conformational transitions of immunoglobulin fragment fc(t) incited by alkyl sulfates of various...
TRANSCRIPT

BIOCHIMICA ET BIOPHYSICA ACTA 631
BBA 35783
CONFORMATIONAL TRANSITIONS OF IMMUNOGLOBULIN FRAGMENT
Fc(t) INCITED BY ALKYL SULFATES OF VARIOUS HYDROPHOBIC
CHAIN LENGTHS
PAK KAI JANE LEE AND BRUNO JIRGENSONS*
The Umvers,ty of Texas, M. D. Anderson Hospital and Tumor Instztute at Houston, Department of Biochem,stry, Houston, Texas 77o25 (U.S.A.)
(Received November 9th, 197 o)
SUMMARY
The effect of alkyl sulfates of various chain lengths on immunoglobulin frag- ment Fc(t) was studied. Circular dichroism measurements indicated that the non- helical polypeptide chains of Fc(t) were ordered by anionic detergents to a certain degree. Resolution of the eUipticity curves into Gaussian peaks showed that the detergents incited the formation of negative bands centered at 206 and at 222 nm and a positive band at 19o-192 nm. This partial transition to a-helical structure was enhanced by increasing amount of detergents until the critical micelie concentrations of the detergents were reached, when no further change was observed with additional detergents. The effect of ionic strength seemed to be on the critical micelle concen- tration. These conformational changes were also influenced by pH and the hydro- phobic chain length of the detergents. In acid solution, anionic detergents appeared to both promote a-helix formation and reduce the amount of disordered structure present in Fc(t). The order of effectiveness of the detergents as helix-forming agents was tetradecyl > dodecyl > decyl > octyl sodium sulfate, indicating the importance of hydrophobic bonding in Fc(t). In near ultraviolet zone, the circular dichroic bands were weakened by the detergents, although the Cotton effects were not completely eliminated.
INTRODUCTION
The interaction of proteins with detergents has been extensively studied and several reviews are available (e.g. refs. I, 2). Depending on the nature of protein
Abbreviations: IgG, Fab and Fc recommended by the World Health Organization 15" Fab(t) and Fc(t) used for the tryptic digestion fragments similar to the papain digestion fragments Fab and Fc; CD, circular dichroism.
* Postal Address: The University of Texas M. D. Anderson Hospital and Tumor Institute, Department of Biochemistry, 6723 Bertner Avenue, Houston, Texas 77o25, U.S.A.
Biochim. Bwphys. Acta, 229 (1971) 631-641

632 P . K . J . LEE, B. JIRGEI~SONS
under investigation and study of such conditions as temperature, pH and ionic strength, detergents can cause dissociation 3, association 4, precipitation 5, complex formation and denaturationS, v.
The conformational changes of the protein molecules induced by detergents have been investigated by methods of optical rotatory dispersion (see refs. 8-12), ultraviolet absorption difference spectrum (e.g. refs. 12, I3), and recently, by circular dichroism (CD) 14.
The present work offers data on the conformational transitions of IgG fragment Fc(t) incited by alkyl sulfates of various chain lengths as shown by CD spectra, The dependence of the conformation on detergent concentrations and on the hydro- phobic chain length of detergents are emphasized.
MATERIALS AND METHODS
Normal human IgG was obtained from Pentex Inc. (Kankakee, Ill.). Highly purified sodium octyl, decyl, dodecyl and tetradecyl sulfates were obtained from Mann Research Laboratories (New York, N.Y.). All other reagents used were of analytical grade.
Preparation of normal human Fc(t). Partial reduction and alkylation of the interchain disulfide bonds of normal human IgG were done according to EDELMAN et al. le. The tryptic digestion fragment Fc(t) was obtained by the procedure of EDEL- MAN et al. le modified by DoI AND JIRGENSONS 17. Symmetrical chromatographic peaks indicated single components in the Fc(t) preparations. Homogeneity of Fc(t) was determined by cellulose acetate electrophoresis and the antigenic specificity was checked by immunoelectrophoresis using goat antiserum against human Fc.
Sedimentation. All determinations were made with a Beckman-Spinco Model E ultracentrifuge at a rotor speed of 59 780 rev./min and at 20.0 °. Sedimentation coefficients were corrected to the standard conditions.
Viscosity. Viscosity experiments were performed with a Cannon-Ubelhode dilution viscometer at 28.0 °. The data were plotted as ~sp/c against c, where ~sp is the specific viscosity and c is the concentration in g/Ioo ml. The intrinsic kinematic viscosity [*/] was obtained by extrapolation to infinite dilution.
Molecular weight. Molecular weight of Fc(t) was calculated from the intrinsic viscosity and sedimentation coefficient by using the equation of SCHERAGA AND MANDELKERN ls, in which fi was assumed to be 2.6.IO e (ref. 19) and the partial specific volume was assumed to be 0.73 cm 3 (ref. 16).
Protein concentration. Protein concentrations were determined using a Beckman DU spectrophotometer. --xEX%cm of 15 at 280 nm was used 17.
CD measurements and resolutions of ellipticity curves. The new Model CD-SP Durrum-Jasco highly sensitive dichrograph was used. The sensitive scale settings of 2. lO -5, 5" lO-5 or I - lO -4 dichroic absorbance difference (zle) per I cm on the recorder chart was used. The optical path in the near ultraviolet was i.o cm and it was o.I or 0.05 cm in the far ultraviolet spectral region from 250 nm down to a little below 19o nm. The errors in this spectral zone, in terms of mean residual molar ellipticities were between -4- 400 and -¢- 600 degrees-cm 2 per decimole. They were much smaller at longer wave lengths. The performance of the instrument was checked by using solutions of camphor sulfonic and poly-L-glutamic acids. All recordings were made
Biochim. Biophys. Acta, 229 (I97I) 631-641

CD OF IgG FRAGMENT Fc(t) WITH ALKYL SULFATES 633
at an ambient room temperature of 23-25 ° . The data were calculated as mean residual molar ellipticities E0]. The mean residue weight of lO 9 was used 17. Reduced mean residual molar ellipticities EO'l, in the dilute salt solution and acid solution were calculated taking the refractive indices of water (see ref. 2o), EO'l = E01" E3/(n2+2)]~2o. The eUipticity curves were constructed from at least three circular dichroism re- cordings by calculating the mean residual molar ellipticities in 2-nm intervals. The ellipticity curves were resolved into Gaussian peaks by means of the DuPont 31o curve resolver.
pH measurement, pH measurements were carried out on a Coming Model 12 research pH meter with a combination electrode. Standard buffer solutions of pH 4.00 and 7.00 (certified by Fisher Scientific Co., Pittsburgh, Pa.) were used to calibrate the pH meter.
Interaction of anionic detergents with Fc(t). Stock protein solutions for the near ultraviolet CD measurements were made up of approx. 0.2% Fc(t) in water. For far ultraviolet measurements, I/IO dilutions of the above stock solution were used. The concentrations were checked from absorbance at 280 nm.
Stock detergent solutions were prepared by dissolving known amounts of sodium octyl, decyl, dodecyl, or tetradecyl sulfates in either water or other solvents, e.g. 0.02 and 0.2 M Na2SO 4, acidified with H2SO 4, whenever necessary to achieve the desired pH.
All samples were prepared by diluting I.O ml of stock Fc(t) solution with known amounts of detergent and the volume made up to 2.0 ml by addition of appropriate salt solutions. Thus the final solutions consisted of either o.I % or o.oi % Fc(t) and various concentrations of detergent in either water, o.oi M or o.I M Na2SO 4 at certain pH. The samples were kept at room temperature, 23-25 °, for 16-24 h or heated in a water bath at 5 °° for 2 h. In the latter case, CD measurements were performed after complete cooling of the solutions to room temperature.
RESULTS
Characterization of Fc(t) The intrinsic viscosity of Fc(t) was o.o512 dl/g and sedimentation coefficient
S~o,w of 3.75 S was obtained, giving a molecular weight of 51 ooo. These values were similar to those obtained by DoI AND .IIRGENSONS 17 indicating that the preparations were practically the same. The far ultraviolet CD spectrum showed a negative minimum at 217 nm with a definite shoulder at 229 nm and a positive peak at 202 nm (Fig. I). Resolution of this curve gave four Gaussian peaks with a positive peak at 202 nm and negative peaks at 217, 232 and 239-240 nm. The reduced mean residual molar ellipticities at 217 and 202 nm were --173o and + lO5O degrees, cm2/decimole, respectively.
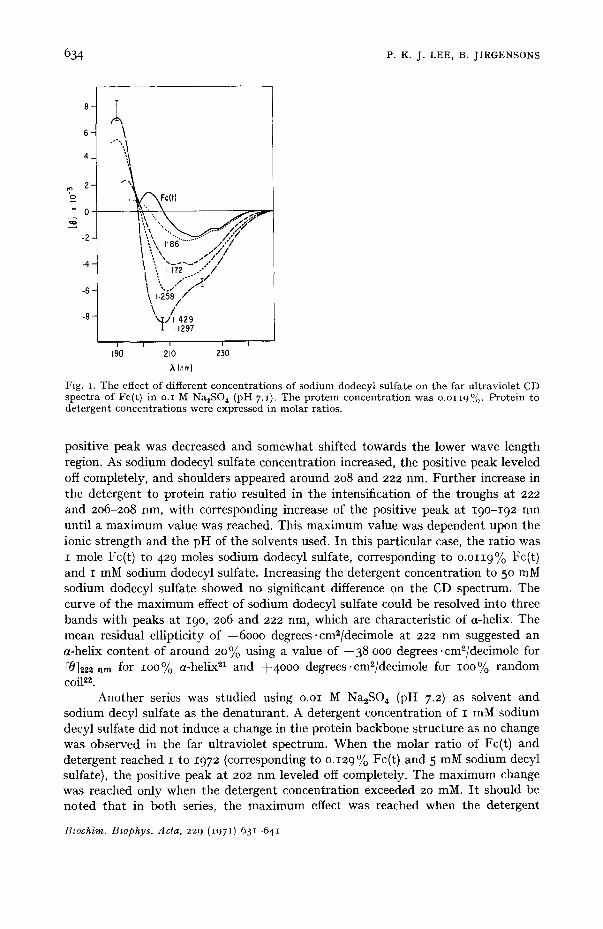
Effect of different concentrations of anionic detergents on Fc(t) The effect of a series of sodium dodecyl sulfate concentrations on o.o119%
Fc(t) in o.I M Na~SO 4 (pH 7.1) in the far ultraviolet is shown in Fig. I. At detergent concentrations below o.2 mM sodium dodecyl sulfate no change was observed. When the protein to detergent ratio was increased to I mole Fc(t) to 86 moles sodium dodecyl sulfate, the negative peak at 217 nm increased very slightly while the
Biochim. Biophys. Acta, 229 (1971) 631-641
634 P . K . J . LEE, B. JIRGENSONS
6- \
4 - """t
o_ (t)
-2- 11', ,.8~ .......... , "V Ix , , ./',-'/
-" ! - i ' r ~ ' - . . . " / ",,. ..,-"
\ 1.25B// \ /
-8 '~T,, / 1 429 .L I1297
I I I I I I
190 2 IO 230 ~ . ( n m )
Fig. I. The effect of different concen t ra t ions of sod i um dodecyl sul fa te on the far u l t rav io le t CD spec t ra of Fc(t) in o.I M NazSO, (pH 7.1). The pro te in concen t r a t ion was o.o119°,o. Pro te in to de te rgen t concen t ra t ions were expressed in molar ratios.
positive peak was decreased and somewhat shifted towards the lower wave length region. As sodium dodecyl sulfate concentration increased, the positive peak leveled off completely, and shoulders appeared around 208 and 222 rim. Further increase in the detergent to protein ratio resulted in the intensification of the troughs at 222 and 206-208 nm, with corresponding increase of the positive peak at 19o-192 nm until a maximum value was reached. This maximum value was dependent upon the ionic strength and the pH of the solvents used. In this particular case, the ratio was I mole Fc(t) to 429 moles sodium dodecyl sulfate, corresponding to o.o119% Fc(t) and I mM sodium dodecyl sulfate. Increasing the detergent concentration to 50 mM sodium dodecyl sulfate showed no significant difference on the CD spectrum. The curve of the maximum effect of sodium dodecyl sulfate could be resolved into three bands with peaks at 19o, 206 and 222 nm, which are characteristic of a-helix. The mean residual ellipticity of --6000 degrees .cm2/decimole at 222 nm suggested an a-helix content of around 20% using a value of --38 ooo degrees .cm2/decimole for [01222 nm for lOO% a-helix 21 and +4000 degrees.cm2/decimole for lOO% random coil iz.
Another series was studied using o.oi M NaiSO4 (pH 7.2) as solvent and sodium decyl sulfate as the denaturant. A detergent concentration of I mM sodium decyl sulfate did not induce a change in the protein backbone structure as no change was observed in the far ultraviolet spectrum. When the molar ratio of Fc(t) and detergent reached I to 1972 (corresponding to o.129 % Fc(t) and 5 mM sodium decyl sulfate), the positive peak at 202 nm leveled off completely. The maximum change was reached only when the detergent concentration exceeded 20 mM. I t should be noted that in both series, the maximum effect was reached when the detergent
B~ochim. Blophys. Acta, 229 (1971) 631-641
C D OF IgG FRAGMENT Fc(t) WITH ALKYL SULFATES 635
concentrations approached their critical micelle concentrations. At 25 °, the critical micelle concentration for sodium dodecyl sulfate in o.I M Na2SO 4 and for sodium decyl sulfate in o.oi M Na2SO 4 are 1.2 and 21 mM, respectively 23.
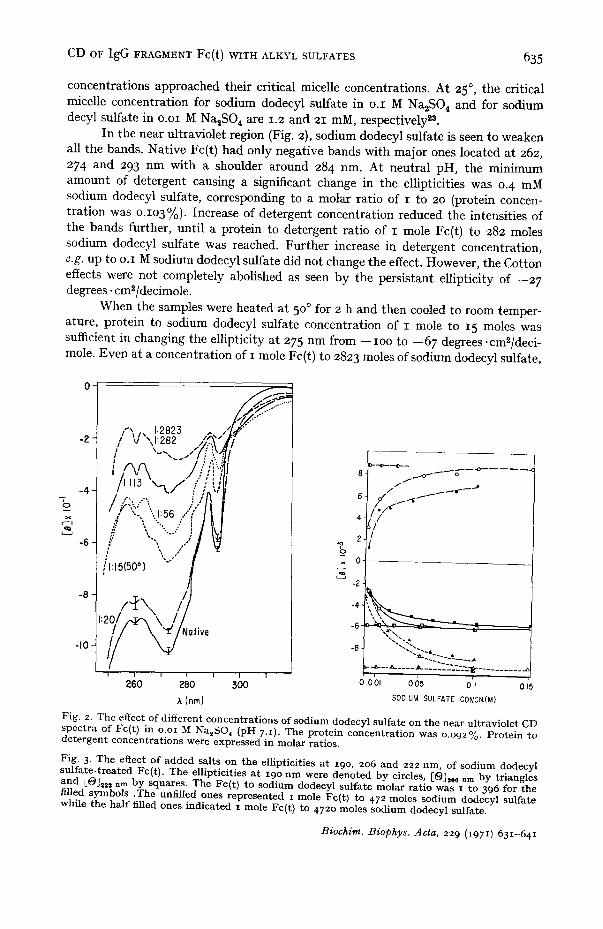
In the near ultraviolet region (Fig. 2), sodium dodecyl sulfate is seen to weaken all the bands. Native Fc(t) had only negative bands with major ones located at 262, 274 and 293 nm with a shoulder around 284 nm. At neutral pH, the minimum amount of detergent causing a significant change in the ellipticities was 0. 4 mM sodium dodecyl sulfate, corresponding to a molar ratio of I to 20 (protein concen- tration was O.lO3%). Increase of detergent concentration reduced the intensities of the bands further, until a protein to detergent ratio of I mole Fc(t) to 282 moles sodium dodecyl sulfate was reached. Further increase in detergent concentration, e.g. up to o.I M sodium dodecyl sulfate did not change the effect. However, the Cotton effects were not completely abolished as seen by the persistant ellipticity of --27 degrees, cm~/decimole.
When the samples were heated at 5 °0 for 2 h and then cooled to room temper- ature, protein to sodium dodecyl sulfate concentration of I mole to 15 moles was sufficient in changing the ellipticity at 275 nm from --IOO to --67 degrees.cm~-/deci - mole. Even at a concentration of I mole Fc(t) to 2823 moles of sodium dodecyl sulfate,
- 2 t ' \ t , 1 2 8 2 3
/ ~ i ' k 1 : 2 8 2
-4 ~ / L . / "
-'::' I / - ' ' ..... !
- - t e , : ] e ~ : , ¢' --, ',, : 0 0 , - ' ;
-6 / "~°'/"J:i ! H5(50 ° )
1 : 2 0 /
- I 0 -
lalive
/ t r" "~
4 -
!
-4 . \ ~
- 6 - ~ , , ~ , . _ ,~
-s . . . . -:. ~ - ~ - - - ~ . . . . ~ . . . . ~---_Z.-.."_.-~ . . . . . . . . . . .2
2 -
o x 0 -
-2
, , i , t t i
260 280 300 o out o()~ o'f o~5 ~ . ( n r n ) SODIUM SULFATE CONCN.(M}
Fig. 2. The effect of different concent ra t ions of sodium dodecyl sulfate on the near ul t raviolet CD spect ra of Fc(t) in o.oi M Na,SO 4 (pH 7.1). The prote in concentra t ion was 0.092%. Prote in to de te rgen t concent ra t ions were expressed in molar ratios.
Fig. 3. The effect o f added salts on the ellipticities a t 19o, 206 and 222 nm, of sodium dodecyl su l fa te - t rea ted Fc(t). The ellipticit ies a t 19o n m were denoted by circles, [6 ) ] , 6 nm by tr iangles and [O],2z nm by squares. The Fc(t) to sodium dodecyl sulfate molar rat io was i to 396 for the filled symbols .The unfilled ones represented I mole Fc(t) to 472 moles sodium dodecyl sulfate while the ha l f filled ones indicated z mole Fc(t) to 472o moles sodium dodecyl sulfate.
Biochim. Biophys. Acta, 229 (1971) 631-64t

636 P.K.J. LEE, B. JIRGENSONS
the [0],~5 am remained around --2o degrees-cm*/decimole and could not be reduced further.
Effect of salt concentration on sodium dodecyl sulfate denaturation of Fc(t) Fc(t) was dissolved in water or Na~SO 4 of various concentrations and the
effect of sodium dodecyl sulfate was observed (Fig. 3). Since the final pH varied only slightly (from pH 7.2 in water and pH 6. 9 in 0.2 M Na2SO4) no adjustment of pH was made. When a detergent concentration of I mM sodium dodecyl sulfate was used, the [0J,22 nm was increased from --3300 degrees .cm2/decimole in water to --6000 degrees .cm2/decimole in o.I M Na,SO a. No further change was observed when the salt concentration was raised to 0.2 M.
At higher detergent concentration (o.oi M) which is above the critical micelle concentration of sodium dodecyl sulfate even in water ~3, no salt effects were observed. Moreover, at this high concentration of detergent, the maximum change in CD spectra of Fc(t) at neutral pH was reached by sodium dodecyl sulfate alone, thus, the presence of salt did neither enhance nor inhibit the maximum conformational transition of Fc(t) incited by sodium dodecyl sulfate.
Also, NaC1 was used as its effect on the critical micelle concentration of sodium dodecyl sulfate has been extensively studied 23-25. Results similar to that of Na2S04 were obtained. That is, at low sodium dodecyl sulfate concentration, the intensities of the three resolved bands of the far ultraviolet spectra increased with the addition of salt until the molarity of NaC1 was 0.25 M; the critical micelle concentration of
T A B L E I
E F F E C T O F O C T Y L (C8) , D E C Y L (Clo) , D O D E C Y L (C12) A N D T E T R A D E C Y L (C14) S O D I U M S U L F A T E S ON Fc( t ) AT NEUTRAL p H
Detergent Concn. Solvent [6)'] * E(~'] * [6) '] * (moles~mole 19o nm 206 nm 22~ nm Fe ( t ) )
Cs 37 44 ° Clo 4 6 8 0 C1~ 479
C 8 46 800 C1° 7 0 2 0 C1, 2 3 6
Cs 93 6 0 0 C10 i i 7 0 0 C12 2 34 ° C14 472
C s 116 97 ° C10 7 889 C1~ I 547 C14 472
C1, 172 C14 79
W a t e r + 3 0 8 0 - - 345 ° - - 2600 W a t e r + 1 4 7 o - - 2 i o o - - 2 1 2 o W a t e r + 147o - - 18oo - - 212o
W a t e r + 2 6 2 0 - - 4 2 8 o - - 325 ° W a t e r + 3 8 2 o - - 5 1 3 ° - - 3 2 5 ° W a t e r + 272o - - 3 0 0 0 - - 325 °
W a t e r + 5 8 7 o - - 6 2 1 o - - 3 8 0 0 W a t e r + 5 4 3 o - - 5 2 o o - - 4 ° 0 0 W a t e r + 5 6 5 0 - - 55 ° 0 - - 4 ° 0 0 W a t e r + 5 4 3 o - - 5 5 ° 0 - - 4 ° 8 0
o . o i M N a ~ S O , + 7 5 0 0 - - 6 0 5 0 - - 4 3 2 0 o . o i M N a 2 S O 4 + 5 2 2 0 - - 4 6 1 o - - 4 1 7 ° o . o i M N a 2 S O 4 + 6 2 5 0 - - 6 6 8 0 - - 4 3 2 0 o . o i M N a 2 S O 4 + 6 7 5 o - - 5 9 0 0 - - 4 6 0 0
o . i M N a 2 S O 4 + 1 8 o o - - 1 8 7 5 - - 2 8 0 0 o . i M N a ~ S O 4 + 2 4 3 o - - 1 8 o o - - 3 o o o
* T h e r e d u c e d m e a n r e s i d u e e l l i p t i c i t i e s a r e e x p r e s s e d in d e g r e e s , c m 2 I d e c i m o l e .
Biochim. Biophys. Acta, 2 2 9 (1971) 6 3 1 - 6 4 1

CD OF IgG FRAGMENT Fc(t) WITH ALKYL S U L F A T E S 637
sodium dodecyl sulfate at 25 ° in o.25 M NaC1 is I . I mM (ref. 25). No such increase was observed at higher (o.oi M) sodium dodecyl sulfate concentration.
Effect of the hydrophobic tail length of detergents on the conformational transition of Fc (t) The effect of anionic detergents with different hydrocarbon chain lengths on
the conformation of Fc(t) is shown in Table I. The CD curves of these systems indi- cated the formation of a-helix as evidenced by the positive peak at 19o nm and negative troughs at 206 and 222 nm. According to the data, tetradecyl sodium sulfate was more effective than dodecyl sodium sulfate; dodecyl sodium sulfate was more effective than decyl sodium sulfate, while the latter was more potent than octyl sodium sulfate.
Tetradecyl sodium sulfate was very difficult to dissolve, especially in 0.2 M Na,SO 4 (pH 6.9), as only concentrations lower than 0.2 mM detergent would form a soluble complex with o.o129% Fc(t). At this ratio of I mole Fc(t) to 79 moles sodium tetradecyl sulfate, the effect was comparable to I mole Fc(t) to 172 moles sodium dodecyl sulfate. The concentration of tetradecyl sodium sulfate needed for complete transition of the conformation of Fc(t) was impossible to determine.
However, tetradecyl sodium sulfate dissolved in water with more ease, although the maximum concentration of detergent that could be used was still limited. At detergent concentration of o.I and 0.2 mM, little difference from the native confor- mation was observed. The effectiveness of this C14 detergent was demonstrated by the 1. 5 mM needed to obtain a change in the far ultraviolet spectra of Fc(t) compar- able to the effect of 6.0 mM sodium dodecyl sulfate.
Octyl and decyl sodium sulfates dissolved in water and salt solutions easily. However, considerably greater amounts of these lower homologues were needed to produce the same effect. For example, 116 97 ° molecules of octyl and 7889 molecules of decyl sodium sulfate had comparable effect to 1547 molecules of sodium dodecyl sulfate on I molecule of Fc(t). As with sodium dodecyl sulfate, the maximum change in the ellipticities in the far ultraviolet spectra of Fc(t) was obtained only when the concentration of the detergents approached their critical micelle concentration.
Effect of acid on the conformational transitions incited by anionic detergents CD spectra in both the far and near ultraviolet regions indicated that acid
enhanced the effect of sodium dodecyl sulfate on Fc(t). The intensities of the bands in the near ultraviolet zone were decreased when the pH was changed from 7.1 to 2.1, indicating that the side chains were less restricted in acid. In the far ultraviolet, the ellipticity at 222 nm increased from --5o00 to --75o0 degrees .cm*/decimole. Thus the helical content was increased from about 21% at pH 7.2 to 27% at pH 2.1. The ellipticities at 19o and 2o6 nm also increased in acid solution, however, [0]xg0 n m
increased proportionally more than [0],o6 nm. The complex formation between protein and detergent was quite evident in
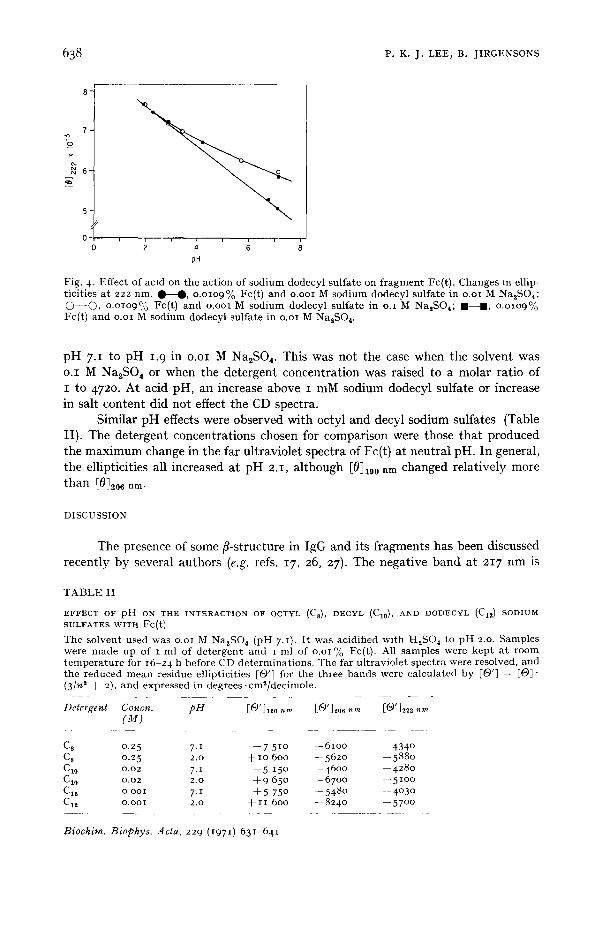
acid solution. Addition of a small amount of detergent caused flocculation, and the precipitate dissolved in excess detergent. Since precipitation occurred below a protein to detergent concentration of i mole Fc(t) to 392 moles of sodium dodecyl sulfate in o.i M Na2SO,, comparisons below this concentration were impossible. Fig. 4 shows the changes in [0]~2 n m with pH. At low detergent concentration (I mole Fc(t) to 472 moles sodium dodecyl sulfate), the increase was linear from
Biochim. Biophys. Acta, 229 (1971) 631-64I
638 P . K . J . LEE, B. JIRGENSONS
8"
7 ,'?
6 qb
| n - - I , , , u ,
2 4 6 8 pH
Fig. 4. Effect of acid on the ac t ion of sod ium dodecyl sul fa te on f r a g m e n t Fc(t). Changes in ellip- t ic i t ies a t 222 rim. q ~ - - t , O.OLO9% Fc(t) and o.ooi M sod ium dodecyl su l fa te in o .oi M Na2SO4; © - - © , O.OlO9°,o Fc(t) and o.ooi M sod ium dodecyl sul fa te in o.I M Na,SO4; l - - l , O.OLO9% Fc(t) and o.oi M sod ium dodecyl sul fa te in 0 .oi M Na,SO 4.
pH 7.1 to pH 1. 9 in o.oi M Na,S04, This was not the case when the solvent was o.I M Na2SO * or when the detergent concentration was raised to a molar ratio of I to 4720. At acid pH, an increase above I mM sodium dodecyl sulfate or increase in salt content did not effect the CD spectra.
Similar pH effects were observed with octyl and decyl sodium sulfates (Table II). The detergent concentrations chosen for comparison were those that produced the maximum change in the far ultraviolet spectra of Fc(t) at neutral pH. In general, the ellipticities all increased at pH 2.1, although [01190 n m changed relatively more than [0120e nm.
DISCUSSION
The presence of some fl-structure in IgG and its fragments has been discussed recently by several authors (e.g. refs. 17, 26, 27). The negative band at 217 nm is
T A B L E I I
EFVECT OV p H ON THE INTERACTION OF OCTYL (Cs), DECYL (Ca0), AND DODECYL (Clz) SODIUM SULVATES WITH Fc(t)
The so lven t used was o.oi M Na2SO 4 (pH 7.1). I t was acidif ied w i t h H2SO 4 to p H 2.o. Samples were made up of I ml of d e t e r g e n t and I ml of o , o i % Fc(t). All samples were k e p t a t room t e m p e r a t u r e for 16-24 h before CD de te rmina t ions . The far u l t r av io l e t spec t ra were resolved, and the reduced mean res idue e l l ip t ic i t ies [(~'] for the three bands were ca lcu la ted b y [6~'] ~ [6)]' (3/n 2 + 2), and expressed in degrees .cm2/dec imole .
Detergent Cohen. (M)
Cs 0.25 Cs 0.25 el0 o,o2 C10 0.02 CI~ o ooi C12 O.OOI
p H [6)'11~o nm [O']206 ,m [tg']z2~ n,n
7 .1 + 7 5 lo - -61oo --434 ° 2.0 + I O 6oo --562o --588o 7 .I + 5 15o --46oo 428o 2.0 + 9 650 --6700 -- 51°° 7 -1 + 5 75 ° - -5480 --403 ° 2.o + I I 600 -- 8240 --5700
Biochim. Biophys. Acta, 229 (1971) 631-641

CD oF IgG FRAGMENT Fc(t) WITH ALKYL SULFATES 639
probably due to n-n* transition of polypeptides in the fl-conformation as predicted from calculations of the ultraviolet optical properties .8. WooDY .9 calculated that some of the exciton component of ze-ze* transitions of antiparallel pleated sheet structure may produce positive CD bands around I9o-21o nm regions and that the actual positions of these bands depend on the sheet length and width in addition to environmental interactions. This could account for the 2o2-nm positive peak of Fc(t) which is different from the I95-nm peak of poly-L-lysine ~2. The weakness of both the negative band at 217 nm and the positive band at 2o2 nm indicates that the amount of fl-structure in Fc(t) is low.
The ability of anionic detergents to induce helix formation in nonhelical proteins has been demonstrated1°, 14. Our CD results suggest that alkyl sulfates are helix-promoting agents in the pH range studied (pH 1.9-7.2 ), however, the amount of helix formed in Fc(t) is low. We showed that this change in conformation also depends on the detergent concentrations, pH, ionic strength, and very noticeably on the length of the hydrocarbon chain of the anionic detergents.
At very low protein to detergent concentrations, no change in the far ultra- violet spectrum was observed, but the near ultraviolet bands were considerably decreased. This weakening of the Cotton effects in the 250-32o-nm region is indicative of the increased rotational freedom of the protein side chains. In fact, the effect in the near ultraviolet approached maximum before any changes in the far ultraviolet spectra were observed. The change in the far ultraviolet is evidenced by a decrease in the positive 2o2-nm peak and a slight increase in the 217-nm negative peak. This transition could be due to the formation of some disordered structure 8 which is characterized by a large negative peak at 197-198 nm 21. I t is expected that a strong negative band around 200 nm is sufficient to cancel the 2o2-nm positive peak com- pletely. Recently, TIFFANY AND KRIMM s0 and DEARBORN AND WETLAUFER 31 have demonstrated that circular dichroism spectra of disordered proteins might be quite different from the CD spectra of unordered poly-L-lysine and poly-L-glutamic acid. They also showed the existence of negative ellipticities at 217-nm region rather than a small positive band, which was observed in poly-L-lysine ~2. This would explain the slight increase in [01317 nm.
With increasing detergent concentration, the formation of a-helical structure is evidenced, as the bands at I90, 206 and 222 nm became clear and intensified. Although a precise quantitation of the helix content is not warranted2m, an esti- mation for comparison is possible from the ellipticities of any of the three bands characteristic of the a-helix 21. The n-~e* transition band centered at 222 nm was used for a first approximation of the a-helix content. The low amount of about 20% a-helix can be expected because of the restrictions of the intrachain disulfide bonds 1.; the high content of valine, threonine, serine and proline residues in Fc(t) 83 may also hinder helix formation 34.
The higher amplitude of the 2o6-nm band than the 222-nm band may be caused by some disordered structures persisting in the detergent-induced structure of Fc(t). This is also suggested by the unproportional increase of [0118o nm and [0]306 nm in acid solutions. The decrease in the amount of disordered structure and its charac- teristic 20o-nm CD extremum would cancel some of the intensity of the 2o6-nm trough while enhancing the I90-nm positive maximum. Moreover, the lowering of pH of the Fc(t)-detergent solutions from neutral to acidic also resulted in the for-
Biochzm. Biophys. Acta, 229 (1971) 631-64I

640 P . K . J . LEE, B. JIRGENSONS
mation of significantly more ordered structure of the protein backbone, evidenced by the marked increase of E012~2 nm (see Fig. 4)-
In acid solution, CD spectra indicated unfolding of Fc(t) due to electrostatic repulsion which overcame the binding forces 17. The increased effect of anionic de- tergent as a-helix former in acid may be attributed to the unfolding of the protein so as to expose more sites for the hydrophobic attack of the detergents. The affinity of positively charged sites of Fc(t) to the negatively' charged detergents may also be a factor promoting the conformational transition incited by the anionic detergents in acid.
The decrease in the intensities of the bands in the near ultraviolet region by the action of anionic detergents indicated that the amino acid side chains are "freer". However, the persistance of the Cotton effects suggested that there are still some restrictions and the side chains are not completely free. This weakening of the CD bands in the near ultraviolet zone by sodium dodecyl sulfate has been shown in other proteins 14.
In the far ultraviolet region, addition of salts did not affect the conformational transition of Fc(t) induced by sodium dodecyl sulfate if the detergent concentrations were above the critical micelle concentration. Fig. I shows the intensity of t0]100 nm, [L01206 nm and E03~2~ nm is maximum when sodium dodecyl sulfate concentration approached the critical micelle concentration, indicating the importance of the presence of detergent micelles in the formation or stabilization of a-helices or other hydrogen-bonded secondary structures. However, at low detergent concentrations, added salts had a marked effect, until the salt concentration reached o.I M for Na~S04 and 0.25 M for NaC1, after which additional salts had no further effect. The critical micelle concentration of sodium dodecyl sulfate at 25 ° in 0.25 M NaC1 and o.I M Na~SO 4 is I . I mM (ref. 22). This suggested that the effect of ionic strength is on the critical micelle concentration. REYNOLDS AND TANFORD 35 also pointed out that the effect of ionic strength appears to be entirely on the concentration of free monomer of sodium dodecyl sulfate, in other words, the critical micelle concentration of sodium dodecyl sulfate. However, they stated that micelles do not bind to the proteins they studied.
The importance of hydrophobic bonds in protein conformation has been em- phasized by many investigators (e.g. refs. 36-38) and the importance of hydrophobic environment on the formation and stability of the hydrogen-bonded a-helix is well known1°,37, 39. Hydrophobic bonds also played a decisive role in the interaction be- tween detergent and proteins (e.g. refs. 4o-43). Results obtained here (Table I) indicated that the interaction of anionic detergents with Fc(t) is indeed affected by the length of the hydrophobic tail of the detergents. This is consistent with previous results% l°. The order of effectiveness of the detergents as helix-forming agents is tetradecyl > dodecyl > decyl > octyl sodium sulfate. Our findings of the confor- mational transitions of Fc(t) incited by anionic detergents strongly supported the dominating importance of hydrophobic bonding in Fc(t). I t seems that the detergent molecules first reached a few sites on the protein surface. This interaction could acquire enough energy to replace the internal hydrophobic interaction with the external ones with detergent residues. As Fc(t) unfolds, more non-polar residues would be brought in contact with the detergent which eventually stabilizes the Fc(t) molecules in a different ordered structure. We have shown that this partial transition
Biochim. B,ophys. Acta, 229 (1971) 631-641

CD OF IgG FRAGMENT Fc(t) WITH ALKYL SULFATES 641
to a-helical structure depended on the amount of detergents present and may be affected by the presence of detergent micelles; it is also influenced by pH, ionic strength and especially by the hydrophobic chain length of the detergents.
ACKNOWLEDGMENTS
This research was supported in part by U.S. Public Health Service Research Grant No. CA-oI785, from the National Cancer Institute and by a grant from The Robert A. Welch Foundation G-osI, Houston, Texas, U.S.A.
P.K.J.L. is a Predoctoral Fellow of The Robert A. Welch Foundation, 1969-
197o.
R E F E R E N C E S
I F. W. PUTNAM, Advan. Protein Chem., 4 (1948) 79. 2 C. TANFORD, Advan. Protein Chem., 23 (1968) 121. 3 F. J. REITHEL, Advan. Protein Chem., 18 (1963) 123. 4 A. RAY, Chem. Soc. Special Rept., 23 (1969) 49. 5 V. W. PUTNAM AND H. ~'~EURATH, J . Am. Chem. Soc., 66 (1944) 692. 6 F. W. PUTNAM AND H. ~NTEURATH, J. Biol. Chem., 15o (1943) 263. 7 J. STEINHARDT AND J. A. REYNOLDS, Multiple Equil,brium in Proteins, Academic Press,
New York, I97 o, pp. 239-302. 8 B. JIRGENSONS, Arch. B~ochem. Biophys., 94 (196I) 59. 9 B. JIEGENSONS, J. Biol. Chem., 241 (1966) 4855.
IO B. JIRGENSONS, J. Biol. Chem., 242 (1967) 912. I I M. L. MEYER AND W. KAUZMANN, Arch. Biochem. Biopkys., 99 (1962) 348. 12 A. IMANISHI, Y. MOMOTANI AND T. ISEMURA, J. Biochem. Tokyo, 57 (1965) 417 • 13 C. C. BIGELOW AND M. SONENBERG, Biochemistry, I (1962) 197. 14 B. JIRGENSONS AND S. CAPETILLO, Bioch,m. Biophys. Acta, 214 (197 o) i . 15 Wor ld H e a l t h Organiza t ion , Bull. W.H.O., 3o (1964) 447. 16 G. M. EDELMAN, W. E. GALL, )¢[. J. WAXDAL AND W. H. KONIGSBERG, Biochemistry, 7 (1968)
195 ° • 17 E. DoI AND B. JIRGENSONS, Biochemistry, 9 (197 o) lO66. 18 H. A. SCHERAGA AND L. MANDELKERN, J. Am. Chem. Soc., 75 (1953) 179. 19 H. K. SCHACHMAN, Methods Enzymol., 4 (1957) 32. 20 G. D. FASMAN, Methods Enzymol., 6 (1963) 928. 21 G. HOLZWARTH AND P. DUTY, J. Am. Chem. Soc., 87 (I965) 218. 22 R. TOWNEND, T. F. •UMOSINSKI, S. N. TIMASHEFF, G. D. FASMAN AND B. DAVIDSON, BiD o
chem. Biophys. Res. Commun., 23 (1966) 163. 23 M. L. CORRIN AND W. D. HARKINS, J. Am. Chem. Soc., 69 (1947) 683. 24 J. POWNEY AND C. C. ADDISON, Trans. Faraday Soc., 33 (1937) 1243. 25 R. J. WILLIAMS, J. N. PHILLIPS AND K. J. MYSELS, Trans. Faraday Soc., 51 (1955) 728. 26 D. L. R o s s AND t3. JIRGENSONS, J. Biol. Chem., 243 (1968) 2829. 27 R. E. CATHOU, A. KULCZYCKI, JR., AND E. MASER, Biochemistry, 7 (1968) 3958. 28 E. ~. PYsH, Proc. Natl. Acad. Sci. U.S., 56 (1966) 825. 29 R. W. WOODY, Biopolymers, 8 (1969) 669. 3 ° M. L. TIFFANY AND S. KRIMM, Biopolymers, 8 (1969) 347. 31 D. G. DEARBORN AND D. B. WETLAUFER, Biochem. Biophys. Res. Commun., 39 (197 °) 314 • 32 B. JIRGENSONS, Biochim. Biophys. Acta, 2o0 (197 o) 9. 33 G. M. EDELMAN, B . A . CUNNINGHAM, W. E. GALL, P . n . GOTTLIEB, e . RUTISHAUSER AND
M. J. WAXDAL, Proc. Natl. Acad. Sci. U.S., 63 (1969) 78. 34 C. M. VENKATACHALAM AND G. N. RAMACHANDRAN, A~n. Rev. Biochem., 38 (1969) 45. 35 J- A. REYNOLDS AND C. TANFORD, Proc. Natl. Acad. Sci. U.S., 66 (197 o) lOO2. 36 W. KAUZMANN, Ado. Protein Chem., 14 (1959) I. 37 C. TANFORD, P. K. DE AND V. G. TAGGART, J. Am. Chem. Soc., 82 (196o) 6208. 38 H. A. SCHERAGA, in H. NEURATH, The Proteins, Vol. I, Academic Press, New York, 1963,
P. 477. 39 S. N. TIMASHEFF AND R. TOWNEND, Biochem. Biophys. Res. Commun., 20 (1965) 360. 4 ° P. JOHNSON AND F. J. JOUBERT, J. Polymer Sci., 7 (1951) 605. 41 V. POMERANZ, Enzymology, 27 (1964) 8. 42 R. V. DECKER AND J. F. FOSTER, Biochemistry, 5 (1966) 1242. 43 J. A. REYNOLDS, S. HERBERT, H. POLET AND J. STEINHARDT, Biochemistry, 6 (1967) 937.
Biochim. Biophys. Acta, 229 (1971) 631-641