cytological comparison different …jb.asm.org/content/78/6/755.full.pdfcytological comparison of...
TRANSCRIPT

CYTOLOGICAL COMPARISON OF SPORES OF DIFFERENT STRAINSOF BACILLUS MEGATERIUM'
PHILIP C. FITZ-JAMES2 AND I. ELIZABETH YOUNG3
Department of Bacteriology and Immunology, and Department of Biochemistry, University ofWestern Ontario, London, Canada
Received for publication May 10, 1959
The content of the phosphorus fractions inbacillus spores reveal a marked difference in thequantity of residue phosphorus (Fitz-James andYoung, 1959). In particular, the spores of twostrains of Bacillus megaterium contained a residuephosphorus of over 1 per cent of the dry weightwhereas in another strain it was essentially nil.This residue fraction remaining after a modifiedSchmidt and Thamnhauer extraction procedure(Fitz-James, 1955) is composed of spore coats,membranes, and some undigested acid-precip-itated protein. Removal of the protein fromthese residues by re-extraction with alkali andof cell wall components by digestion withlysozyme did not alter the phosphorus contentof the residue (Fitz-James, 1955). It thus appearsthat the phosphorus is associated with someinsoluble component of the spore coats. Thisstructural comparison of spores of strains ofB. megaterium revealed that a high content ofresidue phosphorus is associated with the presenceof a heavy outer spore coat and that this extralayer influences the behavior of the spores tostaining procedures.
MATERIALS AND METHODS
The strains of B. megaterium used, includingBacillus 350, have been described (Fitz-Jamesand Young, 1959).
Fixation and embedding for electron microscopy.Spores were suspended overnight in 1 per centosmium (at 2 to 4 C) then held in the fixativefor 1 hr at 37 C. Washed, fixed spores weredehydrated with ethanol and methacrylate, im-pregnated by standard procedures, then workedinto prepolymerized butyl and methyl methacry-lates (10: 1) and polymerized by 4 to 6 hr exposure
1 Supported by grant-in-aid from the NationalResearch Council of Canada.
2 Medical Research Associate, National Re-search Council of Canada.
3 Present address: Department of Bacteriology,University of Alberta, Edmonton, Canada.
to ultraviolet light (Hanovia, Slough, England).When thoroughly hardened, gray to silver coloredsections were cut with glass knives in a Porter-Blum microtome onto either acetone water, 0.5per cent lanthanum nitrate, or 1 per cent phos-photungstic acid. After a variable floating time(5 to 15 min) the flattened sections were pickedup on carbon coated grids, washed with onedrop of distilled water, carefully blotted andexamined in a Philips 100 A electron microscopefitted with a 25 ,u objective aperture and usedat an accelerating voltage of 60 kv.Water washed residues of extracted spores and
carbon replicas of whole spores made by theprocedures described by Bradley and Williams(1957) were shadowed with tungsten oxide beforebeing viewed in the electron microscope.Methods of fixation, hydrolysis, and staining
used for light microscopy are described in detailelsewhere (Robinow, 1951; Fitz-James 1955).
RESULTS AND DISCUSSION
Electron microscopy of shadowed residues. Twotypes of membranes in various stages of disrup-tion could be differentiated in shadowed residuematerial of these spores. It is interesting thatthe electron density of one of these layers, theremnants of the spore coat, varied with thephosphorus content of the residue. The phos-phorus-rich remnants of spores of B. megateriumstrain L (figure 1) were much thicker and moredense than those of strain KM (figure 2). Theselatter were devoid of phosphorus (Fitz-Jamesand Young, 1959). The coats of the Penn strainappeared even more dense than those of strain L.In addition one surface was granular, the otherinterlaced with anastomosing ridges (figures 3aand 3b).The coat residue of the large spores of Bacillus
350 was seen in shadowed preparation, comprisedof disintegrated 5- and 6-sided plaques of rela-tively low density as well as other remnants ofmembranous material. Carbon replicas of these
755
on July 6, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

FITZ-JAMES AND YOUNG
spores revealed the origin of these plaques.Elevations divide the surface of the spore intohexagonal areas, the whole of which is apparentlycovered by a thin loose membrane (figure 4).
Thin-section electron microscopy of spores ofstrains of B. megaterium. More rewarding in-formation regarding the differences in coat struc-ture of these two types of spores was unearthedby thin sections. Robinow (1953) and Mayall andRobinow (1958) have already described thevarious coverings of osmiun fixed spores ofB. megaterium strain L. Such spores possessboth a dense outer coat and a less dense innercoat surrounding a cortex (figures 5 and 6).In contrast, a similar cut of B. megateriumstrain KM (figure 7) shows that the dense outercoat is missing; otherwise the parts of the sporesare quite similar.
Spores of B. megaterium strain Penn alsopossess high density outer coats (figures 8 to10). This coat is equally dense in sections cutonto lanthanum or acetone-water. Moreover,the reoxidation of lower oxides of osmium byfloating sections on peroxide solution (Merriam,1958) did not alter the density of the spore coat.This treatment did, however, remove the smalldark particles seen scattered between the outerand inner coats of figures 5, 6, 8, and 9. Thissuggests that the density of the outer coat isassociated with its composition and not simplya result of the method fixation. If one assumesthat these outer coats account for some 15 percent of the total spore weight, then the concentra-tion of phosphrous in these coats approaches 9per cent and is sufficient to explain the relativelygreater electron density of these coats comparedto that of structures made up of atoms of loweratomic number.From studies of a number of more or less
complete cuts, the outer coats of strains L andPenn appear to be applied to the spore in twohalves which meet at an equatorial ridge likethe hulls of a walnut (figures 5 and 6). The innercoats of strains L and Penn and the single coatof strain KM, on the other hand, encircled thecortex without any apparent major folds. Theappearance of many cuts perpendicular to theequatorial ridge suggested that on one side ofthe spore the outer coat may be open (figure 6)or that the two halves are opposed but notcontinuous over part of the circumference (figure10). The ridge on these two strains of B. mega-terium is probably functionally similar to the
equatorial groove on the surface of the "exine"or outer coat of the spores of Bacillus licheniformisobserved in carbon replicas by Bradley andWilliams (1957) and later shown by Franklin andBradley (1957), to result from a thinning of thecoat.Although it often becomes accentuated during
the embedding process, the space between theouter and inner coats can be quite narrow. It andthe space inside the equatorial ridge appear tocontain a low density material distinguishablefrom the inner coat.
Perpendicular cuts of the spore coats of B.megaterium strain Penn showed a mottled ap-pearance which did not at first suggest theinterlace of strands seen in the spore coat residue(figure 3). Oblique or tangential cuts of thisouter coat, however, revealed a continuous weaveof low density strands, some 100A in width,lying in a background of high density material(figure 1la). Some details of the interlockingbetween the low density strands and the back-ground substance could be detected in a morehighly magnified image (figure llb).
Sections of Bacillus 350 spores revealed thatits coat was single, yet made up of multiplelayers. The outer layer of the coat was responsiblefor the ridging and the covering membrane seenin the replicas appeared to be attached to thepeaks of the ridges (figure 12).By applying the carbon replica technique of
Bradley and Williams (1957) the equatorialridge of the double coated spores could be readilydemonstrated. The single coated spores on theother hand presented either a smooth surfaceor some nonspecific wrinkling (figure 13). In afew of the replicas of spores of the Penn strain,where the ridge was viewed "end on," a finecleavage line could be detected, suggesting ajoining of two halves as already indicated in thethin sections (figure 14).
Effect of acid hydrolysis on spores of differentstrains of B. megaterium. The differences in thecomposition and complexity of the coats en-countered in the spores of different strains ofB. megaterium were also reflected in differences ofresponse to stains and hydrolysis by N HCI(60 C). Spores with the high content of residue-Pin their coats, i. e., the double coated spores,took up considerable surface stain when exposedto crystal violet (figures 15 and 17). Those withthe low residue phosphorus content, the singlecoated spores, were only faintly tinged (figure 16).
756 [VOL. 78
on July 6, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

CYTOLOGICAL COMPARISONS OF SPORES OF B. MEGATERIUM
During acid hydrolysis, resting spores burstand displace their nuclear body through theruptured spore coat (Robinow, 1951; Bisset andHale, 1951; Robinow, 1953; Fitz-James et al.,1954). If it is presumed that the internal pressurerises as a result of an increase in osmotic pressurefrom the breakdown of spore material and thatthe major barrier to this pressure is the sporecoat, then one should be able to demonstrate adifference in response to HCl hydrolysis, de-pending on the number of coats. In keepingwith this, the chromatin of the double-coatedspores (B. megaterium strains L and Penn) wasseldom displaced beyond the outline of the sporesbut was held within the outer coat (figures 18and 20) while that of the single-coated sporesB. megaterium strain KM and Bacillus 350)was found expelled well beyond the cell outline(figures 19 and 21).
Bisset and Hale (1951) likewise found dif-ferent bacilli and clostridia to vary in the extentof their chromatin displacement following HClhydrolysis, but did not relate their results todifferences in spore structure. Hydrolysis withnitric acid-permanganate solutions (Robinow,1951) did not show such a difference, the chro-matin being well extruded in the spores of allfour organisms.
SUMMARY
The alkali and acid insoluble residue phos-phorus fraction of spores of Bacillus megateriumis composed largely of spore coats. Spores ofstrains that are rich in residue phosphorus possesstwo coats, the outer being more electron dense.Spores of a strain (KM) of B. megaterium devoidof phosphorus in its coat residues possessed asingle coat similar to the inner coat of the otherstrains. Carbon replicas emphasized the dif-ferences in surface structure of these spores.The presence or absence of a second coat alteredthe degree of differentiation of resting spores ofB. megaterium to acid hydrolysis.
REFERENCES
BISSET, K. A. AND HALE, C. M. F. 1951 Obser-vations upon the bacterial spore nucleus. J.Hyg., 49, 201-204.
BRADLEY, D. E. AND WILLIAMS, D. J. 1957 Anelectron microscope study of the spores ofsome species of the genus Bacillus usingcarbon replicas. J. Gen. Microbiol., 17, 75.
CAULFIELD, J. B. 1957 Effects of varying thevehicle for OS04 in tissue fixation. J. Bio-phys. Biochem. Cytol., 3, 827-829.
FITZ-JAMES, P. C. 1955 The phosphorus frac-tions of B. cereus and B. megaterium. I. Acomparison of spores and vegetative cells.Can. J. Microbiol., 1, 502-519.
FITZ-JAMES, P. C. AND YOUNG, I. E. 1959 Com-parison of species and varieties of the genusBacillus. Structure and nucleic acid contentof spores. J. Bacteriol., 78, 743-754.
FITZ-JAMES, P. C., RoBINOW, C. F., AND BERGOLD,G. H. 1954 Acid hydrolysis of the spores ofB. cereus. A correlation of chemical andcytological findings. Biochim. et Biophys.Acta, 14, 346-355.
FRANKLIN, J. G. AND BRADLEY, D. E. 1957 Afurther study of the spores of species of thegenus Bacillus in the electron microscopeusing carbon replicas and some preliminaryobservations on Clostridium welchii. J. Appl.Bacteriol., 20, 467-472.
MAYALL, B. H. AND ROBINOW, C. F. 1958 Ob-servations with the electron microscope on theorganization of the cortex of resting andgerminating spores of Bacillus megaterium.J. Appl. Bacteriol., 20, 333-341.
MERRIAM, R.-W. 1958 The contribution of loweroxides of osmium to the density of biologicalspecimens in electronmicroscopy. J. Bio-phys. Biochem. Cytol., 4, 579-582.
ROBINOW, C. F. 1951 Observations on thestructure of Bacillus spores. J. Gen. Micro-biol., 5, 439-457.
ROBINOW, C. F. 1953 Spore structure as re-vealed by thin sections. J. Bacteriol., 66,300-311.
1959] 757
on July 6, 2018 by guesthttp://jb.asm
.org/D
ownloaded from
758 FITZ-JAMES AND YOUNG [VOL. 78
on July 6, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

CYTOLOGICAL COMPARISONS OF SPORES OF B. MEGATERIUM
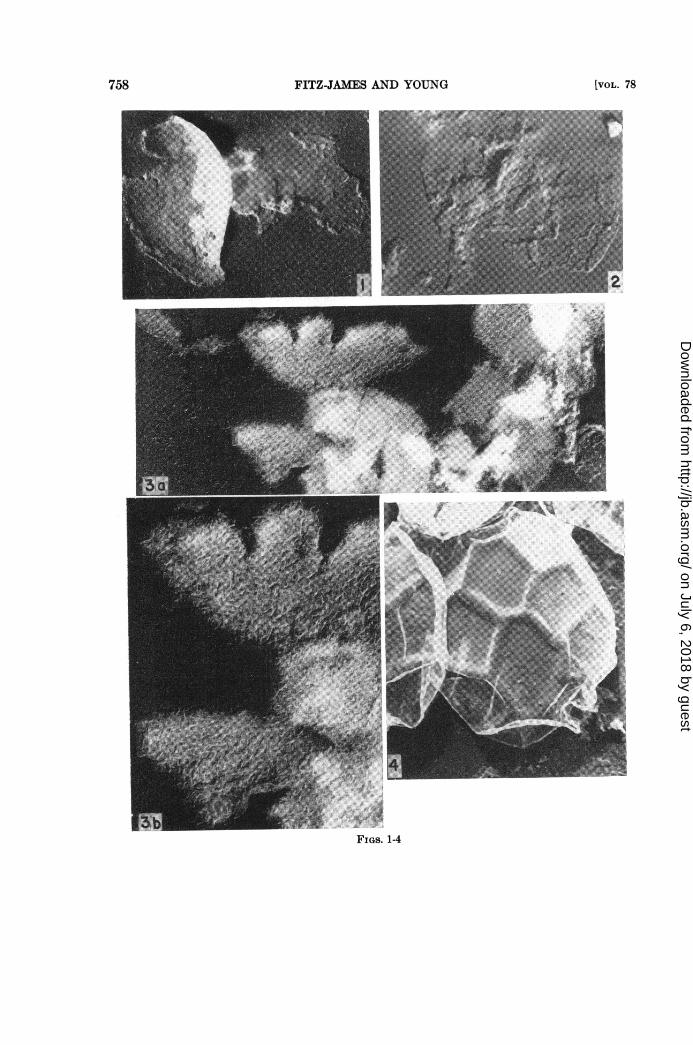
Figures 1 to S. Shadowed electron micrographs of the residues of disrupted spores of Bacillus mega-terium after N NaOH and hot trichloroacetic acid extraction (residue-P fraction). Water-washed,shadowed with tungsten oxide at an angle of 18 degrees.
Figure 1. The high phosphorus-containing residue coats of B. megaterium strain L. The preparationwas composed of relatively dense coats and thinner membranes (X31,000).
Figure 2. The low phosphorus coat residue of B. megaterium strain KM was made up of low densitymembranes much like the thinner component seen in the strain L residues (X23,000).
Figure S. The high phosphorus containing coat residues of B. megaterium strain Penn were, like thoseof strain L, composed of both dense and less dense membranes. The remnants of the dense coats havea woven appearance on one side and granular on the reverse (X23,000). The interlacing weave of thealkali and acid digested coats is shown in figure 3b at higher magnification (X44,000).
Figure 4. A carbon replica of a spore of Bacillus 350 after the method of Bradley and Williams (1957),showing the 5 and 6 sided facets and the ridges. A loose outer membrane appears to cover the sporecoat (X30,000). Shadowed with tungsten oxide at an angle of 22 degrees.
1959] 759
on July 6, 2018 by guesthttp://jb.asm
.org/D
ownloaded from
6
7
8
...9.
FIGS. 5-10
760
on July 6, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

CYTOLOGICAL COMPARISONS OF SPORES OF B. MEGATERIUM
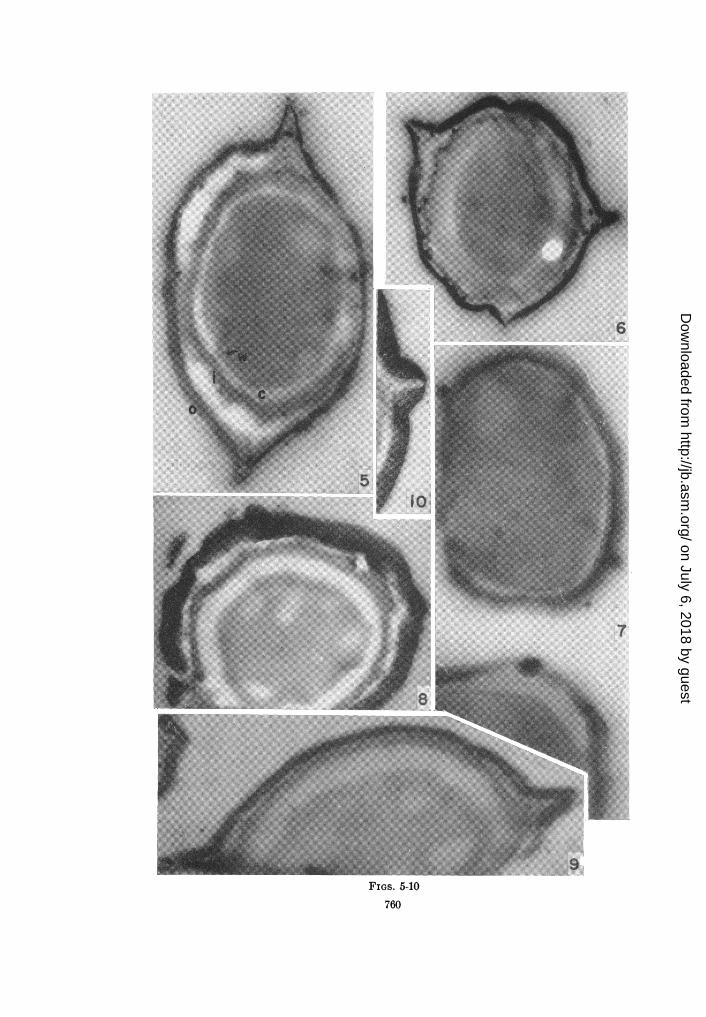
Figures 5 to 10. A comparison of the thin section appearance of resting spores of Bacillus megaterium.Figure 5. A lanthanum treated control section through a spore of B. megaterium strain L showing
the dense outer (o) and less dense inner (i) spore coats, the cortex (c), and the wall (w) of the futurecell. In this and in other sections the low density chromatin structures are seen lying in the more densecytoplasm usually out near the wall. The outer coat is folded into two halves by an equatorial fold(X70,000).Figure 6. A more polar section of a spore of B. megaterium strain L showing heavier lanthanum stain-
ing of the cortex and the high density of the outer coat (X50,000).Figure 7. Portions of two spores of B. megaterium strain KM. The single coat of this strain is similar
in appearance to the inner coat of the Penn and L strains. Lanthanum stained (X66,000).Figures 8, 9, and 10. Sections of B. megaterium strain Penn showing the remarkably dense outer spore
coat and its equatorial fold. Figure 8 was cut on water-acetone (X53,000) and figure 9 onto lanthanumnitrate (X60,000). Figure 10 shows a detail of the equatorial ridge (X72,000).
7611959]
on July 6, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

....._ .t ~~~~~~~~~~~~~ ~ ~ ~~~~~~~~~~~~~~~~~~~~~~~~............_ _-
-. .l l a .- !royrf~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~.....
W g r 1 io .s. EZ Z ... ..............:~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~.. ..... ....
|e:-3;Tt a _SI _ ^ - FIGS. 11-1a#°;ua^ 5 _.ti.p. : - | . !l ~'7i62r-a- -
on July 6, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

CYTOLOGICAL COMPARISONS OF SPORES OF B. MEGATERIUM
Figure lla. Two adjacent spores of Bacillus megaterium strain Penn showing a cross section throughthe outer coat of one and a tangential section through that of the other (X54,000).
Figure lib. The marked area of figure Ila shown in higher magnification (X162,000). The tangentialcut reveals strands in the coat fabric not unlike those encountered in the residue of the alkali-digestedcoats (figure 3b).
Figure 12. A section through one end of a spore of Bacillus 350 (X113,000). We are grateful to Drs.C. F. Robinow and R. G. E. Murray for this electron micrograph.
Figure 13. Carbon replica of the single-coated spores of B. megaterium KM (X30,000).Figure 14. A replica of the double-coated spores of B. megaterium strain Penn showing the prominent
equatorial ridge of the outer coat (X45,000).
1959] 763
on July 6, 2018 by guesthttp://jb.asm
.org/D
ownloaded from
FITZ-JAMES AND YOUNG
16 17
:.4At..: ......
5J-
19
:*.**:
2021 0
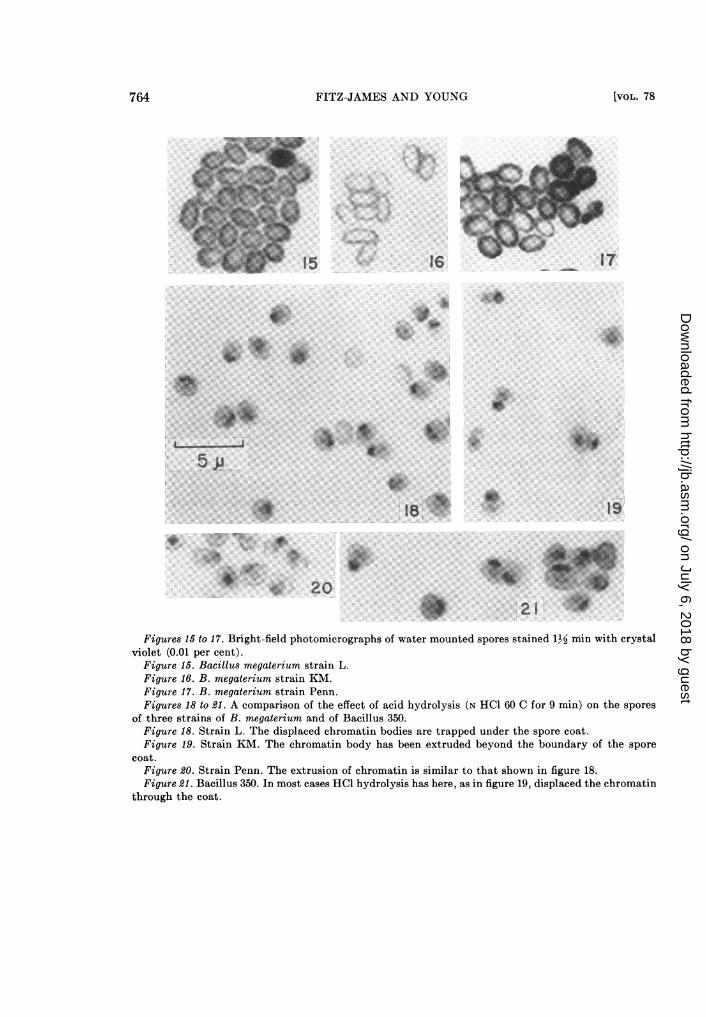
Figures 15 to 17. Bright-field photomicrographs of water mounted spores stained 1½ min with crystalviolet (0.01 per cent).
Figure 15. Bacillus megaterium strain L.Figure 16. B. megaterium strain KM.Figure 17. B. megaterium strain Penn.Figures 18 to 21. A comparison of the effect of acid hydrolysis (N HCl 60 C for 9 min) on the spores
of three strains of B. megaterium and of Bacillus 350.Figure 18. Strain L. The displaced chromatin bodies are trapped under the spore coat.Figure 19. Strain KM. The chromatin body has been extruded beyond the boundary of the spore
coat.Figure 20. Strain Penn. The extrusion of chromatin is similar to that shown in figure 18.Figure 21. Bacillus 350. In most cases HCl hydrolysis has here, as in figure 19, displaced the chromatin
through the coat.
764 [VOL. 78
...
W:* :.j18 .:.....
on July 6, 2018 by guesthttp://jb.asm
.org/D
ownloaded from