cryptosporidiosis in humans - microbiology and molecular...
TRANSCRIPT

MICROBIOLOGICAL REVIEWS, Mar. 1983, p. 84-96 Vol. 47, No. 10146-0749/83/010084-13$02.00/0Copyright © 1983, American Society for Microbiology
Cryptosporidiosis in Animals and HumansSAUL TZIPORI
Attwood Veterinary Research Laboratory, Westmeadows, Victoria 3047, Australia
INTRODUCTION................................. 84CHARACTERISTICS OF THE ORGANISM................................. 84
Classification ................................. 84Life Cycle ................................. 84Species Specificity ................................. 87Diagnosis .................................. 88Studies on Oocysts .................................. 88
CHARACTERISTICS OF THE DISEASE................................. 89The Disease in Calves................................. 89The Disease in Lambs ................................. 89The Disease in Goats ................................. 89The Disease in Humans ................................. 89The Infection in Birds ................................ 90The Infection in Other Species................................ 90
EXPERIMENTAL CRYPTOSPORIDIOSIS ............. ................... 90Studies with Calf Isolates ................................ 90Studies with Human Isolates................................ 91
TREATMENT................................ 92CONCLUSIONS ................................ 92SUMMARY................................ 94LITERATURE CITED ................................ 94
INTRODUCTIONCryptosporidium is a protozoan parasite
which completes its life cycle on intestinal andrespiratory surface epitheliums of mammals,birds, and reptiles (28). The infection, untilrecently, was thought to be uncommon and theorganism was thought to be opportunistic and,like other coccidia, highly host specific.
Cryptosporidium was first recognized in thegastric glands of the laboratory mouse by Tyzzerin 1907 (58). Up to 1975, some 15 reports de-scribing the infection in eight species of animalswere published; only 5 of them associated cryp-tosporidiosis with some illness, 3 of which werein calves (36, 42, 52). Since 1975, over 60scientific publications have appeared in the liter-ature, the great majority being since 1980. Dur-ing this period our concept of cryptosporidiosiswas transformed from that of a rare and largelyasymptomatic infection to an important cause ofenterocolitis and diarrhea in several species,including humans.
In this communication an attempt was madeto gather the available information on the orga-nism, to examine the nature of the infection itproduces, and to demonstrate that Cryptospo-ridium infection can, under certain circum-stances, cause serious disease in some species ofanimals.
CHARACTERISTICS OF THE ORGANISMClassification
Cryptosporidium is a genus in the familyCryptosporidiidae, suborder Eimeriina, orderEucoccidiida, subclass Coccidia, class Sporo-zoa, phylum Apicomplexa (30). At present thesuborder Eimeriina contains 13 families withover 1,500 named species. The great majority ofthese species, however, belong to the generaEimeria and, to a lesser extent, Isospora. Bothare intracellular parasites which primarily infectthe intestinal tract of vertebrates. Toxoplasma, atissue cyst-forming coccidia, is another impor-tant member of the suborder Eimeriina.
Life CycleThe life cycle of Cryptosporidium has been
elucidated for a number of representative hostspecies and generally follows that of other enter-ic coccidia (10, 22, 77). However, there remainareas of discrepancy among authors regardingthe presence (77) or absence (7, 22, 59) of asecond-generation schizont. It has been suggest-ed that the second-generation schizont may sim-ply be sporocysts containing four sporozoites(44). However, sporocysts have not been identi-fied in structures considered to be oocysts (7),and the genus is not considered to have sporo-cysts by some (29). The morphological charac-
84
on May 19, 2018 by guest
http://mm
br.asm.org/
Dow
nloaded from
CRYPTOSPORIDIOSIS IN ANIMALS AND HUMANS
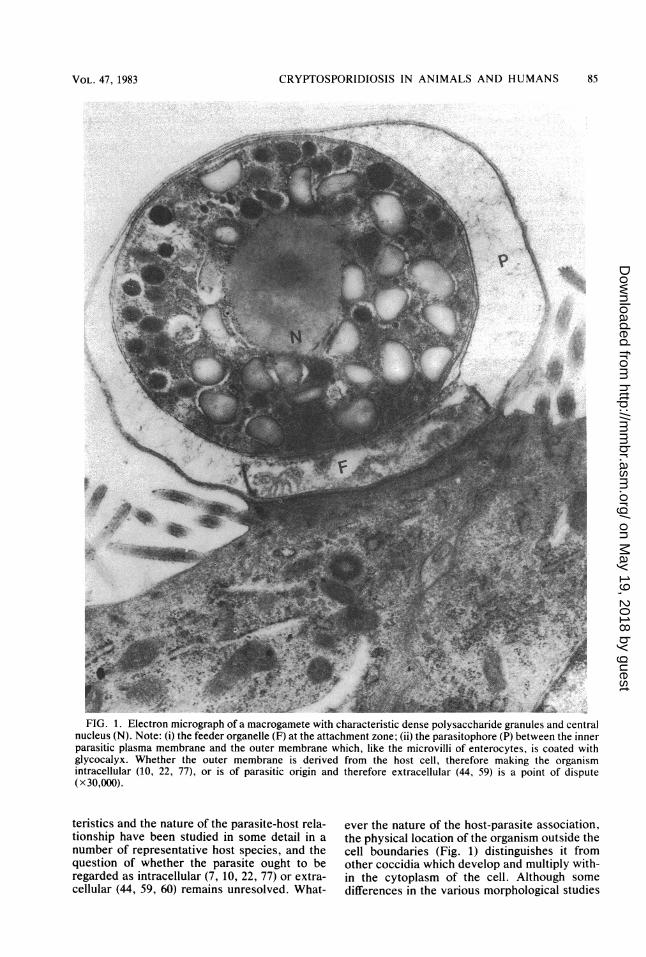
FIG. 1. Electron micrograph of a macrogamete with characteristic dense polysaccharide granules and centralnucleus (N). Note: (i) the feeder organelle (F) at the attachment zone; (ii) the parasitophore (P) between the innerparasitic plasma membrane and the outer membrane which, like the microvilli of enterocytes, is coated withglycocalyx. Whether the outer membrane is derived from the host cell, therefore making the organismintracellular (10, 22, 77), or is of parasitic origin and therefore extracellular (44, 59) is a point of dispute(x 30,000).
teristics and the nature of the parasite-host rela-tionship have been studied in some detail in anumber of representative host species, and thequestion of whether the parasite ought to beregarded as intracellular (7, 10, 22, 77) or extra-cellular (44, 59, 60) remains unresolved. What-
ever the nature of the host-parasite association,the physical location of the organism outside thecell boundaries (Fig. 1) distinguishes it fromother coccidia which develop and multiply with-in the cytoplasm of the cell. Although somedifferences in the various morphological studies
VOL. 47, 1983 85
on May 19, 2018 by guest
http://mm
br.asm.org/
Dow
nloaded from
86 TZIPORI
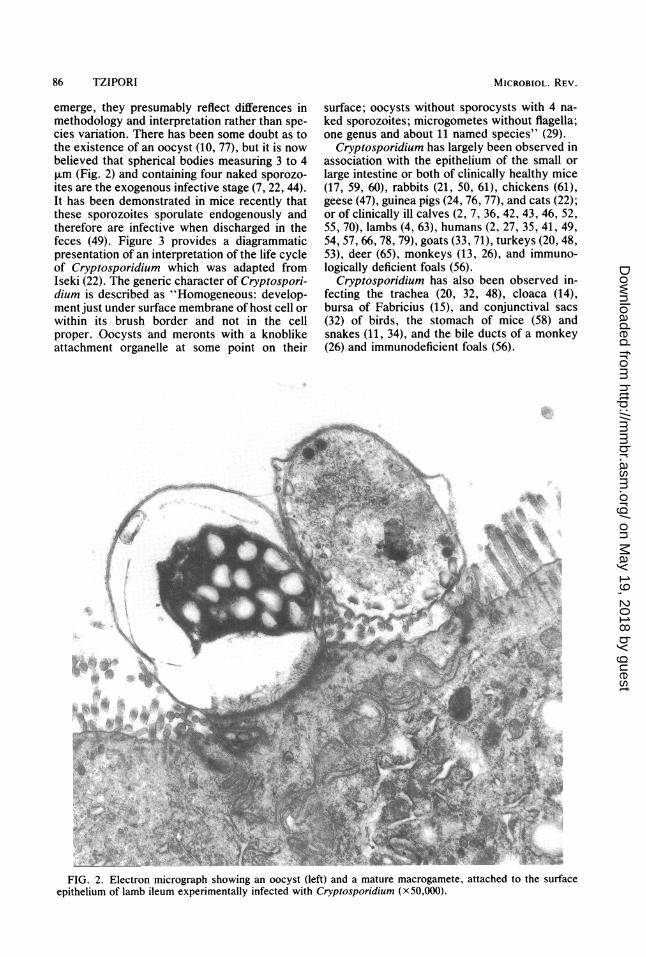
emerge, they presumably reflect differences inmethodology and interpretation rather than spe-cies variation. There has been some doubt as tothe existence of an oocyst (10, 77), but it is nowbelieved that spherical bodies measuring 3 to 4,um (Fig. 2) and containing four naked sporozo-ites are the exogenous infective stage (7, 22, 44).It has been demonstrated in mice recently thatthese sporozoites sporulate endogenously andtherefore are infective when discharged in thefeces (49). Figure 3 provides a diagrammaticpresentation of an interpretation of the life cycleof Cryptosporidium which was adapted fromIseki (22). The generic character of Cryptospori-dium is described as "Homogeneous: develop-ment just under surface membrane of host cell orwithin its brush border and not in the cellproper. Oocysts and meronts with a knoblikeattachment organelle at some point on their
surface; oocysts without sporocysts with 4 na-ked sporozoites; microgometes without flagella;one genus and about 11 named species" (29).
Cryptosporidium has largely been observed inassociation with the epithelium of the small orlarge intestine or both of clinically healthy mice(17, 59, 60), rabbits (21, 50, 61), chickens (61),geese (47), guinea pigs (24, 76, 77), and cats (22);or of clinically ill calves (2, 7, 36, 42, 43, 46, 52,55, 70), lambs (4, 63), humans (2, 27, 35, 41, 49,54, 57, 66, 78, 79), goats (33, 71), turkeys (20, 48,53), deer (65), monkeys (13, 26), and immuno-logically deficient foals (56).
Cryptosporidium has also been observed in-fecting the trachea (20, 32, 48), cloaca (14),bursa of Fabricius (15), and conjunctival sacs(32) of birds, the stomach of mice (58) andsnakes (11, 34), and the bile ducts of a monkey(26) and immunodeficient foals (56).
FIG. 2. Electron micrograph showing an oocyst (left) and a mature macrogamete, attached to the surfaceepithelium of lamb ileum experimentally infected with Cryptosporidium (x50,000).
MICROBIOL. REV.
on May 19, 2018 by guest
http://mm
br.asm.org/
Dow
nloaded from

CRYPTOSPORIDIOSIS IN ANIMALS AND HUMANS
OUTSIDE HOST
FIG. 3. Diagrammatic representation of the life cycle of Cryptosporidium. (1 to 4) Asexual cycle of theendogenous stage: 1, sporozoite or merozoite invading a microvillus of a small intestinal epithelial cell; 2, a fullygrown trophozoite, 3, a developing schizont with eight nuclei; 4, a mature schizont with eight merozoites. (5 and6) Sexual cycle: 5, microgametocyte with many nuclei; 6, macrogametocyte. (7) A mature oocyst containing foursporozoites without sporocyst. (8) Oocyst discharged in the feces. a, Merozoite released from mature schizont;b, sporozoite released from mature oocyst. Adapted from Isaki (22).
Species Specificity
Based on early transmission experimentsCryptosporidium was thought to be not onlyhost (22, 58, 76) but site specific (60). Morerecent experiments, using specific-pathogen-free (SPF) newborn animals, indicate that Cryp-tosporidium from some species of animals caninfect a wide variety of other animals, with orwithout causing illness (64). Thus, isolates fromcalves, humans, deer, goats, and lambs readilyinfect other species such as lambs, calves, andpiglets, causing diarrhea (37, 62, 64, 67, 72, 75;S. Tzipori, P. S. Bhathal, M. Smith, and C.Halpin, submitted for publication), and mice,rats, guinea pigs, chickens, and foals withoutcausing illness (49, 51, 64; Tzipori, Bhathal, etal., submitted; unpublished data). However,some differences between isolates have beennoted which could indicate "strain" variations.For instance, under similar experimental condi-tions calf isolates studied in Scotland infectedthe intestines of chickens (64) and propagatedserially at least 16 times in SPF mice (51),whereas Australian isolates consistently infect-ed the trachea of chickens and could only be
serially passaged three to four times in mice(unpublished data). The human isolates in ourexperience also appear to infect baby mice lessreadily than do isolates from other species (Tzi-pori, Bhathal, et al., submitted); on the otherhand, human isolates studied by Reese and co-workers (49) were said to infect adult mice aswell as newborns. Tyzzer was able to transmitinfection between adult mice (60), whereas abovine isolate only infected mice younger than21 days (51). These differences could be attribut-ed to either host or parasite factors or both. Theexact nature and indeed the existence of suchstrain differences will become clear when moreinformation is available, including the develop-ment of more sensitive serological tests and theavailability of techniques for accurately estimat-ing the infectious dose in experimental animals.
Passage of Cryptosporidium experimentallythrough animals does not appear to alter theirpathogenicity; after 12 serial passages of one calfisolate in mice, the infection remained asympto-matic for mice and highly pathogenic for lambs(51). A calf isolate and a human isolate werepassaged some 20 times intermittently throughSPF calves, pigs, and mice, causing the same
VOL. 47, 1983 87
on May 19, 2018 by guest
http://mm
br.asm.org/
Dow
nloaded from

88 TZIPORI
degree of illness in calves and piglets and sub-clinical infections in mice (unpublished data).
DiagnosisThe most reliable method for diagnosis is
histological demonstration of the endogenousstages attached to the brush border of epithelialcells. Although the existence of oocysts hasbeen known for some time, it was only recentlythat their presence in feces was utilized fordiagnosis. The method used to identify oocystsinclude the modified zinc sulfate centrifugal flo-tation (22), Giemsa-stained fecal smears (Fig. 4)(45, 55), the fecal flotation technique (1), and themodified Ziehl-Neelson technique (19). Theidentity of the organism can be confirmed by theoral inoculation offeces into SPF newborn labo-ratory animals in which the endogenous stagescan then be identified histologically (64). Untilrecently cryptosporidiosis in humans was diag-nosed by intestinal biopsy (27, 35, 41, 54, 57, 78,79). In animals that are clinically affected thereis a close correlation between illness and excre-tion of oocysts in the feces (2, 4, 37, 45, 55, 63,70-73, 75).
Studies on OocystsCryptosporidium oocysts, like those of other
coccidia, are extremely resistant to a variety ofdisinfectants commonly used in laboratories andhospitals. These include iodophore, cresylic
acid, sodium hypochlorite, benzylkonium chlo-ride, sodium hydroxide (12), and two aldehyde-based disinfectants (5). Infectivity of oocystswas destroyed by ammonia, formol saline (12),freeze-drying, and exposure to temperatures be-low freezing and above 65°C for 30 min (unpub-lished data). Infectivity is also lost over a periodof 2 to 6 months at 4°C irrespective of themethod of storage (37, 51). Oocysts can be keptviable for at least 8 to 9 months in containerswhere air is excluded. Excystation seems tooccur shortly after exposure to air, presumablystimulated by appropriate levels of oxygen andcarbon dioxide (unpublished data).Few methods have been suggested for quanti-
tation of infectivity in infectious material foranimal inoculation. These include flotation andcounting (22), titration in mice (51), or countingthe number of oocysts present in a 5-,u suspen-sion smeared and stained with Giemsa (75).
Purification of Cryptosporidium oocysts inmaterial intended for animal inoculation was aproblem. Indeed, most animal experiments wereconducted with preparations which, althoughknown to be free of other enteropathogens,contained enteric microorganisms (6, 37, 49, 62,64, 67, 72). Recently, however, oocysts weresuccessfully purified by treatment of feces withalcohol (60% final dilution) before they wereused for inoculation (74, 75; Tzipori, Bhathal, etal., submitted).
Ag diiPpI. :q.
t~* .. .
.** 0
+
ts
9Sd
FIG. 4. Giemsa-stained fecal smear taken from a human patient with gastroenteritis; two intact Cryptospori-dium oocysts (arrows) can be seen with some internal structures (x3,000).
MICROBIOL. REV.
IMF
'WI
on May 19, 2018 by guest
http://mm
br.asm.org/
Dow
nloaded from

CRYPTOSPORIDIOSIS IN ANIMALS AND HUMANS
CHARACTERISTICS OF THE DISEASE
The number of reports which describe crypto-sporidiosis in calves is by far the greatest andsuggests that the infection is most important inthis species.
The Disease in CalvesThe disease is reported to be common in
North America (9, 39, 45), Europe (18, 40, 46),and Australia (23). Cryptosporidium infectionsin calves occur commonly with other entericpathogens which for a long time made the as-
sessment of their role as an enteropathogenuncertain (39, 45, 55). A few recent reports,however, have described outbreaks of diarrheawhich could only be attributed to Cryptosporid-ium (2, 9, 18, 70). The clinical picture whichemerges from field reports is one of mild tosevere diarrhea occurring in calves aged be-tween 1 and 4 weeks, with high morbidity andlow mortality. The youngest calf reported to beaffected with the disease was 4 days (55) and theoldest was 26 days (70). In one study (70) theillness lasted between 2 and 14 days, the averagebeing 7 days; relapses occurring after apparentrecovery have also been reported. In anotherreport in which a mortality rate of 16% was
recorded, the clinical signs were said to includepersistent diarrhea, dehydration, and weakness(9). Cryptosporidiosis was also reported to havecontributed to the etiology of a syndrome, "Ca-chexia in Veal Calves," described in Holland(16).
The Disease in LambsTwo outbreaks of enteritis attributed to cryp-
tosporidiosis have recently been reported inlambs (4, 63). The first outbreak occurred in 40of 48 artificially reared lambs within 5 to 12 daysof birth, of which 16 subsequently died. Crypto-sporidium was detected in 10 of 16 fecal samplesexamined, and histological sections taken fromthe ileum of dead lambs confirmed the diagnosis.In another outbreak, 200 suckling lambs of atotal of 532 had diarrhea and 58 subsequentlydied. Lambs between 8 and 12 days of agedeveloped diarrhea which resulted in eitherdeath 2 to 3 days later or recovery after anillness of up to 7 days. In lambs, too, there werecases of a second bout of diarrhea after apparentrecovery (4). Two earlier reports (7, 8) whichdescribed Cryptosporidium infection in lambsattributed the diarrhea to concurrent infectionswith pathogenic bacteria.
The Disease in GoatsCryptosporidium was detected in a dead goat
kid with diarrhea (33) and in a herd of 29suckling kids of which 21 suffered severe diar-
rhea, and 3 subsequently died (71). The affectedkids were aged between 5 and 21 days of age andthe diarrhea lasted 3 to 7 days. There were sevencases of relapse shortly after recovery. Youngerkids seemed to have suffered for longer periodsand were more prone to relapses than olderanimals. Gut sections examined from two deadkids showed organisms attached to the brushborder of enterocytes and mucosal lesions con-sistent with cryptosporidiosis seen in calves andlambs.
The Disease in HumansThere have been a dozen or so reports in the
literature on cases of human cryptosporidiosis.Six of these involved patients who had pro-longed illness and were shown to be immunolog-ically deficient. Two of them were 6-year-oldpatients with congenital immunoglobulin defi-ciency who had persistent diarrhea which lasted3 (27) and 6 (54) years, culminating in the deathof the latter child. Four other patients hadimpaired humoral and cellular immunity as aresult of immunosuppressive chemotherapy (35,79) or there was clinical evidence of incompe-tence (57, 78). One of them (57) died, after apersistent diarrhea which lasted 2 years, fromoverwhelming dissemination of toxoplasmosis.However, the underlying cause of death wassaid to have been severe malabsorption due toCryptosporidium infection of the small intestine.One patient recovered within 3 weeks of with-drawal of immunosuppressive therapy (35). Theremaining four reports refer to individuals whowere apparently immunologically normal (2, 41,49, 66). Infection in three of them was diagnosedby demonstration of oocyst shedding in the feces(2, 49, 66). The diarrhea in these patients lastedfrom 1 to 3 weeks and was associated withabdominal pain, fever, and vomiting whichfailed to respond to treatment. In two of thesereports (2, 49) there was circumstantial evidenceto suggest that infection had resulted from calf-human transmission. In patients from whomintestinal biopsies were examined histologically,there were varying degrees of mucosal changeswhich included partial villous atrophy, lengthen-ing of the crypt, low cuboidal surface epitheli-um, and cellular infiltration of the lamina propriaof the jejunum and ileum.A limited serological survey showed that the
infection was as common among humans asamong nine other species of animals tested (68).Studies on the prevalence of infection in hospitalpatients with gastroenteritis showed that, overthe four Australian summer months (February toMay), 26 of 369 examined (7%) were excretingCryptosporidium oocysts in the feces; of these,only 4 showed evidence of infection with otherenteropathogens. For the remainder of the year,
89VOL. 47, 1983
on May 19, 2018 by guest
http://mm
br.asm.org/
Dow
nloaded from

90 TZIPORI
10 of 515 patients (2%) were excreting oocysts;none of 320 hospital patients without gastroen-teritis (S. Tripori, M. Smith, C. Birch, G.Barnes, and R. Bishop, submitted for publica-tion) did so. The most common clinical manifes-tation was diarrhea, mostly accompanied byvomiting, anorexia, and abdominal pain lastingfrom 3 to at least 14 days. The majority of thesepatients were city dwellers, and in some in-stances there were a few cases which occurredwithin one family or a neighborhood, indicatingthat transmission may occur among humans.Infection in humans, therefore, does not neces-sarily originate from contact with animals assome authors believe. This was the first evi-dence that the infection could be an importantcause of mild to severe transient diarrhea inimmunologically normal patients and that theinfection in humans, as in other species, can bediagnosed by the presence of oocysts in thefeces.
The Infection in BirdsTurkeys and peacocks are the only reported
species in which infection was observed in theupper respiratory tract (20, 32, 48). The infectionwas associated with clinical respiratory signs ofvarying severity. Histological changes observedin the trachea included thickened mucosa due toinfiltration of the lamina propria with histio-cytes, lymphocytes, and heterocytes and flatten-ing of the epithelium. In one case the ceca wereheavily infected as well. In some cases theorganisms were seen adhering to the mucosa ofthe nasal cavity and the bronchi. The infectionwas seen in six different flocks within 2 months.The morbidity and mortality in one instancewere up to 30 and 20%, respectively (20).One of 30 peacock chicks which died at about
2 weeks of age from a respiratory disease wasexamined (32). It was depressed and had agurgling respiration, coughing, and sneezingwith a serous ocular discharge. Organisms wereobserved attached to the surface of conjunctival,nasal, sinus, and tracheal epithelium.Other reports in birds include illness with
diarrhea and a low death rate in 10- to 14-day-oldturkey poults due to Cryptosporidium infectionof the small intestine (53), asymptomatic infec-tion of the cloaca of two Red-lored parrots (14),the bursa of Fabricius of three chickens (15), andthe large intestine of one domestic goose (47).
The Infection in Other SpeciesGeneralized Cryptosporidium infection was
observed in five immunodeficient Arabian foalswith diarrhea which were thought to have diedof adenoviral infection (56). In these foals organ-isms were seen adhering to the wall of the
stomach and throughout the small and largebowel, the common bile duct, gall bladder, andmajor pancreatic ducts. It was thought that thestate of immunodeficiency in these foals in-creased susceptibility to the infection.
Cryptosporidiosis was reported in two juve-nile rhesus monkeys, one of which died sudden-ly with severe enterocolitis and a second ofwhich suffered persistent diarrhea (26). The in-fection was also observed in eight rhesus mon-keys which were believed to have died fromother causes (13).
Illness associated with Cryptosporidium infec-tion was also described in snakes (11, 34), in onecase causing prolonged, severe, chronic hyper-trophic gastritis in 14 mature reptiles.Asymptomatic infections were reported to oc-
cur in mice (17, 58, 59), rabbits (21, 50), cats(22), guinea pigs (24, 76, 77), and several-week-old piglets (25, 31).
EXPERIMENTAL CRYPTOSPORIDIOSISExperimental disease was studied in calves
(37, 44, 45, 74), lambs (6, 62-64, 67, 73), piglets(37, 64, 72, 75; Tzipori, Bhathal, et al., submit-ted), rats, guinea pigs, chicks (64, 76, 77), mice(49, 51, 64), monkeys, and foals (unpublisheddata). The most extensively studied isolateswere those obtained from calves and humans.
Studies with Calf IsolatesSeveral calf isolates induced enterocolitis and
diarrhea of equal severity in newborn calves,SPF lambs, and piglets, except that calves re-mained susceptible to a much older age. Therewas no difference in the clinical response orpathological findings between calves fed, ordeprived of, colostrum or whether they wereinfected orally or by contact (74). The diseasewas characterized by depression, anorexia, anddiarrhea. The incubation period varied between3 and 5 days and up to 7 days in calves infectedby contact (74). Excretion of Cryptosporidiumoocysts in the feces coincided with the durationof clinical illness. The small intestine was infect-ed in all calves, and the cecum and colon wereinfected in more than half (37, 74). Mucosalchanges in the small intestine were most severein the ileum, and the effects most frequentlyobserved included stunting and swelling of villiwith variable degrees of villous fusion. Affectedvilli were infiltrated by numerous macrophages,neutrophils, and eosinophils and were oftenlined with immature cuboidal cells. The laminapropria of affected areas contained numerousmononuclear cells, with some eosinophils and afew neutrophils. Areas in the ileum were hyper-emic and exhibited a focal turgidity, with serumprotein leakage at the base of the crypts. Small
MICROBIOL. REV.
on May 19, 2018 by guest
http://mm
br.asm.org/
Dow
nloaded from
CRYPTOSPORIDIOSIS IN ANIMALS AND HUMANS
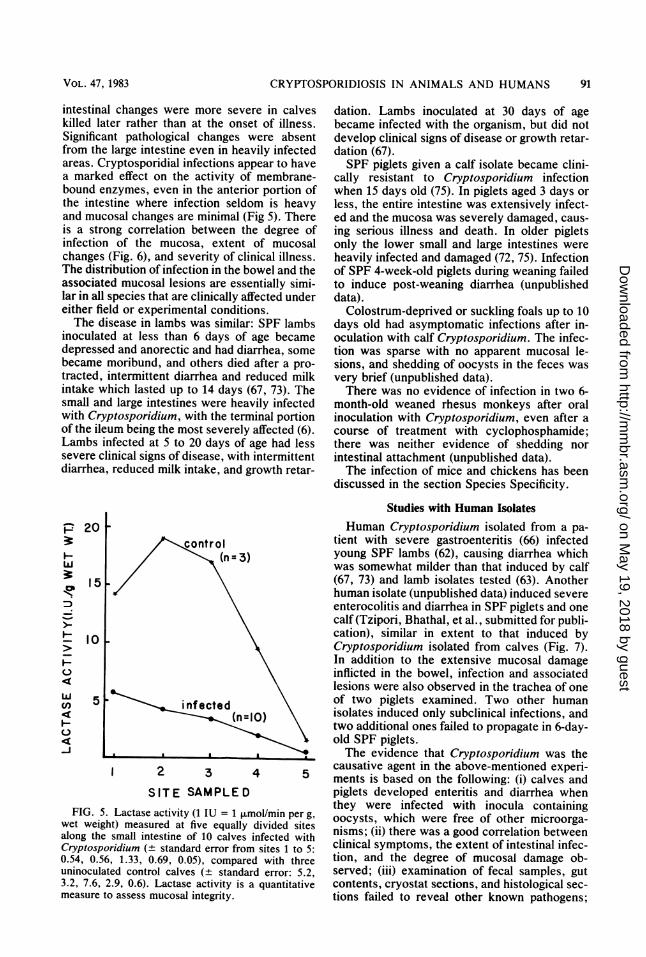
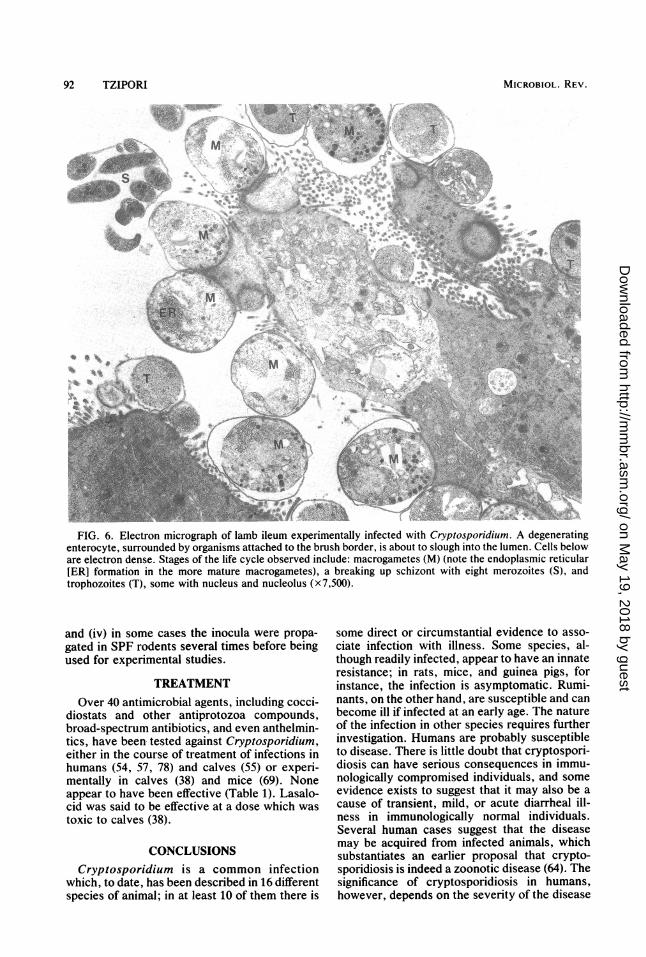
intestinal changes were more severe in calveskilled later rather than at the onset of illness.Significant pathological changes were absentfrom the large intestine even in heavily infectedareas. Cryptosporidial infections appear to havea marked effect on the activity of membrane-bound enzymes, even in the anterior portion ofthe intestine where infection seldom is heavyand mucosal changes are minimal (Fig 5). Thereis a strong correlation between the degree ofinfection of the mucosa, extent of mucosalchanges (Fig. 6), and severity of clinical illness.The distribution of infection in the bowel and theassociated mucosal lesions are essentially simi-lar in all species that are clinically affected undereither field or experimental conditions.The disease in lambs was similar: SPF lambs
inoculated at less than 6 days of age becamedepressed and anorectic and had diarrhea, somebecame moribund, and others died after a pro-tracted, intermittent diarrhea and reduced milkintake which lasted up to 14 days (67, 73). Thesmall and large intestines were heavily infectedwith Cryptosporidium, with the terminal portionof the ileum being the most severely affected (6).Lambs infected at 5 to 20 days of age had lesssevere clinical signs of disease, with intermittentdiarrhea, reduced milk intake, and growth retar-
3
w
w
Cl)
C-)
FIG. 5.wet weiglalong theCryptospo0.54, 0.56uninoculal3.2, 7.6, 2measure t4
dation. Lambs inoculated at 30 days of agebecame infected with the organism, but did notdevelop clinical signs of disease or growth retar-dation (67).SPF piglets given a calf isolate became clini-
cally resistant to Cryptosporidium infectionwhen 15 days old (75). In piglets aged 3 days orless, the entire intestine was extensively infect-ed and the mucosa was severely damaged, caus-ing serious illness and death. In older pigletsonly the lower small and large intestines wereheavily infected and damaged (72, 75). Infectionof SPF 4-week-old piglets during weaning failedto induce post-weaning diarrhea (unpublisheddata).
Colostrum-deprived or suckling foals up to 10days old had asymptomatic infections after in-oculation with calf Cryptosporidium. The infec-tion was sparse with no apparent mucosal le-sions, and shedding of oocysts in the feces wasvery brief (unpublished data).There was no evidence of infection in two 6-
month-old weaned rhesus monkeys after oralinoculation with Cryptosporidium, even after acourse of treatment with cyclophosphamide;there was neither evidence of shedding norintestinal attachment (unpublished data).The infection of mice and chickens has been
discussed in the section Species Specificity.
Studies with Human IsolatesHuman Cryptosporidium isolated from a pa-
control tient with severe gastroenteritis (66) infected/(n-3) young SPF lambs (62), causing diarrhea whichwas somewhat milder than that induced by calf(67, 73) and lamb isolates tested (63). Anotherhuman isolate (unpublished data) induced severeenterocolitis and diarrhea in SPF piglets and onecalf (Tzipori, Bhathal, et al., submitted for publi-cation), similar in extent to that induced byCryptosporidium isolated from calves (Fig. 7).In addition to the extensive mucosal damageinflicted in the bowel, infection and associatedlesions were also observed in the trachea of one
infected of two piglets examined. Two other human(n=10) \ isolates induced only subclinical infections, and
two additional ones failed to propagate in 6-day-old SPF piglets.
.____________________________.The evidence that Cryptosporidium was thel 2 3 4 5 causative agent in the above-mentioned experi-
ments is based on the following: (i) calves andSITE SAMPLED piglets developed enteritis and diarrhea when
they were infected with inocula containingLactase activity (1 IU = 1 smo/min per g, oocysts, which were free of other microorga-ht) measured at five equally divided sites nisms; (ii) there was a good correlation betweensmall intestine of 10 calves infected withridium (± standard error from sites 1 to 5: clinical symptoms, the extent of intestinal infec-, 1.33, 0.69, 0.05), compared with three tion, and the degree of mucosal damage ob-ted control calves (+ standard error: 5.2, served; (iii) examination of fecal samples, gut2.9, 0.6). Lactase activity is a quantitative contents, cryostat sections, and histological sec-o assess mucosal integrity. tions failed to reveal other known pathogens;
VOL. 47, 1983 91
on May 19, 2018 by guest
http://mm
br.asm.org/
Dow
nloaded from
92 TZIPORI
FIG. 6. Electron micrograph of lamb ileum experimentally infected with Cryptosporidium. A degeneratingenterocyte, surrounded by organisms attached to the brush border, is about to slough into the lumen. Cells beloware electron dense. Stages of the life cycle observed include: macrogametes (M) (note the endoplasmic reticular[ER] formation in the more mature macrogametes), a breaking up schizont with eight merozoites (S), andtrophozoites (T), some with nucleus and nucleolus (x7,500).
and (iv) in some cases the inocula were propa-gated in SPF rodents several times before beingused for experimental studies.
TREATMENTOver 40 antimicrobial agents, including cocci-
diostats and other antiprotozoa compounds,broad-spectrum antibiotics, and even anthelmin-tics, have been tested against Cryptosporidium,either in the course of treatment of infections inhumans (54, 57, 78) and calves (55) or experi-mentally in calves (38) and mice (69). Noneappear to have been effective (Table 1). Lasalo-cid was said to be effective at a dose which wastoxic to calves (38).
CONCLUSIONSCryptosporidium is a common infection
which, to date, has been described in 16 differentspecies of animal; in at least 10 of them there is
some direct or circumstantial evidence to asso-ciate infection with illness. Some species, al-though readily infected, appear to have an innateresistance; in rats, mice, and guinea pigs, forinstance, the infection is asymptomatic. Rumi-nants, on the other hand, are susceptible and canbecome ill if infected at an early age. The natureof the infection in other species requires furtherinvestigation. Humans are probably susceptibleto disease. There is little doubt that cryptospori-diosis can have serious consequences in immu-nologically compromised individuals, and someevidence exists to suggest that it may also be acause of transient, mild, or acute diarrheal ill-ness in immunologically normal individuals.Several human cases suggest that the diseasemay be acquired from infected animals, whichsubstantiates an earlier proposal that crypto-sporidiosis is indeed a zoonotic disease (64). Thesignificance of cryptosporidiosis in humans,however, depends on the severity of the disease
MICROBIOL. REV.
on May 19, 2018 by guest
http://mm
br.asm.org/
Dow
nloaded from
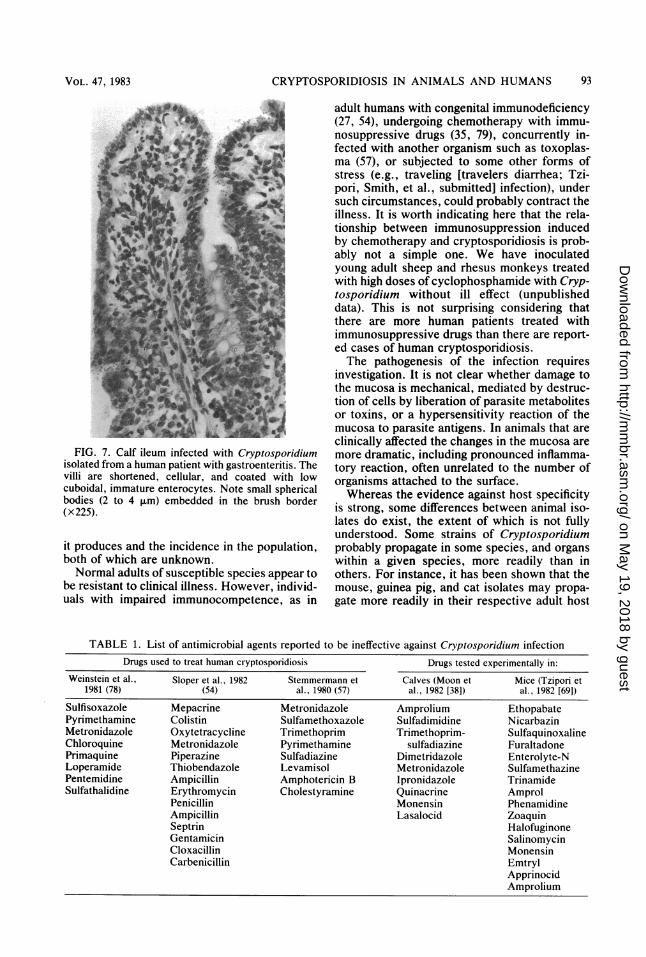
CRYPTOSPORIDIOSIS IN ANIMALS AND HUMANS
FIG. 7. Calf ileum infected with Cryptosporidiumisolated from a human patient with gastroenteritis. Thevilli are shortened, cellular, and coated with lowcuboidal, immature enterocytes. Note small sphericalbodies (2 to 4 ,um) embedded in the brush border(x225).
it produces and the incidence in the population,both of which are unknown.Normal adults of susceptible species appear to
be resistant to clinical illness. However, individ-uals with impaired immunocompetence, as in
adult humans with congenital immunodeficiency(27, 54), undergoing chemotherapy with immu-nosuppressive drugs (35, 79), concurrently in-fected with another organism such as toxoplas-ma (57), or subjected to some other forms ofstress (e.g., traveling [travelers diarrhea; Tzi-pori, Smith, et al., submitted] infection), undersuch circumstances, could probably contract theillness. It is worth indicating here that the rela-tionship between immunosuppression inducedby chemotherapy and cryptosporidiosis is prob-ably not a simple one. We have inoculatedyoung adult sheep and rhesus monkeys treatedwith high doses of cyclophosphamide with Cryp-tosporidium without ill effect (unpublisheddata). This is not surprising considering thatthere are more human patients treated withimmunosuppressive drugs than there are report-ed cases of human cryptosporidiosis.The pathogenesis of the infection requires
investigation. It is not clear whether damage tothe mucosa is mechanical, mediated by destruc-tion of cells by liberation of parasite metabolitesor toxins, or a hypersensitivity reaction of themucosa to parasite antigens. In animals that areclinically affected the changes in the mucosa aremore dramatic, including pronounced inflamma-tory reaction, often unrelated to the number oforganisms attached to the surface.Whereas the evidence against host specificity
is strong, some differences between animal iso-lates do exist, the extent of which is not fullyunderstood. Some strains of Cryptosporidiumprobably propagate in some species, and organswithin a given species, more readily than inothers. For instance, it has been shown that themouse, guinea pig, and cat isolates may propa-gate more readily in their respective adult host
TABLE 1. List of antimicrobial agents reported to be ineffective against Cryptosporidium infectionDrugs used to treat human cryptosporidiosis Drugs tested experimentally in:
Weinstein et al., Sloper et al., 1982 Stemmermann et Calves (Moon et Mice (Tzipori et1981 (78) (54) al., 1980 (57) al., 1982 [38]) al., 1982 [691)
Sulfisoxazole Mepacrine Metronidazole Amprolium EthopabatePyrimethamine Colistin Sulfamethoxazole Sulfadimidine NicarbazinMetronidazole Oxytetracycline Trimethoprim Trimethoprim- SulfaquinoxalineChloroquine Metronidazole Pyrimethamine sulfadiazine FuraltadonePrimaquine Piperazine Sulfadiazine Dimetridazole Enterolyte-NLoperamide Thiobendazole Levamisol Metronidazole SulfamethazinePentemidine Ampicillin Amphotericin B Ipronidazole TrinamideSulfathalidine Erythromycin Cholestyramine Quinacrine Amprol
Penicillin Monensin PhenamidineAmpicillin Lasalocid ZoaquinSeptrin HalofuginoneGentamicin SalinomycinCloxacillin MonensinCarbenicillin Emtryl
ApprinocidAmprolium
VOL. 47, 1983 93
on May 19, 2018 by guest
http://mm
br.asm.org/
Dow
nloaded from

94 TZIPORI
and in a particular organ. They presumablywould also infect, and may even induce, entero-colitis of varying degrees in newborn susceptiblehosts and asymptomatic infection in newbornlaboratory animals. A limited serial passagethrough one host does not appear to alter theiraffinity to that host; it may require much greaternumbers of passages to do so. The nature ofdifferences between isolates has wide implica-tions in terms of our understanding of compara-tive medicine, natural transmission betweenspecies, and the epidemiology of infection ingeneral.The difference in the incubation period be-
tween susceptible and resistant species may be areflection of variation in the life cycle of theorganism which, in turn, influences the outcomeof the infection. For instance, the number ofgenerations of schizonts before the sexual stageand the role, if it indeed exists, of autoinfectioncan influence both the length of the life cycle andthe outcome of infection.The organism appears to be resistant to a great
variety of drugs that have been used successful-ly for treatment against other protozoan infec-tions. It is anticipated that effective drugs maybe those that are taken up by infected cells.
Cryptosporidiosis caused more severe clinicalillness in lambs and calves and in older animalsthan did rotavirus, enterotoxigenic Escherich-ia coli or both (73, 74; unpublished data). Pig-lets, on the other hand, became resistant to theinfection at a much younger age than they did torotavirus or enteropathogenic E. coli (77). Thedisease causes prolonged brush border maldi-gestion and, to a greater extent, malabsorptionresulting from altered mucosal architecture ofthe lower small bowel. Death resulting fromdehydration and acidosis is less common.
Cryptosporidium strains primarily infect thealimentary tract and the upper respiratory tractof some birds, but in one instance infection ofthe trachea in a piglet experimentally inoculatedwith a human isolate was also noted. The fre-quency and significance of respiratory tract in-fection in mammals are not clear and may implythat transmission of infection may be by aerosol,as well as fecal. Indeed, in our experimentalinoculation of chickens, where only the tracheawas apparently infected, no oocysts were de-tected in the feces (unpublished data). The in-volvement of the respiratory tract in an infectionmay also jeopardize the effectiveness of future,nonsystemic, oral chemotherapy.
If our assumption of a single species genus(64), in addition to the diversity of tissue tro-phism, is correct and the preliminary serologicalstudies (69) are a true reflection of prevalence,then Cryptosporidium must be one of the mostsuccessful and adaptable parasites known.
Therefore, it is surprising that in the decadessince its discovery the ability of the organism tocause disease has not received more attention.
SUMMARYThe protozoan Cryptosporidium (family Cryp-
tospiridiidae), which was identified in 1907, hasonly recently been shown to be an importantcause of enterocolitis and diarrhea in a nuber ofmammalian species. The disease in humans wasinitially reported to occur in immunologicallycompromised individuals, but a recent studyamong hospital patients with gastroenteritis indi-cated that the infection is common in immuno-logically normal patients. The organism lackshost specificity, a characteristic uncommonamong other enteric coccidia, and is therefore apotential zoonosis. It is extremely resistant tothe action of common laboratory disinfectantsand to antimicrobial agents tested so far. Apreliminary serological survey indicates that theinfection is prevalent among and within popula-tions of 10 mammalian species examined. Fieldoutbreaks of diarrhea attributed to Cryptospori-dium infection have so far been reported incalves, lambs, deer, and goats and the diseasehad been reproduced experimentally in lambs,calves, and piglets. Sporadic cases of illness inother species have also been reported, and inbirds it has been shown to cause upper respira-tory tract infection. The organism infects theentire bowel but most commonly the lower smallintestine, where extensive mucosal changes oc-cur. The organism completes its life cycle on themucosal lining by adhering to the brush borderof enterocytes. Cryptosporidium causes partialatrophy, fusion, and distortion of villi, resultingin maldigestion in the brush border and malab-sorption.
ACKNOWLEDGMENTSI am grateful to the staff of the Moredun Research Institute,
Edinburgh, U.K., where my involvement with this area ofresearch began, for their enthusiastic support; in particular, Ithank Kenneth W. Angus.
ADDENDUM IN PROOFCryptosporidium infection has recently been de-
scribed in a gray squirrel (J. P. Sundberg, D. Hill, andM. J. Ryan, J. Am. Vet. Med. Assoc. 181:1420-1422,1982) and in a raccoon (B. L. Carlson and S. W.Nielson, J. Am. Vet. Med. Assoc. 181:1405-1406,1982). Additional information regarding natural andexperimental cryptosporidiosis in lambs (B. C. Ander-son, J. Am. Vet. Med. Assoc. 181:151-153, 1982),experimental fecal transmission of Cryptosporidiumbetween humans and piglets (H. W. Moon, A.Schwartz, M. J. Welch, P. P. McCann, and P. L.Runnels, Vet. Pathol. 19:700-707, 1982), and a chronicdiarrhea and wasting in a cat (K. B. Pooncha and C.Pippin, Vet. Pathol. 19:708-710, 1982) is now avail-able.
MICROBIOL. REV.
on May 19, 2018 by guest
http://mm
br.asm.org/
Dow
nloaded from

CRYPTOSPORIDIOSIS IN ANIMALS AND HUMANS
LITERATURE CITED1. Anderson, B. C. 1981. Patterns of shedding of cryptospori-
dial oocysts in Idaho calves. J. Am. Vet. Med. Assoc.178:982-984.
2. Anderson, B. C., and M. S. Bulgin. 1981. Enteritis causedby Cryptosporidium in calves. Vet. Med. Small Anim.Clin. 76:865-868.
3. Anderson, B. C., T. Donndelinger, R. M. Wilkins, and J.Smith. 1982. Cryptosporidiosis in a veterinary student. J.Am. Vet. Med. Assoc. 180:408-409.
4. Angus, K. W., W. T. Appleyard, J. D. Menzies, I. Camp-bell, and D. Sherwood. 1982. An outbreak of diarrhoeaassociated with cryptosporidiosis in naturally rearedlambs. Vet. Rec. 110:129-130.
5. Angus, K. W., D. Sherwood, G. Hutchison, and I. Camp-bell. 1982. Evaluation of two aldehyde-based disinfectantson the infectivity of faecal cryptosporidia for mice. Res.Vet. Sci., 33:379-381.
6. Angus, K. W., S. Tzipori, and E. W. Gray. 1982. Intestinallesions in specific-pathogen-free lambs associated withCryptosporidium from calves with diarrhoea. Vet. Pathol.19:67-78.
7. Barker, I. K., and P. L. Carbonell. 1974. Cryptosporidiumagni sp. n. from lambs, and Cryptosporidium bovis sp. n.from a calf, with observations on the oocysts. Z. Parasi-tenkd. 44:489-498.
8. Berg, I. E., A. C. Peterson, and T. P. Freeman. 1978.Ovine cryptosporidiosis. J. Am. Vet. Med. Assoc.173:1586-1587.
9. Bergeland, M. E., D. D. Johnson, and H. Shave. 1979.Bovine cryptosporidiosis in the north-central UnitedStates, p. 131-138. In 22nd Annual Proceedings, Ameri-can Association of Veterinary Laboratory Diagnosticians.
10. Bird, R. G., and M. D. Smith. 1980. Cryptosporidiosis inman: parasite life cycle and fine structural pathology. J.Pathol. 132:217-233.
11. Brownstein, D. G., J. D. Strandberg, R. J. Montali, M.Bush, and J. Fortner. 1977. Cryptosporidium in snakeswith hypertrophic gastritis. Vet. Pathol. 14:606-617.
12. Campbell, I., S. Tzipori, G. Hutchison, and K. W. Angus.1982. The effect of disinfectants on survival of Crypto-sporidium oocysts. Vet. Rec., 111:414-415.
13. Cockerell, B. Y., M. G. Valerio, and F. M. Garner. 1974.Cryptosporidiosis in the intestines of Rhesus monkeys(Macaca mulatta). Lab. Anim. Sci. 24:881-887.
14. Doster, A. R., E. A. Mahaffey, and J. R. McClearen.Cryptosporidium in the cloacal coprodeum of Red-loredparrots (Amazona autumnalis). Avian Dis. 23:654-661.
15. Fletcher, 0. J., J. F. Munnell, and R. K. Page. 1975.Cryptosporidiosis of the Bursa of Fabricius of chickens.Avian Dis. 19:630-639.
16. Hage-Noordam, A. W., J. M. A. Pol, and P. W. de Leeuw.1982. Cryptosporidium in veal calves affected with ca-chexia. Tijdschr. Diergeneeskd. 107:497-502.
17. Hampton, J. C., and B. Rosario. 1966. The attachment ofprotozoan parasites to intestinal epithelial cells of themouse. J. Parasitol. 52:939-949.
18. Heiene, J. von, and J. Boch. 1981. Kryptosporidien-Infektionen beim Kalb. Nachweis, Vorkommen und ex-perimentelle Ubertragung. Berl. Muenich. Tieraerztl.Wochenschr. 94:289-292.
19. Henriksen, S. A., and J. F. L. Pohlenz. 1981. Staining ofcryptosporidia by a modified Zeihl-Neelsen technique.Acta Vet. Scand. 22:594-596.
20. Hoerr, F. J., F. M. Ranck, and T. F. Hastings. 1978.Respiratory cryptosporidiosis in turkeys. J. Am. Vet.Med. Assoc. 173:1591-1593.
21. Inman, L. R., and A. Takeuchi. 1979. Spontaneous cryp-tosporidiosis in an adult female rabbit. Vet. Pathol. 16:89-95.
22. Iseki, M. 1979. Cryptosporidium felis sp. n. (Protozoa:Eimeriorina) from the domestic cat. Jpn. J. Parasitol.28:285-307.
23. Jerrett, I. V., and D. R. Snodgrass. 1981. Cryptosporidiaassociated with outbreaks of neonatal calf diarrhoea.
Aust. Vet. J. 57:434-435.24. Jervis, H. B., T. G. Merrill, and H. Sprinz. 1966. Coccidi-
osis in the guinea pig small intestine due to a Cryptospori-dium. Am. J. Vet. Res. 27:408-414.
25. Kennedy, G. A., G. L. Kreitner, and A. C. Strafuss. 1977.Cryptosporidiosis in three pigs. J. Am. Vet. Med. Assoc.170:348-350.
26. Kovatch, R. M., and J. D. White. 1972. Cryptosporidiosisin two juvenile Rhesus monkeys. Vet. Pathol. 9:426-440.
27. Lasser, K. H., K. J. Lewin, and F. W. Ryning. 1979.Cryptosporidial enteritis in a patient with congenital hypo-gammaglobulinaemia. Hum. Pathol. 10:234-240.
28. Levine, N. D. 1973. Protozoan parasites of domesticanimals and man, p. 229-230, 2nd ed. Burgess Publica-tions, Minneapolis.
29. Levine, N. D. 1980. Some corrections of coccidian (Api-complexa:Protozoa) nomenclature. J. Parasitol. 66:830-834.
30. Levine, N. D., J. 0. Corliss, F. E. G. Cos, G. Deroux, J.Grain, B. M. Honigberg, G. F. Leedale, A. R. LoeblichIII, J. Lom, D. Lynn, E. G. Merinfeld, F. C. Page, G.Poljansky, V. Sprague, J. Vavra, and F. G. Wallace. 1980.A newly revised classification of the protozoa. J. Proto-zool. 27:37-58.
31. Links, I. J. 1982. Cryptosporidial infection of piglets.Aust. Vet. J. 58:60-62.
32. Mason, R. W., and W. J. Hartley. 1980. Respiratorycryptosporidiosis in a peacock chick. Avian Dis. 24:771-776.
33. Mason, R. W., W. J. Hartley, and L. Tilt. 1981. Intestinalcryptosporidiosis in a kid goat. Aust. Vet. J. 57:386-388.
34. McKenzie, R. A., P. E. Green, W. J. Hartley, and C. C.Pollitt. 1978. Cryptosporidium in a red-bellied black snake(Pseudechis porphyriacus). Aust. Vet. J. 54:365-366.
35. Meisel, J. L., D. R. Perera, C. Meligro, and C. E. Rubin.1976. Overwhelming watery diarrhoea associated with aCryptosporidium in an immunosuppressed patient. Gas-troenterology 70:1156-1160.
36. Meuten, D. J., H. J. van Kruiningen, and D. H. Lein. 1975.Cryptosporidiosis in a calf. J. Am. Vet. Med. Assoc.165:917.
37. Moon, H. W., and W. K. Bemrick. 1981. Faecal transmis-sion of calf cryptosporidia between calves and pigs. Vet.Pathol. 18:248-255.
38. Moon, H. W., G. N. Woode, and F. A. Ahrens. 1982.Attempted chemoprophylaxis of cryptosporidiosis incalves. Vet. Rec. 110:181.
39. Morin, M., S. Lariviere, and R. Lallier. 1976. Pathologicaland microbiological observations made on spontaneouscases of acute neonatal calf diarrhoea. Can. J. Comp.Med. 40:228-240.
40. Nagy, B., A. Antal, and F. Ratz. 1979. Occurrence ofbovine cryptosporidiosis in Hungary. Magy. Allatorv.Lapja 34:585-588.
41. Nime, F. A., J. D. Burek, D. L. Page, M. A. Holscher, andJ. H. Yardley. 1976. Acute enterocolitis in a human beinginfected with the protozoan Cryptosporidium. Gastroen-terology 70:592-598.
42. Panciera, R. J., R. W. Thomassen, and F. M. Garner.1971. Cryptosporidial infection in a calf. Vet. Pathol.8:479-484.
43. Pearson, G. R., and E. F. Logan. 1978. Demonstration ofCryptosporidia in the small intestine of a calf by lighttransmission and scanning electron microscopy. Vet. Rec.103:212-213.
44. Pohlenz, J., W. J. Bemrick, H. W. Moon, and N. F.Cheville. 1978. Bovine cryptosporidiosis: a transmissionand scanning electron microscopic study of stages in thelife cycle and of the host-parasite relationship. Vet.Pathol. 15:417-427.
45. Pohlenz, J., H. W. Moon, N. F. Cheville, and W. J.Bemrick. 1978. Cryptosporidiosis as a probable factor inneonatal diarrhoea of calves. J. Am. Vet. Med. Assoc.172:452-457.
46. Pol, J. M. A., B. E. C. Schreuder, G. J. Kok, and P. W. de
VOL. 47, 1983 95
on May 19, 2018 by guest
http://mm
br.asm.org/
Dow
nloaded from

96 TZIPORI
Leeuw. 1982. Cryptosporidium: a new factor in the aetiol-ogy of neonatal diarrhoea in calves. Tijdschr. Diergen-eeskd. 107:503-510.
47. Proctor, S. J., and R. L. Kemp. 1974. Cryptosporidiumanserinum sp. n. (sporozoa) in a domestic goose Anseranser L. from Iowa. J. Protozool. 21:664-666.
48. Ranck, F. M. 1979. Cryptosporidiosis-a possible cocci-dial respiratory pathogen in turkeys, p. 49-51. In Proceed-ings of the 28th Western Poultry Disease Conference and13th Poultry Health Symposium.
49. Reese, N. C., W. L. Current, J. V. Ernst, and W. S.Bailey. 1982. Cryptosporidiosis of man and calf: a casereport and results of experimental infections in mice andrats. Am. J. Trop. Med. Hyg. 31:226-229.
50. Rehg, J. E., G. W. Lawson, and S. P. Pakes. 1979.Cryptosporidium cuniculus in the rabbit (Oryctolaguscuniculus). Lab. Anim. Sci. 29:656-660.
51. Sherwood, D., K. W. Angus, D. R. Snodgrass, and S.Tzipori. 1982. Experimental cryptosporidiosis in labora-tory mice. Infect. Immun. 38:471-475.
52. Shmitz, J. A., and D. H. Smith. 1975. Cryptosporidiuminfection in a calf. J. Am. Vet. Med. Assoc. 167:731-732.
53. Slavin, D. 1955. Cryptosporidium meleagridis (sp. nov.).J. Comp. Pathol. 65:262-266.
54. Sloper, K. S., R. R. Dourmashkin, R. B. Bird, G. Slavin,and A. D. B. Webster. 1982. Chronic malabsorption dueto cryptosporidiosis in a child with immunoglobulin defi-ciency. Gut 23:80-82.
55. Snodgrass, D. R., K. W. Angus, E. W. Gray, W. A. Keir,and L. W. Clerihew. 1980. Cryptosporidia associated withrotavirus and an Escherichia coli in an outbreak of calfscour. Vet. Rec. 106:458-459.
56. Snyder, S. P., J. J. England, and A. E. McChesney. 1978.Cryptosporidiosis in immunodeficient arabian foals. Vet.Pathol. 15:12-17.
57. Stemmermann, G. N., T. Hayashi, G. A. Glober, N. Oishi,and R. I. Frankel. 1980. Cryptosporidiosis, report of afatal case complicated by disseminated toxoplasmosis.Am. J. Med. 69:637-642.
58. Tyzzer, E. E. 1907. A sporozoan found in the peptic glandsof the common mouse. Proc. Soc. Exp. Biol. Med. 5:12-13.
59. Tyzzer, E. E. 1910. An extracellular coccidium Crypto-sporidium muris (gen. et sp. nov.) of the gastric glands ofthe common mouse. J. Med. Res. 23:487-509.
60. Tyzzer, E. E. 1912. Cryptosporidium parvum (sp. nov.) acoccidium found in the small intestine of the commonmouse. Arch. Protistenkd. 26:394-412.
61. Tyzzer, E. E. 1929. Coccidiosis in gallinaceous birds. Am.J. Hyg. 10:269-383.
62. Tzipori, S., K. W. Angus, and I. Campbell. 1982. Experi-mental infection of lambs with Cryptosporidium isolatedfrom a human patient with diarrhoea. Gut 23:71-74.
63. Tzipori, S., K. W. Angus, I. Campbell, and L. W. Cleri-hew. 1981. Diarrhea due to Cryptosporidium infection inartificially reared lambs. J. Clin. Microbiol. 14:100-105.
64. Tzipori, S., K. W. Angus, I. Campbell, and E. W. Gray.1980. Cryptosporidium: evidence for a single speciesgenus. Infect. Immun. 30:884-886.
65. Tzipori, S., K. W. Angus, I. Campbell, and D. Sherwood.1981. Diarrhoea in young Red Deer associated with infec-tion with Cryptosporidium. J. Infect. Dis. 144:170-175.
66. Tzipori, S., K. W. Angus, E. W. Gray, and I. Campbell.1980. Vomiting and diarrhoea associated with cryptospor-idial infection. N. Engl. J. Med. 303:818.
67. Tzipori, S., K. W. Angus, E. W. Gray, L. Campbell, and F.Allan. 1981. Diarrhoea in lambs experimentally infectedwith Cryptosporidium isolated from calves. Am. J. Vet.Res. 42:1400-1404.
68. Tzipori, S., and I. Campbell. 1981. Prevalence of Crypto-sporidium antibodies in ten species of animals. J. Clin.Microbiol. 14:455-456.
69. Tzipori, S., I. Campbell, and K. W. Angus. 1982. Thetherapeutic effect of 16 antimicrobial agents on Crypto-sporidium infection in mice. Aust. J. Exp. Biol. Med. Sci.60:187-190.
70. Tzipori, S., I. Campbell, D. Sherwood, D. R. Snodgrass,and A. Whitelaw. 1980. An outbreak of calf diarrhoeaattributed to cryptosporidial infection. Vet. Rec. 107:579-580.
71. Tzipori, S., J. Larsen, M. Smith, and R. Lugfl. 1982.Diarrhoea in goat kids attributed to Cryptosporidiuminfection. Vet. Rec. 111:35-36.
72. Tzipori, S., E. McCartney, G. H. K. Lawson, A. C. Row-land, and I. Campbell. 1981. Experimental infection ofpiglets with Cryptosporidium. Res. Vet. Sci. 31:358-368.
73. Tzipori, S., D. Sherwood, K. W. Angus, I. Campbell, andM. Gordon. 1983. Diarrhea in lambs: experimental infec-tions with enterotoxigenic Escherichia coli, rotavirus, andCryptosporidium. Infect. Immun. 33:401-406.
74. Tzipori, S., M. Smith, C. Halpin, K. W. Angus, D. Sher-wood, and I. Campbell. 1983. Experimental Cryptospori-dium in calves; clinical manifestations and pathologicalfindings. Vet. Rec., in press.
75. Tzipori, S., M. Smith, T. Makin, and C. Halpin. 1982.Enterocolitis in piglets caused by Cryptosporidium puri-fied from calf faeces. Vet. Parasitol., 11:121-126.
76. Vetterling, J. M., H. R. Jervis, T. G. Merrill, and H.Sprinz. 1971. Cryptosporidium wrairi sp. n. from theguinea pig Cavea porcellus, with an emendation of thegenus. J. Protozool. 18:243-247.
77. Vetterling, J. M., A. Takeuchi, and P. A. Madden. 1971.Ultrastructure of Cryptosporidium wrairi from the guineapig. J. Protozool. 18:248-260.
78. Weinstein, L., S. M. Edelstein, J. L. Madara, K. R. Fal-chuk, B. M. McManus, and J. R. Trier. 1981. Intestinalcryptosporidiosis complicated by disseminated cytomega-lovirus infection. Gastroenterology 81:584-591.
79. Welsburger, W. R., D. F. Hutcheon, J. H. Yardley, J. C.Roche, W. D. Hills, and P. Charache. 1979. Cryptospori-diosis in an immunosuppressed renal transplant recipientwith IgA deficiency. Am. J. Clin. Pathol. 72:473-378.
MICROBIOL. REV.
on May 19, 2018 by guest
http://mm
br.asm.org/
Dow
nloaded from