curso de botanica
DESCRIPTION
libro sobre plantasTRANSCRIPT

Martes, 22 de junio de 2010
BOTÁNICA ON - LINE
La botánica, es una rama de la biología que trata del estudio de las plantas desde el nivel celular, estableciendo las relaciones entre estructura y función, pasando por el individuo, hasta su distribución geográfica, en los distintos ecosistemas terrestres. La experiencia de muchos años como docente de fisiología vegetal, me ha llevado a enfatizar más los aspectos bioquímicos que son básicos para la comprensión del funcionamiento de las plantas. En la elaboración del temario no se ha respetado una secuencia preestablecida, sino que se han incluido temas considerados importantes por el autor.
Esperamos que éste libro sea de utilidad tanto a estudiantes como a docentes y que despierte el interés por el aprendizaje de la botánica, en sus diferentes especialidades: citología, morfología, fisiología, sistemática, ecología, biogeografía; así como en la botánica aplicada.
Translocación por el Xilema
El proceso de translocación por el xilema puede ocurrir tanto por lapresión radical, la que no tiene la magnirud suficiente para explicar eltransporte de agua y minerales a la copa de los árboles de mayor altitud,que alcanzan más de 100 metros, como por la cohesión de las moléculas deagua, ya que la transpiración genera tensiones lo suficientemente grandescomo para mover una columna líquida hasta la copa de los árboles másgrandes, lo que está de acuerdo con la teoría de Dixon de la cohesión.
Translocación y retranslocación de solutos
En las hojas y tejidos fotosintéticos, se realiza la síntesis de carbohidratos y otros metabolitos que son transportados por el floema desde los sitios de producción hacia los sumideros, donde se utilizan o almacenan en forma de almidón. La hipótesis del flujo de masas o de presiones de Münch, explica con claridad y sencillez como ocurre la translocación.
Crecimiento Vegetal
El sedentarismo de las plantas terrestres, ha sido determinante en sus hábitos de vida, lo que les permite captar la energía lumínica del sol mediante la fotosíntesis y por otro lado obtener los nutrientes del medio a través de las raíces, estimulando su crecimiento y desarrollo. El crecimiento está determinado por la actividad de las células meristemáticas, en los puntos apicales de tallos y raíces. Las plantas desarrollan mecanismos de adaptación dependiendo de las condiciones ambientales, impartiéndole plasticidad a su desarrollo. Así mismo las células vegetales muestran totipotencia. Las hormonas vegetales, auxina y citocinina participan en el ciclo celular, la auxina estimula la replicación del ADN, mientras que la citocinina inicia los eventos de la mitosis
Germinación.

Se muestran los resultados de tres estudios publicados, sobre la germinación de semillas de tres especies de árboles neotropicales
.1.- Cambios anatómicos y químicos que ocurren durante la hidratación de semillas de Enterolobium cyclocarpum Griseb.
2.- Un estudio sobre el efecto de la luz en la germinación isotérmica de Alnus acuminata H.B.K., la cual está controlada por un Sistema de fitocromo.
3.- Un trabajo en que se estudia el efecto de la temperatura y el tamaño de la semilla en la germinación de Heliocarpus popayanensis H.B.K.
Proteínas y Aminoácidos
Las células vegetales están formadas por 9,4 % de proteínas. Estas son macromoléculas compuestas por amino ácidos, unidos mediante enlaces peptídicos, cuya secuencia determina su estructura primaria. Presentan propiedades diversas determinadas por la forma de las macromoléculas. Las proteínas globulares tienen propiedades catalíticas y de defensa, otras forman sistemas largos y fibrosos que suministran fuerza y rigidez a las células y a los organismos. Así mismo, un conjunto largo y delgado pueden contraerse y producir movimiento.
Acidez y pH
En casi todas las células vegetales, el jugo vacuolar es más ácido que el protoplasma debido a la acumulación de ácidos orgánicos, los que se encuentran separados del citoplasma mediante la membrana vacuolar o tonoplasto, impidiendo la inactivación de las enzimas, que son sensibles a los cambios bruscos de pH.
Los ácidos orgánicos en los tejidos vegetales se encuentran en una concentración del 0,1% en base al peso seco y en algunos casos como en los limones es 100 veces mayor. La savia de muchas plantas son ligeramente ácidas con un pH de 6,0; aun que la de algunos frutos puede estar entre pH 2 y 3. Los cambios moderados de pH afectan el estado iónico de las enzimas especialmente alrededor del centro activo y con frecuencia también el del substrato. Cuando se mide la actividad enzimática a diversos pH, la actividad óptima generalmente se observa entre los valores de 5,0 a 9,0; sin embargo algunas enzimas como la pepsina, se activa a valores de pH entre 1,5 a 2,5 , la amilasa de la malta tiene un óptimo de 5,2 y la lipasa de la semilla de tártago(Ricinus communis) de 5,0.
Vegetación de Venezuela
La energía del sol dirige los patrones de circulación global del aire y de las aguas de los océanos. El calentamiento y enfriamiento de las masas de aire y de agua en movimiento explica la mayoría de los patrones climáticos de la Tierra. Los climas, ejercen una gran influencia sobre la abundancia y distribución de las especies. Se considera la distribución de la vegetación en Zonas Climáticas en una escala global, dependiendo del clima zonal y el tipo de suelo.
Así mismo, los tipos de vegetación en Venezuela muestran una gran biodiversidad determinados por diversos factores como son la topografía, exposición al sol, temperatura, precipitación anual, suelos, la velocidad y dirección de los vientos, el drenaje del suelo y la historia geológica del área. Se presenta una clasificación de la vegetación de Venezuela según Julián Steyermark. Introducción a la Ecofisiología Vegetal

La fisiología vegetal es una disciplina que se refugia en el laboratorio para su estudio, manipula las condiciones de crecimiento del individuo y mide la respuesta de un determinado proceso. La ecofisiología, estudia los fenómenos fisiológicos fuera del laboratorio, en su medio ambiente natural, el cual está sujeto a cambios y alteraciones, como resultado de fenómenos naturales o producto de la actividad humana.
Célula Vegetal Todos los organismos vivos están compuestos por células. El inglés, Robert Hooke en 1665, realizó cortes finos de una muestra de corcho y observó usando un microscopio rudimentario unos pequeños compartimentos, que no eran más que las paredes celulares de esas células muertas y las llamó células ( del latín cellula, que significa habitación pequeña ) ; ya que éste tejido le recordaba las celdas pequeñas que habitaban los monjes de aquella época. No fue sino hasta el siglo XIX, que dos científicos alemanes el botánico Matthias Jakob Schleiden y el zoologo Theodor Schwann, enunciaron en 1839 la primera teoría celular : " Todas las plantas y animales están compuestos por grupos de células y éstas son la unidad básica de todos los organismos vivos".
Enzimas
Las enzimas son biocatalizadores de naturaleza proteica. Todas las reacciones químicas del metabolismo celular se realizan gracias a la acción de catalizadores o enzimas. La sustancia sobre la que actúa una enzima se denomina substrato. Pasteur descubrióque la fermentación del azúcar mediante levaduras, con su conversión en alcohol etílico y anhídrido carbónico es catalizada por fermentos o enzimas.
Bioenergética
El concepto de energía libre, llamada también trabajo neto o útil isotérmico fue propuesto independientemente por Gibbs y Helmholtz.. Si conocemos los cambios en energía libre a temperatura y presión constante, podemos predecir si una reacción es espontánea o no. La energía libre es la energía útil, mientras que la entropía es la energía degradada...
ATPEl ATP puede actuar como transportador de energía química, en cientos de reacciones celulares, por lo que se le considera como un compuesto rico en energía; ya que muestra una gran disminución de energía química cuando participa en reacciones hidrolíticas.
RespiraciónLa Respiración aeróbica es un proceso transductor de energia en la cual la molecula de glucosa se oxida en presencia de oxígeno liberando anhídrido carbónico, agua y energia en forma de ATP; mientras que la glucólisis o respiracion anaeróbica es la degración de la glucosa en ausencia de oxígeno molecular formandose alcohol etílico y anhídrido carbónico, o en algunos casos acido láctico y ATP.
Fotosíntesis
La vida en la tierra depende fundamentalmente de la energía solar. La fotosíntesis es el único proceso de importancia biológica que puede atrapar esa energía. Toda la materia orgánica disponible en la tierra ha

sido producida por la fotosíntesis. La materia orgánica comprende los alimentos que consumimos diariamente tanto nosotros como los animales, los combustibles fósiles (petróleo, gas, gasolina, carbón); así como la leña, madera, pulpa para papel, inclusive la materia prima para la fabricación de fibras sintéticas, plásticos, poliester, etc.
Glosario Compost
Es la materia orgánica completamente descompuesta, de color oscuro, inodora, pero abundante en nutrientes. Un poeta como W. Whitman hace la siguiente alabanza al compost: "el compost o humus es el fertilizante más eficiente y práctico que el hombre conoce, le restablece al suelo gastado los nutrientes valiosos y transforma un pedazo de tierra improductiva y árida en un exuberante jardín".
Nutrición Mineral de las Plantas
El estudio de la nutrición mineral de las plantas amerita conocer su composición química, cuyo objetivo se puede alcanzar utilizando los dos métodos siguientes: 1. El análisis elemental, que determina la naturaleza y las proporciones en que se encuentran los elementos minerales en los tejidos vegetales. 2. El análisis inmediato, que trata de reconocer la naturaleza de los compuestos orgánicos que existen en las diversas partes de la planta. Así mismo, es recomendable saber las proporciones de humedad y de materia seca en los órganos sometidos al análisis. Se ha comprobado que 10 elementos son requeridos para el crecimiento normal de las plantas en grandes concentraciones, son los macroelementos constituidos por: carbono, oxígeno, hidrógeno, nitrógeno, calcio, portasio, fósforo, azufre, magnesio y silicio; así mismo las plantas utilizan en pequeñas cantidades que oscilan entre 0,01 a 0,5 ppm, 9 elementos, los micronutrientes u oligoelementos que son: boro, cloro, cobre, hierro, manganeso, molibdeno, níquel, cinc y sodio.
Relaciones Hídricas en las Plantas
No se puede concebir la vida sin la presencia de agua. Es el líquido más común y extraordinario conocido. Tres cuartas partes de la superficie terrestre están cubiertas por agua. En la tierra existen reservas ocultas de agua en el subsuelo, en los casquetes polares se encuentra en forma de hielo y en la atmósfera está presente en forma de vapor de agua . A pesar de que el agua es la molécula más abundante en la superficie terrestre, su disponibilidad es el factor que limita más la productividad vegetal en la tierra, en una escala global. La poca disponibilidad de agua limita la productividad de los ecosistemas terrestres, principalmente en climas secos.
Transpiración
Es la pérdida de agua en forma de vapor a través de los estomas, cutícula, y peridermis de las plantas. Casi toda el agua que se pierde por la hoja lo hace a través de los poros del aparato estomático, que son más abundantes en el envés de la hoja. Las hojas pierden agua irremediablemente a través de los poros estomáticos, como consecuencia de la actividad fotosintética de las células del mesófilo. Se podría decir que la transpiración es un mal necesario, ya que sí los estomas no se abren no penetra el CO2 requerido para la fotosíntesis por las células del parénquima clorofílico.

CONTENIDOS
/ ABSORCIÓN DE AGUA / ABSORCIÓN DE SALES MINERALES / TRANSLOCACIÓN APOPLÁSTICA Y SIMPLÁSTICA / RELACIONES ENTRE ESTRUCTURA Y FUNCIÓN DEL XILEMA / TEORÍA DE DIXON DE LA COHESIÓN TENSIÓN TRANSPIRACIÓN / GRADIENTES DE POTENCIALES HÍDRICOS A TRAVÉS DEL
CONTINUUM RAÍZ-PLANTA-ATMÓSFERA /
PÁGINA PRINCIPAL
ABSORCIÓN DE AGUA
La absorción de agua es un proceso pasivo que se produce por simple difusión, sin el aporte de energía metabólica. El agua siempre se mueve siguiendo un gradiente de potenciales químicos, de zonas de alto potencial hídrico hacia zonas de bajo potencial hídrico. El agua se difunde de la solución del suelo a la raíz a través de la epidermis, luego pasa el cortex, endodermis, periciclo y finalmente penetra en los vasos, siguiendo un gradiente de potenciales hídricos. El gradiente neto es el producido entre los vasos y el medio externo.
En el interior de los vasos se desarrolla una presión hidrostática, inducida osmóticamente, a la que se le da el nombre de presión radical. Las células vivas entre los vasos del xilema de la raíz y la solución del suelo actúan como una membrana semipermeable. Podemos pensar que el xilema de la raíz actúa como un osmómetro que se encuentra sumergido en la solución del suelo; de tal forma que la máxima presión osmótica que se desarrolla depende de la concentración de la savia del vaso y de aquella en la solución del suelo.
Donde Pr es la máxima presión radical; es el potencial osmótico de la solución del suelo
y es el potencial osmótico de la savia vascular.
En muchos casos la savia vascular es muy diluida y sí la solución del suelo es agua pura, se han registrado presiones radicales que varían entre 1 y 3 atm (0,1 a 0,3 MPa).
Las presiones radicales registradas de 2 a 3 atm, permiten que una columna de agua se eleve una altura de hasta 30 metros, lo que no explicaría el ascenso de la savia en árboles gigantes de más de 100 metros.
La absorción activa de solutos debe acompañar la absorción de agua para que se mantenga la presión radical. Experimentalmente la absorción de agua está acompañada de la absorción de oxígeno, lo cual no es sorprendente, ya que la absorción de sales es un proceso activo que garantiza la presión radical, y que ocurre paralelamente con la absorción de agua.
Probablemente la presión radical no juega un papel importante en el ascenso de savia en la planta; ya que la tasa de exudación es menor que la tasa de transpiración; así mismo la savia xilemática se halla bajo tensión en lugar de presión.
¿Cómo se mide la presión radical? Si se corta el extremo de un tallo, se pude observar la exudación de savia. Si a una planta de tomate bien regada se le corta la parte superior del tallo y este se conecta mediante un manguito de goma a un tubo de vidrio que contiene agua, se puede observar que el nivel del agua en el tubo de vidrio sube, poniendo en evidencia la presión radical.

Cuando las plantas se encuentran en un medio saturado de agua o que la humedad relativa es muy alta, el agua se exuda como gutación en forma de gotas a través de los márgenes foliares por estructuras llamadas hidatodos. La gutación se puede parar regando la planta con una solución salina o de azúcar, lo que disminuye la presión radical al disminuir el potencial osmótico del medio donde se encuentra la raíz.
ABSORCIÓN DE SALES MINERALES.
La permeabilidad controla la entrada de sales a la célula solamente sí es causada por difusión. Las membranas celulares son diferencialmente permeables, por lo que permiten el paso selectivo de ciertas sustancias. Por ejemplo, los herbicidas si son solubles en lípidos al asperjarlos sobre las plantas entran a las células y producen el efecto deseado.
La absorción pasiva es causada por simple difusión, siguiendo un gradiente electroquímico, hasta obtener la condición de equilibrio. No requiere de energía metabólica.

La difusión de un soluto a través de una membrana, depende de los siguientes factores:
1) De la agitación térmica de las moléculas a difundir, 2) del gradiente de concentración a través de la membrana, 3) de la permeabilidad de la membrana, que está determinada por la solubilidad en el núcleo hidrofóbico de la doble capa lípidica de la membrana.
La absorción activa se realiza en contra de un gradiente electroquímico, de sitios de menor concentración hacia sitios de mayor concentración y requiere energía metabólica. La fuente principal de la energía metabólica es la hidrolisis del ATP aportado por la respiración. Este transporte se realiza en presencia de proteínas transportadoras o bombas que pueden transportar H+ , Na+, Ca+2 , K+ .
En el transporte activo participan tres tipos de proteínas: Transportadores mono transportadores, mueven un solo soluto en una dirección. Por ejemplo, una proteína que transporta el ion Ca+2 , que se encuentra en la membrana plasmática, mueve este ion hacia zonas de mayor concentración. Las proteínas cotransportadoras mueven dos iones en la misma dirección. Las proteínas de contratransporte mueven

dos solutos en direcciones opuestas, muchas células tienen una bomba Na+ - K+, que mueven el potasio hacia el interior de la célula y el sodio hacia el exterior. Las características de la acumulación metabólica son muy complejas, sin embargo se pueden establecer las siguientes conclusiones: Las moléculas pequeñas y sin carga, pueden entrar las células rápidamente. Aquí podemos mencionar el H2O, CO2 y nitrógeno. Los otros nutrientes penetran las células ya sea como aniones (-) o cationes (+).
Iones que se acumulan rápidamente.
Iones que se acumulan Lentamente.
Aniones NO3- , Cl- PO4-3 , SO4-2
Cationes K+ , Na+ , NH4+ Ca+2, Mg+2
La velocidad de entrada de Fe, Zn y Mn es lenta; sin embargo podemos decir que la entrada de muchos micronutrientes no ha sido bien estudiada.
Durante la acumulación de nutrientes, se debe mantener la neutralidad eléctrica a ambos lados de la membrana, esto resulta en la interacción iónica que se observa comúnmente. Por ejemplo, sí el catión rápido K+ está presente junto al anión rápido NO3- , la velocidad de entrada de ambos iones es rápida, en consecuencia se expulsa un protón y un ion hidroxilo para mantener la electro neutralidad y el pH del medio no se altera. Sin embargo sí el catión K+ , está presente en una solución nutritiva con el anión SO4-2 , que es lento, se expulsan más protones ( H+ ) que hidroxilos ( OH-) y el pH de la solución baja, acidificándose el medio.
Cuando la absorción del anión excede la del catión, como en el caso del CaCl2 , que el anión Cl- , se absorbe más rápido que el calcio (Ca+2 ), se expulsan de la raíz iones hidroxilos (OH-), haciendo el medio básico. Cuando se cultivan plantas en soluciones hidropónicas, los cambios del pH son problemáticos; sin embargo los especialistas en nutrición mineral han tratado de superar este escollo diseñando soluciones nutritivas balanceadas, pero esto no ha sido fácil y todavía se presentan cambios de pH indeseables, lo que ocasiona que se deban cambiar las soluciones nutritivas periódicamente, aumentando los costos de este cultivo.
TRANSLOCACIÓN APOPLÁSTICA Y SIMPLÁSTICA.
Los nutrientes minerales que penetran la raíz a través de los pelos radicales pueden moverse a través del citoplasma celular y atravesar las células mediante los plasmodesmos, hasta llegar al tejido vascular. Las células de la rizodermis están conectadas mediante plasmodesmos o puentes citoplasmáticos, lo que permite el movimiento de solutos de célula a célula. El transporte de los solutos mediante plasmodesmos se denomina translocación simplástica. Sin embargo los solutos se pueden mover a través de las paredes celulares de las células de la raíz, denominándose a esta, translocación apoplástica. El agua y las sales minerales se pueden mover por el apoplásto, hasta llegar a la endodermis, donde el movimiento se realiza a través de la membrana plasmática y continúa a través del simplasto.

Corte transversal de la raíz a nivel de un pelo radical, mostrando el transporte apoplástico y simplástico.
RELACIONES ENTRE ESTRUCTURA Y FUNCIÓN DEL XILEMA
El xilema es el tejido encargado de la conducción de la savia, que es desplazada por la corriente transpiratoria. Las células del xilema son tubulares, carecen de protoplasto, poseen extremos abiertos, tienen paredes celulares lignificadas, atravesadas por punteaduras, lo que impide el aplastamiento de los vasos que están sometidos a tensión (hipopresión) cuando la planta transpira activamente,. Cuando son funcionales, están muertos, son permeables a solutos y solventes, tienen la concentración de la savia baja, no generan presión de turgencia. Cuando funcionan colapsan parcialmente, exceptuando cuando opera la presión radical. Cuando se corta un tallo, absorbe agua o aire. Transportan agua y solutos disueltos. La savia puede adquirir una velocidad promedio de 75 cm por minuto.
En el xilema se encuentran dos tipos de elementos conductores: traqueidas y tráqueas (o vasos). Las traqueidas son células alargadas con forma de huso, de lumen estrecho y abundantes punteaduras. La resistencia a la conducción es mayor en las traqueidas que en las tráqueas, que son más cortas y de lumen ancho y con paredes terminales ausentes o muy perforadas. El diámetro de los vasos leñosos varía de 40 µm (aciculifolias) a 700 µm (lianas), con velocidades máximas de la corriente transpiratoria de 2 cm por minuto en aciculifolias a 250 cm por minuto en lianas.
Las paredes celulares de los vasos pueden adquirir engrosamientos que pueden tener forma anular, helicoidal, espiral, escalariforme o reticular, lo que les imparte resistencia al vaso e impide su colapso bajo hipopresión.

Vasos o tráqueas
Traqueidas
TEORÍA DE DIXON DE LA COHESIÓN TENSIÓN TRANSPIRACIÓN.
La presencia de enlaces de hidrógeno en el agua da como resultado la cohesión, que es la fuerza de atracción mutua de las moléculas, esto es lo que permite que una columna de agua se mueva desde las raíces de un árbol hasta las hojas a más de 100 metros de altura. Una propiedad relacionada a esta es la adhesión, que es la atracción del agua por una fase sólida, como las paredes celulares. El agua presenta una alta tensión superficial, que indica que las moléculas de agua se atraen mucho más fuertemente entre sí, que a la fase gaseosa que está en contacto con el agua. Tanto las fuerzas de cohesión, tensión superficial y adhesión producen como consecuencia la capilaridad que es un ascenso del agua dentro de un tubo capilar (un tubo xilemática), hasta llegar a un punto donde el peso de la columna líquida balancea la fuerza capilar. El ascenso de la savia por capilaridad en un tubo xilemático típico no sobrepasa un metro, lo que es insuficiente para explicar el ascenso de la savia a la corona de los arboles más altos.
En 1914, un botánico Irlandés llamado Henry H. Dixon, formuló la hipótesis que la tensión generada por la transpiración, absorción de agua por ósmosis y la hidratación de la pared celular, producía un movimiento de agua desde la parte de abajo hacia arriba, de la columna líquida que se mantenía unida por fuerzas de

cohesión, esto se conoce como la teoría de Dixon de la cohesión. Actualmente el fenómeno que explica el ascenso de la savia por el xilema recibe el nombre de teoría coheso-tenso-transpiratoria.
Esta teoría, se puede resumir diciendo que las hojas, al perder vapor de agua por transpiración, tienden a disminuir el potencial hídrico de las células del mesófilo, lo que determina la entrada de agua a los elementos vasculares de la hoja, donde es sometida a tensión y se mueve mediante flujo de masas. El agua dentro de los conductos xilemáticos, se extiende formando una columna continua desde la raíz hasta las hojas, por lo que la tensión (hipopresión) en las partes altas del xilema, se transmite hacia abajo a través de dicha columna. Si las fuerzas de cohesión entre las moléculas de agua son lo suficientemente grandes, toda perdida de agua que se produce en la hoja por evapo-transpiración genera una tensión que se traduce en el ascenso de la columna líquida hacia la hoja, mientras que en la porción inferior de la columna ocurre una succión del agua procedente de la raíz. Como resultado de este proceso disminuye el potencial hídrico de las células de la raíz, lo que se traduce en una nueva absorción de agua por estas células.
Resumen de la teoría de la cohesión-tensión-transpiración
Los árboles más grandes sobrepasan los 100 metros, así que para elevar una columna de agua desde el nivel del suelo hasta la corona de esos árboles, se requiere una presión en la base de 10,9 atm (1,11 MPa), más una presión adicional que contrarreste la resistencia friccional al flujo del agua. Sí esta presión es similar a la precisada para el ascenso del agua, se necesita una presión de 2,2 MPa (21,8 atm). Las fuerzas de cohesión entre las moléculas de agua ha sido determinada mediante varios procedimientos y está en el

rango de 10 a 30 MPa, lo que es suficiente para mantener una columna liquida de 100 metros y más, sin que se rompa, aguantando las grandes tensiones que se desarrollan en el tallo, halando la columna liquida que se mueve a través del xilema.
GRADIENTES DE POTENCIALES HÍDRICOS A TRAVÉS DEL CONTINUUM RAÍZ-PLANTA-ATMÓSFERA.
En el movimiento del agua desde el suelo a través de la planta hasta la atmósfera participan diferentes mecanismos de transporte.
1. El agua se mueve en el suelo y en el xilema mediante flujo de masas en respuesta al gradiente de presión.
2. En la fase gaseosa el agua se mueve primeramente por difusión, hasta que alcanza el aire externo.
3. Cuando el agua se transporta a través de la membrana, la fuerza impulsora es la diferencia de potencial hídrico a través de la membrana. El flujo osmótico ocurre cuando las células absorben agua y cuando las raíces transportan agua desde el suelo hasta el xilema.
Potencial hídrico MPaSuelo húmedo 0Raíz -0,6Hoja -1,0Aire seco -100,0
Diferencia de potenciales hídricos en el transporte de agua del continuum suelo-planta-atmósfera.
En la tabla de arriba podemos observar que existe un gradiente de potenciales hídricos decrecientes, desde la solución del suelo, pasando por la raíz, hoja y el aire atmosférico seco, que permite el movimiento de agua por flujo de masas desde el suelo hasta la hoja y por ultimo hacia el aire circundante.
CONTENIDOS
/ INTRODUCCIÓN / RELACIÓN ENTRE ESTRUCTURA Y FUNCIÓN EN EL FLOEMA /HIPOTEIS DEL TRANSPORTE EN EL FLOEMA / CARGA Y DESCARGA DEL FLOEMA /
PÁGINA PRINCIPAL
INTRODUCCION
La planta para su crecimiento normal requiere agua, nutrientes minerales y realizar la transformación de la energía luminosa en energía química mediante el proceso fotosintético. En las hojas y tejidos verdes de la planta se realiza la síntesis de carbohidratos y otros metabolitos que son transportados desde los lugares de producción, hacia los sumideros o sitios de utilización y almacenamiento como son tallos,

raíces, frutos, semillas etc. Los fotoasimilados, se transportan también hacia los tejidos jóvenes, tallos y hojas en proceso de crecimiento, a través del tejido floemático.
Las primeras evidencias del transporte de azucares por el floema fueron presentadas por el italiano Marcello Malpighi en 1675, mediante la técnica del descortezamiento anular del tallo. En un tallo tratado de esta forma ocurre que la parte superior se hincha, por la acumulación de carbohidratos, solamente cuando hay presente hojas fotosintéticamente activas. Este procedimiento puede ser utilizado para eliminar árboles, ya que al interrumpir el flujo de alimentos hacia las raíces, estas mueren. Así mismo, esta técnica es de gran utilidad para la propagación de plantas mediante el enraizamiento de acodos aéreos.
Fuente: http://www.euita.upv.es/VARIOS/BIOLOGIA/images/Figuras_tema13/Figura13_8.jpg

Cuando los compuestos radiactivos estuvieron disponibles para la investigación, se utilizó anhídrido carbónico marcado (14CO2 ) para demostrar que los azucares formados en el proceso fotosintético se translocan a través de los tubos cribosos.
Ir al principio
RELACIÓN ENTRE ESTRUCTURA Y FUNCIÓN EN EL FLOEMA
En plantas con crecimiento secundario, el floema constituye la corteza interna. Las células del floema que transportan azucares y otros compuestos orgánicos a través de la planta son los elementos cribosos. En las angiospermas están diferenciados en tubos cribosos y en las gimnospermas se encuentran las células cribosas menos especializadas. El floema además de elementos cribosos contiene células compañeras y células parenquímáticas (que almacenan reservas de alimentos), en algunos casos el floema puede contener fibras y esclereidas; y en otros lacticíferos. Sin embargo, solamente los elementos cribosos participan en la translocación.
Los elementos cribosos maduros, carecen de núcleo y tonoplasto (membrana vacuolar); así mismo se encuentran ausentes microfilamentos, microtúbulos, cuerpos de Golgi y ribosomas, además presentan una proteína P. Durante el desarrollo de los elementos cribosos se retiene la membrana plasmática, mitocondrias, plastidios y el retículo endoplasmático liso. Los elementos cribosos poseen áreas cribosas o perforadas en sus paredes celulares, donde los poros conectan las células conductoras. Los poros pueden tener un diámetro que varía de 1µm a 15 µm, a diferencia de las células cribosas de las gimnospermas, en las angiospermas se diferencia una placa cribosa, y los elementos de los tubos cribosos se unen longitudinalmente formando un tubo criboso. Las placas cribosas son canales abiertos que permiten el transporte entre células. Íntimamente asociadas a los tubos cribosos o a las células cribosas se encuentran las células compañeras (en angiospermas) y las células albuminoideas (en gimnospermas), que poseen citoplasma denso y núcleo bien diferenciado. Entre las células acompañantes y los tubos cribosos existen abundantes plasmodesmos que los conectan.

Fuente: http://www.euita.upv.es/VARIOS/BIOLOGIA/images/Figuras_tema13/Figura13_4.jpg
Más del 90% del material translocado por el floema son carbohidratos, especialmente azucares no reductores, entre estos la sacarosa (el azúcar de cocina) es la más abundante. En el esclarecimiento de los azucares transportados por el floema ha jugado papel importante, la utilización de insectos chupadores denominados áfidos, que introducen el estilete de la boca, en un tubo criboso, para succionar la savia que le sirve de alimento, cuando cortan el estilete con una hojilla, después de anestesiar el áfido; el estilete continua exudando material a razón de 1 mm3 por hora durante cuatro días. Este procedimiento de obtener savia floemática, ha permitido determinar las sustancias que se translocan en el floema; así como la velocidad de translocación, utilizando marcadores radiactivos. Se registran velocidades de la savia de 5cm por minuto.
En la tabla siguiente se muestra un análisis de la savia floemático y xilemática, la savia xilemática tiene una concentración baja especialmente de sales inorgánicas; mientras que el floema tiene una concentración alta de sólidos totales, principalmente de compuestos orgánicos, en la que la sacarosa es el azúcar más importante, con una concentración aproximada del 20%.
Análisis químico de la savia de un árbol de pera y del exudado floemático de Robinia pseudo-acacia
Substancia Concentración en la savia mg/litro Xilema Floema
Ca 85 720Mg 24 380

SO42- 32 --PO43- 25 --Azucares* -- 200.000N, orgánico -- 425N, inorgánico -- 135(*) Principalmente sacarosa.
Fuente: Bidwell, R.G.S.1979. Plant Physiology. Macmillan Publishing Co., Inc. New York.726 p.
Fuente: http://www.euita.upv.es/.../Figura13_10.jpg
Las células del mesófilo de los arboles tienen un potencial osmótico de -1,3 a -1,8 MPa, mientras que los elementos cribosos en las hojas tienen un potencial osmótico de -2,0 a -3,0 MPa, que es equivalente a una solución de sacarosa de 20 a 30 % , esto nos indica que la concentración de azúcar es aproximadamente 1,5 a 1,7 veces mayor en los elementos cribosos que en las células del mesófilo. El proceso mediante el cual la concentración de azúcar aumenta cerca de las células fotosintetizadoras del mesófilo se denomina carga del floema. El movimiento de azúcares de una célula del mesófilo a otra se realiza a través del simplasto, ya que existen numerosos plasmodesmos conectando las células. En el movimiento de la savia en el floema no se realiza en una sola dirección, sino que se realiza de zonas donde hay un buen suministro (llamada fuente) hacia áreas de utilización o almacenamiento, llamadas sumidero, o áreas de descarga.
HIPOTEIS DEL TRANSPORTE EN EL FLOEMA
Modelo del Flujo de masas o de presiones.
En 1931, Ernest Munch propuso que el movimiento de la savia en la planta, era análoga a la circulación, en la que las células del mesófilo en las hojas actuaban como una bomba, en las que se fabricaban carbohidratos mediante la fotosíntesis, manteniendo alta la concentración de azucares en las células del mesófilo, a pesar de que algunos azucares se transportaran a través del floema. El agua y las sales minerales se movían hacia arriba (sentido acrópeto) por el xilema; mientras que las sustancias orgánicas se movían hacia abajo por el floema. El potencial osmótico en el floema se mantiene bajo producto de la

acumulación de carbohidratos por la fotosíntesis, de tal forma que el agua se mueve por ósmosis del apoplásto (tejido xilemático). De tal forma que se genera una presión hidrostática en el floema que forza la savia hacia las raíces a través de los conductos floemáticos. En las raíces, se descargan los solutos del floema, mediante un transporte activo, lo que resulta en un aumento del potencial osmótico de la savia del floema. En respuesta a este proceso, se difunde agua fuera del floema y se mueve de nuevo hacia las hojas a través del gradiente de potencial osmótico del xilema. El efecto neto es la circulación de agua. La energía que mantiene esta circulación es suministrada por la adición activa de solutos orgánicos a las hojas por la fotosíntesis y su descarga activa en las raíces. En las regiones vertederos (de descarga), los solutos transportados son extraídos activamente de los elementos del tubo criboso, hacia los tejidos vecinos. Esta descarga ayuda a mantener el gradiente de potencial de presión en los tubos cribosos y promueve la formación de azúcares y almidón a concentraciones elevadas en las regiones de almacenamiento, como son los tubérculos, raíces, frutos y semillas en crecimiento.
El aspecto principal que hay que tomar en cuenta en relación a esta teoría, es que requiere una presión hidrostática positiva en el floema, y en consecuencia un suministro permanente de azucares en las hojas, que se utilice para generar la presión. La savia del floema tiene una presión positiva, que se puede demostrar al cortar el estilete de un áfido que se alimente de la savia de una planta, ya que la savia del floema continua exudando a través del estilete durante cierto tiempo, como resultado de la presión interna. Los análisis de la savia floemática confirman la teoría, ya que la concentración de azúcar es alta, pudiendo ser de 20 % de sacarosa. Otro argumento a favor de la teoría del flujo de masas es que, las substancias de crecimiento y las partículas de virales aplicadas a las hojas, se translocan rápidamente sí la hoja se ilumina, pero no así en condiciones de oscuridad.
El segundo argumento importante a favor de esta teoría es que la difusión requiere una baja resistencia, a lo largo de la vía de translocación, ya que la placa cribosa debe estar abierta sin taponaduras.
El movimiento de la savia en el floema no es unidireccional, ya que ciertas sustancias se pueden mover en diferentes direcciones, y con diferentes velocidades, lo que implicaría la participación de diferentes tubos cribosos. El mecanismo de movimiento de la savia en los tubos cribosos, además del flujo de presiones, requeriría de la participación de corrientes citoplasmáticas y difusión, electroósmosis, etc; lo que hace este fenómeno de una mayor complejidad.

Fuente: Figura modificada de Curtis, H. y Barnes, N.S. (1997). "Invitación a la Biología". Ed. Panamericana].
CARGA Y DESCARGA DEL FLOEMA
Los fotoasimilados se mueven desde los cloroplastos del mesófilo de las hojas maduras hacia los elementos de los tubos cribosos, según los siguientes pasos:
1. Las triosas fosfatos que se forman en la fotosíntesis durante el día, se transportan hacia el citosol, donde mediante reacciones enzimáticas se transforman en sacarosa. Durante la noche, el almidón almacenado en el cloroplasto se convierte en maltosa y se mueve hacia el citosol, convirtiéndose en sacarosa.
2. La sacarosa se mueve desde las células del mesófilo, hacia células cercana a los elementos cribosos.
3. En el proceso de carga del floema, los azucares se transportan dentro de los tubos cribosos y las células compañeras. En muchas de las especies estudiadas, los azucares se encuentran más concentrados en los elementos cribosos y las células compañeras que en las células del mesófilo. Una vez que los azucares están en los elementos cribosos, son exportados de la fuente hacia el sumidero, que pueden ser fruto, granos, raíces o tubérculos en crecimiento.
Los azucares se pueden mover a través del simplasto mediante plasmodesmos o pueden entrar al apoplásto antes de ser cargado al floema. La carga de la sacarosa vía apoplástica ocurre mediante el

transporte activo; una ATPasa que bombea protones, establece un gradiente de protones y la sacarosa es transportada dentro de las célula compañeras asociadas con el floema, mediante un mecanismo de cotransporte sacarosa-protón.
La descarga del floema también requiere energía metabólica. La sacarosa puede salir del floema de forma pasiva y convertirse en glucosa y fructosa mediante la enzima invertasa ácida, en ese caso la fructosa y glucosa se transportan hacia el sumidero; alternativamente la sacarosa puede dejar el floema vía plasmodesmos o vía un transportador de sacarosa. Sí los azucares se metabolizan rápidamente en el sumidero (para formar almidón), se favorece la carga del floema al crearse un gradiente de concentración favorable.
El proceso de importación de azucares hacia los sumideros se puede resumir en los pasos siguientes:
1. Descarga del floema. Este es el proceso mediante el cual los azucares importados salen de los elementos cribosos de los tejidos sumidero.
2. Transporte a corta distancia. Después de la descarga, los azucares se transportan a las células en el sumidero mediante un transporte de corta distancia.
3. Almacenamiento y metabolismo. El paso final, implica el almacenamiento de azucares o su metabolismo. El sumidero varía ampliamente desde órganos vegetativos en crecimiento (ápices de raíces y hojas jóvenes) tejidos de almacenamiento (raíces, como la yuca, el apio y tallos como la papa, ocumo, ñame), hasta órganos de reproducción y dispersión (frutos y semillas); por lo que no existe un esquema particular para la descarga del floema y el transporte a corta distancia.
CONTENIDOS
/ PROCESO DE CRECIMIENTO, DESARROLLO Y DIFERENCIACIÓN EN LAS PLANTAS / INTERRELACIONES Y PLANTA Y MEDIO AMBIENTE COMO DETERMINANTES DEL CRECIMIENTO Y
DESARROLLO /ACVTIVADORES QUÍMICOS E INHIBIDORES / FITOHORMONAS / AUXINAS / CITOCININAS / GIBERELINAS / ETILENO / ACIDO ABSCÍSICO (ABA) / BRASINOESTEROIDES (BR) /
ACIDO JASMÓNICO (AJ) / POLIAMINAS /
PÁGINA PRINCIPAL
PROCESO DE CRECIMIENTO, DESARROLLO Y DIFERENCIACIÓN EN LAS PLANTAS
Uno de los aspectos más relevantes de las plantas terrestres es su vida sedentaria, lo que ha determinado sus hábitos de vida, que le permiten captar la energía lumínica del sol mediante el proceso fotosintético y obtener los nutrientes del medio a través de las raíces, esto estimula el crecimiento y desarrollo, ya que las plantas son autótrofas, a diferencia de los animales que se mueven en procura de su alimentación, por su condición de heterótrofos. Las plantas presentan una estructura rígida, determinada por la presencia de una pared celular celulósica, en la que sus células se encuentran muy unidas a sus vecinas mediante plasmodesmos, lo que es característico de una matriz leñosa. El crecimiento de las plantas está determinado por la actividad de células meristemáticas que se encuentran en los ápices de tallos y raíces, son los meristemas apicales y en los meristemas laterales formados por: el cambium vascular y el cambium del corcho o felógeno. Por otro lado, el desarrollo temprano de los animales, se caracteriza por migración celular hacia sitios nuevos hasta formar tejidos específicos. El sedentarismo de las plantas, provoca su adaptación ambiental, que puede ser de tipo morfofisiológica. A diferencia de los animales cuyo patrón corporal se establece durante la embriogenésis, las plantas

construyen sus formas durante toda su vida, a través de programas de desarrollo vegetativo. Es por esto que los tejidos meristemáticos proveen una forma de adaptación dependiendo de las condiciones ambientales, al mantener un reservorio de células no diferenciadas, con características embrionarias, lo que le da una gran plasticidad al desarrollo de las plantas. Las células vegetales muestran totipotencia, que es la capacidad que tiene una sola célula de regenerar un organismo completo.
El crecimiento ocurre cuando se forman nuevas células y tejidos mediante división celular. La división celular ocurre en células meristemáticas y consiste en dos fases: la mitosis en la que se replican los cromosomas y se disponen en dos núcleos hijos, que son cualitativa y cuantitativamente idénticos y la citocinesis, en la que se divide la pared celular, el citoplasma y los organelos.
Las células meristemáticas se caracterizan por tener paredes celulares delgadas, tienen vacuolas pequeñas, núcleos grandes y se dividen constantemente. Las células hijas, pueden dividirse por un tiempo, pero después pierden esa capacidad, se alargan y desarrollan vacuolas grandes y por último ocurre la diferenciación o maduración de estas células, que originan tejidos con funciones específicas, órganos y estructuras especializadas que requiere la planta durante su ciclo de vida.
Las hormonas vegetales, auxina y citokinina participan en el ciclo celular, la auxina estimula la replicación del ADN, mientras que la citocinina inicia los eventos de la mitosis.
Fuente: http://www.biologia.edu.ar/botanica/tema15/images15/proColeus.jpg

El desarrollo de la planta se caracteriza por la división, el alargamiento y la diferenciación celular, todos estos cambios están regulados de una forma compleja, en la que participan cuatro factores:
La planta capta y responde a las señales ambientales. El genoma de la planta codifica enzimas que catalizan las reacciones
bioquímicas del desarrollo, que incluyen las que fabrican hormonas, receptores, participan en la síntesis de proteínas y en el metabolismo energético.
La planta utiliza receptores que detectan las señales ambientales, como los fotorreceptores que captan la luz.
Los mensajeros químicos u hormonas, median los efectos de las señales ambientales captadas por los receptores.
Las hormonas vegetales son compuestos reguladores que actúan en muy bajas concentraciones, son producidas en una parte de un órgano multicelular y transportadas a otro sitio donde ejercen su efecto específico.
Cuando la semilla sale de su estado latente, germina y se transforma en una plántula en crecimiento. Para que el embrión comience a desarrollarse, la latencia de la semilla debe ser interrumpida por la acción de factores físicos, como la exposición a la luz, la abrasión mecánica de la testa, el fuego o el lavado de los inhibidores del crecimiento por el agua. A medida que la semilla germina, primero absorbe agua, lo que desencadena una serie de reacciones bioquímicas que movilizan las reservas de grasas, polisacáridos y proteínas. Los fotorreceptores y las hormonas regulan el desarrollo de la plántula.
Durante la germinación de la cebada y otras semillas de cereales, el embrión segrega giberelinas, una hormona que promueve la liberación de enzimas que digieren proteínas y el almidón almacenado en el

endospermo. Las giberelinas son utilizadas en la industria de la cerveza para aumentar la germinación de la cebada y la degradación del endosperma, produciendo azúcar que es fermentada a alcohol etílico.
Después que la planta alcanza una edad o tamaño apropiado, se produce el fenómeno de floración y la formación de frutos. La floración en algunas plantas es controlada por la longitud de la noche. Es probable que una hormona viaje desde un órgano receptor como una hoja hacia la yema donde se formará una flor. Cuando la flor se forma, las hormonas desempeñan otros papeles, como el crecimiento del tubo polínico a través del estilo de un pistilo, para producir la fertilización. Luego se desarrolla un fruto que madura bajo control hormonal.Las auxinas, giberelinas y el etileno regulan la formación de un fruto.
Por último todas las plantas envejecen y mueren. La muerte, sigue a los cambios del envejecimiento controlados por hormonas como el etileno. Las hojas antes de comenzar el período de sequía, por interacción de hormonas como la auxina y el etileno envejecen y caen. El envejecimiento y muerte de plantas como el agave y algunas leguminosas anuales, se debe a un incremento del ácido abscisico durante la floración.
Ir al principio
INTERRELACIONES PLANTA Y MEDIO AMBIENTE COMO DETERMINANTES DEL CRECIMIENTO Y DESARROLLO
Existe una amplia variedad de formas vegetales, que se pueden clasificar en varias categorías con diferentes características ecológicas. Algunas plantas crecen sin ningún soporte mecánico y estas constituyen los árboles y arbustos leñosos o pueden ser herbáceas sin tejido leñoso.
Los árboles dominan muchos ecosistemas en los que el clima es húmedo y caliente durante parte del año. Sin embargo dejan de ser dominantes cuando el suelo es demasiado escaso para el crecimiento de las raíces, y donde es demasiado seco como en desiertos, o donde el suelo se congela permanentemente como en la tundra, o en sitios anegados durante parte del año, o en sitios dominados por sales o metales pesados o en ambientes alterados por el hombre. Los árboles pueden crecer por muchos años produciendo semillas o solamente por algunos años. La mayoría de los bosques son dominados por angiospermas dicotiledóneas a excepción de los grandes bosques de confieras del hemisferio norte. En muchos bosques los árboles forman estratos determinados por varios factores ambientales entre los que podemos mencionar la luz y el genoma de las especies que lo componen. Los helechos arbóreos, cycadas y monocotiledóneas arbóreas son comunes, pero no son dominantes o en algunas excepciones en pantanos o en situaciones en que son colonizadores. En muchos bosques los árboles forman estratos, compuesto de árboles grandes que emergen del dosel en algunos bosques tropicales y estratos secundarios formados por árboles y arbustos, frecuentemente formando más de una capa. Generalmente las especies tolerantes de la sombra se encuentran en los estratos inferiores y las especies heliófilas que toleran la luz, en los estratos superiores, sin embargo esta separación no es estricta, encontrándose árboles grandes en los estratos inferiores también. En los bordes o límites del bosque donde existen brechas se observa que crecen especies pioneras, de rápido crecimiento, como el balso y el yagrumo.
Muchas plantas herbáceas son anuales y mueren después de una estación de crecimiento, dejando solamente las raíces y algunas veces las hojas hasta la nueva estación de crecimiento. Las herbáceas junto a los arbustos pequeños forman la capa basal en muchas áreas, como en sabanas, mesetas, desiertos y llanos. Las gramíneas son casi todas herbáceas a excepción del bambú. Están bien adaptadas para resistir la presión del pastoreo, ya que en lugar de tener meristemas ápicales, están situados en los nudos, lo que les permite formar tallos por encima de los nudos. Las gramíneas que poseen meristemas subterráneos, pueden resistir la acción inclemente del fuego.

En los bosques tropicales predominan las plantas trepadoras y epifitas que requieren la presencia de un hospedero, que les proporcione soporte para su crecimiento. Las epifitas dependen de abundantes lluvias, ya que estas obtienen el agua de las precipitaciones y de la escorrentía a lo largo de los tallos y ramas.
En los bosques tropicales algunas epifitas son arbustivas, desarrollan un tallo leñoso, que puede formar raíces que alcanzan el suelo hasta hacerse independientes. Los Ficus (matapalos) y los matapalos del género Clusia son los ejemplos más conspicuos de estas epifitas estranguladoras, que se independizan del hospedero, matándolo.
Las Lorantáceas son hemiparásitas, ya que a pesar de realizar la fotosíntesis, penetran el hospedero a través de haustorios succionándole agua y nutrientes, ejemplo de esta tenemos la tiña y el guate pajarito.
Existen varios factores externos que afectan el crecimiento, como son: La Temperatura, la luz, el agua y ciertas sustancias químicas que lo estimulan o inhiben.
TEMPERATURA
El crecimiento de las plantas superiores ocurre en un rango de 0o a 350 C, en ese rango un aumento de 100C (Q10 está entre 2-3) aumenta la tasa de crecimiento 2 a 3 veces. Estas se conocen como las temperaturas cardinales o puntos de crecimiento: la mínima, o la menor temperatura a la que ocurre el crecimiento; el óptimo, la temperatura de máxima velocidad de crecimiento; y el máximo, o la temperatura más alta en la que se detecta el crecimiento. Estas no son temperaturas bien definidas y varían de especie a especie. Las temperaturas cardinales pueden variar con el estado de desarrollo de la planta. La velocidad de crecimiento de un órgano no depende solamente de su propia temperatura sino también del resto de la temperatura de la planta. Por ejemplo cuando se mantuvo constante la temperatura de una hoja de maíz a 250C y con una humedad relativa del 100 %, la tasa de crecimiento se incremento entre 50 y 150C, que están por debajo de la temperatura a la que se mantuvo la hoja. El crecimiento de plantas de maíz jóvenes se inhibe cuando la temperatura superficial del suelo excede los 350C. Así mismo debemos tener presente que el crecimiento depende de otros factores como son la fotosíntesis y la respiración y estos fenómenos poseen también temperaturas cardinales. La tasa de fotosíntesis de muchas plantas con el ciclo de carbono C3 alcanza un máximo entre 200 y 300C. En las plantas que fijan el carbono mediante el ciclo C4 la fotosíntesis aumenta rápidamente a una temperatura foliar entre 300 y 400C y luego a temperaturas más alta disminuye rápidamente. Las temperaturas cardinales para el crecimiento son de una gran importancia práctica, ya que el máximo crecimiento y productividad puede requerir un clima con un lapso de tiempo máximo cerca de la temperatura óptima. Debido a que las reacciones químicas que ocurren en las plantas están controladas por enzimas, una temperatura máxima debe ocurrir tan pronto como las enzimas se desnaturalizan y se inactivan. Esto sucede generalmente a una temperatura ligeramente por encima del máximo para un proceso biológico. Las temperaturas cardinales para el crecimiento deben estar controladas por la desnaturalización de las enzimas de las plantas. El crecimiento es un proceso que depende del tiempo, no solamente afecta la temperatura puntual; sino el número de horas o días en que un organismo se somete a una temperatura determinada. El crecimiento y desarrollo de las plantas, casi siempre responde a un termo período, que consiste en temperaturas alternas entre el día y la noche.El efecto de la temperatura sobre la germinación tiene una gran importancia en la ecología de poblaciones. El rango de temperaturas para la germinación de esporas y semillas debe corresponder a condiciones externas apropiadas, que permiten un rápido desarrollo de las plantas jóvenes. El rango de temperatura para el comienzo de la germinación es amplio en especies que están extensamente distribuidas y en aquellas adaptadas a altas fluctuaciones de temperatura en su hábitat natural.

LUZ
Aunque el crecimiento de las plantas superiores depende de la fotosíntesis, sin embargo la luz no es necesaria para el proceso de crecimiento per se, en la medida en que haya un buen suministro de sustancias orgánicas. Algunas plantas tuberosas y bulbos pueden completar su ciclo de vida en la oscuridad, a expensas de las abundantes reservas de alimentos, sin embargo el tipo de crecimiento es diferente cuando la luz esta ausente. En la oscuridad las plantas crecen largas, débiles y cloróticas, lo que se conoce como etiolación. En el caso de muchas dicotiledóneas, el tallo se encuentra excesivamente alargado y las hojas se desarrollan con deficiencia. La diferenciación es muy poca y los tejidos son principalmente parenquimatosos. Las hojas tienen ausencia de clorofila y el color es amarillo pálido; aunque se presentan excepciones entre las plántulas de las gimnospermas y algunos helechos, que pueden formar clorofila en la oscuridad. Las monocotiledóneas pueden mostrar el alargamiento excesivo del primer entrenudo y un crecimiento normal o un desarrollo excesivo de las hojas en la oscuridad. Una exposición diaria corta a la luz previene la etiolación. La luz retarda el crecimiento excesivo de las plantas etioladas. Las plantas parameras crecen achaparradas o arrosetadas debido a la luz excesiva en las grandes altitudes, la cual es muy rica en radiaciones violeta y ultravioleta, que parecen tener un efecto de enanismo marcado.
El primer efecto de la luz es iniciar los patrones de expresión genética, y la plántula comienza a formar cloroplastos fotosintéticamente activos, alterando sus formas de crecimiento de alargamiento rápido en la producción de hojas y un tallo capaz de soportarlas. Esto se conoce como fotomorfogénesis, que son los cambios de formas en respuesta al factor luminoso. Existen otros ejemplos como son la germinación de algunas semillas que germinan solamente después de ser sometidas a luz roja, ej., Alnus acuminata; así como la morfología foliar. Dependiendo del hábitat las hojas desarrollan una serie de características morfológicas especiales; si están expuestas al sol las hojas son más gruesas que las que crecen a la sombra, y tienen un parénquima en empalizada con células más largas que si las hojas crecen en la sombra. El movimiento de los cloroplastos es controlado por la luz. Bajo condiciones de baja luminosidad los cloroplastos se orientan de una forma perpendicular a la luz incidente y se agrupan en las células paralelamente a la superficie foliar. A altas intensidades luminosas los cloroplastos se mueven hacia la superficie celular que es paralela a la luz incidente, evitando la absorción excesiva de luz. El movimiento de los cloroplastos es una respuesta típica a la luz azul, así como el fototropismo. El crecimiento en respuesta a la luz roja implica la participación del fitocromo.
Fotoperiodismo es una respuesta estacional a la longitud del día y la noche del fenómeno de floración. Algunas plantas florean más rápido cuando la longitud del día fue de 12 horas o menos , son las plantas de días cortos y otras florearon cuando la longitud del día fue de 12 horas o más, plantas de días largos; mientras que otras plantas florearon independientemente de la longitud del día, son plantas neutras. Si la longitud del día no es apropiada para la floración, la planta permanece en estado vegetativo indefinidamente, o simplemente va a tomar un período más largo para florear. Tenemos un caso de los crisantemos ( pompon) que se siembran en la zona andina, a los que se les suministra un período de luz artificial para que continúen con el crecimiento vegetativo por cierto tiempo, luego se retira la iluminación artificial para que ocurra el fenómeno de floración ( es una planta de días cortos). El resultado son plantas largas con flores en un extremo del tallo, que pueden ser colocadas fácilmente en floreros. Este es un ejemplo de una aplicación práctica del fotoperíodo en la floricultura.
Fitocromos son proteínas fotorreceptoras azuladas. Son azules porque absorben la luz roja e infrarroja y transmiten el resto de longitudes de onda. En el citosol hay dos formas interconvertibles de fitocromo, la forma que absorbe luz roja de 660 nm se llama PR, cuando absorbe la luz roja, se transforma en PFR, forma esta que absorbe luz infrarroja de 730 nm y cuando lo hace se transforma en PR . Algunas semillas requieren una exposición a la luz roja para germinar, entre estas están las de lechuga, aliso (Alnus acuminata).
Los fitocromos participan como mediadores en una serie de respuestas de las plantas a la luz, como son:
Etiolación, en la que las plántulas o sus órganos se alargan rápidamente sin la producción de cloroplastos hasta tanto no reciben

luz roja. Al ser expuestas a la luz roja se forman cloroplastos funcionales.
Ritmos circadianos. Una serie de procesos metabólicos y de posicionamiento de las hojas responden a un ciclo periódico de 24 horas. La respuesta al fitocromo asegura la sincronía del ritmo con la longitud del día. Cambios en el turgor celular, como el observado en el movimiento circadiano de hojas y pétalos de algunas plantas.
Germinación. Muchas semillas son estimuladas a germinar por la luz, en una respuesta mediada por el fitocromo.
El fitocromo le permite a las plantas detectar la presencia y distancia de hojas de plantas vecinas.
Muchas plantas que crecen en hábitat abierto y claro del bosque, poseen semillas que germinan solamente cuando expuestas a la luz, con una alta proporción de radiación roja (la luz promueve la geminación). En cielo abierto la luz natural tiene una proporción de Rojo/Rojo lejano de 1,2 a 1,3; mientras que debajo del dosel del bosque la proporción de rojo lejano puede ser 2 a 10 veces mayor que la radiación roja. Las plantas que requieren más radiación roja no germinan y permanecen latentes, hasta tanto no se remueven las ramas y hojas, mediante talado o por acción de un fenómeno natural (tormenta, rayos, etc.). Así mismo las semillas que han sido expuestas al rojo lejano antes de ser enterradas en el banco, requieren de luz roja para germinar.
Los fitocromos también regulan cambios en la expresión del gen. Los fitocromos regulan un número de genes en el cambio de una planta etiolada a la normalidad, lo que resulta en la transformación de plastidios no fotosintéticos (etioplastos), a cloroplastos completamente desarrollados y con función fotosintética.
AGUA
Es indispensable para el crecimiento de las plantas, ya que en su presencia ocurren reacciones metabólicas, que participan en los procesos de crecimiento y desarrollo. El crecimiento depende de la existencia de una presión de turgencia; es por esto que un déficit hídrico lo puede retardar e interrumpir por completo. Un exceso de agua puede resultar en condiciones anóxicas que provocan un crecimiento anormal. En una atmósfera saturada de humedad ocurre un desarrollo pobre de las hojas y se retarda la diferenciación de los tejidos.
Las plantas hidrófilas, que viven en el agua tienen una presión osmótica baja y no poseen una presión de turgencia excesiva. Estas plantas desarrollan un parénquima aerífero con amplios espacios intercelulares, las hojas son delgadas y los estomas cuando presentes solamente se observan en la haz foliar.
A altas temperaturas se afecta el crecimiento por una evapotranspiración excesiva, lo que provoca una disminución de la presión de turgencia. A bajas temperaturas el crecimiento se puede inhibir debido a una disminución de la absorción de agua.
Ir al principio
ACVTIVADORES QUÍMICOS E INHIBIDORES
Las sales nutritivas son requeridas por las plantas para su crecimiento normal; sin embargo cuando se encuentran en soluciones nutritivas desbalanceadas pueden inhibir el crecimiento. Muchos iones son tóxicos para el crecimiento de las plantas a altas concentraciones. Algunos iones como el cadmio (Cd) y el aluminio (Al), son tóxicos inclusive a bajas concentraciones. La concentración de iones tóxicos aumenta cuando se

añaden toxinas al suelo mediante la contaminación atmosférica o en desechos industriales. Así mismo, si las condiciones de acidez del suelo cambia, se libera el aluminio a partir de complejos tóxicos insolubles. La toxicidad se manifiesta con una inhibición del crecimiento o en última instancia la planta deja de completar su ciclo de vida. La toxicidad puede causar:1) la inhibición de la absorción de agua, nutrientes o la fotosíntesis, 2) inhibición de la utilización de recursos metabólicos, como inhibición enzimática, daños a las membranas celulares., etc. En el cuadro siguiente se resume el efecto de algunos iones tóxicos.
Ión tóxico Condiciones Efecto
Aluminio Suelos ácidos debajo depH 4.
Inhibe el crecimiento de la raíz, se enlaza a fosfatos, ADN, ARN, destruye membranas y el metabolismo del ATP.
Boro ( a altas concentraciones)
Suelos contaminados con cenizas residuos de combustión.
Clorosis y necrosis de tejidos.
Cobre ( a altas concentraciones)
Suelos contaminados con residuos de minería.
Daño a membranas celulares de raíces, inhibe el crecimiento.
Magnesio( a altas concentraciones)
Suelos con una alta relación Mg/Ca. Causa deficiencia de Ca.
Manganeso ( a altas concentraciones) Suelos ácidos Causa deficiencia de Ca y Mg, inhibe el
crecimiento del vástago.
Sodio Suelos salinos, suelos irrigados
Compite con la absorción de potasio, efecto osmótico; los estomas permanecen abiertos.
Cloro Suelos salinos, suelos irrigados.
Efecto osmótico, compite por la absorción de otros aniones, produciendo deficiencia.
Algunas plantas pueden sobreviven bien adaptándose a la presencia de iones tóxicos en el suelo. Estas plantas crecen muy lento, pero como carecen de especies competidoras se benefician de esta situación. Hay especies como la Eichhornia crassipes (Bora o jacinto de agua) que acumula grande cantidades de elementos metálicos tóxicos, como son: Cd, Co, Pb, Hg, Ni y Au, a partir de aguas contaminadas, y produce una biomasa de 600 Kg. /Ha/ día, esta planta se puede usar para purificar aguas contaminadas.
La toxicidad por contaminación atmosférica se presenta cuando las plantas se exponen a gases tóxicos, producto de las emanaciones industriales y volcánicas.Entre estos contaminantes podemos mencionar el ozono, dióxido de azufre, oxido de nitrógeno y monóxido de carbono. Su efecto puede ser directo, inhibiendo la apertura estomática, daño a la superficie de las plantas, inhibición de enzimas o indirecto, alterando el pH del suelo, impidiendo la absorción mineral o liberando metales tóxicos a la solución del suelo, como el Al y Mn.
Ir al principio
FITOHORMONAS
Las hormonas vegetales o sustancias de crecimiento actúan en bajas concentraciones, específicamente para regular el crecimiento y el desarrollo de las plantas. Las hormonas vegetales desempeñan múltiples papeles reguladores que afectan aspectos diferentes del desarrollo. En las hormonas vegetales no existe una clara separación entre los sitios de síntesis y de acción.
Ir al principio

AUXINASs
La principal auxina vegetal es el ácido-3-indol acético. Existen otros compuestos con actividad auxínica, entre los cuales podemos mencionar el ácido fenoxi- acético y el ácido 3-indol butírico.
Las auxinas afectan el crecimiento y la forma de las plantas. Si se corta la yema apical en el extremo de una planta de arveja, los brotes laterales inactivos se hacen activos, y desarrollan ramas. Así mismo, cuando se poda un árbol, eliminando las yemas terminales, se aumenta la ramificación. Esto se conoce como dominancia apical. Si se reemplaza la yema apical con auxina, se inhibe el crecimiento de las yemas axilares, sugiriendo esto que una alta concentración de auxina en el ápice inhibe las yemas axilares. El transporte de auxina es polar. Si se corta la lámina foliar, pero se deja el pecíolo unido a la planta, el pecíolo se cae antes que si la hoja estuviera intacta, ya que en la lámina foliar se sintetiza auxina. Si una planta se mantiene en el interior de una habitación, pero cercana a una ventana por donde penetra la luz, la planta crece hacia la luz, esta respuesta se conoce como fototropismo. Los tallos presentan fototropismo positivo; mientras que las raíces crecen alejandose de la luz, mostrando fototropismo negativo. Este fenómeno se puede explicar asumiendo que la auxina se mueve hacia el lado sombreado, donde estimula un crecimiento celular mucho más rápido que en el lado expuesto a la luz, lo que origina la curvatura fototrópica. Cuando un tallo se encuentra acostado horizontalmente sobre el suelo, la auxina se mueve hacia el lado inferior del tallo, lo que se traduce en un crecimiento más rápido de las células del lado inferior, que produce su inclinación hacia arriba, esta inclinación se denomina gravitropismo negativo, mientras que las raíces se inclinan hacia abajo, mostrando gravitropismo positivo.
Las auxinas afectan el crecimiento de varias formas:
Inician el crecimiento de las raíces en estacas. Estimulan la separación de las hojas viejas de los tallos (abscisión). Mantienen la dominancia apical. Promueven el alargamiento del tallo e inhiben al alargamiento de la raíz. La auxina promueve la expansión celular aumentando la plasticidad de las
paredes celulares. Crecimiento de los tallos en relación a la luz, lo que asegura que las hojas
reciban una cantidad de luz óptima para la fotosíntesis (Fototropismo). Crecimiento de las raíces hacia el suelo (gravitropismo positivo) y de los tallos
hacia arriba (gravitropismo negativo). Tigmotropismo o crecimiento en respuesta al contacto con un cuerpo duro, lo
que produce el movimiento de las raíces alrededor de una roca o de los tallos de las plantas trepadoras alrededor de otras estructuras que le sirven de soporte.
La auxina controla el desarrollo de algunos frutos. Normalmente el desarrollo de un fruto requiere la fertilización de un óvulo; sin embargo el tratamiento de un ovario no fertilizado con auxina o giberelina produce la formación de un fruto sin la fertilización de un óvulo, por lo tanto carecen de semillas, este fenómeno se conoce como partenocarpia por ejemplo, fresas, tomates, pepinos, calabaza, naranjas. Los frutos partenocarpicos se forman espontáneamente en algunas plantas, incluidas las uvas sin semillas, las bananas o cambures cultivados.

La auxina promueve la diferenciación celular. Cuando un callo (masa de tejido no diferenciado) se pone a crecer en un medio nutritivo, con una concentración apropiada de auxina se forman raíces. Este efecto se observa también en estacas donde la auxina estimula la formación de raíces laterales. El patrón de formación de los órganos depende de la relación auxina: citocinina, en el medio de cultivo. Una proporción elevada de auxina favorece la formación de raíces; mientras que una concentración elevada de citocinina favorece la formación de brotes.
Las auxinas sintéticas tienen amplia aplicación en agricultura y horticultura. El ácido 2,4-dicloro-fenoxi acético (2,4-D) y el 2, 4,5 tricloro-fenoxi-acético (2, 4,5-T) se utilizan como herbicidas a altas concentraciones, especialmente para controlar eudicotiledóneas, ya que son inofensivos para las monocotiledóneas. Sin embargo por degradarse lentamente, son contaminantes del medio ambiente causando daños a los ecosistemas. El ácido naftaleno acético (NAA) se utiliza para promover el enrraizamiento de estacas.
Las auxinas naturales se sintetizan a partir del amino ácido triptofano, especialmente en hojas jóvenes, meristemas del vástago y frutos en desarrollo, y dondequiera que las células se estén dividiendo rápidamente.
Las auxinas promueven la diferenciación de nuevos tejidos vasculares en la parte del tallo que queda debajo de la yema terminal y de hojas jóvenes en crecimiento, la remoción de las hojas jóvenes impide la diferenciación vascular. En plantas leñosas perennes, la auxina producida por las yemas en crecimiento durante la primavera, estimula la actividad del cambium vascular en dirección basipeta. El nuevo anillo de crecimiento secundario comienza en las ramitas más pequeñas y progresa hacia abajo hasta el ápice de la raíz.
Ir al principio
CITOCININAS
Las citocininas estimulan la formación de brotes, promueven la división celular en tejidos cultivados. Así mismo ayudan a la germinación, inhiben el alargamiento del tallo, estimulan el crecimiento de los brotes laterales y retardan el envejecimiento foliar.
Las citocininas son derivados de la adenina, una base nitrogenada que forma parte de los ácidos nucleicos. Esta hormona se denominó citocinina debido a que promueve la división celular o citocinesis.

La citocinina es sintética y no se encuentra naturalmente, sino como producto de degradación del ADN . En las plantas se encuentra el producto natural citocinina , que fue aislado de la leche de coco y que promueve la división celular. A partir del endosperma inmaduro de maíz (Zea mays) se aisló una sustancia que tiene las mismas propiedades biologicas de la cinetina y se llamo zeatina.
Las citocininas se forman principalmente en los meristemas apicales de las raíces y se mueven hacia otras partes de la planta. Las agallas que forma la bacteria Agrobacterium tumefaciens sintetizan zeatina, que promueve el crecimiento de los tumores que se observan en plantas infectadas con esa bacteria.
Ir al principio
GIBERELINAS
El estudio de las giberelinas comenzó indirectamente cuando un biólogo japonés Kurosawa observó que algunas plantas de arroz enfermas de "bakanae o planta loca", tenian un crecimiento en altura mucho mayor que las plantas sanas, las que mueren antes de producir semillas. Esta enfermedad es causada por un hongo ascomiceto Giberella fujikuroi el cual produce giberelinas.
Se han aislado más de 100 giberelinas de las plantas, pero muchas carecen de actividad biológica. La más estudiada y la que probablemente es la de mayor actividad biológica es la GA3. La giberelina promueve el alargamiento del tallo, regulan la transición de la fase juvenil a la adultez, estimulando la formación de órganos florales.
Las giberelinas pueden sustituir el requerimiento de días largos para la floración en muchas plantas, especialmente en especies arrosetadas.
Las giberelinas regulan el crecimiento de los frutos. Las uvas sin semillas crecen más pequeñas que las uvas con semillas. La eliminación de las semillas de uvas muy jóvenes impidió el crecimiento normal de los frutos, lo que permitió concluir que las semillas producen un regulador del crecimiento del fruto. Luego se demostró que al asperjar uvas jóvenes sin semillas con una solución de giberelina, estas crecen tan grandes como las que tenían semillas. Actualmente se asperjan las uvas sin semillas, para que crezcan grandes y apetecibles. Estudios posteriores mostraron que las semillas en desarrollo producen giberelinas.

Las giberelinas rompen la latencia de las semillas que requieren bajas temperaturas (termoperíodo) o luz para germinar (fotoperíodo), como es el caso de las semillas de aliso (Alnus acuminata) en que 5 ppm de giberelina sustituye el requerimiento de luz. En algunas semillas, las giberelinas estimulan la actividad de enzimas hidrolasas que promueven la movilización de las reservas del endospermo durante el proceso de germinación.
fuente: http://iescarin.educa.aragon.es/depart/biogeo/varios/BiologiaCurtis/Seccion%206/6%20-%20Capitulo%2038.htm
La mayor actividad de las giberelinas se presenta en tejidos jóvenes de rápido crecimiento, como hojas, yemas, y semillas y frutos en desarrollo.Asi mismo estimula la germinación y el desarrollo del tubo polínico de algunas especies, como los lirios.
El transporte se realiza a través del floema y no muestra polaridad. Las giberelinas interactúan con la auxina controlando el alargamiento del tallo.
Ir al principio
ETILENO
El etileno o eteno, es un gas que regula la maduración de frutos. Se ha descubierto que la cercanía de frutos maduros como naranjas o manzanas acelera el proceso de maduración de otros frutos, como tomates y cambures. Es importante cuando se transportan y mercadean frutos, regular la producción de etileno con el propósito de retardar el proceso de maduración. El agricultor que requiere madurar con premura cambures o plátanos para su mercadeo, utiliza carburo de calcio, el cual libera al humedecerse acetileno que luego se convierte en etileno, acelerando el proceso de maduración.

El etileno se forma en la mayoría de los órganos de las plantas superiores, aunque los tejidos en fase de envejecimiento y frutos en maduración producen más etileno que los tejidos jóvenes o maduros. El amino ácido metionina es el precursor de la síntesis del etileno.
El etileno no solamente regula la maduración de frutos, sino los fenómenos asociados con el envejecimiento de flores y hojas, la abscisión de hojas y frutos; así como el desarrollo de los pelos radicales y crecimiento de plántulas. También regula la expresión de los genes de la maduración de frutos y de la patogenecidad.
El etileno induce la formación de aerenquima en raíces bajo condiciones de inundación.La preservación de frutos almacenados se alarga utilizando una atmósfera controlada, con baja concentración de CO2 (3 a 5 %) y bajas temperaturas, que inhiben la síntesis de etileno.
El etileno es una de las hormonas más utilizadas en agricultura, pero como es un gas es difícil su aplicación; sin embargo se han sintetizado compuestos químicos que liberan etileno, como el Etefon (ácido 2-cloroetil fosfónico) conocido también como Etrel, el cual acelera la maduración de manzanas, tomates, cítricas, sincroniza la floración y fructificación en piñas y acelera la abscisión de flores y frutos.
Ir al principio
ACIDO ABSCÍSICO (ABA)
Es una hormona que se encuentra en todas las plantas superiores y musgos, regula el crecimiento y la apertura estomática especialmente cuando las plantas están sometidas a estrés ambiental. Otra función importante es la regulación de la maduración y latencia de semillas.

ABA es transportado en la planta tanto por el xilema como por el floema, pero es más abundante en la savia floemática. Durante las condiciones de estrés provocada por la sequía las raíces sintetizan ABA, que se transporta por el xilema hacia las hojas que responden con el cierre estomático reduciendo la transpiración.
ABA mantiene la latencia de las yemas durante el invierno. Inhibe la germinación de las semillas. Inhibe el alargamiento del tallo. Se le denomina la hormona del estrés de las plantas, ya que se acumula
cuando las plantas son privadas de agua. Regula el intercambio de gas y vapor de agua entre la planta y el medio
ambiente al regular la apertura estomática.
La síntesis de ABA se realiza tanto en cloroplastos como en otros plastidios. En las plantas superiores su biosíntesis se realiza siguiendo la ruta de los terpenos.
Ir al principio
BRASINOESTEROIDES (BR)
Son un grupo de hormonas esteroideas polihidroxiladas que juegan un papel importante en fenómenos de desarrollo de las plantas, como son: división y alargamiento celular de tallos y raíces, fotomorfogénesis, desarrollo reproductivo, envejecimiento foliar, respuestas al estrés y germinación.
Los brasinoesteroides se encuentran presentes en las plantas a muy bajas concentraciones, sin embargo tienen un fuerte efecto como promotores del crecimiento, estimulando tanto la división como el alargamiento celular. Estos actúan conjuntamente con otras hormonas vegetales en la regulación del crecimiento y la diferenciación. Una planta mutante de Arabidopsis es enana, ya que no sintetiza brasinoesteroide; sin embargo al aplicarle brasinoesteroide artificialmente, la planta se alarga y empieza a crecer normalmente, indicando que este es esencial para el alargamiento celular en plantas normales.
BR juega un papel importante en el desarrollo celular, promoviendo la diferenciación del xilema y suprimiendo la del floema. Promueve el desarrollo del tubo polínico desde el estigma y a través del estilo

hasta el saco embrionario. Asi mismo, promueve la germinación de semillas de tabaco, independientemente del ácido giberélico.
Ir al principio
ACIDO JASMÓNICO (AJ)
La defensa de las plantas contra los herbívoros surge como una respuesta al daño ocasionado por insectos. Los insectos que se alimentan de las hojas de tomate dañan las células, conduciendo a la formación de una hormona polipeptídica la sistemina, lo que dispara la formación de otra hormona el ácido jasmónico como producto de la degradación de ácido linolenico, que es un ácido graso constituyente de la membrana celular. El ácido jasmónico entra en el núcleo de la célula, donde activa genes que programan la síntesis de un inhibidor de proteasas, lo que ocasiona que el insecto no pueda digerir las proteínas, retardando su crecimiento y provocando su muerte.
El ácido jasmónico se sintetiza a partir del ácido linolenico mediante la acción de enzimas que se encuentran en dos organelos celulares, los cloroplastos y los peroxisomas. El AJ activa la defensa de las plantas contra insectos y muchos hongos patógenos. Así mismo, regula otros aspectos del crecimiento de las plantas como son: el desarrollo de las anteras y el polen. Promueve la senescencia y abscisión de hojas, inhibe la germinación de semillas y el alargamiento de las raíces.
Ir al principio
POLIAMINAS
Son compuestos que tienen dos o más grupos aminos y son importantes para la vida vegetal. Ejemplos que tienen actividad biológica son:
Putrescina: H2N-(CH2)4-NH2 Espermidina : H2N-(CH2)3-NH--(CH2)3- NH2 Espermina: H2N-(CH2)3-NH--(CH2)3- NH-(CH2)3-NH2
Se originan a partir del amino acido arginina.

Las concentraciones de poliaminas aumentan durante la división celular.
Efectos fisiológicos:
Favorecen la floración. Incrementan la tolerancia al estrés. Promueven la división celular. Estimulan la senescencia en hojas cortadas.
CONTENIDOS
/Proteínas y aminoácidos/ A. Aminoácidos con cadenas laterales hidrofóbicas apolares /B. Aminoácidos con cadenas laterales neutras o hidrofílicas (polares) / C. Aminoácidos con cadenas
laterales básicas o hidrofílicas (polares) / D. Aminoácidos con cadenas laterales ácidas o hidrofílicas / El enlace peptídico / Péptidos / Propiedades ácido-base de los aminoácidos / Estructura de las proteínas / Estructura terciaria especificidad y desnaturalización / Diversidad funcional de las
proteínas/
PÁGINA PRINCIPAL
PROTEÍNAS Y AMINOÁCIDOS Las proteínas son macromoléculas de alto peso molecular formadas por carbono,
hidrógeno, oxígeno y nitrógeno. El contenido de nitrógeno de muchas proteínas es de 16 %, por lo que la cantidad de proteína se puede estimar multiplicando el contenido de nitrógeno por 6,25 (100/16). Algunas proteínas pueden contener azufre, hierro, cobre, magnesio, fósforo (fosfóproteínas), glucidos (glucoproteínas), lípidos (lipoproteínas), ácidos nucleicos (nucleoproteínas). Tienen pesos moleculares que varían de 6000 a 7.000.000, por hidrólisis liberan aminoácidos, que se pueden considerar como los sillares primarios de las moléculas proteicas. El alfabeto de la molécula proteica está constituido por 20 aminoácidos, que se pueden ordenar para formar un polipéptido, de hasta mil aminoácidos de largo.
Los aminoácidos poseen un grupo carboxilo (-COOH) y un grupo amino (-NH2) sobre el
carbono (central) y una cadena lateral o grupo R (resto o residuo) de aminoácidos.
Cuando los cuatro sustituyentes sobre el carbono central son diferentes se dice que el carbono es asimétrico y permite la existencia de isomeros ópticos. Existen dos posibles disposiciones de los
átomos enlazados al carbono (central) en los aminoácido, aunque solamente la estructura L, se

encuentra en las moléculas proteicas. La imagen óptica de la estructura L, el isomero D, se puede hallar en la naturaleza, pero nunca en proteínas típicas. Sin embargo, los D-aminoácidos se encuentran naturalmente en antibióticos y paredes celulares de bacterias.
A los aminoácidos (A-A) se les ha asignado un símbolo formado por tres letras, que se emplean para designar la composición y la secuencia de A-A en las cadenas polipeptídicas. La forma más significativa de clasificación de los diversos A-A se basa en la polaridad del grupo R, cuando se hallan en disolución acuosa a pH 7.0. Las cadenas laterales hidrofóbicas apolares apenas son solubles en agua, mientras que las cadenas laterales hidrofilicas sí lo son. Las cadenas laterales hidrofílicas a su vez se clasifican en neutras, ácidas y básicas. En las proteínas las cadenas hidrofílicas se encuentran expuestas al agua, mientras que las hidrofóbicas se hallan ocultas en el interior de la molécula proteica.
Ir al principio
A. AMINOÁCIDOS CON CADENAS LATERALES HIDROFÓBICAS APOLARES
Alanina, Valina, Leucina, Isoleucina, Prolina, Fenilalanina, Triptófano, Metionina, Cisteína y Glicina o Glicocola.
La alanina, valina, leucina, isoleucina, poseen hidrocarburos saturados como cadena lateral.
La prolina presenta un grupo amino secundario. El nitrógeno está presente en un anillo pirrolidina.
La fenilalanina y el triptófano poseen cadenas laterales aromáticas. El triptófano posee un anillo indol. A partir de este A-A se sintetiza el ácido indol acético (AIA), o auxina, que determina la dominancia apical de los tallos y es importante en el enrraizamiento de esquejes.
La metionina es uno de los dos aminoácidos que contienen azufre, posee un enlace tioéter (-S-CH3) en su cadena lateral.
La cisteína es el otro aminoácido que contiene azufre en forma de sulfhidrilo (-SR). La cisteína puede aparecer como tal o como cistina, en la cual dos moléculas de cisteína se hallan unidas mediante un puente disulfuro (-S-S-), formado por la oxidación de los grupos tiol. La cisteína desempeña un papel importante en la estructura de las proteínas, ya que sus dos mitades moleculares pueden hallarse presentes en dos cadenas polipeptídicas diferentes. Las enzimas con grupos -SH libres en sus centros activos son inhibidas por los elementos As, Pb y Hg.
La glicina o glicocola es el más simple de los aminoácidos, tiene un sabor dulce. Fue aislada a partir de hidrolizados de gelatina. Carece de un carbono asimétrico, por lo que no forma isomeros.

Ir al principio
B. AMINOÁCIDOS CON CADENAS LATERALES NEUTRAS O HIDROFÍLICAS (POLARES).
Asparagina, Glutamina, Serina, Treonina, y Tirosina.
La asparagina y glutamina son dos amidas derivadas de dos aminoácidos dicarboxílicos, el ácido glutámico y el ácido aspártico.
La serina fue aislada de la proteína de la seda sericina, contiene un grupo hidroxilo (-OH). En el centro activo de la quimotripsina el grupo -OH es reactivo.

La treonina también contiene un grupo -OH, pero no es tan reactivo en las enzimas como el de la serina.
La tirosina se forma cuando el hidrógeno de la posición para de la fenilalanina se reemplaza por un grupo hidroxilo (-OH). El resultado es que una cadena apolar se transforma en una polar. La tirosina se encuentra en los centros activos de algunas enzimas.
Ir al principio
C. AMINOÁCIDOS CON CADENAS LATERALES BÁSICAS O HIDROFÍLICAS(POLARES).
Lisina, Arginina y la Histidina
La lisina contiene un grupo amino (-NH2) primario unido a un carbono terminal, por lo que el
grupo amino lateral de la lisina se denomina grupo -amino. Se trata de una base relativamente fuerte, a pH 7.0 la especie iónica predominante es el ion amonio (-NH3+).

La arginina es el aminoácido más básico. A pH 7.0 su grupo guanidina se convierte en el ion gunidinio protonado. Este A-A es un constituyente de proteínas básicas como histonas y protaminas que se encuentran asociadas a ácidos nucleicos.
La histidina tiene en su cadena lateral un grupo imidazol. La histidina por descarboxilación, en presencia de la enzima histidina descarboxilasa produce la histamina. La histamina estimula la secreción gástrica.
Ir al principio
D. AMINOÁCIDO CON CADENAS LATERALES ÁCIDAS E HIDROFÍLICAS.
El ácido aspártico y el ácido glutámico son ácidos dicarboxílicos que tiene cuatro y cinco carbonos respectivamente. A pH 7.0 la cadena lateral se encuentra disociada confiriendo carga negativa a las proteínas de las que forman parte. Para enfatizar en este aspecto, se suelen preferir los términos aspartato y glutamato cuando nos referimos al ácido aspártico y ácido glutámico. Como nota de interés podemos decir que la sal sódica del ácido glutámico se usa como condimento.

Ir al principio
EL ENLACE PEPTÍDICO
Cuando se unen los aminoácidos para formar polipéptidos, el grupo amino (-NH2) de un aminoácido se une con el grupo carboxilo (- COOH) de otro aminoácido, mediante una reacción de condensación, formando un enlace peptídico. Esta unión va acompañada con la eliminación de una molécula de agua.
La cadena polipeptídica comienza con un aminoácido que aporta un grupo amino y se conoce como N-terminal, mientras que el último aminoácido tiene un grupo carboxilo libre, es el C- terminal. El enlace peptídico está formado por la combinación de un grupo carbonilo y un grupo amino.
La unidad que se repite en un péptido puede ser cualquier aminoácido y se denomina residuo de aminoácido.
Cuando se unen dos aminoácidos el producto es un dipéptido, tres es un tripéptido, seis hexapéptido, ocho octapéptido, si son muchos polipéptido.
En la nomenclatura sistemática los péptidos son acil aminoácidos, por lo que se nombran añadiendo la terminación acil, al aminoácido cuyo grupo carboxilo ha formado el enlace peptídico Ej. Glicilalanina, es diferente de Alanilglicina.

Ir al principio
PÉPTIDOS
Existen muchos péptidos de importancia biológica, estos incluyen hormonas como la oxitocina (9 residuos de A-A), estimula la contracción uterina, hormona adrenocorticotrópica (39 residuos de A-A), glóbulo anterior de la pituitaria, glutatión (3 residuos), es un agente reductor. La estructura molecular del glutatión se muestra abajo:
La insulina segregada por el páncreas, con un peso molecular de 5733, tiene 51 residuos de aminoácidos, por lo que se considera como un péptido. Posee dos cadenas de polipéptidos, la cadena A con 21 residuos de A-A y la cadena B con 30 residuos de A-A.
En la cadena A, se forma un puente disulfuro interno entre la cisteína de la posición 6 y la 11. Así mismo existen dos puentes disulfuros que unen las cadenas A y B, entre las cisteínas de la posición 7 y 7, Y las cisteínas de la posición 20 y la 19.

Responder a las siguientes interrogantes:1. ¿En las cadenas Ay B cuales son los aminoácidos con el N-terminal y el C-Terminal? 2. ¿Cuántos residuos de Ala, Val y Leu hay por cadena?3. ¿Cuántos son los residuos de cisteina?4. ¿Cuántos son los residuos de Lisina, Arginina e Histidina?5. ¿Cuántos son los residuos de Serina y Treonina?6. ¿Cuántos son los residuos de A-A polares ácidos?7. ¿Cuántos residuos de Triptófano hay en la molécula de insulina?8. ¿Cuántos residuos de Fenilalanina y Tirosina tiene la molécula de insulina? Estos son aminoácidos aromáticos ¿Por qué?
Ir al principio
PROPIEDADES ACIDO-BASE DE LOS AMINOÁCIDOS
Los aminoácidos cristalizan de sus soluciones acuosas como iones bipolares o iones híbrido s, tienen alto punto de fusión por encima de 200°C, característica de las sales. Los aminoácidos dependiendo del pH pueden actuar como una base, donde el grupo amino (- NH2) acepta un protón transformándose en NH3 + y como un ácido donde el grupo carboxilo (- COOH) se disocia cediendo un protón ( - COO- + H+) se dice que son anfóteros. La forma más correcta de representar la fórmula de un aminoácido es la dipolar o iónica, denominada zwitterion (palabra Alemana que significa ion anfótero).
A continuación se presenta un esquema donde se juega un poco con la adición y sustracción de protones, con la finalidad de visualizar lo que le sucede a la molécula de un aminoácido a medida que el pH se hace ácido o básico.
A un pH determinado un aminoácido se presenta casi exclusivamente en forma de Zwitterion, es decir con las formas iónicas positivas y negativas en iguales proporciones; ese pH se conoce como Punto Isoélectrico. A ese pH un aminoácido no migra en un campo eléctrico; así mismo presenta mínima solubilidad, por lo que se precipita al no formar enlaces de hidrógeno con el agua. En el punto isoélectrico las proteínas precipitan, se separan de sus mezclas y se pueden purificar. El punto isoélectrico se puede calcular a partir de los valores de pK de los grupos ácidos o básicos.

Eg., El aminoácido alanina tiene un pK1 (-COOH) de 2,34 y el pK2 (-NH2) de 9,69; Por lo que el PI = (2,34 + 9,69) / 2 = 6,02. Cuando el pH es 6,02 existe igual proporción de las formas iónicas con carga positiva y negativa. Por debajo de pH 6,02 la alanina se carga positivamente, por lo que migra hacia el cátodo y por encima de pH 6,02 está cargada negativamente por lo que lo hace hacia el ánodo.
Ir al principio
ESTRUCTURA DE LAS PROTEÍNAS
Las moléculas proteicas tienen cuatro niveles de organización.
I. Estructura primaria, está determinada por el enlace peptídico entre residuos de aminoácidos sucesivos. La estructura primaria debe dar respuesta a las siguientes interrogantes para que se pueda comprender la estructura proteica:
1. ¿Qué aminoácido se encuentra en el extremo N-terminal de la cadena polipeptídica (NH2 -terminal)?2. ¿Qué aminoácido se encuentra en el extremo C-terminal de la cadena polipeptídica (COOH-terminal)?3. ¿Qué aminoácidos se encuentran entre los dos extremos de la cadena, cuál es su secuencia, y la localización de los puentes disulfuros? (Observe el ejemplo de la molécula de insulina, en la que se muestran los aminoácidos terminales, la secuencia de A-A en las cadenas A y B Y la ubicación de los puentes disulfuros).
II. Estructura secundaria, se refiere a ciertas estructuras comunes que se repiten en la molécula
proteica. Se consideran dos tipos de estructuras secundarias la hélice y la lámina plegada. El componente más probable de la estructura secundaria de una proteína es la hélice
. La hélice se puede visualizar imaginándose una cadena de polipéptido enrollada en espiral a lo largo de una varilla pequeña. Un giro completo de la cadena está compuesta de 4 residuos de aminoácidos. La estabilidad de la hélice se la dan enlaces de hidrógeno entre los enlaces peptídicos de la cadena proteica (CONH). El enlace de hidrógeno se forma entre un grupo - C=O y un grupo - NH del cuarto (4) aminoácido que le sigue.
El otro tipo de estructura secundaria es la lámina plegada. En este caso los A-A se disponen en zig-zag. Este tipo de estructura se encuentra en la fibroina de la seda, en músculos y fibras

contráctiles. En la lámina se encuentran unidas dos o más cadenas por enlaces de hidrógeno (Eg. Flagelos, mitocondrias, cloroplastos).
III. La estructura terciaria, es la estructura tridimensional plegada de la cadena polipeptídica, que da origen a una conformación globular. La estabilización de la estructura terciaria requiere además de los enlaces de hidrógeno, de enlaces covalentes de disulfuro, enlaces salinos (iónicos), enlaces hidrofóbicos (apolares), fuerzas de van der Waals.
a) enlace salinob) enlace de hidrógenoc) interacción hidrofóbica

d) enlace disulfuroe) interacciones de van der Waals
IV. Estructura cuaternaria, está determinada por el ordenamiento de las subunidades de polipéptidos para formar una súper estructura, que se mantiene unida mediante fuerzas electrostáticas débiles no covalentes. Esta estructura está presente si la proteína tiene más de una cadena de polipéptido. El ejemplo clásico es la molécula de hemoglobina, que tiene cuatro cadenas de polipéptidos, constituidas por dos alfa globinas (con 141 residuos de aminoácidos cada una) y dos beta globinas (con 146 residuos de aminoácidos cada una).
Ir al principio
ESTRUCTURA TERCIARIA ESPECIFICIDAD Y DESNATUZALIZACIÓN
Especificidad. Las alteraciones o cambios en el plegamiento de la hélice alfa pueden alterar la especificidad de una proteína. La especificidad de una proteína significa que cada proteína podrá reaccionar químicamente con una sustancia específica, debido a la configuración externa de la molécula. Una alteración del plegamiento produce un cambio en la forma que afecta su superficie. Para que una reacción tenga lugar el acoplamiento entre la sustancia reaccionante y la proteína debe ser perfecto, para que los átomos de ambas moléculas estén lo suficientemente cerca que les permita reaccionar. Podemos establecer la comparación entre un guante y una mano, el guante debe calzar perfectamente en la mano para que cumpla su función protectora.Cualquier cambio en los niveles de organización de una proteína puede hacerla disfuncional. Las proteínas solubles juegan un papel muy importante en las reacciones bioquímicas a nivel celular, por lo que cualquier cambio en su conformación puede alterar los procesos metabólicos. La configuración espacial de una molécula proteica es un buen ejemplo de la relación entre estructura y función a un nivel molecular.
Desnaturalización. Es la perdida o destrucción de la estructura terciaria de una proteína, en la que la proteína pasa de una forma plegada o enroscada a una forma desenroscada o desplegada. La hélice alfa se estira o despliega, por rompimiento de enlaces de hidrógeno. En la proteína desnaturalizada ocurren cambios estructurales que afectan sus propiedades biológicas, como la hormonal, enzimáticas y la actividad inmunológica. . Se produce una pérdida de solubilidad, ya que se exponen grupos hidrofóbicos que hacen insoluble la proteína. La desnaturalización es generalmente un proceso irreversible, fuertemente endotérmico, que produce un aumento de la entropía o desorden del sistema.Los principales agentes desnaturalizantes son:a) Altas temperaturas, provocan la coagulación. Eg. Huevo cocido.b) Cambios bruscos de pH.c) En el laboratorio se pueden usar altas concentraciones de guanidina o urea (4-8 M), que rompen enlaces de hidrógeno.

DIVERSIDAD FUNCIONAL DE LAS PROTEÍNAS
DIVERSIDA FUNCIONAL DE LAS PROTEÍNAS
Estructurales Queratina, fibroina, colágeno, elastina, mucoproteínas, glucoproteínas.
EnzimasBiocatalizadores.Proteasas, ribonucleasas, amilasas, lipasas.Ficina(higo), papaína( lechosa), Bromelina(piña) huraína (Jabillo).
Hormonas Regulan el metabolismo, Eg, insulina.
Anticuerpos Proteínas formadas en respuesta a un antígeno. Eg., gamma globulinas
Contráctiles Actina y miosina de los músculos
Reserva Ovoalbúmina, ferritina, caseína, gliadina, faseolotoxina (Phaseolus vulgaris).
TransporteHemoglobina, citocromos, mioglobina.Leghemoglobina en nódulos deleguminosas (Fabaceae), ferrodoxina de los cloroplastos.
Toxinas
Ricina (Ricinus communis), Toxina difterica, toxina botulínica.Veneno de alacranes o escorpiones, tienen neurotoxina, cardiotoxina, lecitinasa, hialuronidasa.Veneno de coral (Micrurus corallinus) tiene colinesterasa, hialuronidasa, L-aminoácido oxidasa y otras.Cascabel (Crotalus durissus terrificus). Tiene colinesterasa, hialuronidasa, desoxiribonucleasa, lecitinasa y otras.

CONTENIDOS
/ ACIDEZ Y pH / pH y pOH / ACIDEZ TITULABLE Y SISTEMA TAMPÓN O BUFFER / ECUACIÓN DE HENDERSON-HASSELBALCH / PROBLEMAS PROPUESTOS / PROBLEMAS RESUELTO /
PÁGINA PRINCIPAL
ACIDEZ Y pH
En casi todas las células vegetales, el jugo vacuolar es más ácido que el protoplasma debido a la acumulación de ácidos orgánicos, los que se encuentran separados del citoplasma mediante la membrana vacuolar o tonoplasto, impidiendo la inactivación de las enzimas, que son sensibles a los cambios bruscos de pH.
Los ácidos orgánicos en los tejidos vegetales se encuentran en una concentración del 0,1% en base al peso seco y en algunos casos como en los limones es 100 veces mayor. La savia de muchas plantas son ligeramente ácidas con un pH de 6,0; aunque la de algunos frutos puede estar entre pH 2 y 3. Los cambios moderados de pH afectan el estado iónico de las enzimas especialmente alrededor del centro activo y con frecuencia también el del substrato. Cuando se mide la actividad enzimática a diversos pH, la actividad óptima generalmente se observa entre los valores de 5,0 a 9,0; sin embargo algunas enzimas como la pepsina, se activa a valores de pH entre 1,5 a 2,5 , la amilasa de la malta tiene un óptimo de 5,2 y la lipasa de la semilla de tártago(Ricinus communis) de 5,0.
En todos estos casos, la actividad declina a ambos lados del óptimo.
La reacción de un suelo es el grado de acidez o alcalinidad que este posee y se obtiene al medirle el pH. Los suelos ácidos son típicos de regiones con alta precipitación en la que ocurre el lavado de los cationes básicos ( Ca+2 , Mg+2, Na+ , K+) con un predominio de cationes ácidos ( Al+3, H+), tanto en los sitios de intercambio como en la solución del suelo, mientras que los suelos alcalinos, generalmente están asociados a regiones áridas o semiáridas y con un alto valor de saturación de bases. Los pH alcalinos inducidos por encalamiento pueden originar una clorosis en las plantas, debido a una deficiencia de hierro, ya que el pH elevado provoca la indisponibilidad del hierro para las plantas.
Según Bronsted (1923) un ácido se define como una molécula que libera un protón (H+) y una base como una molécula que acepta un protón (H+) en solución. Por ejemplo:
El agua es una molécula anfiprótica, que se comporta como un ácido y como una base débil.Cuando se comporta como un ácido, libera un protón y forma un ión hidroxilo.
Cuando se comporta como base, acepta un protón para formar el ión hidronio ( H3O+ ). Muchos protones en solución existen como ión hidronio ( H3O+ ).
Para simplificar la exposición, el ión hidronio ( H3O+ ) se escribirá en forma de ( H+ ), aunque sabemos que existe en la forma hidratada.
Ir al principio

pH y pOH
La acidez y la alcalinidad de una solución se expresa frecuentemente como el logaritmo del inverso de la concentración de iones de hidrógeno [ H+] en moles por litro (M). Este valor se conoce como el pH de la solución. Por definición
tenemos:
pH = - log [ H+] .El pH es el logaritmo con signo negativo de la concentración de iones de hidrógeno.
Así mismo la concentración de iones de hidrógeno se calcula de la siguiente forma: .
Por ejemplo si [ H+] es 1x 10-3 M , entonces el Sí el pH de una solución es por ejemplo 4,5; la concentración de iones de hidrógeno es:
Si la concentración de iones de hidrógeno[ H+] se expresa como una potencia de 10 y el exponente es 1, entonces el valor del pH de una solución es igual al exponente con signo positivo.
Similarmente podemos definir el . El pOH es el logaritmo con signo negativo de la concentración de hidroxilos.
Así mismo la concentración de hidroxilos se puede calcular de la siguiente forma:
En soluciones acuosas , esta constante se conoce como producto iónico del agua. El valor de Kw a 250 C es 1.0 x 10-14 . En una solución ácida, la concentración de H+ es relativamente alta y la correspondiente concentración de OH- es baja.
En una solución neutra a 250C, y el pH de esa solución es 7.0.
La siguiente relación es válida: pH + pOH = 14 a 250C.
Por ejemplo, calcular el pH de una solución de NaOH 0,001 M, ,Entonces pH = 14 - pOH = 14-3 = 11. La respuesta el pH 3.Otro ejemplo, sí el pH de una solución es 2,6, su pOH = 14- 2,6 = 11,4. Mientras más pequeño sea el pOH, mayor será la alcalinidad. La tabla que se muestra a continuación resume las relaciones estudiadas.
En términos de pH, el pH de la savia celular varia frecuentemente entre 1 y 7, siendo la de pH 1 un millón de veces más ácida que la de pH 7. Una variación de una unidad de pH representa un aumento o una disminución de la acidez de 10 veces. Así tenemos que al aumentar el pH de 1 a 2 y de 6 a 7, en cada caso representa una reducción de la acidez de un décimo ( 0,1) del valor original. Una disminución de una unidad de pH, representa un aumento de diez veces la acidez:

¿Cuantas veces es una solución de pH 3,2 más ácida que una solución de pH 4,75?
a) [ H+] = 10 - 3,2 = 0,0006309
b) [ H+] = 10 - 4,75 = 0,0000177
c) Se divide
En la siguiente tabla se muestran los valores de pH y acidez de algunos fluidos comunes:

Ir al principio
ACIDEZ TITULABLE Y SISTEMA TAMPÓN O BUFFER
La acidez actual es una medida de la concentración de iones de hidrógeno y se determina con el pH; mientras que la acidez titulable o normalidad del ácido se determina por titulación o valoración, mediante una base de normalidad conocida. En otras palabras la acidez titulable se puede determinar añadiendo suficiente álcali de normalidad conocida hasta neutralizar toda la acidez o llevar el pH al punto neutro. La acidez titulable no se puede predecir a partir de la concentración actual de iones de hidrógeno que se determina con el pH.
ACIDEZ TITULABLE (NORMALIDAD) Y pH DE ALGUNAS SAVIAS DE PLANTASÓrgano Normalidad pH
Fruto de limónPecíolo de ruibarbo
Uvas verdesHojas de OxalisManzana verde
Hojas de Begonia rexHojas de Begonia tuberosa
0,950,220,210,160,130,110,10
2,43,23,02,33,22,22,2

Tomates madurosPecíolos de apio España o Celery.
0,0630,025
4,45,2
Los ácidos débiles no se encuentran totalmente disociados y tienen casi todos sus átomos de hidrógeno en la forma de iones de hidrógeno potenciales. Cuando un ácido débil se mezcla con su sal, mantiene un pH casi constante, aunque se le añada grandes cantidades de iones de hidrógeno o de hidroxilo. La mezcla de un ácido débil y su sal o de una base débil y su sal, que evitan los cambios bruscos de pH, se denomina solución buffer o tampón. Por ejemplo, cuando se añade 0,01 mol de HCl a un litro de agua pura, el pH cambia de su valor inicial que es 7 a 2, un cambio de 5 unidades de pH. El pH se hace 100.000 veces más ácido. Sin embargo, cuando se añade 0,01 mol de HCL a una solución que contiene una mezcla de ácido acético 0,1 M (CH3 COOH) y acetato de sodio (CH3 COONa) 0,1M, el pH cambia de un valor inicial de 4,74 a 4,66, un cambio de solo 0,08 unidades de pH, lo que es aproximadamente 1,2 veces más ácida.El pH se puede determinar mediante un electrodo de vidrio, que es sensible a la concentración de iones de hidrógeno. El electrodo de vidrio se calibra contra un buffer de pH conocido. Otra forma de medir el pH es utilizando indicadores, muchos de los cuales son ácidos débiles (designado HInd). La especie aceptora de protones del indicador (Ind-) tiene un espectro de absorción diferente que la especie donadora de protones:
Los indicadores son colorantes que cambian color a un pH más o menos específico. Las flavonas cambian de incoloras a amarillas por encima de pH 8; las antocianinas son rosadas o rojas por debajo de pH 5 y azul o púrpura por encima de ese pH.
Vamos a realizar algunos ejercicios para reforzar los conceptos estudiados:
1. ¿Calcular la concentración de protones de un vino tinto cuyo pH es 3,7?[H+] = 10 -pH por lo tanto [H+] = 10-3,7 o lo que es lo mismo que: 2,0 x 10-4 M.
2. El pH del agua de lluvia de un área contaminada se determinó que era 3,50. ¿Cuál es el valor de [H+] para esa agua de lluvia? [H+] = 10 -pH por lo que [H+] = 10-3,5 o lo que es lo mismo que: 3,16 x 10-4 M.
3. Se encontró que el pOH del agua de una pecera era de 6,59. ¿Cuál es el valor de [OH-] para el agua? [OH-] = 10-
pOH por lo que [OH-] = 10-6,59 o lo que es lo mismo que:2 x 10-7M. Así mismo podemos calcular el pH usando la relación pH + pOH =14, de donde el pH es 14-6,59 = 7,41.
Ir al principio
ECUACIÓN DE HENDERSON - HASSELBALCH
Esta ecuación relaciona el pH de la disolución y el pKa de un ácido débil (AH).

Cuando la concentración de la sal es igual a la concentración del ácido, el pH = pK; ya que el log de 1 es igual a cero. Las soluciones buffers tienen su máximo poder de amortiguación cuando el pH es igual al pK.El protoplasma celular es amortiguado por sus proteínas, las que son efectivas en un rango de pH de 4 a 10. Algunas savias celulares no tienen un buen poder de amortiguación; ya que su pH puede variar por adición de CO2. En las plantas con el metabolismo ácido de crasuláceas el pH puede estar cercano a 3. En muchas plantas el pH disminuye durante la noche y se hace más básico durante el día. En hojas de Bryophyllum el pH durante el día puede ser 5,5; pero durante la noche puede caer a 3,8, en estas plantas el cambio de pH se debe a la acumulación de ácidos orgánicos durante la noche y su disminución durante el día.
PROBLEMAS PROPUESTOS
1. Calcular el pH de las soluciones que tienen las siguientes concentraciones de [H+] o de [OH-] :
a. 3,5 x 10-4 M [H+]b. 1 x 10-3 M [H+]c. 1 x 10-9 M [H+]d. 1 x 10-4 M [H+]e. 7 x 10-6 M [H+]f. 5 x 10-8 M [H+]g. 3 x 10-11 M [H+]h. 1 x 10-3 M [OH-]i. 7,8 x 10-4 M [OH-]
2. Calcular la concentración molar de [H+] de las soluciones cuyos pH se muestran a continuación:
a. 2,73 ; b. 5,29 ; c. 8,65; d. 11,12
3.¿Calcular cuantas veces es más ácida una solución de pH 2 que una solución de pH 5?
4. Se titularon 25 ml de jugo de naranja con 5,2 ml de KOH 0,5 N. ¿Calcular la normalidad del jugo?
Ir al principio
PROBLEMAS RESUELTOS
1. Se titularon 25 ml de jugo de piña con 5 ml de KOH 0,5 N. ¿Calcular la normalidad del jugo de piña?
Respuesta: Volumen conocido (V1) =5 ml x Normalidad conocida (N1) =0,5 N es proporcional al Volumen del desconocido (V2)= 25 ml x Normalidad del desconocido
.
La normalidad del jugo de piña es 0,1 normal.
2.Calcular el pH de la solución final cuando se añaden 100 ml de NaOH 0,1 M a 150 ml de CH3 COOH 0,2 M (Ka = 1,8 x 10-5 ).
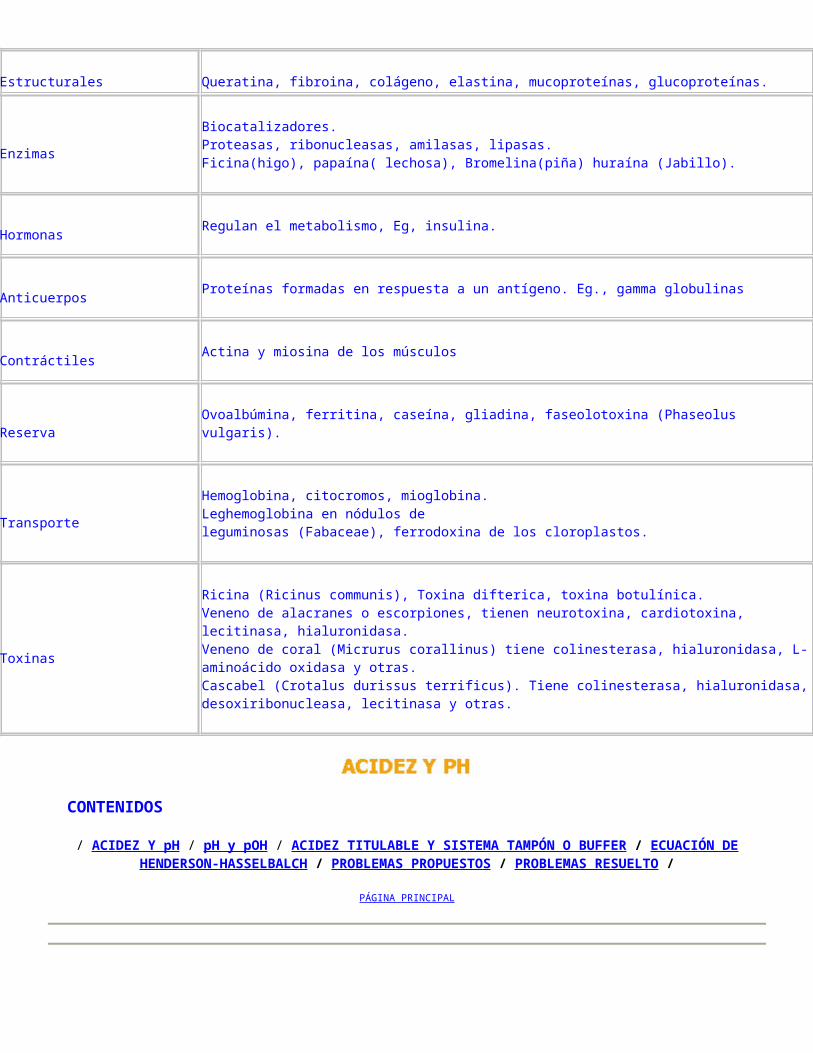
Solución: Sabemos que 150 ml de CH3 COOH 0,2 M tienen 0,03 moles de CH3 COOH. Así mismo 100 ml de NaOH 0,1 M contienen 0,01 moles de NaOH . Al mezclarse estos, 0,01 moles de NaOH neutralizan 0,01 moles de CH3 COOH para formar 0,01 moles de acetato de sodio y permanecerán libres 0,02 moles de CH3 COOH. Ambos están contenidos en un volumen de 250 ml.Aplicando la ecuación de Henderson-Hasselbalch tenemos:
Primero debemos calcular el pKa = - log 1,8 x 10-5 = 4,74 De tal forma que:
3. Determinar la relación de las concentraciones de para preparar un buffer de pH 5,0. El pKa del ácido acético es 4,66.
4. Determinar el pH de un buffer lactato, que tiene una concentración de ácido láctico de 0,050M, y la concentración del ión lactato es 0,32 M, sabiendo que la constante de disociación aparente del ácido láctico ( CH3- CHOH- COOH ) es 1,38 x 10-4 . Se calcula primero el pKa que es igual a: pKa= - log 1,38 x 10-4 = 3,86.
5. El pH del medio interno de una célula es 6,4. Calcular la relación de las concentraciones de: si la segunda constante de disociación para el ácido fosfórico K´ es 6,31 x 10-8.
CRECIMIENTO DE LAS PLANTAS Y pH.
El rango de pH para el crecimiento de las plantas está generalmente entre pH 6 - 7, sin embargo este varia con las diferentes especies de plantas, ya que hay especies que toleran muy bien la acidez y otras los suelos básicos.
CONCEPTO DE pH E IMPORTANCIA EN FERTIRRIGACIÓN
Enlace de interés:
o http://www.infoagro.com/abonos/pH_suelo.htm
Bibliografías:

http://web.ukonline.co.uk/fred.moor/soil/ph/p010601.htmhttp://web.ukonline.co.uk/fred.moor/soil/ph/p010602.htmhttp://web.ukonline.co.uk/fred.moor/soil/ph/p010604.htmhttp://web.ukonline.co.uk/fred.moor/soil/ph/p010603.htmhttp://web.ukonline.co.uk/fred.moor/soil/ph/p010605.htm
CONTENIDOS
/ DISTRIBUCIÓN DE LA VEGETACIÓN EN RELACIÓN AL CLIMA / VEGETACIÓN DE VENEZUELA / TIPO DE ÁREAS BOSCOSAS / SELVAS SIEMPRE VERDES / SELVAS TROPÓFILAS / BOSQUES XERÓFILOS /
MANGLARES / ÁREAS TERRESTRE NO BOSCOSAS / PÁRAMOS / SABANAS DE TIERRAS ALTAS DE GUAYANAS / SABANAS / DUNAS / ÁREAS ACUÁTICAS /
PÁGINA PRINCIPAL
DISTRIBUCIÓN DE LA VEGETACIÓN EN RELACIÓN AL CLIMA
La energía del sol dirige los patrones de circulación global del aire de la atmósfera y de las aguas de los océanos. El calentamiento y el enfriamiento de las masas de aire y de agua en movimiento explica la mayoría de los patrones climáticos de la tierra. Los climas, a su vez ejercen una gran influencia en la distribución, la abundancia y la evolución de las especies.
Los factores climáticos más importantes son la temperatura o factor térmico y la precipitación o factor agua, que varían durante todo el año. La temperatura desciende desde el Ecuador hacia los polos y las oscilaciones anuales son cada vez mayores. En la región de los trópicos, las precipitaciones son lluvias cenitales, se producen en la época del año en que el sol se encuentra en el cenit al mediodía. Por el contrario, en las zonas templadas encontramos lluvias ciclonales, es decir están ligadas a la presencia de zonas de baja presión que se mueven de Oeste a Este y que en el hemisferio Norte parten de Islandia o las Aleutianas.
El clima es un factor importante para el crecimiento de las plantas, determina la extensión de su área de distribución y fija los límites para su supervivencia. El clima regional o macroclima es el área donde se encuentra una estación meteorológica, mientras que el clima local o microclima es el hábitat en el cual crece una comunidad vegetal. Es importante por ejemplo la exposición: en latitudes Norte, la orientación hacia el Sur es más caliente, hacia el Norte más fría y en el hemisferio Sur ocurre lo contrario. En el Ecuador, la orientación hacia el Este está expuesta al sol durante el curso de la mañana, mientras que la orientación hacia el Oeste toda la tarde; las orientaciones hacia el Norte y Sur casi no difieren en cuanto a exposición. Los valles profundos tienen un clima húmedo, con nivel freático poco profundo y con abundantes precipitaciones; sin embargo en áreas con bajas precipitaciones, el nivel freático es más profundo, con el predominio de una vegetación xerofítica.
La energía del sol dirige los patrones de circulación global del aire y de las aguas de los océanos. El calentamiento y enfriamiento de las masas de aire y de agua en movimiento explica la mayoría de los patrones climáticos de la Tierra. Los climas, ejercen una gran influencia sobre, la abundancia y distribución de las especies.
En la publicación de Walter (1977), denominada Zonas de Vegetación y Clima, se considera en una escala global la distribución de la vegetación, según el clima zonal y el tipo de suelo las siguientes Zonas Climáticas (ver cuadro).
ZONAS CLIMÁTICAS DE LA TIERRA, TIPOS DE SUELO Y VEGETACIÓNClima Suelos Vegetación
1. EcuatorialCon oscilaciones diurnas, precipitaciones abundantes.
1.Arcillas marrones suelos ferralíticos. Ultisoles.
2. Arcillas rojas y tierras
1. Bosque lluvioso tropical, formas estacionales casi ausentes.

2. TropicalPrecipitaciones en verano y estación seca fresca ( húmeda hasta árida).
3. SubtropicalClima desértico, árido.
4. MediterráneoLluvias en invierno y seco en verano (semiárido a semihúmedo)
5. Cálido-templado.Con lluvias frecuentes en verano o medio marítimo.
6. Frío-templado.Con inviernos cortos y fríos.
7. Continental.Árido-templado, inviernosfríos.
8. Boreal.Frío-templado, con veranos fríos y largos inviernos.
9. PolarVerano muy corto.
rojas, suelos de sabana ferralíticos. Oxisoles.
3.Suelos desérticos (arenosoles, yermosoles, xerosoles), también suelos salinos. Aridisoles.
4. Tierras Mediterráneas marrones, frecuente terra rossa fósil. Alfisoles
5. Suelos forestales amarillos o rojos, ligeramente podsolizados. Podzoles.
6. Tierras forestales marronesy suelos forestales grises.
7.Suelos de estepa (chernozem o tierras negras).Mollisoles.
8. Podzoles o humus crudo,tierras blanqueadas.
9. Suelos de Tundra muy ricos en humus, con soliflux-ión pronunciada. Aridisoles.
2. Bosques tropicales deciduos o sabanas.
3. Vegetación subtropical desértica, predomina paisaje pedregoso.
4. Plantas leñosas esclerófilas, sensibles a heladas prolongadas.
5. Bosques siempre verdestemplados.
6. Bosques deciduos ( desnudos en invierno).
7. Estepa a desierto, solofrío en invierno.
8. Bosques de confieras.
9. Vegetación de Tundrasin árboles, generalmenteen permafrost.
Ir al principio
VEGETACIÓN DE VENEZUELA
Venezuela tiene una superficie terrestre de 916.445 Km2 . De este total corresponden 1.270 Km2 a las islas venezolanas del Mar Caribe. Está situada entre el ecuador y los 12° 12‘ de latitud norte, en el Cabo San Román de la península de Paraguaná, mientras que el país se extiende entre los meridianos 59° 47‘ y 73° 23‘ de longitud oeste. Presenta alturas que van desde el nivel del mar hasta el pico Bolívar, en la Cordillera andina con 4980 m lo que determina la existencia de cuatro pisos altitudinales que son: la tierra caliente, entre 0-1500 m s.n.m., con temperaturas entre 20° -30° C, la tierra templada entre 1500 - 2500 m s.n.m., con temperaturas entre 12° -20° C, la tierra fría entre 2500 - 3500 m s.n.m., con temperaturas entre 5° -12° C y la tierra helada o paramera, por encima de 3500 m s.n.m. con temperaturas por debajo de 5° C.

Se estima que más del 50% del territorio nacional está ocupado por bosques. La mayor parte de estos bosques están situados al sur del Orinoco, en el Estado Bolívar y Estado Amazonas; otras áreas boscosas están situadas en las Cordilleras de la Costa y de los Andes, Sierra de Perijá, sur del Lago de Maracaibo y Territorio Delta Amacuro. Las áreas no boscosas, como llanos y páramos ocupan aproximadamente el 45% del territorio, y el resto está ocupado por ríos, lagos y áreas cultivadas.Link. http://www.mipunto.com/venezuelavirtual/mapas/mapa_vegetacion.html(Instituto Geográfico de Venezuela Simón Bolívar-2003).
Los tipos de vegetación en Venezuela muestran una gran biodiversidad determinados por diversos factores como son la topografía, exposición al sol, temperatura, precipitación anual, suelos, la velocidad y dirección de los vientos, el drenaje del suelo y la historia geológica del área.
Ir al principio
La vegetación de Venezuela se clasifica según Steyermark(1966) de la siguiente forma:
I. Tipos de áreas boscosas
A.Selvas siempre verdes
Con precitaciones anuales que varían entre 1700 a 3800 mm (1 mm= 1 lítro de agua por metro cuadrado), aunque en algunos años puede descender hasta los 1500 mm y en las partes más altas de los bosques andinos puede variar entre 590 y 780mm. Estos bosques están situados en la parte más baja de la Tierra Caliente (0- 400 m.s.n.m.) o un poco más alto, en altitudes más frías de la Tierra Templada, generalmente en elevaciones de 800 a 3500 m.s.n.m. , donde se conocen con el nombre de selvas nubladas, debido a la alta condensación de nubes en estas altitudes y en la Tierra Fría con altitudes de 4200 metros, para formar el Bosque Altoandino Paramero, donde las temperaturas pueden llegar al punto de congelación.
a. Selvas pluviales.
Como se desarrollan en regiones de alta temperatura, se les conoce también como selvas pluviales macrotérmicas o megatérmicas, y como selvas pluviales higrófilas u ombrófilas por su ambiente siempre húmedo . Estos bosques se presentan en regiones con temperaturas anuales promedio de 26 0 a 28 ° C, generalmente en altitudes desde el nivel del mar hasta más o menos 400 metros, con precipitaciones de 2000 ó más milímetros, distribuida durante todo el año. En algunos casos se presenta un período de sequía de dos meses. Estas selvas se encuentran en Delta Amacuro, Guatopo en el estado Miranda, Estado Yaracuy, Sierra de Perija en el Zulia, la mayor parte del Estado Bolívar y Estado Amazonas.
Una característica de estos bosques es la majestuosidad de los árboles que alcanzan de 40 a 50 metros de altura, algunos pueden llegar a 60 metros. El sotobosque es húmedo y sombrío, con vegetación escasa, las raíces de los árboles son superficiales y presentan raíces tabulares. Se observan tres estratos bien definidos, entre los árboles más altos, que proveen una densa sombra y una alta humedad, manteniendo el suelo sombreado y húmedo. En estos bosques se producen cientos de especies de árboles, arbustos, trepadoras leñosas altas o lianas, que pertenecen en su mayoría a las Bignoniaceae. Estas últimas trepan a las coronas de los árboles más altos donde florean. Estos bosque muestran una gran diversidad, pudiéndose contar 87 especies diferentes de árboles por hectárea de 423 individuos arbóreos, este número puede exceder el centenar en otras áreas. En los árboles de este tipo de bosques se encuentran

muchas epífitas, tales como helechos, orquídeas, bromelias, aráceas, peperomias, musgos y hepáticas. Algunas epífitas de las Aráceas como Philodendron y Monstera desarrollan raíces adventicias aéreas muy largas que pueden tener hasta 45 metros. Las selvas pluviales no presentan periodicidad en los fenómenos de desarrollo. Así la época de floración y fructificación no están bien marcadas, ni tampoco una periodicidad en la pérdida del follaje. Podemos encontrar que diferentes individuos de una misma especie florecen en épocas distintas. Sin embargo, algunas especies muy sensibles pueden mostrar durante breves períodos de sequía, floración y fructificación, así como la pérdida parcial o total del follaje y cierto cambio en la coloración del mismo.
Los árboles de estos bosques poseen raíces superficiales, y desarrollan amplias extensiones laminares a manera de contrafuertes, que se extienden desde la base del árbol, hasta distancias considerables, como en algunas especies de higuera o matapalo (Ficus). En algunas palmas se forman raíces alargadas en forma de zancos, que le dan soporte al tallo. En estos bosques están representadas muy bien la familia Fabaceae, Sapotaceae, Lauraceae, Annonaceae, Moraceae, Lecythidaceae, Vochysiaceae, Bombacaceae, Sterculiaceae, Meliaceae, Simarubaceae, Sapindaceae, Meliaceae, Anacardiaceae, Rutaceae, Combretaceae, Palmae, Guttiferae, Apocinaceae, Myristicaceae, etc. Las selvas pluviales son muy ricas en especies, con una flora exuberante y de gran diversidad, que desconcierta al botánico más avezado que se siente perdido ante tal magnificencia.
b. Selvas nubladas.
Son bosques frecuentemente cubiertos de nubes o neblinas, que reciben además de la lluvia, una cantidad de humedad por medio de intercepción o precipitación horizontal o condensación de pequeñas gotitas de agua , que influyen en el régimen hídrico y en el balance de radiación y así en los demás factores climáticos, edáficos y ecológicos.
Este tipo de bosque, algunas veces denominado selva subtropical y selva pluvial alta, se desarrolla en ambientes de temperaturas moderadas o intermedias, donde entonces se le llama mesotérmica. A estas selvas también se las denomina higrófilas u ombrófilas por su ambiente húmedo. En la cordillera de la Costa, la selva nublada se presenta entre los 700 y los 2.500 msnm. En los Andes, generalmente, desde los 2.000 hasta los 3.000 msnm; en las tierras altas de Guayana, entre los 1.000 y los 2.500 msnm. Normalmente, sus límites de temperatura media anual están comprendidos entre 18 y 24° C, aunque en las partes frías de la cordillera de la Costa la temperatura mínima puede llegar a 4-6° C, mientras que en la cordillera de los Andes las temperaturas mínimas pueden llegar a 0° C. La precipitación promedio anual varía considerablemente según la zona; así, en ciertas partes de los Andes y en tierras altas de Guayana están entre 2.000-3.800 mm; en la cordillera de la Costa, entre 1.500-2.200 mm; y en la zona más alta de los Andes puede alcanzar sólo 550-780 mm.
Las nubes y la neblina rodean diariamente y durante varias horas la selva nublada, que se originan por las corrientes ascendentes de aire calentado por el sol a nivel del mar. Debido a este fenómeno, es difícil distinguir las formas de las hojas y las flores de los árboles altos, así como las trepadoras altas y las plantas epífitas ubicadas entre las ramas superiores. Este ambiente produce una vegetación muy rica y característica, con muchas especies arbóreas. El número de especies presentes en un bosque nublado es usualmente menor que el encontrado en un bosque pluvial. Los árboles alcanzan alturas entre 35-40 m de alto, pero en promedio son más bajos que los del bosque pluvial, y además, hay menos estratos debajo de los árboles altos. Aun cuando están presentes las trepadoras leñosas, son mucho menos frecuentes que en la selva pluvial; sin embargo, la vegetación epífita, mucho más desarrollada y diversificada, está representada por orquídeas, bromelias, aráceas, helechos, peperomias, musgos y hepáticas. Son también abundantes los helechos arborescentes. El suelo es usualmente húmedo y muy rico en materia orgánica. Entre las palmas indicadoras de este tipo de bosque se reportan especies pertenecientes a los géneros: Euterpe, Oenocarpus, Ceroxylon, Geonoma, Catoblastus, Chamaedorea y Dictyocaryum; como Gimnospermas representativas tenemos especies del género Podocarpus.
En estos bosques donde no se observa un período de sequía marcado, los procesos de polinización y dispersión de frutos dependen de factores bióticos. Se observa que hay especies en floración durante todo

el año y que aparentemente cada forma de vida tiene un comportamiento diferente; esto sugiere que cada especie tiene polinizadores específicos, lo que garantiza poca competencia a este nivel. Cuando se alcanza los límites superiores de los bosques nublados, tanto de la Cordillera de la Costa, los Andes, o las tierras altas de la Guayana, las hojas de los árboles y arbustos tienden a hacerse más pequeñas, reducidas, fuertes y gruesas, como una adaptación y resistencia a las mayores exposiciones a los vientos y al sol, combinados con temperaturas prevalentemente más bajas. A mayores altitudes los mismos árboles se hacen más bajos, con coronas más densas, así mismo el número de especies arbóreas se reduce. Otro fenómeno frecuentemente encontrado en bosques nublados es el endemismo, sea de flora y fauna. Un caso típico es la Serranía la Neblina en el Estado Amazonas, en esa área la gran mayoría de plantas encontradas no existen en otras partes del mundo, lo que se puede deber al efecto biogeográfico de isla.
En el trópico húmedo las causas principales de la deforestación y conversión del bosque nublado son: A) La explotación forestal sin planificación ni manejo silvicultural, lo que causa degradación y erosión. B)El avance de la " frontera agrícola " que convierte bosques naturales en áreas de uso agropecuario. C) Nuevas carreteras que atraviesan áreas boscosas, que provocan la colonización espontánea y no controlada a lo largo de su trayecto.
La deforestación de los bosques nublados en zonas montañosas pone en peligro su función protectora en términos hidrológicos y de conservación de los suelos, ya que se sustituye el bosque natural por pastizales para la ganadería extensiva. Lo que causa la compactación de los suelos, disminuye la capacidad de infiltración, provocando un aumento desmedido de la escorrentía superficial. Este cambio en el régimen hídrico en cuencas altas, en combinación con procesos de erosión acarrea un gran peligro de inundación para las cuencas bajas donde muchas veces se concentran poblaciones importantes cerca de los cauces de los ríos.
c. Bosques Parameros
Este tipo de bosque se presenta por encima de las selvas nubladas, y solamente en las partes altas del estado Mérida. Este bosque también es denominado bosque altoandino paramero y se extiende en una forma muy irregular y frecuentemente interrumpido en una faja altitudinal comprendida entre los 2.980 y los 4.200 msnm, donde la temperatura promedio varía entre 2-11,5° C, encontrándose hielo y temperaturas por debajo de 0° C durante todos los meses del año; la precipitación media anual varía entre 650-700 mm. Los árboles propios de este tipo de bosque alcanzan alturas de 3-8 m, siendo dominados por el Coloradito, Polylepis sericea, de la familia Rosaceae. Esta especie puede formar colonias puras o estar entremezclada con otras especies como, Vallea stipularis , Espeletia neriifolia, Aragoa lucidula, Hypericum laricifolium, Baccharis prunifolia, Chaetolepis lindeniana, Hesperomeles pernettyoides, Cestrum parviflorum y varias especies de Ericaceae.
Tales bosques de Polylepis, situados en medio de la vegetación de páramo, son considerados por algunos investigadores como representantes de colonias relícticas de condiciones climáticas previas, cuando el clima era más frío.
Ir al principio

B. Selvas Tropófilas
Estos bosques se conocen también como bosques deciduos, secos, caducifolios o veraneros, debido a la caída de las hojas durante el período de sequía. Se encuentran en regiones calientes, con precipitaciones anuales entre 1.000 y 2.500 mm y temperatura promedio de 25-28° C. Durante la estación seca, comprendida entre noviembre o diciembre hasta abril, la gran mayoría de los árboles pierden sus hojas, comenzando la floración y fructificación de muchas de sus especies; pero durante el período lluvioso, que comienza generalmente en abril o mayo, retoñan de nuevo y el bosque se torna completamente verde, mostrando su exuberante follaje y la escasez de flores en los árboles que lo pueblan. Los bosques de las selvas tropófilas cubren extensas superficies en las partes bajas de los llanos de los estados Apure, Barinas, Portuguesa, Guárico, Cojedes, Anzoátegui, Monagas, Yaracuy y Falcón. Ocupan también grandes extensiones de terreno en la parte norte de Guayana, al oeste del estado Amazonas y en grandes zonas del centro del estado Zulia y del norte del estado Trujillo. Se extienden desde el nivel del mar hasta 1.000 m. Algunos de los árboles comúnmente presentes pertenecen a géneros como Copaifera, Piptadenia, Swietenia macrophylla (caoba), Hura crepitans ( jabillo), Tabebuia rosea ( apamate) , Cederla odorata (cedro), Hymenaea, Virola, Anacardium excelsum(mijao), Protium, Eschweilera, Vitex, y con representantes abundantes de lauráceas y sapotáceas. Los árboles de mayor valor económico se encuentran en estas selvas, y pueden alcanzar alturas hasta de 40 metros, aunque generalmente alcanzan alturas de 20-30 metros. Se observa una gran diversidad de árboles, pareciéndose en este aspecto a la selva pluvial.
a. Bosques de Galería
Este tipo de bosque se desarrolla a lo largo de los ríos y varía en anchura y extensión, estando asociado con las sabanas llaneras. Algunos botánicos ubican este tipo de bosque dentro del concepto de selva tropófila, mientras que otros lo consideran dentro de la vegetación de los Llanos. Se ha generalizado la expresión bosque de galería para designar aquellas formaciones boscosas que corren a lo largo de los ríos llaneros e interrumpen la continuidad de la sabana. En los llanos orientales, por ejemplo, el bosque de galería está nítidamente delimitado por la sabana abierta de suelos arenosos. En cambio, en los llanos occidentales es casi imposible hablar de bosque de galería, debido a lo extenso de las formaciones boscosas allí presentes. En el bajo llano de los estados Apure, Barinas y de la parte sur de Guárico, es bastante pronunciada la formación del bosque de galería, diferenciándose claramente de los esteros y sitios inundables de esta parte del Llano. Los factores climáticos predominantes en estos bosques son los característicos de la región llanera: un clima biestacional típico, períodos de lluvia y sequía muy marcados, estación seca comprendida entre los meses de diciembre y abril, época lluviosa entre mayo y noviembre, y máximo de lluvias durante los meses julio a agosto. En general, la vegetación del bosque de galería se comporta como una típica vegetación tropófila, desprendiéndose del follaje durante la época seca y apareciendo con gran frondosidad durante los períodos de lluvia.
Ir al principio
C. Bosques Xerófilos

Se desarrollan en regiones donde la precipitación anual es inferior a los 1,000 mm y las altitudes van desde el nivel del mar hasta los 1,500 msnm, con variaciones de la temperatura entre 23-29° C, aunque en los Andes puede llegar a los 18° C. Se extienden por la zona norte y costanera del país, penetrando en el interior de los estados Falcón, Lara y Anzoátegui, Se distinguen dos tipos principales: los espinares y los cardonales. Se presentan en algunas zonas andinas.
a. Bosques semi-secos
Son los llamados espinares, chaparrales y cujizales, con árboles cuya altura varía entre 8 y 15 m, y que presentan un aspecto de árboles pequeños y vegetación esparcida. La precipitación anual es irregular, varía de menos de 800 y hasta 1,000 mm. Esta vegetación se encuentra distribuida en la partenorte y costanera del país, principalmente en el estado Falcón, penetrando al estado Lara; en la región occidental del lago de Maracaibo; alrededor de Cariaco, al sur de Barcelona, en el estado Anzoátegui; y en la península de Paria e islas cercanas a la costa. Entre las especies típicas están: cují yaque ( Prosopis juliflora ), dividivi (Caesalpinia coriaria ), espinillo (Parkinsonia aculeata) , cactáceas, como Mammillaria columbiensis y algunas cactáceas columnares. Durante la corta estación lluviosa aparece una cobertura herbácea formada por gramíneas y otras plantas que pertenecen principalmente a las familias leguminosas, malváceas, poligaláceas, convolvulácas, boragináceas, labiadas y esterculiáceas entre otras.
b. Bosques secos
Son los llamados cardonales ya que el principal constituyente de esta vegetación son los llamados cardones (Cereus, Ritterocereus, Subpilocereus, Pilosocereus) y tunas o nopales del género Opuntia y el guamacho (Pereskia guamacho). Están distribuidos en algunas zonas de los estados Lara y Falcón, en las penínsulas de Paria y La Guajira, en algunas zonas de la isla de Margarita y en algunos valles de las montañas andinas, alrededor del Chama, Motatán y Ureña. Reciben una precipitación anual inferior a los 600 mm, con una estación seca que dura de 7 a 10 meses, en algunos casos los suelos son salinos.
Ir al principio
D. Manglares
Constituyen bosques especiales en las zonas salinas costeras del norte y el oriente del país. Se distribuyen en las desembocaduras de los grandes ríos que surten el Golfo Triste, el golfo de Cariaco, las lagunas de Píritu, Unare, Tacarigua y Carenero, la costa de los estados Carabobo y Falcón hasta Chichiriviche, alrededor del lago de Maracaibo y en el delta del Orinoco. Los manglares generalmente aparecen en regiones de abundante precipitación anual, pero también se encuentran en áreas de gran aridez. Las especies de estos bosques retienen sus hojas durante todo el año y son denominadas siempre verdes. Los suelos contienen altas concentraciones salinas por estar mezclados con agua marina. Los árboles forman colonias impenetrables, densas, con raíces aéreas descendentes hasta el agua, que ayudan a la

acumulación y formación de suelo. Las especies constituyentes de la formación manglar pueden alcanzar alturas de hasta 25 m, pero generalmente son más pequeñas. Las plantas de los manglares presentan viviparía, criptoviviparía o por no ser vivíparas; se han reportado embriones verdes, posiblemente fotosintéticos y las plantas adultas desarrollan neumatóforos, que son raíces epigeas con un aerénquima bien desarrollado, cuya función es suministrar oxígeno a los órganossubterráneos carentes de él, por las condiciones edáficas en que se desarrollan estas plantas. El medio donde prosperan los mangles tiene una alta concentración salina de aproximadamente 25 atm. de presión osmótica (potencial osmótico de - 2,5 MPa); registrándose en el jugo vacuolar de algunas hojas de mangle presiones osmóticas de 35 a 55 atm. ( potencial osmótico de -3,5 a -5,5 MPa). Es común encontrar las siguientes especies: Rhizophora mangle (Rhizophoraceae) , Avicennia nítida (Verbenaceae), Laguncularia racemosa (Combretaceae). Se desarrollan desde la desembocadura de los ríos hasta mar adentro. Algunas especies presentan glándulas de secreción salina, como Avicennia y Laguncularia, otras como Rhizophora no las presentan, por lo que se puede pensar en un mecanismo de exclusión de sales más efectivo a nivel radical.
Ir al principio
II. Áreas Terrestres no Boscosas
A. Páramos
Son formaciones características de las montañas andinas, desde el estado Táchira hasta los límites con el estado Lara. Se pueden encontrar a 1.700 msnm en el páramo del Morro, en el estado Mérida(donde se encuentra el Alnus acuminata), pero son más bien propias de altitudes comprendidas entre los 3.000 y los 4.700 m. Áreas con vegetación parecida a la de los páramos se encuentran en las regiones más altas de la cordillera de la Costa, en el cerro Turumiquire del estado Sucre y en partes del Distrito Federal, como el cerro Naiguatá en el parque nacional El Ávila. La temperatura media anual de los páramos es de 5° C o menos, y la formación de hielo, con temperaturas por debajo de 0° C, ocurre durante todo el año. La precipitación anual es baja, con un promedio de 600 mm por año. Los páramos existen bajo condiciones ambientales de intenso frío y poca precipitación, exhibiendo un aspecto xerofítico o desértico; y suelen presentar un color acentuadamente grisáceo, debido a la abundancia de frailejones del género Espeletia (Espeletia Schultzii de flores amarillas y Espeletia neriifolia, que es un arbolito de hojas perennes) , cuyas hojas densamente pilosas y sedosas tienen este color grisáceo o de ceniza, aunque también hay una gran variedad de especies con flores de colores llamativos, de la familia Melastomataceae. Para resistir el frío y la sequedad, las hojas de las plantas parameras son pequeñas y reducidas o densamente cubiertas con un tomento sedoso semejante a la lana.
Ir al principio

B. Sabanas de las tierras altas de Guayana
Son denominadas por algunos autores Sabanas Altas o de Lomas, o Sabanas higrófilas Mesotérmicas. Estas sabanas se presentan en las laderas de arenisca y las cumbres de las formaciones geológicas características de los tepuyes, en los estados Bolívar y Amazonas. Presentan un paisaje sin árboles que se asemeja a una sabana verdadera. Se encuentran en alturas que oscilan entre 1200 a 2800 metros, donde la precipitación promedio anual puede exceder los 3800 mm. Este tipo de paisaje se desarrolla bajo las condiciones combinadas de la formación Roraima, compuesta de areniscas y cristalinas, con suelos altamente ácidos y limosos, temperaturas fías que varían de 4,5 a 240 C. La precipitación abundante distribuida durante todo el año, con un período de sequía muy corto y una intensa exposición a los vientos y a la luz solar. La vegetación es características de estas sabanas, la que es muy diferente a las sabanas llaneras y muestra una gran cantidad de endemismo. Algunas de las familias representadas con especies en estas sabanas son: Rapatacea, Cyperaceae, Sarraceniaceae, Bromeliaceae, Xyridacea, Eriocaulacea, Ochnaceae, Lentibulariaceae, Melastomataceae, Orchidaceae, Ericaceae, Compositae, Bignoniaceae, Theaceae, Rubiaceae, etc.
Ir al principio
C. Sabanas
Están representadas por tierras cubiertas de gramíneas, donde pueden aparecer plantas arbóreas en forma esporádica. La mayor parte de las sabanas ocupan la región llanera, que comprende los estados Apure, Barinas, Portuguesa, Cojedes, Guárico, Anzoátegui y Monagas. También se pueden encontrar sabanas en los estados Bolívar, Sucre, Zulia, Aragua y Miranda y en el Distrito Federal. Las tres cuartas partes de los llanos venezolanos están representadas por vegetación de sabana; la cuarta parte restante está ocupada, en cambio, por bosques tropófilos, de galería, morichales, matas y palmares.El clima de la región llanera se presenta con una temporada de sequía entre noviembre y abril, y un período de lluvia entre mayo y septiembre; con una precipitación anual de 1.000-2.000 mm. La temperatura media anual está entre 26 y 28° C. Los vientos predominantes son los alisios y la altitud varía entre 50 y 300 msnm. Los factores edáficos son muy importantes en la determinación de las sabanas; asimismo el fuego ha sido relacionado con el mantenimiento de la vegetación de sabana. En los Llanos de acuerdo a su composición florística se distinguen los siguientes tipos : sabanas de Trachypogon, sabanas de Paspalum fasciculatum ( chiguirera) y sabanas de banco, bajíos y esteros, morichales, palmares.
1. Sabanas de Trachypogon
Cubren una amplia extensión de la región llanera y se encuentran en los llanos orientales y el Apure meridional. Están dominadas por tres especies de gramíneas; Trachypogon montufari (paja velluda), Trachypogon plumosus (saeta) y Trachypogon vestitus (saeta), aunque pueden estar asociadas a otras especies de gramíneas, algunas cyperaceas y a gran variedad de hierbas y sufrútices donde predominan las leguminosas. Los suelos contienen un alto porcentaje de arena; son pobres en nutrientes y de pH ácido. El terreno tiene buen drenaje, de manera que durante los meses lluviosos no se cubren de agua. La importancia forrajera de los Trachypogon se debe a su gran abundancia, aunque su valor nutritivo es bajo en su estado de madurez.Los chaparrales se encuentran en algunas zonas de Anzoátegui, Guárico y Cojedes, y en el piedemonte de la cordillera de los Andes, representados por Curatella americana (chaparro) , Byrsonima crassifolia (manteco)y Bowdichia virgilioides (alcornoque), etc.Las sabanas de Trachypogon inarboladas se encuentran al sur de los llanos de Monagas, al sur-este de Anzoátegui y al sur de Apure. Son sabanas de gran amplitud, con ríos separados por distancias considerables y de suelos arenosos profundos, donde pueden aparecer especies arbóreas de los chaparrales aun cuando permanecen pequeñas. La zona entre los ríos Guárico y Manapire representa la transición entre los llanos occidentales y los orientales, caracterizados por sabanas de bancos, bajíos y esteros, y sabanas de Trachypogon, respectivamente.
2. Sabanas de Paspalum fasciculatum

También conocidas como sabanas de gamelote apureño o gamelote chiguirero. Conservan su verdor durante los meses secos, de ahí que los gamelotes sean pastizales de sequía, quemados a principios de esa temporada para provocar retoños frescos. Estas sabanas son el principal recurso forrajero durante la estación seca en el llano. Predomina en el Bajo Apure, en el Distrito Arismendi de Barinas, al Sur de Cojedes y al suroeste de Guárico. Estas sabanas están atravesadas por muchos ríos, caños y cañaotes, que desbordan durante los meses lluviosos y aniegan estas sabanas formando un tipo de estero. El terreno es plano, el suelo es aluvial y rico en limo.
3.Sabanas de Bancos, Bajíos y Esteros
Se encuentran distribuidas en el alto Apure, Barinas, Guárico, Portuguesa y Cojedes. Presentan una topografía de altos y bajos que produce diferenciaciones fisonómicas, florísticas y edáficas propias, además de diferentes niveles de inundación durante el período de lluvias. Los bancos son sitios altos que no se inundan durante los meses lluviosos; su vegetación de gramíneas tiene un porte de mediano a bajo y está constituida por hierbas apetecibles por el ganado. Los esteros son sitios de estas sabanas que se inundan de tal manera, que se pueden navegar con canoas o curiaras. Tienen suelos de textura pesada. El estero cambia completamente su fisionomía durante el año: en sequía presenta un terreno seco y una vegetación graminosa baja, y en los meses lluviosos se transforma en una laguna con abundancia de plantas acuáticas.Los esteros se anegan como consecuencia del desbordamiento de los ríos, mientras que los bajíos se forman por el agua de lluvias. En los bajíos y esteros crecen varias plantas espinosas, las cuales muchas veces forman colonias que desvalorizan el pastizal. En esteros anegados aparecen varias especies de Eichornia, de Ludwigia y Neptunia postrata, entre otras plantas hidrófilas.
4. Morichales
Corresponden a sabanas de suelos inundables, dominadas fisonómicamente por la palma Moriche, Mauritia flexuosa. En los Llanos, frecuentemente, se encuentran morichales distribuidos al norte del Orinoco, en los estados Apure, Guárico, Anzoátegui y Monagas, desarrollándose a lo largo de los manantiales. También se desarrollan morichales al sur del Orinoco, en los estados Bolívar y Amazonas. Los suelos de los morichales son siempre húmedos y poseen una flora característica asociada con ellos y no localizable en otros lugares de la sabana.
5. Palmares
Los palmares son sabanas de suelos inundables donde domina la palma llanera, Copernicia tectorum. Se distribuyen en Venezuela en grandes áreas de los Llanos, en el estado Guárico y en algunas regiones de los estados Bolívar, Anzoátegui, Sucre y Apure. Los suelos son pesados, arcillosos, y se mantienen inundados durante la época de lluvia.
Ir al principio
D. Dunas
Corresponden a ambientes muy áridos, distribuidos en Venezuela en el istmo de Paraguaná, en las cercanías de Coro y La Guajira, y en algunas partes de los Llanos; tienen una temperatura media anual mayor de 25° C, y un bajo promedio anual de precipitación. Presentan una vegetación muy rala y están cubiertas temporalmente por arenas movibles por acción del viento. Desde el punto de vista florístico, las dunas se caracterizan por la presencia de la especie Tournefortia gnaphalodes, Tabaco pescador, de la familia Boraginaceae, un arbusto decumbente, de hojas suculentas con pubescencia grisácea, y que forma colonias. El árbol más común en la dunas es el cují yaque( Prosopis juliflora) ,

de la familia Leguminosae. Las gramíneas rabo de zorro Aristida venezuelae, y saladillo ( Sporobolus virginicus) , forman el estrato de mayor cobertura herbácea.
Ir al principio
III. Áreas Acuáticas
a. Plantas acuáticas de agua dulce
Incluyen toda la vegetación desarrollada en, lagos, lagunas, ríos, estuarios, esteros, raudales y saltos. En algunos lagos y ciénagas de los paramos andinos se encuentran los helechos acuáticos Isoetes lechleri y Isoetes triquetra. Dentro de las Angiospermas se reporta, Elatine de la familia Elatinaceae.
En los lagos y lagunas de los estados Bolívar, Guárico y Apure encontramos especies pertenecientes a los géneros Jussiaea, Canna, Mayaca, Neptunia, Eleocharis, Scirpus y Tonina.
Como representantes comunes en el Lago de Valencia se reportan especies de los géneros ValIisneria, Potamogeton, Sagittaria, Najas, Ruppia, Cyperus y Typha.
b. Plantas Marinas
Están formadas fundamentalmente por algas marinas, principalmente feofitas, clorofitas y rodófitas. Estas plantas forman densas colonias que se mueven de manera incesante siguiendo las corrientes del mar. También aparecen fanerógamas bentónicas como: Ruppia marítima, Thalassia testudinum y Halophila baillonis, que forman praderas. Sobre la flora bentónica viven gran variedad de organismos epifitos vegetales y animales.
Ir al principio
Bibliografías:
1. Steyermark,J. A. 1966. Nuestra Vegetación. El Farol. 216:6-15.
2. Lindorf, H. ,L. Parisca y P. Rodríguez. 1985. Botánica. Ediciones de la Universidad Central de Venezuela, Caracas. 584p.
3. Stadtmuller, T. 1987. Los bosque nublados en el trópico húmedo. Centro Agronómico Tropical de Investigación y Enseñanza. Turrialba, Costa Rica. 85 p.
4. Walter, H. 1973. Vegetation of the earth. Springer Verlag, New York. 237p.

http://www.gobiernoenlinea.gob.ve/venezuela/perfil_geografia7.html
http://www.pdv.com/lexico/venezuela/flora.htm
http://www.mipunto.com/venezuelavirtual/mapas/mapa_vegetacion.html
CONTENIDOS
/ BASES CONCEPTUALES DE LA ECOFISIOLOGÍA VEGETAL / RESPUESTAS DE LAS PLANTAS A LOS FACTORES AMBIENTALES / ATMÓSFERA / HIDRÓSFERA / RADIACIÓN Y CLIMA / LA LITOSFERA Y EL SUELO /
PÁGINA PRINCIPAL
BASES CONCEPTUALES DE LA ECOFIOLOGÍA VEGETAL
El famoso naturalista Inglés Charles Darwin propuso la teoría de la evolución (1858), que tiene dos componentes principales :1. Las especies no son inmutables , sino que cambian o se adaptan a lo largo del tiempo. 2. El agente que produce los cambios es la selección natural. Los individuos están comprometidos en una lucha permanente por la existencia, en la que los más aptos sobreviven. Esa lucha puede ser de dos tipos. 1. La obtención de los recursos necesarios para establecerse y crecer en un medio ambiente algunas veces pobre y hostil. 2. La lucha con competidores vecinos de la misma o de especies diferentes. La ecología o la sociología vegetal tratan de las relaciones o interacciones de las especies dentro de las comunidades, o la forma en que poblaciones de una especie se adaptan a diferentes medios ambientes. Mientras que la fisiología vegetal se relaciona con el estudio de las funciones de los individuos, en diferentes niveles de organización y su lucha con el medio ambiente, auxiliándose de varias disciplinas como la bioquímica, biofísica y la biología molecular. La fisiología vegetal es una disciplina que se refugia en el laboratorio para su estudio, manipula las condiciones de crecimiento del individuo y mide la respuesta de un determinado proceso.
La ecofisiología, estudia los fenómenos fisiológicos fuera del laboratorio, en su medio ambiente natural, el cual está sujeto a cambios y alteraciones, como resultado de fenómenos naturales o producto de la actividad humana. En este sentido, podemos mencionar por ejemplo el aumento significativo de la concentración atmosférica de CO2 por efecto de la actividad industrial, la quema de combustibles fósiles y el calentamiento global de la atmósfera. Este factor es difícilmente controlable, como se podría hacer al estudiar las variaciones en la concentración de CO2 sobre la fotosíntesis, lo cual es estudiado por la fisiología vegetal, bajo condiciones de laboratorio. Las plantas en su medio ambiente natural responden a las variaciones de temperatura, intensidad de luz, humedad, concentraciones de CO2 , lo cual depende de sus adaptaciones fisiológicas.
Los ecofisiólogos en un principio estudiaron las respuestas fisiológicas de las plantas en un medio ambiente abiótico, como por ejemplo suelos calcáreos y ácidos o en suelos secos y anegados, interacciones fisiológicas con otras plantas, animales y el beneficio de microorganismos. Mientras que, un ecofisiólogo moderno, requiere una buena comprensión tanto de los procesos moleculares que ocurren a nivel celular, como del funcionamiento de la planta intacta en un contexto ambiental.
Un aspecto relevante en el desarrollo de la ecofisiología se obtuvo de la importancia de la fisiología para la agricultura. Incluso hoy en día, la productividad agrícola en países industrializados está limitada al 25% de su potencial, por condiciones de sequía, suelos infértiles y otros factores ambientales indeseables. Uno de los objetivos principales de las investigaciones agrícolas, ha sido el desarrollar cultivos tolerantes a estados ambientales estresantes, de tal manera que soporten condiciones climáticas adversas o que se puedan cultivar en hábitat desfavorables. Así se han desarrollado variedades de plantas tolerantes a la salinidad o que soporten el estrés hídrico.

La ecofisiología vegetal estudia las respuestas fisiológicas frente a diferentes condiciones ambientales, desarrollando técnicas que permiten medir el micro medio ambiente de las plantas, las relaciones hídricas y los patrones de intercambio gaseoso. En sus inicios, se estudió el crecimiento de las plantas determinando las variaciones en biomasa, sin embargo la invención de equipos portátiles ha posibilitado medir los intercambios de CO2 en una hoja y la conductividad estomática como un índice de la apertura y cierre de los estomas. Mediante los análisis de crecimiento, se ha podido determinar la cantidad de carbono que se deposita en raíces y hojas, así como la tasa de producción y muerte de ciertos tejidos; lo que permite una mejor comprensión de las diferencias en el crecimiento vegetal, en distintas condiciones ambientales. Finalmente, concluimos que la ecofisiología es un componente importante del entrenamiento de un ecólogo vegetal.
Ir al principio
RESPUESTAS DE LAS PLANTAS A LOS FACTORES AMBIENTALES
El ambiente de una planta está formado por la hidrosfera, la atmósfera y la pedósfera, pero además lo integran factores físicos y químicos en su hábitat, y la influencia ejercida por otros organismos. Resumiendo, el ambiente de cada planta es la totalidad de las condiciones externas que actúan sobre un individuo o comunidad de organismos (biocenosis) en un territorio definido (biotopo).
Se entiende por clima el conjunto de los fenómenos meteorológicos que caracterizan el estado medio de la atmósfera y las variaciones regulares de los estados del tiempo. Los distintos climas del mundo vienen dados por las temperaturas y precipitaciones. Las diferencias climáticas se expresan en forma de diagramas climáticos o climadiagramas. En estos diagramas se grafican las variaciones de temperatura en grados centígrados y las precipitaciones en milímetros a lo largo del año ( 1 mm de precipitación equivale a un litro por m2 ).En el eje de las ordenadas se representa la precipitación y la temperatura, la escala de precipitación es el doble de la escala de temperatura ( una división equivale a 100o C o 20 mm de lluvia. Panaquire [3], significa que los datos son el promedio de 3 años de mediciones, tanto de precipitaciones como de temperaturas. El valor 26,3o es el promedio de temperatura anual en grados centígrados y 2719 el promedio de precipitación anual en milímetros. En el eje de las abcísas se representan los meses del año.
CLIMADIAGRAMA
Ir al principio

ATMÓSFERA
La atmósfera es una capa de aire delgada que rodea la tierra. Alrededor de 80% de la capa inferior está en la tropósfera, que se extiende desde la superficie de la tierra hacia arriba unos 17 Km, en los trópicos y subtrópicos, pero solo 10 Km en latitudes más altas . La mayor parte de la circulación global ocurre en la tropósfera, y allí se localiza casi todo el vapor de agua de la atmósfera. En esa capa, la presión atmosférica disminuye hacia arriba, así como la temperatura, que alcanza un mínimo en la tropopausa. La estratósfera, que se extiende hacia arriba desde el límite de la tropósfera hasta unos 50 Km por encima de la superficie terrestre, contiene cerca del 99% de la masa remanente de la atmósfera , pero es extremadamente seca. El ozono ( O3 ) de la estratosfera absorbe la mayor parte de la radiación ultravioleta de onda corta que proviene del sol, generando un escudo protector que permite la vida en la tierra.
Actualmente, la atmósfera está compuesta por 78 % de nitrógeno (N2 ), 21% de oxígeno, 0,95% de gases raros y 0,035% de dióxido de carbono. También contiene trazas de hidrógeno, vapor de agua, metano, dióxido de azufre, haluros, compuestos nitrogenados volátiles, aerosoles, ozono, metano polvo y ceniza. La atmósfera alberga el mayor reservorio de nitrógeno de la Tierra y grandes cantidades de oxígeno. El dióxido de carbono es la fuente principal de carbono utilizada por los organismos autótrofos mediante la fotosíntesis, que produce azucares a partir de CO2, agua y energía solar; así mismo se libera oxígeno a la atmósfera. La respiración de los organismos terrestres y la combustión consume oxígeno, que es reemplazado por la actividad del fitoplancton y de otros organismos fotosintéticos.
Ir al principio
HIDRÓSFERA
El agua es un componente esencial de la biosfera y es la base de todos los procesos vitales. La economía hídrica del planeta se alimenta principalmente del mar constituyendo más del 97%, un 2% se encuentra en forma de hielo y nieve, 0,61% se encuentra en las aguas continentales, como agua freática, pero sólo 1 % está disponible para las raíces de las plantas. En la atmósfera se halla el 0,001 %, en forma de nubes, niebla y vapor. La cobertura vegetal de la tierra firme está formada por plantas superiores que absorben la precipitación que se fija al suelo por las raíces y la retorna a la atmósfera por la transpiración de las hojas.
ECONOMÍA HÍDRICA DEL PLANETAAGUA TOTAL EN LA BIOSFERA (%)
Lagos de agua dulce 0,009Lagos salados y mares continentales 0,008
Ríos 0,0001Humedad del suelo y escorrentía 0,005
Humedad subterránea hasta 4000 m 0,61Casquetes polares y glaciares 2,41
Atmósfera 0,001Océanos 97,2279
En la figura que se observa a continuación se muestra un ejemplo del ciclo del agua. El ciclo hidrológico está determinado principalmente por la evaporación del agua desde la superficie de los océanos. Parte del agua regresa a los océanos en forma de precipitaciones, pero esta cantidad es menor que la que se escapa de estos por evaporación. El resto del agua es arrastrada por los vientos hacia la tierra donde se condensa, formando las nubes y luego cae en forma de lluvia o nieve. El agua se evapora también de los suelos, lagos, ríos y desde la superficie de las hojas, pero la cantidad que se evapora es menor que la que se precipita sobre la tierra. El agua que cae en exceso regresa de nuevo a los océanos por los ríos, la percolación y las corrientes subterráneas.

En muchas regiones pobres en precipitaciones, pero con niebla abundante ocurre el fenómeno de intercepción horizontal, que se caracteriza por la condensación de gotas de agua sobre las plantas, que pueden caer al suelo.
CICLO HIDROLÓGICO
Ir al principio
RADIACIÓN Y CLIMA
La causa principal del clima es la energía radiante del sol recibida por la tierra. La radiación está compuesta de ondas electromagnéticas que viajan a la velocidad de la luz( 300.000 Km/seg ).
La radiación solar es producida en el centro del sol mediante fusión nuclear, en la que se producen elementos pesados a partir de elementos livianos, liberándose energía según la ecuación de Einstein E=mxC2, donde m es la masa y C es la velocidad de la luz . La radiación solar es usada en fenómenos biológicos. En el límite externo de la atmósfera la energía del sol llega a una tasa constante de 1,39 KW/m2 o 2,0 cal x cm-2 x min-1, cuando el sol se encuentra en el cenit, este valor se denomina constante solar. Esta constante es la cantidad de energía radiante de todas las longitudes de onda que atraviesa una unidad de superficie por unidad de tiempo. La radiación tiene propiedades de onda y de partículas, se compone de una serie continua de longitudes ondas que van de < 10-14 m (rayos cósmicos) hasta las ondas largas de radio > 106 m. Este rango de longitudes de ondas se denomina espectro electromagnético.

ESPECTRO ELECTROMAGNÉTICO
El espectro electromagnético. Imagen tomada de Purves et al. Life: The Science of Biology, 4th Edition, by Sinauer Associates (http://www.sinauer.com ad WH Freeman http://www.whfreeman.com).
El espectro visible es muy estrecho ocupa de 3,9 x 10-7 m a 7,7 x 10-7 m o 390 nm a 770 nm (3900 a 7700 Å). Ni la luz ultravioleta(UV) de onda corta, ni la infrarroja de onda larga son visibles; sin embargo algunas personas a las que se les ha removido el lente(cristalino) de los ojos, debido a cataratas pueden ver la luz UV por debajo de 390 nm. Las regiones de longitud de onda de mayor interés en biología son: la ultravioleta, la visible y la infrarroja.
Las longitudes de onda por debajo de 400 nm se conocen como ultravioleta. El rango de longitudes de onda de 320 a 390 nm, se llama UV-A, el rango de 280 a 320 nm como UV-B y el de 200 a 280 nm es la UV-C. La radiación que llega a la tierra contiene cerca de 7 % de UV ( < 400 nm ) en relación a su energía. La capa de ozono absorbe intensamente la radiación de longitud de onda entre 200 y 330 nm. La zona infrarroja ( IR ) tiene longitudes de onda mayores que la roja y se extiende aproximadamente hasta 100µm. La longitud de onda, la velocidad de la luz y la frecuencia, se hallan relacionadas por la siguiente ecuación: λ.V=C; de donde λ es la longitud de onda; V es la frecuencia y C es la velocidad de la luz ( 300.000 Km x S -1 ). La longitud de onda se puede expresar en número de ondas (V), que es el reciproco de la longitud de onda en m -1. El número de ondas se utiliza para los cálculos, ya que es directamente proporcional a la energía de la onda. A medida que aumenta el número de onda la energía asociada con la onda también aumenta. La energía de un fotón es el producto de la frecuencia ( V ) por la constante de Planck ( h )que tiene un valor de 6,6262x10-34JxS.E(fotón)=h.V y donde V= C/λ . La velocidad de la luz C=3,0x 108 m . S-1 ; λ = longitud de onda en metros (m); V= frecuencia en segundos( S-1). E(fotón)= h . C /λ . Calculemos el número de onda ( V ) de la radiación con longitud de onda de 700 nm, 7x 10-5 cm o 7 x 10-7 m. V= 1/λ= 1/7 x 10-5 cm o sea 14286 cm-1 o 1.428.600 m-1 .
Definición y características de varias regiones de longitud de onda de la luz
ColorRango de
longitud de onda(nm)
Longitud de onda representativa
Frecuencia (ciclos/S) o
hertziosEnergía (KJ/mol)
Ultravioleta < 400 254 11.8 x 1014 471Violeta 400-425 410 7.31x 1014 292
Azul 425-490 460 6.52x 1014 260Verde 490-560 520 5.77x 1014 230
Amarillo 560-585 570 5.26x 1014 210Anaranjado 585-640 620 4.84x 1014 193

Rojo 640-740 680 4.41x 1014 176Infrarrojo >740 1400 2.14x 1014 85
Seleccionemos una longitud de onda en la zona azul del espectro con λ= 460 nm.V= C /λ entonces tenemos V= 3.0 x 108m . S-1 / 460 x 10-9 m x ciclo-1, que es igual a 6.52x1014ciclosxS-1.La energía de un fotón se calcula mediante la ecuación siguiente: E(fotón) = h . C /λ.
E(fotón) = 1.9865 x 10-16 /λ ( en nm ) el resultado en Jul o Julios.La energía del fotón se puede expresar también en electrón voltio (eV). Sabiendo que 1 eV tiene una energía de 1.6022 x 10-19 J, la energía del fotón expresada en eV, está dada por la ecuación: E(fotón)=1,2399x103/λ (en nm) el resultado en eV.
De acuerdo a la ley de Einstein, de la equivalencia fotoquímica, una molécula puede reaccionar en una reacción fotoquímica, solamente después de haber absorbido un fotón. De tal forma que, un mol puede reaccionar solamente después de absorber 6,0222 x 1023 fotones ( 6,0222 x 1023 es el número de Avogadro, N, que es el número de moléculas en un mol de cualquier sustancia). Ahora bien, la energía de 6,0222 x 1023 fotones es igual a 6,0222 x 1023 x h. V.
E = N x h x V, de donde E = N x h x C /λ , E = 6,0222 x 1023 h x C /λ , esta cantidad de energía se denomina un Einstein.
Un Einstein, se puede definir como la cantidad de energía radiante de longitud de onda apropiada, que debe ser absorbida por un mol de una sustancia antes que pueda reaccionar en una reacción fotoquímica. La energía de un Einstein se calcula de la siguiente manera:
E(Einstein) = 1,1963 x 108 /λ ( en nm ).
La energía de un fotón varía directamente con la frecuencia( V ) e inversamente con la longitud de onda ( λ ). Así tenemos que, a mayor frecuencia y menor longitud de onda mayor será la energía.
La función de los sistemas de pigmentos que se encuentran en los cloroplastos es la de absorber energía luminosa.
¿Calcular para una longitud de onda de 650 nm, el número de onda ( V ) en cm-1 , la frecuencia (V) en Tera Hertzios(THz) , energía del fotón en attoJul (aJ), eV, y la energía de un Einstein en KJ?
λ= 650 nm ; λ = 6,5 x 10-7 m .
a) V = 1 /λ = 1 / 6,5 x 10-7 m = 15385 x 102 m-1 o 15385 cm-1. b) V = C /λ = 3,0 x 108m x S-1 / 6,5 x 10-7 m o sea 461,2 x 1012 S-1.
Un Tera Hertzio es equivalente a 1012 Hz o 1012 ciclos/segundo, de donde V= 461,2 THz.
c) La energía del fotón en atto Jul ( a J ). Un atto Jul es 10-18 J. E(fotón) = 1,9865 x 10-16 / 650, es igual a 0,3056 x 10-18 J E(fotón) = 0,3056 aJ ( atto Joule).
d) E(fotón) = 1,2399 x 103 / λ (en nm) resultado en eV = 1,2399 x 103 / 650
E(fotón) = 1,9075 eV.
Un electrón voltio tiene una energía de 1,6022 x 10-19 J.
e) E(Einstein) = 1,1963 x 108 / λ (en nm)J = 1,1963 x 108 / 650, esto es igual a 184.046J.
E(Einstein) = 184,0 KJ.
Si sabemos que una caloría es equivalente a 4,184 Jul, entonces tenemos que 184,046 x 103 / 4,184 = 43,98 x 103 calorías o 43,98 Kcal x mol-1.

Toda la vida sobre la tierra es mantenida por el flujo de energía radiante proveniente del sol, que penetra la biosfera. Mediante la fotosíntesis esta energía se fija en forma de energía química latente, que se moviliza hacia todos los componentes de una cadena alimenticia. Para las plantas la radiación no es solamente una fuente de energía (efecto foto energético), es también un estimulo que dirige el desarrollo ( efecto foto cibernético), y ocasionalmente puede actuar como un factor estresante(efecto foto destructivo). Cada uno de estos procesos es activado por la absorción de cuantos de luz, cuya respuesta es mediada por foto receptores, con un espectro de absorción que corresponde a un efecto foto biológico. En este tipo de respuesta son importantes el tiempo de duración del estimulo, la dirección de la radiación incidente, y la composición espectral de la radiación absorbida.
La energía radiante que alcanza diferentes sitios de la superficie terrestre, depende de su orientación con respecto al sol como resultado de los movimientos de rotación y traslación del globo terráqueo. La cantidad de radiación que entra a la tierra fluctúa periódicamente, lo que impone una ritmicidad climática sobre todos los fenómenos terrestres. La alternancia periódica de las noches y los días, es estimulo o disparador responsable que regula los ritmos diurnos (circadianos) y estacionales. Así mismo, la radiación solar al actuar como una señal, controla muchos procesos de desarrollo y diferenciación, tales como la germinación, fototropismo, fototaxis (ciclosis de los cloroplastos), fotomorfogénesis, síntesis de antocianinas, visión, fotosíntesis y movimientos de orientación de los cloroplastos.
Ir al principio
LA LITOSFERA Y EL SUELO
La litosfera es la capa externa de la Tierra y está formada por materiales sólidos, engloba la corteza continental, tiene aproximadamente 80 Km. de espesor, y la corteza oceánica o parte superficial del manto consolidado, de unos 10 Km. de espesor. Se presenta dividida en placas tectónicas que se desplazan lentamente sobre la astenosfera, capa de material fluido sobre la cual se encuentra la litosfera. Las tierras emergidas son las que se hallan situadas sobre el nivel del mar y ocupan el 29% de la superficie del planeta. Su distribución es muy irregular, concentrándose principalmente en el Hemisferio Norte o continental, dominando los océanos en el Hemisferio Sur o marítimo. La atmósfera tiene unos 273 Km. de espesor, mientras que el suelo forma una capa fina entre atmósfera y litosfera. Los suelos son muy importantes para el crecimiento de las plantas y muestran interacciones complejas. Las plantas obtienen el agua y los nutrientes minerales de la solución del suelo o del agua en la que crecen, al igual que el oxígeno para la respiración de las raíces. La aireación varía de acuerdo a las condiciones del suelo, por ejemplo un suelo encharcado o saturado de agua tiene poca aireación. El suelo proporciona sostén mecánico para las plantas terrestres y alberga microorganismos del ciclo del nitrógeno o que realizan la descomposición de la materia orgánica, liberando los elementos químicos que requiere la planta para crecer. Así mismo, en el suelo se encuentran hongos que forman asociaciones con las raíces, micorrizas y organismos patógenos que le causan daños a las plantas.
Resumiendo podemos decir que el suelo le proporciona a la planta: sostén o anclaje, agua, nutrientes y oxígeno para las raíces.
El suelo es un sistema de tres fases: sólido, líquido y gaseoso, y cuatro componentes mineral, orgánico, agua y aire. Un buen suelo para el crecimiento de un cultivo, tendrá 45 % en volumen de materia mineral, 5 % de materia orgánica y 50% de espacio poroso dividido aproximadamente en 25 % de agua y 25% de aire.

COMPONENTES INORGÁNICOS Y ORGÁNICOS DEL SUELO
Purves, et al.Life.The Science of Biology. 7th e
La proporción real de aire y agua varía según las condiciones. Cuando llueve los poros se llenan de agua, que se vacían de nuevo cuando el suelo se seca. En los países tropicales con temperaturas altas, los porcentajes de materia orgánica tienden a ser menores (de 2,5 a 3 %) debido a su rápida descomposición.
Cuando los edafólogos realizan un estudio de suelo, generalmente cavan un pozo en la tierra (calicata), y luego observan sus paredes. Las paredes del pozo revelan un patrón estratificado, formado por capas, cada una de las cuales se denomina horizonte, y al conjunto de capas presentes en las paredes del pozo, perfil del suelo. A pesar de que los suelos difieren mucho, casi todos consisten de dos o más capas horizontales, ubicadas una sobre otras. En el perfil de un suelo típico se reconocen tres horizontes principales: A, B y C. El horizonte superior o horizonte A, posee un mayor contenido de materia orgánica y tiene un color mas oscuro que las capas inferiores. En este horizonte se desarrollan las raíces de las plantas y en él se encuentran presentes lombrices, insectos, nematodos y microorganismos. Por debajo del horizonte A, se encuentra el B o subsuelo, que es la zona de infiltración y acumulación de los minerales lixiviados desde arriba, su color es más claro que el horizonte A y contiene más arcilla. Más abajo está el horizonte C que es la roca madre original de la cual se deriva el suelo.

PERFIL DE UN SUELO TÍPICO MOSTRANDO LOS HORIZONTES A, B Y C.
Purves, et al.Life.The Science of Biology. 7th e
Los componentes inorgánicos de los suelos comprenden pequeños fragmentos de roca y de minerales de varias clases. El tamaño de las partículas varía en tamaño desde grandes cantos rodados hasta partículas pequeñas de arcilla que tienen menos de 2µm de diámetro. Las cuatro clases de partículas inorgánicas son: arena gruesa, arena fina, limo y arcilla.
CLASIFICACIÓN DE LAS PARTÍCULAS DEL SUELO (ISSS)
Tipo de suelo Dimensión Visible usando Tamaño de la partícula (mm)
Arena gruesa Muy gruesas A simple vista 0,2 - 2,0Arena fina Gruesas A simple vista 0,02- 0,2
Limo Finas Al microscopio 0,002 - 0,02
Arcilla Muy finas Al microscopio electrónico < 0,002
Los suelos se forman por desintegración de la roca madre, en parte por la erosión mecánica, que es provocada por cambios de temperatura, la lluvia, el hielo, el viento, el crecimiento de plantas como musgos líquenes y las raíces de las plantas. Sin embargo, los procesos químicos descomponen la roca madre por hidrólisis, hidratación, carbonatación y otros procesos acidificantes. El proceso clave es la formación de arcilla, como resultado de la erosión química.
Los suelos constituyen la fuente de nutrición de las plantas terrestres, la cual depende de la presencia de partículas de arcilla . Los cationes potasio (K+), magnesio (Mg2+) y calcio (Ca2+) están cargados positivamente y son atraídos por las cargas negativas de las partículas de arcilla. Para que los cationes puedan ser absorbidos por las raíces de las plantas, debe ocurrir un intercambio catiónico, lo que se logra con protones (H+) presentes en la solución del suelo producto de la ionización del ácido carbónico y del bombeado por las raíces. . Los protones se unen entonces a las partículas de arcilla y se liberan los cationes (K+), (Mg2+) y (Ca2+) a la solución del suelo quedando disponibles para la absorción por las raíces. Vamos a mostrar un ejemplo de lo que ocurre con el catión potasio (K+) y los protones (H+), que provienen de las raíces o del ácido carbónico ( H2 CO3 ).

DIAGRAMA QUE MUESTRA EL INTERCAMBIO IÓNICO
Purves, et al.Life.The Science of Biology. 7th e.
La fertilidad de un suelo está determinada por su capacidad para aportar nutrientes por intercambio iónico.Las partículas de arcilla intercambian con eficacia los iones cargados positivamente que son retenidos en el horizonte A, mientras que los aniónes como el nitrato ( NO3
-) y el sulfato ( SO42- ) cargados negativamente se lavan rápidamente del
suelo por efecto del agua . La reserva de azufre, fósforo y esencialmente la única fuente de nitrógeno del suelo es la materia orgánica. La materia orgánica del suelo representa una acumulación de plantas parcialmente destruidas y de residuos de animales, que se descompone y mineraliza por acción de los animales y de los microorganismos del suelo. En los buenos suelos forestales, la masa de lombrices es de 20 a 80 g/m2 y la masa de bacterias alcanza 0,3 % del peso del suelo. La actividad de los organismos del suelo depende de las condiciones ambientales. En los climas tropicales, cálido-húmedos, la materia orgánica se descompone rápidamente y se mineraliza por completo. Sí las temperaturas son bajas y el suelo se encharca, la descomposición es lenta y se forma humus bruto o turba, y la respiración edáfica disminuye, la cual es un indicador de la actividad de los organismos del suelo.
El agua con las sales nutritivas disueltas forma la solución del suelo, mucha de las cuales son esenciales para el crecimiento de las plantas. En regiones áridas y semiáridas, la solución del suelo es más concentrada que en aquellas donde la lluvia es más abundante. Bajo condiciones de poca lluvia y mal drenaje, las concentraciones salinas son altas, pudiendo interferir de forma peligrosa con el crecimiento de las plantas.
La acidez del suelo puede influir en la absorción nutritiva y crecimiento de las plantas, ya sea directamente a través de las concentraciones de protones ( H+ ), o indirectamente, por su acción sobre la absorción de nutrientes y la presencia de iones tóxicos. Cuando los valores de pH están por debajo de 5 a 5,5, el aluminio, hierro y manganeso son solubles en cantidades suficientes como para ser tóxicos para algunas plantas. El ion fosfato es muy sensible a los cambios de pH, a pH 6,5 es fácilmente asimilable por las plantas, pero a pH 8, es insoluble y se fija como fosfato cálcico. En suelos con pH menores a 5, el ion fosfato se fija en forma insoluble con aluminio, hierro y manganeso, y no es aprovechable por las plantas. Estos ejemplos ilustran en parte el efecto de la acidez sobre la disponibilidad de algunos nutrientes del suelo.

CONTENIDO
/ CÉLULA VEGETAL / CÓMO SE ESTUDIAN LAS CÉLULAS / / CÉLULAS EUCARIOTICAS Y PROCARIOTICAS / CÉLULAS VEGETALES Y ANIMALES /
/ CÉLULA EUCARIÓTICA / LA PARED CÉLULAR / / LA MEMBRANA PLASMÁTICA / EL PROTOPLASTO / GLOSARIO / IMAGENES /
PÁGINA PRINCIPAL
CÉLULA VEGETAL
Todos los organismos vivos están compuestos por células. El inglés, Robert Hooke en 1665, realizó cortes finos de una muestra de corcho y observó usando un microscopio rudimentario unos pequeños compartimentos, que no eran más que las paredes celulares de esas células muertas y las llamó células ( del latín cellula, que significa habitación pequeña ) ; ya que éste tejido le recordaba las celdas pequeñas que habitaban los monjes de aquella época. No fue sino hasta el siglo XIX, que dos científicos alemanes el botánico Matthias Jakob Schleiden y el zoologo Theodor Schwann, enunciaron en 1839 la primera teoría celular : " Todas las plantas y animales están compuestos por grupos de células y éstas son la unidad básica de todos los organismos vivos". Esta teoría fue completada en 1855, por Rudolph Virchow, quien estableció que las células nuevas se formaban a partir de células preexistentes ( omni cellula e cellula ). En otras palabras las células no se pueden formar por generación espontánea a partir de materia inerte.
Célula fijada con KMnO4
En la frontera de lo viviente, se han descubierto seres aun más pequeños : los virus, que crecen y se reproducen solamente cuando parasitan otra célula. Podemos afirmar que, no hay vida sin célula. Al igual que un edificio, las células son los bloques de construcción de un organismo. La célula es la unidad más pequeña de materia viva, capaz de llevar a cabo todas las actividades necesarias para el mantenimiento de la vida.

La teoría celular actualmente se puede resumir de la siguiente forma :
1. Todos los organismos vivos están formados por células y productos celulares.2. Sólo se forman células nuevas a partir de células preexistentes.
3. La información genética que se necesita durante la vida de las células y la que se requiere para la producción de nuevas células se transmite de una generación a la siguiente.
4. Las reacciones químicas de un organismo, esto es su metabolismo, tienen lugar en las células.
Ir al principio
CÓMO SE ESTUDIAN LAS CÉLULAS
Una de las principales herramientas para el estudio de la célula es el microscopio. En general las células y tejidos vivos son difíciles de estudiar con el microscopio fotónico; ya que los tejidos multicelulares son demasiado gruesos para dejar pasar la luz y las células vivas aisladas suelen ser transparentes, con poco contraste entre los detalles internos. Sin embargo, se pueden realizar estudios de tejidos, realizando cortes a mano alzada con una hojilla bien afilada y haciendo observaciones con el microscopio óptico, previo montaje de la muestra sobre un porta objeto de vidrio, con una gota de agua y cubriendo con un vidrio cubre objeto.
Primeramente el estudio detallado de las células se ha favorecido con el mejoramiento de los microscopios y el desarrollo de métodos y técnicas para preparación y observación de las células. En segundo lugar, se trata de correlacionar los hallazgos estructurales con la información bioquímica.
Además de los avances en la microscopia que se observaron en la segunda mitad del siglo XIX y en el siglo XX, que han mejorado el poder de resolución de estos instrumentos, se han desarrollado también las técnicas básicas de preparación del material para su estudio con el microscopio :
1. Se fijan las células o tejidos con agentes que matan y estabilizan la estructura, p. ej. alcohol, ácido acético, formol, tetróxido de osmio, permanganato de potasio, entre otros.
2. Se deshidratan con alcohol etílico, butanol, acetona,etc
3. Se montan en sustancias duras que actúan como soporte del tejido para ser posteriormente cortados, ya sea con un micrótomo de Minot o con hojilla de diamante, si se requieren cortes ultra finos, para microscopia electrónica.

4. Se tiñen las células con colorantes que actúan sobre algunos organelos, produciendo contraste entre núcleo o citoplasma, o entre mitocondrias y otros elementos del citoplasma.
Existen distintos métodos de preparación para el estudio de ciertas característica celulares específicas En éste siglo, el desarrollo de las técnicas citológicas ha seguido las siguientes líneas : 1) se desarrollaron nuevos aparatos ópticos, como el microscopio de contraste de fase y se perfeccionaron otros como el microscopio de luz polarizada , facilitando así el estudio de las células vivas ; 2) se inventó el microscopio electrónico de transmisión ( TEM, transmission electron microscopy) y el microscopio electrónico de barrido( SEM, scanning electron microscopy) ; 3) se crearon métodos citoquímicos para lograr información química a partir de preparaciones microscópicas, entre estos se pueden citar la inmunofluorescencia y la microrradioautografía; 4) se idearon técnicas para fragmentar las células mediante , ultrasonido, homogeneizado, y el aislamiento de los organelos y otros componentes mediante centrifugación diferencial, para su posterior estudio bioquímico.
Ir al principio
CÉLULAS EUCARIÓTICAS Y PROCARIÓTICAS
En el mundo viviente se encuentran básicamente dos tipos de células: las procarióticas y las eucarióticas. Las células procarióticas (del griego pro, antes de; karyon, núcleo) carecen de un núcleo bien definido . Todas las otras células del mundo animal y vegetal, contienen un núcleo rodeado por una doble membrana y se conocen como eucarióticas ( del griego eu, verdadero y karyon, núcleo ). En las células eucarióticas, el material genético ADN, esta incluido en un núcleo distinto, rodeado por una membrana nuclear. Estas células presentan también varios organelos limitados por membranas que dividen el citoplasma celular en varios compartimientos, como son los cloroplastos, las mitocondrias, el retículo endoplasmático, el aparato de Golgi, vacuolas, etc.
Los organismos procariotes son unicelulares y pertenecen al grupo de las Moneras, que incluyen las bacterias y cianobacterias (algas verde-azules). El ADN de las células procarióticas está confinado a una o más regiones nucleares, que se denominan nucleoides, que se encuentran rodeados por citoplasma, pero carecen de membrana. En las bacterias, el nucleoide esta formado por un pedazo de ADN circular de aproximadamente 1 mm de largo, torcido en espiral, que constituye el material genético esencial. Las células procarióticas son las más primitivas de la tierra, hicieron su aparición en los océanos hace aproximadamente 3,5 millardos de años; mientras que las células eucarióticas fósiles tienen menos de un millardo de años.
Las células procarióticas son relativamente pequeñas, nunca tienen más de algunas micras de largo y no más de una micra de grosor. Las algas verde-azules son generalmente más grandes que las células bacterianas. Así mismo, todas las algas verde-azules realizan la fotosíntesis con la clorofila a, que no se encuentra en las bacterias, y mediante vías metabólicas comunes a las plantas y algas, pero no a las bacterias.

Un gran número de células procarióticas, están rodeadas por paredes celulares, que carecen de celulosa, lo que las hace diferentes de las paredes celulares de las plantas superiores.
En la parte interna de la pared celular, se encuentra la membrana plasmática o plasmalema, la cual puede ser lisa o puede tener invaginaciones, llamados mesosomas, donde se llevan a cabo las reacciones de transformación de energía (fotosíntesis y respiración). En el citoplasma, se encuentran cuerpos pequeños, esféricos, los ribosomas, donde se realiza la síntesis de proteínas. Así mismo, el citoplasma de las células procarióticas más complejas puede contener también vacuolas ( estructuras en forma de saco ), vesículas ( pequeñas vacuolas ) y depósitos de reserva de azucares complejos o materiales inorgánicos. En algunas algas verde-azules las vacuolas están llenas con nitrógeno gaseoso.
Muchas bacterias son capaces de moverse rápidamente gracias a la presencia de flagelos.
Ir al principio
CÉLULAS VEGETALES Y ANIMALES
Tanto las células de las plantas como las de los animales son eucarióticas, sin embargo presentan algunas diferencias:
1. Las células vegetales presentan una pared celular celulósica, rígida que evita cambios de forma y posición.
2. Las células vegetales contienen plastidios, estructuras rodeadas por una membrana, que sintetizan y almacenan alimentos. Los más comunes son los cloroplastos.
3. Casi todas las células vegetales poseen vacuolas, que tienen la función de transportar y almacenar nutrientes, agua y productos de desecho.
4. Las células vegetales complejas, carecen de ciertos organelos, como los centriolos y los lisosomas.

Ir al principio
CÉLULA EUCARIÓTICA
Las plantas son organismos multicelulares formados por millones de células con funciones especializadas. Sin embargo, todas las células vegetales poseen una organización común: tienen un núcleo, un citoplasma y organelos subcelulares; los cuales se encuentran rodeados por una membrana que establece sus límites. Así como una pared celular que rodea el protoplasto (núcleo + citoplasma con sus inclusiones).
Ir al principio
LA PARED CÉLULAR
Aunque las células vegetales y animales son muy parecidas, las células vegetales tienen una pared rígida de celulosa, que le brinda protección, sin impedir la difusión de agua y iones desde el medio ambiente hacia la membrana plasmática, que es la verdadera barrera de permeabilidad de la célula. Una pared celular primaria típica, de una dicotiledónea está formada por 25-30 % de celulosa, 15-25 % de hemicelulosa, 35 % de pectina y 5-10 % de proteínas (extensinas y lectinas), en base al peso seco. La constitución molecular y estructural precisa de la pared celular, depende del tipo de célula, tejido y especie vegetal.
La pared primaria es delgada ( de 1 a 3 micras de grosor) y se forma cuando la célula crece, ejemplo de esta la tenemos en células jóvenes en crecimiento, en el tejido parenquimático, en el clorénquima, epidermis, etc.
La membrana celular está fuertemente adherida a la pared celular, debido a la presión de turgencia provocada por los fluidos intracelulares. Literalmente podemos decir que las células se encuentran abombadas, empujándose entre ellas; en otras palabras se encuentran infladas por una presión hidrostática.
Las macromoléculas de celulosa, en la pared celular esta formada por unidades de glucosa (un azúcar de 6 carbonos) enlazadas covalentemente, formando una estructura en forma de cinta aplanada, que puede tener de 0,25 a 5 micras de largo. Entre 40 a 70 de estas cadenas se mantienen unidas mediante enlaces de hidrógeno, entre los grupos OH de los residuos de glucosa, formando una estructura cristalina llamada microfibrilla, que tiene aproximadamente 3 nm de diámetro. La celulosa es muy estable químicamente e insoluble. Las microfibrillas tienen una alta fuerza tensional, que actua reforzando la pared. Grupos de microfibrillas se disponen como los alambres en un cable, formando macrofibrillas. Las macrofibrillas son los componentes más importantes de la pared celular y se mantienen unidas mediante otros componentes de la pared celular, como son las macromoléculas de hemicelulosa y péctina. Estas sustancias pegan toda la estructura, en capas de fibras. Las primeras microfibrillas que se depositan en la pared celular, forman una red con disposición transversal. Pero, cuando la presión de turgencia produce la extensión celular y la pared crece en área superficial, la otra capa de microfibrillas se deposita paralelamente, al eje longuitudinal de la célula. El efecto final es una apariencia entramada de varias capas.
Dos células adyacentes se mantienen unidas mediante la lámina media, la que se encuentra formada principalmente por sustancias pecticas, que cementan las paredes primarias, a ambos lados de la lámina media. Nosotros podemos extraer la pectina de frutos verdes, como por Ej. el mango y hacer jalea. En muchas plantas posteriormente se puede depositar una pared celular secundaria, que imparte rigidez y fortaleza al tejido, sí se deposita lignina. Por ejemplo los troncos de los árboles, tienen células con gruesas paredes celulares secundarias.

Las plantas multicelulares, se conectan a través de pequeñas perforaciones que comunican las células adyacentes, denominadas campos de punteaduras primarias, a través de los cuales pasan cordones citoplasmáticos denominados plasmodesmos. A pesar de que son muy pequeños para que lo atraviesen organelos celulares, sin embargo las conexiones citoplasmáticas permiten la transferencia de sustancias de una célula a otra. La membrana plasmática es continua y se extiende de una célula a la otra a través de los plasmodesmos, constituyendo lo que se conoce como simplasto ; mientras que el conjunto de las paredes celulares de un tejido, más los espacios intercelulares, se denomina apoplasto. La pared celular es muy permeable a diferentes sustancias, permitiendo el paso de agua y solutos; aunque la verdadera barrera que controla la permeabilidad, al igual que en las células animales, es la membrana plasmática o plasmalema.
Ir al principio
LA MEMBRANA PLASMÁTICA
La membrana plasmática, tanto de las células procarióticas como eucarióticas, son básicamente similares. En ambos casos, regula el flujo de sustancias disueltas hacia adentro y hacia afuera de la célula. La ósmosis, que funciona debido a que el agua pasa a través de las membranas más rápido que los solutos, regula el flujo de agua.
Las membranas plasmáticas tienen aproximadamente 50% de fosfolípidos y 50% de proteínas. La estructura en tres capas de las membranas celulares, consiste de una doble capa de fosfolípidos, con los grupos hidrofóbos(no afines al agua) mirando hacia el centro y los grupos hidrofílicos (afines al agua) orientados hacia las partes externas de la bicapa lípidica. Las moléculas de proteínas, flotan en la bicapa lipídica, con sus terminaciones hidrofílicas penetrando en ambas superficies de la membrana, lo que se conoce como el modelo de mosaico fluido, propuesto por Singer y Nicolson (1972). Se sabe que en las membranas existen dos tipos de proteínas : las proteínas integrales (intrínsecas) y las proteínas periféricas (extrínsecas).
Cuando se estudia la membrana plasmática mediante el microscopio electrónico, después de haber sido apropiadamente fijada con tetróxido de osmio, las capas de proteínas se observan como dos líneas densas (oscuras), con un espacio claro entre ellas. Las líneas oscuras tienen un espesor de aproximadamente 2,5 a 3,5 nm y la línea clara tiene aproximadamente 3,5 nm, para un grosor de aproximadamente 10 nm o100
. La que se conoce como la unidad de membrana. Esto no significa que todas las membranas sean

iguales; ya que ellas pueden presentar diferentes características de permeabilidad. El hecho de que una sustancia pueda atravesar la membrana de un cloroplasto, no significa que lo pueda hacer también a través de una membrana mitocondrial. Las membranas poseen la propiedad de ser selectivas, lo que indica que cada tipo de membrana tiene características moleculares particulares, que les permite funcionar bajo sus propias condiciones. Todas las membranas biológicas que rodean las células, núcleos, vacuolas, mitocondrias, cloroplastos y otros organelos celulares son selectivamente permeables. Las membranas son muy permeables a las moléculas de agua y ciertos gases, incluyendo el oxígeno y el dióxido de carbono ; mientras que otras moléculas pueden tener problemas para atravesar las membranas, debido a su tamaño, polaridad y solubilidad en lípidos. Los iones y las moléculas polares (con carga eléctrica), tienden a moverse a través de la parte proteica de la membrana. Muchas sustancias se mueven mediante difusión simple, por un proceso de transporte pasivo, de zonas de mayor a menor concentración. Sin embargo, en los seres bióticos muchas sustancias atraviesan la membrana mediante transporte activo, moviéndose en contra de un gradiente de concentración, y con la utilización de energía metabólica por la célula , en forma de ATP (adenosin trifosfato), el cual es aportado por la respiración.
Ir al principio
EL PROTOPLASTO
El contenido del protoplasto, se puede dividir en tres partes fundamentales: citoplasma, núcleo y vacuola(s); así mismo se encuentran sustancias ergásticas y órganos de locomoción. Todas las células eucarióticas, al menos cuando jóvenes poseen un núcleo ; el cual puede desaparecer en los tubos cribosos y en otras células vegetales, en la medida que maduran. El protoplasto se encuentra ausente en los elementos xilemáticos maduros ( vasos y traqueidas). La presencia de vacuolas y sustancias ergásticas, es una característica de las células de hongos y de las plantas.
El citoplasma (plasma fundamental), tiene una consistencia viscosa y está compuesto de una mezcla heterogénea de proteínas (enzimas) y es el lugar donde ocurren importantes reacciones metabólicas, como la glucólisis. Debido a su naturaleza coloidal, el citoplasma sufre cambios de estado, puede pasar de sol (fluido) a gel (parecido a la gelatina). El citosol, es la matriz fluida en la que los organelos se encuentran suspendidos, está organizado en una red tridimensional de proteínas fibrosas, llamadas citoesqueleto. El citoesqueleto es mucho más organizado, que la sopa clara que nos podemos imaginar.
Los elementos del citoesqueleto son: los microtúbulos y los microfilamentos. Los microtúbulos son filamentos cilíndricos, huecos que tienen un diámetro externo de 25 nm y varias micras de longitud. Las paredes de los microtúbulos, están formadas por filamentos protéicos lineares o en espiral de aproximadamente 5 nm de diámetro y estos están compuestos de 13 subunidades. En el centro de un microtúbulo se encuentra un lumen (área vacía); sin embargo se pueden observar bastones o puntos. Los microtúbulos estan compuestos por moléculas esféricas de una proteína llamada tubulina. Los microtúbulos pueden formarse o descomponerse rápidamente a conveniencia, y se encuentran formando parte de estructuras celulares que facilitan el movimiento, como el huso mitótico y los flagelos. La colquicina, un alcaloide del cólquico (Colchicum autumnale), destruye la organización de los microtúbulos, impidiendo la formación del huso acromático durante la mitosis celular. Por lo que la colquicina se ha utilizado en genética, en la obtención de células poliploides.
Los microfilamentos son estructuras más pequeñas, pero sólidas de 5 a 7 nm de diámetro, que actúan solos o conjuntamente con los microtúbulos para producir movimiento celular. Estos también están formados por proteínas, específicamente la proteína actina, la que con la miosina son también constituyentes del tejido muscular de los animales. Los microfilamentos causan el movimiento de corriente citoplasmática o ciclosis, la que ocurre en muchas células vegetales, como en las algas Chara y Nitella, donde se han reportado velocidades de 75 por segundo. En las hojas de la Elodea canadensis, se observa muy bien la ciclosis, que produce un movimiento de los organelos celulares, de una forma helicoidal, de un lado hacia abajo y del otro lado hacia arriba. Los microfilamentos también juegan un papel importante en el crecimiento del tubo polínico y en el movimiento ameboidal. En el citoplasma se

encuentra un sistema de endomembranas, que incluye al retículo endoplasmático, el aparato de Golgi, la envoltura nuclear y otros organelos celulares y membranas (tales como los microcuerpos, esferosomas y membrana vacuolar), que tienen sus orígenes en el retículo endoplasmático o en el aparato de Golgi. La membrana celular que ya la hemos estudiado, se considera como una entidad separada; aunque su crecimiento se debe a la adición de vesículas por el aparato de Golgi. Las mitocondrias y plastidios se encuentran rodeados por una doble membrana, que se parece al sistema de endomembranas; aunque estos organelos se autoduplican, por lo que no están relacionados al sistema de endomembranas. Así mismo, los ribosomas, los microtúbulos y los microfilamentos, no forman parte del sistema de endomembranas.
El retículo endoplasmático (RE o ER, del inglés endoplasmic reticulum) es un sistema multirramificado de sacos membranosos planos, denominados cisternas, que presentan la típica estructura de unidad de membrana. El RE es continuo con la membrana externa de la envoltura nuclear, a la que se une en las cercanías del núcleo. El RE puede tener ribosomas, que se encuentran unidos como lo hacen los botones a un pedazo de tela, y se conoce como RE rugoso o puede carecer de ribosomas y se llama RE liso. El RE rugoso sintetiza lípidos de membrana y proteínas de secreción; mientras que el RE liso está implicado también en la producción de lípidos y en la modificación y transporte de las proteínas sintetizadas en el RE rugoso. Los ribosomas, observados en una micrografía electrónica a bajo aumento, aparecen como puntos negros, redondos sobre el RE, pero a altos aumentos se observa que están formados por un cuerpo pequeño esférico y un cuerpo cóncavo grande, tienen de 20 a 30 nm de grosor. Frecuentemente aparecen formando agregados característicos que reciben el nombre de polisomas. Los ribosomas son partículas de ribonucleoproteínas (contienen proteínas y ácido ribonucleíco), donde se produce la síntesis de proteínas a partir de aminoácidos, mediante el mecanismo de la traducción, de la información genética contenida en el ácido ribonucleico mensajero (ARNm). En una célula pueden existir miles de ribosomas, con una capacidad de síntesis prodigiosa, ya que cada ribosoma puede producir una molécula de proteína por minuto.
Complejos de Golgi o aparato de Golgi esta relacionado con el RE; éste sistema de membranas está compuesto por conjuntos de sacos de Golgi, aplanados y llenos de fluido. Se observan como membranas aplanadas, parecidas a una pila de cachapas. En los extremos de estas membranas aplanadas o cisternas, se pueden observar vesículas que contienen las macromoléculas que se usan para la construcción de las membranas y la pared celular. Tanto los polisacáridos hemicelulosa y pectina, como la proteína de la pared celular (extensina) son sintetizados y procesados en el interior de las vesículas de secreción del aparato de Golgi o dictiosoma. Cada aparato de Golgi tiene 4 a 6 cisternas con una separación de 10 nm; no obstante algunas algas pueden tener de 20 a 30. El aparato de Golgi puede tener otras funciones además de contribuir al crecimiento del plasmalema y transporte de material a la pared celular, como es la de segregar mucílago en la parte externa de la punta de la raíz, que actúa como un lubricante permitiendo su movimiento entre las partículas del suelo. El aparato de Golgi es abundante en muchas células secretoras. Los dictiosomas no son estructuras permanentes y en caso de necesidad se forman de novo por el retículo endoplasmático.
Microcuerpos, peroxisomas, glioxisomas. Los microcuerpos son organelos esféricos, rodeados por una sola unidad de membrana. Su diámetro varía de 0,5 a 1,5 y tienen un interior granular; algunas veces con inclusiones cristalinas de proteínas. Se originan a partir del RE, formando parte del sistema de endomembranas. Los peroxisomas son organelos esféricos, especializados en reacciones de oxidación. La enzima catalasa, constituye casi el 40% de las proteínas totales del peroxisoma, esta enzima descompone el peróxido de hidrógeno en agua y oxígeno. En las plantas se conocen los peroxisomas foliares, como organelos de la foto respiración. Los glioxisomas se encuentran en semillas de oleaginosas, y contienen las enzimas que ayudan a convertir las grasas almacenadas, en carbohidratos que son translocados a la planta joven para su crecimiento. Los glioxisomas contienen las enzimas del ciclo del ácido glicólico.
Plastidios. Además del núcleo y las vacuolas, los plastidios constituyen los organelos más conspicuos de una célula vegetal. Los plastidios están rodeados por una doble membrana, con una estructura interna constituida por un sistema de membranas, separadas por una matriz de naturaleza proteica llamada estroma. Los plastidios tienen ADN (DNA) con una estructura similar al encontrado en células procarióticas, así como ribosomas, embebidos en el estroma. Todos los plastidios se desarrollan a partir de proplastidios, que son cuerpos pequeños encontrados en plantas que crecen tanto en la luz como en la oscuridad. Se dividen por fisión o bipartición, como lo hacen las mitocondrias y los organismos procariótes. Los plastidios incoloros se conocen como leucoplastos, contienen enzimas responsables de la síntesis del almidón. Los leucoplastos mejor conocidos son los amiloplastos, que almacenan granos de

almidón, como los encontrados en la raíz de la yuca, el tubérculo de la papa, en granos de cereales, etc. Otros leucoplastos pueden almacenar proteínas, se conocen como proteinoplastos. Los cromoplastos son organelos coloreados, especializados en sintetizar y almacenar pigmentos carotenoides (rojo, anaranjado y amarillo), estos son el origen de los colores de muchos frutos, flores y hojas, por ej. la piel del tomate, la raíz de zanahoria, etc. Los cromoplastos se originan a partir de cloroplastos jóvenes o de cloroplastos maduros, por división.
Los cloroplastos son plastidios que contienen los pigmentos verdes clorofila a y b, así como carotenoides de color anaranjado y xantofilas amarillas, son característicos de los seres fotoautótrofos, que poseen la maquinaria enzimática para transformar la energía solar en energía química, a través de la fotosíntesis. Los cloroplastos son característicos de las células del mesófilo foliar, poseen una doble membrana que los asemeja a las mitocondrias. Tienen una membrana externa y otra interna, el espacio delimitado por la membrana interna está ocupado por un material amorfo, parecido a un gel, rico en enzimas, denominado estroma. Contiene las enzimas que realizan la fijación o reducción del CO2, convirtiéndolo en carbohidratos, como el almidón. La membrana interna de los cloroplastos también engloba un tercer sistema de membranas, que consta de sacos planos llamados tilacoides, en los cuales la energía luminosa se utiliza para oxidar el agua y formar ATP (compuesto rico en energía) y NADPH (poder reductor), usados en el estroma para convertir el CO2 en carbohidratos. En ciertas partes de los cloroplastos, los tilacoides se disponen como monedas apiladas, denominados grana, pero en el estroma permanecen aislados.
Los cloroplastos tienen forma elíptica, con un diámetro de 5 a 10 y su número puede variar de 20 a 100 por célula vegetal. Durante la ciclosis se mueven libremente en el citoplasma. Ellos responden directamente a la energía solar, para llevar a cabo la fotosíntesis, orientándose perpendicularmente a los rayos de luz; sin embargo sí la energía lumínica es muy fuerte, se disponen de tal forma que la radiación incida oblicuamente, recibiendo menos luz.
Los cloroplastos se originan a partir de proplastidios, reacción ésta que es disparada por la luz, que provoca la diferenciación del plastidio, apareciendo los pigmentos y la proliferación de membranas, que origina los tilacoides y grana. Así mismo, en el estroma del cloroplasto se encuentran pequeños pedazos circulares de ADN, dispuestos en doble hélice; parecidos al ADN de las mitocondrias y bacterias. El ADN del cloroplasto regula la síntesis del ARN ribosomal, del ARN de transferencia y de la Ribulosa 1,5 difosfato carboxilasa-oxigenasa (RUBISCO), enzima que cataliza la fijación del CO2 en la fotosíntesis. Sin embargo, la mayoría de las proteínas del cloroplasto, son sintetizadas en el citosol y transportadas al cloroplasto.
Mitocondrias. Las células eucarióticas poseen organelos complejos, denominados mitocondrias. Observadas con el microscopio óptico, se ven como pequeñas esferas, bastones o filamentos, que varían en forma y tamaño, comúnmente miden de 0,5 a 1,0 de diámetro y de 1,0 a 4,0 de longitud. Son más numerosas que los cloroplastos, pudiéndose encontrar hasta 1000 por célula, pero varias algas, incluyendo Chlorella tienen una sola por célula La mitocondria es el organelo responsable de la respiración aeróbica (que utiliza O2), un proceso en el cual un carbohidrato se oxida por completo en presencia de O2, convirtiéndose en CO2, H2 O y energía almacenada en forma de ATP. Las mitocondrias se dividen por fisión o bipartición, y todas se originan a partir de las mitocondrias contenidas en el zigoto; de tal forma que sus membranas no se derivan del sistema de endomembranas. Ellas contienen ADN circular y ribosomas pequeños (15 nm), en la matriz, de tal manera que son capaces de sintetizar algunas de sus propias proteínas. Sin embargo, dependen también de proteínas sintetizadas en el citoplasma que están bajo el control nuclear.
Las mitocondrias tienen una doble membrana, la membrana externa es lisa, y actúa como un colador, permitiendo el paso de muchas moléculas pequeñas; mientras que la membrana interna, muestra plegamientos denominados crestas, que aumentan la superficie interna. La membrana interna es selectivamente permeable, regulando el tipo de moléculas que la atraviesan. El compartimiento interno encerrado por la membrana interna es la matriz, de naturaleza coloidal, que contiene las enzimas del ciclo de Krebs o del ácido cítrico. En la membrana interna de las mitocondrias, se encuentran insertos los transportadores de electrones y la ATP sintetasa, realizándose en ella la fosforilación oxidativa o sea la síntesis de ATP, acoplada al consumo de O2.
Vacuolas. Son organelos característicos de las células vegetales, rodeados por una membrana denominada tonoplasto, que controla el transporte de solutos hacia adentro y hacia afuera de la vacuola; regulando el potencial hídrico de la célula a través de la osmosis. La vacuola contiene iones inorgánicos, ácidos

orgánicos, azucares, enzimas, cristales de oxalato de calcio, y una variedad de metabolitos secundarios (alcaloides, taninos, ), que frecuentemente juegan un papel en la defensa de las plantas. Algunas vacuolas tienen altas concentraciones de pigmentos, hidrosolubles, que le dan la coloración a muchas flores, hojas y a la raíz de remolacha. Los colorantes vacuolares, de hojas y flores sirven para atraer los insectos que transportan el polen y, en parte funcionan como pigmentos protectores del exceso de radiación. Las vacuolas pueden almacenar proteínas, especialmente en legumbres y cereales, es importante señalar los granos de aleurona, en las células de la capa de aleurona de los cereales (trigo, cebada) o en los cotiledones de semillas de leguminosas (caraota, arveja, lenteja). Al germinar las semillas, las proteínas son hidrolizadas y los aminoácidos transferidos al embrión en crecimiento. Algunas vacuolas almacenan grasas como oleosomas o cuerpos grasos, p.ej. el endosperma del Ricinus communis (aceite de ricino). Las vacuolas son ricas en enzimas hidrolíticas, como proteasas, ribonucleasas, y glicosidasas, que cuando se liberan en el citosol, participan en la degradación celular durante la senescencia. Las vacuolas tienen un pH más ácido que el citosol, cualquier exceso de iones de hidrógeno en el citosol es bombeado hacia la vacuola, manteniéndose la constancia del pH citosólico. En vista de la cantidad de sustancias que se acumulan en la vacuola, se ha pensado de ellas por mucho tiempo, que son como el botadero de productos de desechos celulares (sustancias ergásticas).
Las vacuolas se originan a partir de pequeñas vacuolas en células, jóvenes, meristemáticas del ápice del tallo o de la raíz, las que crecen con la célula, absorbiendo agua por osmosis y uniéndose unas con otras, hasta que se forman grandes vacuolas. Las pequeñas vacuolas o provacuolas parecen formarse a partir del aparato de Golgi o del retículo endoplasmático .
El núcleo celular y sus componentes. El núcleo es el organelo celular más conspicuo, tiene forma esférica o globular, con un diámetro de 5 a 15 . Es el centro de control de la célula; sin embargo no es un organelo independiente, ya que debe obtener sus proteínas del citoplasma. El núcleo contiene la mayor cantidad de ADN, al que se le da el nombre de genoma, está rodeado por una envoltura nuclear, compuesta de dos membranas, que se fusionan en algunos puntos formando poros nucleares, que permiten la comunicación del interior del núcleo con el citoplasma celular. Pueden existir desde pocos a miles de poros en una envoltura nuclear. Algunas macromoléculas del núcleo, incluyendo subunidades ribosomales, son capaces de atravesar los poros nucleares hacia el citosol y viceversa. El núcleo ejerce su control sobre las funciones celulares vía ARNm (ácido ribonucleico mensajero), determinando las enzimas que se fabrican en la célula y éstas a su vez determinan las reacciones químicas que se llevan a cabo, y por ende la estructura y función celular.
El núcleo es el sitio de almacenamiento y replicación de los cromosomas, que están compuestos de ADN y proteínas acompañantes. El complejo ADN-proteína (nucleoproteina), se denomina cromatina, que se observa dispersa durante la interfase. Aunque la cromatina pareciera estar desordenada, no es así; ya que está organizada en estructuras llamados cromosomas. La longitud de todo el ADN del genoma de una planta es millones de veces mayor que el diámetro del núcleo donde se encuentra, podemos establecer la analogía con una bola de hilo enrollada varios kilómetros de longitud, metida dentro de una pelota de golf. Cuando una célula se prepara para dividirse, el ADN y las proteínas que forman cada cromosoma se enrollan más estrechamente; los cromosomas se acortan, engruesan y se hacen visibles al microscopio. El núcleo contiene una solución acuosa, repleta de enzimas, el nucleopasma, en el cual se encuentran suspendidos la cromatina o los cromosomas y los nucléolos. Como ya mencionamos, el ADN almacena información, en forma de genes, que son segmentos o secuencias de ADN que contienen toda la información genética para originar un producto génico determinado -ARN, proteína-.
El núcleo contiene uno o más cuerpos esféricos (pueden ser hasta 4), los nucléolos, que pueden tener de 3 a 5 de diámetro. Los nucléolos son masas densas de fibras, de forma irregular, se tiñen de oscuro, que se encuentran suspendidos en el nucleoplasma. En ellos se pueden encontrar áreas claras, llamadas vacuolas nucleolares, que son indicativos de un nucléolo muy activo. Las células meristemáticas, generalmente tienen nucléolos más grandes que las células maduras o latentes. En el nucléolo se fabrica el ARN ribosomal, que junto a las proteínas sintetizadas en el citoplasma, forman los ribosomas. El ARN ribosomal es codificado por regiones especiales en los cromosomas denominadas regiones organizadoras del nucléolo. Los nucéolos se observan bien durante la interfase de la mitosis, que es la fase de descanso de la división celular, pero cuando la célula comienza a dividirse, en la profase, desaparecen los nucléolos y la membrana nuclear, que se reabsorbe en el retículo endoplasmático.

Ir al principio
GLOSARIO
A B C D E F G H I M N O P R S
Abiótico: Contrario a la vida.
Absorción: El proceso de acumulación, como la absorción por las raíces.
Ácido: Un donador de protones ( H+ ), una sustancia que libera protones y por lo tanto causa que el pH de una solución sea menor de 7,0.
Acídico: Que posee un número relativo grande de protones y tiene un pH menor de 7,0.
Ácido desoxirribonucleico ( ADN ): Un ácido nucleico de doble cadena, compuesto de adenina, guanina, citosina, timina, desoxirribosa y fosfato.
Ácido Ribonucleico (ARN): Un ácido nucleico de una sola cadena, compuesto de adenina, guanina, citosina, uracilo, ribosa y fosfato.
Ácido ribonucleico mensajero ( ARNm): Un ácido ribonucleico que se transcribe a partir de la matriz de ADN .
Ácido ribonucleico ribosomal ( ARNr): Un ácido rinucleico que participa en la formación de los ribosomas.
Ácido ribonucleico de transferencia (ARNt): Una molécula pequeña de ácido ribonucleico, que participa en la transferencia de aminoácidos específicos para la síntesis de una proteína.
Actina: Proteína globular, participa en los mecanismos de contracción de los microfilamentos.
Adenina: Base nitrogenada halladas en el ADN y ARN.
Adenosin trifosfato.( ATP ): Compuesto orgánico que contiene, adenina, ribosa y tres grupos fosfatos. La mayor fuente de energía química para las reacciones metabólicas.
Aeróbico: Organismo que metaboliza en presencia de oxígeno molecular.
Aleurona: Producto de naturaleza proteica, que se almacena en las semillas y que se moviliza durante la germinación.
Alcalina: Sustancia que libera iones hidroxilos en solución. Aumenta el pH por encima de 7,0.
Alcaloide: Un grupo de compuestos nitrogenados orgánicos de origen vegetal, de carácter básico. Muchos alcaloides tienen propiedades medicinales, alucinógena o tóxica.
Alga verde azul: Organismo procariote, con pigmentos de ficobilina para realizar la fotosíntesis.
Almidón: Un polisacárido compuesto por moléculas de a -glucosa, es el principal producto de reserva de las plantas.

Aminoácido: Un ácido orgánico con un grupo amino (NH2 ) y un grupo carboxilo ( -COOH ). Los aminoácidos se unen entre sí para formar las moléculas de proteínas.
Amilasa: Una enzima que hidroliza el almidón convirtiéndolo en azucares.
Anaeróbico: Organismo que funciona en ausencia de oxígeno molecular.
Antocianinas: Un grupo de pigmentos flavonoides, solubles en agua, que le dan las coloraciones, azul, rojo y morado a las flores.
Aparato de Golgi: Organelos compuestos de pilas de membranas aplanadas, que funcionan en el empaquetamiento y síntesis de membranas y pared celular.
Apoplasto: Esta formado por la pared celular y los espacios intercelulares, que constituyen el espacio libre del tejido. El agua y los solutos se mueven a través del espacio libre.
Autotrófico: Un organismo que produce sus propios alimentos a través de la fotosíntesis, p. ej. las plantas verdes.
Ir inicio glosario
Buffer: Cualquier sustancia que absorbe o libera protones para mantener el pH de la solución estable, inclusive sí se añade un ácido o una base.
b -caroteno: Un carotenoide vegetal importante, precursor de la vitamina A.
Ir inicio glosario
Campo de punteaduras primario: Región en la pared primaria, donde los plasmodesmos atraviezan la pared celular.
Capa de aleurona: Un grupo de células ricas en gránulos de proteínas y localizada como la capa externa del endosperma de muchas semillas
Carbohidrato: Un compuesto orgánico que contiene carbono, hidrógeno y oxígeno en el patrón básico CH2 O; como los azucares, almidón y celulosa.
Carotenoide: Un terpeno compuesto por ocho unidades de isopreno, sintetizado por muchas plantas. Pigmentos de colores anaranjado rojizo, que se encuentran en la zanahoria, hojas, etc.
Catalizador: Cualquier sustancia que acelera la velocidad de una reacción sin participar en ella. En las reacciones bioquímicas las enzimas, son proteínas que actúan como catalizadores, disminuyendo la energía de activación.
Células somáticas: Todas las células del soma o cuerpo, diferentes de las células sexuales, que contienen al menos los dos conjuntos de cromosomas heredados de ambos padres.
Celulasa: Una enzima que hidroliza la celulosa, convirtiéndola en glucosa.
Celulosa: El principal carbohidrato estructural de las células vegetales. Es un polímero de la glucosa (esta compuesta de muchas unidades de glucosa).
Cigoto: Célula diploide (2n) resultante de la union de dos gametos haploides (n), óvulo fecundado como resultado de la fertilización.
Clorofila: La molécula responsable de captar la energía luminosa en los primeros eventos de la fotosíntesis. Es un pigmento de color verde.

Cloroplastos: Organelos encontrados en las partes superiores de las plantas (tallos, hojas, frutos, etc.), contienen clorofila y realizan la fotosíntesis.
Cromatina: Es el material nuclear que se tiñe de oscuro, presente durante la interfase. Esta compuesta de ADN y proteínas.
Cromoplasto: Organelos rodeados por una membrana que almacenan carotenoides.
Cromosoma: Filamentos microscópicos dentro del núcleo de células eucarióticas, que tienen el ADN responsable de la herencia. Contienen las unidades hereditarias o genes.
Clon: Organismos genéticamente idénticos.
Cresta: Pliegues presentes en la membrana interna de las mitocondrias.
Cianobacteria: (Gr. kyanos, azul oscuro). Algunas veces llamadas algas verde azules, bacterias fotosintéticas; productores importantes de oxígeno para la evolución de la vida sobre la tierra.
Ciclosis: Circulación del citoplasma en el interior de la célula junto con algunos organelos.
Citoplasma: El contenido viscoso de la célula, que se encuentra en la parte interna de la membrana plasmática, excluyendo al núcleo.
Citosol: Es la savia celular, es el medio acuoso, en el que están suspendidos los organelos y las partículas insolubles de la célula.
Ir inicio glosario
Desoxirribosa: Un azúcar de cinco carbonos que forma parte del ADN.
Difusión: El movimiento de moléculas al azar, de una región de alta concentración a otra de baja concentración.
Dióxido de carbono: Una molécula gaseosa compuesta de un átomo de carbono y dos de oxígeno, que participa en la fotosíntesis y es liberada en la respiración.
Doble hélice: Una hélice compuesta de dos cadenas moleculares que se enrollan entre sí, como en el ADN.
Ir inicio glosario
Envoltura nuclear: La membrana que rodea el núcleo en células eucarióticas.
Enzima: Es un biocatalizador de naturaleza proteíca.
Ergástico: Inclusiones de material relativamente puro, frecuentes en plastidios o vacuolas, p ej. cristales como el de oxalato de calcio, grasas, granos de almidón, taninos, cuerpos proteicos.
Estroma: La matriz proteica entre las granas de los cloroplastos. Sitio de las reacciones oscuras de la fotosíntesis.
Etanol: El alcohol etílico es el producto final de la fermentación alcohólica.
Eucariote: Organismo cuyas células poseen núcleo delimitado por membrana.
Ir inicio glosario

Fisión: Proceso de reproducción asexual, en el que un organismo u organelo se divide en dos partes más o menos iguales. La forma más común de reproducción en procariotes.
Fosforilación oxidativa: Producción de ATP por las mitocondrias, acoplada al consumo de oxígeno.
Fotosíntesis: Es la producción de carbohidratos por la combinación de CO2 y H2O, en los cloroplastos, catalizada por la luz, con la liberación de O2 .
Ir inicio glosario
Gen: La unidad de la herencia. Un grupo de nucleótidos en la molécula de ADN responsable por la herencia de un carácter particular. Codifica una proteína.
Genoma: El complemento genético total de un organismo.
Grasas: Moléculas orgánicas que contienen gran cantidad de carbono e hidrógeno, pero poco oxígeno. Los aceites son grasas en el estado líquido.
Glucosa: Un monosacárido de 6-carbonos ( azúcar simple), el primer substrato de la respiración.
Glucólisis: Una serie de reacciones que preceden la respiración aeróbica o anaeróbica, en la que la glucosa es oxidada a ácido pirúvico.
Glioxisoma: Un microcuerpo subcelular presente en el citoplasma de muchas semillas oleaginosas. Las enzimas del glioxisoma convierten lípidos a carbohidratos durante el proceso de la germinación.
Grana: Estructuras en el interior de los cloroplastos, que se observan como gránulos verdes con el microscopio óptico y con el microscopio electrónico como una pila de membranas en forma de discos. La grana contiene las clorofilas y carotenoides y son el sitio de las reacciones luminosas de la fotosíntesis.
Ir inicio glosario
Hemicelulosa: Un polisacárido componente de la pared celular primaria; similar a la celulosa, pero degradado más fácilmente.
Herencia: La transmisión de caracteres genéticamente controlados de padres a hijos a través de la reproducción sexual.
Heterotrófico: Un organismo que obtiene sus alimentos a partir de otros organismos. Histonas. Proteínas básicas que constituyen una porción del material nuclear, asociadas funcionalmente al ADN.
Hidrólisis: El rompimiento de una molécula grande en moléculas pequeñas, mediante la adición de agua.
Hidrofílico: La propiedad que tiene una sustancia de atraer agua.
Hidrofóbico: La propiedad que tiene una sustancia de repeler el agua.
Ir inicio glosario
Impermeable: Que tiene la propiedad de restringir el pasaje de sustancias.
Inorgánico: Un compuesto químico sin carbono en su esqueleto atómico.
Interfase: La condición nuclear entre una mitosis y la próxima. Los cromosomas no son visibles, aunque ocurre una intensa actividad metabólica.

Ion: Un átomo o molécula que ha ganado o perdido un electrón, haciendo que la partícula se cargue eléctricamente.
Ir inicio glosario
Lámina media: La capa cementante de sustancias pécticas entre dos paredes celulares primarias.
Leucoplasto: Organelo rodeado por una membrana, especializado en el almacenamiento de almidón.
Lignina: Una molécula orgánica compleja hallada como componente importante de las paredes secundarias; imparte rigidez y fortaleza a las microfibrillas de celulosa.
Lípido: Un aceite o grasa, formado por glicerol y ácidos grasos.
Lumen: La cavidad central de una célula.
Ir inicio glosario
Macrofibrilla: Un agregado de microfibrillas en la pared celular, visibles con el microscopio óptico.
Macromolécula: Una molécula muy grande. Término generalmente aplicado a polisacáridos, lípidos, proteínas y ácidos nucleicos.
Membrana: En los organismos vivos, una bicapa de fosfolípidos impregnada con proteínas y otros compuestos; funciona en la compartimentalización de la actividad celular.
Membrana diferencialmente permeable: Una membrana que permite el paso de ciertas partículas e impide el paso de otras; también se conoce como membrana selectivamente permeable.
Meristema: Es una zona o tejido, a partir del cual se forman células nuevas por división.
Metabolismo: La suma de todas las reacciones químicas que ocurren en una célula, incluyen tanto las de síntesis, como las de degradación.
Microfibrilla: Un cordón alargado de moléculas de celulosa.
Microtúbulos: Estructuras delgadas cilíndricas, formados por la proteína tubulina, que son importantes en la síntesis de algunas membranas.
Mitocondria: Un organelo celular rodeado por una doble membrana, cuya función es la respiración aeróbica.
Mitosis: La división nuclear de las células somáticas, que da como resultado dos núcleos hijos idénticos.
Mosaico fluido: Se refiere al modelo de la estructura de las membranas, que consiste en una bicapa de lípidos con proteínas globulares flotando y con movimiento lateral.
Mucigel: Un material mucoso segregado por los ápices de las raíces y los pelos radicales.
Mucilaginoso: Que contiene un mucilago, compuesto de mucopolisacáridos.
Ir inicio glosario
Nucleólo: Estructura nuclear especializada, con regiones densas de ADN asociadas a ciertos cromosomas, lugar de síntesis de los precursores de los ribosomas.

Núcleo: Es el organelo más grande de la célula eucarióte, rodeado por una envoltura nuclear, contiene los cromosomas y mucho del ADN celular.
Ir inicio glosario
Organelo: Partícula subcelular que realiza una función determinada en la célula.
Organizador nucleolar: Un área en ciertos cromosomas, asociada con la formación del nucleolo.
Ósmosis: Un caso especial de difusión de agua, a través de una membrana selectivamente permeable.
Ir inicio glosario
Pared celular: La capa rígida más externa encontrada en las células de las plantas, muchos protistas y algunas bacterias. En las plantas formada principalmente de celulosa.
Pared celular primaria: La pared celular celulósica de todas las células vegetales, depositada durante la mitosis y citocinesis.
Pared celular secundaria: Una pared celulósica, impregnada con lignina, depositada en la parte interna de la pared primaria de muchas especies leñosas.
Pectina: La sustancia cementante encontrada en la lámina media, compuesta principalmente de ácido pectico y pectato de calcio.
Permeabilidad: Una propiedad de la membrana de dejar pasar libremente sustancias.
Peroxisoma: Un microcuerpo celular que contiene las enzimas de la fotorrespiración.
pH. Es el logaritmo negativo de la concentración de iones de hidrógeno. Una escala numérica utilizada para medir la acidez y basicidad de una sustancia.
Plasmodesmos: Cordones microscópicos de citoplasma, que atraviesan la pared celular y ponen en comunicación los citoplasmas de dos células contiguas.
Plasmólisis: La pérdida osmótica de agua del citoplasma y la vacuola, causa que el citoplasma se separe de la pared celular.
Plastidios: Organelos de la célula vegetal, entre los cuales están los cloroplastos, cromoplastos y leucoplastos.
Polímero: Una macromolécula formada por la unión de pequeñas moléculas (monómeros) idénticas, p ej. celulosa, almidón.
Polisoma: Un grupo de ribosomas relacionados funcionalmente y unidos por un cordón de ARNm.
Poros nucleares: Regiones perforadas en la envoltura nuclear, a través de las cuales el ARNm migra hacia los ribosomas del citoplasma.
Presión de turgencia: Una presión hidrostática desarrollada en el interior de una célula, como resultado de la ósmosis y que empuja el contenido celular contra la pared celular.
Procariotes: Organismos cuyas células no contienen núcleo, incluyen a las bacterias y cianobacterias.
Profase: Es la primera etapa de la división nuclear, se caracteriza por la desaparición de la envoltura nuclear y la aparición de cromosomas acortados.

Proplastidios: Partículas rodeadas por membrana, que desarrollan una estructura interna; pueden dar origen a cloroplastos, cromoplastos o leucoplastos.
Proteína: Una macromolécula compuesta por una secuencia linear de aminoácidos. Contienen C,H,O,N, S. Las proteínas son los principales componentes estructurales de las células.
Proteínas integrales: Proteínas que penetran en la bicapa lípidica de las membranas celulares o que la atraviesan por completo.
Proteínas periféricas: Proteínas asociadas con la superficie de las membranas biológicas.
Protoplasto: La porción viva de la célula. Se excluye la pared celular.
Ir inicio glosario
Respiración: Es el proceso mediante el cual se convierte la energía de la glucosa en ATP, ocurre en las células de todos los organismos vivos y libera CO2 como un subproducto. La respiración aeróbica requiere la presencia de O2 ; aunque algunos organismos pueden respirar anaeróbicamente.
Retículo endoplasmático: Una red de membranas aplanadas que recorren el citoplasma celular ; si tienen ribosomas adheridos se denomina retículo endoplasmático rugoso; si no se encuentran presentes ribosomas, la membrana se denomina retículo endoplasmático liso.
Ribosa: Un azucar de cinco carbonos importante en el ARN y otros compuestos.
Ribosoma: El organelo celular responsable de la traducción de la síntesis de proteínas
RUDP: Ribulosa 1,5 difosfato, el azúcar de 5 carbonos que se combina con el CO2 en el ciclo de Calvin, de la fotosíntesis.
RUDP carboxilasa: La enzima que fija el CO2 en el ciclo de Calvin de la fotosíntesis.
Ir inicio glosario
Simplástico: Movimiento de agua y sóluto a través de los tejidos atravesando las membranas biológicas
Sistema transportador de electrones: El transporte de electrones excitados a través de una serie de moléculas o transportadores, resultando en la síntesis de ATP.
Ir inicio glosario
Taninos: Compuestos orgánicos de origen vegetal, contienen C,H,O, de sabor astringente. Se usan industrialmente por sus propiedades curtientes en la industria del cuero. Le sirven a la planta como defensa contra el ataque de parásitos o la acción de animales fitófagos.
Tilacoides: Pilas de sacos membranosos aplanados, que forman las granas en el interior del cloroplasto.
Tejido: Un grupo de células similares en origen y estructura, que realizán una función particular.
Tonoplasto: La membrana que rodea la vacuola.
Totipotencia: Este término implica, que todas las células somáticas de un organismo tienen la información genética para completar todo su ciclo vital, o sea pueden formar una nueva planta, si el medio ambiente es adecuado. Esto significa que todas las células tienen un complemento completo de ADN, que puede dar origen a una planta completa, p. ej. Una célula parenquimática de una raíz de zanahoria, puede originar un embrión que se diferencia en una planta adulta de zanahoria. ! La oveja Dolly !

Traducción: Conversión de la información contenida en el ARNm en una secuencia específica de aminoácidos, durante la síntesis de una proteína en la superficie de un ribosoma.
Transporte activo: El movimiento de iones o moléculas hacia el interior de una célula, en contra de un gradiente de concentración, usando energía metabólica.
Tubulina: Proteína que compone los microtúbulos.
Turgencia:. Acción y efecto de hincharse, como consecuencia de la absorción de agua por ósmosis.
Ir inicio glosario
Unidad de membrana: La interpretación mediante el uso de microscopio electrónico de las membranas biológicas, que consisten en dos líneas oscuras con una línea clara en el centro. Tiene un espesor de 7, 5 a 10 nm.
Ir inicio glosario
Vacuola: Una inclusión citoplasmática, con un contenido acuoso, rodeada por el tonoplasto y que almacena iones y moléculas de bajo peso molecular.
Vesícula: Un saco pequeño rodeado de una membrana, que se separa de una membrana mediante una constricción como en el aparato de Golgi.
Virus: Una partícula parecida a un cristal, que tiene una cubierta proteica y un núcleo de ADN o de ARN, pero no de ambos.
Ir inicio glosario
Xilema: En las plantas vasculares es el tejido conductor de agua y sales minerales. En varias plantas el xilema está compuesto por vasos, traqueidas, fibras y parenquima.
Ir inicio glosario
Ir al principio
IMAGENES
Célula fijada con KMnO4D:Golgi
pd:Plasmodesmos CW:Pared Célular
N: Núcleo V: Vacuola
Ch: Cloroplasto M:Mitocondria
I: Espacio Intercelular Nu: Nucléolo
Célula fijada con Os O4 Se observa nucléolo

Cloroplastos despues de la
división.
Cloroplasto en división
CW:Pared Célular M:Mitocondria
Cloroplastos con almidón
S: Almidón
Cloroplastos con abundantes granas
G: Grana P: Glóbulo de Grasa

Célula Vegetal. Figura tomada de
Northington & Schneider (1996
CW: Pared CelularPM: Mambrana Plasmática
C: CitosolN: Núcleo
Ch: CloroplastoM: MitocondriaRER: Retículo
endoplasmático rugoso.G: Aparato de Golgi
V: Vacuola
Núcleo.Figura tomada de Rawn(1989).
Figura tomada de Rawn (1989)

Las Microfibrillas de celulosa en la pared celular estan conpuestas de cadenas crsitalinas de ß-1,4 cadenas de glucanos, mantenidas unidas por enlaces de hidrógeno.
Figura tomada de Taiz & Zeiger (1991).
La membrana plasmática de las células vegetales consiste de proteínas inmersas en una bicapa de
lípidos.Figura tomada de Northington & Schneider (1996)

CONTENIDO
/ INTRODUCCIÓN / ACTIVADORES METÁLICOS / NOMENCLATURA / CLASIFICACIÓN DE LAS ENZIMAS / / CATALISIS MOLECULAR / EFECTO DE LA CONCENTRACIÓN DEL SUBSTRATO SOBRE LA CATALISIS ENZIMATICA /
/ REPRESENTACIÓN GRÁFICA DE LINEWEAVER Y BURK / EFECTO DEL pH SOBRE LA ACTIVIDAD ENZIMATICA / EFECTO DE LA TEMPERATURA / EFECTO DE ACTIVADORES /
/ ESPECIFICIDAD ABSOLUTA / ESPECIFICIDAD RELATIVA / INHIBICIÓN ENZIMATICA /
PÁGINA PRINCIPAL
INTRODUCCIÓN
Las enzimas son biocatalizadores de naturaleza proteica. Todas las reacciones químicas del metabolismo celular se realizan gracias a la acción de catalizadores o enzimas. La sustancia sobre la
que actúa una enzima se denomina substrato. Pasteur descubrió que la fermentación del azúcar mediante levaduras, con su conversión en alcohol etílico y anhídrido carbónico es catalizada por
fermentos o enzimas. En 1897 Buchner logró extraer de las células de levadura las enzimas que catalizan la fermentación alcohólica. Sumner en 1926, aisló en forma cristalina la enzima ureasa, a partir de extractos obtenidos de Cannavalia enzyformis (Fabaceae) la que hidroliza la urea según la siguiente
reacción: UREASA
(NH2)2 CO + H2O CO2 + 2 NH3
En 1930, Northrop aisló en forma cristalina las enzimas digestivas: pepsina, tripsina y quimotripsina. En la actualidad se conocen más de 2000 enzimas que han sido aisladas en forma cristalina.
En términos generales los catalizadores se caracterizan por las siguientes propiedades:
1º Son eficaces en pequeñas cantidades. Tienen un número de recambio alto, que varia entre 100 y 36 millones (anhidrasa carbónica). El número de recambio o actividad molar, se define como la
cantidad de substrato transformado en la unidad de tiempo por una cantidad dada de enzima, por ej. la catalasa hidroliza 5,6 * 106 moléculas de H2 O2 por molécula de enzima por minuto, por lo que
su número de recambio es 5,6 * 106 .
2º No se alteran durante las reacciones en que participan.
3º Aceleran el proceso para la obtención del equilibrio de una reacción reversible.
4º Muestran especificidad. La acción de la enzima es extremadamente selectiva sobre un substrato específico.
Las enzimas tienen pesos moleculares que oscilan entre 12.000 y un millón. Algunas enzimas son proteínas conjugadas; ya que poseen un grupo no proteico o prostético, por Ej. un azucar -glucoproteínas, un lipido -lipoproteinas, un ácido nucléico -nucleoproteinas. Una enzima completa se denomina holoenzima, y está formada por una parte proteica (apoenzima) y un cofactor no proteico (coenzima).

HOLOENZIMA = APOENZIMA + COENZIMA
Entre los cofactores que requieren las enzimas para su funcionamiento están las coenzimas: NADPH+H (nicotinamida adenina dinucleótido fosfato reducido), NAD (nicotinamida adenina dinucleotido), FAD(flavina adenina dinucleótido), piridoxal, biotina, tiamina, ácido tetra hidrofólico , cobalamina, etc. Así mismo, muchas enzimas requieren activadores metálicos, y he de allí la importancia de los minerales para el buen funcionamiento y crecimiento de las plantas.
Ir al principio
ACTIVADORES METÁLICOS
ACTIVADORES METÁLICOS
Elemento Enzima Activada
Zn++ Deshidrogenasas, anhidrasa carbónica, ARN y ADN polimerasas.
Mg++ Fosfohidrolasas, RUBISCO, fosfotransferasas, fosfatasas.
Mn++ Arginasas, peptidasas, quinasas.
Mo Nitratoreductasa, nitrogenasa.
Fe2+, Fe3+ Citocromos, catalasas, ferredoxina, peroxidasas, nitritoreductasa.
Cu2+ Citocromo oxidasa, tirosinasa, ácido ascórbico oxidasa, plastocianina
Ca2+ 1,3 b glucan sintetasa, calmodulina.
K+ Piruvato fosfoquinasa, ATPasa.
Co Vitamina B12 hallada en microorganismos y animales, pero no en plantas. Importante en la fijación simbiótica de nitrógeno.
Ni Ureasa.

Ir al principio
NOMENCLATURA
Antiguamente las enzimas fueron nombradas atendiendo al substrato sobre el que actuaban, añadiéndole el sufijo -asa o haciendo referencia a la reacción catalizada. Así tenemos que la ureasa, cataliza la
hidrólisis de la urea; la amilasa, la hidrólisis del almidón; la lipasa, la hidrólisis de lípidos; la ADNasa, la hidrólisis del ADN; la ATPasa, la hidrólisis del ATP, etc.
Ir al principio
CLASIFICACIÓN DE LAS ENZIMAS
Debido al gran número de enzimas conocidas en la actualidad, se ha adoptado una clasificación y nomenclatura más sistemática, en la que cada enzima tiene un número de clasificación que la identifica.
1. Oxidorreductasas. Reacciones de transferencia de electrones. 2. Transferasas. Transferencia de grupos funcionales. Ej. UDP-glucosa-fructosa-glucotransferasa. 3. Hidrolasas. Reacciones de hidrólisis. Ej. lipasa, proteasa, celulasa. 4. Liasas. Adición a dobles enlaces. Ej. carboxilasa, fenilalanina amonioliasa. 5. Isomerasas. Reacciones de isomerización.Ej. fosfoglucosa isomerasa. 6. Ligasas. Se conocían como sintetasas. Participan en la formación de enlaces con hidrólisis de ATP.
Ir al principio
CATALISIS MOLECULAR
Un catalizador modifica la velocidad de una reacción química sin ser utilizado o aparecer como uno de los productos de la reacción. Una reacción química en la que un substrato(S) se transforma en un producto (P): S P, ocurre por que cierta fracción de moléculas de S, posee mucho más energía que el resto de ellas, lo que es suficiente para que alcancen un estado activado, en el que pueda formarse o romperse un enlace químico y se forme el producto (P).
La energía de activación es la cantidad de energía expresada en calorías, necesaria para que todas las moléculas de un mol, a una temperatura dada alcancen el estado reactivo. Mientras que, el estado de transición es el estado rico en energía de las moléculas que interaccionan en la cima de la barrera de activación. La velocidad de una reacción química es proporcional a la concentración del complejo en el estado de transición.
Una reacción química se puede acelerar de la siguiente forma: 1) Al aumentar la temperatura se incrementa la energía cinética, por lo que es mayor el número de moléculas que alcanzan el estado de transición. Generalmente el Q10 =2,, lo que indica que la velocidad de una reacción química se duplica al

aumentar la temperatura en 100C. 2) Añadiendo un catalizador, que disminuye la energía de activación y aumenta la velocidad de reacción.
La enzima (E), se combina con el substrato (S) formando el complejo de transición, enzima-substrato (E-S), mediante una reacción reversible, cuya energía de activación es menor que la de la reacción no catalizada. Cuando se forma el producto de la reacción (P), se regenera de nuevo la enzima (E) de forma libre, la que puede combinarse de nuevo con otra molécula de substrato (S).
Una enzima reduce más eficientemente la energía de activación (Ea) de una reacción que un catalizador inorgánico, lo que permite que una reacción se realice a menor temperatura.
El siguiente ejemplo ilustra mejor lo que hemos discutido:
Reacción Energía de activación (Kcal*mol-1)
a) el peróxido de hidrógeno se desescompone en:
H2 O2 H2 O + O2 18
b) el hierro catalítico (Fe) realiza la reacción
H2 O2 H2 O + O2 13
c) el platino catalítico (Pt) realiza la reacción
H2 O2 H2 O + O2 12
d) la catalasa una enzima hepática la realiza
H2O2 H2O + O2 5
Una enzima no modifica la energía libre, ni la constante de equilibrio, sino que disminuye la energía de activación de la reacción.

Ir al principio
EFECTO DE LA CONCENTRACIÓN DEL SUBSTRATO SOBRE LA CATALISIS ENZIMATICA
La actividad de una enzima se puede estudiar in vitro, bajo condiciones controladas añadiéndole una enzima a un substrato. Sí se trabaja con la condición de que la concentración del substrato sea saturante, variando entonces la concentración de enzima, se observa que aumenta el producto de la reacción, a pH y temperatura constantes.
Si se mantiene la concentración de la enzima constante y variando la concentración de substrato se obtiene una curva hiperbólica como la de la figura. Al principio un aumento de la concentración de substrato produce un aumento rápido de la velocidad de reacción, pero si se sigue aumentando la concentración de substrato, la velocidad de reacción comienza a disminuir y a muy altas concentraciones de substrato se observa que no cambia la velocidad de reacción, se dice que los centros activos de la enzima se encuentran saturados. La velocidad de reacción que se obtiene a esa alta concentración de substrato se define como la velocidad máxima (V) de la reacción enzimática bajo las condiciones especificadas. La concentración de substrato (S), a la semivelocidad máxima de reacción (V/2) se puede determinar de la figura y representa la constante de Michaelis o Km, la cual es una característica para cada enzima. La inversa de Km, o 1/Km, mide aproximadamente la afinidad de la enzima por el substrato. Mientras más pequeño sea el valor de Km, mayor será la afinidad de la enzima por el substrato. Si varias enzimas compiten en el metabolismo por el mismo substrato, éste será transformado preferentemente por la enzima con mayor afinidad.

La ecuación de Michaelis describe la relación cuantitativa entre la velocidad de reacción y la concentración de substrato [S], si se conoce Vmax o Km:
v=
En donde:
v= es la velocidad de reacción observada a una concentración de substrato determinada [S]
Km= constante de Michaelis en moles/lítro
Vmax= velocidad máxima a concentración saturante de substrato.
Si v= Vmax/2; entonces
=
Km+[S] = 2[S]
Km =[S]
De tal forma que podemos concluir que Km es igual a la concentración de substrato a la semivelocidad máxima de reacción.
Ir al principio
REPRESENTACIÓN GRÁFICA DE LINEWEAVER Y BURK
Si se toma el inverso en ambos miembros de la ecuación de Michaelis tenemos:
= +

que es la ecuación de una línea recta, cuya pendiente es , que intercepta al eje de las ordenadas
en y a la abcisa en el punto Lo que es evidente al hacer , ya que se convierte en
Si se hace una gráfica con los dobles inversos, colocando los valores de en el eje de las ordenadas y en el eje de las abcisas, se obtiene una línea recta a partir de la cual se puede calcular fácilmente Km.
Ir al principio
EFECTO DEL pH SOBRE LA ACTIVIDAD ENZIMATICA
Sabiendo que las enzimas son proteinas, cualquier cambio brusco de pH puede alterar el carácter iónico de los grupos amino y carboxilo en la superficie proteica, afectando así las propiedades catalíticas de una enzima. A pH alto o bajo se puede producir la desnaturalización de la enzima y en consecuencia su inactivación. La fosfatasa ácida es más activa a pH 5,0, mientras que la fosfatasa alcalina lo es a pH 9,0. Muchas enzimas tienen máxima actividad cerca de la neutralidad en un rango de pH de 6 a 8.
El pH per se no afecta la actividad enzimática, sino la concentración de protones. Los protones además de alterar la estructura de la enzima y el substrato, pueden participar también en la reacción como substrato o producto. En esos casos, la concentración de protones afecta directamente la velocidad de la reacción.
ENZIMA pH OPTIMO

Pepsina 1,5
Tripsina 7,7
Catalasa 7,6
Arginasa 9,7
Fumarasa 7,8
Ribonucleasa 7,8
Ir al principio
EFECTO DE LA TEMPERATURA

Un aumento en la temperatura provoca un aumento de la velocidad de reacción hasta cierta temperatura óptima, ya que después de aproximadamente 450 C se comienza a producir la desnaturalización térmica. Las enzimas de muchos mamíferos tienen una temperatura óptima de 370 C, por encima de esa temperatura comienzan a inactivarse y se destruyen Sin embargo existen especies de bacterias y algas que habitan en fuentes de aguas termales y en el otro extremo ciertas bacterias árticas tienen temperaturas óptimas cercanas a 00 C.
Ir al principio
EFECTO DE ACTIVADORES
Ciertas enzimas se encuentran como aproenzimas o zimogenos que son inactivos.
El tripsinógeno es convertido en tripsina por la misma acción de la tripsina, que remueve un hexapéptido. Tripsina Tripsinógeno Tripsina + Val (Asp)4 lisina
Otra forma de activación consiste en mantener los grupos SH reducidos. Estos grupos pueden formar parte de centros activos de ciertas enzimas. Si se oxidan estos grupos a -S-S- la enzima se inactiva. Así tenemos que la enzima papaina, se inactiva después de exponerla al oxígeno; pero cuando se le añade un reductor apropiado la enzima se reactiva. Los grupos -S-S- se convierten en -SH, activándose la enzima.
Otra forma de activación requiere la presencia de otra sustancia que se enlaza al sitio alostérico, el compuesto que se enlaza se denomina un efector alostérico. El centro alostérico es bien específico para el efector o modulador alostérico.
Ir al principio
ESPECIFICIDAD ABSOLUTA
Oxígeno L-aminoácido a -cetoácido + NH3 + H 2 O 2 .
L-aminoácido oxidasa
Oxígeno

D-aminoácido a -cetoácido + NH3 + H 2 O 2 D-aminoácido oxidasa
En este ejemplo se muestra la estereo-especificidad. Una enzima puede tener una especificidad óptica para isomeros D o L.
Hay enzimas que muestran especificidad absoluta como la aspartasa, que cataliza la adición reversible de amoníaco al doble enlace del ácido fumárico, pero no al de otros ácidos insaturados.
HOOC – CH = CH – COO- + NH4 + HOOC – CH2 - CHNH2 - COO-
+ H +
Ir al principio
ESPECIFICIDAD RELATIVA
Ciertas enzimas muestran especificidad relativa, ya que actúan sobre muchos compuestos que poseen un rasgo estructural común. Por ejemplo, la fosfatasa del riñón cataliza la hidrólisis de esteres fosfato del
ácido fosfórico a diferentes velocidades, la amilasa hidroliza el almidón independiente de su origen. Las proteasas y lipasas hidrolizan proteinas y lípidos.
Del estudio de la especificidad del substrato que exhiben las enzimas, surge la idea de una complementaridad, o relación semejante a la de una llave y una cerradura, entre la molécula del substrato y una zona determinada de la superficie de la molécula de la enzima, la cual recibe el nombre de centro activo o centro catalítico.

Ir al principio
INHIBICIÓN ENZIMATICA
Mediante el uso de inhibidores enzimáticos se ha obtenido información muy valiosa sobre la conformación del centro activo de algunas enzimas.
a. Inhibición irreversible: Este tipo de inhibidores forman un enlace covalente con las enzimas cerca del centro activo. Un ejemplo son los gases nerviosos, como el fluorofosfato de diisopropilo (DFP) que forma un complejo con la enzima acetilcolinesterasa. Los animales envenenados con este gas quedan paralizados, debido a la imposibilidad de transmitir adecuadamente los impulsos nerviosos.
Ester di-isopropil fosfórico de la enzima (inactivo)
b) Inhibición competitiva: Es cuando el inhibidor compite con el substrato por la unión con el centro activo de la enzima. Este tipo de inhibición puede reducirse si se aumenta la concentración de substrato.
-OOC - CH2 - CH2 - COO - -OOC - CH = CH - COO- + 2H + Succinato Succinato Deshidrogenasa Fumarato
COO - I
CH2 Malonato (inhibidor competitivo) I
COO - En la inhibición competitiva la enzima se combina con el inhibidor para formar un complejo enzima-
inhibidor:
E + I E I en competencia con la reacción normal E +S ES

La sulfanilamida (bactericida) interfiere con la síntesis del ácido fólico compitiendo con el ácido p-
aminobenzoico, de una forma competitiva.
HOOC-§- NH2 NH2 –SO2 - §- NH2 ácido p- amino benzoico sulfanilamida
§ = fenil
a. Inhibición no competitiva.
Esta inhibición se caracteriza por que no se puede revertir el efecto del inhibidor, aumentando la concentración del substrato.
Por ejemplo la inhibición de la deshidrogenasa del gliceraldehido-3-fosfato por la yodo-acetamida:
Enzima-SH + I-CH2 CONH2 Enzima-SCH2 CONH2 + IH
La deshidrogenasa del gliceraldehido-3 fosfato puede ser inhibida también por el ácido iodo-acético:
Enzima-SH + ICH2 COOH Enzima-SCH2 COOH + IH
c) Inhibición por metales.
Ciertos metales como el plomo, mercurio y arsénico inhiben enzimas que tienen en su centro activo grupos -SH libres
Enlace Recomendado
ENZIMAS: http://www.arrakis.es/~lluengo/biologia.html
CONTENIDO
/ ENERGÍA LIBRE / CAMBIOS EN LA ENERGÍA QUÍMICA STANDARD O TIPO DE LAS REACCIONES QUÍMICAS / REACCIONES ACOPLADAS / LA ECUACIÓN DE NERNST /
PÁGINA PRINCIPAL

ENERGÍA LIBREEl concepto de energía libre, llamada también trabajo neto o útil isotérmico fue propuesto independientemente por Gibbs
y Helmholtz.. Si conocemos los cambios en energía libre a temperatura y presión constante, podemos predecir si una reacción es espontánea o no.
La energía libre es la energía útil, mientras que la entropía es la energía degradada.
El símbolo G se usará para denotar la energía libre, aunque se use también el símbolo F. La definición de D G es:
D G = D H - TD S (1.1)
D G = Al cambio de la energía libre del sistema. D H = A los cambios de una función llamada entalpía o el calor transferido entre el sistema
y el medio que lo rodea.
T = Temperatura absoluta ( t 0 C + 273 0 K ).
D S = Cambios de entropía, es un término que expresa el grado de desorden en un sistema.
D = Se refiere al valor final menos el valor inicial.
Los cambios de entalpía o cambios en el contenido calórico D H se definen de la siguiente manera:
D H = D E + D PV (1.2)
D E = Es el cambio en la energía total del sistema .
P = Presión .
V = Volumen .
En los sistemas biológicos, las reacciones químicas se llevan a cabo en soluciones acuosas diluidas, en los cuales T, P y V son constantes. Bajo estas condiciones el término D PV en la ecuación (1.2) es igual a
cero y D H = D E. Sustituyendo en la ecuación (1.1) tenemos que:
D G = D E - T D S o D E = D G + T D S.
El cambio de energía total del sistema D E es la suma algebraica de TD S, que es siempre positivo en cualquier proceso real y D G que es siempre negativo en cualquier proceso real .
Los cambios en la energía libre se pueden definir como aquella parte de los cambios en la
energía total que se utilizan para realizar un trabajo, a medida que el sistema procede hacia la condición de equilibrio a temperatura y presión constante. A medida que el sistema se acerca
al equilibrio DG decrece al mínimo y se hace igual a cero.
Ir al principio
CAMBIOS EN LA ENERGÍA QUÍMICA STANDARD O TIPO DE LAS REACCIONES QUÍMICAS

Para una reacción reversible:
a A + b B c C + d D (2.1)
En donde a,b,c y d son número de moles de A,B,C y D que participan en la reacción. Existe una constante que expresa el equilibrio químico alcanzado por el sistema, la llamada constante de equilibrio termodinámica que se puede escribir de la manera siguiente:
(2.2)
(A), (B), (C) y (D) son las concentraciones molares de A, B, C y D. La constante de equilibrio debe ser una función matemática de los cambios de energía libre de los componentes de la reacción. El cambio de energía libre está dado por la ecuación:
D G = D Gº + RT ln K (2.3)
K = Constante de equilibrio. R = la constante de los gases 1.987 cal *mol-1 * 0K-1
T = Temperatura absoluta . D Gº = Cambios en energía libre o standard.
Cuando se alcanza la condición de equilibrio D G = 0 , entonces la ecuación (2.3) se convierte en :
0 = D Gº + RT ln K (2.4)
D G º = - RT ln K (2.5)
La relación que existe entre la constante de equilibrio de una reacción y la energía libre standard calculada a 25º C y a pH 7.0 se presenta en la siguiente tabla:
K ln K DG' cal/mol
0.001 -6.9 4.085.6
0.01 -4.6 2.751.4
0.1 -2.3 1.375.6
1 0 0
10 +2.3 -1.375.6
100 +4.6 -2.751.4
1000 +6.9 -4.085.6
Podemos entonces observar del cuadro de arriba que cuando la constante de equilibrio es alta, los cambios en energía libre son negativos (reacción espontánea). Cuando la constante de equilibrio es baja, los cambios en la energía libre son

positivos, lo que indica que hay que suministrar energía al sistema para transformar un mol de reaccionante en un mol de producto, bajo condiciones donde cada compuesto se encuentre en una concentración 1 molal.
Si la constante de equilibrio es 1.0, la energía libre es cero, lo que corresponde a la condición de equilibrio.
D Gº Se puede calcular a partir de la constante de equilibrio como lo demostraremos con un ejemplo:
E 10- glucosa - 1 -fosfato Glucosa - 6 - Fosfato.
E = La enzima fosfoglucomutasa.
Se comienza la reacción con una concentración de glucosa - 1 - fosfato de 0.020 M , a 25ºC. Cuando se alcanza la condición de equilibrio la concentración de glucosa - 1 - fosfato decrece a 0.001M, mientras que la concentración de glucosa - 6 - fosfato aumenta a 0.019M. Con estos valores calculamos la constante de equilibrio K.
De tal forma que la energía libre:
D G º = - RT ln K
= - 1.987 * 298 * ln 19 =
= - 1761.2 cal / mol.
20- La conversión del ácido fumárico en ácido málico es catalizada por la enzima fumarasa. Bajo la condición de equilibrio a 25ºC se tienen las siguientes concentraciones: ácido málico 0.0801M y de ácido fumárico 0.0159M. Con estos valores calculamos la constante de equilibrio K.
K = (Ácido fumárico) = 0.0801 5.03 = 5.03 (Ácido málico)(H20) 0.0159*1
D G º = - RT ln K
= - 1.987 * 298 * ln 5.03 =
= - 592.13* ln 5.03
= - 956.54 cal / mol

Los cambios en la energía libre estándar de una reacción se pueden calcular también obteniendo la diferencia entre los valores de D G º de las sustancias reaccionantes y los valores de D G º de los productos:
D G º reacción = D G º productos - D G º reaccionantes
30- Calcular la energía libre standard de la siguiente reacción a 25ºC
L - Aspartato - + Fumarato- + NH4+
La energía libre standard de formación (D Gº f) en solución acuosa a 25ºC a una actividad 1M es:
SUSTANCIA D G º f kilocalorías*mol-1
L – aspartato -166.99
Fumarato -144.41
NH4+ -19.00
La energía de formación será:
D G º reacción = D G º productos - D G º reaccionantes
D G º reacción = -163.41 - (-166.99)
= 3.58 kilocalorías * mol-1
Ir al principio
REACCIONES ACOPLADAS
En la célula, la energía liberada o que se hace disponible en una reacción exergónica ( que libera energía), es utilizada para mover otras reacciones endergónicas ( que consumen energía), en otras palabras la energía es utilizada para realizar trabajo. Ejemplos de procesos endergónicos son la síntesis de moléculas complejas a partir de moléculas simples, el transporte de sustancias en contra de un gradiente de concentración, o la elaboración de estructuras celulares a partir de sustancias simples. Podemos decir que la vida se mantiene gracias a procesos endergónicos con el suministro de energía libre. Según la primera ley de la termodinámica, la energía requerida para un proceso endergónico debe ser aportada por un proceso que la suministre. La única forma de que esto pueda ocurrir es mediante sustancias reaccionantes comunes, en un proceso conocido como acoplamiento de reacciones.
Un proceso de acoplamiento energético se puede observar en la figura de abajo:

Como un ejemplo consideraremos las siguientes reacciones:
A B K = 0.1 D G º = +1.375,6 cal * mol-1
B C K = 1.000 D G º = -4.085,6 cal * mol-1
Si sumamos ambas reacciones podemos obtener la constante de equilibrio de todas la reacciones.
A C K = C =100 D G º = -2.751,4 cal * mol-1 A
Lo que nos da una reacción exergónica, ya que la energía libre es negativa.
Estudiemos la conversión de malato.
Fumarasa Malato Fumarato + H2O
D G º = (310 º K) = + 700 cal * mol-1
Esta reacción endergónica, requiere un aporte de + 700 cal * mol-1. Sí introducimos un catalizador que provoque la eliminación del fumarato en una reacción con D Gº
muy negativo, la concentración eficaz del fumarato descenderá con el siguiente desplazamiento de la reacción de la derecha hasta que casi todo el malato haya desaparecido. Una reacción que pueda acoplarse a la conversión endergónica del malato en fumarato es la reacción entre el fumarato y el ión amonio para formar aspartato:
ASPARTASA Fumarato + NH4
+ Aspartato
La aspartasa se encuentra en los extractos de Escherichia coli , y la constante de equilibrio a pH 7.4 y 37 ºC es de 417.

La energía libre de esta reacción se puede calcular ahora:
D G º= - 310 * 1.987 ln 417
= -615,9 ln 417
= - 3.715,7 cal * mol -1
A 37 ºC , D G º(310) para la conversión de malato en fumarato y agua es de +700 cal.
Malato Fumarato + H2O D G º = +700 cal * mol -1 Fumarato + NH4
+ Aspartato DG º = -3.715,7 cal * mol -1 Malato + NH4
+ Aspartato + H2O
D G º formación = -3.715,7 +(+700) = -3.015,7 cal * mol (Reacción Exergónica).
Ir al principio
LA ECUACIÓN DE NERNST
Las reacciones bioquímicas en los organismos vivos son de transferencia de energía. Frecuentemente ocurren acopladas, en lo que se conoce como reacciones de oxido-reducción(redox). La reducción se puede definir como la ganancia de un electrón o de un protón; mientras que la oxidación es la perdida de un electrón o de un átomo de hidrógeno. En una reacción redox, una sustancia es oxidada, y sus electrones se transfieren como una papa caliente a otra sustancia que se reduce. En el metabolismo celular ocurren reacciones redox en la transferencia de electrones acoplada a la síntesis de ATP de la respiración y de la fotosíntesis.
Vamos a estudiar la oxidación del ion ferroso (Fe+2 ) a ion férrico ( Fe+3 ) por perdida de un electrón.:
Fe+2 - e- Fe+3
Así mismo, el ion férrico se reduce a ion ferroso por ganancia de un electrón:
Fe+3 + e- Fe+2
Otro ejemplo lo tenemos en el que un metabolito reducido (AH2 ) se oxida a (A), mediante la reacción redox con NAD (ox) que en el proceso se reduce a NADH2 :
AH2 (reductor) + NAD(oxidante) A(oxidante) + NADH2 (reductor).
La relación que existe entre la energía libre y la energía eléctrica es:
D G º = -N* F* D E o (4.1)
N = Número de electrones transferidos en la reacción de oxido reducción.
E = Potencial eléctrico en voltios
F = El Faradio, una constante de proporcionalidad = 23.060 calorías por voltios
= 96. 487 coulomb * equiv -1
D E0= R*T * ln K Ecuación de Nernst (4.2)

N*F
N * F * D E o = R * T * ln K (4.3)
Pero como D G o = -R * T * ln K (4.4)
Sustituyendo en (4.3) tenemos que: D G º = -N * F * D E o (4.5)
Para los sistemas biológicos es normal expresar el potencial redox (E0 ' ) a pH 7,0 en el cual el potencial de electrodo de
hidrógeno es de - 0,42 voltios.
Como regla general los compuestos reductores transfieren electrones a los oxidantes. Un reductor fuerte tiene un potencial eléctrico en voltios muy negativo, mientras que un oxidante lo tiene positivo.
El potencial de reducción standard de varios sistemas de oxidación - reducción nos permite predecir la dirección del flujo de electrones de un sistema a otro, bajo condiciones standard.
Potenciales eléctricos standard de algunas sustancias de interés Bioquímico (soluciones acuosas a 25ºC y pH 7.0)
REACCIÓN E0
'
Ferrodoxina(ox)/ Ferrodoxina(red)cloroplastos -0,432
H+/ H2 -0.420
NADP + / NADP + H+ -0.324
NAD+ / NADH + H+ -0.320
Piruvato / Lactato -0.180
Acetaldehido / Etanol -0.160
Oxalacetato / L - malato -0.166
Flavoproteina (ox) / Flavoproteina (red) -0.060
Coenzima Q (ox) / Coenzima Q (red) +0.010
Citocromo C (ox) / Citocromo C (red) +0.260
Citocromo Oxidasa (ox) / citocromo oxidasa (red) +0.550
½ O2 / H2O +0.820
En el cuadro de arriba el reductor más fuerte es el par Ferrodoxina(ox)/Ferrodoxina(red) con -0,432 voltios y el más oxidante el par 1/2 O2 / H2O con + O,820 voltios.
Calcular la energía libre estándar de la reacción:

Lactato + Acetaldehido Piruvato + Etanol (reduc) (oxidante)
A pH 7.0 E0' para el sistema piruvato - lactato es de -0.180 voltios. Para el acetaldehido- etanol es de -0.160 voltios , NAD
es la coenzima para los dos sistemas.
D G º = -N * F* D E0' D E0
' = ( E 0
' oxidante - E0' reductor)
N = 2 D E0' = -0.160 - (-0.180) = 0.02 voltios
D G º = -2 * 23.062 * 0.02 = -922,5 cal * mol-1
Calcular el cambio de energía libre standard cuando se mueve un par de electrones de NADH al oxígeno molecular para formar agua a pH 7.0 y 25°C.
E0' de NADH = -0.32 voltios.
E0' O2 = + 0.82 voltios.
D G º ¨= -2 * 23.062 * (0.82 - (-0.32)) = -52.700cal*mol¨¹
= -52,7 Kcal * mol¨¹
El paso de dos electrones del NADH al oxígeno va acompañado por una gran disminución en la energía libre, que pueda hacer posible la síntesis de 7,5 moles de ATP, si se asume una energía libre de hidrólisis para el ATP de 7,0 Kcal*mol -1
( 52,7/7,0 =7,5).
EL ATP COMO FUENTE DE ENERGÍA CELULAR / MOLÉCULA DE ATP / ¿CUÁL ES LA RAZÓN DE QUE EL ATP TENGA ALTA ENERGÍA DE HIDRÓLISIS? / ¿CÓMO SE SINTETIZA EL ATP? / FOSFORILACIÓN FOTOSINTÉTICA /
PÁGINA PRINCIPAL
EL ATP COMO FUENTE DE ENERGÍA CELULAR
El ATP (adenosina trifosfato), químicamente es un nucleótido formado por una base nitrogenada, la molécula de adenina, unida a un azúcar de 5-carbonos, la ribosa y a tres grupos fosfatos. El ATP puede actuar como transportador de energía química, en cientos de reacciones celulares, por lo que se le considera como un compuesto rico en energía; ya que muestra una gran disminución de energía química cuando participa en reacciones hidrolíticas. La energía que se libera cuando se hidroliza el ATP, es utilizada en la síntesis de biomoléculas, en el transporte activo de iones en contra de un gradiente de concentración, en movimientos de ciclosis citoplasmática, en la contracción muscular, en la emisión de luz por bacterias, luciérnagas y en el movimiento de flagelos y cilios. El ATP a nivel celular funciona como una batería, que almacena energía por períodos cortos de tiempo; en otras palabras se puede considerar como la moneda de intercambio de energía de la célula.
La energía liberada cuando se hidroliza enzimáticamente el ATP, convirtiéndose en ADP y Pi, se utiliza para mover reacciones endergónicas (que requieren energía) de biosíntesis en cualquier parte de la célula. Estos procesos juegan una parte vital en establecer orden biológico. La energía liberada es de aproximadamente 7,3 Kcal/mol
Pi=HPO4-2

Ir al principio
MOLÉCULA DE ATP.
El ATP es inestable a ácidos, álcalis y al calor. A pH 7.0 el ATP se encuentra como un anión con cuatro cargas negativas. El fosfato terminal del ATP se puede decir que existe en un estado activado, cuando este fosfato se hidroliza se forma ADP y Pi, dos moléculas de menor contenido energético. El enlace químico que se rompe en esa reacción de hidrólisis se conoce algunas veces como un enlace de alta energía.
Ir al principio

¿CUÁL ES LA RAZÓN DE QUE EL ATP TENGA ALTA ENERGÍA DE HIDRÓLISIS?
Los productos resultantes de la hidrólisis, ADP-3 y HPO4-2 se hallan cargados negativamente, por lo que tienen poca tendencia a aproximarse, debido a la repulsión de sus cargas. Es por eso que son más estables que el ATP.
La molécula de ATP-4 a pH 7.0 tiene cargas negativas muy próximas entre sí, lo que ocasiona una fuerte repulsión de sus cargas eléctricas.
Cuando se hidroliza el grupo fosfato terminal desaparece parte de la tensión creada por la repulsión de las cargas eléctricas.
Los productos de la hidrólisis se estabilizan por resonancia.
Ir al principio
¿CÓMO SE SINTETIZA EL ATP?
El ATP se puede sintetizar a partir de ADP y Pi mediante dos procesos:
Fosforilación a nivel de substrato.
Mecanismo quimiosmótico de Mitchell.
La fosforilación a nivel de substrato ocurre en el citoplasma celular. Por ejemplo, la formación del ácido pirúvico a partir del ácido fosfoenol pirúvico, catalizada por la enzima piruvatoquinasa produce ATP.
Según la teoría quimiosmótica de Mitchell( ganador del premio Nobel en 1978), el sistema transportador de electrones produce un gradiente de protones entre los dos lados de la membrana mitocondrial interna, que crea una diferencia de pH y un potencial de membrana. De acuerdo a esta teoría, los protones son bombeados de la matriz mitocondrial hacia el compartimiento intermembranal, a medida que los electrones del NADH2 se mueven a través de una cadena transportadora de electrones, la cual forma parte de la membrana mitocondrial. Cada par de electrones cruza la membrana tres veces, transportando en cada una de ellas dos protones hacia el espacio intermembranal. Se forma así un gradiente de protones y de potencial eléctrico ( gradiente electroquímico) , que origina un movimiento inverso de protones hacia la matriz, a través de canales de difusión formados por la ATP asa ( ATP sintetiza). A medida que los protones (H+) pasan a través de la ATP sintetiza, la energía libre liberada potencia la síntesis de ATP a partir de ADP y fosfato. Por cada NADH2 que se oxida, se traslocan 6 protones los que al regresar a la matriz generan 3 moléculas de ATP. La síntesis de ATP acoplada al consumo de oxígeno se llama fosforilación oxidativa.

Ir al principio
FOSFORILACIÓN FOTOSINTÉTICA
La síntesis de ATP producida en los cloroplastos, mediante la utilización de energía luminosa, se denomina fotofosforilación:

En la membrana tilacoidal como resultado de la fotólisis del agua ( ) y de la oxidación de la
plastoquinona ( ) se generan protones (H+ ), que originan un alto gradiente de concentración de protones, al ser transportados del lumen tilacoidal hacia el estroma. Ese gradiente de pH a través de la membrana es responsable de la síntesis de ATP, catalizada por la enzima ATP sintetasa.
FIGURA TRANSPORTE DE ELECTRONES EN LA MEMBRANA TILACOIDAL
CONTENIDO
/ INTRODUCCIÓN / COCIENTE RESPIRATORIO / FORMACIÓN DE GLUCOSA A PARTIR DE CARBOHIDRATOS DE RESERVA /GLUCÓLISIS / BALANCE DE ATP EN LA GLUCOLISIS / RESPIRACIÓN AERÓBICA /
/ DESCARBOXILACIÓN OXIDATIVA DEL ÁCIDO PIRÚVICO (C-3) / CADENA TRANSPORTADOR DE ELECTRONES /GRADIENTE DE PROTONES Y FOSFORILACIÓN OXIDATIVA / BALANCE ENERGETICO DE LA RESPIRACIÓN AERÓBICA /
/ CICLO DE LAS PENTOSAS / EL CICLO DEL GLIOXILATO Y LOS GLIOXISOMAS / CICLO DEL GLIOXILATO /
PÁGINA PRINCIPAL

INTRODUCCIÓN
Antes de comenzar el tema, definamos primero el metabolismo como el conjunto de todos los cambios químicos que ocurren en una célula o en un organismo. Los procesos de síntesis como la fotosíntesis o formación de sustancias orgánicas por organismos fotosintéticos, en presencia de luz, a partir del CO2 y H2O pertenece al anabolismo; mientras que la respiración, en la que se forman sustancias simples como el CO2 y H2O, a partir de compuestos orgánicos complejos como la glucosa, pertenece al catabolismo. Cuando el anabolismo y el catabolismo ocurren a la misma velocidad, la planta se encuentra en un estado estacionario, se dice que está latente o en reposo. En esta situación no se reflejan cambios de masa. Pero cuando el anabolismo predomina sobre el catabolismo, la planta crece. En caso de que las reacciones catabólicas excedan las anabólicas, la planta pierde masa y se está muriendo.
La respiración es un proceso catabólico en el que se oxida una molécula combustible, la glucosa, cuya energía es atrapada en forma de ATP (fuente universal de energía).
La respiración puede ser anaeróbica en ausencia del oxígeno (glucólisis) o aeróbica en presencia de oxígeno molecular.
La reacción general de oxidación de la glucosa es:
C6 H 12O6 + 6 O2 6 CO2 + 6H2O + 686 Kcal * mol-1 (1)
Esto significa que 180 g (1 mol) de glucosa es oxidada por 192 g (6moles) de oxígeno, con la formación de 264 g (6moles) de CO2, 108 g (6 moles) de agua y la liberación de 686 Kcal * mol-1 (2872 KJ * mol-1), ésta es una reacción fuertemente exergónica, con una energía libre ( DGo ) negativa, y representa la respiración aeróbica. Esto significa que por cada mol de oxígeno absorbido (32g.), se producen 114 Kcal de energía. De la ecuación (1) se deduce que la respiración se puede medir por la cantidad de oxígeno absorbido o de anhídrido carbónico liberado.
Ir al principio
COCIENTE RESPIRATORIO (CR)
Sí como sustratos respiratorios se oxidan por completo, sacarosa, fructosa, glucosa o almidón, el volúmen de O2
absorbido es igual al volúmen de CO2 liberado, el cociente respiratorio es uno. Las semillas de cereales y de muchas legumbres tienen valores cercanos a uno, cuando se oxidan grasas y aceites durante la respiración, a menudo el cociente respiratorio es 0.70. Por ejemplo cuando se oxida el tripalmitato de glicerol:
2 C51H 98O6 + 145 O2 102 CO2 + 98 H2O
El CR es: 102/145 = 0.70
Si se oxida un sustrato más rico en oxígeno, el CR es mayor de 1, por ejemplo: la oxidación de ácido málico:
C4H 6O5 + 3 O2 4 CO2 + 3 H2O
El CR es: 4/3 = 1,33
El cociente respiratorio de proteínas es cercano a 1,0. Conociendo los valores del cociente respiratorio, se puede obtener información del tipo de sustrato que es usado en la respiración.
Ir al principio
FORMACIÓN DE GLUCOSA APARTIR DE CARBOHIDRATOS DE RESERVA

Aunque la glucosa es el carbohidrato más comúnmente usado como sustrato respiratorio, se deriva del almidón que se almacena en forma de gránulos insolubles en agua, mediante las siguientes reacciones:
Ir al principio
GLUCOLISIS
La glucólisis se localiza en el citosol y consiste en la degradación de glucosa, glucosa – 1 - fosfato o fructosa para formar ácido pirúvico. La glucólisis se puede considerar biológicamente como un proceso primitivo, ya que surgió antes de la aparición de los organelos celulares.
La glucólisis consiste de dos fases. En la primera fase se transforma la glucosa (C6) originando dos moléculas de tres carbonos (C3); en la segunda se oxidan las moléculas C3 transformándose en dos moléculas de ácido pirúvico, ATP y NADH + H+
Glucosa + 2 NAD+ + 2ADP + 2H2 PO 4 - 2 Ác pirúvico + NADH + 2H+ + 2 ATP+ 2 H2O
Bajo condiciones anaeróbicas, sin participación del oxígeno molecular, el ácido pirúvico se transforma en alcohol etílico, mediante la fermentación alcohólica:
C6H 12O6 2 CH3 – CO - COOH glucosa ácido pirúvico.
2 CH3 – CO – COOH 2 C2H5 OH + 2 CO2 ácido pirúvico alcohol etílico.
Cuando la glucosa es oxidada a alcohol etílico y anhídrido carbónico, se liberan 52 Kcal * mol-1 (217,71 KJ * mol-1). Muchos organismos pueden fermentar el ácido pirúvico en ácido láctico (fermentación láctica).
C6H 12O6 2 ácido láctico + 52 Kcal * mol-1
En ciertas condiciones los tejidos vegetales pueden estar sujetos a bajas concentraciones de oxígeno (hipoxia) o ausencia total de oxígeno (anoxia). Condiciones que se dan bajo suelos inundados, en los que la concentración de oxígeno es muy baja, dificultando su difusión a las raíces, que entonces operan formando ácido láctico o alcohol etílico.
Podemos resumir la transformación de ácido pirúvico en el proceso glucolítico de la manera siguiente:

La fermentación alcohólica consiste de 11 reacciones enzimáticas, mostradas a continuación y en la Figura 1.
FIGURA 1.

Ir al principio
BALANCE DE ATP EN LA GLUCOLISIS
En la reacción de fosforilación de la glucosa mediante la acción de la glucokinasa, con la formación de la glucosa 6 – P se consume una molécula de ATP; así mismo en la conversión de la fructosa - 6 – P a fructosa 1, 6 di – P, se consume otra molécula de ATP.
En la reacción No. 6 catalizada por la enzima fosfoglicerokinasa, en la que el ácido 1, 3 difosfoglicérico se convierte en Ác.3 – fosfoglicérico se producen dos moléculas de ATP, así mismo se forman otras dos moléculas de ATP en la reacción 9, en la que el ácido fosfoenolpirúvico se transforma en ácido pirúvico, catalizada por la enzima kinasa pirúvica.
El balance neto es: 4ATP producidos menos 2ATP consumidos = 2ATP, si la energía de hidrólisis del ATP varía entre 7 – 8 Kcal * mol-1 (29,3 – 33,5 KJ * mol-1), entonces el balance energético de la glucólisis es de 16 Kcal * mol-1 (71 KJ * mol-1).
La eficiencia de la glucólisis se puede calcular entonces de la siguiente forma: (16/686) *100=2.3%.

La energía almacenada en la glucosa es de 686 Kcal * mol-1, en la glucólisis, se conserva solamente 2,3% de esta energía en forma de ATP, como se calculó anteriormente.
Ir al principio
RESPIRACIÓN AERÓBICA
En presencia de oxígeno, el ácido pirúvico proveniente de la glucólisis, se oxida totalmente en la mitocondria, este proceso se divide en dos fases. En la primera, el ácido pirúvico ingresa a la mitocondria donde es fraccionado y oxidado completamente hasta liberar CO2. Como oxidantes actúan coenzimas , que a su vez son reducidas. El hidrógeno unido a las coenzimas es transferido en la segunda fase al oxígeno molecular, con formación de agua.
La degradación del ácido pirúvico se lleva a cabo en la matriz mitocondrial, donde se encuentran las enzimas del ciclo de Krebs ( Hans Krebs postuló esta vía metabólica en 1937 y posteriormente recibió el premio Nobel por su trabajo) o de los ácidos tricarboxílicos, conocido también como ciclo del ácido cítrico. Los sistemas Red-0x del transporte de electrones se encuentran adosados a las crestas mitocondriales, terminando con la fosforilación oxidativa, la que ocurre tanto en bacterias aeróbicas como en las mitocondrias de células eucarióticas. Una versión resumida de las dos fases la vemos en la figura siguiente:
Ir al principio
DESCARBOXILACIÓN OXIDATIVA DEL ÁCIDO PIRÚVICO (C-3)

Mediante la descarboxilación oxidativa del piruvato se forma acetil coenzima A ( Acetil CoA) o " ácido acético activado", como se muestra en la reacción N0 1 del ciclo del ácido cítrico, catalizada por el complejo enzimático de la deshidrogenasa pirúvica:
Ácido pirúvico + NAD + CoASH Acetil CoA + NADH + H+ + CO2
La función primaria del ciclo del ácido cítrico es oxidar los grupos acetilo de la Acetil CoA, después que reaccionan con el ácido oxalacético, con la formación de ácido cítrico (C-6) ( reacción No 2). Después de una serie de reacciones enzimáticas , dos de los seis carbonos del ácido cítrico se oxidan a CO2 , regenerando por último una molécula de ácido oxalacético (C-4) para repetir de nuevo el ciclo. En cada vuelta del ciclo del ácido cítrico se producen tres moléculas de CO2; por lo que la oxidación de las dos moles de ácido pirúvico formadas en la fase glucolítica de la respiración aeróbica, requiere dos vueltas completas para formar 6 moles de CO2 . El CO2 producido se difunde fuera de la mitocondria o de la bacteria dejando la célula.
Por cada vuelta del ciclo se convierten 2 moléculas de NAD+ en 2 de NADH2 , una de FAD se transforma en FADH2 , una molécula de NADP se convierte en NADPH2 y una molécula de GDP(guanina difosfato) forma GTP(guanina trifosfato). En el citosol se generan como resultado de la vía glucolítica 2 moléculas de NADH2(NADH + H+ ).
La energía almacenada por la transferencia de electrones en esas moléculas , se va atransformar mediante la reacción de la fosforilación oxidativa en ATP ( como se observa en la Figura No 2).
El proceso de la oxidación del NADH2 , se puede representar de la siguiente forma:
NADH + H+ + 0,5 O2 NAD+ + H2O


Ir al principio
CADENA TRANSPORTADORA DE ELECTRONES
Las enzimas que catalizan las reacciones del ciclo de Krebs se hallan en la matriz mitocondrial ; mientras que el sistema transportador de electrones se encuentra inmerso en las crestas mitocondriales. Mediante una serie de reacciones de oxido-reducción, los electrones se transfieren en cascada, ya sea desde el NADH o del FADH2 al oxígeno molecular para que se forme H2O. Parte de la energía del electrón es usada para fabricar ATP y el resto se libera como calor. En la reacción de oxidación del NADH se produce una separación de cargas, los protones (H+) permanecen en la solución acuosa, mientras que los electrones se transfieren a través de transportadores de electrones, que incluyen la ubiquinona y un sistema de citocromos.
Los citocromos son moléculas proteícas que poseen un anillo de porfirina con un átomo de hierro, denominado grupo heme, difieren entre si en su cadena proteíca y en la afinidad por los electrones .Así mismo, los citocromos transportan un solo electrón sin el respectivo protón. Se puede decir que los citocromos pasan "las papas calientes" ( electrones) a lo largo de la cadena transportadora de electrones, mientras que la energía liberada en el proceso es capturada en forma de ATP.
Debido a que cada molécula de citocromo contiene un átomo de hierro, por cada electrón transportado se requiere solamente un citocromo.
La secuencia de los transportadores de electrones en la respiración es:
Los electrones se transfieren desde el NADH al O2 a través de una serie de transportadores de electrones como se ilustra arriba. El complejo de citocromo (a+a3 ) se conoce como citocromo oxidasa; al final se transfieren 4 electrones, que con 4 H+ y una molécula de O2 forman 2 moléculas de H2O:
O2 + 4 H+ + 4 e- 2H2O
El flujo de electrones de la citocromo oxidasa al oxígeno, se puede inhibir con monóxido de carbono (CO ), ión cianuro (CN-) y azida (N-
3 ). La rotenona inhibe el transporte de electrones del NADH a la coenzima Q.
Durante el transporte de electrones a través de la cadena respiratoria(transportadora de electrones) se forma ATP, mediante la fosforilación oxidativa en tres sitios: 1) cuando se transfieren dos electrones del NADH a la NADH deshidrogenasa, 2) cuando se teransfiere un electrón del Cit b al Cit c1 , 3) cuando se transfiere un electrón de la citocromo oxidasa al O2 .
Ir al principio

GRADIENTE DE PROTONES Y FOSFORILACION OXIDATIVA
La hipótesis quimio- osmótica de Peter Mitchell (1961), puede explicar la síntesis de ATP tanto en cloroplastos como en mitocondrias. La cual es un mecanismo de conservación de energía a través de las membranas biológicas, que se basa en la orientación asimétrica de los transportadores de electrones en el interior de la membrana interna de la mitocondria, lo que permite una transferencia de protones ( H+ ) desde la matriz mitocondrial hacia el espacio intermembranal. Como la membrana interna de la mitocondria es impermeable a los protones ( H+ ), se establece un gradiente electroquímico, en el que el espacio intermembranal puede tener un pH de 5,5; mientras que el pH de la matriz, justo en la cara interna de la membrana puede llegar a 8,5, la diferencia ( 8,5 – 5,5 = 3) es de 3 unidades de pH, lo que significa que el espacio intermembranal es 1000 veces más ácido que la matriz, el gradiente de protones, fuerza protonmotriz, representa energía potencial. La única forma de que los protones entren a la matriz, es a través del complejo proteico de la ATP sintetása o ATPasa mitocondrial, sintetizando ATP según la reacción:
ADP + Pi ATP
Este proceso se conoce como fosforilación quimiosmótica o fosforilación oxidativa.

Ir al principio
BALANCE ENERGETICO DE LA RESPIRACIÓN AERÓBICA
A estas alturas estamos preparados para calcular que cantidad de energía, de la presente originalmente en la molécula de glucosa se recupera en forma de ATP. En la vía glucolítica en presencia de oxígeno, se forman directamente 2 ATP, más dos moléculas de NADH2 (de donde se forman 6 moléculas adicionales de ATP), la ganancia total no son 8 ATP como se podría calcular, sino 6; ya que el transporte de los dos electrones que aportan las 2 moléculas de NADH2 a través de la membrana mitocondrial implican un costo de 2 ATP.
La conversión del ácido pirúvico en acetil CoA origina 2 moléculas de NADH2 en el interior de la mitocondria, de tal forma que se originan 6 moléculas de ATP. En el ciclo de Krebs, por cada molécula de glucosa se forman 2 GTP que generan 2 ATP, 6 moléculas de NADH2 ( originan 18 moléculas de ATP) y 2 moléculas de FADH2 ( originan 4 moléculas de ATP), para un total de 24 ATP.
Resumiendo por molécula de glucosa oxidada, el número de moléculas de ATP formados es de 36 . Si la energía de hidrólisis del ATP varía entre 7-8 Kcal * mol –1 (29,3- 33,5 KJ* mol-1), entonces el balance energético de la respiración aeróbica es de 288 Kcal * mol-1 (1205 KJ* mol-1 ) . La eficiencia de la respiración aeróbica se puede calcular de la siguiente forma: 288/ 686 x 100= 42 %. En la respiración aeróbica se conserva aproximadamente el 42 % de la energía de la glucosa en forma de ATP.
Ir al principio
CICLO DE LAS PENTOSAS

Es una vía metabólica alternativa del metabolismo de los carbohidratos presente en tejidos vegetales. Las enzimas del ciclo de las pentosas se encuentran en el citoplasma soluble o citosol. Una de las funciones principales de este ciclo es actuar como una fuente de pentosas. Se conoce también como la vía catabólica de oxidación directa de la glucosa, formada por una serie de reacciones que transforman la glucosa en triosa fosfato y CO2. Por cada molécula de glucosa se produce solamente una molécula de CO2 . Algunas de las reacciones y enzimas de éste ciclo son comunes a la glucólisis. La reacción inicial es la conversión oxidativa de la glucosa-6-fosfato en 6-fosfogluconato, con la formación de NADPH2. La reacción siguiente es la oxidación del 6-fosfogluconato mediante una enzima que requiere NADP con la producción de ribulosa-5-fosfato y CO2. Las reacciones siguientes del ciclo llevan a cabo la conversión de la ribulosa-5-fosfato a otros metabolitos de la glucólisis como son el gliceraldehido-3-fosfato y la fructosa-6-fosfato. En estudios realizados en que se midió la liberación de 14CO2 a partir de glucosa marcada con 14C, se encontró que la vía metabólica dominante del flujo de carbono en muchos tejidos vegetales es la glucólisis representando de 80 a 95 % . Sin embargo la vía de los fosfatos de pentosas realizan su contribución, y ésta es más importante en la medida en que las plantas pasan de un estado meristemático a otro más diferenciado. Los tejidos vegetales enfermos generalmente oxidan la glucosa mediante el ciclo de las pentosas.
EL CICLO DEL GLIOXILATO Y LOS GLIOXISOMAS
Un gran número de semillas poseen como sustancias de reserva grasas o aceites(Tabla 11-1), las que existen principalmente como triglicéridos, en los que los ácidos grasos se encuentran enlazados mediante enlaces esteres a los tres grupos hidroxilos del glicerol. Los aceites son líquidos a temperatura ambiente , debido a la presencia de enlaces insaturados en los ácidos grasos ; mientras que las grasas son sólidas, por tener una alta proporción de ácidos grasos saturados.
Contenido de grasas de algunas semillas
Especie Nombre común % grasa en base al peso seco
Cocos nucifera Coco 65
Ricinus communis Ricino o tartago 60
Helianthus annuus Girasol 50
Linum usitatissimum Lino 35
Glycine max Soya 20
Zea mays Maíz 5
Triticum vulgare Trigo 2
Cuando las semillas de estas plantas germinan, las grasas no se convierten en anhídrido carbónico y agua como sería de esperar, sino que en los tejidos de reserva como son los cotiledones y el endosperma se transforman en sacarosa,

que es transportada al eje embrional. En el eje embrional se quema parcialmente y se incorpora parcialmente a los componentes orgánicos de los tejidos recientemente formados.
En el ciclo del glioxilato el combustible es la Acetil CoA, que se condensa con el oxalacetato para formar isocitrato. En este punto el ácido isocítrico se rompe en una reacción catalizada por la enzima isocitritasa, formando ácido succínico y ácido glioxílico. El glioxilato formado se condensa con otra molécula de Acetil CoA, para formar malato, reacción esta catalizada por la enzima malato sintetasa. El malato así formado se oxida formando oxalacetato, el cual se puede condensar con otra molécula de Acetil CoA, reiniciando el ciclo. Por cada vuelta del ciclo del glioxilato entran dos moléculas de Acetil CoA, y se forma una molécula de succinato que se usa en otros procesos de biosíntesis.
Ir al principio
CICLO DEL GLIOXILATO
Las enzimas del ciclo del glioxilato, principalmente la isocitritasa y la malato sintetasa se encuentran localizadas en unos organelos especiales llamados glioxisomas, que se encuentran rodeados por una membrana. Estos organelos están presentes solamente en células de plantas capaces de convertir grasas en azucares.
Recomendamos visitar: http://www.arrakis.es/~lluengo/biologia.html
CONTENIDO
/ INTRODUCCIÓN / NATURALEZA DE LA LUZ / CLOROFILA Y PIGMENTOS ACCESORIOS / ESTRUCTURA DEL CLOROPLASTO Y DE LAS MEMBRANAS FOTOSINTETICAS / FASES DE LA FOTOSINTESIS / REACCIONES DE LUZ/FOTOSISTEMAS / FOTOFOSFORILACION / EFECTOS DE HERBICIDAS EN EL TRANSPORTE FOTOSINTÉTICO DE LOS ELECTRONES /
REACCIONES DE OSCURIDAD / PLANTAS CON EL CICLO DICARBOXÍLICO / FOTORESPIRACION /PLANTAS CON EL METABOLISMO ÁCIDO DE CRASULÁCEAS (MAC O CAM / CICLO DEL CARBONO
PÁGINA PRINCIPAL
INTRODUCCIÓN
La vida en la tierra depende fundamentalmente de la energía solar, la cual es atrapada mediante el proceso fotosintetico, que es responsable de la producción de toda la materia organica que conocemos. La materia orgánica comprende los alimentos que consumimos diariamente tanto nosotros como los

animales, los combustibles fósiles (petróleo, gas, gasolina, carbón); así como la leña, madera, pulpa para papel, inclusive la materia prima para la fabricación de fibras sintéticas, plásticos, poliester, etc.
La cantidad de carbono fijado por la fotosíntesis es espectácular, como lo demuestran las cifras de la producción anual de materia orgánica seca, estimada en 1,55 x 1011 toneladas, con aproximadamente 60% formada en la tierra, el resto en océanos y aguas continentales.
Los organismos que en el curso de la evolución aprendieron a usar la energía solar y a transformarla en energía química son los llamados autótrofos, que están representados por bacterias y organismos del Reino Vegetal.
Ir al principio
NATURALEZA DE LA LUZ
La luz blanca se separa en diferentes colores(longitudes de ondas) al pasar a través de un prisma. La longitud de onda ( ) se define como la distancia entre dos crestas o dos valles de una onda. La energía es inversamente proporcional a la longitud de onda; las longitudes de onda largas tienen menos energía que las de longitudes de onda cortas. La energía de un fotón se puede calcular con la ecuación:
E= ,. Donde h es la constante de Planck con valor de 6,6262 x 10-34 J.S, C la velocidad de la luz 3,0 x 108 m .S-1 y l la longitud de onda en metros (m). La energía del fotón es inversamente proporcional a la longitud de onda. El ordenamiento de los colores del espectro luminoso, está determinado por las longitudes de onda de la luz. La luz visible es una pequeña parte del espectro electromagnético comprendida entre 390 nm y 770 nm (nanómetro).
DIFINICION Y CARACTERISTICAS DE VARIAS REGIONES DE LONGITUD DE ONDA DE LA LUZ
Color Rango de longitud de onda (nm)
Longitud de onda representativa
Frecuencia (Ciclos/S) o hertzios
Energía(KJ/mol)
Ultravioleta <400 254 11.8 x 1014 471 Violeta 400-425 410 7.31 x 1014 292
Azul 425-490 460 6.52 x 1014 260
Verde 490-560 520 5.77 x 1014 230
Amarillo 560-585 570 5.26 x 1014 210
Anaranjado 585-640 620 4.84 x 1014 193
Rojo 640-740 680 4.41 x 1014 176
Infrarrojo >740 1400 2.14 x 1014 85

Mientras la longitud de onda de la luz visible sea más larga, más rojo es el color y sí la longitud de onda es más corta ésta, estará más cerca del lado violeta del espectro. Las longitudes de onda mayores que las rojas, se conocen como infrarojas y las más cortas que las violetas son ultravioletas.
La luz se comporta como una onda y como una partícula. Las propiedades de onda de la luz incluyen la curvatura de la onda cuando pasa de un medio a otro (Ej. A través de un prisma, el arcoiris, un lápiz introducido en un vaso de agua, etc.). Las propiedades de partícula se demuestran mediante el efecto fotoeléctrico. Por ejemplo cuando un átomo de Zn se expone a la luz ultravioleta, se carga positivamente (Zn+), debido a que la energía luminosa expulsa electrones del Zinc. Estos electrones pueden crear una corriente eléctrica. Los elementos sodio, potasio y selenio tienen una longitud de onda crítica, es la longitud de onda máxima (visible o invisible) que produce un efectro fotoeléctrico. En 1905, Albert Einstein desarrolló una teoría en la que se propuso que la luz estaba compuesta de partículas llamadas fotones, cuya energía es inversamente proporcional a la longitud de onda de la luz. La luz tiene propiedades que se pueden explicar tanto por el modelo de onda como por el de partícula.
Ir al principio
CLOROFILA Y PIGMENTO ACCESORIOS

Un pigmento es cualquier sustancia que absorbe luz. El color de un pigmento es el resultado de la longitud de onda reflejada (no absorbida ). La clorofila, el pigmento verde de todas las células fotosintéticas, absorbe todas las longitudes de onda de la luz visible excepto el verde, el cual es reflejado y percibido por nuestros ojos. Un cuerpo negro absorbe todas las longitudes de onda que recibe. El pigmento blanco o colores claros reflejan todo o casi todas las longitudes de onda. Las sustancias coloreadas tienen su espectro de absorción característico, que es el patrón de absorción de un pigmento dado.
FORMULA ESCTRUCTURAL DE LA CLOROFILA
La clorofila es una molécula compleja, formada por cuatro anillos pirrólicos, un átomo de magnesio y una cadena de fitol larga (C20H39OH).
En las plantas y otros organismos fotosintéticos existen diferentes tipos de clorofilas. La clorofila a se encuentra en todos los organismos fotosintéticos (plantas, ciertos protistas, proclorobacterias y cianobacterias). Los pigmentos accesorios absorben energía que la clorofila es incapaz de absorber. Los pigmentos accesorios incluyen clorofila b (en algas y protistas las clorofilas c,d y e), xantofila(amarilla) y caroteno, anaranjado ( como el beta caroteno, un precursor de la vitamina A ). La clorofila a absorbe las longitudes de ondas violeta, azul, anaranjado- rojizo, rojo y pocas radiaciones de las longitudes de onda intermedias ( verde-amarillo-anaranjado ).

Los pigmentos accesorios actúan como antena, conduciendo la energía que absorben hacia el centro de reacción. Una molécula de clorofila en el centro de reacción puede transferir su excitación como energía útil en reacciones de biosíntesis.
Los carotenoides absorben la longitud de onda azul y un poco en el verde, estos pigmentos tienden a ser rojos, amarillos o anaranjados. La clorofila b absorbe en el azul, y en el rojo y anaranjado del espectro ( con longitudes de ondas largas y baja energía ). La parte media del espectro compuesta por longitudes de onda amarilla y verde es reflejada y el ojo humano la percibe como verde. La distribución de los organismos fotosintéticos en el mar se debe a esto. La longitud de onda corta (más energética ) no penetra más allá de 5 métros de profundidad. La habilidad de absorber parte de la energía de longitud de onda larga (menos penetrante ) debe haber sido una ventaja para las algas fotosintéticas primitivas, que eran incapaces de encontrarse todo el tiempo en la zona superior ( fótica) del mar. Las algas verdes y pardas se instalan en la zona litoral superior, en tanto que en la zona profunda predominan las algas rojas.
Podemos decir que, el espectro de acción de la fotosíntesis es la eficiencia relativa en la generación de una respuesta biológica en función de la longitud de onda, de los diferentes colores, como por ejemplo la liberación de oxígeno. Mediante el estudio de los espectros de acción se descubrió, la existencia de dos fotosistemas en organismos que liberan O2 fotosintéticamente.

Cuando la clorofila absorbe energía luminosa pueden ocurrir tres cosas: l) que la energía sea atrapada y convertida en energía química como en la fotosíntesis, 2) que se disipe como calor, 3) que sea emitida inmediatamente como una longitud de onda mayor con perdida de energía como fluorescencia. La clorofila es capaz de disparar una reacción química cuando se encuentra asociada a proteínas inmersas o embebidas en la membrana de los tilacoides de los cloroplastos, o en las membranas plegadas que se encuentran en organismos procariotes fotosintéticos, como son las cianobacterias y las proclorobacterias.
Ir al principio
ESTRUCTURA DEL CLOROPLASTO Y DE LAS MEBRANAS FOTOSINTETICAS
La unidad estructural de la fotosíntesis es el cloroplasto. Los organismos fotosintéticos procariotes y eucariotes poseen sacos aplanados o vesículas llamadas tilacoides, que contienen los pigmentos fotosintéticos; pero solamente los cloroplastos de los eucariotes están rodeados por una doble membrana. Los tilacoides se disponen como una pila de panquecas, que recibe el nombre de grana. El interior del cloroplasto entre las granas es el estroma proteico, donde se encuentran las enzimas que catalizan la fijación del CO2 . Las mitocondrias constituyen un sistema con dos membranas como los cloroplastos, pero los cloroplastos tienen tres compartimentos: el estroma, el espacio tilacoidal y el espacio entre las membranas. El cloroplasto en su interior tiene un ADN circular y ribosomas.
Ir al principio
FASES DE LA FOTOSINTESIS

La fotosíntesis es un proceso que ocurre en dos fases. La primera fase es un proceso que depende de la luz (reacciones luminosas), requiere la energía directa de la luz que genera los transportadores que son utilizados en la segunda fase. La fase independiente de la luz (reacciones de oscuridad), se realiza cuando los productos de las reacciones de luz son utilizados para formar enlaces covalentes carbono-carbono (C-C), de los carbohidratos. Las reacciones oscuras pueden realizarse en la oscuridad, con la condición de que la fuente de energía (ATP) y el poder reductor (NADPH) formados en la luz se encuentren presentes. Investigaciones recientes sugieren que varias enzimas del ciclo de Calvin, son activadas por la luz mediante la formación de grupos -SH ; de tal forma que el termino reacción de oscuridad no es del todo correcto. Las reacciones de oscuridad se efectúan en el estroma; mientras que las de luz ocurren en los tilacoides.
Ir al principio
REACCIONES DE LUZ
En los procesos que dependen de la luz (reacciones de luz), cuando un fotón es capturado por un pigmento fotosintético, se produce la excitación de un electrón, el cual es elevado desde su estado basal respecto al núcleo a niveles de energía superior, pasando a un estado excitado. Después de una serie de reacciones de oxido-reducción, la energía del electrón se convierte en ATP y NADPH. En el proceso ocurre la fotólisis del agua, la que se descompone según la ecuación:
H2 O + cloroplasto + fotón 0,5 O2 + 2 H+ + 2 electrones.
En la reducción de un mol de CO2 se utilizan 3ATP y 2 NADPH, que a través de una serie de reacciones enzimáticas producen los enlaces C-C de los carbohidratos, en un proceso que se efectúa en la oscuridad.

En las reacciones de oscuridad, el CO2 de la atmósfera (o del agua en organismos fotosintéticos acuáticos/marinos) se captura y reduce por la adición de hidrógeno (H+ ) para la formación de carbohidratos [ ( CH2 O )] . La incorporación del dióxido de carbono en compuestos orgánicos, se conoce como fijación o asimilación del carbono. La energía usada en el proceso proviene de la primera fase de la fotosíntesis. Los seres vivos no pueden utilizar directamente la energía luminosa, sin embargo a través de una serie de reacciones fotoquímicas, la pueden almacenar en la energía de los enlaces C-C de carbohidratos, que se libera luego mediante los procesos respiratorios u otros procesos metabólicos.
Ir al principio
FOTOSISTEMAS
En la fotosíntesis cooperan dos grupos separados de pigmentos o fotosistemas, que se encuentran localizados en los tilacoides. Muchos organismos procariotes solamente tienen el fotosistema I (es el más primitivo desde el punto de vista evolutivo).
Los organismos eucariotes poseen los fotosistemas I y II. El fotosistema I está asociado a las formas de clorofila a, que absorbe a longitudes de onda de 700 nm ( P700 ), mientras que el fotosistema II tiene un centro de reacción que absorbe a una longitud de onda de 680 nm ( P680 ). Cada uno de estos fotosistemas se encuentra asociado a polipeptidos en la membrana tilacoidal y absorben energía luminosa independientemente. En el fotosistema II, se produce la fotólisis del agua y la liberación de oxígeno; sin embargo ambos fotosistemas operan en serie, transportando electrones, a través de una cadena transportadora de electrones. En el fotosistema I se transfieren dos electrones a la molécula de NADP+ y se forma NADPH, en el lado de la membrana tilacoidal que mira hacia el estroma..

Ir al principio
FOTOFOSFOLIRACIÓN
Es la síntesis de ATP que se produce cuando se exponen cloroplastos aislados a la acción de la luz, en presencia de ADP y fosfato. La formación de ATP a partir de la reacción de ADP y fosfato, es el resultado del acoplamiento energético de la fosforilación al proceso de transporte de electrones inducido por la luz, de la misma forma que la fosforilación oxidativa está acoplada al transporte de electrones y al consumo de oxígeno en las mitocondrias.

En el fotosistema I se realiza la síntesis cíclica de ATP, que es independiente de la fotólisis del agua y de la formación de NADPH; mientras que la fotofosforilación no cíclica, está acoplada al transporte de electrones desde el agua, en el fotosistema II a través de una cadena transportadora de electrones hacia el fotosistema I, donde la ferrodoxina cede dos electrones al NADP+ para que se reduzca a NADPH.
H2O + NADP+ + Pi + ADP+ cloroplastos + luz ½ O2 + NADPH + H+ + ATP + H2 O
La molécula de H2 O del lado izquierdo de la ecuación, cede los dos electrones necesarios para la reducción del NADP+ y el átomo de oxígeno que se libera en forma de ½ O2. La molécula de H2O del lado derecho de la ecuación procede de la formación de ATP a partir de la reacción de ADP + Pi.
En la membrana tilacoidal como resultado de la fotólisis del agua y de la oxidación de la plastoquinona ( PQH2 ) se generan protones ( H+ ); que originan un fuerte gradiente de concentración de protones( H+ ) al ser transportados del lumen tilacoidal hacia el estroma. Este gradiente de pH a través de la membrana es responsable de la síntesis de ATP, catalizada por la ATPsintasa (o sintetasa) o conocida tambien como factor de acoplamiento; ya que acopla la síntesis de ATP al transporte de electrones y protones a través de la membrana tilacoidal. La ATPsintasa existe en los tilacoides del estroma y consta de dos partes principales: un tallo denominado CFo, que se extiende desde el lumen de la membrana tilacoidal hasta el estroma y una porción esférica ( cabeza) que se conoce como CF1 y que descansa en el estroma. Esta ATPasa es similar a la de las mitocondrias donde sintetiza ATP.
El flujo cíclico de electrones tiene lugar en algunos eucariotes y bacterias fotosintéticas primitivas. No se produce NADPH , sino ATP solamente. Esto puede ocurrir cuando las células pueden requerir un suministro de ATP adicional, o cuando no se encuentre presente NADP+ para ser reducido a NADPH. En el fotosistema II, el bombeo de iones H+ dentro del tilacoide crea un gradiente electroquímico que culmina con la síntesis de ATP a partir de ADP +Pi.

Las halobacterias, que crecen en agua extremadamente salada, son aerobias facultativas; ya que pueden crecer en ausencia de oxígeno. Los pigmentos púrpuras conocidos como retinal (pigmento encontrado en el ojo humano) funcionan como las clorofilas . La bacteriorodopsina es un complejo formado por retinal y proteínas de la membrana, la que genera electrones que establecen un gradiente de protones que activa una bomba ADP-ATP, que produce ATP en presencia de la luz, pero en ausencia de clorofila. Este comportamiento ayuda a sustentar la universalidad de la teoría quimio-osmótica de Mitchell, en la función de sintetizar ATP.
Ir al principio
EFECTOS DE LOS HERBICIDAS EN EL TRANSPORTE FOTOSINTETICO DE LOS ELECTRONES
Algunos derivados de la urea, como el monurón o CMU (3-p-clorofenil-1,1 di metil urea) y el DCMU [3-(3,4 di cloro fenil) –1,1 dimetil urea] , se aplican al suelo y se desplazan por el xilema hasta las hoja, donde bloquean el transporte de electrones entre las plastoquinonas QA y QB . Inhiben la reacción de Hill en el fotosistema II¸ por lo que no ocurre la fotólisis del agua ni la liberación de O2 .
Ciertos herbicidas a base de triazinas, como la simazina y atrazina bloquean el transporte de electrones entre QA y QB .El maíz y el sorgo son tolerantes a las triazinas, pero no así a los derivados de la urea; ya que contienen enzimas que detoxifican dichos compuestos. Los herbicidas como el diquat y paraquat ( gramoxone), actúan inhibiendo el flujo de electrones entre la ferrodoxina y el NADP, y reduce el oxígeno a un radical superoxido (O2- ), que produce la perdida de la actividad de los cloroplastos.

Ir al principio
REACCIONES DE OSCURIDAD
Las reacciones de fijación o reducción del carbono, son conocidas también como reacciones de oscuridad (son independientes de la luz), sin embargo dos sustancias producidas en la luz, como son el NADPH y el ATP participan en la reducción del CO2.
El CO2 pasa al interior de organismos unicelulares y de otros autótrofos acuáticos por difusión y no a través de estructuras especiales; mientras que las plantas terrestres deben protegerse de la desecación y en ese sentido han desarrollado estructuras llamadas estomas, que permiten el intercambio gaseoso.
En el estroma de los cloroplastos se encuentran presentes las enzimas que intervienen en el Ciclo de Calvin. El Ciclo de Calvin fue estudiado y descubierto en un alga verde unicelular, llamada Chlorella.
El CO2 se combina con la ribulosa 1,5 bifosfato (RUBP- es un azúcar de 5 carbonos ), mediante la acción de la enzima ribulosa bifosfato carboxilasa oxigenasa o rubisco. La rubisco constituye aproximadamente el 50% de las proteínas del cloroplasto y se piensa que es la proteína más abundante en la tierra. El primer producto estable de la fijación de CO2 es el ácido-3-fosfoglicérico ( PGA), un compuesto de 3 carbonos. En el ciclo se fijan 6 moles de CO2 a 6 moles de ribulosa 1,5 bifosfato, y se forman 12 moles de PGA. La energía del ATP, producido en la luz es utilizada para fosforilar el PGA y se forman 12 moles de ácido 1,3 difosfoglicérico, el cual es reducido luego mediante la acción de 12 NADPH a gliceraldehido-3-fosfato( PGAL). Dos moles de gliceraldehido-3-fosfato son removidas del ciclo para fabricar glucosa. El resto de los moles de PGAL se convierten en 6 moles de ribulosa-5-fosfato, que al reaccionar con 6 ATP, regenera 6 moles de ribulosa 1,5 bifosfato, que da comienzo al ciclo de nuevo.
El gliceraldehido-3-fosfato producido en los cloroplastos sirve de intermediario en la glucólisis. Una gran parte del PGAL que permanece en los cloroplastos se transforma en el estroma, en almidón, que es un carbohidrato de reserva. Otra parte del PGAL es exportado al citosol, donde se convierte en fructosa-6-fosfato y glucosa-1-fosfato. La glucosa-1-fosfato se transforma en el nucleótido UDP-glucosa, que al combinarse con la fructosa-6-fosfato forma la sacarosa fosfato, que es el precursor inmediato de la

sacarosa. El disácarido sacarosa es la principal forma en que los azucares se transportan a través del floema, desde las hojas hasta los sitios de la planta donde son requeridos. Es bueno hacer notar que todas las reacciones del Ciclo de Calvin, son catalizadas por enzimas específicas.
Ir al principio
PLANTAS CON EL CICLO DICARBOXÍLICO
Las plantas cuyo primer producto de la fijación de CO2 tiene tres átomos de carbono (C-3), como el ácido-3-fosfoglicérico, poseen el Ciclo de Calvin. Sin embargo, existen otras especies en los que la fijación del CO2 tienen cuatro átomos de carbono (C-4), concretamente ácidos oxalacético, málico y aspártico.Entre las plantas con fotosíntesis C-4, se encuentran la caña de azúcar, el maíz, el sorgo y el amaranto (bledo o alegría).
Las plantas C-3 muestran en general, una anatomía foliar con mesófilo esponjoso en el envés y mesófilo en empalizada en la haz, con tejidos epidermicos en ambas caras y con poros estomáticos para el intercambio gaseoso. Las plantas C-4 se caracterizan por presentar una anatomía en corona o con vaina amilífera, que rodea los conductos o haces vasculares. Los cloroplastos de las células de la vaina son más grandes que los del mesófilo, acumulan mucho almidón y poseen pocas granas o son agranales.
La captura del CO2 en las plantas C-4, comienza con la reacción del CO2 con el ácido fosfoenol pirúvico (PEP), catalizada por la enzima PEP-carboxilasa , con la formación de ácido oxalacético (OAA). El OAA se convierte a ácidos málico o aspártico (C-4), que luego son transportados desde las células del mesófilo, hacia las células de la vaina amilífera. En las células de la vaina el ácido málico (C-4) es descarboxilado, produciéndose CO2 y ácido pirúvico (C-3). Luego el CO2 entra al Ciclo de Calvin y el ácido pirúvico después se convierte en PEP que retorna a las células del mesófilo. Los azucares formados durante este proceso, se transportan por las nervaduras en los conductos del floema a toda la planta.

Ir al principio
FOTORESPIRACIÓN
Una de las propiedades más interesantes de la rubisco es que además de catalizar la carboxilación de la ribulosa 1,5 bifosfato, también produce su oxigenación; proceso conocido como fotorrespiración.
La fotorrespiración da como resultado la liberación de CO2 , después de una serie de reacciones enzimáticas. ¡Es admirable que la rubisco de bacterias anaeróbicas autótrofas, cataliza la reacción de la oxigenasa! La reacción de la carboxilación es favorecida a la oxigenación en una proporción de 3:1; lo que indica un 33% de ineficiencia en la carboxilación. El metabolismo del glicolato requiere la participación de las mitocondrias y de los peroxisomas. Sin embargo, es en las mitocondrias donde el aminoácido glicina, producido en los peroxisomas es descarboxilado liberando CO2 .
El ritmo de la fotorrespiración de las plantas C-3 es bastante elevado, siendo 5 veces superior al de la respiración en la oscuridad; lo cual es perjudicial para estas plantas. Las plantas C-4, que muestran muy poca o ninguna fotorrespiración, son considerablemente más eficientes; ya que realizan la fotosíntesis a concentraciones más bajas de CO2 y a más elevadas tensiones de oxígeno.
Las plantas C-4 son de origen principalmente tropical, habitan en condiciones de alta luminosidad y altas temperaturas. Esto les permite competir más eficientemente con las plantas C-3, al tener que cerrar los estomas para economizar agua y evitar la desecación; sin embargo pueden realizar la fotosíntesis a bajas tensiones de CO2 , debido a que la enzima PEP-carboxilasa muestra una mayor afinidad por el CO2 que la rubisco.
Ir al principio

PLANTAS CON EL METABOLISMO ÁCIDO DE CRASULACEAS (MAC O CAM)
La fotosíntesis es el principal proceso en que el CO2 es fijado por las plantas verdes; sin embargo hay ciertas plantas suculentas y semisuculentas, que fijan el CO2 de noche o en la oscuridad, con un incremento de la ácidez vacuolar, como resultado de la acumulación de ácido málico.Estas plantas pueden fijar el CO2 en la oscuridad, a velocidades superiores de la que lo expulsan mediante la respiración, resultando en una acumulación neta de CO2 .Sí estas plantas se someten a la luz la acidez disminuye. La variación diurna en la acidez fue descubierta en Bryophyllum calycinum, especie perteneciente a la familia Crassulaceae y en consecuencia se denominó "Metabolismo ácido de crasuláceas(MAC o CAM)".
El MAC se encuentra presente en algunos géneros de las Bromeliaceae ( piña, barba de palo), Agavaceae (sisal), Orchidaceae, Cactaceae, Compositae, Amaryllidaceae, Euphorbiaceae y por supuesto en la familia Crassulaceae. Hasta ahora se conocen más de 28 familias con plantas MAC, entre las monocotiledóneas y dicotiledóneas.
Las plantas con MAC habitan en regiones áridas y seca, donde el factor limitante es el agua, por lo que han desarrollado un mecanismo adaptativo, que les ofrece una ventaja ecológica, como es el cierre de los estomas de día y su apertura nocturna. Estas plantas presentan un ritmo circadiano (dura aproximadamente 24 horas), que consta de dos fases: 1) una oscura que produce una acidificación de la vacuola, por acumulación de ácido málico (C-4), con los estomas abiertos, 2) una luminosa en la que ocurre una desadificación, producida por la descarboxilación del ácido málico (C-4) , su conversión en ácido pirúvico (C-3) y CO2 , con los estomas cerrados.
El CO2 producido a partir del ácido málico, se fija en el ciclo de Calvin en la luz con los estomas cerrados. El CO2 se fija en la oscuridad a través de una reacción con PEP(C-3), catalizada por la PEP-carboxilasa. El producto de esta reacción es el ácido oxalacético (OAA, C-4), el cual se reduce a malato(C-4). El PEP viene de la glucólisis, de tal forma que a medida que se forma malato, el almidón disminuye de noche.
El "truco" que emplean las plantas MAC, es que incorporan CO2 de noche, con los estomas abiertos y con el mínimo peligro de desecarse por evapotranspiración; ya que la humedad relativa es más alta y las temperaturas son más bajas. Durante el día, por el contrario, cuando la transpiración es mayor, las plantas MAC cierran los estomas, impidiendo la pérdida de agua.
Reacciones del Metabolismo Acido de Crassuláceas

Ir al principio
CICLO DEL CARBONO
La atmósfera que rodea el globo terráqueo suministra el CO2 a las plantas y el oxígeno a todos los organismos vivos. La atmósfera primitiva contenía grandes cantidades de dioxído de carbono , amonio, y metano, en otras palabras era fuertemente anóxica ( carente de O2 ). Actualmente , los componentes principales de la tropósfera son: 78 vol % nitrógeno, 21 vol % oxígeno, 0,95 vol % gases raros y 0,035 vol % anhídrido carbónico.
Las plantas capturan el dioxído de carbono de la atmósfera y de los océanos, fijándolo en compuestos orgánicos ( son consumidoras de CO2 ). Las plantas producen también CO2 mediante la respiración, el cual es rápidamente usado por la fotosíntesis. Las plantas convierten la energía del sol en energía química, almacenada en los enlaces C-C, de los compuestos orgánicos.
Los animales liberan CO2 , como producto final de la respiración, en la que se degradan carbohidratos sintétizados en la fotosíntesis. El balance entre el CO2 fijado y el CO2 producido es mantenido por la formación de carbonatos en los océanos. Lo que remueve el exceso de CO2 del aire y del agua ( que están en equilibrio en relación al CO2 ). Desde mediados del siglo XVIII, el contenido del CO2 atmosférico ha ido aumentando, primero lentamente , pero desde mediados del siglo XX el incremento ha sido rápido (en promedio de 1,3 µl x L-1 por año ). Durante ese lapso de tiempo se han destruido extensas regiones boscosas tanto en norteamérica, como en las regiones tropicales de la tierra, dando paso a grandes urbes humanas. Así mismo, se han quemado cantidades apreciables de madera, de combustibles fósiles, como el carbón y el petróleo. Las actividades industriales , así como las guerras han destruido enormes cantidades de materia orgánica. Todos estos acontecimientos han reducido las reservas de carbono en la biomasa y el suelo; y han incorporado cantidades excesivas de CO2 a la atmósfera. El dióxido de carbono en la atmósfera, al lado del vapor de agua , metano, ozono y óxido de nitrógeno (N2 O ) ejercen una influencia

negativa en el clima, produciéndo un calentamiento global de la atmósfera, que se conoce como efecto invernadero . Así mismo, como resultado de la actividad humana se han agregado a la atmósfera, hidrocarburos halogenados ( cloro-fluoro-carbonos) y otros gases en pequeñas cantidades, que destruyen la capa de ozono, que proteje a los seres vivos de los efectos dañinos de la radiación ultravioleta.
Como resultado de la combustión de los vehiculos automotores, se liberan a la atmósfera dióxido de azufre, óxidos de nitrógeno y CO2 , que al combinarse con el vapor de agua de la atmósfera, generan ácidos, que al ser lavados por las aguas de lluvia , nieve o niebla producen las lluvias ácidas o precipitaciones ácidas, con valores de pH que están entre 3 y 4. Esta lluvia es causante de grandes daños a los bosques cercanos a áreas industrializadas y de enfermedades cronicas de la vegetación. Los daños antropogénicos a los bosques son el resultado de la actividad contaminante de los seres humanos. La lluvia ácida produce alteraciones en los suelos y en las aguas, afectando la microflora, la macro y microfauna; asi como los procesos de nitrificación y disponibilidad de cationes básicos. Al lado del efecto tóxico de sus componentes químicos, el deposito de lluvia ácida, puede causar efectos directos a los órganos fotosintéticos, tales como necrosis de los bordes foliares, destrucción de la cutícula, y de las ceras cuticulares de las acículas de las coniferas.
Una alternativa que reducidiría la cantidad de anhídrido carbónico atmosférico sería capturando el CO2 al plantar bosques que actúen como sumideros de CO2 reduciendo las concentraciones de éste gas mediante su fijación en la fotosintesis y su conversión en materia orgánica.
El problema del calentamiento global de la atmósfera puede producir que se derritan los casquetes polares de Groenlandia y del polo sur , elevando el nivel del mar a una altura hasta de 120 metros. Los cambios en temperatura y en el nivel de los mares, podrá afectar el clima, alterando la producción de cultivos alimenticios, así como los regímenes de lluvias, ocasionando inundaciones, perdida de vidas humanas, de cultivos agrícolas y dejando grandes masas de población desamparadas y sin hogares.
Ir al principio