsbefore use.26
RESULTS
Effects of CsA, FK-506 and rapamycin on the MLR
CsA, FK-506 and rapamycin were all potent inhibitors of themouse MLR (Fig. la). The IC50 values for FK-506 and CsAwere 2-5 x 10-10 M and 10-8 M, respectively. The IC50 forrapamycin in the MLR was essentially identical to that for FK-506 (Fig. la). The same results were obtained using humanPBMC (data not shown) and, in both systems, inhibition byCsA and FK-506 approached 100%. Interestingly, rapamycindiffered from FK-506 in that the maximum inhibition did notexceed 90% (Fig. la).
Effects of CsA, FK-506 and rapamycin on IL-2 production byhuman T-lymphocyte cell lines
Human T-lymphocyte cell lines such as Jurkat and Hut-78 canbe induced to produce IL-2 by stimulation with Ca2+ ionophoreand PMA.27 Recent reports have shown that CsA and FK-506both inhibit this response.'0'28 This system was used to comparethe actions of rapamycin with CsA and FK-506. As expected,
317
D. J. Henderson et al.
(a)
100
80
60-
40
20
0 -
-20
-12 -10 -8 -6 -4
(b)
- 100
C
C.
Cj
100o
,80
i 60
540-
520=-0
-20-12 -10 -8 -6 -4
Conc. drug (log M)
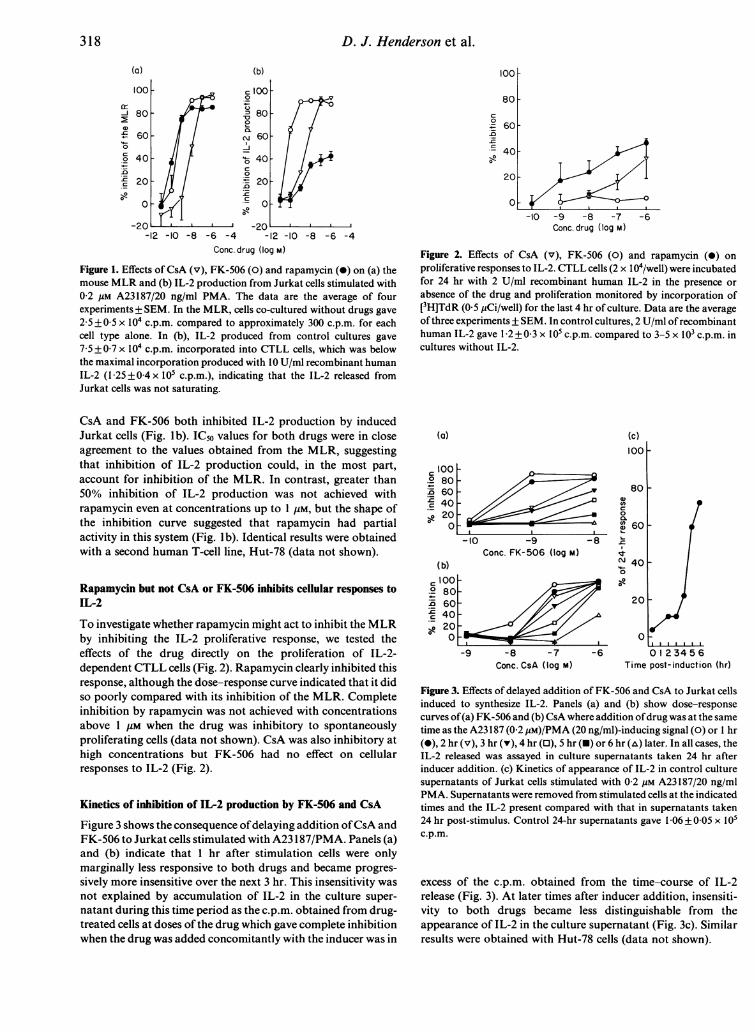
Figure 1. Effects of CsA (v), FK-506 (0) and rapamycin (0) on (a) themouse MLR and (b) IL-2 production from Jurkat cells stimulated with02 gm A23187/20 ng/ml PMA. The data are the average of fourexperiments + SEM. In the MLR, cells co-cultured without drugs gave
2 5 + 0 5 x 104 c.p.m. compared to approximately 300 c.p.m. for eachcell type alone. In (b), IL-2 produced from control cultures gave
7-5+07 x 104 c.p.m. incorporated into CTLL cells, which was belowthe maximal incorporation produced with 10 U/ml recombinant humanIL-2 (1 25+0-4 x 105 c.p.m.), indicating that the IL-2 released fromJurkat cells was not saturating.
80
C:
.
-10 -9 -8 -7 -6Conc. drug (log M)
Figure 2. Effects of CsA (v), FK-506 (0) and rapamycin (0) on
proliferative responses to IL-2. CTLL cells (2 x 104/well) were incubatedfor 24 hr with 2 U/ml recombinant human IL-2 in the presence or
absence of the drug and proliferation monitored by incorporation of[3H]TdR (0 5 pCi/well) for the last 4 hr of culture. Data are the average
ofthree experiments + SEM. In control cultures, 2 U/ml ofrecombinanthuman IL-2 gave 1-2+0-3 x 105 c.p.m. compared to 3-5 x 103 c.p.m. incultures without IL-2.
CsA and FK-506 both inhibited IL-2 production by inducedJurkat cells (Fig. lb). IC50 values for both drugs were in closeagreement to the values obtained from the MLR, suggestingthat inhibition of IL-2 production could, in the most part,account for inhibition of the MLR. In contrast, greater than50% inhibition of IL-2 production was not achieved withrapamycin even at concentrations up to 1 gM, but the shape ofthe inhibition curve suggested that rapamycin had partialactivity in this system (Fig. Ib). Identical results were obtainedwith a second human T-cell line, Hut-78 (data not shown).
Rapamycin but not CsA or FK-506 inhibits cellular responses toIL-2
To investigate whether rapamycin might act to inhibit the MLRby inhibiting the IL-2 proliferative response, we tested theeffects of the drug directly on the proliferation of IL-2-dependent CTLL cells (Fig. 2). Rapamycin clearly inhibited thisresponse, although the dose-response curve indicated that it didso poorly compared with its inhibition of the MLR. Completeinhibition by rapamycin was not achieved with concentrationsabove 1 gm when the drug was inhibitory to spontaneouslyproliferating cells (data not shown). CsA was also inhibitory athigh concentrations but FK-506 had no effect on cellularresponses to IL-2 (Fig. 2).
Kinetics of inhibition of IL-2 production by FK-506 and CsA
Figure 3 shows the consequence ofdelaying addition ofCsA andFK-506 to Jurkat cells stimulated with A23 1 87/PMA. Panels (a)and (b) indicate that I hr after stimulation cells were onlymarginally less responsive to both drugs and became progres-sively more insensitive over the next 3 hr. This insensitivity wasnot explained by accumulation of IL-2 in the culture super-natant during this time period as the c.p.m. obtained from drug-treated cells at doses of the drug which gave complete inhibitionwhen the drug was added concomitantly with the inducer was in
(a)
a 100.2 80.5 60'c 40S 20
0
(b)
(c)
100
01)
0.0)cL
-10 -9
Conc. FK-506 (log M)
-i -7
Conc. CsA (log M)
-8 %.
ci-0
;at
0 1 2345 6Time post-induction (hr)
Figure 3. Effects of delayed addition of FK-506 and CsA to Jurkat cellsinduced to synthesize IL-2. Panels (a) and (b) show dose-responsecurves of(a) FK-506 and (b) CsA where addition ofdrug was at the sametime as the A23 187 (0-2 gM)/PMA (20 ng/ml)-inducing signal (0) or I hr(0), 2 hr (v), 3 hr (v), 4 hr (0), 5 hr (-) or 6 hr (tx) later. In all cases, theIL-2 released was assayed in culture supernatants taken 24 hr afterinducer addition. (c) Kinetics of appearance of IL-2 in control culturesupernatants of Jurkat cells stimulated with 0 2 pm A23187/20 ng/mlPMA. Supernatants were removed from stimulated cells at the indicatedtimes and the IL-2 present compared with that in supernatants taken24 hr post-stimulus. Control 24-hr supernatants gave 1-06 +005 x 105c.p.m.
excess of the c.p.m. obtained from the time-course of IL-2release (Fig. 3). At later times after inducer addition, insensiti-vity to both drugs became less distinguishable from theappearance of IL-2 in the culture supernatant (Fig. 3c). Similarresults were obtained with Hut-78 cells (data not shown).
318
-J
00)
c
w

Immunosuppressive drugs and IL-2 production
(a) NFAT site (b) NFKR site
coCsBFmAs m m~ MAc% LSy 2. -IT 1 2jX=!LLLL LL WLAL O wLzLz z LzcNz Z Zzzz Cu°Iu e> ZZZ-vv0 a
D D:D D up0 en u0 L :wtnnn>LO
Figure 4. (a) Gel retardation of the NFAT oligonucleotide binding site.Nuclear protein extracts were from unstimulated Jurkat cells (U),stimulated cells (S) and stimulated cells treated with drugs. Twoconcentrations of FK-506 (FK) were used (see Fig.), CsA was used at80 nm and rapamycin (Rap) at 10 nm. Competition with excess
unlabelled oligonucleotides is indicated with the prefix c. (b) Gelretardation of the NFKI3 oligonucleotide binding site.
Effects of immunosuppressive drugs on factors binding to the IL-2gene promoter/enhancer
The data from the experiments described above establish thatIL-2 production is a target for both FK-506 and CsA and thatrapamycin acts by a different mechanism. Since induction of IL-2 mRNA is inhibited by CsA and FK-506 but not byrapamycin,8 we were interested in the effects of the three drugson the control of the IL-2 gene promoter. Their effects on theappearance of two inducible IL-2 gene promoter bindingfactors, NFAT and NFKfl,'4-'6 were investigated and theactivity of the constitutive binding factor, NFIL2A,'2 was
assayed as a control.As expected, nuclear extracts from both unstimulated and
stimulated Jurkat cells contained activity binding to and thusretarding the NFIL2A binding site oligonucleotide in gel-shiftexperiments. This was a specific reaction since binding was
competed out with a 20-fold excess of unlabelled NFIL2Abinding site oligonucleotides but not by NFAT or NFKf3binding sites. This activity was not affected by addition of CsA,FK-506 or rapamycin to stimulated cells (not shown).
NFAT binding activity was detected in extracts fromstimulated but not unstimulated cells (Fig. 4a). The appearance
of this binding activity was repressed by 80 nm CsA as well as by2 nm FK-506 but not by 10 nm rapamycin (Fig. 4a). Thespecificity of binding for the NFAT site was demonstrated bycompetition with unlabelled NFAT binding site oligonucleo-tides. An excess of unlabelled NFKf3 binding site oligonucleo-tide did not compete out the binding but, interestingly, theNFIL2A binding site oligonucleotide partially competed outNFAT binding activity (Fig. 4a), although the converse did notoccur. This suggests that the NFAT protein is able to recognizethe NFIL2A binding site sequence.
Specific, inducible NFKfl binding activity was not notice-ably affected by any ofthe drugs at concentrations which inhibit
T-cell activation (Fig. 4b). Again, specificity of the reaction wasdemonstrated by competition experiments.
DISCUSSION
In the MLR, allogeneic stimulation of T cells leads to cytokineproduction and proliferation. This response is thought to mimicevents occurring during transplant rejection. We found thatrapamycin, like CsA and FK-506, is a potent inhibitor of thisproliferative response with an ICso very similar to that of FK-506. However, inhibition by rapamycin was never greater than90%, whereas both CsA and FK-506 gave essentially completeinhibition. This suggests that the mechanism of action ofrapamycin differs from that of FK-506 even though the twodrugs are structurally related and are known to bind to acommon cellular receptor.2223 Distinct modes of action weredemonstrated by their different effects on IL-2 production bytwo separate T-cell lines: Jurkat and HuT-78. A recent report2has also suggested that the mode of action of rapamycin differsfrom that of FK-506 on murine T cells and here we extend thisfinding to include human cells.
The dose-responses for inhibition of IL-2 production byFK-506 and CsA closely parallel the doses required to inhibitthe MLR. This activity can, therefore, account for inhibition ofthe MLR by these two compounds. Rapamycin does not act byblocking IL-2 production. It has recently been reported that,unlike FK-506 and CsA, rapamycin inhibits cellular responsesto IL-2, providing an alternative explanation for its immuno-suppressive effects.'6 We confirm that rapamycin inhibits IL-2action, although the dose-response of the drug was inconsistentwith its effects in the MLR. Rapamycin was unable tocompletely inhibit IL-2-induced proliferation; greater than 50%inhibition was not achieved until toxic concentrations of thedrug were used (> 1 tM). This activity by itself therefore doesnot explain the potent activity of rapamycin in the MLR.Rapamycin does display activity consistent with the dosesrequired to inhibit the MLR in lectin or antibody plus PMA-induced T-cell proliferation2' and in PMA-induced CTLL cellproliferation (our unpublished data). Other assays in whichrapamycin is active, which include Dl0.11.10 hybridoma cellproliferation, DIO.G4 cell proliferation in response to varioussignals2' and IL-4 activity (our unpublished data),2' all give IC50values for the drug in the nanomolar range, 1-2 orders ofmagnitude greater than that required to inhibit the MLR. All ofthese activities suggest that rapamycin is acting to block cellproliferation, although it cannot be discounted that the drugmay inhibit the acquisition of competence to respond to aproliferation signal. Since rapamycin is not generally cytostatictowards spontaneously proliferating cells until micromolarconcentrations are used (unpublished observations), its effectsare selective. It remains to be determined which proliferativesignal in the MLR is the rapamycin-sensitive step, althoughmore than one signal may be involved since a number of growthpromoting cytokines (e.g. IL-2, IL-4) are released by activated Tcells.
In view of the apparent central importance of inhibition ofIL-2 production by FK-506 and CsA in suppression of theMLR, we were particularly interested in the effects of the threedrugs on the appearance of regulatory factors binding to the IL-2 gene promoter. FK-506, like CsA, suppresses the appearanceof IL-2 mRNA following stimulation.8 The mechanism of this
319

320 D. J. Henderson et al.
suppression appears, at least in part, to be due to the absence ofthe IL-2 gene transcription factor NFAT in treated cells. FK-506 inhibited the stimulation-specific appearance ofNFAT buthad no effect on the appearance of another stimulation-specificfactor, NFKf. In this respect, the mechanism of suppression ofIL-2 production by FK-506 appears to be the same as that ofCsA, which also inhibits the appearance of NFAT bindingactivity but does not affect NFK/ except at much higherconcentrations than those used in this study or that are requiredfor immunosuppression.'7" These data confirm the importanceof the NFAT transcription factor in controlling IL-2 geneexpression. Thus, two distinct immunosuppressive drugs, FK-506 and CsA, which act through different receptors, both affectappearance of the same transcriptional regulator. These resultsagree with those of Granelli-Piperno et al., published duringpreparation of this manuscript.29 Unlike FK-506, rapamycinhad no effect on the appearance of NFAT binding activity instimulated Jurkat cells and did not affect the other twotranscription factors investigated. This is consistent with itsreported lack of effect on the induction of IL-2 gene transcrip-tion.8 The suppressive activity of rapamycin on IL-2 production(Fig. lb) appears to be mediated through a different system tothat of FK-506.
Our observation that the NFIL2A binding site partiallycompetes with the NFAT site for NFAT bindings suggests thatNFAT may have binding activity at the NFIL2A site. TheNFIL2A binding site has been shown to activate transcriptionfrom a linked promoter in a stimulation-specific manner.30 Thisactivity is sensitive to CsA although changes in the DNase Iprotection pattern in this region following stimulation were notseen.30 Our result suggests that NFAT interaction with the sitemight be responsible for its stimulation-dependent activity.
NFAT binding activity has been reported to appear less than30 min after T-cell activation and maximum levels are foundafter about 3 hr.'5 The observed kinetics of loss of sensitivity ofJurkat cells to both FK-506 and CsA were slower than this andcorresponded more to the appearance of IL-2 and thus thefunctioning of this transcription factor than to its production.Insensitivity to FK-506 and CsA is probably due to accumu-lation of IL-2 mRNA, which peaks around 6 hr after induc-tion3'1-33 and to the establishment of an autostimulatory IL-2production/response cycle.
ACKNOWLEDGMENTS
We thank Drs E. Wells, R. Hutchinson and R. Eady for helpfuldiscussions and Mrs E. Holness for technical assistance.
REFERENCES
1. THOMSON A.W. (1989) FK 506-how much potential? Immunol.Today, 10, 6.
2. MARTEL R. R., KLICIUS J. & GALET S. (1977) Inhibition of theimmune response by rapamycin, a new antifungal antibiotic. Can. J.Physiol. Pharmacol. 55, 48.
3. THOMSON A.W. (1989) Cyclosporin-Mode of Action and ClinicalApplication. Kluwer Academic Publishers.
4. SAWADA S., SUZUKI G., KAWASE Y. & TAKAKU F. (1987) Novelimmunosuppressive agent FK 506. In vitro effects on the cloned Tcell activation. J. Immunol. 139, 1797.
5. KINo T., HATANAKA H., MIYATA S., INAMURA N., NISHIYAMA M.,YAJIMA T. et al. (1987) FK 506. A novel immunosuppressantisolated from a Streptomyces. II. Immunosuppressive effect of FK506 in vitro. J. Antibiot. 40, 1256.
6. KAY J.E., BENZIE C., GOODIER M., WICK C. & DOE S.E. (1989)Inhibition of T lymphocyte activation by the immunosuppressivedrug FK 506. Immunology, 67, 473.
7. YOSHIMURA N., MATSUI S., HAMASHIMA T. &OKA T. (1989) Effect ofa new immunosuppressive agent, FK 506, on human lymphocyteresponses in vitro. I. Inhibition of expression of alloantigenactivated suppressor cells as well as an induction of alloreactivity.Transplantation, 47, 351.
8. Tocci M.J., MATKOVICH D., COLLIER K., KWOK P., DUMONT F.,LIN S., DEGUDICIBUS S., SIEKIERA J., CHIN J. & HUTCHINSON N.(1989) The immunosuppressant FK 506 selectively inhibits ex-
pression of early T cell activation genes. J. Immunol. 143, 718.9. YOSHIMURA N., MATSUI S., HAMASHIMA T. & OKA T. (1989) Effect of
a new immunosuppressive agent, FK 506, on human lymphocyteresponses in vitro. II. Inhibition of the production of IL-2 andgamma IFN but not B cell stimulating factor 2. Transplantation, 47,356.
10. BIERER B.E., SCHREIBER S. & BURAKOFF S. (1990) Mechanisms ofimmunosuppression by FK 506. Preservation of T cell transmem-brane signal transduction. Transplantation, 49, 1168.
11. ANDRUS L. & LAFFERTY K. (1981) Inhibition of T cell activity bycyclosporin A. Scand. J. Immunol. 15, 449.
12. DURAND D.B., SHAW J.-P., BUSH M., REPLOGLE R., BELAGAJE R. &CRABTREE G.R. (1988) Characterisation of antigen receptor re-
sponse elements within the Interleukin-2 enhancer. Mol. Cell. Biol.8, 1715.
13. SERFLING E., BARTHELMAS R., PFEUFFER I., SCHENK B., ZARIUS S.,SWOBODA R., MERCURIO F. & KARIN M. (1989) Ubiquitous andlymphocyte specific factors are involved in the induction of themouse Interleukin-2 gene in T lymphocytes. EMBO J. 8, 465.
14. BRUNVAND M.W., SCHMIDT A. & SIEBENLIST U. (1988) Nuclearfactors interacting with mitogen responsive regulatory region of theInterleukin-2 gene. J. Biol. Chem. 263, 18904.
15. SHAW J.-P., UTZ P., DURAND D., TOOLE J., EMMEL E. & CRABTREEG.R. (1988) Identification of a putative regulator of early T cellactivation genes. Science, 241, 202.
16. HoYos B., BALLARD D., BOHNLEIN E., SIEKEVITZ M. & GREENE W.C.(1989) Kappa-fl specific DNA binding proteins: role in regulation ofhuman Interleukin-2 gene expression. Science, 244, 457.
17. EMMEL E.A., VERWEIJ C., DURAND D., HIGGINS K., LACY E. &CRABTREE G.R. (1989) Cyclosporin A specifically inhibits functionof nuclear proteins involved in T cell activation. Science, 246, 1617.
18. RANDAK C., BRABLETZ T., HERGENROTHER M., SOBOTTA I. &
SERFLING E. (1990) Cyclosporin A suppresses the expression of theinterleukin-2 gene by inhibiting binding of lymphocyte specificfactors to the IL-2 enhancer. EMBO J. 9, 2529.
19. MORRIS R.E. & MEISER B.M. (1989) Identification of a new
pharmacologic action for an old compound. Med. Sci. Res. 17, 609.20. CALNE R.Y., COLLIER D.ST J., LIM S., POLLARD S., SAMAAN A.,
WHITE D. & THIRU S. (1989) Rapamycin for immunosuppression inorgan allografting. Lancet, ii, 227.
21. DUPONT F.J., STARUCH M., KOPRAK S., MELINO M. & SIGAL N.H.(1990) Distinct mechanisms of suppression of murine T cellactivation by the related macrolides FK 506 and rapamycin. J.Immunol. 144, 251.
22. DUMONT, F.J., MELINO M., STARUCH M., KOPRAK S., FISCHER P. &SIGAL N.H. (1990) The immunosuppressive macrolides FK 506 andrapamycin act as reciprocal antagonists in murine T cells. J.Immunol. 144, 1418.
23. HARDING M.W., GALAT A., UEHLING D. & SCHREIBER S.L. (1989) Areceptor for the immunosuppressant FK 506 is a cis-trans peptidylprolyl isomerase. Nature, 341, 758.
24. GORSKI K., CARNEIRO M. & SCHIBLER U. (1986) Tissue-specific in

Immunosuppressive drugs and IL-2 production 321
vitro transcription from the mouse albumin promoter. Cell, 47, 767.25. RHODES D. (1987) Analysis of sequence specific DNA binding
proteins. In: Methods in Enzymology 152. Guide to MolecularCloning (eds S. L. Berg and A. R. Kimmel). Academic Press.
26. SAMBROOK J., FRITSCH E. & MANIATIS T. (1989) Molecular Cloning.A Laboratory Manual. Cold Spring Harbor Laboratory Press.
27. MANGER B., WEISS A., WEYAND C., GORONZY J. & STOBO J.D.(1985) T cell activation. Differences in the signals required for IL-2production by non-activated and activated T cells. J. Immunol. 135,3669.
28. WISKOCIL R., WEISS A., IMBODEN J., KAMIN-LEWIS R. & STOBO J.D.(1985) Activation of a human T cell line: A two stimulus require-ment in the pretranslational events involved in the co-ordinateexpression of Interleukin-2 and gamma interferon genes. J. Immu-nol. 134, 1599.
29. GRANELLI-PIPERNO A., NOLAN P., INABA K. & STEINMAN R.M.(1990) The effects of immunosuppressive agents on nuclear factors
that bind to sites on the Interleukin-2 promoter. J. exp. Med. 172,1869.
30. ULLMAN K.S., NORTHROP J., VERWEU C. & CRABTREE G.R. (1990)Transcription signals from the T lymphocyte antigen receptor to thegenes responsible for cell proliferation and immune function: Themissing link. Ann. Rev. Immunol. 8, 421.
31. REED J.C., ALPERS J., NOWELL P. & HOOVER R.G. (1986) Sequentialexpression of protooncogenes during lectin stimulated mitogenesisof normal human lymphocytes. Proc. natl. Acad. Sci. U.S.A. 83,3982.
32. WEiss A., SCHIELDS R., NEWTON M., MANGER B. & IMBODEN J.(1987) Ligand receptor interactions required for commitment to theactivation of the Interleukin-2 gene. J. Immunol. 138, 2169.
33. JuNE C.H., LEDBETTER J., LINDSTEN T. & THOMPSON C.B. (1989)Evidence for the involvement of three distinct signals in theinduction of IL-2 gene expression in human T lymphocytes. J.Immunol. 143, 153.