conifer chemical defense: regulation of bark beetle - diva portal
TRANSCRIPT

Akademisk avhandling som med tillstånd av Kungliga Tekniska Högskolan i Stockholm framlägges till offentlig granskning för avläggande av doktorsexamen i kemi med inriktning mot organisk kemi Måndagen den 23 Maj 2011, kl 10.00 i sal F3, KTH, Lindstedtsvägen 26, Stockholm. Avhandlingen försvaras på engelska. Opponent är Irena Valterová, Institute of Organic Chemistry and Biochemistry, Academy of Sciences of the Czech Republic.
Conifer Chemical Defense: Regulation of Bark
Beetle Colonization and Pheromone Emission
Tao Zhao
Doctoral Thesis
Stockholm 2011

Cover: Spruce bark beetle Ips typrographus
(by Niklas Björklund)
ISBN 978-91-7415-884-7 ISSN 1654-1081 TRITA-CHE Report 2011:16 © Tao Zhao, 2011 E-Print, Stockholm

Tao Zhao, 2011. Conifer Chemical Defense: Regulation of Bark Beetle Colonization
and Pheromone Emission. Royal Institute of Technology, Sweden Abstract
Terpenes and phenols are of importance in conifer defense against
insects and pathogens. Knowledge about tree chemical defense is vital for
developing practical methods to maintain healthy forests. With the aims of
characterizing the defensive chemical induction in Norway spruce Picea abies
and demonstrating its ecological function to spruce bark beetle Ips
typographus, we measured the terpenoid and phenolic content in the bark of
mature Norway spruce trees suffering windstorm, inoculated with
Ceratocystis polonica, or treated with methyl jasmonate (MeJA), and
investigated the colonization and pheromone emission of I. typographus.
All three stressors altered the chemical profile in the bark of Norway
spruce. Trees damaged by windstorm had lower proportions of (+)-3-carene
and two unidentified stilbenes, and a higher taxifolin glycoside content than
trees without apparent windstorm damage; C. polonica inoculation induced
extremely strong quantitative terpene accumulation in the wound reaction
zone, but only increased the levels of (+)-3-carene, sabinene and terpinolene
in the bark near the reaction zone; MeJA treatment generally elicited
quantitative terpene accumulation, but the induction differed extensively
between individual trees. In addition, logs from MeJA-treated tree showed
much stronger wounding response compared to control logs.
The chemical profile of Norway spruce affected the colonization and
pheromone emission of I. typographus. In response to fungal inoculation,
terpene present in the reaction zone inhibited the colonization of I.
typographus in a dose-dependent manner. Host defense elicited by MeJA
treatment reduced emissions of 2-methyl-3-buten-2-ol and (S)-cis-verbenol,
the two key aggregation pheromone components of I. typographus, and
altered the ratio between the two components.
Keywords: Picea abies, Ips typographus, Ceratocystis polonica, methyl jasmonate,
windstorm, stress response, terpenes, phenols, colonization, pheromone
emission.


List of Publications
This thesis is based on the following papers, referred to in the text by their
Roman numerals:
I. Storm-induced tree resistance and chemical differences in Norway spruce
(Picea abies)
Zhao T., Solheim H., Långström B., and Borg-Karlson A.-K.
Annals of Forest Science, 2011
II. The influence of Ceratocystis polonica inoculation and methyl jasmonate
application on terpene chemistry of Norway spruce, Picea abies
Zhao T., Krokene P., Björklund N., Långström B., Solheim H., Christiansen
E., and Borg-Karlson A.-K.
Phytochemistry, 2010, 71, 1332-1341
III. Induced terpene accumulation in Norway spruce inhibits bark beetle
colonization in a dose dependent manner
Zhao T., Krokene P., Hu J., Christiansen E., Björklund N., Långström B.,
Solheim H., and Borg-Karlson A.-K. Submitted
IV. Host resistance elicited by methyl jasmonate reduces emission of aggregation
pheromones by the spruce bark beetle, Ips typographus
Zhao T., Borg-Karlson A.-K., Erbilgin N., and Krokene P.
Oecologia, 2011, in press
Papers not included in this thesis:
V. Odours of Norway spruce (Picea abies) seedlings: differences due to age and
chemotype and their relevance for weevil resistance
Kännaste A., Zhao T., Lindström A., Stattin E., Borg-Karlson A.-K., and
Långström B. Manuscript
VI. Volatile emissions of pine seedlings, age related changes and chemotypes
Kännaste A., Zhao T., Lindström A., Stattin E., Borg-Karlson A.-K., and
Långström B. Manuscript for Phytochemistry
VII. Chemical and transcriptional responses of Norway spruce clones with
different susceptibility to Heterobasidion spp. infection
Danielsson M., Lundén K., Arnerup J., Hu J., Zhao T., Swedjemark G.,
Elfstrand M., Borg-Karlson A.-K., and Stenlid J. Manuscript

Abbreviations
2D-GC Two-dimensional gas chromatography
ANOVA Analysis of variance
AO Antioxidant
Cp Ceratocystis polonica
CT Control
cV (S)-cis-Verbenol
DT Diterpene
DVB
DW
Divinylbenzene
Dry weight
GC Gas chromatography
HPLC High pressure liquid chromatography
IS Internal standard
MB 2-Methyl-3-buten-2-ol
MeJA Methyl jasmonate
MEP Methyl-D-erythritol phosphate
MS Mass spectrometry
MT Monoterpene
MVA Mevalonate
PC Principal component
PCA Principal component analysis
PDMS Polydimethylsiloxane
PP cell Polyphenolic parenchyma cell
RTD Traumatic resin duct
SD Standard deviation
SE Standard error
SPME Solid phase microextraction
ST Sesquiterpene
TPS Terpene synthase
TPS-Bis E-α-Bisabolene synthase
TPS-Car (+)-3-Carene synthase
TPS-Far (E,E)-α-Farnesene synthase
TPS-Iso Isopimaradiene synthase
TPS-LAS Levopimaradiene/abietadiene synthase
TPS-Lim (-)-Limonene synthase
TPS-Lon Longifolene synthase
TPS-Pin (-)-α/β-pinene synthase
UV Ultraviolet

Table of Contents
1. Introduction .................................................................................................... 1
1.1. Aims ........................................................................................................ 1
1.2. Background ............................................................................................. 1
1.3. Bark beetles and their associated fungi .................................................... 2
1.4. Conifer resistance .................................................................................... 4
1.5. Terpenoid and phenolic compounds in conifers ...................................... 5
1.5.1. Terpenoid compounds and their biosynthetic pathway .................... 5
1.5.2. Phenolic compounds ........................................................................ 9
1.5.3. Defensive roles of phytochemicals against bark beetles .................... 9
2. Materials and Methods ................................................................................ 11
2.1. Biological materials and procedures ...................................................... 11
2.1.1. Plant material and experimental site .............................................. 11
2.1.2. Impacts of windstorm in standing trees .......................................... 11
2.1.3. MeJA application and C. polonica inoculation ................................ 12
2.1.4. Impacts of MeJA treatment on beetle pheromone emission .......... 13
2.2. Sample preparation ................................................................................ 14
2.2.1. Phloem sample extraction ............................................................... 14
2.2.2. Collection of volatiles released from beetle entrance holes ............ 15
2.3. Sample analysis ...................................................................................... 16
2.3.1. Gas chromatography-mass spectrometry ....................................... 16
2.3.2. Two dimensional gas chromatography ........................................... 16
2.3.3. Liquid chromatography ................................................................. 16
2.3.4. Terpene and phenolic identification and quantification ................. 17
2.4. Data analyses ......................................................................................... 17
3.Effect of Windstorm on Tree Resistance and Chemical Profile ................... 19
3.1. Tree resistance to fungal inoculation ..................................................... 19
3.2. Terpene and phenolic composition ........................................................ 22
3.3. Correlation between resistance and chemical composition ................... 23
4. Fungal Inoculation and MeJA Application Induced Terpene Responses ..... 25
4.1. C. polonica inoculation induced terpene responses ................................. 26
4.1.1. Terpene response in reaction zone ................................................. 26

4.1.2. Terpene responses near reaction zone ............................................ 28
4.2. MeJA treatment induced terpene responses .......................................... 30
4.3. Comparison between MeJA and C. polonica induced terpene response 33
4.4. MeJA and C. polonica induced terpene response lasted one year ........... 33
4.5. Does Norway spruce process systemic terpene induction? .................... 35
5. Terpene Accumulation Reduced Bark Beetle Colonization ......................... 37
5.1. Variation in terpene response and beetle colonization .......................... 38
5.2. Terpene accumulation in trees with different susceptibility .................. 38
5.3. Terpene accumulation inhibited I. typographus colonization in a dose-
dependent manner ....................................................................................... 40
6. Host Resistance Reduced Bark Beetle Pheromone Emission ...................... 43
6.1. MeJA treatment elicited terpene defense in Norway spruce................. 44
6.2. The performance of I. typographus in MeJA treated logs ...................... 46
6.3. MeJA treatment altered pheromone quantity and quality .................... 46
6.4. MeJA treatment altered host volatiles emitted from entrance holes ..... 49
7. Concluding Remarks ....................................................................................... 51
8. Acknowledgment ............................................................................................. 53
9. References ........................................................................................................ 55
Author’s Contributions .................................................................................... 71
Appendix B1 .................................................................................................... 72
Appendix B2................................................................................................... 73
Appendix C..................................................................................................... 74
Appendix D..................................................................................................... 75

1
1. Introduction
1.1. Aims
• To characterize the chemical defense in Norway spruce elicited by
abiotic and biotic stressors
• To understand the role of chemical defense in bark beetle colonization
and pheromone production
• To find out the possible chemical markers related to conifer resistance
or susceptibility to bark beetle-fungus complex
• To extend our general knowledge on conifer-bark beetle interactions.
Figure1. Scheme of the thesis.
1.2. Background
Conifers are the dominant plants in boreal forests. Providing about 45
percent of the world’s annual timber production (FAO 2006), conifers are
of great economical importance in these areas. In Sweden, forest products
deliver half of the net national income. According to The Swedish National
Chemical defense (Papers I-IV)
Norway spruce
Associated fungus (Paper I and II)
Beetle colonization (Papers II-IV)
Beetle pheromone emission
(Paper IV)
Windstorm (Paper I)
Fungus, MeJA (Papers II-IV)

2
Forest Inventory, Norway spruce (Picea abies) and Scots pine (Pinus
silvestris), the two major conifer species in the country, each makes up of
~40 % of its forest growing stock.
However, conifers are often threatened by pests such as bark beetles
and their associated fungi (Christiansen and Bakke 1988; Kurz et al. 2008;
Raffa et al. 2008). In Scandinavia spruce bark beetle Ips typographus have
killed more than 50 million m3 of Norway spruce trees in several outbreaks
since the late 1940’s (Christensen and Bakke 1988; Wermelinger 2004;
Worrell 1983). In North America, the outbreak of the mountain pine beetle
Dendroctonus ponderosae in British Columbia between the mid 1990s and
2006 killed ca. 10 million hectares of pine trees (Kurz et al. 2008). Global
climate change alters the physiological condition of the trees, facilitates
insect establishment in new areas, and accelerates the development of bark
beetle, increasing the risk of bark beetle outbreak (Bidart-Bouzat and
Imeh-Nathaniel 2008; Jonsson et al. 2009; Kurz et al. 2008).
Outbreaks of bark beetles not only result in great economic losses,
but also disturb ecosystem dynamics, elevate volatile (Bidart-Bouzat and
Imeh-Nathaniel 2008; Laothawornkitkul et al. 2009; Tao and Jain 2005)
and CO2 emissions (Kurz et al. 2008), and thus generate considerable
impacts on the forest ecosystem and global environment. So far there is no
environmentally friendly and economically acceptable way to control these
pests. A deeper understanding of conifer
defense mechanisms and functions is clearly
needed in order to develop practical methods
to maintain healthy forests.
1.3. Bark beetles and their associated fungi
Bark beetles (Coleoptera: Scolytinae) include
more than 6000 species from 225 genera
(Knizek and Beaver 2004). These insects
generally reproduce and develop in recently
killed trees, but some aggressive species are
Figure 2. Norway spruce killed by spruce bark beetle, Ips typographus (by Paal Krokene).

3
capable of attacking, mating and laying eggs in living trees. After hatching,
the young larvae feed in the inner bark, forming extensive larval galleries
there. The successful reproduction of aggressive bark beetles destroys the
tissues transporting nutrition and water, and finally kills the tree (Figure 2).
The subcortical tissue of a living conifer tree is rich in nutrition, but
it is also full of various defensive compounds and structures (Franceschi et
al. 2005). To successfully reproduce in this tissue, bark beetles have evolved
several strategies to overcome tree defense. The important strategies
include cooperative mass attack behavior directed by aggregation
pheromones and the ability to vector phytopathogenic fungi (Raffa et al.
1993).
Bark beetles colonizing living conifers are often associated with
phytopathogenic fungi. When bark beetles construct mating chambers and
galleries in the phloem, they introduce fungi to the tree. These fungi invade
the tree’s vascular system and produce toxins, helping beetles to deplete tree
resistance (Klepzig et al. 2009; Lieutier et al. 2009; Paine et al. 1997).
Therefore, the associated fungi are thought to play an important role in the
successful colonization of bark beetles (Lieutier et al. 2009; Paine et al.
1997; Six and Wingfield 2011). Infection by these fungi often discolors the
sapwood to blue-grey. For this reason bark beetle associated fungi are
commonly called bluestain fungi.
Besides the fungal associates, many aggressive bark beetle species use
oxygenated hemi- and mono- terpenoids as aggregation pheromones to
coordinate mass attacks (i.e. gather conspecifics to attack the tree
simultaneously) and thus collectively exhaust the resistance of a living tree
(Raffa and Berryman 1983b). The pheromones are mainly produced de
novo through the mevalonate pathway in the midgut cell of the bark beetle
(Blomquist et al. 2010; Ivarsson et al. 1997; Seybold et al. 2000), or
detoxified from the terpenes ingested from the host tissue (Lindström et al.
1989). In addition to the aggregation pheromone, bark beetles also use
anti-aggregation pheromones to regulate population density and reduce
competition (Byers 1993; Miller et al. 1995). The pheromone system thus

4
regulates the host colonization, mating and reproduction of these bark
beetles.
1.4. Conifer resistance
Under selective pressure from bark beetles and other pests, conifers have
evolved multiple constitutive and inducible resistance mechanisms
(Franceschi et al. 2005; Keeling and Bohlmann 2006). The constitutive
defense, mainly including chemicals (e.g. terpenoids and phenolics) and the
anatomical structures regularly existing in the tree, forms the first line of
defense against the potential invaders by combining chemical toxicity with
physical deterrence.
After the beetles bore through the bark the inducible anatomic and
biochemical defenses are triggered quickly around each attack point
(Franceschi et al. 2005; Keeling and Bohlmann 2006). Traumatic resin
ducts (RTD) are formed de novo, polyphenolic parenchyma cells (PP cells)
are activated and mass defensive compounds are produced. The induced
resistance is usually elicited locally, but systemic induced resistance is also
observed in tissue far away from the attack point (Bonello et al. 2008; Eyles
et al. 2010), which may prepare the tissue for potential challenges. In
addition to the response within the tree, conifers also release large amounts
of airborne volatiles upon stress ( Jost et al. 2008; Martin et al. 2003). These
volatiles may interfere with the host selection of bark beetles and their
predators and parasitoids, indirectly influencing the resistance of the trees
(Boone et al. 2008; Erbilgin and Raffa 2000; Wallin and Raffa 2000a).
Collectively, these mutually overlapping defenses may stop beetle
invasion, impede fungal growth and seal the wounded tissues (Franceschi et
al. 2005; Keeling and Bohlmann 2006). However, induced defenses are
thought to be more important in conifer resistance against bark beetle–
fungus complex than constitutive ones (Eyles et al. 2010; Raffa and Smalley
1995).
Plant defense responses are governed by complex signaling networks
(Figure 3), which are regulated by phytohormones. Jasmonic acid ( JA),

5
salicylic acid (SA) and ethylene are the major phytohormones mediating
plant defense against abiotic stresses, pathogens and herbivores (Kunkel
and Brooks 2002). It is generally suggested that the JA-mediated metabolic
biosynthetic pathway is associated with the response to feeding by
herbivores and the SA-mediated pathway is typically activated by
pathogens (Smith et al. 2009). However, since chemical defense is
energetically costly, plants are thought to employ cross-talk networks to
minimize the cost associated with defense (Koornneef and Pieterse 2008;
Kunkel and Brooks 2002; Smith et al. 2009).
Figure 3. Simplified signal transduction pathway in conifer.
In conifers, Hudgins and Franceschi (2004) found that the formation
of traumatic resin ducts was induced via ethylene signaling in Douglas fir
(Pseudotsuga menziesii) and giant redwood (Sequoiadendron giganteum) after
methyl jasmonate (MeJA, for structure see chapter 4) application. Ralph et
al. (2006) demonstrated that insect herbivory or mechanical wounding in
Sitka spruce (Picea sitchensis) affects the genes involved in the octadecanoid
and ethylene pathways. How the signaling compounds regulate the
expression of defense genes is not clear.
1.5. Terpenoid and phenolic compounds in conifers
1.5.1. Terpenoid compounds and their biosynthetic pathway
As the largest class of natural products, terpenes include approximately
30000 known structures (Gershenzon and Dudareva 2007; Keeling and
Bohlmann 2006). In nature terpenes are exploited by both plants and
Stress elicitor
Octadecanoid pathway
Shikimic acid pathway
Jasmonic acid
Receptors/Sensores
Gene activationInduced defenses
Ethylene pathway
Salicylic acid
Ethylene

6
insects, and play a variety of roles in plant-insect interactions (Gershenzon
and Dudareva 2007; Reddemann and Schopf 1996; Seybold et al. 2006).
Conifer oleoresin is a complex mixture of mono-, sesqui- and
diterpenes (Keeling and Bohlmann 2006; Phillips and Croteau 1999), of
which mono- and diterpenes account for more than 95% of the total
constitutive terpenes in the stem of Norway spruce (Martin et al. 2002).
Figure 4 shows the structures of the major terpenes studied in this thesis.
Figure 4. Structure of the major terpenes in the phloem of Norway spruce (Picea abies). A. mono-, B. sesqui-, and C. diterpenes.
Terpenes are synthesized by terpene synthases (Bohlmann and
Keeling 2008; Chappell 1995; Fäldt et al. 2003; Keeling and Bohlmann
2006; Martin et al. 2004; Ro et al. 2006) (Figure 5). The methyl-D-

7
erythritol phosphate (MEP) pathway in plastids and melvanoate (MVA)
pathway in cytosol (Bochar et al. 1999) supply isopentenyl diphosphate
(IPP) and dimethylallyl diphosphate (DMAPP), common substrates for all
the terpenes. These substrates are then condensed to form the precursors of
monoterpenes (geranyl diphosphate), sesquiterpenes (farnesyl diphosphate)
and diterpenes (geranylgeranyl diphosphate). The terpene synthases (TPS)
further transform the precursors to diverse terpenes (Chappell 1995).
So far nine TPSs have been characterized in Norway spruce (Fäldt et
al. 2003; Martin et al. 2004). Two sesqi-TPSs and one di-TPS only produce
a single terpene, while one sesqui-TPS, one di-TPS and all of four mono-
TPSs produce multiple terpenes in different proportions (Figure 6). For
example, TPS-Lim produces 87.7 % (-)-limonene, 5.2 % myrcene, 4.4 %
(-)-α-pinene and 2.1 % (+)-limonene (Martin et al. 2004); TPS-pin
produces 57.1 % (-)-β-pinene, 27.4 % (-)-α-pinene, 11.0 % (-)-β-
phellandrene and 2.1 % myrcene (Martin et al. 2004).
Zulak et al. (2009) investigated the activity and induction of TPS
enzyme in Norway spruce treated with MeJA on multiple levels. They
found a good correlation between induction of transcript, proteins and
enzyme activities. Most of the mono- and diterpene accumulation were in
similar temporal patterns to molecular induction. This result confirmed
that defensive terpene production is regulated by multiple steps in conifers.
The formation of IPP and DMAPP through the MEP pathway is
catalyzed by DXS (1-deoxy-D-xylulose 5-phosphate synthase) (Lange et al.
1998). In cell cultures, one subtype of DXS (PaDXS2A) was induced by
treatment with methyl salicylate and C. polonica, and another subtype
(PaDXS2B) was induced by MeJA treatment, suggesting that MeJA and C.
polonica inoculation may induce different responses, at least in the early
stage of terpene synthesis (Phillips et al. 2007). How these differences
influence the terpene metabolites are still unknown.

8
Figu
re 5
. M
ain
path
way
s of
terp
ene
bios
ynth
esis
in N
orw
ay s
pruc
e (P
icea
abi
es).
ME
P: m
ethy
l-D-e
ryth
ritol
pho
spha
te; M
VA:
mev
alon
ate;
IPP:
isop
ente
nyl d
ipho
spha
te;
DM
AP
P: d
imet
hyla
llyl d
ipho
spha
te; G
PP
: ger
anyl
dis
phos
phat
e; F
PP: f
arne
syl d
ipho
spha
te; G
GP
P: g
eran
ylge
rany
l dip
hosp
hate
; TPS
: ter
pene
syn
thas
es (
mod
ified
from
Zu
lak
et a
l. 20
09).

9
1.5.2. Phenolic compounds
In addition to terpenoids, phenols such as stilbenes and flavonoids have
also been implicated in conifer resistance against bark beetles and their
associated fungi (Faccoli and Schlyter 2007; Lieutier 2004; Ralph et al.
2006). Plant phenols include ca. 4000 structurally different compounds
which are biosynthesized by different pathways, but the shikimic acid
pathway is the major pathway in conifers (Ralph et al. 2006). Figure 6
shows the structures of major stilbenes and flavonoids in Paper I.
Figure 6. Structures of major phenols in the phloem of Norway spruce (Picea abies).
1.5.3. Defensive roles of phytochemicals against bark beetles
Because oleoresin entraps and impedes invaders, the resin system in
conifers has long been considered as the major component of tree resistance
against insects (Webb 1906). Later on lab assays suggested that oleoresin is

10
toxic or repellant to bark beetles and their associated fungi. For terpenes,
Smith (1965) found that saturated monoterpene vapor is toxic to bark
beetles; Solheim (1991) reported that induced resin inhibited the growth of
beetle associated fungi on malt agar; Raffa and Smalley (1995)
demonstrated that phloem powder containing high concentrations of
synthetic monoterpenes led to considerable beetle mortality. For phenols,
Faccoli et al. (2007) demonstrated catechin, taxifolin and resveratrol are
antifeedants to I. typographus; Salle et al. (2005) reported that catechin and
taxifolin inhibited the growth of Ophiostoma piceaperdum and O. bicolor
(fungi associated with I. typographus) on malt-agar medium.
In addition to the direct contribution, conifer terpenes may also
indirectly affect a tree’s resistance by interfering with the attraction of bark
beetle to its pheromone sources (Seybold et al. 2006), or by influencing its
pheromone synthesis. For example, host monoterpenes, such as α-pinene,
are attractive to many bark beetles (Gallego et al. 2008; Miller and Rabaglia
2009); but also enhances or inhibits the attraction of bark beetle to its
aggregation pheromone, depending on the concentrations (Erbilgin et al.
2007; Erbilgin et al. 2003; Miller and Borden 2000; Wallin and Raffa
2000b). There is also evidence that the ratio of cis/trans-verbenol in the gut
of Ips bark beetles is correlated with the enantiomeric composition of
α-pinene in host tissue (Klimetzek and Francke 1980; Lindström et al.
1989).

11
2. Materials and Methods
2.1. Biological materials and procedures
2.1.1. Plant material and experimental site
The chemical compositions and defense responses of conifers upon
invasion depend on the age, genetic background and physiological
condition of the trees (Danielsson et al. 2008; Laracinepittet and Lebreton
1988; Zeneli et al. 2006). I. typographus, in particular, colonizes mature
Norway spruce, and prefers trees which have been exposed to certain
physiological stress. It can only attack healthy trees when its population
reaches epidemic levels.
To obtain a general pattern of induced chemical responses in Norway
spruce, and relate them with tree resistance against I. typographus, it is
necessary to choose genetically and physiologically different mature trees as
experimental materials with which to screen different chemical responses,
and an experimental site with a high bark beetle population density in
order to investigate how the chemical defenses affect the colonization of I.
typographus.
In Papers I-III, we used naturally regenerated Norway spruce trees
growing at Tönnersjöheden Experimental Forest (56°41' N, 13°4' E,
Sweden) as plant material. This area was damaged by the “Gudrun” wind-
storm in January 2005 and I. typographus populations have erupted since
2006 (Långström et al. 2009). The high beetle population provided a good
opportunity to test the ecological function of chemical defenses in the P.
abies – I. typographus system.
2.1.2. Impacts of windstorm in standing trees
To evaluate the impacts of windstorm on the chemistry and resistance of
standing trees, we selected 10 of each Norway spruce with different
physiological conditions (i.e. leaning-stems, trees with short-leader-shoot,
and normal-growing trees with no apparent damage, representing severe,

12
visible and no storm-damage, respectively) at a storm damaged stand edge
in June 2006, 17 months after the “Gudrun” storm.
We took one stem bark sample from each experimental tree for
terpene and phenolic analysis, and then inoculated the stem of half the
experimental trees with C. polonica to assess their resistance levels. The
fungal performances in the trees were investigated 11 weeks after fungal
inoculation (Paper I).
2.1.3. MeJA application and C. polonica inoculation
Figure 7. Diagram shows the experimental design and field procedures. Cp: C. polonica; CT:control; Day 0 represents the day of treatment. Day 14, Day 30, Day 35 and Day 362 represent 14, 30, 35, and 362 days after treatment.
In May 2007, 24 triplets of neighboring trees of similar size were randomly
assigned to either C. polonica inoculation (Cp), MeJA application, or used as
untreated control (CT). Two bark samples, one from the treated (ca. 1.3 m
stem height) and another from the untreated stem sections (ca. 0.5 m stem
height, 30 cm below the treated section), were taken from each tree three
times: on the day of treatment, 30 days, and one year after treatment
(Figure 7). Samples from fungal-inoculated trees (in the treated stem
sections) were taken 5 cm above the inoculation holes, which is near (ca. 1-
3 cm above) the reaction zone (Figure 8). A pheromone dispenser tape
(Hercon® type) was attached to the stem of each tree to attract bark
beetles, and a sticky trap (Pherobank®, 10 × 15 cm) was used to monitor
Vaccination project
2007 experiment 2008 experiment
72 spruce trees 60 spruce trees
CT MeJA Cp inoculation CT
Samples: Day 0, Day 30, Day 362 Attack density, gallery length, landing rate
Samples: Day 14, Day 35
Paper II Paper III
MeJA Cp inoculation

13
the landing rate of bark beetles on each tree one month after the treatment.
The number of beetles landing on the sticky trap was recorded regularly.
The trees were inspected for beetle colonization at the end of the season
(Figure 7) (Paper II).
Because we started the experiment late in the season, we didn’t
receive the expected amount of beetle colonization in 2007. In April 2008,
we repeated the experiment with a new set of 60 trees in the same stand
(Figure 7). Two bark samples (one from the treated section and another
from the untreated section) were taken from each tree 14 days and 35 days
after treatment respectively. For fungal-inoculated trees, the samples from
the treated stem sections were taken immediately above the inoculation
holes (Figure 8). A sticky trap was attached to the stem of each tree to
monitor beetle landing, and a pheromone dispenser was placed on a pole
standing ca. 40 cm from southwest side of each tree to attract bark beetles,
one month after the treatment. The landing rate and colonization of bark
beetle in each tree were inspected in the same way as 2007 (Paper III).
Paper II Paper III
: Inoculation hole; : Sample for terpene analyses
Figure 8. Scheme shows the different sampling positions in the trees inoculated
with Ceratocystis polinica in 2007 and 2008 experiments.
2.1.4. Impacts of MeJA treatment on beetle pheromone emission Two 48-year-old clonal trees within the same Norway spruce clone, one
treated with MeJA and one left as a control, were felled in early June 2009
(Paper IV). In the lab, adults I. typographus were introduced to the logs to
initiate attacks, and solid phase microextraction (SPME) was employed to
adsorb pheromone components and host volatiles emitted from the
2007
5cm
2008

14
entrance holes. The intact and wounded bark samples were taken from the
experimental logs to elucidate the chemical defense in Norway spruce after
MeJA treatment. The number of maternal galleries with or without a
mating chamber was recorded and the gallery length was measured two
months later (Figure 9).
Figure 9. Diagram shows the experimental procedures.
2.2. Sample preparation 2.2.1. Phloem sample extraction
Plant materials are often extracted with a series of solvents, usually
increasing in polarity, to selectively dissolve compounds with different
properties (Millar and Haynes 1998). Plant samples are often freeze-dried
and ground to powder for efficient extraction (Millar and Haynes 1998).
However, the sample pretreatment procedures are time consuming, and
more importantly, they increase the risk of contamination, chemical
changes and loss of highly volatile compounds during the process. In
addition, adverse changes may also occur in the plant material during
transportation if the sampling site is far from the lab.
Most of the bark samples investigated in this thesis were taken from
the forest, ca. 400 km away from the lab. For this reason we adopted a
simple sample extractive method, minimizing cell destruction and suitable
for handling a large number of samples in the forest. Hexane, a nonpolar
solvent which can effectively extract non-polar terpenoid compounds
(Fäldt et al. 2006; Persson et al. 1993), was used to extract terpenes (Papers
I-IV). The fresh phloem plugs were taken from living Norway spruce
stems and submerged in hexane containing pentadecane (Lancaster
synthesis, English) as internal standard and 3-tert-butyl-4-hydroxyanisole
MeJA treated logs
Untreated logs
Introduce adult bark beetles
Entrance holes of bark beetle
Intact and wounded bark Hexane extracts GC-MS SPME extraction
Beetle performance

15
(Fluka, Switzerland) as antioxidant, immediately after sampling. After
hexane extraction, the phloem plugs were extracted again by 80 % methanol
for phenolic compounds (Paper I). Similar phenolic extractive methods
have been used in several other studies (Bois and Lieutier 1997; Brignolas
et al. 1995a; Lieutier et al. 2003).
2.2.2. Collection of volatiles released from beetle entrance holes
SPME is a simple and versatile technique for sampling volatile organic
compounds in vivo (Bartelt 1997; Vas and Vekey 2004). With the
advantage of being non-destructive and free of solvent, SPME has been
widely used to collect monoterpene, sesquiterpene and other volatile
compounds released by living organisms (Baker and Sinnott 2009; Fäldt et
al. 2000; Rocha et al. 2001; Rochat et al. 2000; Tholl et al. 2006; Vas and
Vekey 2004; Yassaa and Williams 2007).
Different types of SPME fibers are commercially available, of which
65 μm polydimethylsiloxane/divinylbenzene (PDMS/DVB) SPME fiber
(Supecol, PA, USA) shows a good adsorptive capacity to a wide range of
small and medium sized molecules. For this reason
it has been commonly used to collect plant
volatiles (Bouvier-Brown et al. 2007; Yassaa and
Williams 2005; 2007) and insect pheromone
emissions (Rochat et al. 2000; Villaverde et al.
2007) for the purposes of identification and
quantification. In Paper IV we used this fiber to
collect aggregation pheromone released by bark
beetle tunneling in untreated logs and logs treated
with MeJA, using the setup shown in Figure 10.
The extraction was done at room temperature, and
extraction time was 120 min.
The calibrations of the aggregation
pheromone components (2-methyl-3-buten-2-ol
and (S)-cis-verbenol) were done by comparison with response curves
derived from the collection of volatiles from synthetic references in
Figure 10. SPME setup for beetle pheromone collection and calibration.

16
identical conditions (Paper IV). To reduce the possible impact of
competitive effects (of host volatiles or water evaporated from the entrance
hole) on quantification, we tried to mimic the true sampling condition by
placing the reference chemicals in the predrilled holes in fresh Norway
spruce logs during calibration (Paper IV).
2.3. Sample analysis
2.3.1. Gas chromatography-mass spectrometry (GC-MS)
The volatile, semi-volatile and non-volatile terpenes such as mono-, sesqui-
and non-polar diterpenes dissolved in hexane (Papers I-IV) or extracted by
SPME (Paper IV) were analyzed by a Varian GC equipped with an SPB-1
(Supelco, 30 m × 0.25 mm × 0.25 μm, a DB-wax ( J&W Scientific, 30 m ×
0.25 mm × 0.25 μm) or a CP-Chirasil-DEX CB (Varian, 25 m × 0.25 mm
× 0.25 μm) column. Different temperature programs were used to achieve
optimal separation. A Finnigan SSQ 7000 MS was used to identify and
quantify the individual terpenes.
2.3.2. Two dimensional gas chromatography (2D-GC)
A 2D-GC system consisting of two Varian 3400 GCs (Borg-Karlson et al.
1993)(Papers I-III) was employed to separate and quantify enantiomers of
chiral monoterpenes of interest. In the 2D-GC, the first GC was equipped
with a DB-wax column for pre-separation, and the second GC with a β-
cyclodextrin column ( J&W Scientific, 30 m × 0.25 mm × 0.25 μm) for the
chiral separation.
2.3.3. Liquid chromatography (LC)
An HP 1090 high performance liquid chromatography (HPLC)
instrument fitted with an ultraviolet/visible diode array detector and a
Discovery® C18 HPLC column (Supelco, 25 cm × 4.6 mm, 5 μm particle
size) was used to separate the phenolic components dissolved in 80 %
methanol. Similar step gradient program and ultraviolet (UV) wave length
(280 nm) were employed for separation and quantification, to make the

17
results comparable with previous studies (Brignolas et al. 1995a; 1998;
1995b) (Paper I).
2.3.4. Terpene and phenolic identification and quantification
The terpene hydrocarbons were identified by comparing retention times
and mass spectra with available authenticated standards, or by comparing
Kovats retention indexes (RIs) and mass spectra with Massfinder 3
(Hochmuth Scientific Consulting, Germany) and the reference libraries of
NIST (National Institute of Standards and Technology) (Papers I-IV). The
phenolic compounds were identified by comparing the retention time and
UV spectrum with authenticated standards (Paper I).
The absolute amounts of terpenes were calculated relative to the
internal standard, and expressed as mg (or μg) g-1 dry weight (wt) relative
to the mass of pentadecane (Papers II-IV). The relative amounts of
terpenes (Paper I, II and IV) and phenols (Paper I) were calculated as the
ratio of the area of each peak to the sum of all the areas of terpenes or
phenolic hydrocarbons in a defined GC or LC fraction.
2.4. Data analyses
Both GC and LC separate more than thirty compounds in each analysis. In
order to determine the overall chemical changes after treatments/storm,
and to find out the potential compounds for further investigation, it is
necessary to include all the compounds in the data analyses (Pettersson
2007). To achieve this, principal component analysis (PCA) was used to
evaluate the overall chemical differences of experimental trees (Paper I, II
and IV). This method estimates the similarity of the samples based on all
the quantified compounds. The samples with similar chemical profiles are
close to each other in the PCA score plot. The axis of the plot, principal
component (PC), indicates how much of the sample variation is explained
in percent by this principal component. The multivariate data analysis
software Canoco 4.5 (Biometris Plant Research International, The
Netherlands) was used for PCA.

18
The changes of individual compounds between treatments or
sampling times were compared by various parametric (t-test, ANOVA) or
non-parametric (Mann-Whitney U test, Fisher’s exact test) methods
included in Statistica 6.0 (Statsoft, Inc. USA), based on the property of the
data set and the purpose of data analysis. Data transformation was adopted
to correct for unequal variance and departures from normality before
parametric testing. The relative amounts of terpenes were arcsin-
transformed and the absolute amount data were square-root or logarithm
transformed. The correlation between chemicals (Paper II) or between
chemicals and fungal performance (Paper I) was analyzed by Pearson-
product-moment correlation.

19
3.Effect of Windstorm on Tree Resistance and Chemical Profile
Paper I
Windstorm is one of the major threats for forest management in boreal
forests (Nilsson et al. 2004). Storms not only directly fell the trees, but also
disturb the ecosystem triggering further damage to the forest in subsequent
years (Everham and Brokaw 1996; Glitzenstein and Harcombe 1988). In
Europe, windstorm leads to more than 50 % of the all forest damage
(Gardiner et al. 2010).
Windstorm in Europe is often followed by bark beetle outbreak
(Komonen et al. 2011). In January 2005, the "Gudrun" storm felled ca. 75
million m3 of mostly Norway spruce forest in southern Sweden, and
resulted in an outbreak of I. typographus in the area in the following years.
Ca. 3 million m3 of Norway spruce trees were killed by the beetles between
2006-2008 (Långström et al. 2009).
The outbreak of bark beetles after a windstorm is largely because the
plentiful wind-fall provides abundant breeding material (Bouget and Duelli
2004; Schroeder 2007). In addition, the bending force of windstorm may
damage the roots and other parts of the trees, impair their water uptake and
photosynthesis (Fredericksen et al. 1995; Nielsen and Knudsen 2004), and
finally render the trees more susceptible to beetle attack. However, little is
known about the physiological and chemical changes in storm-damaged
trees and how these may affect tree susceptibility to fungi and insects.
Studies are clearly needed to understand the influence of windstorm on
living trees for sustainable forest development.
3.1. Tree resistance to fungal inoculation
The resistance of the experimental trees was evaluated by means of artificial
C. polonica inoculation. Eleven weeks after fungal inoculation the overall
proportions of dead cambium, occluded sapwood and bluestained sapwood

20
in leaning-stem and short-leader trees were significantly lower than the
seemingly undamaged trees, although the phloem necrosis lengths did not
vary extensively between the three tree categories (Figure 11).
Proportions of dead cambium, occluded sapwood, and bluestained
sapwood have been regularly used to evaluate tree resistance (Christiansen
et al. 1999; Krokene et al. 2001; Sandnes and Solheim 2002; Zeneli et al.
2006). After fungal inoculation the trees with relatively high resistance
usually generate lower proportions of dead cambium, occluded sapwood
and bluestained sapwood than trees with relatively low resistance, because
they have a greater ability to restrain fungal infection. Our result, contrary
to expectation, clearly indicated that short-leader and leaning-stem trees
had higher resistance to C. polonica than seemingly undamaged trees.
By simulating wind stress (bending stems and pruning the
branches), Fredericksen et al. (1995) studied the possible impacts of wind
on loblolly pine (Pinus taeda) resistance against Dendroctonus and Ips bark
beetles. Although the treatment reduced the radial growth of experimental
trees by 67 %, the treated trees did not show more susceptibility to beetle
attacks than controls during the two year observation.
Figure 11. Symptoms of Ceratocystis polonica infection in Norway spruce trees with no apparent damage (white bars), short-leader (black bars) and leaning-stem (grey bars), 11 weeks after inoculation. The data were collected from each of 5 trees and expressed as mean + 1 SE. Bars with different letters were significantly different by LSD test at p = 0.05.
a
b
a
ab
b
bb
ba
b
a
ab
b
bb
b
10
20
30
40
50
60
70
Occluded sapwood Dead cambium Bluestained sapwood
Perc
ent o
f tis
sue
b b
a
a
b b
bb
a

21
The impacts of abiotic- and biotic stressors on conifer resistance have
been well documented, but often with contrary conclusions. For example, it
is well agreed that drought is a factor reducing tree vigor and predisposing
trees to beetle attacks (Negron et al. 2009; Nilsson et al. 2004; Rouault et al.
2006; Wermelinger 2004), but moderate drought has been repeatedly
observed to render trees more resistant to bark beetles and their associated
fungi (Christiansen and Glosli 1996; Lorio 1986; Salle et al. 2008).
Similarly, although fungal pathogens and insects kill conifers frequently,
low density fungal inoculation enhanced the resistance of Scots pine and
Norway spruce to subsequent mass inoculation with the same fungal
species (Christiansen et al. 1999; Krokene et al. 1999; 2000). Furthermore,
previously unsuccessful beetle attacks provided resistance to Norway spruce
against later beetle colonization (P. Krokene et al. unpublished).
Constitutiveresistance
Induced responseTime
Resis
tanc
e
a: With adequate carbon reserve (Induced resistance)
b: Carbon reserve dep leted
Induced susceptibili ty
(Resistance decreases)
Figure 12. Possible ecological consequences of plant induced response (modified from Bonello et al. 2006), showing the interplay between plant carbon reserve and induced resistance levels.
Bonello et al. (2006) proposed that plant induced response is a
dynamic interplay between tree-stressor(s), which may lead to an ecological
consequence of induced resistance or induced susceptibility (Figure 12).
Upon invasion, the induced response starts rapidly and the concentration of
secondary defensive compounds systemically increases for a period. If the
induced response can constrain the invasion before the available carbon
reserve is depleted it would lead to induced resistance, otherwise it could
result in induced susceptibility.

22
3.2. Terpene and phenolic composition
Simultaneous with the relatively poor performance of the fungus in trees
with apparent damage, we found that the terpene (Figure 13A) and
phenolic (Figure 13B) compositions in the phloem were different between
tree groups. The relative amounts of (+)-3-carene and two unidentified
stilbenes were significantly higher, and taxifolin glycoside was lower in no-
damage trees than short-leader and leaning-stem trees (Appendix B1).
A B
Resveratrol is an antifeedant to I. typographus (Faccoli and Schlyter
2007) and it inhibits the growth of the bark beetle associated fungi
Ophiostoma piceaperdum and O. bicolor on malt agar medium (Salle et al.
2005). This compound was found to be rare or absent in unwounded
Norway spruce phloem, but was highly induced by fungal infection
(Brignolas et al. 1995a; 1998; 1995b; Evensen et al. 2000; Lieutier et al.
2003; Viiri et al. 2001). Similarly, taxifolin glycoside was only detected in
the phloem of resistant Norway spruce clones and the amount was related
Figure 13. PCA plot based on the relative amounts (normalized to 100%) of all the quantified terpenes (A) or phenols (B) in the phloem of Norway spruce trees with no apparent damage (triangle), short-leader (circle) and leaning-stem (cross), 17 months after the storm “Gudrun”. Each symbol represents one tree. The arrows indicate the contribution of terpenes to the principal components. In panel A the first principal component (PC1) explained 27.9%, and the second component (PC2) explained 17.4% of the sample variation. In panel B, PC1 explained 39.4% and PC2 explained 22.6% of the sample variation.
-1.0 1.0
-1.0
1.0
(-)-α-Pinene
(+)-α-Pinene
β-Pinene(+)-3-Carene
Myrcene
(+)-Limonene
β-Phellandrene
Longifolene
Caryophyllene
(-)-Germacrene D
δ-CadineneGermacrene D-4-ol
Thunbergol
Unknown DT
Unknown DT
α-Longipinene
Sabinene
Terpinolene
(-)-Limonene21
4
PC1, 27.9%
PC2,
17.
4%
-1.0 1.0
-1.0
1.0
Catechin
Unown flavonoid
Astrigin
Taxifolin glycoside
Piceid
TaxifolinIsorhapontigenin
Unknown stilbene
Unknown stilbene
Resveratrol
Isorhapontin
PC1: 39.4%
PC2:
22.
6%

23
to tree resistance against C. polonica (Brignolas et al. 1995a; 1998; 1995b;
Lieutier et al. 2003). In our experiment, we observed much higher
proportions of resveratrol and taxifolin glycoside in the phloem of the
experimental trees compared with our previous study of healthy spruce,
probably indicating that the phenolic defense in all the experimental trees
had been induced.
3.3. Correlation between resistance and chemical composition
In addition to the differences between tree categories, some chemicals
showed either positive or negative correlation with resistant related
parameters. The proportion of (-)-α-pinene was negatively correlated with
the percentage of occluded sapwood and dead cambium. (+)-3-Carene,
sabinene, (-)-germacrene D, thunbergol and the two unidentified stilbenes
were positively correlated with one or more symptoms of fungal infection
(Appendix B2). These results further indicate that the resistance to fungal
inoculation is more or less related to the chemical composition in the
experimental trees.
In Norway spruce, (+)-3-carene has been induced by MeJA
application (Zulak et al. 2009), C. polonica infection (Paper II) and
Heterobasidion spp. inoculation (Paper VII), but its function in tree
resistance is still poorly understood. We found that the proportion of (+)-3-
carene is positively correlated with the growth of C. polonica, indicating that
this compound might be associated with susceptibility to C. polonica
inoculation in Norway spruce suffering windstorm damage.

24

25
4. Fungal Inoculation and MeJA Application Induced Terpene Responses
(Paper II, Paper III)
As controlled levels of bark beetle attacks are difficult to achieve
experimentally, fungal inoculation has long been used to mimic beetle
attacks for characterizing tree responses to such attack (Christiansen 1985;
Sandnes and Solheim 2002). Quantitative terpene accumulations, a varied
phenolic composition and anatomical changes after fungal inoculation have
been observed in a number of conifers including Norway spruce (Raffa and
Berryman 1982a; Raffa and Berryman 1983a; Raffa and Smalley 1995;
Viiri et al. 2001). MeJA, a signaling phytohormone which plays an
important role in various developmental processes in plants (Beale and
Ward 1998; Turner et al. 2002), has recently been observed to induce
similar anatomical, terpenoid and molecular responses in the bark of
conifers to fungal inoculation (Franceschi et al. 2002; Hudgins et al. 2004;
Martin et al. 2002; Zeneli et al. 2006; Zulak et al. 2009), but no notable
phenolic change was found after MeJA treatment (Zeneli et al. 2006).
MeJA
Figure 14 .The structure of MeJA and the perithecium of Ceratocytis (Photographed by H. Solheim).
In the field, exogenous MeJA application or fungal inoculation
enhanced conifer resistance to white root rot pathogen Pythium ultimum
(Kozlowski et al. 1999), large pine weevil Hylobius abietis (Heijari et al.
2005), bluestain fungus C. polonica (Christiansen et al. 1999; 1999; Krokene
O
O
O

26
et al. 2008b; Zeneli et al. 2006) and spruce bark beetle I. typographus
(Erbilgin et al 2006). Hence it would be possible to develop vaccination
methods based on MeJA or artificial inoculation for future forest
protection. However, there are still gaps in our knowledge about the
mechanism and ecological function of MeJA and C. polonica induced
defenses. For example, little is known about the systemic terpene induction
and how long the chemical induction could last in conifers. In addition, no
detailed study has been done comparing the terpene content induced by
fungal inoculation and exogenous MeJA application.
4.1. C. polonica inoculation induced terpene responses
4.1.1. Terpene response in reaction zone (Paper III)
After fungal inoculation or beetle attack, the formation of callus tissues
results in an elliptical dark brown zone called the reaction zone. This zone
is highly lignified and full of defensive compounds, which may protect the
trees from further damage (Franceschi et al. 2005).
In accordance with the previous observations from Norway spruce
and other conifer species (Fäldt et al. 2006; Klepzig et al. 2005; Raffa and
Berryman 1982a; 1983a; Viiri et al. 2001), C. polonica inoculation also
elicited very strong terpene accumulation in the reaction zone in the
present experiment. The levels of all the terpenes in the reaction zone were
significantly higher than in untreated stem sections at both sampling times
(14 days and 35 days after fungal inoculation), and an extremely strong
terpene accumulation was observed at the second sampling time (Figure
15). The levels of total terpene in the reaction zone were 7- and 95- fold
higher 14 and 35 days after treatment, respectively, than in untreated stem
sections.

27
In general, the terpene response to fungal inoculation was more
quantitative than qualitative at both sampling times, but the relative
proportions of some minor compounds (constituting < 8 % of the total
terpene amount in untreated bark) increased after inoculation. 14 days after
inoculation, the proportions of myrcene, (E)-β-caryophyllene, α-terpineol
and an unidentified diterpene in the reaction zone were significantly higher
than in the untreated phloem; proportions of (+)-3-carene, camphene,
terpinolene, bornyl acetate, (E)-β-caryophyllene, α-terpineol, (-)-
germacrene D and two unidentified diterpenes were higher 35 days after
fungal inoculation.
In Scots pine, monoterpene induction started one day after
inoculation, and increased up to 800-fold and 30-fold 28 days after
Leptographium wingfieldii and Ophiostoma canum inoculation, respectively
(Fäldt et al. 2006). In red pine (Pinus resinosa), O. ips inoculation rapidly
increased the terpene level for two weeks after which it began to reduce
(Raffa and Smalley 1995). In the present experiment, we detected much
higher terpene levels in the second sampling time than the first. These
70
60
50
40
30
20
10
0
(+)-
3-C
aren
eβ-
Phe
lland
rene
Abi
enol
β-P
inen
e
Tric
ycle
neC
amph
ene
Sab
inen
e
(-)-
Lim
onen
e(+
)-Li
mon
ene
(+)-
α-P
inen
e(-
)-α-
Pin
ene
Myr
cene
Terp
inol
ene
Unk
now
n M
TU
nkno
wn
ST
α-Lo
ngip
inen
eB
orny
l ace
tate
Long
ifole
ne
α-Te
rpin
eol
Hum
ulen
e
(E)-
β-C
aryo
phyl
lene
(E)-
β-Fa
rnes
ene
Ger
mac
rene
Dδ-
Car
dien
e
Ger
mac
rene
D-4
-ol
Thun
berg
ene
Thun
berg
olU
nkno
wn
DT
Unk
now
n D
T
Cub
enol
Unk
now
n S
T
Terp
ene
amou
nt (m
g g-
1 dr
y w
t)
(-)-
Figure 15. Levels of individual terpenes in untreated bark (black bar) and in the reaction zone in Norway spruce bark 14 (grey bar) and 35 days (white bark) after inoculation with Ceratocystis polonica. Data are collected from the experiment carried out in 2008 and expressed as means + 1 SE. n = 20 trees.

28
results suggest that the rate and intensity of terpene defenses in conifers
greatly depends on the tree species and virulence of fungal species.
Although fungal inoculation generally increased the terpene level in
inoculated trees, the intensity of terpene induction differed extensively
among the 20 experimental trees. The total terpene levels in the reaction
zone varied from 0.9 to 42.5 mg terpene g-1 dry phloem 14 days after
fungal inoculation, and from 49.2 to 462.1 mg terpene g-1 dry phloem 35
days after fungal inoculation (Figure 21). The different inductive intensities
may reflect a diverse genetic background and physiological condition (Raffa
and Berryman 1983b), and probably dissimilar defense abilities among the
trees.
4.1.2. Terpene responses near reaction zone (Paper II)
200
300
Abs
olut
e am
ount
(ug
g -1
dry
wt)
0
50
100
150
200
100
0
CT Cp CT Cp
(+)-α-Pinene Sabinene
(+)-3-Carene Terpinolene
**
**
Figure 16. Absolute amount of selected terpenes in Norway spruce phloem before (white bars) and 30 days after (grey bars) stem inoculation with Ceratocystis polonica (Cp). CT denotes untreated control trees. Data are collected from the experiment carried out in 2007 and expressed as Means + 1SD. n = 16 trees except (+)-α-pinene where n = 8. Bars with stars were significantly different by t-test (p < 0.05).

29
In contrast to the strong quantitative terpene accumulation in the reaction
zone, fungal inoculation increased the amount of (+)-3-carene, sabinene
and terpinolene, and decreased (+)-α-pinene and (+)-limonene, but did not
markedly change the amount of other terpenes 5 cm above the inoculation
hole (ca. 1-3 cm above reaction zone) 30 days after fungal inoculation
(Figure 16, Appendix D). (+)-3-Carene, sabinene and terpinolene are
products of (+)-3-carene synthase (Fäldt et al. 2003). These results indicate
that fungal infection activates (+)-3-carene synthase, inhibits some terpene
synthases but does affect others in this sampling position. Induction of 3-carene is repeatedly observed upon abiotic- and biotic
stressors. For example, 3-carene is the major monoterpene induced in Scots
pine roots upon infection with the ectomycorrhizal fungus, Boletus
variegatus (Krupa and Fries 1971). Infection of Ceratocystis clavigera in
lodgepole pine (Pinus contorta)(Croteau et al. 1987) and Leptographium
wingfieldii in Scots pine (Fäldt et al. 2006) phloem also induced high (+)-
3-carene production. In addition, significantly higher (+)-3-carene
accumulation has also been observed in Norway spruce saplings after MeJA
treatment (Zulak et al. 2009, Fäldt et al. 2003). In our experiment, MeJA
treatment only increased the (+)-3-carene level in a few trees (section 4.2),
but this terpene was greatly induced by C. polonica inoculation in most of
the experimental trees, indicating that (+)-3-carene may be an interesting
chemical marker of conifer resistance or susceptibility. Previous
investigations have already associated 3-carene levels with conifer resistance
to some insects and fungi (Pasquier-Barre et al. 2001; Reed et al. 1986;
Rocchini et al. 2000). More studies should be done to demonstrate the
function of (+)-3-carene induction to spruce bark beetle and its fungal
associates.
Ips typographus had significantly stronger electroantennogram
responses to (-)-α-pinene than to the (+)-enantiomer (Dickens 1978).
More importantly, I. typographus uses ingested (-)- and (+)-α-pinene as
precursors to synthesize cis- and trans-verbenol respectively. The
enantiomeric composition of α-pinene in the host tissue therefore
correlates with the ratio of the two verbenol isomers in the gut of bark

30
beetles (Klimetzek and Francke 1980; Lindström et al. 1989). In this
experiment, fungal inoculation increased the enantiomeric ratio of α-pinene
and limonene (Figure 17). The altered α-pinene chirality in C. polonica
infected bark may change the ratio of cis-/trans-verbenol produced by bark
beetles when they excavate galleries in the infected area. In the same
experiment, we found that the relative amount of (-)-limonene was
significantly higher, and enantiomeric ratio of (-)-α-pinene was lower in
the seven relatively unattractive trees (< 5 beetles landing or entering) than
in the six relatively attractive ones (> 20 bark beetle landing on, and /or >
10 beetle entering the bark), indicating the altered enantiomeric
composition of α-pinene and limonene may have certain impacts on host
selection by I. typographus.
4.2. MeJA treatment induced terpene responses
MeJA elicited terpene accumulation in the bark of most experimental trees
in both the 2007 and 2008 experiments. In 2007 all the terpenes in the
experimental trees increased somewhat (Appendix D), but only (-)-
limonene and (-)-β-pinene had significantly increased 30 days after
treatment (Paper II).
40
20
0
60
80
Cp MeJA CT Cp MeJA CT
Per
cent
age
of (-
)-en
antio
mer
**
*
Limoneneα-Pinene
Figure 17. Enantiomeric composition (mean + 1 SD) of α-pinene and limonene before (white bars) treatment with methyl jasmonate (MeJA) or Ceratocystis polonica (Cp) (grey bars) and one month after treatment. CT denotes untreated control trees. Data are expressed as the percentage of the (-)-enantiomer to the sum of both enantiomers. n = 8 trees. Bars with stars were significantly different by t-test (p < 0.05).

31
In 2008 we detected much stronger terpene induction than in the
2007 experiment. Most of the major individual terpenes were extensively
induced at both sampling times (Figure 18). Compared with the control
trees, total terpene level in MeJA-treated trees was 2.2-fold higher 14 days,
and 2.6-fold higher 35 days after treatment.
MeJA treatment seemed to activate the synthesis of individual
terpenes similarly 35 days after treatment. Although the total terpene level
was significantly higher, only the relative amount of abienol, but not other
terpenes, changed. In contrast, MeJA treatment seemed to activate the
synthesis of some terpenes more than others 14 days after treatment. The
overall relative amounts of (-)-germacrene D, germacrene D-4-ol, δ-
cadinene, thunbergol and abienol were significantly higher, and α-pinene
and β-pinene were significantly lower than the control at this sampling
time (Figure 18). High (+)-3-carene induction was also observed in some
experimental trees (Figure 19).
0
20
0
0.5
1
1.5
2MeJA (Day14)MeJA (Day35)Control (Day14)
Tric
ycle
ne
α-P
inen
e
Cam
phen
e
β-P
inen
e
Sab
inen
e
Myr
cene
Lim
onen
e
(+)-
3-C
aren
e
β-P
hella
ndre
ne
Terp
inol
ene
Unk
now
n S
T
α-Lo
ngip
inen
e
Bor
nyl a
ceta
te
α-Te
rpin
eol
Ger
mac
rene
D(-
)-
δ-C
ardi
ene
Cub
enol
Ger
mac
rene
D-4
-ol
Thun
berg
ene
Thun
berg
ol
Abi
enol
Unk
now
n D
T
Unk
now
n D
T
p-C
ymen
e
Long
ifole
ne
(E)-
β-C
aryo
phyl
lene
Abs
olut
e am
ount
(mg
g-1
dry
wt)
Rel
ativ
e am
ount
(%)
Figure 18. The absolute and relative amount of terpenes in the bark of MeJA treated and control trees. Day 14 and Day 35 represent 14 days and 35 days after MeJA treatment respectively.

32
Terpene accumulation following MeJA treatment has been observed
repeatedly in the bark of Norway spruce (Martin et al. 2002; Zeneli et al.
2006; Zulak et al. 2009). In a seedling based experiment, Zulak et al. (2009)
reported that the molecular and biochemical responses of Norway spruce to
MeJA were time dependant. During the 35 day observation period, the
highest mono- and diterpene productions were detected 16 days after
treatment. In our experiment, the total terpene level didn’t vary between the
two sampling times, but the levels of sesquiterpenes (-)-germacrene D,
germacrene D-4-ol and δ-cadinene and diterpene thunbergol were
significantly higher 14 days after treatment than the later sampling (Figure
18 and 19).
Figure 19. Representative total ion chromatograms from treated and untreated section of MeJA treated trees. A. treated section (14 days after treatment); B. treated section (35 days after treatment); and C. untreated section (14 days after treatment) in the same tree.
In a field study, Zeneli et al. (2006) applied MeJA to the stems of
clonal trees and compared the terpene responses between clones. They
found that the terpene induction varied extensively among the six Norway
spruce clones, four weeks after treatment. Trees from one clone showed
little response, the terpene levels in two clones increased 3-4 fold and the
other three clones had intermediate terpene responses. Similarly, we found
that the intensities of terpene induction varied extensively among the 20
4 8 12 16 20 24 28 32 36 40 44 48
Time (min)
50
100
50
100
Rel
ativ
eA
bund
ance
50
100
Treated_35 days
Untreated_14 days
Treated_14 day
α-Pineneβ-Pinene
β-PhellandreneTerpinolene
Internal standard Germacrene D
Germacrene D-4-olδ-Cadinene MeJA
Antioxidant
Thunbergol3-Carene
A.
B.
C.
(+)-

33
experimental trees at both sampling times. Fourteen days after treatment, 3
trees showed extremely strong terpene responses (Figure 19) whereas 5
trees exhibited minor changes.
In the above-mentioned experiment, Zeneli (2006) observed little
terpene response in all the experimental trees 8 days after treatment.
Considering both results (Zeneli et al. and our own), we can speculate that,
in natural conditions, mature Norway spruce trees might need 8-14 days to
activate their terpene synthesis. In contrast, Zulak et al. (2009) detected
terpene accumulation in the bark of Norway spruce seedlings 5 days after
treatment. The different observations might be due to different
temperature and moisture conditions, MeJA application methods, or
because terpene response in Norway spruce seedlings is faster than in
mature trees.
4.3. Comparison between MeJA and C. polonica induced terpene response
Miller et al. (2005) compared the terpene responses of Sitka spruce (Picea
sitchensis) following attack by white pine weevils (Pissodes strobe), or MeJA
application. The pine weevil induced stronger terpene accumulation in the
stem, as well as a more complex terpene emission than MeJA application.
They concluded that weevils and MeJA induced similar, but not identical
terpenoid responses in Sitka spruce.
In this study we found that the terpene accumulation in the reaction
zone after fungal inoculation was much stronger than that elicited by MeJA
application (details see Figure 15 and 18), and fungal infection induced a
completely different terpene profile in a position 5 cm above the
inoculation hole (Figure 16, Appendix D) (Paper II).
4.4. MeJA and C. polonica induced terpene response lasted one year
Krokene et al. (2003) found that C. polonica inoculation could protect
Norway spruce for at least one year, but terpene induction one year after
fungal inoculation has not been previously reported. In our study, we found

34
that the terpene profile in the bark of inoculated trees was still different
from that of control trees one year after inoculation (Figure 20), indicating
that the terpene defense induced by fungal inoculation may last for at least
one year (Paper II).
Figure 20. PCA plots based on absolute amounts (A) and relative amounts (B) of all the quantified terpenes in Norway spruce phloem 362 days after stem treatment with Ceratocystis polonica (circles) or methyl jasmonate (triangles). + = untreated control trees that were also sampled at the start of the experiment; × = untreated control trees with no previous sampling history.
Similarly, the terpene induction elicited by MeJA also lasted over one
year. Most of the treated trees had a terpene profile indicating an increase
in (+)- and (-)-α-pinene, (-)-β-phellandrene, (-)-β-pinene, myrcene and
(+)-limonene compared with the untreated control (Figure 20) (Paper II).
This conclusion is in agreement with Erbilgin et al. (2006).
A
-0.4 1.0
-0.6
0.8
(+)-α-Pinene
Sabinene
(-)-β-Pinene
(+)-3-Carene
(-)-β-Phellandrene
(-)-Limonene
Terpinolene
4b
7b
7c
10b
13c
16b22C
19a
2b
2c
5b
5C
14b
17c20c
3c
6b
9a
9b
15b
21a
21b
21c
CT1CT3
CT4
CT5
CT6
CT7
CT8
CT2
Thunbergol
PC1, 74.5%
PC2,
14.
5%
(-)-α-Pinene(+)-Limonene
Myrcene
Camphene
B
-1.0 1.0
-0.8
0.8
(-)-α-Pinene(+)-α-Pinene
Camphene
Sabinene
(-)-β-Pinene
Myrcene
(+)-3-Carene
(-)-β-Phellandrene
(+)-Limonene
Terpinolene
δ-Cadinene
Germacrene D
Thunbergol
4b
7b
7c
10b
13c16b
22c
19C
2b
2c
5b
5c
14b
17c
20c
8C3c
6b
9a
9b
15b
21a
21b
21c
CT1
CT3CT4
CT5
CT6
CT7CT8
CT2
PC1, 36.5%
PC2,
21.
6%

35
4.5. Does Norway spruce process systemic terpene induction?
Wounding, fungal infection and MeJA treatment induced traumatic resin
duct formation in tissues far away from the treated section in Norway
spruce (Franceschi et al. 2000; Krekling et al. 2004; Krokene et al. 2008a).
Systemic phenolic induction has also been observed in ponderosa pine
(Pinus ponderosa) phloem (Bonello et al. 2003), demonstrating that conifers
have the ability to generate systemic induction.
In this study, we observed terpene induction in a sampling position 5
cm above the inoculation hole (ca. 1-3 cm away from the reaction zone),
but little chemical change was observed in the position 30 cm below the
treated zone in both fungal and MeJA-treated trees. These results suggest
that Norway spruce has a minor ability to induce terpene in un-infested
positions (Appendix D).

36

37
5. Terpene Accumulation Reduced Bark Beetle Colonization
Paper III
Terpenes have long been considered to play a central role in conifer
resistance against the bark beetle-fungi complexes (Keeling and Bohlmann
2006; Phillips and Croteau 1999). Many studies have demonstrated their
biosynthesis pathways and metabolic responses to various stress elicitors
(for reference see Keeling and Bohlmann 2006). However, the evidence for
the defensive role of terpenes is largely indirect and rests on the facts that
synthetic monoterpenes are lethal to bark beetles (Raffa and Smalley 1995;
Smith 1965) and inhibit the growth of beetle associated fungi in laboratory
bioassays (Solheim 1991). The lack of field studies is probably due to the
difficulty associated with manipulating tree resistance in the field (Erbilgin
et al. 2006).
Recently, some efforts have been made to demonstrate the role of
terpenes in conifer resistance against bark beetles in the field, by means of
fungal inoculation or exogenous MeJA treatment. In Norway spruce, both
treatments induced terpene accumulation in the bark (Viiri et al. 2001;
Zeneli et al. 2006) and enhanced the resistance of Norway spruce to I.
typographus and its associated fungus C. polonica (Erbilgin et al. 2006;
Krokene et al. 1999; Zeneli et al. 2006). However, since anatomical
(Franceschi et al. 2002; Krokene et al. 2003) and other chemical changes
(e.g. phenolics) (Brignolas et al. 1995b; Evensen et al. 2000; Viiri et al.
2001) were also induced during these treatments, it is difficult to conclude
that the elevated terpene level after treatment is directly responsible for the
observed resistance. More precise knowledge about the function of terpene
accumulation under field conditions is needed to better understand the role
of terpenes in conifer resistance.

38
5.1. Variation in terpene response and beetle colonization
Fungal inoculation induced different intensities of terpene accumulation
among the 20 experimental trees (Figure 21). Additionally, the colonization
of bark beetles differed extensively between the trees. Attack density
reached up to 185.7 attacks per m2 bark, and the gallery length reached 3.0
m per m2 bark, but in 5 experimental trees no attacks were found. The
differences between terpene and bark beetle colonization levels provide a
possibility to more precisely evaluate the effect of terpenes on tree
resistance.
5.2. Terpene accumulation in trees with different susceptibility
Figure 21. Terpene levels in the wound reaction zone in Norway spruce bark 14 and 35 days after Ceratocystis polonica inoculation and baseline levels in uninoculated bark. Based on the extent of colonization by Ips typographus, trees were classified as susceptible (114.4 ± 70.3 (SD) attacks and 1.5 ± 1.0 m beetle galleries m-2 bark; n = 5) or resistant (1.1 ± 2.6 beetle attacks and 0.02 ± 0.04 m beetle galleries m-2 bark; n = 6). Intermediate trees had 31.6 ± 16.8 attacks and 0.2 ± 0.26 m beetle galleries m-2 (n = 9).
In order to closely investigate the relationship between terpene
accumulation and beetle colonization, we distinguished three levels of tree
Baseline Day 14 Day 35
Terp
ene
leve
l (m
g g-1
dry
wt)
0
100
200
300
400
500Resistant treesSusceptible treesOther treesIntermediate trees

39
resistance using different lines (solid, dashed and dotted; Figure 21). We
marked the 6 most resistant trees (1.1 ± 2.6 beetle attacks and 0.02 ± 0.04
m beetle galleries m-2 bark) with solid lines, the 5 most susceptible trees
(114.4 ± 70.3 attacks and 1.5 ± 1.0 m beetle galleries m-2 bark) with dashed
lines and the other 9 trees with intermediate resistance (31.6 ± 16.8 attacks
and 0.2 ± 0.26 m beetle galleries m-2) with dotted lines. The figure clearly
shows that tree resistance is correlated with the terpene level in the reaction
zone 35 days after fungal inoculation. The 5 most resistant trees showed
the highest terpene levels, the 5 most susceptible trees showed relatively
low terpene levels while the remaining 9 trees exhibited low to moderate
terpene levels at this sampling time (Figure 21).
Figure 22. Total mono- (MT), sesqui- (ST) and diterpene (DT) amounts in the stem bark of susceptible, intermediate and resistant (for details see Figure 21) Norway spruce, 35 days after inoculation with the bluestain fungus Ceratocystis polonica. Data are expressed as means + 1 SE. Bars with different letters were significantly different by LSD test at p = 0.05 following ANOVA.
Raffa and Berryman (1982b; 1983a) found that the resistant
lodgepole pine trees produced more quantities of resin than susceptible
ones after fungal inoculation; similar results were obtained from Abies
grandis upon inoculation with the fungus associated with Scolytus ventralis
(Raffa and Berryman 1982a; Raffa et al. 1985). In our experiment, the
levels of mono-, sesqui- and diterpene in the reaction zone 35 days after C.
polonica inoculation increased correspondingly in the three groups with
50
100
150
200
250
300
MT ST DT
a
bc
a ab
b
c
5
10
15
20
MT ST DT
Terp
ene
amou
nt (m
g g
-1 d
ry w
t)
Susceptible treesIntermediate treesResistant trees
a
a
a
aa
a
aa
a c a
Day 14 Day 35

40
increasing resistance to I. typographus. The mono- and diterpene levels
varied significantly between the three groups (Figure 22). This result
convincingly showed that the terpene levels induced by C. polonica
inoculation were directly responsible for the observed increase in tree
resistance to I. typographus.
In contrast, the terpene levels in the reaction zone 14 days after
fungal inoculation showed less impact on beetle colonization (Figure 21
and 22 ), probably because the pheromone dispensers were set in place one
month after treatment and the beetle colonization in the trees was
investigated at the end of the season. However, this observation is in
agreement with Krokene et al. (2003) which demonstrated that Norway
spruce needs 1-3 weeks to activate the induced resistance after C. polonica
inoculation.
Although significantly different terpene levels were observed in the
three tree groups, no notable difference on the relative amounts of any
major terpenes was observed between the three groups, or between
susceptible and resistant trees at both sampling times. These results
correspond with previous studies on pine trees (Raffa and Smalley 1995;
Wallin and Raffa 2000b), and suggest that the total quantity of terpenes,
rather than any qualitative difference, inhibited the entry and gallery
excavation of I. typogaphus in Norway spruce.
5.3. Terpene accumulation inhibited I. typographus colonization in a dose-
dependent manner
Although total terpene accumulation 35 days after inoculation inhibited
beetle colonization, the correlation between terpene level and beetle
colonization was not linear (Figure 23). Instead, there seemed to be two
threshold concentrations. Above a high threshold concentration (ca. 200
mg terpene g-1 dry phloem) trees more or less escaped beetle attack, as
evidenced by the few galleries and entrance holes in these trees. Above a
lower threshold concentration (ca. 100 mg terpene g-1 dry phloem) trees
experienced moderate beetle colonization, and attack density and gallery

41
construction were never high (Figure 23), whereas beetle colonization was
high in some of the trees below this lower threshold.
Figure 23. Attack densities and gallery lengths in 20 Norway spruce trees with different induced
terpene levels in the bark. Dotted lines show two potential thresholds in terpene levels (200 and 100
mg g-1 dry wt) with different inhibitive effects on beetle colonization. Chemical samples were taken
from the reaction zone 35 days after Ceratocystis polonica inoculation.
To illustrate the inhibitory effects of terpenes on beetle colonization
more precisely, we divided the 20 experimental trees into three groups using
the threshold levels above, and compared beetle performance in these three
groups. The attack density in trees with low terpene levels (<100 mg g-1 dry
phloem) was 3.9-fold and 14.1-fold higher than trees with medium (100-
200 mg g-1 dry phloem) and high (> 200 mg g-1 dry phloem) terpene levels
respectively. Correspondingly, the gallery length in the low terpene trees
was 2.8- and 37.9-fold higher than the trees with medium and high
terpene levels, whereas there was no difference in beetle attraction (trap
catches) to trees with different susceptibility. These results indicated that
the terpene accumulation, induced by C. polonica inoculation, inhibited the
host entry and gallery excavation of I. typographus in a dose-dependent way
(Figure 24).
Terpene level (mg g-1 dry wt)
0 100 200 300 400 500
Atta
cks
m-2
0
50
100
150
200
0 100 200 300 400 500
Gal
lery
leng
th (m
m-2
)
0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
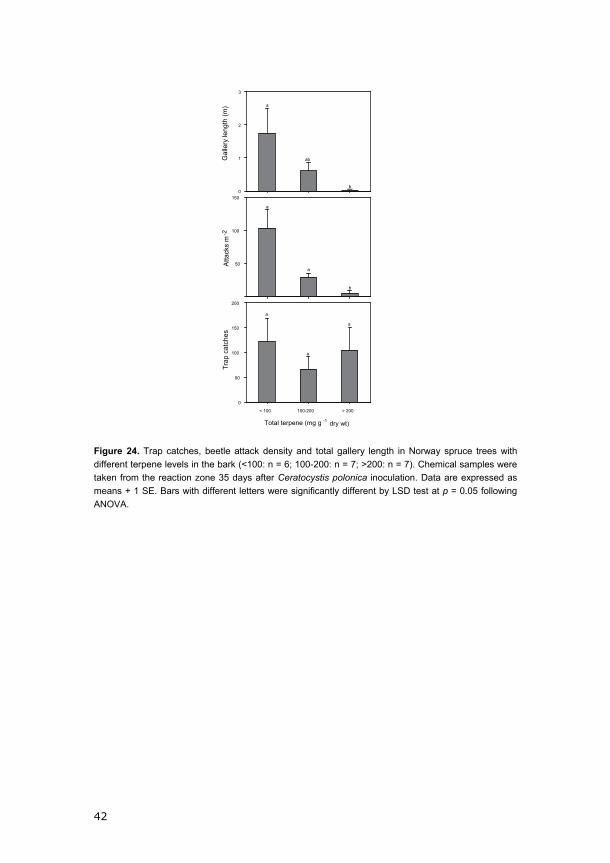
42
Figure 24. Trap catches, beetle attack density and total gallery length in Norway spruce trees with different terpene levels in the bark (<100: n = 6; 100-200: n = 7; >200: n = 7). Chemical samples were taken from the reaction zone 35 days after Ceratocystis polonica inoculation. Data are expressed as means + 1 SE. Bars with different letters were significantly different by LSD test at p = 0.05 following ANOVA.
Total terpene (mg g -1 dry wt)
< 100 100-200 > 200
Trap
cat
ches
0
50
100
150
200
Atta
cks
m-2
50
100
150
Gal
lery
leng
th (m
)
0
1
2
3
a
a
a
a
a
b
a
b
ab

43
6. Host Resistance Reduced Bark Beetle Pheromone Emission
(Paper IV)
Like other tree-killing bark beetles, I. typographus uses pheromones in the
host colonization process (Birgersson et al. 1984; Schlyter et al. 1987a).
Shortly after a bark beetle male has landed and started to feed on a tree, he
starts to produce aggregation pheromones to gather conspecifics to the tree.
Ips typographus can even kill healthy trees if a sufficient number of beetles
respond to the pheromone signals (Christiansen and Bakke 1988). The
quantity and quality of aggregation pheromone plays an essential role in the
successful colonization of I. typographus (Schlyter and Birgersson 1989).
Nine compounds (2-methyl-3-buten-2-ol, cis- and trans-verbenol,
ipsenol, ipsdienol, verbenone, myrtenol, trans-myrtanol and 2-
phenylethanol) have been identified from the hindgut of male I.
typographus (Bakke 1976; 1977; Birgersson et al. 1988; 1984) and tested as
pheromone candidates. The mixture of 2-methyl-3-buten-2-ol (MB) and
(S)-cis-verbenol (cV) results in significant trap catches in field experiments
(Bakke 1976; 1977), and the two compounds are commonly used as the key
components in commercial pheromone dispensers such as Ipslure, Typolure
and Pheroprax® (Schlyter et al. 1987a). Ipsdienol is also slightly attractive
to I. typographus, but it is only produced by the male after mating
(Birgersson et al. 1984). Ipsenol, tran-verbenol and verbenone have been
reported as antiaggregation pheromone for I. typographus (Byers 2004), but
myrtenol, trans-myrtanol and 2-phenylethanol showed no clear function in
the field (Schlyter et al. 1987b).
Birgersson et al. (1988) found that the pheromone production varied
extensively between individuals and speculated the pheromone production
may be regulated by host resistance (Birgersson 1989; Birgersson et al.
1988). However, whether host resistance directly interferes with the bark
beetle pheromone production remains unknown. Relevant knowledge is
clearly needed for an integrated understanding of host resistance.

44
(S)-cis-verbenol (cV) 2-methyl-3-buten-2-ol (MB)
OH HO
Figure 25. Structures of (S)-cis-verbenol and 2-methyl-3-buten-2-ol.
6.1. MeJA treatment elicited terpene defense in Norway spruce (Paper IV)
Regarding the 2008 field experiment, some trees showed minor terpene
response to MeJA treatment at both sampling times (see section 4.2), but
the MeJA treatment protected all the treated trees from bark beetle attack.
The attack densities and gallery lengths in MeJA-treated trees were
significantly lower than in the control. No trees treated with MeJA died,
but 9 out of the 20 control trees in the same stand were killed by I.
typographus. One possible explanation for this observation could be that, in
addition to the observed terpene accumulation in the intact phloem, MeJA-
treated trees may release more resin than control trees upon wounding by
the bark beetle.
To test this hypothesis, we mimicked bark beetle wounding by
puncturing the bark with a pushpin, sampled the wounded bark after 24 hrs
and analyzed the terpene content by GC-MS and 2D-GC (Paper IV). As
expected, whilst MeJA treatment itself only induced a moderate increase in
terpene levels in the bark, MeJA-treated logs showed a much stronger
quantitative response to mechanical wounding than untreated logs (Figure
26). In wounded phloem all terpene classes were significantly more
abundant in MeJA-treated than in untreated phloem and diterpenes
increased more than the other two terpene classes. Consequently, MeJA-
treated logs had higher diterpene and lower monoterpene proportions than
untreated logs (Figure 26, for the differences in individual terpenes see
Appendix C).

45
Norway spruce has specialized anatomic structures such as resin
ducts to store oleoresin (Franceschi et al. 2005). Upon invasion, new
traumatic resin ducts (TRD) are formed in the secondary xylem (Nagy et
al. 2000), providing extra resin flow to inhibit the invaders. Similar to
fungal infection or bark beetle attack, MeJA application also induces TRD
formation in the stem of Norway spruce trees (Hudgins et al. 2004; Martin
et al. 2002). Therefore, the particularly strong wounding response in MeJA-
treated logs might just be because the terpenes stored in the RTD were
passively released, or it may reflect that the MeJA treatment primed
Norway spruce against subsequently invasion. Similarly, the enhanced
diterpene proportion in MeJA-treated bark after wounding might be
simply due to the evaporation of monoterpenes, but there is also a
Figure 26. Absolute and relative amounts (normalized to 100%) of monoterpenes (MT), sesquiterpenes (ST) and diterpenes (DT) in intact (white bars) and wounded (grey bars) phloem of control logs and methyl jasmonate (MeJA) treated Norway spruce logs. Error bars denote one standard error. Wounded phloem was intensively wounded using a push pin 24 hrs before sampling. n = 3 logs, except for wounded phloem in MeJA treated logs where n = 5.
MT
5
10
15
20
25
30
ST DT
Control MeJA0
20
40
60
Control MeJA Control MeJA
Rel
ativ
e am
ount
(%)
Abso
lute
am
ount
(mg
g-1
dry
wt)

46
possibility that the enzymes related to diterpene synthesis were activated
more than others.
6.2. The performance of I. typographus in MeJA treated logs (Paper IV)
Similar to Erbilgin et al. (2006), we observed that beetles performed much
better in untreated than in MeJA treated logs, probably because they
encountered less terpene there. In untreated logs, all the analyzed gallery
systems contained mating chambers, but only 38.5 % of the analyzed
gallery systems contained mating chamber in MeJA treated logs.
Additionally, I. typographus constructed much longer galleries in untreated
logs than in treated ones (7.9 ± 1.6 cm, and 2.3 ± 0.4 cm respectively).
6.3. MeJA treatment altered pheromone quantity and quality (Paper IV)
The poorer bark beetle performance, higher terpene level in the phloem
and the much stronger wounding response in MeJA-treated than in
untreated logs indicated that the MeJA treatment elicited a defense
response in Norway spruce.
Raffa and Berryman (1983) reported that Dendroctonus ponderosae
Hopkins was unable to initiate aggregation on vigorous lodgepole pine
trees. Similarly, reduced aggregation by I. typographus was observed in
MeJA treated trees which exhibited a greatly elevated terpene level
(Erbilgin et al. 2006). It has been speculated that this failed aggregation can
be attributed to the impaired pheromone production by beetles exposed to
sticky resin and other tree defenses (Raffa and Berryman 1983; Erbilgin et
al. 2006). However, very few studies have investigated the actual
mechanisms of failed aggregation, particularly how induced tree defenses
affect the quality and quantity of pheromone production (Birgersson 1989;
Birgersson and Leufven 1988). How the host resistance affects the
pheromone production, especially the production of MB, by I. typographus
is still unknown.

47
Figure 27. Amounts of 2-methyl-3-buten-2-ol and (S)-cis-verbenol emitted from Ips typographus entrance holes in control logs (CT, n = 7 holes) and methyl jasmonate treated (MeJA, n = 13 holes) Norway spruce logs the first six days after beetle entry. Error bars denote one standard error.
In our experiments, we found that the induced defense in Norway
spruce not only negatively affected host colonization, but also reduced the
pheromone production by I. typographus. MB and cV were detected from
all the entrance holes in control logs, but only from 46.2 % and 61.5 % of
the entrance holes in MeJA-treated logs. Overall, beetle tunneling in
control logs produced 30 and 13.5 times more MB and cV respectively
than beetle tunneling in MeJA-treated logs during the six analyzed days
(Figure 27). The response of I. typographus to MB and cV is concentration
dependent (Dickens 1981; Schlyter et al. 1987b; 1987c). The reduced MB
and cV emission may therefore decrease the number of beetles attracted to
the tree, resulting in a failed attack. This observation may explain the
reduced aggregation after MeJA treatment (Erbilgin et al. 2006).
In addition, MeJA treatment also altered the ratio between MB and
cV. The ratio of MB to cV in the untreated logs was similar to that
observed by Birgersson and Bergström (1989), but much lower MB:cV
ratios were found for beetles tunneling in MeJA treated logs 2-6 days after
Days after beetle entry
1 2 3 4 5 60
1
2
3
4
5
1 2 3 4 5 6
0.02
0.04
0.06
0.08
0.10CT
MeJA
Am
ount
(ug-1
hol
e-1)
2-Methyl-3-buten-2-ol (S)-cis-Verbenol

48
beetle entry. MB and cV are predicted to have different roles in the
aggregation behavior of I. typographus. cV is used for long-range orientation
towards a potential forest, and MB is used for short-range host finding
(Dickens 1981; Schlyter et al. 1987b). The low MB:cV ratio in treated trees
suggested that tree defenses elicited by the MeJA treatment might affect
beetle landing on a specific tree more than its orientation to a stand.
-O
O
OH
OHOPPOPP
2-Methyl-3-buten-2-ol
MevalonateIsopentenyl DiphosphateDimethylallyl Disphosphate
OH
Figure 28. Proposed mevalonate pathway leading to 2-methyl-3-buten-2-ol in Ips bark beetle.
MB is synthesized de novo in the gut of the beetles, under strict
hormonal control (Lanne et al. 1989) (Figure 28). The MB production is
greatly dependent on the physiological condition of the bark beetle, and is
regulated by its tunneling and feeding activities (Birgersson et al. 1988).
These behaviors are probably regulated by terpenes (Erbilgin et al. 2006).
The mono- and diterpenes are thought to work together in conifer
defense. Upon invasion, the volatile monoterpenes act as solvents to take
diterpenes to the wounding sites (Gershenzon and Dudareva 2007), and
then mono- and diterpenes collectively deter the invader. After
monoterpenes have evaporated, diterpenes rapidly polymerize to create a
hardened barrier, which can seal the wound and stop the invasion of bark
beetles and their associated fungi (Phillips and Croteau 1999).
The terpene accumulation in the intact phloem and the strong
wounding response in MeJA treated bark suggest that bark beetles boring
into MeJA-treated bark encountered higher terpene concentrations and a
harder barrier compared with beetles tunneling in a control tree. The tough

49
environment negatively influenced the colonization of bark beetles in the
tree. The lower feeding activity (Bearfield et al. 2009) and the negative
antenna feedback (Ginzel et al. 2007) may inhibit the hormone related to
MB synthesis and reduce its emission.
In contrast, I. typographus uses digested (-)-α-pinene as a precursor to
synthesize cV. Hence, the production of cV may be regulated by the
amount of (-)-α-pinene consumed by the beetle. The lower proportion of
(-)-α-pinene in MeJA-treated bark after wounding, as well as the fact that
I. typographus fed less in MeJA treated logs may explain the reduced cV
emission in MeJA treated logs.
6.4. MeJA treatment altered host volatiles emitted from bark beetle
entrance holes
With the purpose of developing an effective pheromone collection and
analysis method, we started a pre-study in 2008. We chose two trees from a
Norway spruce clone (different from the clone we used in 2009), treated
the trees with MeJA, and collected the volatiles emitted from entrance
holes by I. typographus in the same way as we did in 2009 (described in page
15). We detected clearly different volatile emissions from MeJA-treated
and untreated logs (Figure 29).
The emissions from the entrance holes in MeJA-treated logs
contained lower proportions of α-pinene and β-pinene but higher
proportion of β-phellandrene than that in untreated logs. Interestingly,
higher proportions of aromatic compounds and oxygenated monoterpenes
such as borneol, bornyl acetate, terpinen-4-ol, α-terpineol, estragole and 2-
methylanisole were also detected from the entrance holes in MeJA-treated
logs (Figure 29).
The emissions of plant volatiles act as infochemicals in plant-
carnivore communication (Dicke et al. 2009) and may influence the
olfactory recognition of bark beetles to the host tree (Zhang and Schlyter
2004). So far little is known about the ecological function of the observed
aromatic compounds and oxygenated monoterpenes regarding I.

50
typographus, but 2-methylanisole elicited a strong antifeedant response to
the pine weevil Hylobius abietis (Borg-Karlson et al. 2006). In addition, 4-
allylanisole α-terpineol, and terpinene-4-ol significantly inhibited the
growth of mycangial fungi Ceratocystis minor and an unidentified fungal
species associated with southern pine beetle (Bridges 1987); terpinen-4-ol
and α-terpineol elicited a high GC-EAD (electroantennographic
detection) response to the bark beetle parasitoid (Pettersson et al. 2000),
suggesting that these volatile compounds may play a role in the host
selection of I. typographus.
However, we didn’t observe these differences in the clone we used in
the 2009 experiment-probably due to genetic differences between the two
clones. It would be interesting to repeat this experiment with more clones.
α-pinene
methyl butenol
methyl butenol
β‐pinene
β‐phellandrene
Limonene
β‐pinene
methylanisole
methylanisole
Estragole
Linalool oxide
myrcene
CT
MeJA
α-pinene
Estragole
Linalool oxide
myrcene
Figure 29. Representative total ion chromatograms charactering volatiles released from entrance holes by Ips typographus in methyl jasmonate (MeJA) treated and control (CT) logs.

51
7. Concluding Remarks
• Windstorm, and its damage to forests, is increasing, but little is known
about its impacts on the physiology and resistance of surviving trees. By
analyzing the terpene and phenolic composition in the phloem of the
Norway spruce tree with different levels of damage following the
"Gudrun" storm and evaluating their resistance to C. polonica
inoculation, we demonstrated that the trees apparently damaged by
windstorm showed more resistance to C. polonica inoculation. In
addition, the proportions of (+)-3-carene and two unknown stilbenes
were lower, and taxifolin glycoside was higher in the phloem of
apparently damaged trees than in the seemingly healthy trees.
• C. polonica inoculation strongly increased the amount of all the terpenes
in the reaction zone in Norway spruce stem, but only induced (+)-3-
carene, sabinene and terpinolene in the bark near the reaction zone. This
result reflected the different terpene biosynthetic activities in the two
sampling positions. This difference probably suggests that Norway
spruce employs different terpene defense strategies at different stages of
fungal invasion.
• MeJA elicited similar intensities of terpene induction 14 days and 35
days after treatment. More changes to relative amounts were observed
14 days after treatment. In addition, MeJA-treated logs showed a much
stronger response to mechanical wounding than untreated ones. This is
the first time a wounding response primed by MeJA treatment has been
demonstrated.
• Phytochemicals have long been considered to be important in conifer
defense against bark beetles, but the defensive role of phytochemicals
such as terpenes is still largely indirect. In this thesis, we demonstrated

52
that the induced terpene levels were significantly different between
resistant, intermediate and susceptible Norway spruce trees.
Furthermore, the terpene accumulation in response to fungal
inoculation inhibited the colonization of I. typographus in a dose-
dependent manner. Two threshold terpene concentrations, one resulting
in escape from beetle attack, and the other effectively reducing beetle
colonization were suggested. To the best of our knowledge, this is the
first study demonstrating a dose-dependent relationship between
induced terpenes and tree resistance to bark beetle colonization under
field conditions.
• Aggregation pheromones play an essential role in the life history of tree-
killing bark beetles, but how tree resistance influences the pheromone
production and composition is still unknown. In this thesis, we found
MeJA treatment elicited high terpene accumulation, reduced the
amount of pheromone emission and altered the ratio of the two key
pheromone components. These results indicate that host resistance can
directly influence the pheromone emission of I. typographus.

53
8. Acknowledgment
First of all, I would like to express my sincere gratitude to my supervisors,
Prof. Anna-Karin Borg-Karlson and Bo Långström, for accepting me as
PhD student, for involving me in the interesting multidiscipline projects,
for sharing your vast knowledge, and for all of your inspiring guidance,
discussions and comments. It is great experience to work with you.
The projects provide me the opportunity to work with Prof. Paal Krokene,
Nadir Erbilgin, Niklas Björklund, Halvor Solheim and Erik Christiansen.
Thank you so much for all kinds of contributions in the thesis and
publications. I do enjoy working with you.
Prof. Tobjörn Norin, Dr. Katinka Pålsson and Dr. Marie Danielsson are
gratefully acknowledged for valuable comments on this thesis; Dr. Douglas
Jones for linguistic revision and nice comments; Annie Yart for providing
phenolic references; Karolin Axelsson, Morgan Erixon, Ulf Johansson and
Olaug Olsen for technical assistance in the field.
Secondly, I would like to thank all the former and present members in
ecological chemistry group: Karolin Axelsson for the help in forest; Astrid
Kännaste for the kind help at the beginning of my PhD and in 2D-GC;
Raimondas Mozuraitis and Azeem Muhammad for help in instrument;
Yuxin Pei for comments on chemical structure; Marie Danielsson for
interesting discussion, for sharing the phenolic structures and references.
Additionally, thanks you all for the nice birthday party. It is a wonderful
surprise, and of course, a good memory in my life.
Thirdly, I would like to thank all the people in organic chemistry division
for various help, especially thanks to Lena Skowron and Henry Challis for
your help with practical stuff.

54
FORMAS, KTH and the Aulin-Erdtman foundation are gratefully
acknowledged for financial support.
My Chinese colleagues are gratefully acknowledged for the good
collaboration and for your trust and support.
At last, I would like to thank my family and friends, for your love,
understanding and support; especially thanks to Mengzhou for being such
a fantastic daughter, to Jiang and my parents for allowing me to study in
Sweden.

55
9. References
Baker B, Sinnott M (2009) Analysis of sesquiterpene emissions by plants using
solid phase microextraction. Journal of Chromatography A 1216:8442-8451
Bakke A (1976) Spruce bark beetle, Ips typographus, pheromone production and field response to synthetic pheromone. Naturwissenschaften 63:92-92
Bakke A, Froyen P, Skattebol L (1977) Field response to a new pheromonal compound isolated from Ips typographus. Naturwissenschaften 64:98-99
Bartelt RJ (1997) Calibration of a commercial solid-phase microextraction device for measuring headspace concentrations of organic volatiles. Analytical Chemistry 69:364-372
Beale MH, Ward JL (1998) Jasmonates: key players in the plant defence. Natural product reports 15:533-548
Bearfield JC, Henry AG, Tittiger C, Blomquist GJ, Ginzel MD (2009) Two regulatory mechanisms of monoterpenoid pheromone production in Ips spp. bark beetles. Journal of Chemical Ecology 35:689-697
Bidart-Bouzat MG, Imeh-Nathaniel A (2008) Global change effects on plant chemical defenses against insect herbivores. Journal of Integrative Plant Biology 50:1339-1354
Birgersson G (1989) Host resistance influencing pheromone production in Ips typographus. Holarctic Ecology 12:451-456
Birgersson G, Bergstrom G (1989) Volatiles released from individual spruce bark beetle entrance holes: Quantitative variation during the first week of attack. Journal of Chemical Ecology 15:2465-2483
Birgersson G, Leufven A (1988) The influence of host tree response to Ips typographus and fungal attack on production of semichemicals (Coleoptera, Scolytidae). Insect Biochemistry 18:761-770
Birgersson G, Schlyter F, Bergstrom G, Lofqvist J (1988) Individual variation in aggregation pheromone content of the bark beetle, Ips typographus. Journal of Chemical Ecology 14:1737-1761
Birgersson G, Schlyter F, Lofqvist J, Bergstrom G (1984) Quantitative variation of pheromone components in the spruce bark beetle Ips

56
typographus (Coleoptera, Scolytidae) from different attack phases. Journal of Chemical Ecology 10:1029-1055
Blomquist GJ et al. (2010) Pheromone production in bark beetles. Insect Biochemistry and Molecular Biology 40:699-712
Bochar DA, Stauffacher CV, Rodwell VW (1999) Sequence comparisons reveal two classes of 3-hydroxy-3-methylglutaryl coenzyme A reductase. Molecular Genetics and Metabolism 66:122-127
Bohlmann J, Keeling CI (2008) Terpenoid biomaterials. The Plant Journal 54:656-669
Bois E, Lieutier F (1997) Phenolic response of Scots pine clones to inoculation with Leptographium wingfieldii, a fungus associated with Tomicus piniperda. Plant Physiology and Biochemistry 35:819-825
Bonello P, Capretti P, Luchi N, Martini V, Michelozzi M (2008) Systemic effects of Heterobasidion annosum s.s. infection on severity of Diplodia pinea tip blight and terpenoid metabolism in Italian stone pine (Pinus pinea). Tree Physiology 28:1653-1660
Bonello P, Gordon TR, Herms DA, Wood DL, Erbilgin N (2006) Nature and ecological implications of pathogen-induced systemic resistance in conifers: A novel hypothesis. Physiological and Molecular Plant Pathology 68:95-104
Bonello P, Storer AJ, Gordon TR, Wood DL (2003) Systemic effects of Heterobasidion annosum on ferulic acid glucoside and lignin of presymptomatic ponderosa pine phloem, and potential effects on bark-beetle-associated fungi. Journal of Chemical Ecology 29:1167-1182
Boone CK, Six DL, Zheng YB, Raffa KF (2008) Parasitoids and dipteran predators exploit volatiles from microbial symbionts to locate bark beetles. Environmental Entomology 37:150-161
Borg-Karlson A-K, Lindström M, Norin T, Persson M, Valterova I (1993) Enantiomeric composition of monoterpene hydrocarbons in different tissues of Norway spruce, Picea abies (L) Karst. - A multidimensional gas-chromatography study. Acta Chemica Scandinavica 47:138-144
Borg-Karlson A-K, Nordlander R, Mudalige A, Nordenhem H, Unelius CR (2006) Antifeedants in the feces of the pine weevil Hylobius abietis: Identification and biological activity. Journal of Chemical Ecology 32:943-957

57
Bouget C, Duelli P (2004) The effects of windthrow on forest insect communities: a literature review. Biological Conservation 118:281-299
Bouvier-Brown NC, Holzinger R, Palitzsch K, Goldstein AH (2007) Quantifying sesquiterpene and oxygenated terpene emissions from live vegetation using solid-phase microextraction fibers. Journal of Chromatography A 1161:113-120
Bridges JR (1987) Effects of terpenoid compounds on growth of symbiotic fungi associated with the southern pine beetle. Phytopathology 77:83-85
Brignolas F et al. (1995a) Induced response in phenolic metabolism in two Norway spruce clones after wounding and inoculations with Ophiostoma polonicum, a bark beetle associated fungus. Plant Physiology 109:821-827
Brignolas F, Lieutier F, Sauvard D, Christiansen E, Berryman AA (1998) Phenolic predictors for Norway spruce resistance to the bark beetle Ips typographus (Coleoptera: Scolytidae) and an associated fungus, Ceratocystis polonica. Canadian Journal of Forest Research-Revue Canadienne De Recherche Forestiere 28:720-728
Brignolas F, Lieutier F, Sauvard D, Yart A, Drouet A, Claudot AC (1995b) Changes in soluble-phenol content of Norway-spruce (Picea abies) phloem in response to wounding and inoculation with Ophiostoma polonicum. European Journal of Forest Pathology 25:253-265
Byers JA (1993) Avoidance of competition by spruce bark beetles, Ips typographus and Pityogenes chalcographus. Experientia 49:272-275
Chappell J (1995) Biochemistry and molecular biology of the isopernoid biosynthatic pathway in plants. Annual Review of Plant Physiology and Plant Molecular Biology 46:521-547
Christensen E, Bakke A (1988) The spruce bark beetle of Eurasia. In: Berryman AA (ed) Dynamics of Forest Insect Populations. Plenum Publishing Corporation, New York, pp 479-503
Christiansen E (1985) Ceratocystis polonica inoculated in Norway spruce-blue staining in relation to inoculum density, resinosis and tree growth. European Journal of Forest Pathology 15:160-167
Christiansen E, Bakke A (1988) The spruce bark beetle of Eurasia. In: Berryman AA (ed) Dynamics of Forest Insect Populations. Plenum Publishing Corporation, New York, pp 479-503

58
Christiansen E, Glosli AM (1996) Mild drought enhances the resistance of Norway spruce to a bark beetle-transmitted blue-stain fungus. USDA For. Serv. Gen. Tech. Rep. NC-183
Christiansen E et al. (1999) Mechanical injury and fungal infection induce acquired resistance in Norway spruce. Tree Physiology 19:399-403
Croteau R, Gurkewitz S, Johnson MA, Fisk HJ (1987) Biochemistry of oleoresinosis1:Monoterpene and diterpene biosynthesis in lodgepole pine saplings infected with Ceratocystis clavigera or treated with carbohydrate elicitors. Plant Physiology 85:1123-1128
Danielsson M et al. (2008) Mini-seedlings of Picea abies are less attacked by Hylobius abietis than conventional ones: Is plant chemistry the explanation? Scandinavian Journal of Forest Research 23:299-306
Dicke M, van Loon JJA, Soler R (2009) Chemical complexity of volatiles from plants induced by multiple attack. Nature Chemical Biology 5:317-324
Dickens J (1981) Behavioural and electrophysiological responses to the bark beetle Ips typographus to potential pheromone components. Physiol Entomol 6:251-261
Erbilgin N, Krokene P, Christiansen E, Zeneli G, Gershenzon J (2006) Exogenous application of methyl jasmonate elicits defenses in Norway spruce (Picea abies) and reduces host colonization by the bark beetle Ips typographus. Oecologia 148:426-436
Erbilgin N, Krokene P, Kvamme T, Christiansen E (2007) A host monoterpene influences Ips typographus (Coleoptera: Curculionidae, Scolytinae) responses to its aggregation pheromone. Agricultural and Forest Entomology 9:135-140
Erbilgin N, Powell JS, Raffa KF (2003) Effect of varying monoterpene concentrations on the response of Ips pini (Coleoptera: Scolytidae) to its aggregation pheromone: implications for pest management and ecology of bark beetles. Agricultural and Forest Entomology 5:269-274
Erbilgin N, Raffa KF (2000) Effects of host tree species on attractiveness of tunneling pine engravers, Ips pini, to conspecifics and insect predators. Journal of Chemical Ecology 26:823-840
Evensen PC, Solheim H, Hoiland K, Stenersen J (2000) Induced resistance of Norway spruce, variation of phenolic compounds and their effects on fungal pathogens. Forest Pathology 30:97-108

59
Everham EM, Brokaw NVL (1996) Forest damage and recovery from catastrophic wind. Botanical Review 62:113-185
Eyles A, Bonello P, Ganley R, Mohammed C (2010) Induced resistance to pests and pathogens in trees. New Phytologist 185:893-908
Faccoli M, Schlyter F (2007) Conifer phenolic resistance markers are bark beetle antifeedant semiochemicals. Agricultural and Forest Entomology 9:237-245
Fäldt J, Eriksson M, Valterova I, Borg-Karlson AK (2000) Comparison of headspace techniques for sampling volatile natural products in a dynamic system. Journal of Biosciences 55c:180-188
Fäldt J, Martin D, Miller B, Rawat S, Bohlmann J (2003) Traumatic resin defense in Norway spruce (Picea abies): Methyl jasmonate-induced terpene synthase gene expression, and cDNA cloning and functional characterization of (+)-3-carene synthase. Plant Molecular Biology 51:119-133
Fäldt J, Solheim H, Langstrom B, Borg-Karlson AK (2006) Influence of fungal infection and wounding on contents and enantiomeric compositions of monoterpenes in phloem of Pinus sylvestris. Journal of Chemical Ecology 32:1779-1795
Franceschi VR, Krekling T, Christiansen E (2002) Application of methyl jasmonate on Picea abies (Pinaceae) stems induces defense-related responses in phloem and xylem. American Journal of Botany 89:578-586
Franceschi VR, Krokene P, Christiansen E, Krekling T (2005) Anatomical and chemical defenses of conifer bark against bark beetles and other pests. New Phytologist 167:353-375
Franceschi VR, Krokene P, Krekling T, Christiansen E (2000) Phloem parenchyma cells are involved in local and distant defense responses to fungal inoculation or bark-beetle attack in Norway spruce (Pinaceae). American Journal of Botany 87:314-326
Fredericksen TS, Hedden RL, Williams SA (1995) Susceptibility of loblolly pine to bark beetle attack following simulated wind stress. Forest Ecology and Management 76:95-107

60
Gallego D, Galian J, Diez JJ, Pajares JA (2008) Kairomonal responses of Tomicus destruens (Col., Scolytidae) to host volatiles alpha-pinene and ethanol. Journal of Applied Entomology 132:654-662
Gardiner B, Schelhaas M-J, Blennow K, Orazio C, Landmann G (2010) Wind strom and forest losses: A brief history of wind damage to Europe forests.
Gershenzon J, Dudareva N (2007) The function of terpene natural products in the natural world. Nature Chemical Biology 3:408-414
Ginzel MD, Bearfield JC, Keeling CI, McCormack CC, Blomquist GJ, Tittiger C (2007) Antennally mediated negative feedback regulation of pheromone production in the pine engraver beetle, Ips pini. Naturwissenschaften 94:61-64
Glitzenstein JS, Harcombe PA (1988) Effect of the December 1983 Tornado on forest vegetation of the big thicket, southeast Texas, USA Forest Ecology and Management 25:269-290
Heijari J, Nerg AM, Kainulainen P, Viiri H, Vuorinen M, Holopainen JK (2005) Application of methyl jasmonate reduces growth but increases chemical defence and resistance against Hylobius abietis in Scots pine seedlings. Entomologia Experimentalis Et Applicata 115:117-124
Hudgins JW, Christiansen E, Franceschi VR (2004) Induction of anatomically based defense responses in stems of diverse conifers by methyl jasmonate: a phylogenetic perspective. Tree Physiology 24:251-264
Hudgins JW, Franceschi VR (2004) Methyl jasmonate-induced ethylene production is responsible for conifer phloem defense responses and reprogramming of stem cambial zone for traumatic resin duct formation. Plant Physiology 135:2134-2149
Ivarsson P, Blomquist GJ, Seybold SJ (1997) In vitro production of the pheromone intermediates ipsdienone and ipsenone by the bark beetles Ips bini (Say) and I. paraconfusus Lanier (Coleoptera: Scolytidae). Naturwissenschaften 84:454-457
Jonsson AM, Appelberg G, Harding S, Barring L (2009) Spatio-temporal impact of climate change on the activity and voltinism of the spruce bark beetle, Ips typographus. Global Change Biology 15:486-499

61
Jost RW, Rice AV, Langor DW, Boluk Y (2008) Monoterpene emissions from lodgepole and jack pine bark inoculated with mountain pine beetle-associated fungi. Journal of Wood Chemistry and Technology 28:37-46
Keeling CI, Bohlmann J (2006) Genes, enzymes and chemicals of terpenoid diversity in the constitutive and induced defence of conifers against insects and pathogens. New Phytologist 170:657-675
Klepzig KD, Adams AS, Handelsman J, Raffa KF (2009) Symbioses: a key driver of insect physiological processes, ecological interactions, evolutionary diversification, and impacts on humans. Environmental Entomology 38:67-77
Klepzig KD, Robison DJ, Fowler G, Minchin PR, Hain FP, Allen HL (2005) Effects of mass inoculation on induced oleoresin response in intensively managed loblolly pine. Tree Physiology 25:681-688
Klimetzek D, Francke W (1980) Relationship between enantiomeric composition of alpha-terpene in host trees and the reproduction of verbenol in Ips species. Experientia 36:1343-1345
Knizek M, Beaver R (2004) Taxonomy and systematic of bark and ambrosia beetles. In: Lieutier F, Day KR, Battisti A, Grégoire JC, Evans HJ (eds) Bark and wood boring insects in living trees in Europe, a synthesis. Kluwer Acad. Pub, Dordrecht, pp 41-54
Komonen A, Schroeder LM, Weslien J (2011) Ips typographus population development after a severe storm in a nature reserve in southern Sweden. Journal of Applied Entomology 135:132-141
Koornneef A, Pieterse CMJ (2008) Cross talk in defense signaling. Plant Physiology 146:839-844
Kozlowski G, Buchala A, Metraux JP (1999) Methyl jasmonate protects Norway spruce [Picea abies (L.) Karst.] seedlings against Pythium ultimum Trow. Physiological and Molecular Plant Pathology 55:53-58
Krekling T, Franceschi VR, Krokene P, Solheim H (2004) Differential anatomical response of Norway spruce stem tissues to sterile and fungus infected inoculations. Trees-Structure and Function 18:1-9
Krokene P, Christiansen E, Solheim H, Franceschi VR, Berryman AA (1999) Induced resistance to pathogenic fungi in Norway spruce. Plant Physiology 121:565-569

62
Krokene P, Nagy NE, Krekling T (2008a) Traumatic resin ducts and polyphenolic parenchyma cells in conifers. Induced Plant Resistance to Herbivory:147 - 169
Krokene P, Nagy NE, Solheim H (2008b) Methyl jasmonate and oxalic acid treatment of Norway spruce: anatomically based defense responses and increased resistance against fungal infection. Tree Physiology 28:29-35
Krokene P, Solheim H, Christiansen E (2001) Induction of disease resistance in Norway spruce (Picea abies) by necrotizing fungi. Plant Pathology 50:230-233
Krokene P, Solheim H, Krekling T, Christiansen E (2003) Inducible anatomical defense responses in Norway spruce stems and their possible role in induced resistance. Tree Physiology 23:191-197
Krokene P, Solheim H, Langstrom B (2000) Fungal infection and mechanical wounding induce disease resistance in Scots pine. European Journal of Plant Pathology 106:537-541
Kunkel BN, Brooks DM (2002) Cross talk between signaling pathways in pathogen defense. Current Opinion in Plant Biology 5:325-331
Kurz WA et al. (2008) Mountain pine beetle and forest carbon feedback to climate change. Nature 452:987-990
Lange BM, Wildung MR, McCaskill D, Croteau R (1998) A family of transketolases that directs isoprenoid biosynthesis via a mevalonate-independent pathway. Proceedings of the National Academy of Sciences of the United States of America 95:2100-2104
Långström B, Lindelöw Å, Schroeder M, Björklund N, Öhrn P (2009) The spruce bark beetle outbreak in Sweden following the January-storms in 2005 and 2007. In: Kunca A, Zubrik M (eds) Proc. IUFRO Working Party 7.03.10, pp 3-19.
Lanne BS, Ivarsson P, Johnsson P, Bergstrom G, Wassgren AB (1989) Biosynthesis of 2-methyl-3-buten-2-ol, a pheromone component of Ips typographus (Coleoptera, Scolytidae) Insect Biochemistry 19:163-167
Laothawornkitkul J, Taylor JE, Paul ND, Hewitt CN (2009) Biogenic volatile organic compounds in the Earth system (vol 183, pg 27, 2009). New Phytologist 184:276-276
Laracinepittet C, Lebreton P (1988) Flavonoid variability within Pinus sylvestris. Phytochemistry 27:2663-2666

63
Lieutier F (2004) Host resistance to bark beetles and its variations. In: Lieutier F, Day KR, Battisti A, Gregoire J-C, Evans HF (eds) Bark and wood boring insects in living trees in Europe, a synthesis. Kluwer Academic Publishers, Dordrecht/Boston/London, pp 135-180
Lieutier F et al. (2003) Intra- and inter-provenance variability in phloem phenols of Picea abies and relationship to a bark beetle-associated fungus. Tree Physiology 23:247-256
Lieutier F, Yart A, Salle A (2009) Stimulation of tree defenses by Ophiostomatoid fungi can explain attack success of bark beetles on conifers. Annals of Forest Science 66:22
Lindström M, Norin T, Birgersson G, Schlyter F (1989) Variation of enantiomeric composition of alpha-pinene in Norway spruce, Picea abies, and its influence on production of verbenol isomers by Ips typographus. . Journal of Chemical Ecology 15:541-548
Lorio PL (1986) Growth-differentiation balance: a basis for understanding southern pine beetle-tree interaction. Forest Ecology and Management 14:259-273
Martin D, Tholl D, Gershenzon J, Bohlmann J (2002) Methyl jasmonate induces traumatic resin ducts, terpenoid resin biosynthesis, and terpenoid accumulation in developing xylem of Norway spruce stems. Plant Physiol 129:1003 - 1018
Martin DM, Fäldt J, Bohlmann J (2004) Functional characterization of nine Norway spruce TPS genes and evolution of gymnosperm terpene synthases of the TPS-d subfamily. Plant Physiology 135:1908-1927
Martin DM, Gershenzon J, Bohlmann J (2003) Induction of volatile terpene biosynthesis and diurnal emission by methyl jasmonate in foliage of Norway spruce. Plant Physiology 132:1586-1599
Millar GJ, Haynes KF (eds) (1998) Methods in chemical ecology. Kluwer Academic publishers, Norwell
Miller B, Madilao LL, Ralph S, Bohlmann J (2005) Insect-induced conifer defense. White pine weevil and methyl jasmonate induce traumatic resinosis, de novo formed volatile emissions, and accumulation of terpenoid synthase and putative octadecanoid pathway transcripts in Sitka spruce. Plant Physiology 137:369-382

64
Miller DR, Borden JH (2000) Dose-dependent and species-specific responses of pine bark beetles (Coleoptera: Scolytidae) to monoterpenes in association with pheromones. Canadian Entomologist 132:183-195
Miller DR, Borden JH, Lindgren BS (1995) Verbenone does dependent interruption of pheromone based attraction of three sympatric species of pine bark beetles (Coleoptera, Scolytidae). Environmental Entomology 24:692-696
Miller DR, Rabaglia RJ (2009) Ethanol and (-)-alpha-pinene: Attractant kairomones for bark and ambrosia beetles in the southeastern US. Journal of Chemical Ecology 35:435-448
Nagy NE, Franceschi VR, Solheim H, Krekling T, Christiansen E (2000) Wound-induced traumatic resin duct development in stems of Norway spruce (Pinaceae): Anatomy and cytochemical traits. American Journal of Botany 87:302-313
Negron JF, McMillin JD, Anhold JA, Coulson D (2009) Bark beetle-caused mortality in a drought-affected ponderosa pine landscape in Arizona, USA. Forest Ecology and Management 257:1353-1362
Nielsen CN, Knudsen MA (2004) Stormstabilitet og sundhed i en rødgransskærm. 7 års resultater efter skærmstillingen. Dansk Skovbrugs Tidsskrift 89:115-128
Nilsson C, Stjernquist I, Barring L, Schlyter P, Jonsson AM, Samuelsson H (2004) Recorded storm damage in Swedish forests 1901-2000. Forest Ecology and Management 199:165-173
Paine TD, Raffa KF, Harrington TC (1997) Interactions among scolytid bark beetles, their associated fungi, and live host conifers. Annual Review of Entomology 42:179-206
Pasquier-Barre F, Palasse C, Goussard F, Auger-Rozenberg MA, Geri C (2001) Relationship of Scots pine clone characteristics and water stress to hatching and larval performance of the sawfly Diprion pini (Hymenoptera : Diprionidae). Environmental Entomology 30:1-6
Persson M, Borg-Karlson A-K, Norin T (1993) Enantiomeric composition of six chiral monoterpene hydrocarbons in different tissues of Picea abies. Phytochemistry 33:303-307
Pettersson EM, Sullivan BT, Anderson P, Berisford CW, Birgersson G (2000) Odor perception in the bark beetle parasitoid Roptrocerus xylophagorum

65
exposed to host associated volatiles. Journal of Chemical Ecology 26:2507-2525
Pettersson M (2007) Stress related emissions of Norway Spruce plants. In: KTH, School of Chemical Science and Engineering (CHE), Chemistry, vol. Licentiate. KTH, Stockholm, p 38
Phillips MA, Croteau RB (1999) Resin-based defenses in conifers. Trends in Plant Science 4:184-190
Phillips MA et al. (2007) Functional identification and differential expression of 1-deoxy-D-xylulose 5-phosphate synthase in induced terpenoid resin formation of Norway spruce (Picea abies). Plant Mol Biol 65:243 - 257
Raffa KF et al. (2008) Cross-scale drivers of natural disturbances prone to anthropogenic amplification: The dynamics of bark beetle eruptions. Bioscience 58:501-517
Raffa KF, Berryman AA (1982a) Accumulation of monoterpenes and associated volatiles following inoculation of grand fir with a fungus transmitted by the fir engraver, Scolytus ventralis (Coleoptera, Scolytidae). Canadian Entomologist 114:797-810
Raffa KF, Berryman AA (1982b) Physiological differences between lodgepole pines resistant and susceptible to the mountain pine beetle (Coleoptera, Scolytidae) and associated moicroorganisms. Environmental Entomology 11:486-492
Raffa KF, Berryman AA (1983a) Physiological aspects of lodgepole pine wound responses to a fungal symbiont of the mountain pine beetle, Dendroctonus ponderosae (Coleoptera, Scolytidea). Canadian Entomologist 115:723-734
Raffa KF, Berryman AA (1983b) The role of host plant resistance in the colonization behavior and ecology of bark beetles (Coleoptera, Scolytidae). Ecological Monographs 53:27-49
Raffa KF, Berryman AA, Simasko J, Teal W, Wong BL (1985) Effects of Grand fir monoterpenes on the fir engraver, Scolytus ventralis (Coleoptera, Scolytidae), and its symbiotic fungus. Environmental Entomology 14:552-556
Raffa KF, Phillips TW, Salom SM (1993) Strategies and mechanisms of host colonization by bark beetles. In: Schowalter T, Filip G (eds) Beetle-pathogen interaction in conifer forests. Academic Press Ltd, pp 103-128

66
Raffa KF, Smalley EB (1995) Interaction of pre-attack and induced monoterpene concentrations in host conifer defense against bark beetle-fungal complexes. Oecologia 102:285-295
Ralph SG et al. (2006) Conifer defence against insects: microarray gene expression profiling of Sitka spruce (Picea sitchensis) induced by mechanical wounding or feeding by spruce budworms (Choristoneura occidentalis) or white pine weevils (Pissodes strobi) reveals large-scale changes of the host transcriptome. Plant Cell and Environment 29:1545-1570
Reddemann J, Schopf R (1996) The importance of monoterpenes in the aggregation of the spruce bark beetle Ips typographus (Coleoptera: Scolytidae: Ipinae). Entomologia Generalis 21:69-80
Reed AN, Hanover JW, Furniss MM (1986) Douglas-fir and western larch: chemical and physical properties in relation to Douglas-fir bark beetle attack. Tree Physiology 1:277-287
Ro DK, Ehlting J, Keeling CI, Lin R, Mattheus N, Bohlmann J (2006) Microarray expression profiling and functional characterization of AtTPS genes: Duplicated Arabidopsis thaliana sesquiterpene synthase genes At4g13280 and At4g13300 encode root-specific and wound-inducible (Z)-gamma-bisabolene synthases. Archives of Biochemistry and Biophysics 448:104-116
Rocchini LA, Lindgren BS, Bennett RG (2000) Effects of resin flow and monoterpene composition on susceptibility of lodgepole pine to attack by the Douglas-fir pitch moth, Synanthedon novaroensis (Lep., Sesiidae). Journal of Applied Entomology-Zeitschrift Fur Angewandte Entomologie 124:87-92
Rocha S, Ramalheira V, Barros A, Delgadillo I, Coimbra MA (2001) Headspace solid phase microextraction (SPME) analysis of flavor compounds in wines. Effect of the matrix volatile composition in the relative response factors in a wine model. Journal of Agricultural and Food Chemistry 49:5142-5151
Rochat D, Ramirez-Lucas P, Malosse C, Aldana R, Kakul T, Morin JP (2000) Role of solid-phase microextraction in the identification of highly volatile pheromones of two Rhinoceros beetles Scapanes australis and

67
Strategus aloeus (Coleoptera, Scarabaeidae, Dynastinae). Journal of Chromatography A 885:433-444
Rouault G, Candau JN, Lieutier F, Nageleisen LM, Martin JC, Warzee N (2006) Effects of drought and heat on forest insect populations in relation to the 2003 drought in Western Europe. Annals of Forest Science 63:613-624
Salle A, Monclus R, Yart A, Lieutier F (2005) Effect of phenolic compounds on the in vitro growth of two fungi associated with Ips typographus. Forest Pathology 35:298-304
Salle A, Ye H, Yart A, Lieutier F (2008) Seasonal water stress and the resistance of Pinus yunnanensis to a bark-beetle-associated fungus. Tree Physiology 28:679-687
Sandnes A, Solheim H (2002) Variation in tree size and resistance to Ceratocystis polonica in a monoclonal stand of Picea abies. Scandinavian Journal of Forest Research 17:522-528
Schlyter F, Birgersson G (1989) Individual variation in bark beetle and moth pheromone: A comparasion and evolutionary background. Holarctic Ecology 12:457-465
Schlyter F, Birgersson G, Byers JA, Lofqvist J, Bergstrom G (1987a) Field response of spruce bark beetle, Ips typographus (Coleoptera, Scolytidae) to aggregation pheromone candidates. Journal of Chemical Ecology 13:701-716
Schlyter F, Birgersson G, Byers JA, Lofqvist J, Bergstrom G (1987b) Field response of spruce bark beetle, Ips typographus, (Coleoptera, Scolytidae) to aggregation pheromone candidates. Journal of Chemical Ecology 13:701-716
Schlyter F, Lofqvist J, Byers JA (1987c) Behavioral sequence in the attraction of the bark beetle Ips typographus to pheromone sources. Physiological Entomology 12:185-196
Schroeder LM (2007) Escape in space from enemies: a comparison between stands with and without enhanced densities of the spruce bark beetle. Agricultural and Forest Entomology 9:85-91
Seybold SJ, Bohlmann J, Raffa KF (2000) Biosynthesis of coniferophagous bark beetle pheromones and conifer isoprenoids: Evolutionary perspective and synthesis. Canadian Entomologist 132:697-753

68
Seybold SJ, Huber DPW, Lee JC, Graves AD, Bohlmann J (2006) Pine monoterpenes and pine bark beetles: a marriage of convenience for defense and chemical communication. Phytochemistry Review 5:143-178
Simth RH (1965) Effect of monoterpene vapors on the westem pine beetle. Journal of economic entomology 58:509-510
Six DL, Wingfield MJ (2011) The role of phytopathogenicity in bark beetle-fungal symbioses: A challenge to the classic paradigm. Annual Review of Entomology 56
Smith JL, De Moraes CM, Mescher MC (2009) Jasmonate- and salicylate-mediated plant defense responses to insect herbivores, pathogens and parasitic plants. Pest Management Science 65:497-503
Smith RH (1965) Effect of monoterpene vapors on the westem pine beetle. Journal of economic entomology 58
Solheim H (1991) Oxygen deficiency and spruce resin inhibition of growth of blue stain fungal associated with Ips typographus. Mycological Research 95:1387-1392
Tao ZN, Jain AK (2005) Modeling of global biogenic emissions for key indirect greenhouse gases and their response to atmospheric CO2 increases and changes in land cover and climate. Journal of Geophysical Research-Atmospheres 110
Tholl D, Boland W, Hansel A, Loreto F, Rose USR, Schnitzler JP (2006) Practical approaches to plant volatile analysis. Plant Journal 45:540-560
Turner JG, Ellis C, Devoto A (2002) The jasmonate signal pathway. Plant Cell 14:S153-S164
Vas G, Vekey K (2004) Solid-phase microextraction: a powerful sample preparation tool prior to mass spectrometric analysis. Journal of Mass Spectrometry 39:233-254
Viiri H, Annila E, Kitunen V, Niemela P (2001) Induced responses in stilbenes and terpenes in fertilized Norway spruce after inoculation with blue-stain fungus, Ceratocystis polonica. Trees-Structure and Function 15:112-122
Villaverde ML, Juarez MP, Mijailovsky S (2007) Detection of Tribolium castaneum (Herbst) volatile defensive secretions by solid phase micro extraction-capillary gas chromatography (SPME-CGC). Journal of Stored Products Research 43:540-545

69
Wallin KF, Raffa KF (2000a) Influences of host chemicals and internal physiology on the multiple steps of post landing host acceptance behavior of Ips pini (Coleoptera: Scolytidae). Environmental Entomology 29:442-453
Wallin KF, Raffa KF (2000b) Influences of host chemicals and internal physiology on the multiple steps of postlanding host acceptance behavior of Ips pini (Coleoptera : Scolytidae). Environmental Entomology 29:442-453
Wermelinger B (2004) Ecology and management of the spruce bark beetle Ips typographus: a review of recent research. Forest Ecology and Management 202:67-82
Worrell R (1983) Damage by the spruce bark beetle in south Norway 1970–80: a survey, and factors affecting its occurrence. Medd Nor Skogforsoeksves 38:1-34
Yassaa N, Williams J (2005) Analysis of enantiomeric and non-enantiomeric monoterpenes in plant emissions using portable dynamic air sampling/solid-phase microextraction (PDAS-SPME) and chiral gas chromatography/mass spectrometry. Atmospheric Environment 39:4875-4884
Yassaa N, Williams J (2007) Enantiomeric monoterpene emissions from natural and damaged Scots pine in a boreal coniferous forest measured using solid-phase microextraction and gas chromatography/mass spectrometry. Journal of Chromatography A 1141:138-144
Zeneli G, Krokene P, Christiansen E, Krekling T, Gershenzon J (2006) Methyl jasmonate treatment of mature Norway spruce (Picea abies) trees increases the accumulation of terpenoid resin components and protects against infection by Ceratocystis polonica, a bark beetle-associated fungus. Tree Physiology 26:977-988
Zhang QH, Schlyter F (2004) Olfactory recognition and behavioral avoidance of angiosperm nonhost volatiles by conifer-inhabiting bark beetles. Agricultural and Forest Entomology 6:1-19
Zulak KG et al. (2009) Targeted proteomics using selected reaction monitoring reveals the induction of specific terpene synthases in a multi-level study of methyl jasmonate-treated Norway spruce (Picea abies). Plant J 60:1015 - 1030

70

71
Author’s Contributions
The following is a description of my contribution to Publications included in
the thesis (I to IV), as requested by KTH.
Paper I: I performed chemical parts of the experiments and wrote most of the
manuscript.
Paper II: I performed all the chemical experiments, and wrote most of the
manuscript.
Paper III: I performed the chemical experiments and wrote the manuscript.
Paper IV: I performed the chemical experiments and wrote the manuscript.

72
Appendix B1.
Relative amounts of terpene and phenol hydrocarbons in stem bark of Norway spruce trees with no apparent damage, short-leader or leaning-stem, 17 months after the storm “Gudrun”. Data are collected from the trees with fungal inoculation (n = 5) and presented as untransformed means ± 1 SE. Means followed by different letters in a row
are significantly different by the LSD test at p = 0.05.
Compounds No apparent damage Short-leader Leaning-stem
Terpenoid compounds (-)-α-Pinene 14.15 ± 0.68 21.28 ± 2.42 17.84 ± 1.44 (+)-α-Pinene 11.22 ± 1.35 14.24 ± 1.59 12.48 ± 1.83 β-Pinene 35.31 ± 3.57 31.38 ± 3.61 31.16 ± 1.78 Sabinene 0.87 ± 0.32 0.41 ± 0.17 0.21 ± 0.14 (+)-3-Carene 1.29 ± 0.32 a 0.31 ± 0.15 b 0.36 ± 0.15 b Myrcene 2.05 ± 0.26 2.04 ± 0.26 1.42 ± 0.46 (-)-Limonene 0.84 ± 0.10 1.57 ± 0.53 1.02 ± 0.20 (+)-Limonene 0.69 ± 0.06 1.01 ± 0.32 0.71 ± 0.11 β-Phellandrene 7.66 ± 1.24 5.99 ± 0.71 6.71 ± 1.62 Terpinolene 0.27± 0.08 0.31 ± 0.09 0.17 ± 0.08 α-Longipinene 0.43 ± 0.18 0.46 ± 0.22 0.45 ± 0.12 α-Gurjunene 0.72 ± 0.32 0.60 ± 0.20 0.52 ± 0.30 Longifolene 0.96 ± 0.35 0.68 ± 0.34 1.65 ± 0.71 (E)-β-Caryophyllene 0.30 ± 0.12 0.52 ± 0.39 0.93 ± 0.28 (-)-Germacrene D 3.80 ± 1.12 1.49 ± 0.66 1.56 ± 0.55 δ-Cadinene 1.12 ± 0.39 1.67 ± 0.56 0.85 ± 0.40 Germacrene D-4-ol 1.82 ± 0.78 a 5.40 ± 2.21 b 2.30 ± 0.55 a Thunbergol 3.75 ± 1.60 2.29 ± 1.08 3.14 ± 1.45 Abienol 6.33 ± 1.72 5.97 ± 3.30 9.61 ± 0.78 Others 4.05 ± 1.48 3.18 ± 0.93 2.34 ± 1.37 Phenolic compounds Catechin 7.34 ± 30.5 13.76 ± 7.61 3.82 ± 0.92 Unknown flavone 2.63 ± 1.75 2.74 ± 0.39 3.27 ± 0.72 Astringin 1.76 ± 0.41 3.15 ± 0.56 4.59 ± 2.99 Taxifolin glycoside 3.55 ± 1.45 a 16.51 ± 1.33 b 26.76 ± 9.01 b Piceid 14.53 ± 4.38 6.76 ± 0.37 8.28 ± 1.59 Taxifolin 5.01 ± 0.54 5.34 ± 0.85 4.74 ± 1.37 Isorhapontin 3.04 ± 0.30 2.65 ± 0.32 2.95 ± 0.41 Unknown stilbene 10.38 ± 1.56 a 3.03 ± 0.59 b 3.66 ± 0.13 b Unknown stilbene 11.72 ± 4.76 a 2.94 ± 0.46 b 3.97 ± 0.68 b Resveratrol 33.70 ± 4.61 32.91 ± 1.39 31.98 ± 5.42 Isorhapontinen 6.27 ± 1.20 10.21 ± 2.18 5.98 ± 1.69
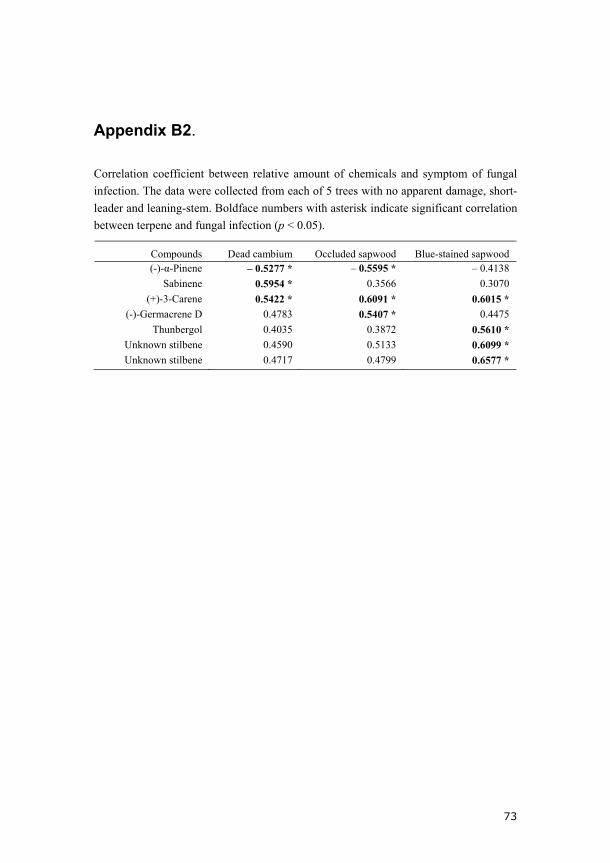
73
Appendix B2. Correlation coefficient between relative amount of chemicals and symptom of fungal infection. The data were collected from each of 5 trees with no apparent damage, short-leader and leaning-stem. Boldface numbers with asterisk indicate significant correlation between terpene and fungal infection (p < 0.05).
Compounds Dead cambium Occluded sapwood Blue-stained sapwood (-)-α-Pinene – 0.5277 * – 0.5595 * – 0.4138
Sabinene 0.5954 * 0.3566 0.3070 (+)-3-Carene 0.5422 * 0.6091 * 0.6015 *
(-)-Germacrene D 0.4783 0.5407 * 0.4475 Thunbergol 0.4035 0.3872 0.5610 *
Unknown stilbene 0.4590 0.5133 0.6099 * Unknown stilbene 0.4717 0.4799 0.6577 *

74
Appendix C Relative amounts of terpenes (normalized to 100%) in intact and wounded phloem of control logs (CT) and methyl jasmonate (MeJA) treated Norway spruce logs. Wounded phloem was sampled 24 hr after intensive wounding using a pushpin. Data is presented as means ± SE. n = 3, except for wounded phloem in MeJA logs where n = 5.
Compounds Intact phloem Wounded phloem
CT MeJA CT MeJA 1 (-)-α-Pinene 16.62 ± 2.79 a 14.32 ± 2.32a 15.01 ± 2.83 a 7.86 ± 0.55 b 2 (+)-α-Pinene 12.83 ± 2.18 a 11.86 ± 1.98a 11.17 ± 2.12 a 7.18 ± 0.62 b 3 Camphene 0.53 ± 0.20 ab 0.25 ± 0.16a 0.99 ± 0.59 b 0.16 ± 0.01 a 4 β-Pinene 22.87 ± 1.54 a 21.60 ± 2.64ab 20.52 ± 4.63 ab 17.00 ± 0.88 b 5 Sabinene 0.62 ± 0.26 a 0.59 ± 0.33a 0.20 ± 0.06 b 0.19 ± 0.01 b 6 (+)-3-Carene 0.31 ± 0.10 0.54 ± 0.14 0.42 ± 0.06 0.48 ± 0.02 7 Myrcene 2.46 ± 0.10 2.14 ± 0.40 1.66 ± 0.44 1.26 ± 0.10 8 Limonene 1.58 ± 0.26 1.63 ± 0.27 1.52 ± 0.37 0.88 ± 0.11 9 β-Phellandrene 9.51 ± 0.79 8.85 ± 1.67 7.37 ± 2.02 6.13 ± 0.36
10 Terpinelene 0.30 ± 0.06 0.16 ± 0.03 0.19 ± 0.02 0.19 ± 0.01 11 1,8-Cineole 0.35 ± 0.12 0.52 ± 0.14 0.49 ± 0.13 0.38 ± 0.09 12 Bornyl acetate 0.11 ± 0.08 0.15 ± 0.08 0.24 ± 0.11 0.12 ± 0.05 13 Other monoterpenes 0.51 ± 0.07 0.56 ± 0.08 0.47 ± 0.08 0.35 ± 0.03 14 α-Longipinene 0.17 ± 0.01 0.20 ± 0.05 0.20 ± 0.09 0.18 ± 0.01 15 α-Gurjunene 0.73 ± 0.07 1.03 ± 0.37 1.07 ± 0.39 0.73 ± 0.03 16 Longifolene 0.17 ± 0.11 0.37 ± 0.12 0.48 ± 0.14 0.54 ± 0.05 17 Bergamotene 0.40 ± 0.11 0.16 ± 0.08 0.27 ± 0.06 0.19 ± 0.02 18 (-)-Germacrene D 3.82 ± 0.76 3.39 ± 0.27 2.97 ± 0.54 2.61 ± 0.22 19 δ-Cadinene 1.27 ± 0.10 1.96 ± 0.70 1.80 ± 0.80 1.32 ± 0.08 20 Germacrene D 4-ol 0.75 ± 0.12 0.51 ± 0.11 0.63 ± 0.22 0.79 ± 0.04 21 Cubenol 0.40 ± 0.06 0.53 ± 0.33 0.64 ± 0.35 0.41 ± 0.02 22 Other sesquiterpenes 2.48 ± 0.23 3.13 ± 0.55 3.16 ± 0.10 3.26 ± 0.15 23 Thunbergene 0.73 ± 0.30 1.17 ± 0.17 1.05 ± 0.37 1.47 ± 0.11 24 Thunbergol 8.31 ± 2.91 a 10.18 ± 2.60a 9.78 ± 2.84 a 17.71 ± 0.81 b 25 Abienol 2.22 ± 0.31 a 2.89 ± 0.23a 6.19 ± 2.12 b 11.89 ± 0.91 c 26 Neoabienol 1.14 ± 0.07 1.80 ± 0.26 1.54 ± 0.08 2.50 ± 0.24 27 Unknown diterpene 1.28 ± 0.18 1.79 ± 1.28 0.98 ± 0.52 1.99 ± 0.37 28 Unknown diterpene 1.37 ± 0.18 a 1.22 ± 0.17a 1.08 ± 0.09 a 1.95 ± 0.16 b 29 Abietadienal 0.26 ± 0.13 a 0.63 ± 0.09ab 0.76 ± 0.20 ab 0.97 ± 0.10 b 30 Dehydroabietal 0.29 ± 0.12 a 0.43 ± 0.28a 0.82 ± 0.33 ab 1.20 ± 0.15 b 31 Methyl dehydroabieta 0.18 ± 0.12 a 0.22 ± 0.11a 0.49 ± 0.31 ab 0.73 ± 0.05 b 32 Other diterpenes 7.46 ± 2.32 7.59 ± 0.68 5.64 ± 0.39 8.10 ± 0.46
Means followed by different letters in a row are significantly different at P < 0.05, based on LSD test following ANOVA.
Boldface: compounds with statistical significant treatment differences.

75
Appendix D Absolute amounts of major terpene hydrocarbons (µg g -1 dry wt equivalent to pentadecane) in Norway spruce phloem before (day 0) and after (day 30) stem treatment with
methyl jasmonate (MJ) or inoculation with the bluestain fungus Ceratocystis polonica (CT are untreated control trees). Samples were taken at 1.3 m stem height within the treated stem section (TS), but on day 30 an additional sample was taken 0.3 m below the treated stem section (UTS) at 0.5 m height. Data are expressed as means ± 1 SD, n = 16 trees.
Compound RI 2
C. polonica MJ CT
Day 0 Day 30
Day 0 Day 30
Day 0 Day 30
TS UTS TS UTS TS UTS
Tricyclene 918 2.0 ± 0.3 2.1 ± 0.3 2.2 ± 0.2 2.1 ± 0.4 2.3 ± 0.3 2.1 ± 0.2 2.5 ± 0.5 2.7 ± 0.2 2.0 ± 0.4
α-Thujene 924 1.2 ± 0.2 1.5 ± 0.5 0.4 ± 0.2 0.9 ± 0.2 0.8 ± 0.5 0.4 ± 0.2 1.1 ± 0.2 1.2 ± 0.6 0.6 ± 0.3
α-Pinene 933 436.6 ± 44.4 370.7 ± 63.3 466.3 ± 36.4 438.2 ± 22.0 537.3 ± 37.7 420.1 ± 22.0 507.8 ± 36.1 454.4 ± 30.0 484.1 ± 82.6
Camphene 943 8.7 ± 0.9 7.0 ± 1.7 8.1 ± 0.7 9.8 ± 2.4 9.2 ± 1.9 7.7 ± 0.6 11.2 ± 2.8 9.2 ± 2.0 7.4 ± 1.6
Sabinene 967 6.6 ± 0.6 15.6 ± 1.0 5.3 ± 0.2 6.9 ± 0.7 6.3 ± 0.5 6.4 ± 0.3 5.0 ± 0.8 6.3 ± 1.2 4.3 ± 1.6
β-Pinene 971 480.8 ± 53.3 516.3 ± 48.8 422.3 ± 43.1 494.2 ± 25.9 707.1 ± 96.3 422.3 ± 51.8 424.9 ± 54.2 425.0 ± 58.3 434.2 ± 95.0
Myrcene 985 26.3 ± 2.6 34.3 ± 2.8 28.6 ± 1.7 32.2 ± 2.6 41.7 ± 10.4 24.6 ± 2.4 26.2 ± 3.5 28.9 ± 4.3 22.0 ± 4.0
(+)-3-Carene 1002 26.0 ± 12.1 166.7 ± 37.4 11.7 ± 4.5 17.8 ± 15.5 23.5 ± 9.1 11.5 ± 5.9 8.9 ± 2.2 18.2 ± 17.8 5.6 ± 3.8
β-Phellandrene 1022 88.6 ± 9.9 104.4 ± 10.9 99.9 ± 8.1 99.6 ± 11.4 152.4 ± 30.9 76.8 ± 8.8 84.8 ± 19.1 88.0 ± 15.7 71.3 ± 12.3
Limonene 1030 24.4 ± 2.8 28.2 ± 8.3 19.0 ± 2.1 29.9 ± 8.2 44.8 ± 12.5 26.3 ± 11.6 33.3 ± 3.1 33.5 ± 2.5 21.9 ± 5.4
α-Terpinolene 1079 5.5 ± 1.3 20.7 ± 2.8 5.2 ± 0.5 6.2 ± 1.6 7.8 ± 1.1 5.7 ± 0.7 4.7 ± 0.3 5.0 ± 0.8 4.0 ± 0.6
Bornyl acetate 1268 1.6 ± 0.7 a
2.5 ± 3.3 1.8 ± 1.3 3.4 ± 1.7 b
4.1 ± 2.5 4.3 ± 1.8 1.6 ± 0.9 a
1.6 ± 1.4 2.3 ± 1.2
α-Longipinene 1347 5.1 ± 1.5 3.5 ± 0.8 3.1 ± 0.6 6.6 ± 0.8 7.0 ±0.4 3.4 ± 0.2 4.7 ± 2.7 4.9 ± 1.4 2.9 ± 1.4
Longifolene 1396 16.2 ± 3.3 14.6 ± 3.7 9.5 ± 1.2 8.5 ± 8.3 13.4 ± 7.4 4.3 ± 3.5 8.2 ± 6.7 8.2 ± 5.7 5.0 ± 3.6
(E)-β-Caryophyllene 1411 12.2 ± 6.8 13.2 ± 6.7 9.1 ± 3.7 5.7 ± 3.6 7.3 ± 4.1 3.8 ± 2.5 10.5 ± 2.0 8.9 ± 2.5 6.9 ± 1.9
(-)-Germacrene D 1473 15.2 ± 15.2 13.6 ± 14.1 9.2 ± 5.9 31.8 ± 15.2 36.5 ± 19.8 10.6 ± 6.5 13.6 ± 12.4 10.3 ± 8.2 8.1 ± 7.1
δ- Cadinene 1506 9.1 ± 1.0 6.4 ± 1.8 3.0 ± 1.3 12.6 ± 3.5 12.3 ± 8.0 3.0 ± 1.7 11.7 ± 8.9 9.1 ± 6.9 3.6 ± 2.2
Germacrene D-4-ol 1562 11.2 ± 4.7 11.0 ± 8.3 5.2 ± 2.1 23.2 ± 18.8 36.5 ± 33.5 8.4 ± 5.4 7.1 ± 3.4 11.3 ± 11.1 6.8 ± 3.3
Thunbergol 2037 56.2 ± 24.2 102.6 ± 35.1 41.9 ± 14.6 21.9 ± 15.7 33.3 ± 33.1 23.4 ± 14.1 45.3 ± 39.2 38.7 ± 23.1 18.6 ± 8.9
Neoabienol 2123 98.1 ± 23.2 104.6 ± 24.3 94.7 ± 6.9 102.0± 17.1 151.3± 16.5 91.6 ± 7.9 91.4 ± 38.1 107.5± 10.2 93.9± 11.9
Abienol 2126 48.5 ± 4.9 43.6 ± 10.7 40.0 ± 2.7 56.0 ± 6.3 61.5 ± 6.1 38.1 ± 2.6 57.9 ± 11.8 49.1 ± 4.9 36.2 ± 4.5
Total 1500.8 ± 120.5 1789.4 ± 143.3 1545.9 ± 89.5 1549.2 ± 86.7 2070.2 ± 216.5 1327.6 ± 57.3 1517.3 ± 165.7 1452.3 ± 131.3 1338.8 ± 127.6
Boldface numbers indicate significant differences between day 0 and day 30 within treatments; different letters indicate significant differences between treatments on day 0 (p < 0.05).