current protocols in cell biology || differentiation of mouse embryonic stem cells and of human...
TRANSCRIPT

UNIT 23.4Differentiation of Mouse Embryonic StemCells and of Human Adult Stem Cells intoAdipocytes
Severe obesity is the result of increases in fat cell size and increased fat cell number.Mature adipocytes do not undergo cell division, and new fat cells arise from a pre-existing pool of adipose stem cells that are present irrespective of age. The developmentof established preadipose cell lines has facilitated the study of different steps leadingto terminal differentiation of preadipocytes into adipocytes. The key events have beencharacterized by the identification of transcription factors that play a regulatory role in thedifferentiation process. The best characterized transcription factors shown to be importantin the development of mature adipocytes are members of the CCAAT/enhancer bindingproteins (C/EBPs) and peroxisome proliferative-activated receptors (PPARs) families(Tontonoz et al., 1995; Mandrup and Lane, 1997). However, established preadipose celllines are limited for studying early events of differentiation as they represent cells that arealready committed to the adipogenic lineage. Master genes that commit multipotent stemcells toward adipocytes remain to be identified. In vitro differentiation of multipotentstem cells to the adipogenic lineage provides an alternative source of adipocytes for studyin tissue culture and offers the possibility to investigate regulation of the first steps ofdifferentiation and to test effects of drugs on adipose cell development.
This unit includes a protocol for culture conditions in which mouse embryonic stem(mES) cells can be maintained at an undifferentiated state (Support Protocol 1) or com-mitted to undergo adipocyte differentiation at a high rate and in a highly reproduciblefashion (Basic Protocol 1). There are also protocols for maintaining (Support Protocol 2)and differentiating human adult stem cells, isolated form adipose tissue (hMADS cells)and from bone marrow (hMS cells), into adipocytes (Basic Protocol 2). These culturesystems provide a powerful means for studying the first step of adipose cell developmentand the effects of drugs on the biology of adipocytes. There are also protocols for visu-alization of adipocytes (Support Protocol 3) and analysis of adipocyte gene expression(Support Protocol 4).
NOTE: All solutions and equipment coming into contact with living cells must be sterile,and aseptic technique should be used accordingly.
NOTE: All culture incubations should be performed in a humidified 37◦C, 5% CO2
incubator unless otherwise specified.
BASICPROTOCOL 1
DIFFERENTIATION OF MOUSE ES CELLS TO ADIPOCYTES
Adipocyte differentiation of mES cells is initiated by aggregation of ES cells to formembryoid bodies (EBs). The authors routinely use the hanging drop method for theformation of EBs. Two phases can be distinguished in adipogenesis from ES cells.The first phase, between day 2 and 5 after EB formation, corresponds to the permissiveperiod for the commitment of ES cells; this phase requires retinoic acid (RA). The secondphase corresponds to the permissive period for terminal differentiation and is influencedby adipogenic factors (e.g., insulin, triiodothyronine, and rosiglitazone) as previouslyshown for the differentiation of cells from preadipose clonal lines. The treatment leadsto 60% to 80% of outgrowths containing adipose cells compared to 2% to 5% in theabsence of RA treatment.
Contributed by Brigitte Wdziekonski, Phi Villageois, and Christian DaniCurrent Protocols in Cell Biology (2007) 23.4.1-23.4.14Copyright C© 2007 by John Wiley & Sons, Inc.
Stem Cells
23.4.1
Supplement 34

Differentiation ofStem Cells into
Adipocytes
23.4.2
Supplement 34 Current Protocols in Cell Biology
Materials
ES cellsGrowth medium for mES cells (see recipe)Leukemia inhibitory factor (LIF)CMF-PBS (phosphate-buffered saline, calcium and magnesium free; Cambrex)Trypsin solution (see recipe)Fetal bovine serum (FBS, see recipe)Retinoic acid (RA, see recipe)Differentiation medium for mES cells (see recipe)
60- and 100-mm bacteriological grade petri dishes (Greiner)20-ml conical sterile tube100-mm gelatinized-tissue culture dishes (see recipe)
Additional reagents and equipment for cell counting (UNIT 1.1)
Collect ES cellsES cells are originally plated at 106 cells per 25-cm2 flask and are grown for 2 or 3 daysuntil they reach a confluency of 80%. At this point, cells are still maintained from theoriginal plating in the presence of LIF, in order to keep ES cells in the undifferentiatedstate. ES cell cultures may contain a proportion of “differentiated” cells which have losttheir pluripotency. It is crucial to minimize this proportion of differentiated cells. Thisis achieved by the addition of LIF in a high-quality culture medium, i.e., an adequatebatch of serum. Identification of pluripotent stem cells is difficult unless one is familiarwith the appropriate cellular morphology. Pluripotent stem cells: (1) are small, (2) have alarge nucleus containing prominent nucleoli structures, and (3) have minimal cytoplasm.Pluripotent stem cells, in contrast to differentiated cells, grow rapidly.
1. Change medium on ES cells with complete growth medium supplemented with1000 U/ml LIF 2 hr before subculture.
2. Aspirate medium and wash twice, each time with 5 ml of CMF-PBS.
3. Aspirate the CMF-PBS and add 1 ml of trypsin solution. Incubate 2 to 3 min.
4. Add 5 ml of growth medium to stop trypsinization and suspend the cells by vigorouspipetting. Transfer the cells to a sterile 20-ml tube and centrifuge 5 min at 250 × g,room temperature.
5. Aspirate the medium and resuspend the cell pellet in 10 ml of growth mediumwithout LIF to prepare a single-cell suspension. Count the cells (UNIT 1.1). After cellcounting, adjust the suspension to a concentration of 5 × 104 cells/ml.
Induce EB formation6. Place aliquots of 20 µl of this suspension onto the lids of 10-cm bacteriological grade
dishes.
This is defined as day 0 of EB formation.
Bacterial grade petri dishes are used to prevent cell attachment to the substrate. EBs havea tendency to attach to the bottom of the plastic dish. EBs that are firmly attached to thedish should be eliminated as these EBs seem to have no adipogenic capacity.
7. Invert the lid and place it over the bottom of a bacteriological petri dish filled with8 ml CMF-PBS containing a few drops of FBS to decrease the surface tension of theliquid.
It is essential to cover the bottom of the dish with the liquid to prevent the evaporationof the hanging drops. When the lid is inverted, each drop hangs and the cells fall to thebottom of the drop where they aggregate into a single clump (EB).
Stem Cells
23.4.3
Current Protocols in Cell Biology Supplement 34
Attachment of EBs on petri dishes can be eliminated by coating the dish with poly (2-hydroxyethylmethylacrylate) supplied as Cellform polymer (ICN Biomedicals). Dishesare filled with a film of Cellform working solution (0.5 g of Cellform dissolved in 42 ml of100% ethanol) and left in a culture hood, with the cover of the dish removed, to evaporateethanol. Dishes are rinsed with CMF-PBS before use.
8. At a time point 2 days later, remove the lid, invert it and collect drops containingEBs in a 20-ml conical sterile tube. Let stand for 5 min at room temperature to allowthe aggregates to sediment.
9. Aspirate the supernatant and resuspend the pellet in 4 ml of growth medium sup-plemented with 10−7 M retinoic acid (RA). Transfer the suspension to 60-mmbacteriological grade petri dishes.
RA is light-sensitive.
10. Incubate for 3 days in the presence of RA changing the growth medium every day.
Plate EBs on gelatin and differentiate.
Induce differentiation11. At day 5 after EB formation, change the medium to growth medium without the
addition of RA and plate 2 to 4 EBs per cm2 in 100-mm gelatinized-tissue culturedishes.
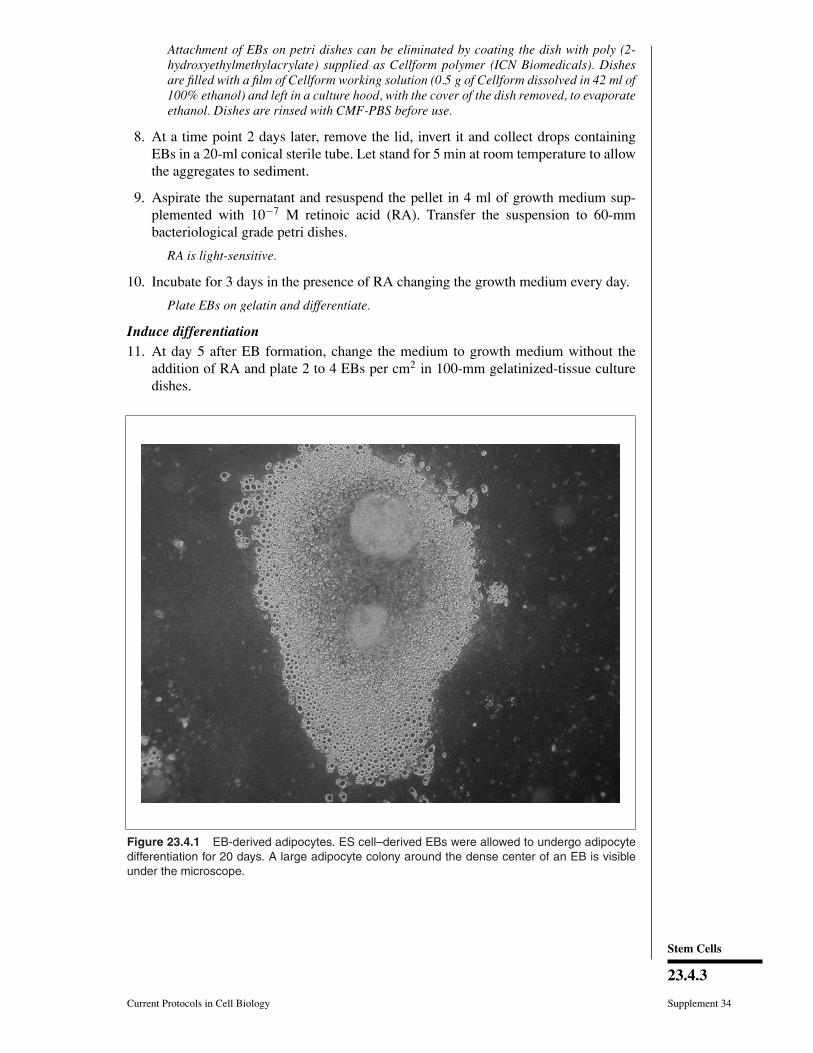
Figure 23.4.1 EB-derived adipocytes. ES cell–derived EBs were allowed to undergo adipocytedifferentiation for 20 days. A large adipocyte colony around the dense center of an EB is visibleunder the microscope.

Differentiation ofStem Cells into
Adipocytes
23.4.4
Supplement 34 Current Protocols in Cell Biology
12. The day after plating, change the growth medium to differentiation medium. Changemedium every other day.
The addition of 0.5 µM rosiglitazone, which is a PPARγ activator (Lehmann et al.,1995), to the differentiation medium dramatically stimulates the terminal differentiationof RA-treated EBs into adipocytes (Phillips et al., 2003).
13. After 10 to 20 days in the differentiation medium, evaluate the cultures for adipocytecolonies (see Fig. 23.4.1).
After 10 to 20 days at least 50% to 70% of EB outgrowths should contain adipocytecolonies.
SUPPORTPROTOCOL 1
MAINTENANCE OF MOUSE ES CELLS WITHOUT FEEDER LAYERS
The conditions outlined below are applicable for the maintenance of feeder layer–independent ES cell lines. Cells can be grown on gelatin-coated tissue culture flasksand maintained in a multipotent undifferentiated state by addition of leukemia inhibitoryfactor (LIF; Smith, 1992) to the growth medium.
Materials
ES cells in 25-cm2 flasks (up to passage 25)CMF-PBS (phosphate-buffered saline, calcium and magnesium free; Cambrex)1× trypsin solution (see recipe)Growth medium for mES cells (see recipe)Leukemia inhibitory factor (Chemicon, Invitrogen)
20-ml conical centrifuge tubes, sterileGelatinized tissue culture 25-cm2 flasks (see recipe)
Additional reagents and equipment for cell counting (UNIT 1.1)
1. For a 25-cm2 flask containing ES cells, aspirate medium and wash twice each timewith 5 ml CMF-PBS.
2. Aspirate the CMF-PBS and add 1 ml of 1× trypsin solution. Ensure the trypsincovers the cell monolayer and incubate for 2 to 3 min. Check that the cells aredissociated in single-cell suspension under an inverted microscope.
It is critical to produce a single-cell suspension for subcultures. This is achieved by tappingthe flask several times to ensure complete dissociation during the trypsin treatment.
3. Add 5 ml of growth medium to stop trypsinization and suspend the cells by vigorouspipetting.
4. Transfer the cells to a sterile 20-ml tube and centrifuge 5 min at 250 × g, roomtemperature.
5. Aspirate the medium and resuspend the cell pellet in 5 ml of growth medium bypipetting up and down two to three times. Count cells (UNIT 1.1).
6. Add 106 cells to 10 ml of prewarmed growth medium containing 1000 U/ml LIFthen transfer to a freshly gelatinized 25-cm2 flask.
LIF is required to maintain pluripotent ES cells and is omitted to induce the commitmentof ES cells towards the adipogenic lineage.
7. Change growth medium with LIF (1000 U/ml) every day.
8. Trypsinize the cultures 2 days later as in step 2.
Cultures should be subcultured before cells have reached confluence.
Stem Cells
23.4.5
Current Protocols in Cell Biology Supplement 34
BASICPROTOCOL 2
DIFFERENTIATION OF HUMAN MULTIPOTENT ADIPOSE-DERIVEDSTEM (hMADS) CELLS TO ADIPOCYTES
Adipocyte differentiation of human multipotent adipose-derived stem (hMADS) cells andhuman mesenchymal stem (hMS) cells in vitro does not require pretreatment with RA.Induction of differentiation of hMADS cells and hMS cells is performed in monolayerculture. However, in contrast to hMADS cells, differentiation of hMS into adipocyterequires a differentiation medium supplemented with 10% fetal bovine serum (FBS).
Materials
Adult stem cells: hMADS (for isolation of hMADS cells see Rodriguez et al.,2005a; Zaragosi et al., 2006)
Growth medium for hMADS cells (see recipe)Differentiation medium for hMADS cells (see recipe)Tissue culture 100-mm dishes (Greiner, S.A. Dutscher) or 6-well plates or 12-well
plates
1. Plate adult stem cells at a high density, e.g., 40,000 cells/cm2 in 10 ml, 2 ml, or1 ml growth medium, on tissue culture 100-mm dishes or 6- or 12-well plates,respectively.
2. When cells reach confluence (2 or 3 days after plating), change growth medium todifferentiation medium containing 0.5 µM rosiglitazone.
The addition of 0.5 µM rosiglitazone in the differentiation medium is required to differ-entiate cells into adipocytes.
3. Three days later, change cells to differentiation medium without IBMX anddexamethasone.
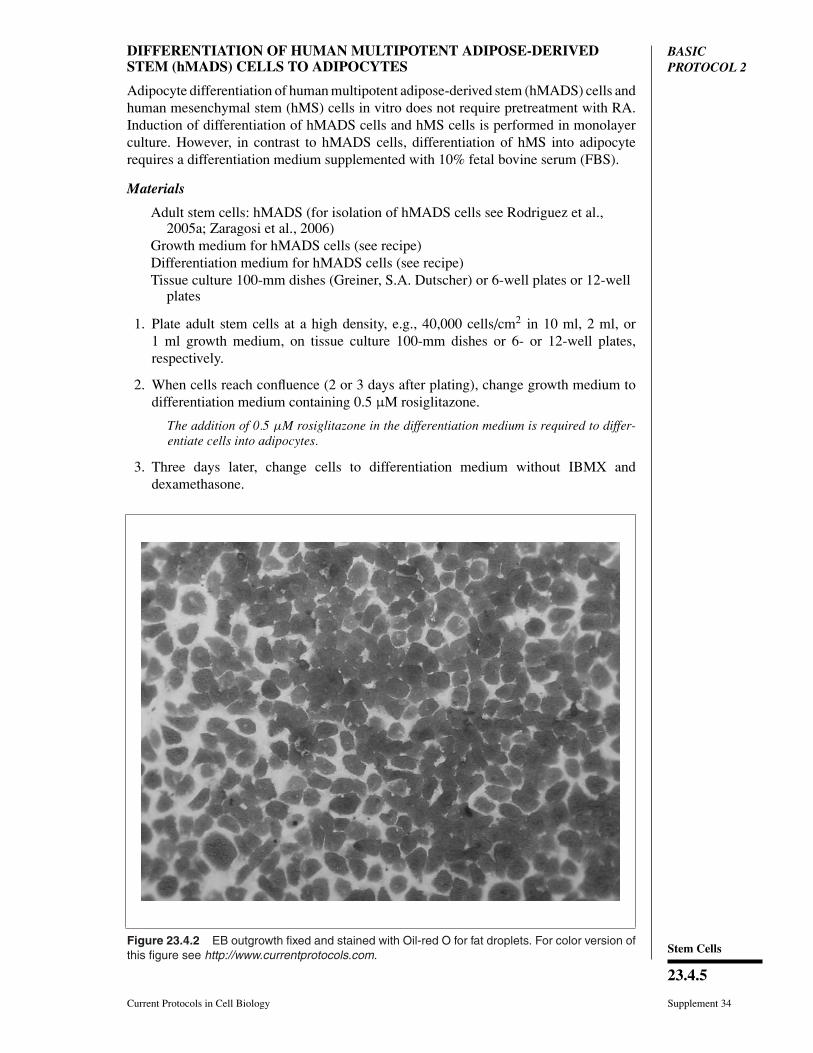
Figure 23.4.2 EB outgrowth fixed and stained with Oil-red O for fat droplets. For color version ofthis figure see http://www.currentprotocols.com.

Differentiation ofStem Cells into
Adipocytes
23.4.6
Supplement 34 Current Protocols in Cell Biology
4. Check the cultures for the appearance of adipocytes.
Adipose cells contain lipid droplets which can be easily visualized microscopically, espe-cially under bright-field illumination. Lipid droplets can also be visualized after stainingwith Oil-red O as indicated in Figure 23.4.2.
Adipocytes should appear 5 to 7 days after induction of differentiation.
ALTERNATEPROTOCOL
DIFFERENTIATION OF HUMAN MS CELLS TO ADIPOCYTES
The same growth medium is used for hMADs cells and for hMS cells (see Reagents andSolutions). In contrast, the differentiation medium is different because hMS cells require10% FBS to undergo differentiation into adipocytes, whereas adipocyte differentiationof hMADS cells occurs under a serum-free condition.
Materials
Adult stem cells: hMS cells (for isolation of adult stem cells from bone marrow seePittenger et al., 1999)
Growth medium for hMS cells (see recipe)Differentiation medium for hMS cells (see recipe)Tissue culture 100-mm dishes or 6-well plates or 12-well plates (Greiner; S.A.
Dutscher)
1. Plate cells at a high density, e.g. 40,000 cells/cm2 in growth medium on tissue culture100-mm dishes (or 6- or 12-well plates).
2. When cells reach confluence (2 or 3 days after plating), change growth medium todifferentiation medium, containing 0.5 µM rosiglitazone.
The addition of 0.5 µM rosiglitazone in the differentiation medium is required to differ-entiate cells into adipocytes
3. Three days later, change cells with differentiation medium without IBMX and dex-amethasone.
4. Check the cultures for the appearance of adipocytes.
Adipose cells contain lipid droplets which can be easily visualized microscopically, espe-cially under bright-field illumination. Lipid droplets can also be visualized after stainedwith Oil-red O as indicated in Figure 23.4.2.
Adipocytes should appear 5 or 7 days after induction of differentiation.
SUPPORTPROTOCOL 2
MAINTENANCE OF HUMAN MADS AND HUMAN MS CELLS WITHOUTFEEDER LAYER
No feeder layer and no gelatin-coated dishes are required to maintain the proliferation ofhMADs cells and of hMS cells. The same growth medium is used to maintain both celltypes.
Materials
hMADS or hMS cells (for isolation of hMADS see Rodriguez et al., 2005a; forisolation of hMS cells see Pittenger et al., 1999)
Phosphate-buffered saline, calcium and magnesium free (CMF-PBS, Cambrex)1× trypsin -EDTA solution (Invitrogen no:25300-054)Growth medium for hMADS and hMS cells (see recipe)
Tissue culture 100-mm dishes (Greiner; S.A. Dutscher)20-ml conical centrifuge tubes, sterile
Additional reagents and equipment for cell counting (UNIT 1.1)

Stem Cells
23.4.7
Current Protocols in Cell Biology Supplement 34
1. For a 100-mm dish of hMADS or hMS cells, aspirate medium and wash twice eachtime with 10 ml CMF-PBS.
2. Aspirate the CMF-PBS and add 1 ml of 1× trypsin solution. Ensure the trypsincovers the cell monolayer and incubate for 2 to 3 min. Check that cells are detachedfrom the bottom of the dish by examining under an inverted microscope.
3. When the cells are detached, add 10 ml of growth medium to stop trypsinization andsuspend the cells by pipetting.
4. Transfer the cells to a 20-ml sterile tube and centrifuge 5 min at 250 × g, roomtemperature.
5. Aspirate the medium and resuspend the cell pellet with 5 ml of growth medium bypipetting up and down 2 to 3 times. Count cells (UNIT 1.1).
6. Add 250 × 103 cells to 10 ml of prewarmed growth medium then transfer to a100-mm dish.
7. Change medium every other day.
8. Trypsinize the cultures 3 to 4 days later as in step 1.
Cultures should be subcultured before cells have reached confluence.
SUPPORTPROTOCOL 3
VISUALIZATION OF ADIPOCYTES
Adipose cells with lipid droplets are easily visualized microscopically, especially underbright-field illumination. Nonadipose cells containing structures resembling droplets areoften detectable in untreated- and RA-treated ES cultures. Therefore, it is essential toidentify droplet-like structures as triglyceride droplets. Staining of cultures with Oil-redO, a specific stain for triglycerides, gives a good indication of adipocyte differentiation.
Materials
Cultures to be examinedCMF-PBS (phosphate-buffered saline, calcium and magnesium free; Cambrex)Fixation buffer (see recipe)Oil-red O solution (see recipe)Storage solution: 70% glycerol (v/v) in H2OOil-red O elution buffer (see recipe)Spectrophotometer
1. Aspirate medium from cultures and wash cells once with 10 ml CMF-PBS.
2. Fix cells for 15 min in 10, 2, or 1 ml fixation buffer in 100-mm dishes, 6- or 12-wellplates, respectively, at room temperature.
3. Wash twice for 10 min each time with 10 ml Milli-Q water (for a 100-mm dish).
4. Stain with 10 ml Oil-red O solution for 15 min (for a 100-mm dish; stain with 2 mlor 1 ml Oil-red O for a 6- or 12-well dish, respectively).
5. Wash twice each time with 10 ml Milli-Q water. Cover cells with a film of storagesolution (see Fig. 23.4.2) or elute the bound stain (step 6).
Cells covered with storage solution can be kept at room temperature for several months.
6. Elute bound stain to quantify the formation of adipocytes. Incubate the 100-mm dishwith 10 ml Oil-red O elution buffer (see reicpe) for 30 min at room temperature.Remove the solution.

Differentiation ofStem Cells into
Adipocytes
23.4.8
Supplement 34 Current Protocols in Cell Biology
7. After elution of the bound stain, measure the amount of bound Oil-red O spectropho-tometrically at 490 nm.
SUPPORTPROTOCOL 4
ANALYSIS OF ADIPOCYTE GENE EXPRESSION
Finally, expression of a-FABP (adipocyte-fatty acid binding protein) an adipocyte-specific gene, in 20-day-old ES cell–derived EB or in 10-day-old differentiated humanstem cells can be detected by northern blotting using 20 µg of total RNA. However,detection of the expression of these genes in early differentiating cells requires a moresensitive method such as RT-PCR. This protocol gives conditions to reveal mouse andhuman a-FABP gene expression.
Materials
TRI Reagent (Molecular Research Center, Euromedex, France)ChloroformIsopropanol100% ethanol5 M NaCl, sterileTES buffer (see recipe)RT-PCR kit (available from several companies)Primers to reveal mouse a-FABP gene expression:
forward: 5′-GATGCCTTTGTGGGAACCTGG-3′reverse: 5′-TTCATCGAATTCCACGCCCAG-3′
Mouse hypoxanthine phosphoribosyltransferase (HPRT, as a standard to balancethe amount of RNA and cDNA used):forward: 5′-GCTGGTGAAAAGGACCTCT-3′reverse: 5′-CACAGGACTAGAACACCTGC-3′
Primers to reveal human a-FABP gene expression:forward: 5′-GCTTTGCCACCAGGAAAGTG-3′reverse: 5′-ATGACGCATTCCACCACCAG-3′
Human β-actin as a housekeeping gene standard:forward: 5′-AGCCATGTACGTTGCTA-3′reverse: 5′-AGTCCGCCTAGAAGCA-3′
RNAs are prepared using TRI Reagent according to the supplier’s protocol.
Days of differentiation are indicated as 20 days for ES cells and 10 days for AS cells.The authors routinely obtain 100 to 200 µg RNA from one 100-mm tissue culture platecontaining 20-day-old EB outgrowths or 10-day-old hMADS cells and hMS cells.
Using the abovementioned primers to reveal mouse a-FABP gene expression PCR isperformed using an annealing temperature of 56◦C and 25 cycles of PCR. The size ofthe expected cDNA is 213 bp. Mouse hypoxanthine phosphoribosyltransferase (HPRT)is used as a standard to balance the amount of RNA and cDNA used. With the above-mentioned primers PCR is performed using an annealing temperature of 60◦C and 25cycles of PCR. The expected size of this cDNA is 249 bp.
Using the abovementioned primers to reveal human a-FABP gene expression PCR isperformed using an annealing temperature of 55◦C and 25 cycles of PCR. The size of theexpected cDNA is 290 bp. Human β-actin is used as a standard. With the abovementionedprimers PCR is performed using an annealing temperature of 55◦C and 25 cycles of PCR.The expected size of this cDNA is 656 bp.
PCR products are separated and visualized by electrophoresis on a 2% (w/v) agarose gelcontaining ethidium bromide (see Voytas, 2000).

Stem Cells
23.4.9
Current Protocols in Cell Biology Supplement 34
Figure 23.4.3 PCR analysis of ex-pression of an adipocyte-specific gene.EBs were pretreated from day 2 to day5 with 0.1% DMSO (1) or 10−7 RA (2).RNAs were prepared at day 20 after EBformation and transcripts for a-FABP.HPRT was used to monitor expressionof a housekeeping gene.
REAGENTS AND SOLUTIONSUse deionized or distilled water in all recipes and protocols except as noted. For common stocksolutions, see APPENDIX 2A; for suppliers, see SUPPLIERS APPENDIX.
Dexamethasone, 1 mM
Dissolve 3.9 mg dexamethasone (powder; Sigma) in 10 ml ethanol. Store 0.5-mlaliquots up to 6 months at −20◦C.
Differentiation medium for hMADS cells
To 25 ml of low-glucose DMEM (Invitrogen) add:25 ml Ham’s F12 (Cambrex)0.5 ml 200 mM glutamine0.5 ml 1 M HEPES-buffered saline (Cambrex)0.5 ml 5000 IU/ml/5000 µg/ml penicillin/streptomycin (Cambrex)0.5 ml 1 mg/ml transferrin0.25 ml 1 mg/ml insulin (see recipe)0.05 ml 100 mM IBMX (see recipe)0.05 ml 1 mM dexamethasone (see recipe)0.005 ml 2 µM T3 (see recipe)Prepare fresh
The differentiation medium must be prepared just before use. Therefore, prepare only thevolume of medium required for changing cells.
Differentiation medium for hMS cells
To 45 ml low-glucose DMEM (Invitrogen) add:5 ml FBS0.5 ml 200 mM glutamine0.5 ml 1 M HEPES-buffered saline (Cambrex)0.5 ml 5000 IU/ml/5000 µg/ml penicillin/streptomycin (Cambrex)0.05 ml 100 mM IBMX (see recipe)0.05 1 mM dexamethasone (see recipe)0.25 1 mg/ml insulin (see recipe)0.005 ml 2 µM T3 (see recipe)Prepare fresh
The differentiation medium must be prepared just before use. Therefore, prepare only thevolume of medium required for changing cells.

Differentiation ofStem Cells into
Adipocytes
23.4.10
Supplement 34 Current Protocols in Cell Biology
Differentiation medium for mES cells
Supplement growth medium for mES cells with:0.5 µg/ml insulin (see recipe)2 nM triiodothyronine (add from 2 µM T3 solution; see recipe)0.5 µM rosiglitazone (provided by GlaxoSmthKline)Prepare fresh
The differentiation medium must be prepared just before use. Therefore, prepare only thevolume of medium required for changing cells.
Fetal bovine serum
Fetal bovine serum is tested for maintenance of the undifferentiated state. The au-thors select a batch of serum that is able to support the growth of stem cells. For thatpurpose, plate 106 ES cells/25-cm 2 flask in 10% of each set of FBS, supplementedwith LIF, and subculture the cells every two days for 4 passages. For a high-qualityserum, a flask should yield 5 to 10 × 106 cells at each passage. Furthermore, notoxicity of the selected serum should be observed at a 30% concentration.
The supplier used for this unit is Dutscher, France, but other suppliers can be used.
Fixation buffer
Dilute 2.5% glutaraldehyde (Sigma) to 0.25% (v/v) in CMF-PBS (Cambrex). Storeup to 1 month at 4◦C.
Gelatin 0.1% (w/v)
Dilute 2% (w/v) gelatin to 0.1% with CMF-PBS (Cambrex). Store up to 2 weeksat 4◦C.
Gelatinized-tissue culture dishes/flasks
Add 5 or 10 ml of 0.1% gelatin (see recipe) per 25-cm2 flask (Corning or Greiner) or100-mm tissue culture dishes (Corning or Greiner Cell Star), respectively. Incubatefor at least 10 min at room temperature and aspirate gelatin. Prepare fresh.
Growth medium for hMADS and hMS cells
To 500 ml low-glucose DMEM add:5 ml 200 mM glutamine5 ml 1 M HEPES -buffered saline (Cambrex)5 ml 5000 IU/ml-/5000 µg/ml penicillin/streptomycin50 ml FBSStore up to 2 weeks at 4◦C.
Growth medium for mES cell maintenance, 1×To 440 ml Glasgow MEM/BHK21 medium (Invitrogen) add:5 ml of 100× nonessential amino acids (stored at 4◦C)5 ml of 200 mM glutamine (stored at −20◦C)5 ml 100 mM sodium pyruvate (stored at −20◦C)0.5 ml 0.1 M 2 -mercaptoethanol (stored at 4◦C; see recipe)50 ml of FBS (stored at −20◦C)5 ml 5000 IU/ml/5000 µg/ml penicillin/streptomycin (Cambrex)Store up to 2 weeks at 4◦C

Stem Cells
23.4.11
Current Protocols in Cell Biology Supplement 34
IBMX, 100 mM
Prepare IBMX (3-isobutyl-1methylxanthine; Sigma) at 100 mM in water. Dissolve250 mg IBMX in 9 ml water and 5 to 10 µl of 10 N NaOH. Adjust volume to 11 mlwith water and sterilize by filtration with a 0.22-µm filter. Store 0.1-ml aliquots upto 2 months at −20◦C.
Insulin (for mES cells)
Dissolve lypholized insulin, —bovine for mouse cells; recombinant human (Sigmano. I-9278) for human cells—at 1 mg/ml in cold 0.01 N HCl. Mix gently andsterilize by filtration with a 0.22-µm filter. Store 0.1-ml aliquots up to 6 months at−20◦C. Store thawed aliquots up to 2 weeks at 4◦C.
2-mercaptoethanol, 0.1 M
100 µl 2-mercaptoethanol14 ml sterile H2OStore up to 3 weeks at 4◦C
Oil-red O elution buffer
4 M guanidinium thiocyanate (Sigma)25 mM sodium citrate, pH 70.5% (v/v) sarcosyl20% (v/v) isopropanolStore up to 1 month at room temperature
Oil-red O solution
Oil-red O stock solution: Dissolve Oil red O (Sigma) at 0.5% (w/v) in isopropanol.Store up to several months at room temperature.
Oil red O working solution: Mix 6 vol of stock solution with 4 vol water. Mix andfilter using a 0.45-µm filter. Store up to 1 month at room temperature.
Retinoic acid
Dilute all-trans retinoic acid (RA; Sigma) in the dark in dimethyl sulfoxide toprepare a 10 mM RA stock solution. Store 0.1-ml aliquots up to 1 year at −20◦C,protected from the light.
Prepare subsequent dilutions of RA in ethanol and use for one experiment only.
After dilution into the culture medium, the concentration of ethanol should neverexceed 0.1%.
TES buffer
10 mM Tris·Cl, pH 7.4 (APPENDIX 2A)0.1 mM EDTA0.1% (w/v) sodium dodecyl sulphateSterilize by autoclavingStore up to several months at room temperature
Transferrin, 1 mg/ml
Dissolve apo-transferrin (human; Sigma) at 1 mg/ml in PBS (APPENDIX 2A) andsterilize by filtration with a 0.22-µm filter. Store 1-ml aliquots up to several monthsat −20◦C. Store thawed aliquots up to 1 week at 4◦C.

Differentiation ofStem Cells into
Adipocytes
23.4.12
Supplement 34 Current Protocols in Cell Biology
Triiodothyronine (T3), 2 µM
Dissolve 5 mg T3 (culture tested, powder; Sigma) into 7.43 ml ethanol to preparea 1 mM T3 solution. Then, dilute 200 µl of 1 mM solution into 4 ml ethanol toprepare a 50 µM T3 solution. Finally, dilute 3.2 ml of 50 µM T3 solution into 80 mlethanol to prepare a 2 µM T3 solution. Store up to 1 year at −20◦C.
Trypsin solution, 1× for mES cells
1 ml 2.5% (w/v) trypsin (Invitrogen)1 ml 100 mM EDTA1 ml chicken serum100 ml CMF-PBS (APPENDIX 2A)Store 10-ml aliquots up to 2 months at −20◦CStore thawed aliquots up to 2 weeks at 4◦C
COMMENTARY
Background InformationMouse embryonic (ES) cells have been pre-
viously shown to differentiate spontaneouslyinto various lineages in culture (Doetschmanet al., 1985). The first morphological observa-tion of adipocyte-like cells derived from EScells was reported by Field et al. (1992). How-ever, the number of ES cell–derived adipocyteswas low because spontaneous commitment ofES cells to the adipogenic lineage is rare. Inorder to use ES cells to study adipocyte de-velopment it was essential to determine condi-tions of culture that committed ES cells to theadipogenic lineage at a high rate. This step hasbeen achieved by showing that a prerequisiteis to treat ES cell–derived embryoid bodies(EBs) at an early stage of their differentiationwith all-trans-retinoic acid (RA) for a shortperiod of time (Dani et al., 1997; Dani, 1999).Adipocytes derived from ES cells display bothlipogenic and lipolytic activities in responseto insulin and to β-adrenergic agonists respec-tively, indicating that mature and functionaladipocytes are formed.
In regard to models of human cellspresently available, for primary preadipocytesderived from stromal-vascular cells of hu-man adipose tissue, with increasing pas-sage number, the ability to differentiate intoadipocytes under appropriate conditions un-dergoes a dramatic decrease before the cellsbecome growth arrested and enter replicativesenescence (Ailhaud, 2002). This limitationhas been partly circumvented with cells thatare immortalized either genetically or sponta-neously while preserving their ability to un-dergo adipose conversion. Overexpression ofSV40T and T antigens in human SV cells from
infant brown adipose tissue (Zilberfarb et al.,1997) or coexpression of human telomerasereverse transcriptase and papilloma E7 onco-protein in SV cells from adult white adiposetissue (Darimont et al., 2003) led to establish-ment of PAZ6 and Chub-S7 preadipocyte celllines, respectively. Cells isolated from subcu-taneous adipose tissue of an infant sufferingfrom Simpson-Golabi-Behmed syndrome andfrom an adult suffering from liposarcoma wereused to obtain SGBS preadipocyte cell strainand LiSa-2 preadipocyte cell line, respectively(Wabitsch et al., 2000, 2001). Upon differ-entiation, cells from these various lines ex-pressed some of the characteristic markers ofhuman adipocytes but the lipolytic responsesspecific of human adipocytes and the secre-tion of specific adipocytokines were not re-ported. Moreover, except for Chub-S7 cellswhich were diploid and showed no chromo-somal alterations, cells from other lines exhib-ited chromosomal abnormalities. Recently, theestablishment of multipotent stem cells fromthe stromal-vascular fraction of infant adiposetissue (Rodriguez et al., 2005a) has led the au-thors to examine, under serum-free adipogenicconditions, whether these human multipotentadipose-derived stem (hMADS) cells enter theadipose lineage and differentiate into cells thatexhibit characteristics of human fat cells. Theauthors have shown that hMADS cells are aunique cell model in displaying the key fea-tures of human adipocytes after differentiationunder adipogenic conditions (Rodriguez et al.,2004, 2005b). The human adult bone marrowalso contains mesenchymal stem (hMS) cellsthat are able to undergo differentiation intoadipocytes in vitro (Pittenger et al., 1999).

Stem Cells
23.4.13
Current Protocols in Cell Biology Supplement 34
Critical Parameters andTroubleshooting
The first critical parameter for the commit-ment of mES cells into adipocytes is the forma-tion of EBs. The authors use the hanging dropmethod for the formation of EBs. The forma-tion of EBs in mass culture by maintaining EScells in suspension at a high density in non-tissue culture grade plastic is rapid and givesrise to a high number of EBs formed. Howeverin the hands of the authors, this method leadssubsequently to a low number of outgrowthscontaining adipocyte colonies.
The, treatment of EBs with RA betweenday 2 and 5 after EB formation is prerequi-site. The authors have observed that treatmentof EBs with RA between days 2 and 5 afterEB formation or between days 3 and 6 givessimilar results. Owing to the high instabilityof RA, the concentration of RA able to com-mit ES cells into the adipogenic lineage at ahigh rate should be determined for each newpreparation of RA (try 10−8 to 10−6 M). RAis light-sensitive.
Adipocyte differentiation of hMADS cellsand hMS cells in vitro does not require pre-treatment with RA. However, activators ofPPARγ such as rosiglitazone, is required forthe terminal differentiation of these stem cellsinto adipocytes.
hMS cells can be expanded for several pas-sages in vitro. However, in the hands of the au-thors these cells lose their capacity to undergodifferentiation after 15 passages. In contrast,hMADS cells can be maintained for 30 pas-sages with no loss of their ability to undergoadipocyte differentiation.
Anticipated ResultsThe authors usually pool 2-day-old EBs
from four lids of 100-mm bacteriological petridishes into one 60-mm bacteriological gradepetri dish. After RA treatment, EBs containedin one 60-mm dish are plated into one 100-mmtissue culture dish. Under these conditions de-velopment of more than 100 EBs can be ob-served. Twenty days after EB formation, 50%to 70% of EBs contain adipocyte colonies.Over 25 passages, adipocyte differentiationmay decrease. In regard to the differentiationof adult stem cells, the authors usually performdifferentiation in 24-well plates when mon-itoring differentiation by Oil-red O staining,and into 60-mm plates, or larger plates, forRNA preparation. Ten days after induction ofdifferentiation, 50% to 90% of hMADS andhMS cells are differentiated into adipocytes.
The level of differentiation may decrease over25 passages of hMADs cells and over 10 pas-sages of hMS cells.
Time ConsiderationsFrom mES, adipocytes can be observed
10 days after EB formation. However, 20 daysin differentiation medium are required to com-plete adipocyte differentiation. Additions ofPPARγ activator dramatically increase the ki-netics and the level of adipocyte differentiationof ES cells. From human adult stem cells, adi-pogenesis is completed within 2 weeks. In thatcase PPARγ activator is required.
Literature CitedAilhaud, G. 2002. Autocrine/paracrine effectors of
adipogenesis. Ann. Endocrinol. (Paris) 63:83-85.
Dani, C. 1999. Embryonic stem cell-derivedadipogenesis. Cells Tissues Organs 165:173-180.
Dani, C., Smith, A., Dessolin, S., Leroy, P., Staccini,L., Villageois, P., Darimont, C., and Ailhaud, G.1997. Differentiation of embryonic stem cellsinto adipocytes in vitro. J. Cell Sci. 110:1279-1285.
Darimont, C., Zbinden, I., Avanti, O., Leone-Vautravers, P., Giusti, V., Burckhardt, P., Pfeifer,A.M., and Mace, K. 2003. Reconstitution oftelomerase activity combined with HPV-E7 ex-pression allow human preadipocytes to preservetheir differentiation capacity after immortaliza-tion. Cell Death Differ. 10:1025-1031.
Doetschman, T.C., Eistetter, H., Katz, M., Schmidt,W., and Kemler, R. 1985. The in vitro devel-opment of blastocyst-derived embryonic stemcell lines: Formation of visceral yolk sac, bloodislands and myocardium. J. Embryol. Exp.Morphol. 87:27-45.
Field, S.J., Johnson, R.S., Mortensen, R.M.,Papaioannou, V.E., Spiegelman, B.M., andGreenberg, M.E. 1992. Growth and differenti-ation of embryonic stem cells that lack an in-tact c-fos gene. Proc. Natl. Acad. Sci. U.S.A.89:9306-9310.
Lehmann, J.M., Moore, L.B., Smith, O.T.,Wilkison, W.O., Willson, T.M., and Kliewer,S.A. 1995. An antidiabetic thiazolidinedione is ahigh affinity ligand for peroxisome proliferator-activated receptor gamma (PPAR gamma).J. Biol. Chem. 270:12953-12956.
Mandrup, S. and Lane, M.D. 1997. Regulating adi-pogenesis. J. Biol. Chem. 272:5367-5370.
Phillips, B.W., Vernochet, C., and Dani, C. 2003.Differentiation of embryonic stem cells forpharmacological studies on adipose cells.Pharmacol. Res. 47:263-268.
Pittenger, M.F., Mackay, A.M., Beck, S.C., Jaiswal,R.K., Douglas, R., Mosca, J.D., Moorman,M.A., Simonetti, D.W., Craig, S., and Marshak,D.R. 1999. Multilineage potential of adult

Differentiation ofStem Cells into
Adipocytes
23.4.14
Supplement 34 Current Protocols in Cell Biology
human mesenchymal stem cells. Science284:143-147.
Rodriguez, A.-M., Elabd, C., Delteil, F., Astier,J., Vernochet, C., Saint-Marc, P., Guesnet, J.,Guezennec, A., Amri, E.-Z., Dani, C., andAilhaud, G. 2004. Adipocyte differentiation ofmultipotent cells established from human adi-pose tissue. Biochem. Biophys. Res. Commun.315:255-263.
Rodriguez, A.-M., Pisani, D., Dechesne, C.A.,Turc-Carel, C., Kurzenne, J.-Y., Wdziekonski,B., Villageois, A., Bagnis, C., Breittmayer, J.-P.,Groux, H., Ailhoud, G., and Dani, C. 2005a.Transplantation of a multipotent cell popula-tion from human adipose tissue induces dys-trophin expression in the immunocompetentmdx mouse. J. Exp. Med. 201:1397-1405.
Rodriguez, A.-M., Elabd, C., Amri, E.Z., Ailhaud,G., and Dani, C. 2005b. The human adiposetissue is a source of multipotent stem cells.Biochimie 87:125-128.
Smith, A.G. 1992. Mouse embryo stem cells: Theiridentification, propagation and manipulation.Semin. Cell Biol. 3:385-399.
Tontonoz, P., Hu, E., Devine, J., Beale, E.G.,and Spiegelman, B.M. 1995. PPAR gamma 2regulates adipose expression of the phospho-enolpyruvate carboxykinase gene. Mol. CellBiol. 15:351-357.
Voytas, D. 2000. Agarose gel electrophoresis. Curr.Protoc. Mol. Biol. 51:2.5A.1-2.5A.9.
Wabitsch, M., Bruderlein, S., Melzner, I., Braun,M., Mechtersheimer, G., and Moller, P. 2000.LiSa-2, a novel human liposarcoma cell line witha high capacity for terminal adipose differentia-tion. Int. J. Cancer 88:889-894.
Wabitsch, M., Brenner, R.E., Melzner, I., Braun,M., Moller, P., Heinze, E., Debatin, K.M., andHauner, H. 2001. Characterization of a hu-man preadipocyte cell strain with high capacityfor adipose differentiation. Int. J. Obes. Relat.Metab. Disord. 25:8-15.
Zaragosi, L.E., Ailhaud, G., and Dani, C. 2006.Autocrine fibroblast growth factor 2 signalingis critical for self-renewal of human multipotentadipose-derived stem cells. Stem Cells 24:2412-2419.
Zilberfarb, V., Pietri-Rouxel, F., Jockers, R., Krief,S., Delouis, C., Issad, T., and Strosberg, A.D.1997. Human immortalized brown adipocytesexpress functional beta3-adrenoceptor coupledto lipolysis. J. Cell Sci. 110:801-807.
Contributed by Brigitte Wdziekonski, PhiVillageois, and Christian Dani
CNRS Universite de Nice SophiaAntipolis
Nice, France