conservation of igfbp structure during evolution: cloning of chicken insulin-like growth factor...
TRANSCRIPT

Pergamon
Progress in Growth Factor Research, Vol. 6. Nos. 2-4, pp. 159-165, 1995 Copyright © 1996 Elsevier Science Ltd. All rights reserved
Printed in Great Britain 0955--2235/95 $29.00 + .00
PII: S0955-2235(96)00011-7
CONSERVATION OF IGFBP STRUCTURE DURING EVOLUTION: CLONING OF CHICKEN
INSULIN-LIKE GROWTH FACTOR BINDING PROTEIN-5
Susanne V. Allander,* Ewa Ehrenborg,* Holger Luthman* and David R. Powellt
*Department of Molecular Medicine, Karolinska Institute, S-171 76 Stockholm, Sweden tDepartment of Pediatrics, Baylor College of Medicine, Houston, TX 77030, U.S.A.
The insulin-like growth factor binding proteins (IGFBPs) have conserved charac- teristics o f their genomic organization, including similar locations o f exon borders rela- tive to nucleotides encoding conserved cysteine residues. Furthermore, the human IGFBP genes, as well as the human homeobox (HOX) genes, are localized to chro- mosomes 2, 7, 12, and 17. Although little is known about the evolution of the IGFBP genes, the association of human IGFBP and homeobox ( HOX) genes at four chromo- somal loci may indicate that their ancestral genes were linked prior to the first dupli- cation o f chromosomal DNA containing the ancestral H O X cluster. The hypothesis that IGFBPs are ancient proteins is supported by the reported detection of IGFBP activity in serum from the Agnathan species, Geotria australis, a primitive vertebrate.
Further studies o f IGFBPs in different species are needed to understand the evolu- tion of this protein/gene family. Chicken provides a good intermediate model, since birds diverged from mammals ~300 million years ago. A complementary DNA (cDNA) clone encoding chicken insulin-like growth factor binding protein-5 (clGFBP-5) was isolated. The deduced amino acid sequence is 83% identical to human IGFBP-5 and encodes a mature polypeptide o f 251 amino acids. The conservation of IGFBP-5 primary structure across vertebrate species suggests maintenance of impor- tant functions during evolution.
Keywords: Gene structure, chromosomal localization, homeobox genes, evolution, insulin-like growth factors, chicken, complementary DNA.
INTRODUCTION
Insul in- l ike growth fac tor b ind ing pro te ins ( I G F B P s ) cons t i tu te a separa te p ro te in family with six ident if ied m e m b e r s [1-3], which b ind insulin-l ike growth
Correspondence to: Dr S. V. Allander. Tel. +46 (8) 729-2096; Fax: +46 (8) 32 77 34. Acknowledgements--This work was supported by Caroline Weiss Law Karolinska-Baylor Research
Fellowship, "F6renade Liv" Mutual Group Life Insurance Company, the Swedish Medical Research Council (9109), and National Institute of Health Grant R01-DK38773.
159

160 S. V. Allander et al.
factors (IGFs) with high affinity and thereby regulate the mitogenic and metabolic activities of IGFs [4]. The six IGFBPs have clearly distinct amino acid sequences, but share regions of homology including cysteine-rich sequences in their N- and C- terminal regions [5]. The diversity within this gene family may have evolved through duplication of an ancestral gene and subsequent transposition to other chromo- somes. IGFBP-5 is the most conserved of the mammalian IGFBPs and a short cDNA fragment from Xenopus suggests a high degree of conservation also in non- mammalian vertebrates [6]. The highly conserved nature of IGFBP-5 makes it a good candidate for initial studies of IGFBP structure and function in non- mammalian species.
RESULTS AND DISCUSSION
Gene Structure and Chromosomal Localization of Human IGFBP Genes
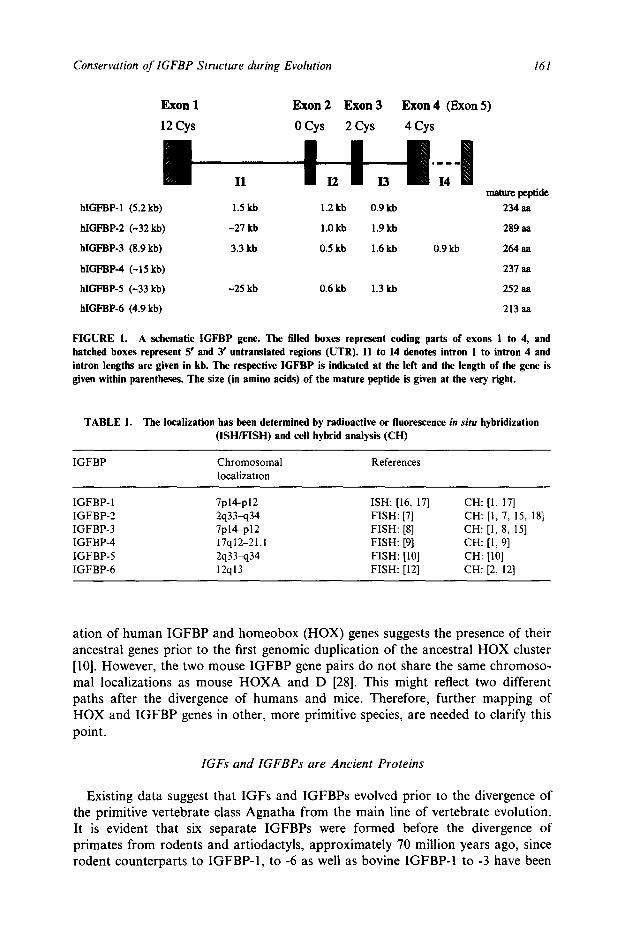
Cosmid clones encompassing the genes for human IGFBP-I to -6 were isolated from a placental cosmid library [7-12]. The authenticity of the cosmid clones were ascertained by further characterization of the genomic organization and DNA sequence of the respective IGFBP gene. The IGFBP genes have their coding parts divided into four exons (Fig. 1), an arrangement now recognized in all members of the human IGFBP family [7, 10-15]. The hlGFBP-3 gene has an additional fifth 3' untranslated exon [14]. The IGFBP genes also have similar locations of exon borders relative to nucleotides encoding conserved cysteine residues [7, 12]. The locations of the 18 conserved cysteine residues relative to exons 1 to 4, is shown in Fig. 1. These cysteines are present in all human IGFBPs, except for IGFBP-6, which is lacking two cysteines in exon 1 [2, 5, 12].
Fluorescence in situ hybridization (FISH) and subsequent quinacrine (Q)-banding localized the hlGFBP-1 to -6 genes to chromosomes 7, 2, 7, 17, 2, and 12, respec- tively [7-10, 12]. The chromosomal assignments determined by FISH analysis were confirmed by PCR amplifications of somatic cell hybrids with gene specific primers. Table 1 summarizes the reported localizations of hlGFBP-1 to -6.
Association Between Human Homeobox (HOX) and IGFBP Genes
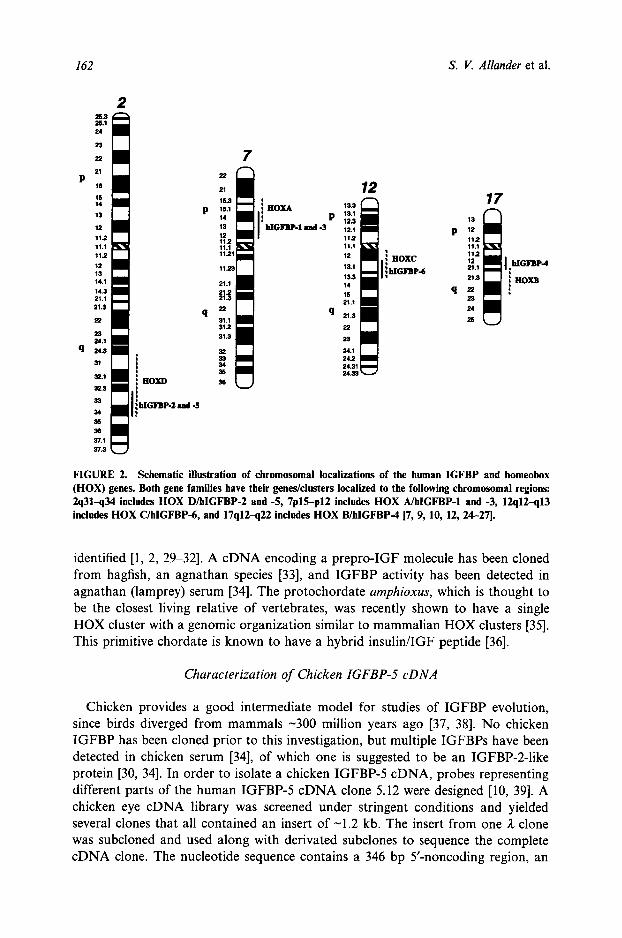
The vertebrate HOX genes are suggested to be true homologs to the insect homeotic gene complexes and to have arisen from duplications of a single ancestral gene cluster [19-22]. The human homeobox (HOX) gene family consists of four gene clusters, HOX A-D, localized to chromosomes 2, 7, 12, and 17 [23-27] (Fig. 2). As shown in Fig. 2, the HOX and IGFBP genes/clusters are localized to the same chromosomal regions. Further, the human IGFBP-2/-5 and IGFBP-1/-3 gene pairs are closely linked, with opposite transcriptional orientations [8, 10]. Similarly, the two mouse IGFBP gene pairs (IGFBP-2/-5) and IGFBP-1/-3) are also closely linked [28]. IGFBP-3 and -5 are more closely related to each other than to other IGFBPs, and IGFBP-1 and -2 are more closely related to each other than to IGFBP-3 or -5, which suggests that duplication of an ancient IGFBP gene preceded dispersion of this gene pair to multiple chromosomal loci. Thus, the evolution of the HOX and IGFBP gene families appear to follow a similar pattern. The associ-
Conservation of IGFBP Structure during Evolution 161
E x o n 1 E x o n 2 E x o n 3 E x o n 4 (Exon 5)
12 Cys 0 Cys 2 Cys 4 Cys
I! =-| l I1
mature peptide hlGFBP-I (5.2 kh) 1.5 kb 1.2 kh 0.9 kb 234 aa
hIGFBP-2 (-32 kb) -27 kb 1.0 kb 1.9 kb 289 aa
hIGFBP-3 (8.9 kb) 3.3 kb 0.5 kb 1.6 kb 0.9 kb 264 aa
hIGFBP-4 (-15 kb) 237 aa
hlGFBP-5 (-33 kb) -25 kb 0.6 kb 1.3 kb 252 aa
hlGFBP-6 (4.9 kb) 213 aa
FIGURE 1. A schematic IGFBP gene. The filled boxes represent coding parts of exons 1 to 4, and hatched boxes represent 5" and 3' untranslated regions (UTR). I1 to 14 denotes intron 1 to intron 4 and intron lengths are given in kb. The respective IGFBP is indicated at the left and the length of the gene is given within parentheses. The size (in amino acids) of the mature peptide is given at the very right.
TABLE 1. The localization has been determined by radioactive or fluorescence in situ hybridization (ISH/FISH) and cell hybrid analysis (CH)
IGFBP Chromosomal References localization
IGFBP-I 7p14-p12 ISH: [16, 17] CH: [1, 17] IGFBP-2 2q33-q34 FISH: [7] CH: [1, 7, 15, 18] IGFBP-3 7p14-p12 FISH: [8] CH: [1, 8, 15] IGFBP-4 17q12-21.1 FISH: [9] CH: [1, 9] IGFBP-5 2q33~134 FISH: [10] CH: [10] IGFBP-6 12q13 FISH: [12] CH: [2, 12]
a t ion of h u m a n I G F B P and h o m e o b o x ( H O X ) genes suggests the presence of their ancestral genes pr ior to the first genomic dupl ica t ion o f the ancestral H O X cluster [10]. However , the two mouse I G F B P gene pairs do not share the same ch romoso- mal localizations as mouse H O X A and D [28]. This might reflect two different pa ths af ter the divergence of humans and mice. Therefore , fur ther m a p p i n g o f H O X and I G F B P genes in other, more primit ive species, are needed to clarify this point .
IGFs and IGFBPs are Ancient Proteins
Existing da ta suggest tha t I G F s and I G F B P s evolved pr ior to the divergence of the primit ive ver tebra te class Agna tha f rom the ma in line of ver tebra te evolution. I t is evident that six separa te I G F B P s were fo rmed before the divergence o f p r imates f rom rodents and art iodactyls , approx ima te ly 70 mill ion years ago, since rodent counte rpar t s to IGFBP-1 , to -6 as well as bovine I G F B P - I to -3 have been
162 S. V. A/lander et al.
2 ~,x.3 23.1 24
23
23
21
13
IS 14
13
12
11.2 11.1 11.2
12 13 14.1 14.3 21.1 21.3
23
23 24.1
24.3
31
32.1
32.3
33
34 36 33 37.1 37.3
HOXD
hlGFBP-2 sad -$
7 22
21
15.3 p 15.1
14 13 12 112 11.1 11.21
31.1 21.2 21.3
q 22 31.1 31.2 31.3
32 33 34 35
3e
t : lloxA 13.3
p 13.1 12.3 12.1 11.2 11,1 12
13,1
13.3 14 16 21.1
q 21.3
22
23
24.1 24.2 24.31 ! 24.33
12
H O X C
hIGM'BP..6
13
12 11.2 11.1 112 12 21.1
21.3
22 23
24
25
17
, h lGFBP-4
H O X B
FIGURE 2. Schematic illustration of chromosomal localizations of the human IGFBP and homeobox (HOX) genes. Both gene families have their genes/clusters localized to the following chromosomal regions: 2q31-q34 includes HOX D/hlGFBP-2 and -5, 7plS-p12 includes HOX A/hlGFBP-1 and -3, 12q12-q13 includes HOX C/hlGFBP-6, and 17q12-q22 includes HOX B/hlGFBP-4 [7, 9, 10, 12, 24-27].
identified [1, 2, 29-32]. A cDNA encoding a prepro-IGF molecule has been cloned from hagfish, an agnathan species [33], and IGFBP activity has been detected in agnathan (lamprey) serum [34]. The protochordate amphioxus, which is thought to be the closest living relative of vertebrates, was recently shown to have a single HOX cluster with a genomic organization similar to mammalian HOX clusters [35]. This primitive chordate is known to have a hybrid insulin/IGF peptide [36].
Characterization of Chicken IGFBP-5 cDNA
Chicken provides a good intermediate model for studies of IGFBP evolution, since birds diverged from mammals ~300 million years ago [37, 38]. No chicken IGFBP has been cloned prior to this investigation, but multiple IGFBPs have been detected in chicken serum [34], of which one is suggested to be an IGFBP-2-1ike protein [30, 34]. In order to isolate a chicken IGFBP-5 cDNA, probes representing different parts of the human IGFBP-5 cDNA clone 5.12 were designed [10, 39]. A chicken eye cDNA library was screened under stringent conditions and yielded several clones that all contained an insert of ~1.2 kb. The insert from one ~ clone was subcloned and used along with derivated subclones to sequence the complete cDNA clone. The nucleotide sequence contains a 346 bp 5"-noncoding region, an

Conservation of IGFBP Structure during Evolution 163
C h L o k e n ZGIPI3P- S
R a t w _ w
]li3~lllQ w _ w Ze:no~8 m _ •
i ¢t * Q It. ~ Q Q *
L~FVQCBPCD~KLSLCPPPPLQC~L KGQPC~r~T . . . . . H . . . . . II . . . . M . . . 8 . . . . . . . . . . . . . . . II . . . . J U I . . 8 . . . . . . . . . . H . . . . . 31 . . . . M . . . 8 . . . . . . . . . . . . . . . N . . . . K l l . . 8 . . . . .
. . . . . H . . . . . I I . . . . M . . . 8 . . . . . . . . . . . . . . . N . . . . J U I . . 8 . . . . .
• . D . V ] I . . . . M . . . 8 . . . . . . . . . . . H R . . N . . . l i K Z X . 8 . . . . !
FIGURE 3. Comparison of IGFBP-5 amino acid sequences among different vertebrates. The deduced amino acid sequence for chicken IGFBP-5 is compared with the reported sequences for human [39], rat [1], mouse, and Xenopus [6] IGFBP-5, in the region of amino acids 7 to 51.
810 bp open read ing frame, and 80 bp o f 3' un t rans la ted sequence. The chicken
I G F B P - 5 sequence codes for a ma tu re pep t ide o f 251 a m i n o acids, lacking one amino acid as c o m p a r e d with m a m m a l i a n I G F B P - 5 sequences [1, 6, 39]. The a m i n o acid subst i tu t ions in the chicken sequence, as c o m p a r e d with m a m m a l i a n I G F B P - 5 sequences, are scat tered t h roughou t the coding region, with pockets o f highly
conserved sequence which may suggest i m p o r t a n t funct ional regions. C o m p a r i s o n to human , rat , and mouse amino acid sequences show an overal l amino acid s imilar i ty o f 82-83% between chicken and either human, rat, or mouse I G F B P - 5 sequences. In a stretch o f a m i n o acid 7 to 51 (Fig. 3) bo th chicken and Xenopus sequences have seven amino-ac id subst i tu t ions c o m p a r e d with the m a m m a l i a n sequences, whereas human , ra t and mouse sequences are 100% conserved in this region. W e suggest tha t the isola ted c D N A clone encodes the chicken coun te rpa r t to I G F B P - 5 .
R E F E R E N C E S
1. Shimasaki S, Shimonaka M, Zhang H-P, Ling N. Identification of five different insulin-like growth factor binding proteins (IGFBPs) from adult rat serum and molecular cloning of a novel IGFBP- 5 in rat and human. J Biol Chem. 1991; 266: 10646-10653.
2. Shimasaki S, Gao L, Shimonaka M, Ling M. Isolation and molecular cloning of insulin-like growth factor binding protein 6. Mol Endocrinol. 1991; 5: 938-948.
3. Drop SLS. Report on the nomenclature of the IGF binding proteins. Endocrinology. 1992; 130: 1736-1737.
4. Sara VR, Hall K. Insulin-like growth factors and their binding proteins. Physiol Rev. 1990; 70: 591~14.
5. Drop SLS, Schuller AGP, Lidenbergh-Kortleve D J, Groffen C, Brinkman A, Zwarthoff EC. Structural aspects of the IGFBP family. Growth Reg. 1992; 2: 69-79.
6. James PL, Jones SB, Busby WH, Clemmons DR, Rotwein P. A highly conserved insulin-like growth factor-binding protein (IGFBP-5) is expressed during myoblast differentiation. J Biol Chem. 1993; 268: 22305-22312.
7. Ehrenborg E, Vilhelmsdotter S, Bajalica S, Larsson C, Stern I, Koch J, Brondum-Nielson K, Luthman H. Structure and localization of the human insulin-like growth factor-binding protein 2 gene. Biochem Biophys Res Commun. 1991 ; 176:1250-1255.
8. Ehrenborg E, Larsson C, Stern I, Janson M, Powell DR, Luthman H. Contiguous localization of the genes encoding human insulin-like growth factor binding proteins 1 and 3 on chromosome 7. Genomics. 1992; 12: 497-502.
9. Bajalica S, Allander SV, Ehrenborg E, Brondum-Nielsen K, Luthman H, Larsson C. Localization of the human-insulin like growth-factor binding protein 4 gene to chromosomal region 17q12-21.1. Human Genetics. 1992; 89: 234-236.
10. Allander SV, Larsson C, Ehrenborg S, Suwanichkul A, Weber G, Morris SL, Bajalica S, Kiefer MC, Luthman H, Powell DR. Characterization of the chromosomal gene and identification of a functional promoter for human insulin-like growth factor binding protein-5. J Biol Chem. 1994; 269: 10891-10898.

164 S. V. Allander et al.
11. Zazzi H, Nikosjkov A, Hall H, Luthman H. Complete sequence of human IGFBP-4 Third International Symposium on insulin-like growth factors. Abstract 11342; 1994.
12. Ehrenborg E. Genetic characterization of human insulin-like growth factor binding proteins. Thesis. Karolinska Institutet. ISBN 91-628-1363-3; 1994.
13. Cubbage ML, Suwanichkul A, Powell DR. Structure of the human chromosomal gene for the 25 kilodalton insulin-like growth factor binding protein. Mol Endocrinol. 1989; 3: 846-851.
14. Cubbage ML, Suwanichkul A, Powell DR. Insulin-like growth factor binding protein-3: Organization of the human chromosomal gene and demonstration of promoter activity. J Biol Chem. 1990; 265: 12642-12649.
15. Binkert C, Margot JB, Landwehr J, Heinrich G, Schwander J. Structure of the human insulin-like growth factor binding protein-2 gene. Mol. Endocrinol. 1992; 6: 826-836.
16. Alitalo T, Kontula K, Koistinen R, Aalto-Set~il~t K, Julkunen M, J~inne OA, Sepp~tl~t M, de la Chapelle A. The gene encoding human low molecular weight insulin-like growth factor binding protein (IGF-BP25): Regional localization to 7p12-p13 and description of a DNA polymorphism. Human Genetics. 1989; 83: 335-338.
17. Ekstrand J, Ehrenborg E, Stern I, Stellan B, Zech L, Luthman H. The gene for insulin-like growth factor binding protein-1 is localized to chromosomal region 7p14-p12. Genomics. 1990; 6: 413418.
18. Agarwal N, Hsieh C-L, Sills D, Swaroop M, Desai B, Francke U, Swaroop A. Sequence analysis, expression and chromosomal localization of a gene, isolated from a substracted human retina cDNA library, that encodes an insulin-like growth factor binding protein (IGFBP2). Exp Eye Res. 1991; 52: 549-561.
19. Akam M. Hox and HOM: homologous gene clusters in insects and vertebrates. Cell. 1989; 57: 347-349.
20. Schugart K, Kappen C, Ruddle FH. Duplication of large genomic regions during the evolution of vertebrate homeobox genes. Proc Natl Acad Sci USA. 1989; 86: 7067-7071.
21. Schubert FR, Nieselt-Struwe K, Gruss P. The Antennapedia-type homeobox genes have evolved from three precursors separated early in metazoan evolution. Proc Natl Acad Sci USA. 1993; 90: 143-147.
22. Pendleton JW, Nagai BK, Murtha MT, Ruddle FH. Expansion of the Hox gene familly and the evolution of chordates. Proc Natl Acad Sci USA. 1993; 90: 6300~304.
23. Scott MP. Vertebrate homeobox gene nomenclature. Cell. 1992; 71: 551-553. 24. Cannizzaro LA, Croce CM, Griffin CA, Simeone A, Boncinelli E, Huebner K. Human homeo-box
containing genes located at chromosome regions 2q31-q37 and 12q12-12q13. Am J Hum Genet. 1987; 41: 1-15.
25. Acampora D, D'Esposito M, Faicella A, Pannese M, Migliaccio E, Morelli F, Stornaiuolo A, Nigro V, Simeone A, Boncinelli E. The human HOX gene family. Nucleic Acids Res. 1989; 17: 10385-10402.
26. 10.5 HgM. Update to the Tenth International Workshop on Human Gene mapping. Cytogenet Cell Genet. 1990; 55:1-785.
27. Lu-Kuo J, Ward DC, Spritz RA. Fluoresence in situ hybridization mapping of 25 markers on distal human chromosome 2q surrounding the human Waardenburg syndrome, type I (WSI) locus (PAX3 gene). Genomics. 1993; 16: 173-179.
28. Kou K, James PL, Clemmons DR, Copeland NG, Gilbert DJ. Jenkins NA, Rotwein P. Identification of two clusters of mouse insulin-like growth factor binding protein genes on chro- mosomes 1 and 11. Genomics. 1994; 21: 653-655.
29. Sneyers M, Kettman R, Massart S, Renaville R, Burny A, Portetelle D. Cloning and characteriza- tion of a cDNA encoding the bovine insulin-like growth factor binding protein 1 (blGFBP-I). DNA Seq. 1991; 1: 4074-408.
30. Upton FZ, Szabo L, Wallace JC, Ballard FJ. Characterization and cloning of a bovine insulin-like growth factor-binding protein. J Mol Endocrinol. 1990; 5: 77-84.
31. Shimasaki S, Shimonaka M, Ui M, Inouye S, Shibata F, Ling N. Structural characterization of a follicle-stimulating hormone action inhibitor in porcine ovarian follicular fluid: its identification as the insulin-like growth factor binding protein. J Biol Chem. 1990; 265: 2198-2202.
32. Spratt SK, Tatsuno GP, Sommer A. Cloning and characterization of bbvine insulin-like growth factor binding protein-3 (blGFBP-3). Biochem Biophys Res Commun. 1991; 177: 1025-1032.
33. Nagamatsu S, Chan SJ, Falkmer S, Steiner DF. Evolution of the insulin gene superfamily: Sequence of a preproinsulin-like growth factor cDNA from the Atlantic hagfish. J Biol Chem. 1991; 266: 2397-2402.

Conservation of IGFBP Structure during Evolution 165
34. Upton Z, Chan S J, Steiner DF, Wallace JC, Ballard FJ. Evolution of insulin-like growth factor binding proteins. Growth ReguL 1993; 3: 29-32.
35. Garcia-Fern~ndez J, Holland PWH. Archetypal organization of the amphioxous Hox gene cluster. Nature. 1994; 370: 563-566.
36. Chan SJ, Cao Q-P, Steiner DF. Evolution of the insulin superfamily: cloning of a hybrid insulin/insulin-like growth factor cDNA from amphioxus. Proc Natl Acad Sci USA. 1990; 87: 9319-9323.
37. Dickerson RE. The structure of the cytochrome c and the rates of molecular evolution. J Molec. Evol. 1971; l: 26-45.
38. Nei M. Molecular evolutionary genetics. New York: Columbia University Press; 1987. 39. Kiefer MC, Ioh RS, Bauer DM, Zapf J. Molecular cloning of a new human insulin-like growth
factor binding protein. Biochem Biophys Res Commun. 1991; 176; 219-225.