correlative microscopy
TRANSCRIPT

Correlative microscopy
ALBERTO LUINI
INSTITUTE OF PROTEIN BIOCHEMISTRY, CNR
ROMAN POLISHCHUCK
TIGEM
NAPLES

The Castellino Campus comprises a number of Institutes of the National Research Council (CNR), among
which the main ones are the IBP and the Institute of Genetics and Biophysics (IGB). Other Castellino
Institutes operate in the fields of photonics (IMM), informatics and computational modeling, and imaging
(IAC, ICAR). The Campus also hosts a large Telethon research Institute, the Telethon Institute for
Genetics and Medicine (TIGEM).
The IBP, IGB and TIGEM each possess an imaging facility. The IMM, IAC and ICAR develop technology
in the field of photonics and computational image analysis, and collaborate with the imaging facilities. In
addition, the IBP hosts the Telethon service Facility for advanced electron microscopy, tomography and
correlative microscopy. All of these facilities are managed in a coordinated manner that is regulated by
formal agreements among the Institutes, and are open to all of the members of the Castellino Campus.
The Telethon facility at the IBP has extensive experience in access, as it has offered service and assistance
to dozens of Telethon laboratories for a number of years. In many cases the support provided by facility
allowed Telethon-funded scientist to publish their data in top journals (see selected publications below).
he criteria and practice of access developed by the Telethon EM facility at IBP will be used to run the
EuroBioImaging proof of concept study (PCS).
THE IMAGING FACILITY AT THE CASTELLINO CAMPUS

The instrumentation available at the integrated Castellino facilities include
Light microscopy
- Confocal microscope Zeiss 710
- Confocal microscope Leica SP5
- Confocal microscope Leica SP2
- Fully motorized Leica DMI6000 light microscope equipped with incubation system for live cell imaging
- 3 Leica DMI6000 light microscopes equipped with image acquisition and analysis systems
- 2 Zeiss Axiophot microscopes equipped with image acquisition and analysis systems
Electron Microscopy
- FEI TEcnai G2 Spirit BioTWIN for EM tomography
- FEI TEcnai G2 Spirit BioTWIN for EM tomography (accessible in the CNR Institute ICTB)
- JEOL JEM-1011 electron microscope
- Leica Ultracut UCT ultramicrotome
- Leica EM FC7 ultramicrotome
- Leica EM TP automated tissue processor
Other equipment
- The accessory equipment comprises centrifuges, ovens, shakers and steromicroscopes for specimen preparation
- Eppendorf microinjection station
- 5 off line PC stations for EM and light microscopy image analysis
PERSONNEL EXPANSION

Short list of the diseases studied by TeEMCoF
Juvenile nephropathy
Ocular albinism
Juvenile hemochromatosis
Prion protein disease
Lysosomal storage diseases
Muscular dystrophy
Diabetes
Neurodystrophy
Optic atrothy
Roma
Napoli
Bari
Chieti
Siena
Padova
Milano
Perugia
Over 30 projects in
2005-2010
SERVICE
The Telethon Electron Microscopy Core Facility (TeEMCoF)
About 40 papers published in
2005-2010 comprising top journals
(Cell, Nature, etc.)
The Main Goal
To help Telethon funded studies of
genetic disease with electron
microscopy
Services
CLEM
EM tomography
Immuno-EM
Routine EM
Morphometry
Equipment use
Training

Advanced light microscopy is instrumental to
reach the ultimate goal of biological imaging,
to visualise single biomolecules and their
functions and interactions within the context of
live biological systems. The specific nodes will
provide the following key technologies:
Euro-BioImaging.
European Biomedical Imaging Infrastructure- from Molecule to Patient. A project on the European Roadmap for Research Infrastructures under coordination of EMBL and EIBIR
ADVANCED LIGHT MICROSCOPY NODES
Superresolution light microscopy
This node will provide access to
methods that improve the spatial
(and also temporal) resolution of
light microscopy imaging with an
emphasis on technologies
applicable for biological
applications and in live
specimens. Key technologies will
include stimulated emission
depletion (STED),
photoactivation localization
microscopy (PALM) as well as
the use of structured
illumination.
Functional imaging of live cells
This node will provide access to
methods that visualise
molecular function in live cells.
Key technologies will include
fluorescence lifetime imaging
(FLIM), fluorescence (cross)
correlation spectroscopy
(FC[C]S), photoactivation and
photobleaching (PA, FRAP),
single molecule imaging, and
novel fluorescent reporters of
biochemical reactions.
Correlative light and
electron microscopy
In this node it will be
possible to combine
dynamic functional assays
in live cells directly with
high resolution 3D
morphology at molecular
resolution by EM (cryo)
tomography. This node
will be intimately linked
to EM activities in the
ESFRI initiative
INSTRUCT.
High throughput microscopy
for systems biology
This node will contribute to
systems biology and rational
drug development by
providing access to automation
and high throughput in
advanced light microscopy
methods including ultra high
content screening of genome
level systematic perturbations
of biological systems such as
RNA interference over-
expression or small molecule
screening.
UNIQUENESS

CORRELATIVE MICROSCOPY
INTEGRATED MICROSCOPY
Attempts to apply different microscopy approaches
to the very same object of interest to integrate
information about the dynamics, fine structure and
composition of that object
VIDEO – EM
VIDEO - IF
VIDEO - IF - EM
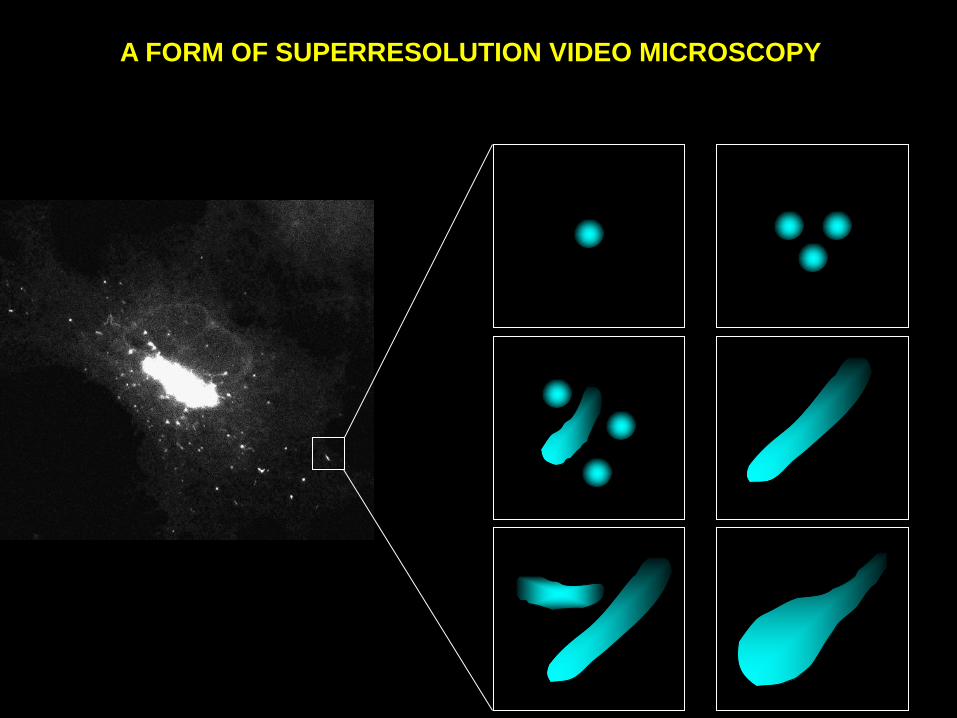
A FORM OF SUPERRESOLUTION VIDEO MICROSCOPY

Principle
Characterize dynamics by video microscopy
Switch to electron microscopy
Characterizing a dynamic process in live cells
Choosing a particular structure (stage) of interest in the process being
studied
Taking two pictures of the same structure, one at the LM, and the other
at the EM level.
Integrating the information
Advantages
Magnification
Structure (reference space)
Disadvantages
Fixation
Probes
Retracing
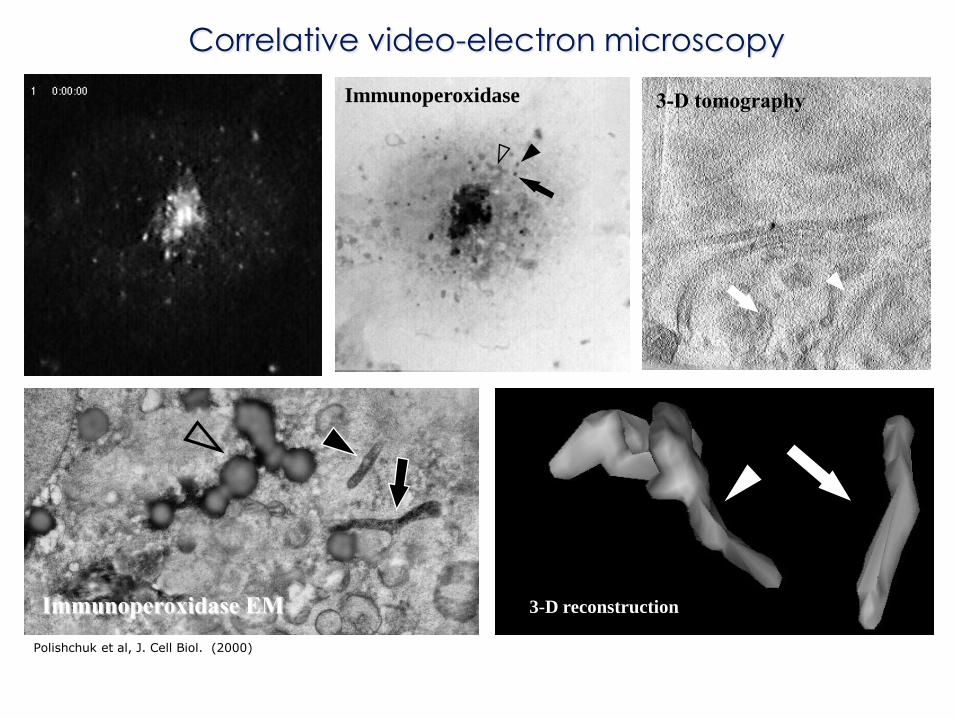
Immunoperoxidase
Immunoperoxidase EM 3-D reconstruction
3-D tomography
Correlative video-electron microscopy
Polishchuk et al, J. Cell Biol. (2000)

A COMPLEX INVOLVED IN CARRIER FORMATION AND FISSION
BY SERIAL BLEACHING AND LONG IMMUNO-FRET

COMBINING VIDEO MICROSCOPY WITH
EM TOMOGRAPHY
CRYO-FIXATION
IMMUNO-GOLD LABELING AND TOMOGRAPHY
DETECTION OF MOLECULAR COMPONENTS AND OF
MOLECULAR COMPLEXES IN OBJECTS OF INTEREST
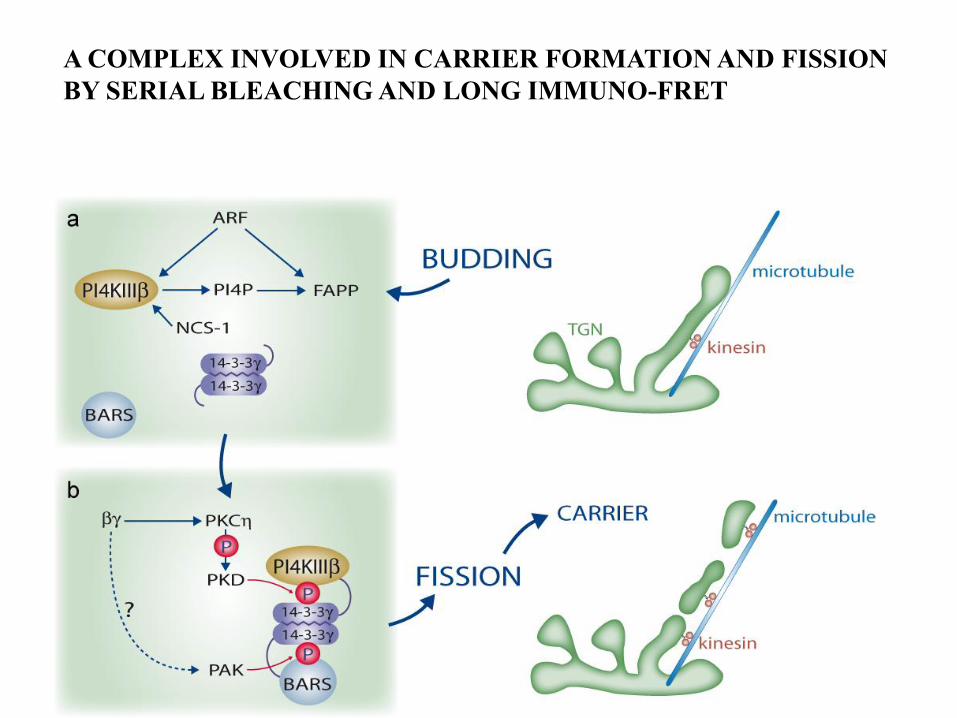
A COMPLEX INVOLVED IN CARRIER FORMATION AND FISSION
BY SERIAL BLEACHING AND LONG IMMUNO-FRET

protein complex
FRET distance (Å) from D to A
D
A

Förster Resonance Energy Transfer

THE IDEAL CONFIGURATION FOR LONG RANGE FRET MULTIPLE ACCEPTORS, LONG FRET DISTANCE
Single donor molecule allows an higher efficiency of energy transfer for a
cumulative effect and eliminates the problem of homotransfer
single
donor
multiple acceptors
Molecular complex
distance (Å) from D to A
D
A

Increasing
FRET efficacy
DONOR-ACCEPTOR distance (Å)

SERIAL LABELING – ACQUISITION – BLEACHING
DIFFICULTY: THE SIZE OF THE COMPLEXES
DETECTION OF MOLECULAR COMPLEXES BY FRET

A COMPLEX INVOLVED IN CARRIER
FORMATION AND FISSION

CORRELATIVE MICROSCOPY INTEGRATES INFORMATION ON :
DYNAMICS
ULTRASTRUCTURE
MOLECULAR COMPOSITION
ASSEMBLY OF MOLECULAR MACHINERY

VSVG-GFP
FAPP-2
CERT
PI4K
VSVG-GFP
Release from 20°C block
Multiple labelling of budding post-Golgi carriers by serial bleaching

A COMPLEX INVOLVED IN CARRIER FORMATION AND FISSION
BY SERIAL BLEACHING AND LONG IMMUNO-FRET

SERIAL BLEACHING OF PRIMARY ANTIBODIES
Schubert W, 2006

Protein complexes have different sizes, number of components (often many) and stability over time
a proteasome, a large
molecular complex
clathrin, a transient
protein assembly

A few moments of time-lapse video are enough to resolve an
Issue that years of microscopy on fixed cells have failed
to settle- Hugh R.B. Pelham (Nature. 1997)
Unfortunately, light microscopy cannot achieve a sufficiently
high resolution, so spectacular through it is, GFP technology
has its limits-Hugh R.B. Pelham (Nature. 1997)

The development of correlative microscopy can be somewhat arbitrarily
divided divided into two stages
1) An early stage from 1960 or earlier.
The goal is generally to look at the same field by both light (imuno-fluorescence)
and electron microscopy, to exploit the advantages of the two techniques:
the broad field of view of light and the resolution of electron microscopy.
2) A recent stage from 2000 onward.
Correlative microscopy as the first kind of GFP-based super-resolution video
microscopy with some disadvantages and a few substantial advantages over other super-
resolution video techniques that are being developed today

1986
Hayat MA. Correlative microscopy in Biology instrumentations and methods.
Academic Press. 1987

1995

TRANSPORT CARRIERS What kind of fine structure do they have?
With what organelles do they interact?

GFP-BASED CORRELATIVE LIGHT-ELECTRON MYCROSCOPY (CLEM):
EXPERIMENTAL PROCEDURE
1. DNA transfection of cells grown on
CELLocate coverslip.
2. GFP-based time-lapse confocal microscopy
and fixation of the cells.
3. Immunoperoxidase or gold labelling and
embedding in resin.
4. Cutting of serial sections and identification
of structure of interest at EM level.
knife
resin block
section

Perfect Loop
Slot grid
Collection of serial sections on the slot grids and their
analysis at the electron microscope
Serial sections
3D reconstruction Analysis of serial sections

Post-Golgi transport carriers in living cell
ER
Golgi
Plasma membrane
VSVG-GFP

Questions to address
• How are post-Golgi carriers
organized during different
stages of their life-cycle?
1. Formation
2. Transition through cytosol
3. Fusion
Golgi
Plasma membrane
1
2
3

Lumen
Cytosol
VSVG-GFP
VSVG is a widely used protein to study membrane transport
VSVG protein is a ts045 mutant strain of vesicular
stomatitis virus
40°C-VSVG is in the ER
20°C-VSVG is in the Golgi (TGN)
32°C-VSVG is moved out of the ER or the Golgi

Correlative light-electron microscopy of VSVG-GFP
labelled post-Golgi membrane carrier
VSVG-GFP
Polishchuk R.S. et al. ( 2000, JCB)

a b
c
d f g
r s
l k
n o p u
t
e
q
h i j
m
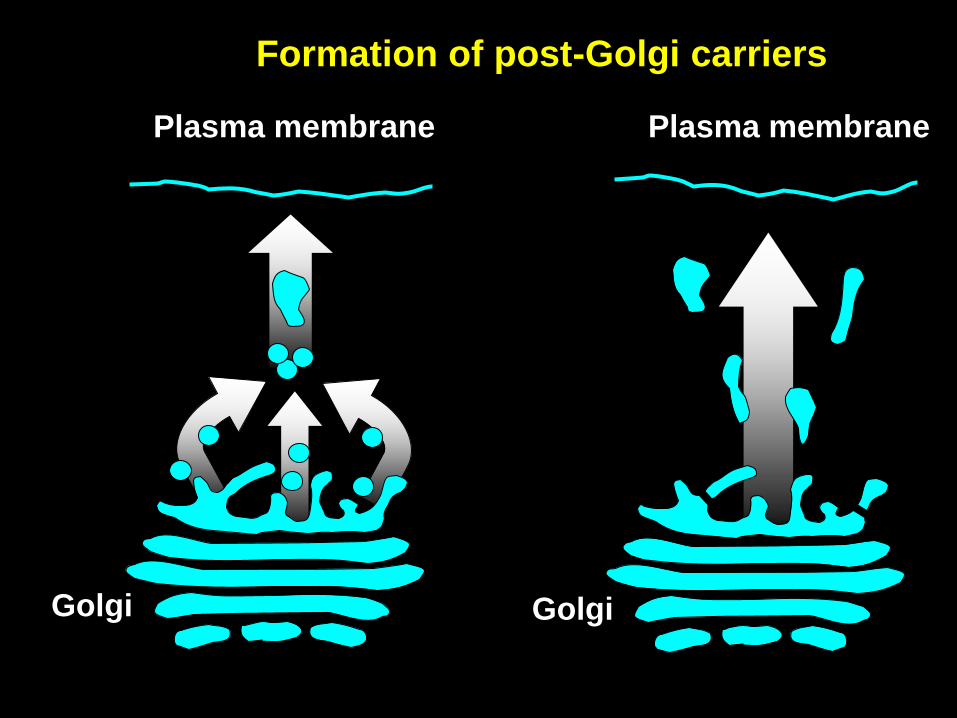
Golgi
Plasma membrane
Golgi
Plasma membrane
Formation of post-Golgi carriers

Formation of post-Golgi carriers
VSVG-GFP

VSVG-GFP
Ultrastructure of exit site of post-Golgi carrier
Polishchuk E.V. et al. ( 2003, MBC)

Ultrastructure of post-Golgi carrier formation site
Polishchuk E.V. et al. ( 2003, MBC)

Ultrastructure of post-Golgi carrier formation site
1
2
3
4
Polishchuk E.V. et al. ( 2003, MBC)

Ultrastructure of post-Golgi carrier formation site
1
2
3
4
Polishchuk E.V. et al. ( 2003, MBC)

Visualization of post-Golgi carrier fusion with
the plasma membrane by transmission EM VSVG-GFP VSVG-GFP Anti-VSVG HRP
Anti-VSVG HRP
Polishchuk R.S. et al. ( 2000, JCB)
VSVG-GFP
confocal microscopy Anti-VSVG gold
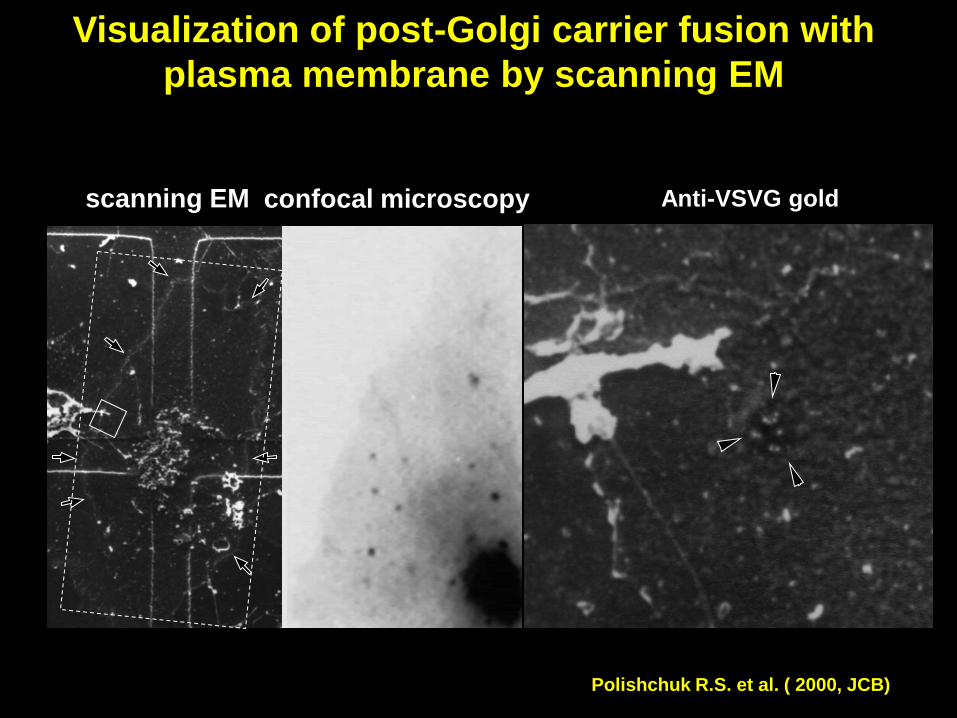
Visualization of post-Golgi carrier fusion with
plasma membrane by scanning EM
Polishchuk R.S. et al. ( 2000, JCB)
scanning EM

Constitutive transport from the Golgi to the
plasma membrane
Golgi
Plasma membrane
Polishchuk R.S. et al. ( 2000, JCB)

Golgi
Apical surface
Basolateral surface
ER
QUESTIONS TO ADDRESS
• Is ultrastructure of apical and basolateral
carriers different?
• Are apical and basolateral cargoes ever
packed into the same post-Golgi carrier?
•If so, how do they distribute within these
structures?
Transport of cargo proteins from the Golgi to
the apical and basolateral surfaces

Fluorescent proteins with apical and basolateral sorting
signals show polarized distribution in MDCK cells
GPI-GFP
Anti-occludin
XY
XZ
VSVG-GFP
Anti-occludin
XY
XZ
Apical marker Basolateral marker GPI-GFP
VSVG-GFP
Lumen
Cytosol
Lumen
Cytosol
Polishchuk R.S. et al. ( 2004, NCB)

Dynamics of Golgi-to-plasma membrane transport
of apical and basolateral markers in living cells
GPI-YFP
VSVG-CFP
merge
Polishchuk R.S. et al. ( 2004, NCB)

Ultrastructure of post-Golgi carriers containing both apical and
basolateral markers
GPI-YFP
VSVG-CFP
2
1
merge
Polishchuk R.S. et al. ( 2004, NCB)
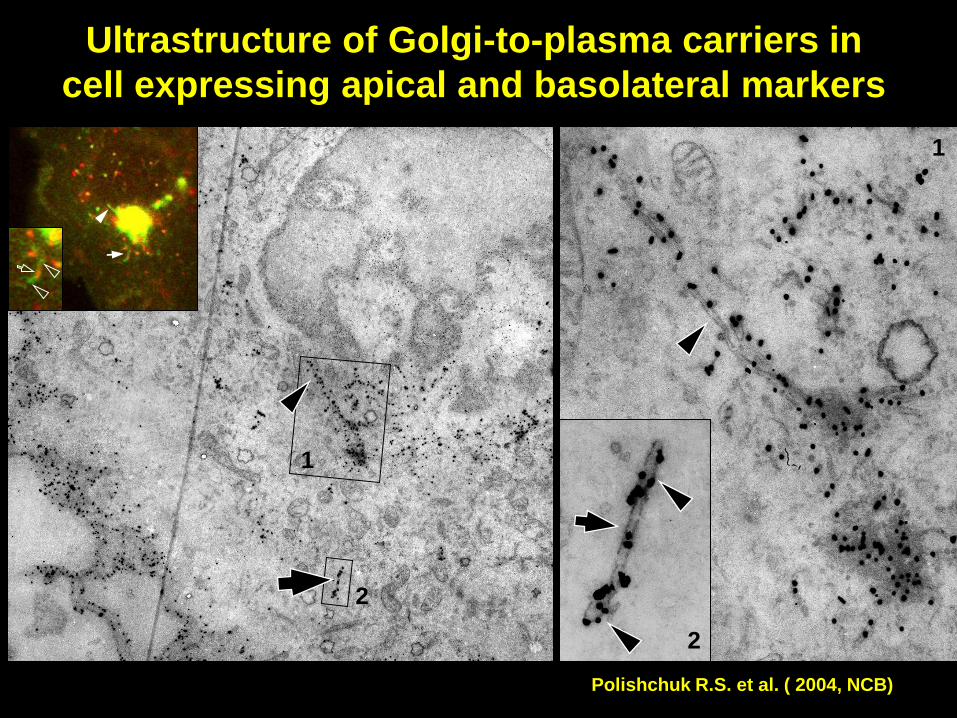
Ultrastructure of Golgi-to-plasma carriers in
cell expressing apical and basolateral markers
2
2
1
1
Polishchuk R.S. et al. ( 2004, NCB)

Intracellular membrane transport
trans-Golgi
network (TGN)
Golgi
Endosomes
Lysosomes
Secretory
granules
Plasma membrane
ER
Clathrin
AP1
GGAs
MPR 1
2
3

Structure of Golgi-to-endosome carriers
GFP view
- pleiomorphic carriers
- frequently tubules
- tubular or vesicular clusters
EM view
- clathrin coated vesicles
- or vesicular clusters

CLEM as a tool to characterize newly-formed
endosomal transport carriers
Constructs
CD-MPR-GFP-Cation-Dependent Mannose 6-Phosphate Receptor-GFP
GGA1-GFP-Golgi-localized Gamma-ear -containing ARF-binding
protein1-GFP
Clathrin light chain-GFP

GGA-GFP positive carriers in living cells
Polishchuk R.S. et al. ( 2006, Traffic)
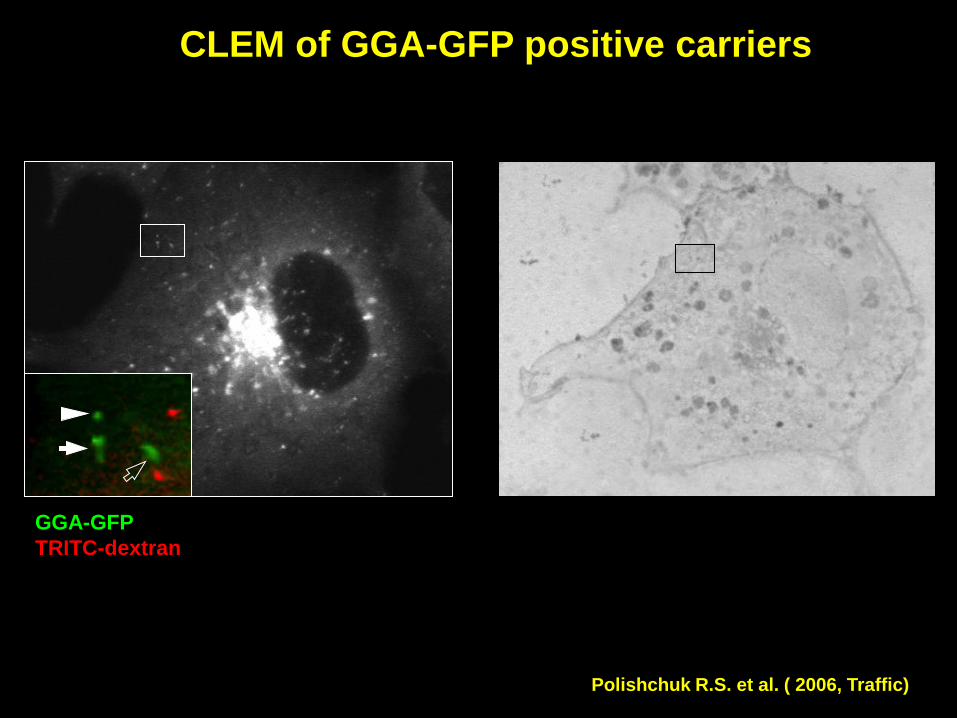
CLEM of GGA-GFP positive carriers
GGA-GFP
TRITC-dextran
Polishchuk R.S. et al. ( 2006, Traffic)

CLEM of GGA-GFP positive carriers
GGA-GFP
TRITC-dextran
Polishchuk R.S. et al. ( 2006, Traffic)
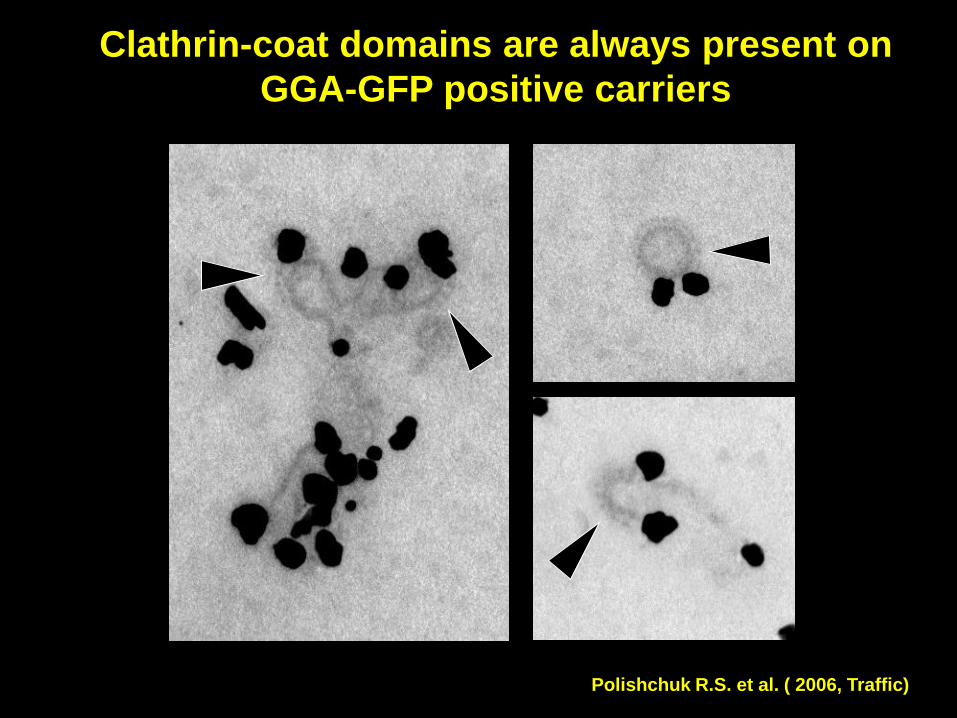
Clathrin-coat domains are always present on
GGA-GFP positive carriers
Polishchuk R.S. et al. ( 2006, Traffic)
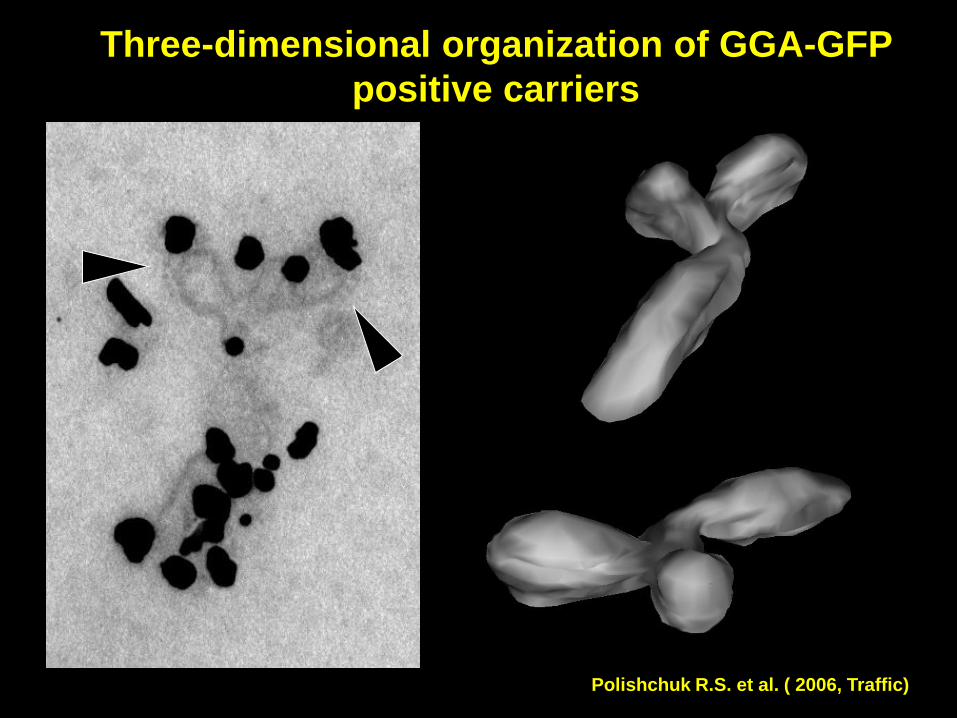
Three-dimensional organization of GGA-GFP
positive carriers
Polishchuk R.S. et al. ( 2006, Traffic)

Variability of shapes through GGA-GFP and
MPR-GFP positive TC populations
vesicle
ovoid
tubule
grape-like
Polishchuk R.S. et al. ( 2006, Traffic)

SUMMARY
• TGN-to-endosome TCs exhibit various morphology ranging
from vesicle-like to complex grape-like
• Frequently such TCs only partially covered with clathrin
• GGA adaptors are not restricted to the clathrin-coated domains
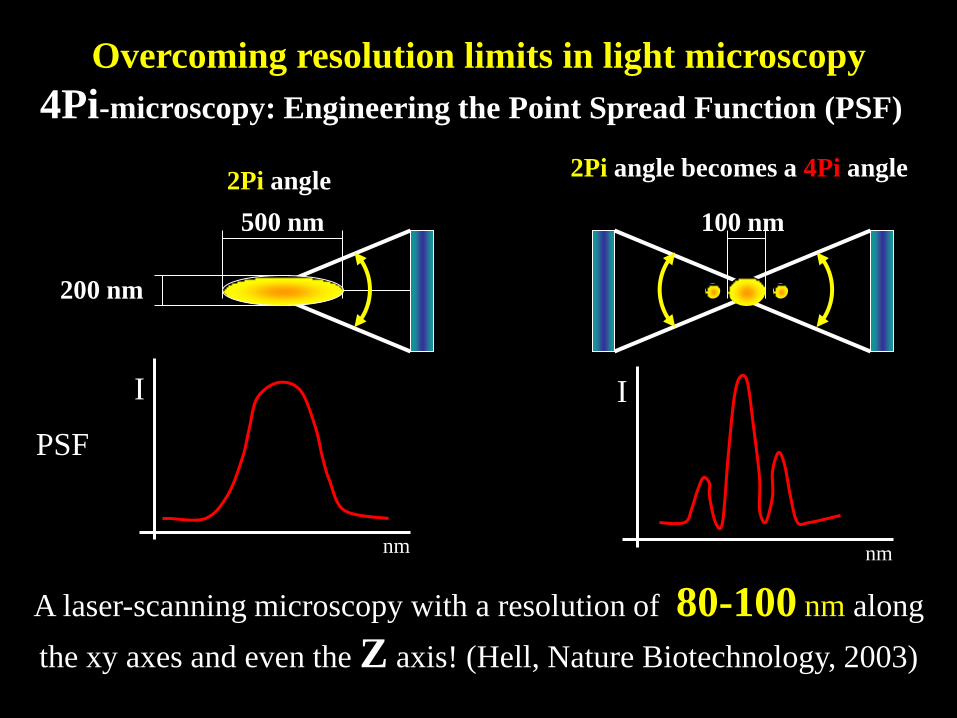
A laser-scanning microscopy with a resolution of 80-100 nm along
the xy axes and even the Z axis! (Hell, Nature Biotechnology, 2003)
2Pi angle 2Pi angle becomes a 4Pi angle
100 nm
PSF
nm
I
nm
4Pi-microscopy: Engineering the Point Spread Function (PSF)
Overcoming resolution limits in light microscopy
I
500 nm
200 nm

z
x
z
y
z
x
z
y
Confocal
4Pi
Perinetti G. et al. ( 2009, Traffic)

4Pi-CLEM Technique
DNA transfection EM processing
Serial Sectioning
3D Reconstruction
Comparison
Deconvolution EM serial recording
4Pi-microscopy Electron microscopy
3D Reconstruction
Confocal microscopy
Confocal Recording
Threshold Zero-Crossing
Threshold Zero-Crossing
Comparison
3D Reconstruction
4Pi Recording
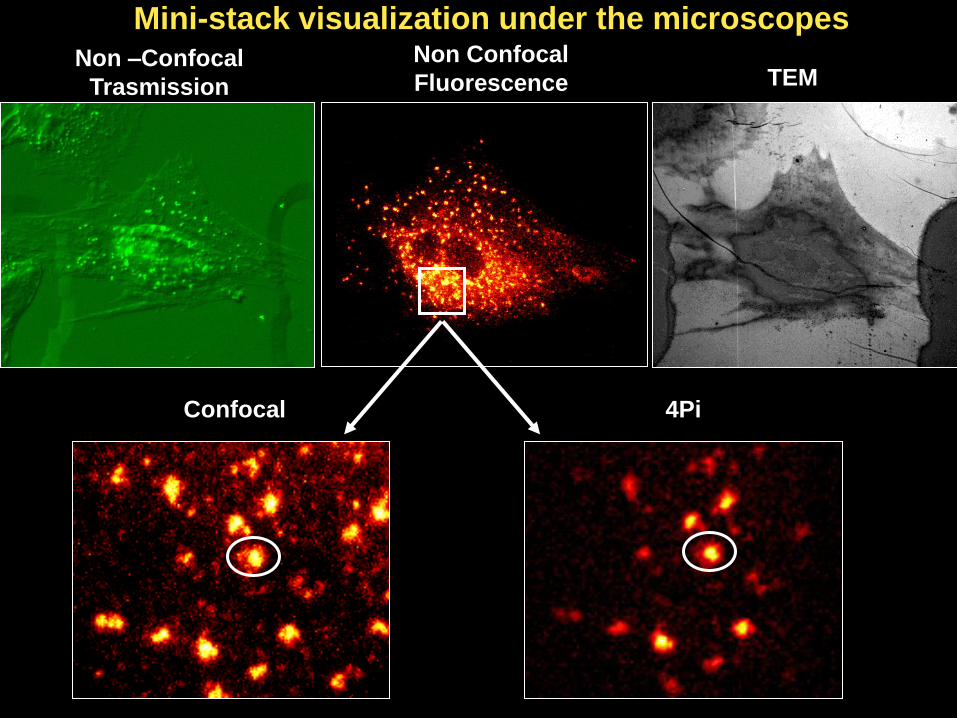
Non –Confocal
Trasmission
Non Confocal
Fluorescence TEM
Confocal 4Pi
Mini-stack visualization under the microscopes

Mini-stack identification under the TEM
recording
MS01
MS03 MS02
MS04
MS05
MS01
MS02
MS04
MS05
MS03
Perinetti G. et al. ( 2009, Traffic)

TEM serial recording and Imod labelling

4Pi-EM overlap
Threshold 20% Zero-Crossing
z
y
z
y
y
x
y
x

4Pi-EM overlap
z
y
z
y
y
x

Perspectives
• Combination with advanced light microscopy methods
• Development of the new light-electron microscopy
probes
• Combination with proteomics approaches

Mario Negri Sud Institute and TIGEM
Alexander Mironov
Albeto Luini
Elena Polishchuk
Alexander Mironov Jr.
Giuseppe Perinetti
Alexander Spaar
Acknowledgements
MPI (Goettingen)
Tobias Muller
Alexander Egner
Stefan Hell
NICHD (NIH) Bethesda
Jennifer Lippincott-Schwartz
Juan Bonifacino
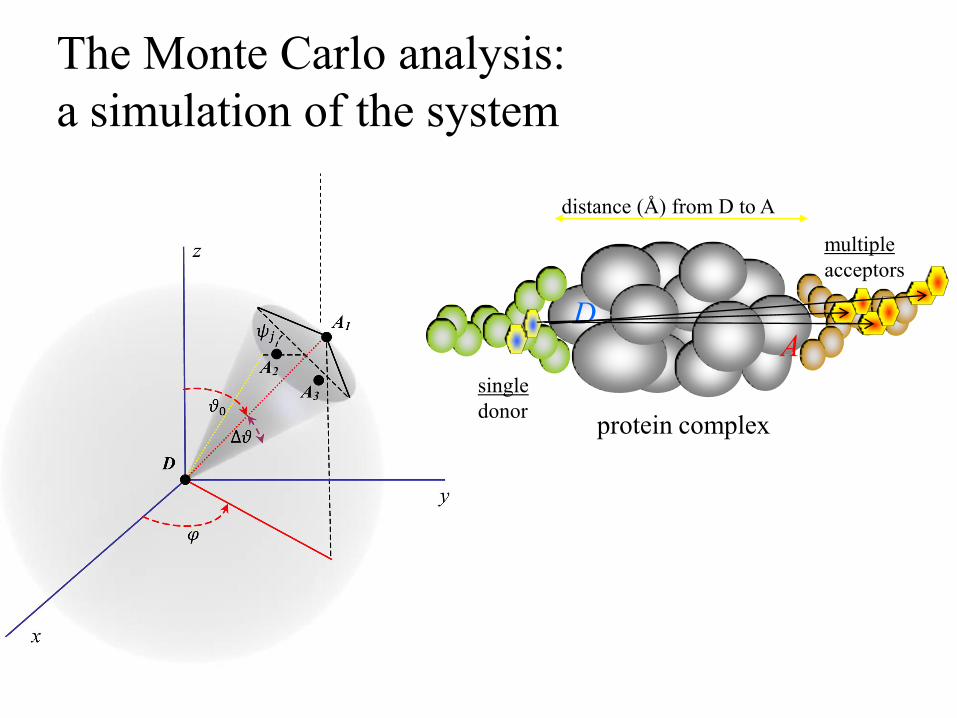
The Monte Carlo analysis:
a simulation of the system
single
donor
multiple
acceptors
protein complex
distance (Å) from D to A
D A

video-microscopy to survey
the cellular behaviour upon
the stimulating event
cells are fixed at the time of interest,
then sequential steps of:
immunolabeling with a donor and various acceptors
long range FRET assessments
selective bleaching of acceptors
staining another protein with new acceptors
TIME AND SPACE ONLY SPACE NOR TIME NEITHER SPACE INFORMATIVE
complexes composition should
be determined using standard
biochemical techniques
A typical experiment:
1994