dde ett ecccciiÓÓnn nddeell avviirruuss ddell...
TRANSCRIPT

INSTITUTO POLITÉCNICO NACIONAL
ESCUELA SUPERIOR DE MEDICINA
SECCION DE ESTUDIOS DE POSGRADO E INVESTIGACION
“DDEETTEECCCCIIÓÓNN DDEELL VVIIRRUUSS DDEELL OOEESSTTEE DDEELL NNIILLOO EENN AAVVEESS
SSIILLVVEESSTTRREESS MMIIGGRRAATTOORRIIAASS YY RREESSIIDDEENNTTEESS EENN MMEEXXIICCOO..”
TESIS QUE PARA OBTENER EL GRADO DE MAESTRO EN CIENCIAS DE LA SALUD
CON ESPECIALIDAD EN MEDICINA DE CONSERVACIÓN PRESENTA:
M.V.Z. AN T ON IO AC IN I V Á S QU E Z AGU IL AR
DIRECTOR DE TESIS
DR. JOSÉ LEOPOLDO AGUILAR FAISAL
DR. HÉCTOR M. ZEPEDA LÓPEZ
MÉXICO, D. F. DICIEMBRE 2009



Este trabajo se realizó en el Laboratorio de Medicina de Conservación de la Escuela Superior
de Medicina del Instituto Politécnico Nacional bajo la dirección del Dr. Héctor M. Zepeda López
y el Dr. José Leopoldo Aguilar Faisal. Fue apoyado por el proyecto SIP 20091093. El alumno
fue becario del Instituto Politécnico Nacional de Agosto de 2008 Diciembre de 2009 y del
Programa Institucional de Formación de Investigadores del IPN de Agosto de 2008 a Diciembre
de 2009.

Dedicatoria
A mi Padre, Delfino Vásquez H. por enseñarme que nada es imposible, por tener tanta
confianza en mí y por exhortarme siempre a superarme para ser mejor en todos los ámbitos de
la vida. Papá, mis triunfos también son tuyos.
A mi esposa, mí amada Dolores. Por estar a mi lado y darme ese empujón justo en el momento
indicado cuando la desesperación me llevaba al hastío y por todas esas noches que te
desvelaste conmigo. Ahora vas tú. Te amo.
A mi hija, Paula Acic por la alegría de hacerme papá y esperando que te sirva como ejemplo a
seguir y a superar.
A mi hermana Acitlali, como muestra de lo mucho que te quiero y aunque no tengo que ponerte
ningún ejemplo, espero que esto sirva como un impulso para que sigas siempre adelante por el
buen camino que hasta hoy haz recorrido. Sigue siempre así.

Agradecimientos
Al Dr. Héctor M. Zepeda por darme esta oportunidad y por regalarme esas experiencias con la
fauna silvestre que no cualquier mortal tiene la suerte de gozar. Gracias Doc.
Al Dr. José Leopoldo Aguilar Faisal por todos los consejos, por todo ese apoyo, por el tiempo
invertido en esta Tesis y por compartir sus conocimientos.
Al Dr. Jorge Bravo Madrigal por esos consejos tan objetivos justo en el momento indicado y por
ser un ejemplo a seguir.
A todos los docentes que me formaron a lo largo de este postgrado, Dra. Evangelina Muños,
Dr. Asbun Bojail, Dr. Aaron Domínguez, Dr. J. Leopoldo Aguilar F. y Dr. Héctor M. Zepeda,
gracias por sus enseñanzas.
A mi queridísimo papá por todos esos consejos, por ese apoyo incondicional que siempre me
ha dado y que siempre me dará.
A mi mamá por todo ese apoyo que me da y por que a pesar de todo siempre creyó en mí y
creo que a final de cuentas no la defraude. Te quiero ma.
A Amalia, pues estos son triunfos compartidos ¿no? Gracias por todo el apoyo, los consejos,
por creer en mí y por no dejar de exhortarme.
A Dolores, mi novia por siempre, por todo el tiempo y esfuerzo que invirtió en esta tesis en las
tantas revisiones para checar la coherencia de mis párrafos jajaja, y pues aquí si aplica….
gracias totales.
A mi hija, por toda la felicidad que trajo a mí y por ser un complemento más en mi vida para
realizar mejor las cosas.
A mi otra hija, jajaja. A Cobalamina, por ser mi fiel compañía todas las veces que me toco
desvelarme durante el transcurso de esta Maestría.
A mis hermanos Maritza, Ana, Sandra, Víctor y Acitlali, por de una u otra alguna forma me
apoyaron para salir adelante con esta empresa.

A toda la familia Vásquez Heredia, a mi abuelita Amparo, a mi tío Víctor, a mi tía Rosa, a mi tía
Emi, A mi tía Mary y a mi tío Toño, por que todos son ejemplos a seguir y de una u otra manera
me inspiran.
A mis primos Víctor, Olu, Gaby, Yure, Inti, Martha y Emiliano por los buenos momentos que
espero no se acaben y pues haber si les sirvo de ejemplo, bueno a los menores jajajaja.
A mis queridos amigos del Laboratorio de Medicina de Conservación, Arturo (Gibran), Yukie
(Zuzu), Flor y Víctor; realmente logramos crear un equipo interdisciplinario y multidisciplinarlo
jajaja. Gracias por su amistad, sus consejos, esas discusiones tan interesantes e
enriquecedoras y por todas esas veces que me ayudaron para poder sacar adelante este
trabajo.
A mi cuñain Paulo Efraín por todo el apoyo que me a brindado desde que lo conozco,
fundamental para realizar esta Maestría.
A la familia Hernández Rodríguez, Felipe, Luz, Javí, Pandín y Tere, por todo el apoyo que eh
recibido de todos desde que llegue a la familia.
Al Dr. Fernando González por darme ese empujoncito que me faltaba con el cuál llegue a este
laboratorio a realizar me servicio social, y gracias eso acabe en esta magnifica Maestría.
A mi queridísima UAM-X por ese magnifico sistema modular, por inculcarme el autoaprendizaje
y por que gracias a eso soy el Medico Veterinario Zootecnista que soy.

ÍNDICE
I
Índice
Índice I
Índice de figuras II
Índice de cuadros III
Resumen IV
Abstract V
1. Introducción 1
1.1Antecedentes 2
1.1.2 Diseminación del Virus del oeste del Nilo en el continente Americano 4
1.2 Las aves migratorias y el Virus del Oeste del Nilo 6
1.2.1 Virus del Oeste del Nilo en México 10
1.3 Características del Virus 14
1.4 Replicación del Virus del Oeste del Nilo 17
1.5 Ciclo de transmisión 18
1.6 Patogénesis 21
1.7 Signos clínicos 22
1.8 Diagnóstico 24
1.9 El Virus del Oeste del Nilo en Otras Especies 25
2. Justificación 26
3. Objetivos 27
3.1 Objetivo General 27
3.2 Objetivos específicos 27
4. Materiales y métodos 28
4.1 Obtención de muestras 28
4.2 Extracción de RNA 28
4.3 Reacción de la RT-PCR punto final 29
5. Resultados 31
6. Discusiones 38
7. Conclusiones 46
8. Referencias 47

ÍNDICE
II
Índice de figuras
Figura 1. Incidencia de enfermedad neuroinvasiva por Virus del Oeste del Nilo por grupo de Edades – Estados Unidos 2007.
4
Figura 2. Diseminación del Virus del Oeste del Nilo en Estados Unidos.
5
Figura 3. Distribución del Virus del Oeste del Nilo Estados Unidos.
5
Figura 4. Distribución del Virus del Oeste del Nilo en Estados Unidos durante el 2009..
6
Figura 5. Ruta migratoria del Gaviotín Ártico (Sterna paradisaea).
8
Figura 6. Mapa de las rutas migratorias de las aves silvestres.
9
Figura 7. Entidades Federativas positivas al VON en la republica Mexicana.
12
Figura 8. Casos de Virus del Oeste del Nilo en Humanos en México.
13
Figura 9. Representación esquemática de la estructura del Virus del Oeste del Nilo.
14
Figura 10. Árbol Filogenético del virus del Oeste del Nilo.
16
Figura 11. Esquema del ciclo de replicación del Virus del Oeste del Nilo.
17
Figura 12. Esquema del ciclo de amplificación y mecanismos de conservación del Virus del Oeste del Nilo.
19
Figura 13. Número de casos de enfermedad neuroinvasiva por VON en los Estados Unidos durante el 2007.
21
Figura 14. Equipos que se utilizarán para el desarrollo del trabajo.
30
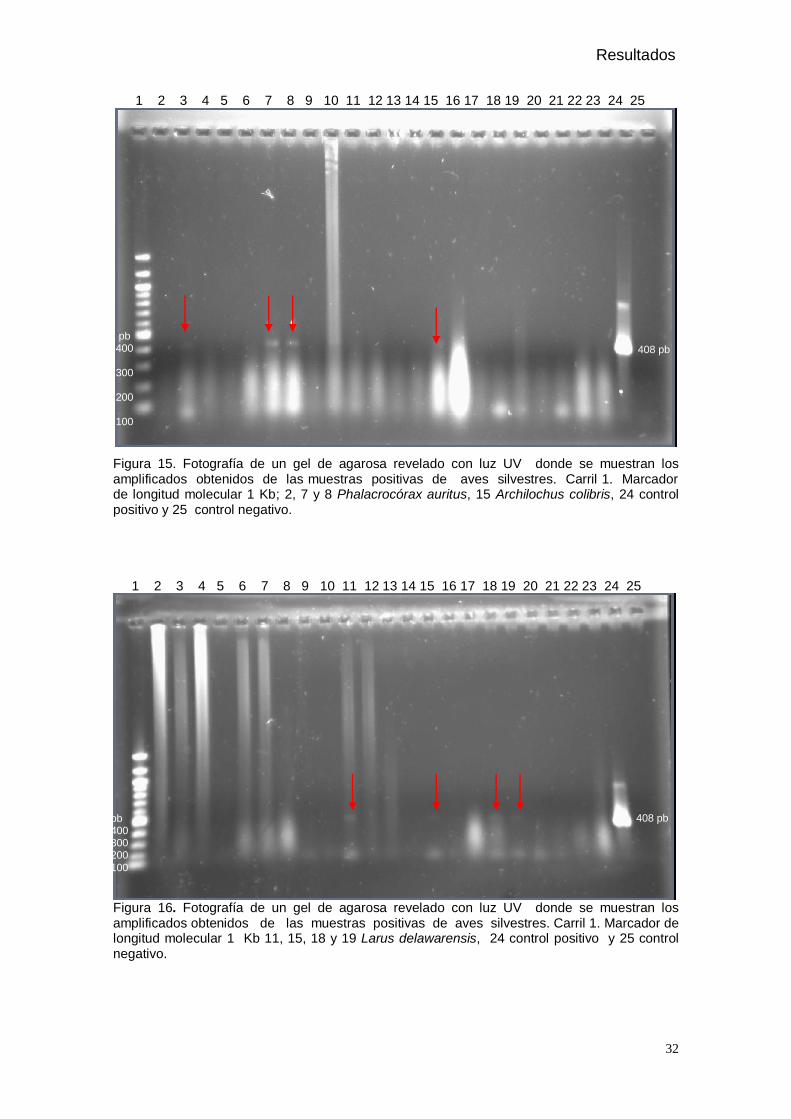
Figura 15. Fotografía de un gel de agarosa revelado con luz UV donde se muestran los amplificados obtenidos de las muestras positivas de aves silvestres.
32
Figura 16. Fotografía de un gel de agarosa revelado con luz UV donde se muestran los amplificados obtenidos de las muestras positivas de aves silvestres.
32
Figura 17. Fotografía de un gel de agarosa revelado con luz UV donde se muestran los amplificados obtenidos de las muestras positivas de aves silvestres.
33
Figura 18. Cormorán bicrestado (Phalacrocórax auritus).
33
Figura 19.Colibrí garganta de rubí (Archilochus colibris).
33
Figura 20. Gaviota pico anillado (Larus delawarensis).
34
Figura 21. Paloma Huilota (Zenaida macroura).
34
Figura 22. Presencia del virus del Oeste del Nilo en la República Mexicana.
36
Figura 23. Área de distribución de la Gaviota pico anillado (Larus delawarensis).
40
Figura 24. Área de distribución de la Paloma huilota (Zenaida macroura).
41
Figura 25. Área de distribución del Cormorán bicrestado (Phalacrocórax auritus).
42
Figura 26. Área de distribución del Colibrí garganta de rubí (Archilochus colibris).
43

ÍNDICE
III
Índice de cuadros
Cuadro 1. Contenido de la mezcla de reacción para Amplificación (RT-PCR). 30
Cuadro 2. Número de muestras obtenidas por estado. 31
Cuadro 3. Especies de aves por Entidad Federativa positivas a VON por RT-PCR 35
Cuadro 4. Prevalencia obtenida de las distintas Entidades Federativas monitoreadas. 37
Cuadro 5. Prevalencia obtenida de las distintas especies de aves monitoreadas. 37

Resumen
IV
RESUMEN
En los últimos años se ha observado la emergencia y reemergencia de algunas zoonosis,
fenómeno relacionado con cambios ecológicos y climáticos, aumento de la población humana
en los trópicos, hacinamiento en las urbes y cambios socioculturales que han determinado que
la población animal comparta su hábitat con el hombre. El Virus del Oeste del Nilo (VON) fue
aislado en 1937 en Uganda. Su ecología fue caracterizada en Egipto en 1950. Pertenece al
género flavivirus donde se ubican virus como el de la Encefalitis Japonesa, Fiebre Amarilla y
Dengue. Se caracteriza por tener tropismo por el sistema nervioso central; y en el caso de
aves, también es cardiotrópico. El VON es transmitido por mosquitos del género Culex y
Aedes, y tiene su ciclo enzootico entre aves y mosquitos. Ha circulado en África, Asia, sur de
Europa y Australia. En septiembre de 1999 el VON se reportó en Nueva York. La introducción
del VON al Continente Americano no se conoce, una hipótesis es que el virus haya entrado a
través de la importación de aves infectadas o se halla introducido por la migración natural de
las aves. En agosto de 2002 se reporto el primer caso de un Mexicano infectado con el VON,
pero se determino que había adquirido la enfermedad en un viaje que realizo a Houston,
Texas. Distintos experimentos han demostrado que las aves son excelentes centinelas ya que
las aves son los hospederos naturales y amplificadores del Virus del Oeste del Nilo por lo que
se utilizan para monitorear la actividad del Virus del Oeste del Nilo en diversas zonas,
realizando de esta forma un diagnóstico epidemiológico oportuno para evitar brotes que
pongan en riesgo la salud humana. El objetivo de este trabajo fue determinar la prevalencia del
VON en aves silvestres residentes y migratorias de México, para conocer la dispersión del
VON en el país, identificar cual es la especie de ave con mayor prevalencia al VON, determinar
cual es el estado con mayor prevalencia al VON e identificar las diferencias en la prevalencia
entre aves residentes y migratorias. Se colectaron muestras cloacales y orales de 200 aves
silvestres migratorias y/o residentes de distintas zonas del país con hisopos estériles con punta
de Dracon, Las muestras fueron congeladas a –20º C y se transportaron al Laboratorio de
Medicina de Conservación de la Escuela Superior de Medicina del Instituto Politécnico Nacional
donde se realizaron las pruebas de laboratorio (Extracción de RNA y RT-PCR). Dieciséis
aves representantes de tres Entidades Federativas y cuatro especies resultaron positivas al
VON: Phalacrocórax auritus, Archilochus colibris, Larus delawarensis y Zenaida macroura. La
prevalencia para el país fue de 8 por cada 100 ejemplares monitoreados, el estado con mayor
prevalencia fue Oaxaca con 33.33 por cada 100 ejemplares monitoreados y las especies con
mayor prevalencia fueron el Phalacrocórax auritus y Archilochus colibris, con 33.33 por cada
100 ejemplares monitoreados. La prevalencia comparada fue 1:1.

Abstract
V
ABSTRACT
In recent years have witnessed the emergence and reemergence of certain zoonoses,
phenomenon related with ecological and climatic changes, increased human population in the
tropics, overcrowding in cities and sociocultural changes that have determined that animal
populations share their habitat with human. The West Nile Virus (WNV) was isolated in 1937 in
Uganda. Its ecology was characterized in Egypt in 1950. Belongs to the genus Flavivirus wich
includes viruses such as Japanese Encephalitis, Yellow Fever and Dengue. It is characterized
by tropism for the central nervous system and for birds, is also cardiotropic. WNV is transmitted
by mosquitoes of the genus Culex and Aedes, and its enzootic cycle is between birds and
mosquitoes. It has circulated in Africa, Asia, southern Europe and Australia. In September 1999,
WNV was reported in New York. The introduction of WNV to the American continent is
unknown, one hypothesis is that the virus has entered through the importation of infected birds
or was introduced by the natural migration of birds. In August 2002 the first case of a Mexican
infected with WNV was reported, but it was determined that it had acquired the disease in a trip
to Houston, Texas. Various experiments have shown that birds are excellent sentinels because
they are the natural hosts and amplifiers of West Nile virus and therefore are used to monitor
the activity of West Nile Virus in various areas, thereby making one appropriate epidemiological
diagnosis in preventing outbreaks that endanger human health. The aim of this study was to
determine the prevalence of WNV in resident and migratory birds in Mexico, to know the
dispersion of WNV in the country, to identify which bird species is most prevalent at WNV,
determine which is the state with the highest prevalence at WNV and identify differences in
prevalence between resident and migratory birds. Cloacal and oral samples were collected
from 200 migratory wild birds and / or residents of different areas of the country with sterile
swabs with Dacron tip, samples were frozen at -20 ° C and were transported to the “Laboratorio
de Medicina de Conservación de la Escuela Superior de Mediciana del Instituto Politécnico
Nacional” where the laboratory tests were conducted (RNA extraction and RT-PCR). Sixteen
birds representing three states and four species tested positive for WNV: Phalacrocorax auritus,
Archilochus colibris, Zenaida macroura and Larus delawerensis. The prevalence for the country
was 8 per 100 birds monitored, the state with the highest prevalence was Oaxaca with 33.33
per 100 birds monitored and the most prevalent species were Phalacrocorax auritus and
Archilochus colibris, with 33.33 per 100 birds monitored. Comparative prevalence was 1:1.

Introducción
1
1. Introducción
La enfermedad es un evento que es parte de la vida; muchas de ellas nos ponen en alerta
cuando los índices de morbilidad y mortalidad aumentan en la población y amenazan el
bienestar, la supervivencia, la productividad y la economía global (García, 2007).
En los últimos años se ha observado la emergencia y reemergencia de algunas zoonosis,
fenómeno estrechamente relacionado a la acelerada industrialización, cambios ecológicos,
cambios climáticos, el aumento de la población humana en los trópicos, el hacinamiento en las
grandes urbes y los cambios socioculturales que han determinado que la población animal
comparta su hábitat con el hombre cada vez con mayor frecuencia (Daszak, et al., 2000;
Barriga et al., 2003; Dabanch, 2003).
El cambio climático puede tener efectos en las zoonosis de dos formas: a) puede aumentar el
rango o abundancia de las aves reservorio y/o de los insectos vectores y b) puede prolongar el
ciclo de transmisión de la enfermedad (Greer et al., 2008).
Los agentes infecciosos involucrados en zoonosis pueden ser transmitidos por distintos
mecanismos, entre ellos: por contacto directo, ingestión, inhalación, por vectores intermediarios
o mordeduras y raramente se transmiten entre los seres humanos, pero algunos agentes
pueden ser transmitidos por transfusión de derivados sanguíneos o trasplante de órganos o
tejidos (Dabanch, 2003).
La importancia de la fauna silvestre y doméstica en las enfermedades zoonóticas, se ha
reportado que de las 175 especies microbianas consideradas patógenas para los humanos, el
75% son zoonóticas, esto es, que se encuentran en los animales como reservorio natural.
Muchos de estos patógenos son transmitidos a los humanos por vectores como los artrópodos
(Tsao et al., 2004), siendo las aves uno de los principales reservorios naturales.
En México las aves acuáticas representan el 25% de la avifauna y el 70.6% de éstas son aves
migratorias de la familia anseriformes y se estima que 3,800,000 aves arriban a nuestro país en
invierno, concentrándose en las lagunas del centro del país (Aguilar, 1998).
Las aves silvestres son importantes en la salud pública por que pueden estar infectadas con un
número importante de microorganismos patógenos que pueden ser transmitidos al humano.
Esto, debido a que las aves migratorias cruzan países y fronteras intercontinentales
transportando con ellas bacterias, virus o parásitos. Esto crea un problema potencial debido a
que se pueden establecer nuevas enfermedades como la infección con Virus del Oeste del Nilo
(VON) a lo largo de las rutas migratorias (Reed, et al., 2002).

Introducción
2
1.1 Antecedentes
El VON fue aislado por primera vez en el año de 1937, en la sangre de una mujer adulta con
un cuadro febril, en la provincia del Oeste del Nilo en Uganda. Su ecología fue caracterizada
por primera vez en Egipto en 1950 (van der Meulen, et al., 2005; O´Donnell y Travis, 2007), y
hay evidencia de la infección por VON en equinos desde 1956 en Egipto y en 1960 en Israel
(van der Meulen, et al., 2005).
La distribución geográfica de este virus se ha reconocido en el viejo mundo incluyendo África,
este de Europa, oeste de Asia y Medio Oriente. Las primeras infecciones del VON se
reportaron en Argelia, Rumania, República Checa, República Democrática del Congo y Rusia
(Barriga et al., 2002; Jawetz et al., 2002; Reed et al., 2003).
El VON es transmitido por mosquitos del género Culex y Aedes, la infección puede causar
mortalidad entre equinos, así como en ciertas aves domésticas y silvestres. Ha circulado
durante mucho tiempo en diferentes partes del mundo, principalmente en África, Asia, sur de
Europa y Australia, siendo el responsable de epidemias importantes, entre las que destacan las
ocurridas en: Israel (en los años 1950’s), Israel (1957), Francia (1962), Rusia (1963), Sudáfrica
(1974), Republica Centro africana (1993), Bielorusia (1977), Ucrania (1985), Rumania (1996 -
1997), Republica Checa (1997), Belgrado (1999), Rusia (1999) e Israel en 2000 (Lanciotti et al,
2000; Rappole y Hubálek, 2003; O´Donnell y Travis, 2007).
Las enzootias por VON en equinos se han reportado en el Reino de Marruecos en 1996 y
2003, en Italia en 1998, en Israel en 2000 y en Francia en 2000, 2003 y 2004 (Hayes et al.,
2005a).
Es importante mencionar que desde 1990, la frecuencia y la severidad de la infección con VON
en humanos se ha incrementado, al igual que el número de reportes de VON en una gran
variedad de vertebrados incluyendo mascotas, animales de granja y animales de vida silvestre.
Argelia fue el primer país en ilustrar este punto, reportando 50 casos en humamos y 8 muertes.
En 1996 se presentó uno de los brotes mas importantes de VON en equinos con 94 casos
reportados y una tasa de mortalidad del 45% en Marruecos (van der Meulen, et al., 2005;
O´Donnell y Travis, 2007).
A finales de agosto y principios de septiembre de 1999, Nueva York y sus alrededores
presentaron un brote de encefalitis humana consistente con una etiología arboviral; al igual que
en humanos se presentó una encefalitis con el mismo agente etiológico en cuervos americanos
(Corvus brachyrhinchos) y cuervos pescadores (Corvus assifragus) en la misma área
geográfica (10 Kilómetros de radio), de este modo el VON hizo un inesperado aviso del
impacto que pueden tener las enfermedades infecciosas sobre la salud pública. El 23 de

Introducción
3
septiembre de 1999 un estudio identificó el Virus del Oeste del Nilo por RT-PCR y se secuenció
el DNA de tejido de aves muertas colectadas en el zoológico del Bronx a principios de
septiembre de ese mismo año (Lanciotti et al, 1999; Rappole y Hubálek, 2003).
Nunca antes se había presentado la enfermedad en América y el resultado inicial fue una
epidemia de 62 casos de encefalitis en humanos, incluyendo 7 muertos (Reed et al. 2003); 27
casos de encefalitis en equinos y miles de aves muertas de 18 especies diferentes incluyendo
a 3000 cuervos americanos (Corvus brachyrhynchos) en el área de Nueva York (Rappole y
Hubálek, 2003).
A la cepa del VON aislada en Nueva York en 1999 se le nombró como WN-NY99 (Gen Bank
AFI96835) y tenia una estrecha relación con una cepa aislada del cerebro de un ganso en
Israel en 1998, por un 99.8% de homología, ya que únicamente se diferenció por dos
nucleótidos (Lanciotti et al, 1999).
Para el 2002, se presentaron 4156 casos con 284 muertes y en 2003, más de 9000 casos y
220 muertes. En 2006 se reportaron 171 muertes con un total de 4261 casos de enfermedad
por VON, de los cuales, 1491 fueron casos de enfermedad neuroinvasiva, aumentando un 14%
con respecto al 2005 en los Estados Unidos (Lindsey et al 2007).
Actualmente la enfermedad se ha distribuido rápidamente y se ha identificado en la mayor
parte de Estados Unidos (Gea- Banacloche et al., 2004), tomando una estacionalidad
endémica (Hayes et al., 2005a) sumando desde su aparición en 1999 más de 23,500 casos en
humanos y mas de 900 muertes (Snapinn et al., 2007; Nielsen et al., 2008), 24,000 casos de
encefalitis equina y miles de aves muertas (Farfán et al., 2006; O´Donnell y Travis, 2007).
En el 2007 la transmisión en humanos se reportó en sitios donde no se había reportado
previamente la presencia del virus y recurrió en 1148 condados de 44 estados de los Estados
Unidos. Se reportaron 117 muertes y 3630 casos de VON de los cuales 1227 (34%) fueron
casos de enfermedad neuroinvasiva, 2350 (65%) fueron casos de fiebre del Oeste del Nilo y 53
(1%) fueron síndromes clínicos inespecíficos. Se reportaron 352 casos de viremia en
donadores sangre, siendo 281 (80%) asintomáticos, 5 (1%) desarrollaron enfermedad
neuroinvasiva y 66 (19%) presentaron fiebre del Oeste del Nilo. La incidencia de la enfermedad
(Figura 1), en los Estados Unidos para el 2007 fue de 0.4 por cada 100,000 habitantes y la
incidencia de la enfermedad neuroinvasiva varió según la edad de los pacientes como se
muestra en la figura 1 (Lindsey et al., 2008).

Introducción
4
Figura 1. Incidencia de enfermedad neuroinvasiva por Virus del Oeste del Nilo por grupo de Edades – Estados Unidos 2007. La gráfica muestra la incidencia por cada 100,000 habitantes y se incluyen dentro de la enfermedad neuroinvasiva la meningitis, la encefalitis y la parálisis flácida aguda (Lindsey et al., 2008).
1.1.1 Diseminación del Virus del oeste del Nilo en el continente
Americano.
La introducción del Virus del Oeste del Nilo al Continente Americano no se conoce con certeza,
una posibilidad es que hayan arribado mosquitos infectados a los Estados Unidos en productos
comerciales o por bioterrorismo. Otra hipótesis, la más aceptada, es que el virus haya entrado
a través de la importación de aves infectadas (Rappole y Hubálek, 2003). Otra posibilidad es
que el VON se haya introducido por la migración natural de la aves, ya que el estallido inicial
esta asociado con el arribo de aves infectadas a los Estados Unidos, lo que puede ocurrir por
las rutas normales de migración trasatlánticas de especies de patos y gaviotas (Lanciotti, et al.,
1999; Reed et al., 2003).
El VON se ha esparcido con mucha rapidez desde su llegada al nuevo continente en agosto de
1999. Durante 1999 el virus se esparció a 150 kilómetros a la redonda del sitio de reporte de
las primeras infecciones (Figura 2A), 300 kilómetros para el año 2000 (Figura 2B), y 500
kilómetros para el 2001 (Figura 2C), lo que demuestra la rapidez de expansión del virus y su
adaptabilidad al nuevo hábitat con un promedio de dispersión de 75 kilómetros por mes
(Rappole y Hubálek, 2003).

Introducción
5
Figura 2. Diseminación del Virus del Oeste del Nilo en Estados Unidos. A. Distribución del
Virus del Oeste del Nilo en 1999. Desde la llegada a Nueva York, el Virus del oeste del Nilo se esparció a 150 kilómetros a la redonda de donde se reportó el brote inicial durante 1999. B. Distribución del Virus del Oeste del Nilo en 2000. Durante el 2000, el Virus del oeste del Nilo se esparció a 300 kilómetros a la redonda del brote inicial en 1999. C. Distribución del Virus del Oeste del Nilo en 2001. Durante el 2001, el Virus del oeste del Nilo se esparció a 500 kilómetros a la redonda del brote inicial en 1999. (Rappole y Hubálek, 2003).
Para 2004, el VON se había esparcido prácticamente por todos los Estados de los Estados
Unidos como se muestra en la figura 3 (Allan, et al., 2007), convirtiéndose en la mayor causa
arboviral de encefalitis en el 2007 (Dai, et al., 2008; Lindsey et al., 2008).
Figura 3. Distribución del Virus del Oeste del Nilo. Se muestra la distribución del VON en
los Estados Unidos y su movimiento de 2002 a 2004 (Allan et al., 2007).

Introducción
6
La principal causa de expansión del VON es el gran número mosquitos ornitofílicos en conjunto
con las aves que distribuyen al virus en sus movimientos migratorios normales. Actualmente el
VON se ha esparcido de costa a costa en la unión americana (figura 4), y el virus se ha
reportado en humanos, equinos, reptiles, mamíferos no humanos y en varias especies de aves.
Figura 4. Distribución del Virus del Oeste del Nilo durante el 2009. Se muestra la distribución del VON en los Estados Unidos; en color guinda se muestran los estados con casos de enfermedad en humanos, y en color lila se muestran los estados con aves, animales o mosquitos infectados (CDC, 2009).
Dentro de los países de América que han reportado al VON se encuentran Cuba, Puerto Rico,
Islas Caimán, Republica Dominicana, Islas Guadalupe, Jamaica, Estados Unidos, El Salvador,
Canadá, Argentina y México (Dupuis et al., 2005; Hayes et al., 2005a Farfán et al., 2006;
Kramer, et al., 2007; Torres, et al., 2008; Díaz, et al., 2008 Lindsey et al., 2008),
1.2 Las aves migratorias y el Virus del Oeste del Nilo
Las aves migratorias han sido durante mucho tiempo sospechosas de la introducción del VON
en nuevas regiones debido a que los nuevos brotes del virus en las regiones con climas
templados generalmente ocurren hacia finales del verano y comienzos del otoño, coincidiendo
con la llegada de grandes concentraciones de aves migratorias. Estos brotes a menudo
ocurren entre humanos que viven cerca de tierras húmedas donde hay altas concentraciones
de aves que entran en contacto con grandes números de mosquitos ornitofílicos. Además, las
aves migratorias han sido vinculadas con el transporte de otros virus relacionados en el

Introducción
7
hemisferio occidental y la migración causa un sustancial stress en las aves el cual puede
promover la inmunosupresión y favorecer la replicación del VON (Rappole et al., 2000).
Según el Centro de Control y Prevención de Enfermedades, CDC por sus siglas en ingles, se
han detectado más de 300 especies de aves silvestres y domésticas infectadas con VON,
(Medica, et al., 2007). Los Passeriformes son el orden más susceptible y desarrollan altos
valores de viremia y altas concentraciones del virus en sus fluidos orales y cloacales; también
presentan signos neurológicos y altas tasas de mortalidad. Otras especies que se ven
afectadas son las del orden Charadriiforme, Strigiformes, Falconiformes y Anseriforme, como
los halcones y los búhos, ya que son muy susceptibles a la infección y a la enfermedad, a
diferencia de los Psittacidos y Gallinaceos que son poco susceptibles (van der Meulen, et al.,
2005; Saito et al., 2007; Ellis et al., 2007,
La migración de las aves consiste en largos desplazamientos que coinciden principalmente con
las épocas invernales de los hemisferios norte y sur del globo terráqueo. El fenómeno de la
migración es conocido por el hombre desde épocas remotas. La temporada de migración de las
aves es uno de los fenómenos naturales más espectaculares. Se estima que cada otoño, cinco
mil millones de aves representantes de 300 especies migran de Norteamérica (Canadá y
Estados Unidos) a México, Centro América y América del Sur, y un número similar viaja del
este de Europa a África (Reed et al., 2003).
La migración de las aves se puede dividir en cuatro categorías: a) especies locales, b)
migraciones de cortas distancias, c) migraciones de largas distancias y d) migraciones
erráticas.
a) Las especies locales son aves que habitan en climas tropicales y subtropicales y que solo se
desplazan pocos kilómetros para aparearse, criar a sus polluelos o en busca de alimento; como
es el caso de los gorriones (Passer domesticus) que viajan entre 15 y 90 kilómetros al año
según su edad (Rappole y Hubálek, 2003).
b) Los viajes migratorios de distancias cortas son viajes de solo pocos kilómetros a sitios o
áreas para invernar o reproducirse. Para muchas especies, la migración está asociada con los
cambios en la altitud más que con las distancias horizontales. Un resultado de este patrón de
migración es que estas especies de aves están confinadas a un área restringida durante el
invierno. La densidad de población es importante en la epidemiología y transmisión de
enfermedades de las aves y tiene una correlación directa con la densidad de mortalidad del
huésped.
c) Los viajes de migraciones de largas distancias son viajes de miles de kilómetros; un ejemplo
es el gaviotín ártico (Figura 5), el cual viaja 17,000 kilómetros de polo a polo. Uno de los

Introducción
8
aspectos más severos de las migraciones largas es que puede contribuir a la adquisición de
patógenos zoonóticos por la migración de estas aves. El estrés fisiológico asociado con la
migración es un factor que causa inmunosupresión e incrementa la susceptibilidad a
enfermedades infecciosas por lo que las aves son un foco latente de infección.
Figura 5. Ruta migratoria del Gaviotin Ártico (Sterna paradisaea). La figura muestra la ruta de migración del gaviotín ártico de más de 17000 kilómetros de polo a polo de America del Norte a América del Sur. En color rojo se muestra el Área de reproducción y anidamiento, el área azul es en donde se encuentra en invierno, el área en amarillo es la ruta por la que migra durante el invierno y el área en color verde muestra la zona en donde se encuentran por migraciones erráticas (Base de datos de aves del mundo, 2009).
d) Por último, las migraciones erráticas, son migraciones hacia nuevos sitios, debido a la
abundancia inusual de alimento en estas nuevas zonas o al cambio de la ruta debido a
desplazamientos por tormentas u otros fenómenos meteorológicos, con lo que hay un riesgo
latente de infección (Rappole et al., 2000; Reed et al., 2003).

Introducción
9
Las rutas de migración de las aves silvestres de Norte América a México, Centro y Sur América
se da por cuatro rutas distintas Estas rutas son: La ruta del Pacifico, la ruta del centro, la ruta
del Golfo y la ruta del Atlántico, como se muestra en la figura 6. Dentro de estas rutas existen
intercambios laterales en las poblaciones durante el proceso de migración, por lo que el
concepto de rutas de migración se usa como una herramienta de manejo para poblaciones de
aves acuáticas durante su desplazamiento (DUMAC, 2006).
Por lo tanto, la interacción entre las aves durante sus viajes pueden esparcir rápidamente las
enfermedades de las cuales son portadoras como es el caso de las enfermedades infecciosas
emergentes, específicamente el Virus del Oeste del Nilo el cual se ha esparcido de costa a
costa en los Estados Unidos y en gran parte del continente (Reed et al., 2003).
En Cuba y Puerto Rico se han reportado aves positivas a anticuerpos contra VON, en el caso
de Cuba dos aves fueron residentes y una migratoria y en el caso de Puerto Rico diez aves
representantes de seis especies fueron positivas; un ave fue residente y nueve aves
migratorias. Estas aves pertenecen a especies migratorias que viajan por la ruta del Atlántico lo
que demuestra el papel tan importante que juegan las aves en el esparcimiento del VON a
nuevos nichos ecológicos (Dupuis et al., 2005). También se ha reportado aves silvestres
infectadas en Canadá, Estados Unidos, México, Centro América, Jamaica, Republica
Dominicana, Isla Guadalupe, Cuba ,Puerto Rico y Argentina (Davis et al., 2005; Dupuis et al.,
2005; Díaz, 2008).
Figura 6.Mapa de las rutas migratorias de las aves silvestres. Ruta del Pacifico.
Ruta del Centro. Ruta del Golfo. Ruta del Atlántico. DUMAC, 2009
Al igual que los casos en humanos, las primeras aves que se documentaron infectadas por el
Virus del Oeste del Nilo en el hemisferio occidental fueron identificadas en agosto de 1999 en

Introducción
10
Nueva York. Como en varios brotes en Europa, el principal vector identificado fue el mosquito
ornitofílico Culex pipiens. Más aún, el brote en humanos ocurrió en siete sitios urbanos
cercanos a tierras húmedas donde se concentraban aves migratorias, mosquitos ornitofílicos y
humanos. Estas circunstancias, conjuntamente con la ecología del virus en el viejo mundo
apoyan la conjetura de que tanto aves silvestres, domésticas y de zoológico, fueron
responsables de la introducción del virus en el nuevo mundo, tanto a través de la migración
normal, el desplazamiento debido a condiciones atmosféricas, o la importación legal o ilegal; y
aunque aun no se sabe con certeza como llegó el virus al continente Americano, las aves
silvestres y las rutas migratorias juegan un papel importante en la distribución del virus
(Rappole et al., 2000; Reed et al., 2003).
En el 2007 se reportaron 2182 aves muertas por VON en 315 condados de los Estados Unidos
pertenecientes a 35 estados y en Puerto Rico. El pico de infección en las aves se reportó
durante la primera semana de septiembre y nuevamente los córvidos fueron la especie más
susceptible con 1690 aves muertas que corresponden al 77% del total de aves muertas. Desde
su aparición en 1999 la infección por VON se a reportado en 321 especies de aves (Lindsey et
al., 2008).
También, en 2007 se reportó una alta mortalidad de aves rapaces debido a la infección con el
VON. Las aves más comunes fueron el halcón cola roja (Buteo jamaicensis) y el gran búho
cornudo (Bubo virginianus), 25 de 56 (45%) aves del estudio resultaron positivas para el VON,
lo que indica la importancia en el consumo de presas infectadas (Saito, et al., 2007).
De igual forma se reporto que en Georgia, la principal causa de muerte en aves rapaces fue el
VON, presentando una tasa de mortalidad de 10% con rangos que van del 3 al 30% (Ellis, et
al., 2007).
1.2.1 Virus del Oeste del Nilo en México
En 2002 en México, sobre todo en los estados del noreste (Tamaulipas, Nuevo León y
Coahuila) se activó un estado de alarma epidemiológica en los sistemas de salud por los
informes de casos humanos, equinos infectados, aves silvestres y especies de mosquitos
vectores positivos al Virus del Oeste del Nilo notificados en los estados de Texas y Louisiana
de Estados Unidos, ya que constituyen un alto riesgo de potenciales epidemias de Virus del
Oeste del Nilo en esta región. Estó resaltó la gran importancia de detectar de modo oportuno la
presencia de este agente viral (Fernández et al., 2007).

Introducción
11
En agosto de 2002 se reportó el primer caso de un Mexicano infectado con el VON, el paciente
tenia 78 años de edad y era residente de Torreón, Coahuila.; sin embargo, se determino que
había adquirido la enfermedad en un viaje que realizó a Houston, Texas y falleció con cuadro
neurológico el 28 de agosto de 2002 (Barriga, et al., 2002).
En noviembre del 2002 se confirmó la presencia del Virus del Oeste del Nilo, en dos estados de
la frontera norte del país: Tamaulipas (un equino) y Coahuila (17 equinos). Al cierre del 2002
se registraron 36 casos de encefalitis atribuible a VON en humanos, todos se descartaron; dos
aves positivas por serología y 21 con serología positiva en equinos (Secretaria de Salud,
2003).
El 5 Mayo de 2003, CPA-SAGARPA recibieron el reporte de la muerte de un cuervo (Corvus
corax) cautivo en el parque zoológico Yum - Ka, en la Ciudad de Villahermosa, en el Estado de
Tabasco. El cadáver del ejemplar fue analizado y se aisló el VON en muestras de tejido
cerebral. El RNA viral fue aislado y caracterizado en el INDRE y por la Universidad de
Galveston, Texas. La cepa se nombró TM171-03 (Gen Bank AY371271) y tuvo un alto grado
de homología con la cepa Flamingo 382. Estudios filogenéticos indican que este aislamiento, el
primero en México, está relacionado con las cepas aisladas en la parte central de Estados
Unidos, en Texas (Estrada, et al., 2003).
Otro estudio mostró que la secuencia genómica para TM171-03 fue distinta a la secuencia
prototipo de la cepa NY99 por 46 nucleótidos (0.42%), también se reportó que la secuencia de
prM y E de TM171-03 tuvo mutaciones por sustitución de aminoácidos. Durante el 2004 no
hubo casos de encefalitis por VON en humanos pero se identificaron caballos seropositivos,
aunque pocos con enfermedad clínica. La secuencia de nucleótidos del VON obtenida de
caballos infectados en estados fronterizos sugiere que hay una estrecha relación con los
aislamientos en Texas (Beasley et al., 2004).
También en 2003 se reportaron 91 muestras positivas a VON determinadas por ELISA IGg en
caballos de distintos municipios de 6 estados de la republica como se muestra en la figura 7
(Estrada, et al., 2003).
En 2004 se analizaron 3856 muestras de sangre de los bancos de sangre de 29 de los 31
estados de la República Mexicana, la muestra de un donador asintomáico fue positiva para
VON en Chihuahua (Téllez et al., 2006)
En una investigación realizada en 2005, se reportaron aves, mamíferos, mosquitos y reptiles
seropositivos al VON en el parque zoológico la venta y en el Parque zoológico Yum-Ka de la
ciudad de Villa Hermosa Tabasco (Hidalgo et al., 2008).

Introducción
12
Se ha reportado la presencia de anticuerpos contra el virus en mamíferos no humanos en la
península de Yucatán. 70 caballos fueron positivos al virus en Cozumel Quintana Roo, un
coyote y un jaguar en el zoológico El Centenario en Mérida Yucatán y en cocodrilos de Ciudad
del Carmen Campeche (Farfán et al., 2006).
Figura 7. Entidades Federativas positivos al VON en la republica Mexicana. El mapa
muestra los estados positivos por serología a VON en 2003 (Estrada, et al., 2003).
En cocodrilos de granja también se han reportado pruebas positivas para anticuerpos anti-
VON, y la cepa aislada que se encontró tuvo un 94% de homología con la cepa NY 2000
Crow/ AF404756. Las muestras positivas fueron recolectadas de zonas por donde transitan
aves migratorias lo cual relaciona estrechamente a las aves como portadoras de este virus
(Sandoval, 2006).
En un estudio realizado en 2006, se menciona que la cepa aislada en Tabasco en 2003
TM171-03, es parecida en a una cepa aislada en florida en 2001, a otra aislada en Lousiana en
2002 y un poco distinta a la aislada en Nueva York en 2000. Este estudio demostró que la cepa
TM177-03 se introdujo a México por la migración de aves del sureste de los Estados Unidos al
Golfo de México o al Caribe pasando por México (Dardorff, et al., 2006).
Hasta diciembre del 2006, la Secretaria de Salud ha reportado 553 serologías positivas a Virus
del Oeste del Nilo de 1586 muestras estudiadas en equinos, 117 serologías positivas de 943
estudiadas en aves y uno con evidencia a Virus del Oeste del Nilo de 203 casos estudiados en

Introducción
13
humanos. El caso positivo para el Virus del Oeste del Nilo fue reportado en el estado de
Oaxaca en el municipio de Santa María Tonameca (Figura 8).
Figura 8. Casos de Virus del Oeste del Nilo en Humanos en México. En la figura se
muestran los datos de el monitoreo del virus del oeste del Nilo en México realizados por la Secretaria de Salud, los cuales solo llegan hasta el 2006 y reporta una muestra positiva a VON de 203 analizadas. La muestra positiva fue en el estado de Oaxaca (CENAVECE, 2009).
Otros estudios reportan 7 casos que ocurrieron en los estados de la frontera, Chihuahua (4),
Sonora (1), y Nuevo León (1) en 2003; de los 6 casos 3 fueron febriles y 3 encefalitis; sin
embargo no hubo casos fatales; y en Sonora (1) en 2004 (Ramos y Falcón, 2004; Dardorff, et
al., 2006).
Por otra parte, en estudios realizados por Fernández et al., (2007), se observa una gran
prevalencia de seropositividad para anticuerpos IgG contra el Virus del Oeste del Nilo en
personas. Se observó que el diagnóstico serológico de pacientes infectados con el Virus del
Oeste del Nilo se complica en la población del norte de México debido a que las pruebas para
la detección de anticuerpos sufren reacción cruzada con el virus del dengue, el cual es
endémico en esta región.
La migración de diferentes especies de aves provenientes de regiones del norte de los Estados
Unidos y Canadá a distintos estados del territorio mexicano durante la temporada invernal,
hace suponer que nuestro país está en riesgo potencial de presentar en algún momento brotes
de esta arbovirosis.

Introducción
14
El riesgo de la dispersión del VON es a través de las aves migratorias. La introducción de este
virus a zonas endémicas del dengue puede estar asociada con cierto grado de protección hacia
la población ya que gran parte de ella tiene anticuerpos antidengue que podrían conferir
protección cruzada temporal o casos menos severos (Ramos y Falcón, 2004), lo cual se vería
reflejado en el bajo número de casos, no solo en México sino también en el resto de
Latinoamérica y el Caribe, ya que hay condiciones de transmisión de arbovirosis en esta área
(Hayes et al., 2005a)
En resumen, la baja incidencia del VON en México se debe a la resistencia de la población
mexicana, posiblemente a través de la inmunidad cruzada por las infecciones de flavivirus
como el Dengue (Dardorff et al., 2006).
1.3 Características del Virus
El VON pertenece al genero flavivirus donde se ubican virus como el de la Encefalitis
Japonesa, Fiebre Amarilla, Encefalitis transmitida por garrapata y Dengue. Se caracteriza por
tener tropismo por el sistema nervioso central; y en el caso de aves, también es cardiotrópico
(Ramos y Falcón, 2004; Ellis et al., 2007).
Es un Arbovirus que pertenece a la familia Flaviviridae, donde se encuentran tres géneros:
flavivirus, epaciavirus y pestivirus. El VON es esférico, cubierto por una membrana lipidica de
45 a 60 nm de diámetro (Figura 9), presenta una cadena de RNA de polaridad positiva
(Lanciotti et al., 1999; Barriga et al., 2002; Jawetz et al., 2002; Gea- Banacloche et al., 2004;
McMurtrey et al., 2008).

Introducción
15
Figura 9. Representación esquemática de la estructura del Virus del Oeste del Nilo.
Muestra la forma esférica del virus.
Se caracteriza por estar envuelto, tiene una nucleocapside icosaédrica de 30 a 35 nm
compuesta de una proteína de 12,000 Daltons. Traduce su genoma a una poliproteína de 3400
aminoácidos que se dividen proteoliticamente en 3 proteínas estructurales: cápside (C),
membrana (prM/M) y envoltura (E), y 7 proteínas no estructurales (NS1, NS2A, NS2B, NS3,
NS4A, NS4B y NS5), las cuales participan en la replicación viral (Barriga et al., 2002; Hayes et
al., 2005; Chung et al., 2006; McMurtrey et al., 2008).
La proteína E, es la proteína estructural de mayor importancia ya que es la encargada de
mediar la unión del virus a la célula, tiene capacidad de aglutininar eritrocitos e induce la
formación de anticuerpos (Barriga et al., 2002), y las proteínas no estructurales del VON
suprimen la respuesta inmune antiviral del hospedero, sin embargo el mecanismo es
desconocido (Kramer et al., 2007),
El genoma del Virus del Oeste del Nilo es de 11.029 nucleótidos y contiene un fragmento de
lectura abierta de 10301 nucleótidos (Lanciotti et al., 1999; Barriga et al., 2002).
Los más de 70 virus del género flavivirus se caracterizan por ser neurotrópicos y
hepatotrópicos; son de importancia en humanos porque pueden ser transmitidos por artrópodos
al igual que el dengue, la fiebre amarilla y la encefalitis de San Luís (Jawetz et al., 2002; Gea-
Banacloche et al., 2004).
El VON es miembro del serogrupo del virus de la encefalitis japonesa (JE), el cual también
incluye los virus de: Cacipacore, Koutango, JEV, encefalitis del Valle de Murray, encefalitis de
Sant Luis, Usutu y Yaounde (O’Donnel y Travis, 2007).
Tiene dos linajes genéticos (I y II): el I es una estirpe de Norte América, Europa, África; Asia, y
Australia; el linaje II es una estirpe aislada en el sur del Sahara y Madagascar, los virus
pertenecientes a este linaje son exclusivos de África y no están relacionados con brotes en
humanos. Basado en homología de secuencias, el virus Kunjin el cual es endémico desde
Australia hasta Asia, es ahora considerado un subtipo del Virus del Oeste del Nilo (Lanciotti et
al, 1999).
En el linaje I la estirpe se anuló y se dividió en 4 clases: Kunjin, India, A y B (Hayes et al.,
2005). En la figura 10 se muestra el árbol filogenético del Virus del Oeste del Nilo.

Introducción
16
Figura 10. Árbol Filogenético del virus del Oeste del Nilo. Se muestra las divisiones por
linaje, y como están clasificadas las clases del linaje 1 y 2 del VON. (Lanciotti et al., 1999).

Introducción
17
1.4 Replicación del Virus del Oeste del Nilo
La infección con el virus comienza con la unión del virus a la célula del hospedero, a través de
la proteína E que reconoce un receptor celular todavía no identificado. Sin embargo, como el
virus infecta tanto células de insectos como de vertebrados, el receptor debe ser una proteína
celular altamente conservada. El virus entra a la célula por endocitosis; luego cuando el
lisosoma se une al endosoma, las enzimas lisosomales permiten la liberación de la
nucleocápside hacia el citoplasma y el RNA del virus. En las células infectadas se observa una
extensa proliferación y reorganización de membranas citoplasmáticas y perinucleares del
retículo endoplásmico (Hayes et al., 2005).
El ensamble de los viriones ocurre en asociación con el retículo endoplásmico rugoso (Figura
11). Viriones intracelulares inmaduros, los cuales contienen heterodímeros de E y prM que se
acumulan en vesículas y se transportan a través del hospedero por la vía secretora en donde
se dan las glicosilaciones en E y prM (Hayes et al., 2005).
Una vez que las glicoproteínas maduran, los viriones son transportados hacia la membrana
citoplasmática en vesículas y son liberadas por exocitosis provocando lisis y muerte celular
(Jawets et al., 2002).
Figura 11. Esquema del ciclo de replicación del Virus del Oeste del Nilo. 1. Se muestra la unión del virus a la célula. 2. Se da una endocitosis mediada por receptores. 3. Se muestra la fusión entre el virión y el endosoma de la membrana celular. 4. Se da la pérdida de la cubierta. 5. Posteriormente se da la traducción y procesamiento de las poliproteínas. 6. Se replica el RNA asociado a membranas. 7. y comienza la morfogénesis del virión en las vesículas intracelulares. 8 Posteriormente el virión es transportado y las glicoproteínas maduran. 9. Finalmente se da la fusión de las vesículas en la membrana plasmática y el virión es liberado. (Jawets, et al., 2002).

Introducción
18
Durante la infección viral, el proteoma de las células es alterado, como ocurre con los péptidos
presentados a los linfocitos T citotóxicos por la HLA. Aunque los linfocitos T citotóxicos
reconocen a las células infectadas, los epitopos presentados a las moléculas clase I HLA no se
saben con certeza. Recientes estudios demuestran que el epitopo dominante que reconoce al
VON es SVGGGVFTSV con una homología de 97.2% en la secuencia de la cepa NY 99
(McMurtrey et al., 2008)
1.5 Ciclo de transmisión
El virus del Oeste del Nilo tiene un ciclo de transmisión entre diferentes especies de aves y
mosquitos del género Culex (Dupois et al., 2005), mosquito - ave - mosquito (Figura 12), lo
que es una amenaza para la salud humana, veterinaria y de la vida silvestre (Komar et al.,
2005).
Sin embargo, el VON se ha detectado en 75 especies de artrópodos, incluyendo las 60
especies de mosquitos que habitan en América (McGee et al., 2007).
Las principales especies de mosquitos transmisoras del VON son Culex pipiens, Culex
quinquefasciatus y Culex tarsalis. Experimentalmente se ha demostrado que hay transmisión
vertical en mosquitos y el virus se ha aislado en hembras hibernantes, lo que sería un
mecanismo de persistencia viral reemergiendo en verano (Hayes et al., 2005ª).
La especie de mosquito mas ornitofílica es la Culex pipiens, aunque también algunas especies
de Aedes, como es el caso Aedes vexans, que juega un papel importante en la transmisión a
mamíferos por que ayudan a propagar la enfermedad con mayor rapidez infectando a más
aves, las cuales se consideran reservorios naturales y amplificadores del virus (Bernard et al.,
2001; Barriga, 2002; O´Donnell y Travis, 2007).
Las garrapatas Argas arboreous, que experimentalmente infectan a pollos y ratones, o la
garrapata Ixodes scapularis, se han encontrado infectadas con el virus, sin embargo, no se ha
demostrado que puedan infectar a través de la mordedura (Hayes et al., 2005a).
El mosquito vector ingiere el virus de la sangre de las aves virémicas, y para que la transmisión
del virus ocurra, 5 días después de que el mosquito ingirió la sangre infectada el virus se debe
de replicar en las células de las glándulas salivales, de este modo el mosquito secretara el
virus en la saliva. El VON es inoculado en el hospedero cuando el mosquito secreta saliva para
facilitar la succión de la sangre. El pico de la viremia lo alcanzan a los 9 días con 107
UFP/ml y
secretan saliva con el VON hasta los 28 días post infección (Girard et al., 2007).

Introducción
19
Figura 12. Esquema del ciclo de amplificación y mecanismos de conservación del Virus del Oeste del Nilo. El ciclo se da Ave -Mosquito - Ave, siendo estas ultimas el
reservorio y amplificador natural del virus. Los humanos, equinos, bovinos, ovinos y demás animales domésticos son hospederos terminales ya que no transmiten la enfermedad (Jawetz et al., 2002; Barriga et al., 2002.)
La mortalidad y la morbilidad en las aves es el sello de los brotes de Virus del Oeste del Nilo en
Norte América. Usualmente las aves sobreviven a la viremia y desarrollan inmunidad, sin
embargo hay especies muy susceptibles como los miembros de la familia Corvidae como es el
caso de los cuervos y los gorriones (Langevin et al., 2005). Estas especies del orden
Passeriforme logran alcanzar niveles de viremia de 1012
UFP/ml (van der Meulen et al., 2005),
y las especies del orden Charadriiforme , Strigiformes, y Falconiforme, desarrollan niveles de
viremia suficientes para infectar a los mosquitos. Especies de Columbiformes, Pisiformes y
Anseriformes no desarrollan viremias suficientes. Ciertos Passerinos como el Quiscalus
quiscula y Corvidos como el cuervo (Corvus brachyrhynchos), la urraca y el gorrión doméstico
(Passer domesticus), son altamente infecciosos para los mosquitos y tienen altas tasas de
mortalidad > 40% (Hayes et al., 2005a).
Las aves passerinas, a diferencia de otras aves, tienen comportamientos defensivos muy
pobres que facilitan que los mosquitos se alimenten de ellas; este comportamiento sumado a
las altas viremias que desarrollan contribuyan a la rápida diseminación del VON entre otras
aves (Darbroand and Harrington, 2007).
Se han realizado estudios que demuestran que del 75 al 100% de los mosquitos Culex tarsalis
que han consumido sangre infectada, con concentraciones de 107.1
UFP/ml, pueden infectar a
las aves y del 0 al 36% con concentraciones de 104.9
UFP/ml (Hayes et al., 2005a).

Introducción
20
Otro punto importante del vector Culex tarsalis es que tiene afinidad para alimentarse de los
animales domésticos (mascotas), y es considerado uno de los vectores mas eficientes del VON
con altas tasas de transmisión oral y vertical. Culex tarsalis no solo se alimenta de aves,
también juega un papel importante en la amplificación del virus; los mamíferos son su fuente de
alimento oportunista y la abundancia de estos mosquitos en áreas peridomésticas lo hacen
vector para humanos y animales domésticos (Venkatesan et al., 2007).
Sin embargo, es importante recalcar que el comportamiento defensivo del hospedero contra los
mosquitos puede facilitar o dificultar la transmisión del VON por vectores infectados, como
también es cierto que la transmisión del virus es facilitada si un mosquito infectado se alimenta
de múltiples hospederos que no se defienden. Diversos estudios han demostrado que la
alimentación de los mosquitos se reduce cuando aumenta la respuesta defensiva de los
vertebrados como las palomas (Columba liva), los gorriones domésticos (Passer domesticus),
conejos, (Oryctolagus cuniculus), ardilla gris, (Sciurus carolinensis), ratón de los campos de
algodón (Pecomysais gossypinus) y el hombre (Homo sapiens), entre otros (Darbroand and
Harrington, 2007)
Se ha demostrado que la transmisión del virus de ave a ave es posible, también se ha
reportado que las aves rapaces se pueden infectar con el virus cuando se alimentan de presas
que se encuentran infectada como otras aves, mamíferos y ratas (Sandoval, 2006).
Datos de campo y experimentales indican que el VON puede ser transmitido por aerosoles,
ingestión y contacto directo, como se ha demostrado en trabajos donde se utilizaron Hamsters
dorados que se alimentaron de ratones neonatos infectados con el VON, presentando el pico
de la viremia con 105.5
UFP/ml a los 4 días (Sbrana et al., 2005).
Otra vía de transmisión pueden ser las heces de aves contaminadas con el virus; además,
experimentalmente los hamsters pueden tener transmisión oral (Sbrana et al., 2005;
O´Donnell y Travis, 2007), y también se ha reportado la transmisión oral en aves y gatos
(Kramer et al., 2007).
Los seres humanos y los equinos son considerados como hospederos terminales por que no
intervienen en el ciclo natural de transmisión (Petersen y Oehrign, 2001) La intensidad de
transmisión a humanos depende de la abundancia y los patrones de alimentación de los
mosquitos infectados, así como de la ecología local. La concentración máxima detectada en
humanos es de 103.2
UFP/ml, la cual se considera baja para que los mosquitos se infecten
(Hayes et al., 2005a).

Introducción
21
En los Estados Unidos y Puerto Rico se han presentado casos de transmisión del Virus del
Oeste del Nilo por el trasplante de órganos, la transfusión sanguínea, vía placentaria (con
severas secuelas en el niño), el amamantamiento y la transmisión percutánea con una herida
por aguja hipodérmica (Whitley, 2003; Torres et al., 2008).
Cuando una persona es transplantada con órganos infectados con el VON, el riesgo de
enfermedad neuroinvasiva aumenta a 40%, a diferencia de la población normal que es de 1%
(Kramer et al., 2007).
Durante el 2007, los casos de VON se presentaron de junio a septiembre (Figura 13),
alcanzando el pico de casos en humanos en la primer semana de agosto lo que coincide con la
estacionalidad que ha demostrado el VON desde su aparición en 1999 (Lindsey et al., 2008).
Figura 13. Número de casos de enfermedad neuroinvasiva por VON en los Estados Unidos durante el 2007. La gráfica muestra el número de casos de enfermedad neuroinvasiva
por semana durante el 2007. Incluye meningitis, encefalitis y parálisis flácida aguda. N = 1227.
1.6 Patogénesis
Se piensa que el Virus del Oeste del Nilo se replica en el sitio de inoculación al hospedero, en
las células dendríticas; posteriormente, las células infectadas migran hacia los nódulos
linfáticos en donde ocurre la amplificación viral, lo que resulta en la viremia primaria y la
diseminación a otros tejidos periféricos por el torrente sanguíneo (Shrestha and Diamond,
2007).
Después de varias replicaciones, el VON se acumula en la sangre y se da la penetración del
virus al sistema nervioso central por la estimulación de los receptores, con lo que se
incrementan los niveles de factor α de necrosis tumoral que aumenta la permeabilidad de la

Introducción
22
barrera sangre cerebro y permite la infiltración de CD4, CD8, NK, células T y macrófagos al
sistema nervioso central (Kramer et al., 2007). El virus infecta y lesiona directamente el
parénquima neuronal, la médula espinal y el tallo encefálico (Hayes et al., 2005b ; Shrestha
and Diamond, 2007).
Las células T CD8+ tienen la función de controlar la infección aguda por virus neurotrópico. En
general los CD8+ tienen funciones antivirales por dos mecanismos: a través de la vía no
citolítica de secreción de citocinas antivirales, como el interferón gamma (IFN-γ) y el factor α de
necrosis tumoral; y por la vía citolitica con el uso de perforinas o por la interacción de Fas-FasL
(Shrestha and Diamond, 2007).
Recientes estudios han demostrado que ICAM -1 juega un papel importante en la entrada del
VON al sistema nervioso Central, esto es debido a que ICAM -1 es una molécula de adhesión
para los leucocitos y transmite señales que influyen en la permeabilidad sangre - cerebro lo que
facilita la infiltración de los leucocitos al cerebro y a la medula espinal (Dai et al., 2008).
El VON obtiene resistencia del sistema inmune innato y adquirido, el sistema inmune adquirido
genera anticuerpos neutralizantes que protegen de la infección, mientras que una deficiencia
de linfocitos T citotóxicos o el Complejo Mayor de Histocompatibilidad exhibe un incremento en
la carga viral e incrementa la mortalidad (McMurtrey et al., 2008)
En personas aparentemente sanas, el Virus del Oeste del Nilo ha sido aislado en muestras de
suero tomadas varios días antes del inicio de los síntomas, pero la viremia desapareció
rápidamente al haberse elevado los títulos séricos de IgM e IgG neutralizantes del Virus
(Campbell et al., 2002; Carrada, 2004).
Los Daños histopatológicos en tejidos de pacientes con enfermedad neuroinvasiva incluyen
inflamación perivascilar, necrosis variable y pérdida de neuronas. El tallo espinal y la medula
espinal pueden ser las más afectadas. Los pacientes con VON pueden tener infiltración
linfocítica perivascular en la médula espinal y nódulos microgliales. La inflamación endonuclear
mononuclear de las raíces de los nervios craneales y espinales se presentan en menor
frecuencia (Hayes et al., 2005).
1.7 Signos clínicos
Aunque las infecciones humanas en áreas endémicas de VON son comunes, la mayoría de
éstas son generalmente leves o subclínicas, mientras que la enfermedad severa se relaciona
generalmente con personas de la tercera edad (Lanciotti et al., 2000; Carrada, 2004).

Introducción
23
Clínicamente la fiebre del Oeste del Nilo es una infección severa con participación significativa
del sistema nervioso central y elevada mortalidad; con período de incubación de 3 a 14 días;
uno de cada 150 desarrolla enfermedad neurológica (Barriga et al., 2002). Aproximadamente
del 70 al 80% de los casos son asintomáticos (Hayes et al., 2005, Torres et al., 2008), y el
20% al 30 % de los individuos infectados desarrolla un cuadro clínico moderado (Barriga et al.,
2002; Ramos y Falcón, 2004; Torres et al., 2008).
El síndrome clínico compatible con la infección del Virus del Oeste del Nilo es fiebre de
aproximadamente 39 ºC, dolor de garganta, anorexia, meningitis aséptica, debilidad muscular,
malasia, mialgia, artralgia, alteración del estado mental, signos gastrointestinales, dolor
abdominal, náuseas, vómito, diarrea, linfoadenopatias, cefalea, rigidez o parálisis del cuello,
parálisis flácida aguda, taquicardia, taquipnea, ataxia, eritomatosis macular, fotofobia,
meningitis, encefalitis, parálisis de los nervios craneales y parálisis muscular diafragmática que
provoca un paro respiratorio (Papperell et al., 2003; Ramos y Falcón, 2004; Kramer et al.,
2007; Hall et al., 2008; Torres et al., 2008).
El 1% de los casos presenta enfermedad neuroinvasiva con signos de encefalitis, meningitis y
parálisis flácida, en ocasiones se puede presentar meningoencefalitis o presentar los tres
padecimientos (Kramer et al., 2007).
Aunque no esta demostrado, después de la infección por VON pueden quedar secuelas
neurológicas como parkinsonismos, tremor, dificultad para concentrarse y síntomas de
ansiedad y depresión (Hall et al., 2008).
Las manifestaciones clínicas no neurológicas que pueden ocurrir durante la infección del Virus
del Oeste del Nilo incluyen hepatitis, pancreatitis, miocarditis, orquitis, manifestaciones oculares
como coreoretinitis, neumonía, y pielonefritis (Papperell et al., 2003; Hayes et al., 2005b).
La letalidad registrada en pacientes hospitalizados ha variado de 4% a 18%, pero el factor de
riesgo más importante es la edad, por ejemplo, en los mayores de 70, años la letalidad
hospitalaria varió de 15% a 29%. La encefalitis con debilidad muscular, el estado de coma, y el
antecedente de diabetes o de inmunodeficiencia son factores de riesgo importantes, asociados
a la discapacidad residual y la muerte del enfermo (Carrada, 2004; Hayes et al., 2005a).
Entre los animales domésticos en que se han observado manifestaciones clínicas, se
encuentran las aves y los equinos, aunque en estos últimos la mayor parte de las infecciones
son asintomáticas. Dentro de los signos clínicos que se pueden observar en los animales están
la anorexia, deshidratación, depresión, pérdida de peso, letargia, tremor muscular,
faciculaciones, desorientación, erizamiento de plumas, miocarditis, dolor abdominal, visión
impar y muerte (van der Meulen et al., 2005; O´Donnell y Travis, 2007, Medica et al., 2007).

Introducción
24
En las aves los signos neurológicos son ataxia, parálisis, volteretas, movimientos de carrera,
opistótonos e incoordinación (van der Meulen et al., 2005).
1.8 Diagnóstico
Los estudios rutinarios de laboratorio no distinguen la infección del Virus del Oeste del Nilo de
otras infecciones virales. La detección del virus por inmunoglobulinas específicas IgM en suero
o en fluido cerebroespinal proporciona una fuerte evidencia para las infecciones del virus
(Hayes et al., 2005b).
Hay 2 pruebas básicas para el diagnóstico del Virus del Oeste del Nilo: 1) la prueba serológica
preferida es la prueba inmunoenzimática (ELISA, enzyme-linked immunosorbent assay) para
IgM o la prueba indirecta de anticuerpos fluorescentes (IFA), y se utiliza la prueba de reducción
de placas para confirmación utilizando suero o líquido cefalorraquídeo (Lanciotti, 2003; Hayes
et al., 2005b). El resultado positivo de la prueba de ELISA para IgM indica que la infección del
virus es reciente ya que se encuentra en el suero del paciente y es detectable durante los 8
días post-síntomas. La IgM puede persistir hasta por un año en los pacientes (Kramer et al.,
2007).
2) La transcripción inversa o retotranscripción seguida de reacción en cadena de la polimerasa
(RT-PCR, reverse transcription polymerase chain reaction), amplificación basada en la
secuencia de ácidos nucleicos (NASBA, nucleic acid sequence-based amplification) o cultivo.
Esta es la prueba más recomendada, por su rapidez, sensibilidad y especificidad, para la
detección del virus, tanto en humanos como en aves, animales y mosquitos (Barriga et al.,
2002)
La secuencia de eventos virales y clínicos depende de la viremia en los períodos de incubación
con picos a varios días antes de la aparición de los síntomas. La IgM aparece cuando se
resuelve la viremia y con la aparición de los síntomas (Lanciotti, 2003).
Para la realización de las pruebas anteriores se requiere de muestras de sangre o de tejidos de
órganos como corazón, hígado, bazo, riñón y cerebro. Sin embargo, estudios realizados por
Komar han demostrado que no hay diferencias significativas en los resultados obtenidos en la
toma de muestras cloacales y orales tomadas con hisopos con punta de Dracon y muestras de
tejido en la RT-PCR en aves infectadas experimentalmente. Por lo cual es el método de
muestreo utilizado para la tomo de muestras de aves hoy en día, de este modo, este método
facilita la toma de muestras, reduce el tiempo y disminuye los riesgos para el personal (Komar
et al., 2002).

Introducción
25
1.9 El Virus del Oeste del Nilo en Otras Especies.
Se han encontrado anticuerpos en 30 especies distintas de vertebrados; por ejemplo en
Sudáfrica la prevalencia en perros va de 8 a 37%, mientras que en Missouri y Nueva York es
de 2.4 y 5% respectivamente. En un estudio que se realizó con una población de osos de
Croacia la prevalencia fue de 36% y en Nueva York es de 6%; en el conejo silvestre de Francia
fue de 0.37%, en el cerdo doméstico de la india de 2.6 a 9.8, en ganado bovino en México
1.5% y en ganado bovino de Nigeria 6%, en camellos, ovejas y cabras de Nigeria fue de 26%,
20% y 8% respectivamente. En Louisiana 36% en primates y en Israel 70% en cocodrilos (van
der Meulen, et al., 2005)
Otras especies en las que se han identificado anticuerpos y algunos casos clínicos son los
murciélagos (Myotis licifugus), gatos (Felis catus), perros (Canis lupus familiariss), lobo ártico
(Canis lupus) ardillas (Sclurus niger), oso negro (Ursus americanus), conejos (Oryctolagus
cuniculus), ovinos (Ovis aries), caprinos (Capra spp), llamas (Lama glama) y alpacas (Lama
pacos) y macacos (Maccaca spp) en cautiverio (Cárdenas y Vargas, 2002; Blitvich et al., 2003;
Ǿlberg, et al., 2004; Davis et al., 2005; O´Donnell y Travis, 2007; Lindsey et al., 2008).
Hay especies que a pesar de que no se conoce su papel en la transmisión del VON, pueden
contribuir de manera importante a su distribución y dispersión, como es el caso de roedores en
Nigeria y murciélagos en la India ya que son especies que alcanzan altos niveles de virémicos
al igual que los cocodrilos y las ranas, Rana ridibunda (Hayes et al., 2005a).

Justificación
26
2. Justificación
La infección por el Virus del Oeste del Nilo ya se ha reportado en aves, mosquitos, caballos,
humanos y otros mamíferos en el República Mexicana; la fiebre del Oeste del Nilo y la
encefalitis causada por el Virus del Oeste del Nilo son enfermedades emergentes por que se
están presentando en zonas geográficas donde antes no se habían presentado, por lo que
deben ser diagnosticadas de forma veraz y oportuna ante la posibilidad de que surja una
epidemia en humanos en México debido al incremento de la actividad del Virus del Oeste del
Nilo en la mayor parte de Estados Unidos
Distintos experimentos han demostrado que las aves son excelentes centinelas en la vigilancia
epidemiológica del VON ya que las aves son los hospederos naturales y amplificadores del
virus. Las aves se utilizan para monitorear la actividad del Virus del Oeste del Nilo en diversas
zonas, realizando de esta forma un diagnóstico oportuno para evitar brotes que pongan en
riesgo la salud humana. Para este fin, es necesario que en México se monitoreen las aves
silvestres residentes y migratorias para mejorar la vigilancia de dicho virus. Esta información
complementará y comprenderá un estudio epidemiológico para conocer la prevalencia del virus
en las aves estudiadas en los distintos estados de la república. El conocer la dispersión del
virus en el país por medio del uso de animales centinelas aportara información para
implementar las medidas de prevención oportunas y mantener bajo control dicha enfermedad,
y de este modo mantener la salud humana.

Objetivos
27
3. Objetivos
3.1Objetivo general
Detectar la presencia del Virus del Oeste del Nilo en aves silvestres residentes y migratorias
de México.
3.2 Objetivos específicos
1. Identificar cual es la especie de ave con mayor prevalencia al Virus del oeste del Nilo.
2. Establecer la Entidad Federativa con mayor prevalencia al Virus del Oeste del Nilo
3. Establecer la prevalencia diferencial entre aves residentes y migratorias.

Materiales y Métodos
28
4. Materiales y métodos
4.1 Colección de muestras
Los criterios para seleccionar a las aves fueron: aves que se han reportado como susceptibles
a la infección por VON. Se recolectaron muestras cloacales y orales de 200 aves silvestres
migratorias y/o residentes de distintas zonas del país con hisopos estériles con punta de
Dracon, de acuerdo a la técnica descrita por Komar (Komar et al., 2002), y se depositaron en
tubos con 5 ml de infusión cerebro corazón (BHI por sus siglas en ingles Brain heart Infusión)
con antibióticos (Penicilina y Estreptomicina) Las muestras fueron congeladas a –20º C y se
transportaron al Laboratorio de Medicina de Conservación de la Escuela Superior de Medicina
del Instituto Politécnico Nacional donde se realizaron las pruebas de laboratorio (Extracción de
RNA y RT-PCR).
4.2 Extracción de RNA
La extracción de RNA se realizó a partir de las muestras cloacales y orales para las cuales se
siguió el procedimiento de extracción del kit Qiagen Rneasy®: método de centrífuga.
Se tomaron 500µl de la muestra (BHI con hisopo), se le adicionaron 500µl del regulador RLT
(adicionado con ß-mercaptoetanol) y se homogenizó en el vortex durante 15 seg. Después
de este paso se da un “quick spin” en la microcentrífuga para recuperar el líquido que quede
disperso en el tubo eppendorf, después se adicionaron 500µl de Etanol al 70 % y se agitó en
el vortex durante 15 seg. Se centrifugó a 5,000 x g durante 5 min. y se agregó a la columna
la mezcla, primero 700µl se centrifugaron durante 15 seg. a 8,000 x g, se desechó el filtrado
y después los restantes 700µl se centrifugaron durante 15 seg. a 8,000 x g, se desechó el
filtrado. Se adicionaron a la columna 700µl de regulador RW1 (1X) y se centrifugó durante
15 seg. a 8,000 x g., se desechó el filtrado y se colocó la columna en un tubo eppendorf
nuevo de 2ml. Se adicionaron a la columna 500µl de regulador RPE y se centrifugó a 8,000
x g durante 15 seg. y se desecho el líquido filtrado. Se agregaron a la columna otros 500µl
de regulador RPE y se centrifugó a 8,000 x g durante 15 segundos, se desecho el líquido
filtrado y se colocó la columna en un tubo colector nuevo de 2ml. Posteriormente se
centrifugo a máxima 11,000 xg durante 2 minutos para remover el exceso de etanol y se
colocó la columna en un tubo eppendorf nuevo de 1.5 ml libre de RNAsas. Se adicionaron 50
µl de agua destilada libre de RNAsas directamente en el centro de la membrana de sílica
gel, sin tocar la membrana y se incubó de 1-5 minutos a temperatura ambiente. El RNA fue
eluido por centrifugación a 11,000 xg durante 1 min; posteriormente se mantuvo a -70º C
hasta su uso.

Materiales y Métodos
29
4.3 Reacción de la RT-PCR punto final
Se amplificó un fragmento de 408 pares de bases (nt 233 al nt 640) el cual contiene la porción
C-terminal del gen C y en la región N-terminal del gen prM del Virus del Oeste del Nilo, lo cual
se llevo a cabo utilizando el kit “One step RT-PCR” de Qiagen ®, utilizando el protocolo descrito
por Shi (Shi et al., 2001).
Los iniciadores que se utilizaron son los descritos por Lanciotti et al., 2000, los cuales son
específicos para el VON y son el 3F 5’-TTGTGTTGGCTCTCTTGGCGTTCTT-3’ y el 4R 5’-
CAGCCGACAGCACTACATTCATA-3’.
La reacción de amplificación se llevo a cabo según las instrucciones del fabricante pero con
ciertas modificaciones en el volumen final de la mezcla de reacción. En el cuadro 1 se muestra
el contenido de la mezcla de reacción que se utilizó.
La reacción de la RT-PCR se realizó en el termociclador Biometra (Figura 14), y consistió en
50º C durante 30 minutos para la síntesis de la cadena de cDNA, 95º C durante 15 minutos
para inactivar la transcriptasa reversa y para activar la DNA taq polimeras, 35 ciclos a 94º C
durante 45 segundos para la desnaturalización, 56º C durante 45 segundos para el
alineamiento de los iniciadores, 72º C durante 1 minuto para la amplificación y un alargamiento
final de 72º C durante 10 minutos.
Una vez terminados los ciclos se mezclaron 3 µl de Buffer de carga de cDNA y 15 µl del
amplificado, 3 µl de Buffer de carga y 6 µl de marcador de longitud molecular de 100 pb
y se colocaron en un gel de agarosa al 1.5 % utilizando TBE 1X (Tris-Borato-EDTA) como
regulador de corrimiento para realizar la electroforesis por una hora con 30 minutos a 110 Volts
(V). Una vez terminada la electroforesis se tiñó con una solución de bromuro de etidio a una
concentración de 0.5 μg/ml y se observó en un Transiluminador Ultravioleta BIO –RAD (Figura
14).

Materiales y Métodos
30
Cuadro 1. Contenido de la mezcla de reacción para la amplificación.
Mezcla de reacción Concentración final Volumen
Regulador 5X 1X 5 μl
dNTPs (10mM) 0.4 mM 1 μl
Primer 3F (15 μM) 0.6 μM 1 μl
Primer 4R (15 μM) 0.6 μM 1 μl
Sol. Q 5X 1X 5 μl
Mezcla de enzimas 1 μl .5 μl
H2O --- 6.5μl
RNA molde --- 5 μl
Volumen total 25 μl
Figura 14. Equipos que se utilizaron para el desarrollo del trabajo. En la fotografía del lado derecho se observa el transiluminador de luz ultravioleta BIO –RAD, y en la fotografía del lado izquierdo se muestra el termociclador Biometra.

Resultados
31
5. Resultados
De las 200 aves muestreadas en 11 estados de la república Mexicana (Tabla 1),
pertenecientes a 7 especies, solo 16 aves correspondientes a 4 especies presentaron
amplificados de 408 pb, es decir, fueron positivas al Virus del Oeste del Nilo en la RT-PCR,
como se muestra en la figura 15, 16, y 17. Las aves positivas fueron de los estados de
Chiapas, Oaxaca y Tamaulipas, las especies afectadas fueron Phalacrocorax auritus (Figura
18), Archilochus colibris (Figura 19), Larus delawarensis (Figura 20), y Zenaida macroura
(Figura 21). En el gel de la electroforesis se observan restos de un exceso de iniciadores
propios del método, el cual fue estandarizado para tener la máxima sensibilidad (Shi et al.,
2001). Así mismo se presentan para algunas muestras debido a su origen cloacal y que el
método de purificación no es muy selectivo, si no que extrae RNA total.
Cuadro 2. Muestras obtenidas por Entidad Federativa
Estado muestreados Muestras obtenidas Muestras Positivas
Baja California 10 0
Chiapas 10 1
Jalisco 15 0
Michoacán 10 0
Nayarit 12 0
Oaxaca 13 3
Sinaloa 20 0
Sonora 13 0
Tamaulipas 70 12
Veracruz 17 0
Yucatán 10 0
Resultados
32
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25
Figura 15. Fotografía de un gel de agarosa revelado con luz UV donde se muestran los amplificados obtenidos de las muestras positivas de aves silvestres. Carril 1. Marcador de longitud molecular 1 Kb; 2, 7 y 8 Phalacrocórax auritus, 15 Archilochus colibris, 24 control positivo y 25 control negativo. 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25
Figura 16. Fotografía de un gel de agarosa revelado con luz UV donde se muestran los amplificados obtenidos de las muestras positivas de aves silvestres. Carril 1. Marcador de longitud molecular 1 Kb 11, 15, 18 y 19 Larus delawarensis, 24 control positivo y 25 control negativo.
pb 400 300 200
100
408 pb
408 pb
pb 400 300 200 100

Resultados
33
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25
Figura 17. Fotografía de un gel de agarosa revelado con luz UV donde se muestran los amplificados obtenidos de las muestras positivas de aves silvestres. Carril 1. Marcador de longitud molecular 1Kb; 10,13, 16, 17 y 19 Zenaida macroura, 21, 22 y 23 Larus delawarensis, 24 control positivo y 25 control negativo.
Figura 18. Cormorán bicrestado (Phalacrocórax auritus). En la imagen se muestra un
Cormorán bicrestado, esta especie se reportó como positiva al Von en este trabajo.
Figura 19. Colibrí garganta de rubi (Archilochus colibrís). En la imagen se muestra
un Colibrí garganta de rubí, esta especie se reportó como positiva al VON en este trabajo.
pb 400 300 200 100
408 pb

Resultados
34
Figura 20. Gaviota pico anillado (Larus delawarensis). En la imagen se muestra un gaviota
pico anillado, esta especie se reportó como positiva al VON en este trabajo.
Figura 21. Paloma Huilota (Zenaida macroura). En la imagen se muestra una Paloma huilota,
esta especie se reportó como positiva al VON en este trabajo
Finalmente de los 11 estados de la República Mexicana en donde se recolectaron muestras
para realizar el diagnóstico del VON, sólo en tres estados se encontraron aves positivas al
virus después de realizar la RT-PCR; y correspondieron a dieciséis aves de 4 especies
distintas. En el cuadro 4 se pueden observar las los resultados de la RT-PCR.
Estas especies provienen de los estados de Oaxaca, Chiapas y Tamaulipas (Figura 21). En
estos estados ya se han hecho reportes de la presencia del virus del Oeste del Nilo. En Oaxaca
hay reportes en cocodrilos (Sandoval, 2006), y por parte de la secretaria de Salud es el único
estado donde se tiene reportado un caso en humanos (Secretaria de Salud, 2006). En Chiapas,
hasta diciembre de 2006, la Secretaría de Salud no había reportado ningún caso de Virus del
Oeste del Nilo en aves, mamíferos o en humanos. Sin embargo, otros estudios lo reportan en
cocodrilo (Sandoval, 2006). En el caso de Tamaulipas también se han reportado casos de
cocodrilos infectados (Sandoval, 2006), y hasta diciembre de 2006, la Secretaría de Salud
reporta 26 muestras positivas en equinos en el municipio de Matamoros, 10 en Nuevo Laredo y
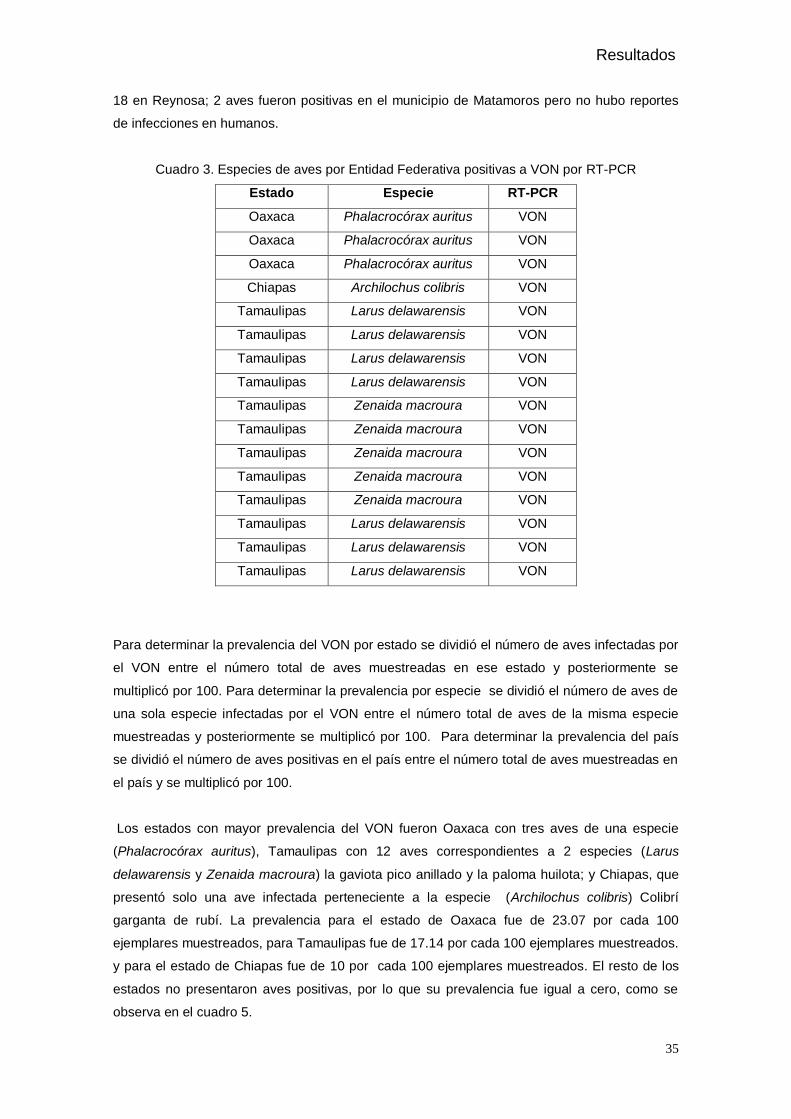
Resultados
35
18 en Reynosa; 2 aves fueron positivas en el municipio de Matamoros pero no hubo reportes
de infecciones en humanos.
Cuadro 3. Especies de aves por Entidad Federativa positivas a VON por RT-PCR
Estado Especie RT-PCR
Oaxaca Phalacrocórax auritus VON
Oaxaca Phalacrocórax auritus VON
Oaxaca Phalacrocórax auritus VON
Chiapas Archilochus colibris VON
Tamaulipas Larus delawarensis VON
Tamaulipas Larus delawarensis VON
Tamaulipas Larus delawarensis VON
Tamaulipas Larus delawarensis VON
Tamaulipas Zenaida macroura VON
Tamaulipas Zenaida macroura VON
Tamaulipas Zenaida macroura VON
Tamaulipas Zenaida macroura VON
Tamaulipas Zenaida macroura VON
Tamaulipas Larus delawarensis VON
Tamaulipas Larus delawarensis VON
Tamaulipas Larus delawarensis VON
Para determinar la prevalencia del VON por estado se dividió el número de aves infectadas por
el VON entre el número total de aves muestreadas en ese estado y posteriormente se
multiplicó por 100. Para determinar la prevalencia por especie se dividió el número de aves de
una sola especie infectadas por el VON entre el número total de aves de la misma especie
muestreadas y posteriormente se multiplicó por 100. Para determinar la prevalencia del país
se dividió el número de aves positivas en el país entre el número total de aves muestreadas en
el país y se multiplicó por 100.
Los estados con mayor prevalencia del VON fueron Oaxaca con tres aves de una especie
(Phalacrocórax auritus), Tamaulipas con 12 aves correspondientes a 2 especies (Larus
delawarensis y Zenaida macroura) la gaviota pico anillado y la paloma huilota; y Chiapas, que
presentó solo una ave infectada perteneciente a la especie (Archilochus colibris) Colibrí
garganta de rubí. La prevalencia para el estado de Oaxaca fue de 23.07 por cada 100
ejemplares muestreados, para Tamaulipas fue de 17.14 por cada 100 ejemplares muestreados.
y para el estado de Chiapas fue de 10 por cada 100 ejemplares muestreados. El resto de los
estados no presentaron aves positivas, por lo que su prevalencia fue igual a cero, como se
observa en el cuadro 5.

Resultados
36
Con respecto a la prevalencia por especie, dos especies compartieron la mayor prevalencia,
el Colibrí garganta de rubí, ya que sólo se monitorearon tres ejemplares de los cuales uno
resultó positivo al Virus del Oeste del Nilo, con lo que la prevalencia fue elevada. La
prevalencia para el Colibrí garganta de rubí (Archilochus colibris) 33 por cada 100 ejemplares
examinados. La segunda especie fue el Cormorán bicrestado (Phalacrocórax auritus) que
tambien presento una prevalencia de 33 por cada 100 ejemplares examinados.
En el caso de la gaviota pico anillado (Larus delawarensis), la prevalencia fue de 28 por cada
100 ejemplares examinados. La menor prevalencia de las especies positivas fue de 27.7 por
cada 100 ejemplares monitoreados y correspondió a la paloma Huilota (Zenaida macroura). La
prevalencia para el país en este trabajo es baja en comparación con la prevalencia por especie
y por estado, siendo de 8 por cada 100 ejemplares monitoreados. En el cuadro 6 se muestran
las prevalencias a la infección por VON en las especies monitoreadas en este trabajo.
Figura 22. Mapa de la República Mexicana donde se representa la presencia del virus del Oeste del Nilo. La figura muestra la presencia del VON en miembros de 4 especies de aves en
tres diferentes estados. Chiapas con una muestra positiva en un Colibrí garganta de rubí (Archilochus colibris), Oaxaca con tres muestras positivas de tres ejemplares de Cormorán bicrestado (Phalacrocórax auritus) y en Tamaulipas con doce muestras positivas correspondientes a siete ejemplares de gaviotas pico anillado (Larus delawerensis) y cinco ejemplares de palomas Huilotas (Zenaida macroura).

Resultados
37
Estas especies ya han sido reportadas como portadoras del Virus del Oeste del Nilo en
Estados Unidos, durante la temporada de transmisión del año 2000 en el estado de Nueva
York (Bernard et al., 2001).
Cuadro 4. Prevalencia obtenida de las distintas Entidades Federativas monitoreadas.
Estado Prevalencia
Baja California 0
Chiapas 10
Jalisco 0
Michoacán 0
Nayarit 0
Oaxaca 23.07
Sinaloa 0
Sonora 0
Tamaulipas 17.14
Veracruz 0
Yucatán 0
Cuadro 5. Prevalencia obtenida de las distintas especies de aves monitoreadas.
Especie Prevalencia
Colibrí Garganta de Rubí.
(Archilochus colibris)
33.33
Cormorán Bicrestado
(Phalacrocórax auritus)
33.33
Gaviota pico anillado
(Larus delawarensis)
28
Paloma Huilota
(Zenaida macroura)
27.7
Los resultados muestran que de las cuatro especies de aves con resultados positivos al Virus
del Oeste del Nilo, dos de ellas; el Colibrí garganta de rubí (Archilochus colibris), y la gaviota
pico anillado (Larus delawarensis) son migratorias. Las otras dos especies, la Paloma Huilota
(Zenaida macroura), y el Cormorán bicrestado (Phalacrocórax auritus), son consideradas aves
residente; por lo tanto la prevalencia comparada entre aves migratorias y residentes es de 1:1.

Discusiones
38
6. Discusiones
Es importante recalcar que las aves son herramientas importantes en el diagnóstico de
enfermedades emergentes y reemergentes ya que son importantes en la salud pública por que
pueden estar infectadas con un número importante de microorganismos patógenos que pueden
ser transmitidos al humano (Reed, et al., 2002), no solo el Virus del Oeste del Nilo, también
otras enfermedades como la influenza y la enfermedad de Lyme. De acuerdo con Rappole et
al., (2000), las aves migratorias han sido vinculadas con el transporte de otros virus
relacionados en el hemisferio occidental y la migración causa un sustancial stress en las aves
el cual puede promover la inmunosupresión y favorecer la replicación del VON, sin que sea
necesaria la presentación de un cuadro clínico ya que en el 80% de los casos, al igual que en
los humanos, las aves son asintomáticos.
La importancia de este estudio radica en que el VON ha sido detectado en más de 300
especies de aves silvestres y domésticas, según el CDC (Medica, et al., 2007), esto es un
problema para la salud pública ya que aproximadamente 6.4 millones de personas en los
Estados Unidos tienen un ave como mascota en casa, lo que aumenta el riesgo de
antropozonosis, incluyendo las metazoonosis (Jorn, et al., 2009)
Por otro lado, como se ha mencionado, el Virus del Oeste del Nilo es un virus transmitido por
mosquitos, principalmente del género Culex, sin embargo hay otros géneros de mosquitos,
como el Aedes, que también pueden participar como vectores del virus (O´Donnell y Travis,
2007).
En México se estima que hay más de 300 especies de mosquitos ornitofilicos, de los cuales
solo se han identificado 207 especies, que son las de interés medico, sin embargo falta mucho
trabajo por hacer con respecto a la identificación taxonómica de otras especies que pueden
participar en el ciclo enzootico de transmisión del virus en zonas selváticas como las que hay
en diversas zonas de nuestro país, para poder conocer los posibles vectores del VON, así
como la interrelación con los distintos reservorios y amplificadores como es el caso de los
cocodrilos y las aves. De acuerdo con Ibáñez (2009), primero se requiere realizar una
clasificación taxonómica de los mosquitos en las áreas con prevalencia de metazonosis para
conocer cual es el vector o cuales son los vectores ya que cada artrópodo tiene su propio
comportamiento.
Si bien no se sabe con exactitud que otros especies de mosquitos podrían participar como
vectores en México si se puede realizar un monitoreo del VON en aves ya que son el reservorio
y hospedero natural del virus.

Discusiones
39
En este trabajo se utilizó la infraestructura del monitoreo de Influenza aviar ya que es una
enfermedad emergente que cuenta con recursos económicos a partir de la cual se pueden
aprovechar las muestras para utilizarla en el diagnóstico de otra enfermedad emergente de
menos importancia en la salud publica de México como es el VON, ya que el método de
muestreo para influenza aviar es el mismo método de hisopados cloacales y orales descrita
por Komar et al., (2002), para el diagnóstico del virus del Oeste del Nilo ya que hace el trabajo
más fácil, rápido, eficiente y seguro.
Como se comentó anteriormente, las especies positivas al VON obtenidas en este trabajo ya
habían sido reportadas como positivas en los Estados Unidos en el 2000 (Bernard et al., 2001);
y a pesar de que son especies menos susceptibles que los córvidos también pueden presentar
la infección.
Según van der Meulen, et al., (2005), las especies que pertenecen al Orden Charadriiforme,
como es el caso de la gaviota pico anillado (Larus delawarensis), son más susceptibles a la
enfermedad a diferencia de los Psittacidos y Gallinaceos. Sin embargo, no presentan las tasas
de morbilidad y mortalidad que los córvidos ya que a pesar de que se han reportado varias
aves de este orden como positivas al VON no han desarrollado cuadros severos e incluso, en
muchos casos son asintomáticos.
Con respecto a la prevalencia para del VON en el país, fue baja en comparación con otros
trabajos. En este estudio se obtuvo una prevalencia de 8 por cada 100 ejemplares
muestreados a diferencia de lo reportado en otros países como Nigeria, en donde la tasa de
prevalencia es de 60 por cada 100 ejemplares muestreados, esto podría deberse a que en
África es una zona endémica de VON (van der Meulen et al., 2005),
De las cuatro especies de aves positivas, dos, la gaviota pico anillado (Larus delawerencis) y
la paloma huilota (Zenaida macroura), se reportaron en el estado de Tamaulipas. Este estado
esta íntimamente relacionado con la ruta migratoria del Golfo y del Atlántico, por la cual migran
aves de la costa este de los Estados Unidos, zona en la cual el Virus del Oeste del Nilo esta
reportado en distintas especies de láridos (Figura 23).
A pesar de que las palomas huilotas están consideradas como residentes en México, su
distribución comprende además varios estados de los Estados Unidos, como se observa en la
figura 24, lo que favorece su interacción con otras especies de aves que podrían estar
infectadas con el VON ya que realizan migraciones de distancias cortas en busca de alimento
(Reed, et al., 2002).

Discusiones
40
Además en el estado de Tamaulipas ya hay reportes previos de la presencia del VON; en
noviembre del 2002 la Secretaría de Salud confirmó la presencia del Virus del Oeste del Nilo en
un equino de este estado por medio de serología.
También en 2002, Estrada, et al., (2003); reportó cuatro caballos positivos al VON en
Tamaulipas, después de realizar estudios filogenéticos se comprobó que las cepas aisladas de
los Caballos estaban íntimamente relacionadas con las cepas aisladas en Texas durante la
temporada de infección, lo que significa que tiene un alto porcentaje de similitud, lo que hace
suponer que posiblemente esta cepa provenga de la cepa de Texas.
Figura 23. Área de distribución de la Gaviota pico anillado(Larus delawerensis). En color
rojo se muestra el Área de reproducción y anidamiento, el área azul es en donde se encuentra en invierno, el área en amarillo es la ruta por la que migra durante el invierno y el área en color verde muestra la zona en donde se encuentran por migraciones erráticas (Base de datos de aves del mundo, 2009, 2009).
Actualmente hay un programa de vacunación contra VON para equinos en el estado de
Tamaulipas, sin embargo la SAGARPA no realiza un monitoreo contra esta enfermedad y no
tiene reportes de seroprevalencias en este estado. Sin embargo de acuerdo con Medica et al
(2007), la razón por la cual no se reportan casos de VON en los países latinoamericanos, es la
deficiencia de sus métodos de diagnóstico por parte de los centros de salud.

Discusiones
41
La prevalencia obtenida en este estudio para el estado de Tamaulipas fue de 17.14 por cada
100 ejemplares, lo que se alta con respecto a la obtenida en otros trabajos; por ejemplo para la
prevalencia obtenida en 2006 por la Secretaria de Salud para el Estado de Tamaulipas fue de
1.36 positivos por cada 100 ejemplares analizados. Estos resultados se deben a que debido
que el tamaño de muestra es obligado no se monitoreó un gran numero de aves, por otro lado
las aves que se monitorearon son aves que se han reportado con anterioridad como
susceptibles a la infección por el VON lo que amplía la probabilidad de encontrar muestras
positivas.
Figura 24. Área de distribución de la Paloma Huilota (Zenaida Macroura). En color verde claro se muestra el Área en donde se encuentra en verano, el área azul es en donde se encuentra en invierno y el verde oscuro es la zona en donde se encuentra todo el año (Base de datos de aves del mundo, 2009).
La tercer especie positiva al VON en este trabajo se reportó en el estado de Oaxaca y fue un
Cormorán bicrestado (Phalacrocorax auritus); en el caso de este ejemplar es incierta su ruta
de entrada ya que debido a la distribución geográfica de nuestro país, lo que corresponde al
sureste, juega el papel de un embudo en donde convergen las cuatro rutas migratorias, por lo
cual sería necesario realizar estudios filogenéticos para inferir la ruta de entrada de dicho
virus, ya que a pesar de que el Cormorán bicrestado esta reportado como especie residente en
la zona de Oaxaca, interactúa con otras aves que podrían estar infectadas con el virus y de
esta forma infectarse ya que se ha demostrado que la transmisión del virus de ave a ave es
posible, también se ha reportado que las aves rapaces se pueden infectar con el virus cuando
se alimentan de presas infectadas como aves, mamíferos y ratas (Komar, 2000) por lo que es
importante conocer la etología de las especies. En la figura 25 se muestra la distribución del
Cormorán bicrestado (Phalacrocorax auritus).
La cuarta especie positiva al VON en este trabajo se reportó en el estado de Chiapas y fue un
colibrí garganta de rubí (Archilochus colibris); en el caso de este ejemplar, al igual que en los
Cormoranes bicrestados, es incierta la ruta de entrada ya que como se mencionó

Discusiones
42
anteriormente, debido a la distribución geográfica de nuestro país, convergen las cuatro rutas
migratorias, por lo cual también sería necesario realizar estudios filogenéticos para inferir la
ruta de entrada de dicho virus. En la figura 26 se muestra la distribución del colibrí garganta
de rubí (Archilochus colibris). Sin embargo, tomando en cuenta la etología del Colibrí garganta
de rubí se podría intuir que entró por la ruta del centro o del golfo, aunque es importante
mencionar que hay una gran interacción entre las aves durante su ruta migratoria y este
ejemplar se pudo haber infectado con el virus en cualquier zona que abarque su ruta
migratoria.
Figura 25. Área de distribución del Cormorán bicrestado (Phalacrocorax auritus) En color verde se muestra el Área de reproducción y anidamiento, el área rosa es en donde se encuentra accidentalmente (Base de datos de aves del mundo, 2009).
En Chiapas, al igual que en Tamaulipas y Oaxaca había reportes de la presencia del virus. En
el 2006 se reportó la presencia del virus en un cocodrilo (Crocodylus moreletti), en cautiverio
(Sandoval, 2006). Estos datos coinciden con los obtenidos en este trabajo con respecto a la
presencia del virus en estos estados, lo que demuestra que el virus ha estado circulando en
esta región por lo menos desde el 2006, año en el que se reportó el cocodrilo positivo, sin
embargo no hay reportes por parte de la secretaria de Salud. En el caso de las aves no se ha
realizado monitoreo y en el caso de equinos solo se monitoreó un ejemplar y el resultado fue
negativo, lo que refleja la mala organización del diagnóstico epidemiológico en el país por parte
de la Secretaria de Salud, tomando en cuenta que el VON es una enfermedad emergente de
reporte obligatorio
Estudios realizados en el sureste mexicano han reportado la presencia del VON en estados
vecinos a Chiapas; por ejemplo, en 2003 se hace el primer aislamiento de una cepa en un
cuervo del zoológico de Yum-Ka en el estado de Tabasco. En 2005, se reportaron aves,
mamíferos, mosquitos y reptiles seropositivos al VON en el parque zoológico la Venta y en el

Discusiones
43
parque zoológico Yum-Ka de la ciudad de Villa Hermosa Tabasco (Hidalgo, et al., 2008). Si
tomamos en cuenta la interacción de los mosquitos con las aves en estos dos zoológicos
donde las aves silvestres en cautiverio interactúan con las aves silvestres, se esperaría que el
virus se haya esparcido a los estados vecinos por medio de las rutas migratorias de las aves
en vida libre, como seria el caso del colibrí garganta de rubí en Chiapas, esparciéndose a
nuevos nichos ecológicos (Dupuis et al., 2005).
En 2006, se reporta que la cepa TM171-03, aislada en Tabasco en 2003 es parecida a una
cepa aislada en Florida en 2001, a otra aislada en Lousiana en 2002 y un poco distinta a la
aislada en Nueva York en 2000. A través de este estudio se propuso que la cepa TM177-03
entro a México por la migración de aves del sureste de los Estados Unidos al golfo de México o
al Caribe pasando por México (Dardorff et al., 2006). Por lo anterior, es recomendable la
realización de aislamientos virales para conocer la filogenia del virus y proponer las distintas
rutas de entrada del virus según su porcentaje de similitud de las cepas.
Figura 26. Área de distribución del Colibrí garganta de rubí (Archilochus colibris). En color rojo se muestra el Área de reproducción y anidamiento, el área azul es en donde se encuentra en invierno y el área en amarillo es la ruta por la que migra durante el invierno (Base de datos de aves del mundo, 2009).

Discusiones
44
La prevalencia para el estado de Chiapas fue de 10 por cada 100 ejemplares, al igual que en el
estado de Tamaulipas, es una prevalencia alta debido a las pocas aves monitoreadas, ya que
en el trabajo con especies de fauna silvestre la toma de muestras es obligada. Con respecto a
la prevalencia por especie no hay datos en México con los que se puedan comparar, sin
embargo se han reportado prevalencias que van desde .5 hasta 71 por cada 100 ejemplares,
esto depende de la temporada del año en que se realice el estudio, la cantidad o disponibilidad
del vector, la especie que es monitoreada y la zona de monitoreo (van der Meulen, et al., 2005)
Por ejemplo; los brotes entre humanos a menudo ocurren en las personas que viven cerca de
tierras húmedas donde hay altas concentraciones de aves que entran en contacto con grandes
números de mosquitos ornitofílicos (Rappole et al., 2000). También se han e han realizado
estudios que demuestran que del 75 al 100% de los mosquitos Culex tarsalis que se han
consumido sangre infectada, con concentraciones de 107.1
UFP/ml, pueden infectar a las aves y
del 0 al 36% con concentraciones de 104.9
UFP/ml (Hayes, et al., 2005a).
Como se ha mostrado en este estudio las aves migratorias son importantes en la dispersión y
amplificación del VON, un ejemplo claro es el reporte de la presencia del virus en Cuba y
Puerto Rico, en el caso de Cuba dos aves fueron residentes (66.66%) y una migratoria
(33.33%) y en el caso de Puerto Rico diez aves representantes de seis especies fueron
positivas; un ave fue residente (10%) y nueve aves migratorias (90%), como se observa la
mayor porcentaje de aves positivas fueron migratorias a diferencia de lo obtenido en este
trabajo en donde las especies de aves migratorias y residentes positivas al VON presentaron el
mismo porcentaje (50%). Las aves de Cuba y puerto Rico, pertenecen a especies migratorias
que viajan por la ruta del Atlántico lo que demuestra el papel tan importante que juegan las
aves en el esparcimiento del VON a nuevos nichos ecológicos (Dupuis et al., 2005).
Se debe recalcar que un punto íntimamente ligado con la tasa de prevalencia de VON, no solo
en aves, si no también en humanos y otros mamíferos, es el comportamiento defensivo del
hospedero contra los mosquitos ya que este comportamiento puede facilitar o dificultar la
transmisión del VON por vectores infectados, como también es cierto que la transmisión del
virus es facilitada si un mosquito infectado se alimenta de múltiples hospederos que no se
defienden. Con respecto a las especies de aves positivas al VON en este trabajo, no hay
reportes del comportamiento defensivo que nos brinden un punto de partida o comparación
para saber como influye en la prevalencia del VON pero con respecto a otras especies hay
diversos estudios que han demostrado que la alimentación de los mosquitos se reduce cuando
aumenta la respuesta defensiva de los vertebrados como las palomas (Columba liva), las
cuales pertenecen al mismo genero y familia que la paloma Huilota (Zenaida macroura),
especie reportada positiva al VON en este trabajo. Los gorriones domésticos (Passer
domesticus), conejos, (Oryctolagus cuniculus), ardilla gris, (Sciurus carolinensis), ratón de los
campos de algodón (Pecomysais gossypinus) y el hombre (Homo sapiens), son otras especies

Discusiones
45
donde se a estudiado el comportamiento defensivo del huesped (Darbroand and Harrington,
2007).
En México, a pesar de que ya se ha reportado en distintas especies de animales, el VON no ha
causado las grandes mortalidades que en Estados Unidos, por citar un ejemplo; en Georgia, la
principal causa de muerte en aves rapaces es el VON, presentando una tasa de mortalidad de
10% con un rango del 3 al 30% (Ellis, et al., 2007). A pesar de que en general la prevalencia ha
sido baja y se han reportado casos en humanos, la infección por el VON no ha causado la
mortalidad y morbilidad que en otros países; posiblemente por la presencia de otros flavivirus
en el país como el caso del virus del Dengue y la Fiebre Amarilla, lo que podría causar un
cruce antigénico.
En el 2007 el pico de infección en las aves en los Estados Unidos se reportó durante la
primera semana de septiembre y nuevamente los córvidos fueron la especie más susceptible
con 1690 aves muertas que corresponden al 77% del total de aves muertas, mientras que en
México no se reportó ningún caso de mortalidad en aves atribuido al VON (Lindsey et al.,
2008), sin embargo, la obtención de muestras de este estudio fue a partir del mes de octubre, y
si tomamos en cuenta que las aves comienzan su migración en septiembre por lo que se
pudría suponer que las aves infectadas no migren, o si migran no lleguen a su destino ya que
el viaje las inmunosuprime y podrían morir en el trayecto o recuperarse y de este modo llegar a
su sitio de invernación sin los niveles suficientes de viremia para poder infectar a los mosquitos
vectores (Rappole et al., 2000, Reed et al., 2003).
La baja prevalencia de este virus puede estar asociada con cierto grado de protección hacia la
población en zonas endémicas del dengue ya que gran parte de ella tiene anticuerpos
antidengue que podrían conferir protección cruzada temporal o casos menos severos (Ramos y
Falcón, 2004), no solo en México sino también en el resto de Latinoamérica y el Caribe, ya que
hay condiciones de transmisión de arbovirosis en estas áreas (Hayes et al., 2005a), de este
modo e hipotéticamente, aunque las aves sean portadoras del VON y los vectores se alimenten
de los huéspedes humanos o mamíferos no se presentaría la infección. En resumen, la baja
incidencia del VON en México se debe a la resistencia de la población humanamexicana,
posiblemente a través de la inmunidad cruzada por las infecciones de flavivirus como el
Dengue (Dardorff, et al., 2006).
El riesgo potencial para México, América Central, el Caribe y el Sur de América está latente
hoy día. A pesar de que no se han presentado epidemias en humanos ni en equinos el virus ya
se ha reportado, en México, Cuba, Puerto Rico, Republica Dominicana, Bahamas y Argentina,
lo que habla de la rapidez con la que el virus se ha esparcido por las aves silvestres
migratorias, ya que como se ha reportado previamente (Reed et al., 2003;Dazac y Huabalek,

Discusiones
46
2006), las aves son los principales transportadores naturales de patógenos a través d e sus
rutas migratorias.
De acuerdo con lo anterior, minimizar los efectos del virus en humanos constituye una actividad
de primera línea que sólo será posible a través de un monitoreo intensivo y continuo de las
concentraciones de aves migratorias estableciendo un sistema de detección útil y sensible
para el Virus del Oeste del Nilo como ya se ha realizado y se esta realizando en estados como
Colorado en los Estados Unido (Nemeth et al., 2007). Para poder hacer frente a una posible
epidemia del Virus del Oeste del Nilo es importante conocer la historia natural de la
enfermedad para poder tomar medidas de control, es decir, realizar la prevención primaria
para no entrar en el periodo patogénico.

Conclusiones
47
7. Conclusiones
1. La prevalencia de VON para el país fue de 8 por cada 100 ejemplares monitoreados.
2. La especie de ave con mayor prevalencia al VON fue el Cormorán Bicrestado
(Phalacrocórax auritus) y el colibrí garganta de Rubí (Archilochus colibris) con 33.33 por
cada 100 ejemplares examinados.
3. El estado con mayor prevalencia al VON fue el estado de Oaxaca con una prevalencia de
23.07 por cada 100 ejemplares examinados.
4. El número de especies residentes y migratorias positivas al VON fueron iguales, dos
especies en cada caso, por lo que la prevalencia comparada fue de 1:1.

Anexo I
48
8. Referencias
1. Aguilar V.B.1998 Hábitat y abundancia de anátidos en el lago de Cuitzeo, Michoacán,
Tesis de Maestría del colegio de postgraduados, pags 1-60.
2. Allan, B. F., Langerhans, R. B., Ryberg, W. A., Landesman, W. J. Griffin, N. W., Katz,
R. S., Oberle, B. J., Schutzenhofer, M. R., Smyth, K. N., de St Maurice, A., Clark, L.,
Crooks, K. R., Hernandez, D., McLean, R. G., Ostfeld, R. S. and Chase, J.M. 2007.
Ecological Correlates of Risk and Incidence of West Nile Virus in the United States.
Community Ecology. Oecología. 0029-8549 (Print) 1432-1939 (Online)
3. Barriga, G. A., Arumir, C. E. y Mercado, G. F. 2002. Actualidades sobre la Fiebre del
Nilo Occidental. Revista Mexicana de Patología Clínica 49(4): 203-211.
4. Base de datos de aves del mundo. (Publicación en línea) 2009. Hallado en
URL:http://avibase.bsceoc.org/species.jsp?lang=ES&avibaseid=DEC3D8D0C74C598B
&sec=natureserve
5. Beasley, D.W.C., Davis C.T., Estrada, F.J., Navarro, L.R., Campomanes, C.A., Tesh,
R.B., Weaver, S.C. and Barrett, D.T. 2004. Genome Secuence and Attenuating
Mutations in West Nile Virus from Mexico. Emerging Infectious Diseases 10(12):2221-
2224.
6. Bernard, K. A., Maffei, J. G., Jones, S. A., Kauffman, E. B., Ebel, G. D., Dupois, A. P.,
Ngo, K. A., Nicholas, D. C., Young, D. M., Shi, P. Y., Kulasekera, V. L., Eidson, M.,
Vhite, D. J., Stone, W. B., Ny State West Nile Virus Surveillance Team and Kramer
L.D. 2001. West Nile Virus Infection in Birds and Mosquitoes, Ney York State, 2000.
Emerging Infectious Disease 7(4): 674-685.
7. Blitvich, B. J., Bowen, R. A., Marlenee, N. L, Hall, R. A., Bunning, M. L. and Baety, B. J.
2003. Epitope-Bloking Enzyme Inmunosorbent Assays for Detection of West Nile Virus
Antibodies in Domestic Mammals. Journal of Clinical Microbiology 41 (6): 2676-2679.
8. Campbell, G.L., Marfin, A. A., Lanciotti, R. S. and Gubler, D. J. 2002. West Nile virus.
Lancet Infect Disease 2: 519-529
9. Cárdenas, L. J. y Vargas, G. R. 2002. Epidemiología de la EON en Equinos en los
EE.UU. Imagen Veterinaria 2(8): 22-25.

Anexo I
49
10. Carrada, B., T. 2004. Encefalitis por Virus Nilo – Oeste: Epidemiología, ecología,
diagnóstico y prevención. Revista Mexicana de Patología Clínica. 51 (1): 6-15.
11. Centro Nacional de Vigilancia Epidemiologica y Control de Enfermedades. (Publicación
en línea) 2009. Hallado en URL http:www.cenave.com/von/default.asp?id=24 .
12. Chung, K. M., Nybakken, G. E., Thompson, B. S., Engle, M. J., Marri, A. Fremont, D. H.,
and Diamond, M. S. 2006. Antibodies Against West Nile Virus Nonstructural Protein
NS1 Prevent Lethal Infection Trough Fc y Receptor Dependent and Independent
Mechamisms. Journal of Virology 80(3):1340-1351.
13. Dabanch, P. J.2003. Zoonosis. Revista Chilena de Infectología. 20 (1):47 – S51
14. Daszac, P., Cunningham, A.A., Hyatt, A. D. 2000 Emerging Infectious Diseases of
Wildlife – Threats to Biodiversity and Human Health. Review Wildlife Ecology.
Science´s Compass Vol. 287 443- 449.
15. Dardorff, E., Estrada, F.J.G., Brault, A.C., Navarro, L.R., Campomanes, C.A., Paz, R.P.,
Solis, H.M., Ramey, W.N., Davis, C.T., Besley, D.W.C., Tesh, R.B. Barret, A.D.T. and
Weaver, S.C. 2006. Introduction of West Nile Viirus Strains to Mexico. Emerging
Infectious Diseases 12(2): 314 -318.
16. Darbro,M.J. and Harrington, L.C. 2007 Avian defense Behavior and blood-feeding
success of West Nile Vector Mosquito, Culex pipiens. Behavioral Ecology 18:750-757.
17. Davis, A., Bunning, M, Gordy, P., Panella, N., Blitvich, B. and Bowen, R. 2005.
Experimental and Natural infection of North American Bats with West Nile Virus.
American Journal Tropical Medicine and Hygiene. 73(2): 467-469.
18. Díaz, L.M., Komar, N., Visintin, A., Dantur, J,M.J., Stein, M., Lobo, A.R., Spinsanti, L.,
Konigheim, B., Aguilar, J., Laurito, M., Almirón, W. and Contigiani M. 2008. West Nile
Virus in Birds, Argentina. Emerging Infectious Diseases. 14(4): 689-691.
19. Dupuis, A., Marra, P., Reitsma, R., Jones, M. Louis, K and Kramer, L. 2005. Short
report: Serologic Evidence for West Nile Virus Transmition in Puerto Rico and Cuba.
Journal Tropical of Medicine and Hygiene. 73 (2): 474-476.

Anexo I
50
20. Ellis, A.E., Mead, D.G., Allison, A.B., Stallknecht, D.E. and Howerth, E.W. 2007.
Pathology and Epidemiology of Natural West Nile Viral Infection of Raptors in Georgia.
Journal of Wildlife diseases 43(2):214-223.
21. Estrada, F.J.G., Navarro, L.R., Beasley, D.W.C., Coffey, L., Carrara, A.S., Travassos
da Rosa, A., Clements, T., Wang, E., Ludwing,G.V., Camponês, C.A., Paz, R.P.,
TeshR.B., Barret, A. and WeaverS.C. 2003. West Nile Virus in Mexico: Evidence of
Widespread Circulation since July 2002. Emerging Infectius Diseases 9 (12): 1604-
1607.
22. Farfán, A.J.A., Bradley, J.B., Blitvich, N.L., Marlenne, M. A., Loroño, P.A., Puerto, M. F.,
Flores, F. L. F., García , R. J. E., Ortega, S.A., Chávez, M. J., Cremieux, G.J.C.,
Correa, M .F., Hernández, G. G., Méndez, G. J. F. and Beaty, B. J. 2006. Antibodies to
West Nile Virus in Asyntomatic Mammals, Birds and Reptiles in the Peninsula of
Yucatán. American Journal Tropical Medicine and Hygiene 74(5): 908-914.
23. Fernández, S.I., Garza, R. M de L., Barry, J.B., Ramos, J. J. y Rivas, E. M. 2007.
Presencia del Virus del Oeste del Nilo en el Noreste de México. Salud Pública de
México. 49(3):210-217.
24. García, E.G. 2007 Aves silvestres en el surgimiento de enfermedades Infecciosas:
Escenario en México. Memorias de las XIII Jornadas medico Avícolas p. 59-63.
25. Gea- Banacloche et. al., 2004. West Nile Virus: Pathogenesis and therapeutic options.
Ann. Int. Med. 140(7): 545-554.
26. Girard, Y.A., Schneider, B.S., McGee, C.E., Wen, J., Han, V.C., Popov, V., Mason,
P.W. and Higgs, S. 2007. Salivary Gland Morphology and Virus Transmission During
Long-Term Cytopathologic West Nile Virus Infection in Culex Mosquitoes. American
Journal Medicine Tropical and Hygiene 76(1):118-128.
27. Greer, A., Ng BS, V., and Fisman, D. 2008. Climate Change and Infectius Diseases in
North America: The Road ahead. Canadian Medical Association Journal178(6): 715-
722.
28. Hall, D. A., Tyler, K. L., Frey, K. L. and Kozora, E. 2008. Journal Neuropsychiatry
Clinical Neurosci. 20 (1): 122-123.

Anexo I
51
29. Hayes, E.B., Komar, N., Nasci, R.S., Susan, P., O´Leary, D. and Campbell, G.L. 2005
Epidemiology and Transmission of West Nile Virus Disease. Emerging Infectious
Diseases 11(8): 1167-1173.
30. Hayes, E. B., Sejvar, J. J., Saki, R. S., Lanciotti, R. S., Bode, A. V. and Campbell, G.L.
2005.Virology, Phatology and Clinical Manifestatio of West Nile Virus Disease.
Emerging Infectious Disease 11(8): 1172-1179.
31. Hidalgo, M.A., Puerto, F., Farfan, A.J.A., Gárcia, R.J.E., Rosado, P.E., Méndez, G.J.,
Figueroa, O.R., Takashima, I. y Ramos,C. 2008 Prevalencia de Infección por el virus
del Nilo Occidental en dos Zoológicos del Estado de Tabasco. Salud Publica de
México. 50(1):76-85.
32. Jawetz, Melnick y Adelberg, 2002. Microbiología Médica. Manual moderno 17ª edición.
p. 363,365,547-562.
33. Jorn, M.S., Thompson, K.M., Larson, J.M. and Blair, J.E. 2009 Polly can make you sick:
Pet bird-associated diseases. Cleveland Clinic Journal of Medicine 76(4). 235-243.
34. Komar, N. 2000 West Nile Viral Encephalitis. Rev. Sci. Teech Off. Int. Epiz. 19:166-
176.
35. Komar, N., Lanciotti, R., Bower, R., Langevin, S. and Bunning, M. 2002. Detection of
West Nile Virus in Oral and Cloacal Swabs Collected From Birds Carcasses. Emerging
Infectius Disease 8(7): 741-742.
36. Komar, N., Panella, N. A., Langevin, S. A., Brault, A. C., Amador, M., Edwards, E. and
Owen, J.C. 2005. Avian Host For West Nile Virus In St. Tammany Parish, Lousiana,
2002.American Journal Medicine Tropical and Hygiene. 73(6): 1031-1037.
37. Kramer, L..D., Li,J and Shi, P.2007 West Nile Virus. Lancer Neurology 6:171-181.
38. Lanciotti, R. S., Roehrig, J. T., Deubel, V. Smith, J. M. Parker, M. Steele, K. Crise, B.
Volpe, K. E., Crabtree, M. B., Scherret, J. H. Hall, R. A., MacKenzie, J. S., Cropp, C.
B., Panigrahy, B., Ostlund, E., Schmitt, B., Malkinson, M., Banet, C., Weissman, J.,
Komar, N. N. Savage, N., Stone, W., McNamara, T. and Gubler, D. J. 1999. Origin of
the West Nile Virus Responsible for an Outbreak of Encephalitis in the Northeastern
United States. Science, 286: 2333.

Anexo I
52
39. Lanciotti, R.S. 2003. West Nile Virus Genetics and Diagnostic Methods. Plan to attend
ICAAC ASM´s Premier Annual Meeting on Infectious Diseases.
40. Lanciotti, RS; Kerst AJ, Nasci, RS, Godsey MS, Mitchell CJ, Savage HM, Komar N,
Panella NA, Allen BC, Volpe KE Davis BS, Roehrig JT, 2000. Rapid detection of West
Nile virus from human clinical specimens, field-collected mosquitoes, and avian
samples by a Taq Man reverse transcriptase-PCR assay. J Clin Microbiol 38: 4066-
4071.
41. Lanciotti, R.S., Roering, J. T., Deubel, V., Smith, J., Parker, M., Steele, K., Crise, B.,
Volpe, K. E., Crabtree, M. B., Scherret, J. H., Hall, R. A., MackenzieJ. S., Cropp, C. B.,
Panigrahy, B., Ostlund, E., Schmitt, B., Malkinson, M., Banet, C., Weissman, J., Komar,
K., Savage, H. M., Stone, W., Mcnamara, T. and Gubler, D.J. 1999. Origin of the West
Nile Virus Responsible for the Outbrake of Encephalitis in the Northeastern United
States. Science 286: 2333-2337.
42. Lindsey, N.P., Hayes, E.B., Nasci, R.S., Komar, M., Petesen, Z. R. and CDC. 2007.
West Nile Virus Activity United States, 2006 . JAMA 298 (6): 619-621.
43. Lindsey, N.P., Lehman, J.A., Staples, J.E., Komar, M., Zielñinzki-Gutierrez, E., Hayes,
E.B., Nasci, R.S. and Fischer, M. 2008. West Nile Virus Activity United States, 2007
JAMA 300 (12): 1406-1408
44. McGee, C.E., Schneider, B.S., Girard, Y.A., Vanlandingham, D.L. and Higgs, S. 2007.
Nonviremic Transmission of West Nile Virus: Evaluation of the Efects os Space, Time
and Mosquito Species. American Journal Tropical Medecine and hygiene 76(3): 424-
430.
45. McMurtrey, C., Lelic, A., Piazza, P., Chakrabarti, A. K., Yablonsky, E. J., Wahl, A.,
Bardet, W., Eckerd, A., Cook, R. L., Hess, R., Buchil, R., Loeb, M., Rinaldo, C. R.,
Bramson, J. and Hildebrand, W. H. 2008. Proccedings of the National Academy of
Science 15 (8): 2981-2986.
46. Medica , D.L., Clauser, R. and Bildstein. 2007. Prevalence of West Nile Virus Antibodies
in a Breeding Population of American Krestel (Falco sparverius) in Pennsylvania.
Journal of Wildlife Diseases 43(3): 538.541.

Anexo I
53
47. Nemeth, N., Beckett, S., Edwards, E., Klenk, K. and Komar, N. 2007. Avian Mortality
Survilance for West Nile Virus in Colorado. American Journal Medicine and Hygiene
76(3): 431-437.
48. Nielsen, C.F., Armijo, M.N., Wheeler, S., Carpenter, T.E., Boyce, W.M., Kelley, K.,
Brown, D., Scott, T.W. and Reisen, W.K. 2008. Risk Factors Associated with Human
Infection during the 2006 West Nile Virus Outbreak in Davis, a Residential Community
in Northern California. American Journal Medicine Tropical and Hygiene 78(1):53-62.
49. O´Donnell, C. R. and Travis, D.A. 2007 West Nile Virus. Int. Zoo Yb. The Zoological
Society of London. 41: 75-84.
50. Ǿlberg,R.M., Barker, I.K., Crawshaw, G.J., Bertelsen, M.F., Drebot, M.A. and Andonov,
M. 2004. Emerging Infectius Diseases 10(4): 712-714.
51. Papperell, C., Rau, N., Krajden, S., Kern, R., Humar, A., Mederski, B., Simor, A., Low,
D. E., McGeer, A., Mazzulli, T., Burton, J., Jaigobin, C., Fearon, M., Artsob, H., Drebot,
M. A., Halliday, W. and Brunton, J. 2003. West Nile Virus Infection in 2002: Morbidity
and Mortality Among Patients Admitted to Hospital in Southcentral Ontario. Canadian
Medical Association or its Licensors Journal. 168(11): 1399-1405.
52. Petersen, L. R. and Roehrign, J. T. 2001. West Nile Virus: a reemerging global
pathogen. Emerging Infectius Disease. 7: 611-614.
53. Rappole, J. H.. Derrickson, S.R. and Hubálek, Z. 2000. Migratory Birds and Spread of
West Nile Virus in the Western Hemisphere. Emerging Infections Diseases 6 (4): 319-
328
54. Rappole, J. H. and Hubálek, Z. 2003. Migratory Birds and West Nile Virus. Journal of
Applied Microbiology 94: 47S-58S
55. Ramos, C. y Falcón, L.J.A. 2004.La fiebre del Nilo Occidental: uNa realidad en México.
Salud Pública de México46(5): 488-490.
56. Reed, K. D., Mecce, J. K., Henkel, J. S. and Shukla, S. K. 2003. Birds, migration and
emerging zoonoses: West Nile Virus, Lyme disease, Influenza A and Enteropathogens.
Clinical Medicine and research 1 (1): 5-12

Anexo I
54
57. Saito, E.K., Sileo, L., Green, D.E., Meteyer, C.U., Mclaughlin, G.S., Converse, K.A. and
Docherty,D.E. 2007. Raptor Mortality Due to West Nile Virus in the United States, 2002.
Journal of Wildlife Diseases43(2):206-213.
58. Sandoval, S. A. L. 2006. Virus del Oeste del Nilo en México. Vida Silvestre. Tesis
Doctoral. Instituto Politécnico Nacional. p. 9-18.
59. Sbrana, E., Tonry J.H., Xiao,S.Y., Trvasos da Rosa, A.P.A., Higgs, S. and Tesh, R.B.
2005. Oral Transmition of West Nile Virus a Hamster Model. American Journal Tropical
Medicine and Hygiene. 72(3): 325-329.
60. Shi, P. Y., Kauffman, B., Ren, P., Felton, A., Tai, H., Dupuis, A. L., Jones, S. A., Ngo, K.
A., Nicholas, D. C., Maffei, J., Ebel, G, D., Bernard, K. A. and Kramer, L. D. 2001. High-
Troughput Detection of West Nile Virus RNA. Journal of Clinical Microbiology. 39(4):
1264-1271.
61. Shrestha, B. and Diamond, M.S. 2007. Fas Ligand Interactions Contribute to CD8+ T-
Cell-Mediated Control of West Nile Virus Infection in the Central Nervous System.
Journal of Virology 81(21): 11749-11757.
62. Snapinn,K.W., Holmes, E.C., Young, D.S., Bernard, K.A. Kramer, L.D. and Ebel, G.D.
2007. Declining Growth Rate West Nile Virus in North America. Journal of Virology 81
(5): 2531-2534.
63. Téllez, I., Calderón, O., Franco, P.C. y Del Río, C. 2006. El Virus del Oeste del Nilo:
Una realidad en México. Gac. Med. Mex. 142 (6): 493-499.
64. Torres, A. j., Garcia, r. E., Stramer, S., Casanova, R., Foster, G., Luce, R., Tomashek,
K., Mohammed, H., Sun, W., Hunsperger, E., Muños-Jordan, J.L., Colon, C., Vergne, E.
and Verduin, M. 2008 Detection of West Nile Virus in Blood Donations- Puerto rico,
2007.JAMA 300 (11): 1293- 1294.
65. Tsao J. I., Wootton, J. T., Buñikis; J., Luna, M. G. Fish D. and Barbour A. G., 2004. An
Ecological Approach to preventing human infection: Vaccinating wild mouse reservoirs
intervenes in the Lyme disease cycle. PNAS 101(52): 18159-18164.
66. van der Meulen, K.M., Pensart, M.B. and Nauwynck, H.J. 2005. West Nile Virus in the
Vertebrate Word. Archives of Virology 150:637-657

Anexo I
55
67. Venkatesan, M., Westbrook, C.J., Hauer, M.C. and Rasgon, J.L. 2007. Evidence for
Population Expansion in the West Nile Virus Vector Culex tarsalis. Molecular Biology
Evolution 24(5):1208-1218.
68. Whitley, R. J. 2003. West Nile virus clinical overview and prospects for development of
therapeutics and a human vaccine. Program and abstracts of the 43rd Annual ICAAC;
September 14-17, 2003; Chicago, Illinois. Abstract 1769
69. Zepeda, H. 2006 Zoonosis y salud pública. Memorias del primer curso de actualización
en medicina, manejo y conservación de fauna silvestre. p. 52 – 53.