decoupling the influence of leaf and root hydraulic conductances
TRANSCRIPT

Decoupling the influence of leaf and root hydraulicconductances on stomatal conductance and its sensitivityto vapour pressure deficit as soil dries in a drainedloblolly pine plantation
JEAN-CHRISTOPHE DOMEC1, ASKO NOORMETS1, JOHN S. KING1, GE SUN2, STEVEN G. MCNULTY2,MICHAEL J. GAVAZZI2, JOHNNY L. BOGGS2 & EMRYS A. TREASURE2
1Department of Forestry and Environmental Resources, North Carolina State University, Raleigh, NC 27695, USA and2Southern Global Change Program, USDA Forest Service, Raleigh, NC 27606, USA
ABSTRACT
The study examined the relationships between whole treehydraulic conductance (Ktree) and the conductance in roots(Kroot) and leaves (Kleaf) in loblolly pine trees. In addition,the role of seasonal variations in Kroot and Kleaf in mediatingstomatal control of transpiration and its response to vapourpressure deficit (D) as soil-dried was studied. Compared totrunk and branches, roots and leaves had the highest loss ofconductivity and contributed to more than 75% of the totaltree hydraulic resistance. Drought altered the partitioningof the resistance between roots and leaves. As soil moisturedropped below 50%, relative extractable water (REW),Kroot declined faster than Kleaf. Although Ktree depended onsoil moisture, its dynamics was tempered by the elongationof current-year needles that significantly increased Kleaf
when REW was below 50%. After accounting for the effectof D on gs, the seasonal decline in Ktree caused a 35%decrease in gs and in its sensitivity to D, responses that weremainly driven by Kleaf under high REW and by Kroot underlow REW. We conclude that not only water stress but alsoleaf phenology affects the coordination between Ktree and gs
and the acclimation of trees to changing environmentalconditions.
Key-words: Pinus taeda; coastal plain; conductivity; embo-lism; LAI; leaf phenology; soil moisture; water potential.
INTRODUCTION
Understanding what controls the diffusive conductance ofstomata to water vapour (gs) has been a critical area ofresearch in plant physiology because of the close relation-ship between gs and net carbon assimilation (Franks & Far-quhar 1999). The most important environmental variableto which stomata respond is the vapour pressure deficitbetween leaf and air (D). Some studies suggest thatstomatal closure occurs with increasing D as a feedback
response to leaf and whole plant water status in order tolimit transpiration and water use, rather than as a directresponse to humidity (Meinzer et al. 1995; Monteith 1995;Mott & Buckley 1998). In the gas phase and under steady-state conditions, when water storage is zero such as duringmidday (Phillips et al. 2002), transpiration on a leaf areabasis (E in mmol m-2 s-1) can be expressed mathematicallyusing the Ohm’s law analogy:
E g D= s (1)
and in the liquid phase as:
E K h= − −( )tree soil leafΨ Ψ 0 01. (2)
where Ktree (mmol m-2 s-1 MPa-1) is the leaf-specific hydrau-lic conductance of soil tree atmosphere continuum,(Ysoil - Yleaf - 0.01 h) represents the driving force for watermovement between the soil and the leaf over a tree heighth corrected for the hydrostatic gradient of 0.01 MPa m-1
(Whitehead 1998). Equations 1 and 2 indicate that Ktree andD interact to control gs, and therefore, an understandingof Ktree is essential to interpret the response of gs to D, andto quantify patterns and controls of the soil-to-rootand canopy-to-atmosphere interactions (Oren et al. 1999;Sperry et al. 2002; Ewers et al. 2005). However, the interac-tion between Ktree and D makes it difficult to partition theirindividual effects on gs. Oren et al. (1999) showed that sto-matal sensitivity to D is proportional to the magnitude of gs
at low D (<1 kPa), and therefore, when D is fixed, here at1 kPa, gs = gs-ref and becomes proportional to E,
g Es-ref ≈ (3)
from equations 2 and 3, it is clear that any change in Ktree
will induce changes in E and gs-ref, and any reductions in Ktree
should result in a proportional reduction in stomatal sensi-tivity to D.We propose to use this framework to investigatethe nature of the relationship between Ktree and gs-ref, andhow this relationship affects the sensitivity of gs to D. Weacknowledge that the Ohm’s law analogy is a simplification
Correspondence: J.-C. Domec. Fax: +1 919 513 2978; e-mail:[email protected]
Plant, Cell and Environment (2009) 32, 980–991 doi: 10.1111/j.1365-3040.2009.01981.x
© 2009 Blackwell Publishing Ltd980

of processes governing the long-distance water transport inplants (Katul, Leuning & Oren 2003; Chuang et al. 2006).However, it provides a simple and useful way to interpretthe correlation between the liquid and the gas phase, as wellas to study the inter-relationships among gs and Ktree understeady-state conditions (see review by Meinzer 2002).
Experimental studies show a fast stomatal response toartificial manipulations in Ktree (Sperry & Pockman 1993;Cochard et al. 2002). However, while many studies haveexamined the relationship between Ktree and gs, Ktree remainsa somewhat mysterious factor because it is mainly depen-dent on Kroot and Kleaf. These two components can accountfor more than 70% of the whole tree hydraulic resistance(Cruiziat, Cochard & Ameglio 2002; Sack et al. 2003; Sack &Holbrook 2006), and yet remains largely unstudied in manyspecies and ecosystems (Brodribb & Holbrook 2003;Schenk& Jackson 2005; Collins & Bras 2007). Therefore, under-standing the partitioning of these two conductivities andtheir respective contributions to Ktree under various environ-mental conditions is crucial to understand how it influencesgs and its sensitivity to D. A mechanistic depiction of theeffect of variation in Ktree, gs-ref, and the sensitivity of gs to Drequires isolating the location of the main resistances towater flow and their seasonal dynamics,which is rarely done.Without knowing how these resistances change with soildrying and plant water status, we cannot understand orpredict the response of plant water use to current and futureenvironmental conditions (Maseda & Fernández 2006;McDowell, White & Pockman 2008).
Maintaining a sufficient water supply to leaves ischallenging because the mechanism driving water flux(cohesion–tension) during transpiration places xylem undertension, making it vulnerable to cavitation-induced embo-lism. Evidence that stomata are able to sense and respondrapidly to changes in hydraulic conductance associated withvariation in embolism (Salleo et al. 2000; Brodribb et al.2003) implies that embolism in certain plant organs mayplay a rather different role than originally believed(Cochard et al. 2002; Domec et al. 2004). Reports of dailycycles of embolism formation, especially in leaves and roots,suggest that embolism is not necessarily avoided and thatxylem hydraulic properties are dynamic, reflecting thebalance between the two distinct processes of embolismand its reversal (Zwieniecki et al. 2000; Bucci et al. 2003;Domec et al. 2006).
The conversion of wetlands to intensively managed forestlands in eastern North Carolina was historically wide-spread, and yet the consequences on plant hydraulic prop-erties and in turn on water balances have not been wellstudied. This study compares the hydraulic functioning ofroot, trunk and leaves, and quantifies their contributionto the integrated functioning of plants through the linkbetween Ktree and the response of gs to D in loblolly pine(Pinus taeda L.) trees. In this species, water availability is amajor factor in determining leaf photosynthesis and leaftranspiration (Ewers, Oren & Sperry 2000). In 2007, a longsummer drought was used to observe how the hydraulicsystem functioned under prolonged low soil moisture. Our
global hypothesis was that reversible changes in Ktree causedby short-term changes in Kroot and Kleaf would act as valvesresponding to soil water availability and constitute hydrau-lic regulator that govern leaf gas exchanges. Using in situfield measurements, we tested the following specific hypoth-eses: (1) The partitioning of Ktree is mostly affected by Kroot
and Kleaf; (2) The reduction in gs-ref and in gs sensitivity to Dare direct responses to decreased Ktree and is influenced bysoil moisture; (3) Without water limitation, Kleaf limits Ktree,but as soil dries, Kroot declines faster than Kleaf and regulatesKtree, and therefore, gs-ref and gs sensitivity to D.
MATERIAL AND METHODS
Site description
The study site was a 16-year-old loblolly pine plantation witha canopy height of 14.1 m, and located at 35°48′N, 76°40′W,on the lower coastal plain of North Carolina, USA(Noormets et al. 2009). The area is <5 m above sea level ondeep Belhaven series histosol soil (http://www.epa.gov/wed/pages/ecoregions/ncsc_eco.htm) that belongs to the outercoastal plain mixed forest province. The mean (1971–2000)annual precipitation is 1320 mm,mean temperature in July is26.6 °C, and in January 6.4 °C. Leaf area index (LAI, pro-jected leaf area above a unit of ground) was measured with aLAI-2000 Plant Canopy Analyzer (Licor Inc., Lincoln, NE,USA). All LAI-2000 measurements were corrected forclumping of needles and for woody surface area (Thérézienet al. 2007; Iiames et al. 2008). To confirm that most of theseasonal variations in LAI were related to loblolly pine trees,we also measured the seasonal change in leaf needle lossfrom 30 litter traps [0.18 m2 laundry baskets suspended onpolyvinyl chloride (PVC) tubes] from which pine and hard-wood leaves were sorted by hand. The litter was collectedevery 60 d during spring and summer, and every 2 weeksduring fall and early winter.The samples were separated intoneedles, leaves and twigs, and then oven-dried at 65 °C for48 h.The total leaf mass collected during the year,divided bythe area of the litter baskets, provided an estimate of annualleaf litter mass production per square meter. This needlemass was converted to LAI using an averaged value of leafmass per area determined from needles harvested from thebottom, middle and top part of the canopy.
Microclimate, soil moisture content and sapflow measurements
The following micrometeorological parameters were mea-sured above canopy: relative humidity and air temperature(HMP45AC, Vaisala, Finland), photosynthetically activeradiation (LI-190, Licor) and precipitation (TE-525, Camp-bell Scientific, Logan, UT, USA). Soil volumetric watercontent (q) was measured using a multisensor frequencydomain capacitance technique (Domec et al. 2004; Warrenet al. 2005). Probes consist of multiple annular capacitancesensors (Sentek PTY LTD, Adelaide, Australia) separatedby 10 cm or more and placed in weatherproof PVC access
Influence of leaf and root hydraulic conductances on stomatal conductance 981
© 2009 Blackwell Publishing Ltd, Plant, Cell and Environment, 32, 980–991

tubes. These probes are relatively insensitive to fluctuationsin soil temperature (Warren et al. 2005). The plot had twoprobes installed to a depth of 1.2 m with nine independentsensors per probe. However, to compare across sites and toerase the influence of soil texture on q, drought intensity isbest quantified in the form of relative extractable soil water[relative extractable water (REW) dimensionless], and qwas converted to REW as defined by Granier, Loustau &Bréda (2000).
Stem sap flux measurements were made on six trees at1.4 m above the ground and at 4–5 radial positions using 1 cmheat dissipation probes (James et al. 2002), modified afterGranier (1987).Preliminary results showed that there was nosignificant difference in azimuthal Js within trees (P = 0.39,students paired t-test, n = 5), probably because of the homo-geneity of pine trunks, so we inserted all probes on thenorth–north-west side of the trees.Thirty-minute averages oftemperature difference data were computed and stored indata loggers (CR10, Campbell Scientific Inc.). The sensorsignal was converted to sap flux density (Js, g m-2 s-1) accord-ing to Granier (1987), which assumes that natural tempera-ture gradients between sensors are small and accounted forthe effects of non-zero night-time fluxes on the signal base-line (Oishi et al. 2008). Using tree sapwood area, stand LAI(converted to all-sided LAI using a conversion factor of 2.36;Rundel & Yoder 1998) and stand tree density (645 trees perhectare), Js was scaled and converted to a tree-scale averagetranspiration per unit leaf area (E, in mmol m-2 s-1).The totalsapwood area of the trees equipped with sapflow probeswas estimated from the relationship between sapwoodarea (from increment cores) and diameter at breast height(DBH) determined on 22 other trees and the DBH of themeasured trees. Sapwood depth was determined by visualinspection of the core and converted to sapwood area basedon the area of a circle, subtracting the areas represented bythe heartwood and bark. In all cases, sapwood depth (ca.75–90 mm) was greater than the depth at which the probeswere installed,and when scaling Js to the entire sapwood, theflux at 6–7 cm was used to represent the flux beyond 7 cm.Todetermine the seasonal increase in sapwood area as a func-tion of the existing sapwood area, 5 trees were also coredonce a month from May to December.All values of E used tocalculate hydraulic conductances accounted for the seasonaldynamics of sapwood and leaf area.
Canopy conductance and analysis of itsresponse to vapour pressure deficit
Canopy conductance derived from sapflow measurementscomprises the total water vapour transfer conductance fromthe ‘average’ stomata of the tree canopy to the measure-ment height of vapour pressure deficit (D), which includesboth surface boundary-layer and stomatal components.Canopy stomatal conductance (Gs) was calculated from Eand D, using the simplification of the inversion of Penman–Monteith model:
G RT E Ds a= ( )ρ (4)
where R is the universal gas constant adjusted for watervapour (0.46 m3 kPa K-1 kg-1), Ta is air temperature indegrees K and r is density of water (998 kg m3). This sim-plified calculation was sufficient because in all treatments Dwas close to the leaf-to-air vapour pressure deficit due tohigh boundary-layer conductance. Indeed, given that >90%of the daytime mean wind velocity was >0.7 m s-1, and thatleaf thickness never exceeded 1.8 mm, we estimated thatthe mean daytime boundary-layer conductance averaged 60times Gs (Jones 1992). Basal sap flow can be used as ameasure for diurnal crown transpiration if the measure-ments are corrected for the time lag with transpirationin order to more closely approximate the relationshipbetween D and transpiration (Phillips et al. 1997; Ewers &Oren 2000). A commonly used method to account for thetime-lag is to artificially shift the sap flow time series tomaximize correlation with D or radiation (Oren et al. 1998;Granier et al. 2000; Chuang et al. 2006). In our system, weused a half-hour lag time that was derived from theobserved time-lag of Js with respect to D.
For isohydric species and under saturated light, Orenet al. (1999) showed that the decrease in gs with increasingD is proportional to gs at low D. Therefore, the sensitivityof the stomatal response to D when PAR was >800 mmolm-2 s-1 (light-saturated Gs) was determined by fitting thedata to the functional form:
G b m Ds = − × ( )ln (5)
where b is gs at D = 1 kPa (hereafter designated as refer-ence canopy stomatal conductance, Gs-ref), and m is the rateof stomatal closure and reflects the sensitivity of Gs to D[-dGs/dlnD, in mmol m-2 s-1 ln(kPa)-1]. The slope, -dGs/dlnD is proportional to Gs-ref with the proportionality aver-aging 0.60 across a wide range of species, and varyingpredictably depending on the range of D used in the analy-sis (Oren et al. 1999; Ward et al. 2008).
Although, it has been shown that the stomatal sensitivityto D does not vary whether gs or Gs is used (Addington et al.2004; Kim, Oren & Hinckley 2008), leaf-based gs were usedto provide an independent assessment of stomatal controlof transpiration to compare with values of Gs derived fromsapflow data. Measurements of gs were conducted every60–90 min with a LI-6400 portable photosynthesis system(Li-Cor Inc., Lincoln, NE, USA) on detached fasciclessampled from the upper crown section (height of 13 m).Our tests within this study was conformed with the reportby Maier, Palmroth & Ward (2008) that the gas exchange ofdetached fascicles remained unchanged compared to that ofattached needles for at least 20 min after excision. Diurnalmeasurements commenced prior to dawn and continueduntil after 1700 h once a month from 22 May to 3 October2007.
Leaf water potential, Kleaf, Ktree and Kroot
Field Kleaf was calculated as Kleaf = E/(Ybranch - Yleaf), whereYleaf is the leaf water potential of transpiring leaves and
982 J.-C. Domec et al.
© 2009 Blackwell Publishing Ltd, Plant, Cell and Environment, 32, 980–991

Ybranch is the stem water potential estimated from non-transpiring covered shoots (Meinzer 2002). Under theseconditions, leaf water potential is generally agreed to equili-brate to that of the adjacent xylem (Richter 1997).We mea-sured Yleaf and Ybranch on the same three trees used for gs.From each of these trees, three terminal branches (<2 cm indiameter) located in the upper crown section were sampled(height of 13 m). Late morning (1000–1130 h) values ofYleaf, Ybranch and E were used because at this time light issaturating and leaves are likely to have transpired all storedwater so that steady-state water flow conditions are likely toexist. To analyse the effect of Kleaf on whole tree hydraulicconductance (Ktree), Ktree was calculated from the slope ofthe relationship between E and Yleaf (Loustau, Domec &Bosc 1998).The regression method was used rather than thesingle point method [Ktree = E/(Ypre-dawn - Yleaf)] becausethe slope of the regression line represents the whole treehydraulic resistance (1/Ktree) and the potential at zero flowrepresents the soil water potential in contact with the roots(Cohen, Moreshet & Fuchs 1987). This approach was pos-sible because the hysteresis effects resulting from plantcapacitance were small, as previously documented foryoung loblolly pine trees (Ewers & Oren 2000). The cumu-lative daily water storage determined from the hysteresisbetween E and Yleaf (Loustau et al. 1998) never exceededmore than 9, 7 and 5% of the daily transpiration in May,August and September, respectively. Using the ohm (elec-trical resistance) analogy applied to a hydraulic circuit, rootto branch hydraulic conductance (Kroot-branch) was calculatedas: Kroot-branch = (Ktree ¥ Kleaf)/(Kleaf - Ktree). Measurements ofYleaf and Ybranch were conducted every 2 weeks from May toSeptember, and then once a month through mid Decemberwith a pressure chamber (Model 1000; PMS, Albany, OR,USA). Additionally, we were able to separate the effect oftrunk and branch hydraulic conductance [Ktrunk-branch = E/(Ytrunk - Ybranch)] from Kroot-branch by measuring water poten-tial (Ytrunk) at the trunk base. Measurements of Ytrunk weredone once every 4 weeks by bagging one entire branch pertree on five adjacent trees bearing low branches (1.7 metersin height). Root hydraulic conductance (Kroot) wascomputed using the electrical analogy as Kroot = (Kroot-branch ¥Ktrunk-branch)/(Ktrunk-branch - Kroot-branch) (Nardini et al. 2003).
Loblolly pine trees carry 2 years of foliage during thegrowing season. To better understand the effect of eachleaf cohort on Kleaf, we also determined the Kleaf ofprevious- and current-year needles (Kleaf-current and Kleaf-
previous, respectively). Kleaf-current and Kleaf-previous were calcu-lated using the porometry-based transpiration and Yleaf
measured on either current- or previous-year needles.Further, to separate the contribution of current-yearversus previous-year needle to the mean Kleaf determinedusing sapflow-based transpiration, we calculated an inte-grated Kleaf (Kleaf-mean) from Kleaf-current and Kleaf-previous
weighted by the proportion of the total leaf area repre-sented by current and previous needles. The leaf area ofeach cohort of needles was determined by comparing thefractional change in litter-fall (representing previous-yearneedle fall) to the total LAI (representing both current-
and previous-year needles). The progression of LAI ofcurrent-year needle through the year was calculated bysubtracting LAI of falling needles from the total LAI. Tofurther confirm that most of the seasonal variations inLAI were related to loblolly pine trees, the seasonalcurrent-year needle production was documented by mea-suring needle elongation every 2 weeks on 18 needles pertree from five trees.
Monitoring seasonal occurrence of embolism inroot, trunk and branches
To compare with the seasonal variations in Kroot, Ktrunk-branch
and Kleaf, specific hydraulic conductivity (ks) and embolismwere measured in roots and branches collected at the endof July, at the beginning of September and in the middleof October, 2007. Sampling started when REW was above50% and ended when it was below 20%. At midday ofeach sampling date, lateral medium roots 2–4 mm in diam-eter and greater than 25 cm in length were excised nearthe base of five different trees at about 25 cm depth in thesoil. Two terminal branches per tree were collected fromthe upper canopy level of three trees. Roots and brancheswere wrapped in aluminium foil and placed in a sealedplastic bag with a moist towel and immediately trans-ported to the laboratory in a cooler, to be processedwithin 3 h of sampling (Domec et al. 2004). In the labora-tory, a 10–15 cm long section of each root and branch wascut, and the proximal end was attached to a tubing systemand perfused at a pressure of 5 kPa with filtered (0.22 mm)water. The hydraulic pressure head was adjusted to avoidany refilling of embolized tracheids. The rate of efflux wasmeasured in a 1 ml graduated pipette. Specific conductiv-ity was calculated as the mass flow rate of the perfusionsolution divided by the pressure gradient across thesegment, normalized by the xylem cross-sectional area.Native embolism or percent loss of specific conductivity ofroot and branch segments (PLks-root and PLks-branch) wasdetermined by comparing the initial (or native) conduc-tivity to the maximum conductivity after removal of airemboli by soaking the samples in perfusion solution undervacuum for 48 h.
On the same days and on the same trees from which rootswere collected, trunk xylem percent loss of conductivity(PLks-trunk) was estimated from the change in relative watercontent (RWC) of trunk increment cores using publishedrelationships relating PLks-trunk to RWC. Indeed, acrossseveral conifer tree species it has been shown that PLks-trunk
is nearly equal as twice the relative trunk water deficit(100-RWC) (Domec & Gartner 2002; Domec et al. 2006;Rosner et al. 2006; Domec & Pruyn 2008). Cores werecollected at midday and wrapped in plastic film.Back at the laboratory, we determined Mf (fresh mass),Vf (fresh volume) by water displacement and Md (dry mass)for the cores. Assuming a cell-wall material densityof 1.53 g cm-3 (Siau 1984), RWC was calculated asRWC = (Mf - Md)/(Vf - Md/1.53).
Influence of leaf and root hydraulic conductances on stomatal conductance 983
© 2009 Blackwell Publishing Ltd, Plant, Cell and Environment, 32, 980–991

Error estimates and statistical analysis
Variance estimates of E and Gs were computed according toOren et al. (1998) to include the variances of Js and LAI andtheir mutual dependency. Similarly, variance estimates ofKroot, Kleaf and Ktree were also computed using the varianceestimate of E, and mean variance of Y (Phillips et al. 2002).Analysis of variance (ANOVA) was performed to test fordifferences between Kroot, Kleaf and Ktree. To assess the dif-ference in native embolism or percent loss of hydraulicconductivity at each date, we used a two-way ANOVA withone repeated measure factor. Variations in q, REW andhydraulic conductivities over the measurements periodwere analysed by repeated measures ANOVA. The rela-tionships among E, Gs and D were investigated, andbetween-REW differences in regression curves were testedby using F-test statistics for extra sum of squares (Motulsky& Christopoulos 2004). Relative errors in Gs, caused byinstrument limitations were kept to <10%, by limiting Gs
calculations to D > 0.6 kPa (Ewers & Oren 2000). Statisticalanalyses were performed using SAS (Version 9.1, Cary, NC,USA) and curve fits were performed using Sigmaplot(version 9.0, SPSS Inc., San Rafael, CA, USA).
RESULTS
The 2007 growing season experienced an extreme drought,with soil water content (q) reaching its minimum in lateAugust, and lasting until major rain events occurred in lateOctober (Fig. 1a). Total precipitation for the measurementperiod was 510 mm, which was 53% of normal based on a30-year mean for the area. In the upper 60 cm, q varied
significantly over the measurement period (P < 0.001;Fig. 1a). At the beginning of the study in May the soil wasclose to field capacity and mean q of the 0–60 cm layerranged from 0.36 to 0.28 m3 m-3 (Fig. 1a). In June andbetween August and November, gradual soil dryingoccurred, with q falling to ca. 0.08 m3 m-3. About half of theseasonal decline in q had occurred by the end of August. Inlate June, q had already fallen to ca. 0.11 m3 m-3, when lightrain events caused a transient partial recovery of q. Duringthe drought, q in the upper 60 cm of soil was reduced to27% of maximum, which corresponded to a decrease inREW of 80%. The seasonal decline in pre-dawn Y cor-rected for the gravitational component was very responsiveto changes in soil moisture and as the soil dried out, pre-dawn Y declined in parallel (Fig. 1b). Pre-dawn Y was closeto -0.4 MPa at the beginning of the study period andshowed little change after late August when q droppedbelow 50% REW (0.2 m3 m-3, Fig. 1b). The water potentialat zero flow derived from the regression between Y and Erepresented the pre-dawn Y in contact with the roots. Thisvalue was not significantly different than measured pre-dawn Y (P = 0.74). The minimum midday Yleaf averaged-2.2 MPa from May to October, and varied significantlythroughout the growing season (P < 0.01), but never fellbelow -2.4 MPa (Data not shown).
We observed natural variation in Ktree during the mea-surement period (Fig. 2a). Mean Ktree was decreased from0.82 mmol m-2 s-1 MPa-1 in May to 0.60 mmol m-2 s-1 MPa-1
in October. Variation in Ktree, Kroot and Kleaf over the mea-surement period was, in all cases, highly significant(P < 0.001). From May to mid July, Kleaf decreased by 23%(P = 0.02), whereas Kroot increased by 12% (P = 0.04)(Fig. 2b) during the same period. As soon as new leavesbecame fully expanded in early August, Kleaf increased backto spring values. As q declined from September to Novem-ber, Kleaf was reduced by 18% and Kroot by 40%, respec-tively. This decline in Kleaf was mainly due to the sharpdecrease in Kleaf from the previous year foliage (Kleaf-previous),which lost 50% of conductivity between July and Septem-ber. In contrast, Kleaf from current-year foliage (Kleaf-current)almost increased by about 47% (Table 1) during the sameperiod. The sap-flow based Kleaf compared well with theporometry-based Kleaf (Kleaf-mean in Table 1), which was cal-culated using Kleaf-previous and Kleaf-current and the leaf arearepresented by previous- and current-year needles.
The root resistance (i.e. inverse of conductance) declinedfrom 61% to 46% of the total root-to-leaf resistancebetween May and August, and then increased back to 58%for the rest of the year (Fig. 2c). Averaged over the growingseason, 1/Kleaf and 1/Ktrunk-branch represented 33% and 11% of1/Ktree, respectively. For the whole measuring period, 1/Kleaf
never exceeded more than 39% of 1/Ktree. Note that mea-surements of midday Ytrunk carried out simultaneously onbasal and on upper branches (13 m above ground level)showed an average midday Ytrunk difference rangingbetween 0.02 and 0.05 MPa after correcting for the gravita-tional component. Thus, the Ytrunk difference between thebase and the top of the trunk was negligible compare to
So
il w
ate
r co
nte
nt,
q (
m3
m–
3)
0.0
0.1
0.2
0.3
0.4
Pre
cip
itation
(m
m d
–1 )
0
20
40
60
80
1000–30 cm
30–60 cm
rain
2007
May Jun Jul Aug Sep Oct Nov Dec Jan
RE
W (
%)
0
20
40
60
80
100
Pre
-da
wn
Y(M
Pa)
–0.7
–0.6
–0.5
–0.4REW (0–60 cm)Pre-dawn water potential
(a)
(b)
Figure 1. Seasonal variations from May to December 2007 in(a) soil volumetric water content (q) and precipitation, and in(b) relative extractable water averaged over 60 cm of soil (REW)and pre-dawn leaf water potential (pre-dawn Y) corrected forthe gravitational component. All measurements are means � SE.
984 J.-C. Domec et al.
© 2009 Blackwell Publishing Ltd, Plant, Cell and Environment, 32, 980–991

Ybranch, and so most of the resistance in the trunk-branchcompartment was due to branch wood. The percent lossof conductance of roots (PLKroot) increased significantly(P = 0.02) from July to October, whereas the percent loss ofconductance of the trunk-branch pathway (PLKtrunk-branch)did not show any significant changes (P = 0.54). Theseresults were confirmed by the measurements of nativeembolism of individual root, branch and trunk samples(PLks-root, PLks-trunk, PLks-branch). The average loss of specificconductivity of each different organ PLks-root-trunk-branch ateach sampling date was close to PLKroot-trunk-branch (Table 1).The partitioning of native embolism indicated that thedecrease in Kroot-trunk-branch was due to a reduction in the root
conductivity rather than branch or trunk conductivity andso the seasonal change in the contribution of root to Ktree
was linked to increase root embolism (Table 1).Stand LAI determined optically varied by about 35%
over a year with a maximum of 5.1 � 0.2 m2 m-2 in August,to a minimum of 3.5 � 0.3 m2 m-2 December (Fig. 2d) inNovember. This minimum value compared well with theLAI of 3.6 � 0.3 m2 m-2 determined from litter fall basketsover a 3-year period, and which corresponds to the meanLAI of a single needle cohort carried by the trees in winter.LAI increased because of new foliage production duringthe spring, reaching a peak in July and August. The devel-oping foliage of the first growth flush of the spring hadalmost attained maximum length by September (Fig. 2d)and the previous year foliage was retained at 50% untilSeptember, and was almost completely lost by November.As a consequence, previous and current needles had thesame LAI in late July, but by October, total LAI was domi-nated at 80% by current needles (Table 1).
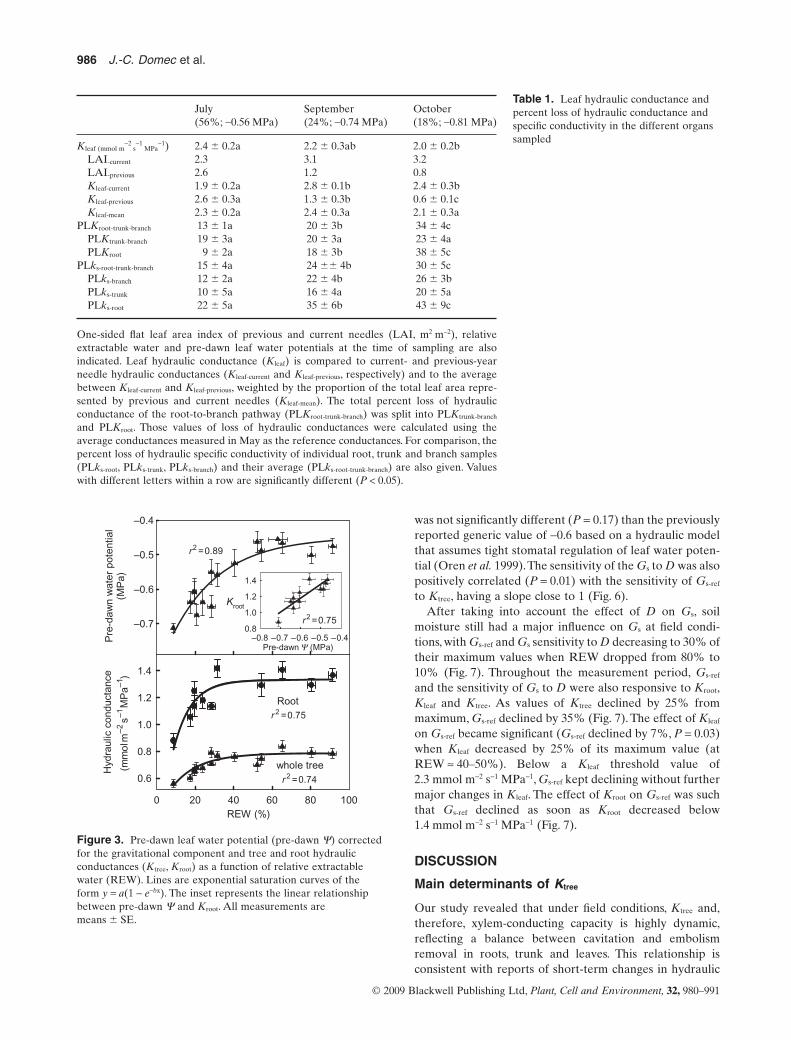
The variation in pre-dawn Y, Kroot and Ktree were relatedto soil moisture (Fig. 3). Ktree decreased by 25% of itsmaximum value as soil water potentials approached-0.6 MPa and soil water content dropped below 40–50%REW (Fig. 3). Kroot was similarly responsive to soil dryingonce the 40–50% REW threshold was reached with Kroot
decreasing by 36–38% of its maximum values between Mayand November.
Stomatal and canopy conductances were linearly related(P < 0.001; Fig. 4a), although mean gs was 13% higher thanGs.This good agreement allowed us to use Gs as an accuratepredictor of leaf level gas exchange. Diurnal variations inthe two independent parameters, gs and Kleaf also suggesta strong correlation between the gas and liquid phaseconductances (Fig. 4b). The daily response of gs to Kleaf wasinfluenced by soil moisture such that maximum gs decreasedby 25% when REW dropped below 50% (P = 0.02).However, the response of gs to midday Yleaf was not affectedby REW (P = 0.11) (Fig. 4c).
Daytime-averaged D showed significant seasonal varia-tion (P < 0.01) from 1.4 kPa in May to 1.8 kPa and 0.91 kPain August and November, respectively. Stomatal conduc-tance and tree transpiration showed marked declines inresponse to increases in D (Fig. 5). The response of Gs andtree transpiration to increasing D was influenced by soilmoisture (Fig. 5).At D = 1 kPa, the decrease in reference Gs
(Gs-ref) and E for days with REW below 50% were, respec-tively, 27 and 23% lower (P < 0.03) than for days whenREW was above 50% (Fig. 5).
The shape of the relationship between Gs and D couldadequately be described using Equation 5. Relationshipsbetween monthly averaged Gs and lnD were generallystrong (minimum r2 = 0.75). The sensitivity of the Gs to D(-dGs/d ln D) was linearly related to stomatal conductanceat D = 1 kPa (Gs-ref; P < 0.001; Fig. 6). Stomatal conductancedeclined in response to increasing D, and the magnitudeof the reduction varied over the measurement period asshown by the decline in Gs-ref. The slope of the relationshipbetween Gs and the sensitivity of the Gs to D (-0.57 � 0.5)
Re
sis
tan
ce
to
wh
ole
tr
ee
wa
ter
flo
w (
%)
0
20
40
60
80
100
May Jun Jul Aug Sep Oct Nov Dec 0
20
40
60
80
100
LA
I (m
2 m
–2)
2.5
3.0
3.5
4.0
4.5
5.0
Elong.
LAI
Loss
Roots
Leaves
Hydra
uulic
conducta
nce
(mm
ol m
–2
s–
1 M
Pa
–1)
1.2
1.6
2.0
2.4
2.8Roots
Leaves
Hydra
uulic
conducta
nce
(m
mo
l m–
2 s
–1
MP
a–
1)
0.6
0.7
0.8
0.9Whole tree
Ne
ed
le e
lon
ga
tio
n
Needle
lo
ss (
%)
Trunk-Branches
(a)
(b)
(c)
(d)
Figure 2. Seasonal variations in (a) soil-to-leaf hydraulicconductance (Ktree), in (b) root (Kroot) and leaf (Kleaf) hydraulicconductances, in (c) the proportion of leaf, trunk to branches(Trunk-Branches) and root hydraulic resistances to the overalltree hydraulic resistance (1/Ktree), and in (d) the patterns inneedle elongation (Elong.), needle loss and one-sided flat leafarea index. All measurements are means � SE.
Influence of leaf and root hydraulic conductances on stomatal conductance 985
© 2009 Blackwell Publishing Ltd, Plant, Cell and Environment, 32, 980–991
was not significantly different (P = 0.17) than the previouslyreported generic value of -0.6 based on a hydraulic modelthat assumes tight stomatal regulation of leaf water poten-tial (Oren et al. 1999).The sensitivity of the Gs to D was alsopositively correlated (P = 0.01) with the sensitivity of Gs-ref
to Ktree, having a slope close to 1 (Fig. 6).After taking into account the effect of D on Gs, soil
moisture still had a major influence on Gs at field condi-tions, with Gs-ref and Gs sensitivity to D decreasing to 30% oftheir maximum values when REW dropped from 80% to10% (Fig. 7). Throughout the measurement period, Gs-ref
and the sensitivity of Gs to D were also responsive to Kroot,Kleaf and Ktree. As values of Ktree declined by 25% frommaximum, Gs-ref declined by 35% (Fig. 7). The effect of Kleaf
on Gs-ref became significant (Gs-ref declined by 7%, P = 0.03)when Kleaf decreased by 25% of its maximum value (atREW ª 40–50%). Below a Kleaf threshold value of2.3 mmol m-2 s-1 MPa-1, Gs-ref kept declining without furthermajor changes in Kleaf. The effect of Kroot on Gs-ref was suchthat Gs-ref declined as soon as Kroot decreased below1.4 mmol m-2 s-1 MPa-1 (Fig. 7).
DISCUSSION
Main determinants of Ktree
Our study revealed that under field conditions, Ktree and,therefore, xylem-conducting capacity is highly dynamic,reflecting a balance between cavitation and embolismremoval in roots, trunk and leaves. This relationship isconsistent with reports of short-term changes in hydraulic
Table 1. Leaf hydraulic conductance andpercent loss of hydraulic conductance andspecific conductivity in the different organssampled
July(56%; -0.56 MPa)
September(24%; -0.74 MPa)
October(18%; -0.81 MPa)
Kleaf (mmol m-2
s-1
MPa-1) 2.4 � 0.2a 2.2 � 0.3ab 2.0 � 0.2b
LAI-current 2.3 3.1 3.2LAI-previous 2.6 1.2 0.8Kleaf-current 1.9 � 0.2a 2.8 � 0.1b 2.4 � 0.3bKleaf-previous 2.6 � 0.3a 1.3 � 0.3b 0.6 � 0.1cKleaf-mean 2.3 � 0.2a 2.4 � 0.3a 2.1 � 0.3a
PLKroot-trunk-branch 13 � 1a 20 � 3b 34 � 4cPLKtrunk-branch 19 � 3a 20 � 3a 23 � 4aPLKroot 9 � 2a 18 � 3b 38 � 5c
PLks-root-trunk-branch 15 � 4a 24 �� 4b 30 � 5cPLks-branch 12 � 2a 22 � 4b 26 � 3bPLks-trunk 10 � 5a 16 � 4a 20 � 5aPLks-root 22 � 5a 35 � 6b 43 � 9c
One-sided flat leaf area index of previous and current needles (LAI, m2 m-2), relativeextractable water and pre-dawn leaf water potentials at the time of sampling are alsoindicated. Leaf hydraulic conductance (Kleaf) is compared to current- and previous-yearneedle hydraulic conductances (Kleaf-current and Kleaf-previous, respectively) and to the averagebetween Kleaf-current and Kleaf-previous, weighted by the proportion of the total leaf area repre-sented by previous and current needles (Kleaf-mean). The total percent loss of hydraulicconductance of the root-to-branch pathway (PLKroot-trunk-branch) was split into PLKtrunk-branch
and PLKroot. Those values of loss of hydraulic conductances were calculated using theaverage conductances measured in May as the reference conductances. For comparison, thepercent loss of hydraulic specific conductivity of individual root, trunk and branch samples(PLks-root, PLks-trunk, PLks-branch) and their average (PLks-root-trunk-branch) are also given. Valueswith different letters within a row are significantly different (P < 0.05).
REW (%)
0 20 40 60 80 100
Hydra
ulic
conducta
nce
(m
mol m
–2
s–
1–
1 M
Pa
)
0.6
0.8
1.0
1.2
1.4
Pre
-da
wn
wate
r p
ote
ntia
l (
MP
a)
–0.7
–0.6
–0.5
–0.4
r2 = 0.89
r 2 = 0.74
whole tree
Root
r 2 = 0.75
Pre-dawn Y (MPa)–0.8 –0.7 –0.6 –0.5 –0.4
Kroot
0.8
1.0
1.2
1.4
r2 = 0.75
Figure 3. Pre-dawn leaf water potential (pre-dawn Y) correctedfor the gravitational component and tree and root hydraulicconductances (Ktree, Kroot) as a function of relative extractablewater (REW). Lines are exponential saturation curves of theform y = a(1 - e-bx). The inset represents the linear relationshipbetween pre-dawn Y and Kroot. All measurements aremeans � SE.
986 J.-C. Domec et al.
© 2009 Blackwell Publishing Ltd, Plant, Cell and Environment, 32, 980–991

conductivity in different plant parts (Zwieniecki et al. 2000;Brodribb et al. 2003; Bucci et al. 2003; Domec et al. 2004).Aspreviously reported for several tree species (Magnani &Borghetti 1995; Irvine et al. 1998;Addington et al. 2004), theseasonal decrease in Ktree was associated with variation insoil water availability. However, and as hypothesized, the35% decline in Ktree throughout the season was highly influ-enced by the variation of Kleaf and Kroot. This respectiveeffect of Kleaf and Kroot on Ktree have seldom been explicitlyreported. Interestingly, at the beginning of the season, Ktree
decreased because of the sharp decline in Kleaf. Later in theseason, once REW dropped below 40–50%, the noticeabledecrease in Ktree was mostly dominated by Kroot, in accor-dance with several studies attributing a major role to roots
in regulating Ktree as soil dries (Steudle 2000; Domec et al.2004).
We postulate that seasonal changes in Kroot and Kroot-branch
were mainly attributable to changes in the embolism ofroots rather than to modifications in hydraulic properties atthe soil-root interface or in the trunk and branch parts(Table 1). In conifers, the components Kbranch and Ktrunk havebeen shown not to vary seasonally (Domec, Pruyn &Gartner 2005; Rosner et al. 2008). In the current study, theseasonal drop in soil moisture and pre-dawn Y were relatedto Kroot. It has been shown that loblolly pine roots lose25–45% of conductivity at water potentials ranging from-0.5 to -0.75 MPa (Ewers et al. 2000; Hacke et al. 2000),which corresponded to the decline in pre-dawn Y whenREW was below 50%, and to our maximum field loss ofroot specific conductivity (ks-root, Table 1). Seasonal changesin Kroot indicated that embolized tracheids were partiallyrefilled by late fall before the soil had been fully rechargedby precipitation, consistent with previous studies showingthat a relatively small amount of water can markedlyincrease root conductivity (Domec et al. 2006). It is unlikelythat the conductance recovery in the late fall reflected theproduction of new roots because it has been shown in
Canopy conductance
(Gs, mmol m−2 s−1)
0 20 40 60 80 100
Sto
ma
tal c
on
du
cta
nce
( gs, m
mo
l m−2
s−1
)
0
20
40
60
80
100
REW > 50%
REW < 50%
Leaf hydraulic conductance
(Kleaf, mmol m−2 s−1 MPa
−1)
0 1 2 3 4
Sto
ma
tal c
on
du
cta
nce
( gs, m
mo
l m−2
s−1
)
0
20
40
60
80
100
gs
= 1.13Gs
r2 = 0.86
(a)
(b)
Leaf water potential (Yleaf, MPa)
–2.5 –2.0 –1.5 –1.0 –0.5
Sto
ma
tal c
on
du
cta
nce
( gs, m
mo
l s−1
m−2
)
0
20
40
60
80
100
r2 = 0.74
(c)
r2 = 0.65
Figure 4. (a) Relationship between leaf- and canopy-levelstomatal conductances (gs and Gs, respectively), (b) between gs
and leaf hydraulic conductance (Kleaf) and (c) between gs andleaf water potential for two relative extractable water ranges. In(b) and (c), quadratic curves were used to fit the data. Allmeasurements are means � SE.
Vapour pressure deficit (kPa)
0.0 0.5 1.0 1.5 2.0 2.5 3.0 3.5
Tre
e t
ran
sp
ira
tio
n
(E,
mm
ol m
−2 s
−1)
0.0
0.2
0.4
0.6
0.8
1.0
1.2
Canopy c
onducta
nce
(Gs,
mm
ol m
−2 s
−1)
0
20
40
60
80
100 REW > 50%
REW < 50%
r2 = 0.95
r2 = 0.45
r2 = 0.91
r2 = 0.34
Figure 5. Responses of light-saturated canopy stomatalconductance (Gs) and transpiration (E) to water vapour pressuredeficit (D) as a function of the relative extractable water (REW).Equation 5 was used to fit the relationships between Gs and D.The vertical dashed-line at D = 1 kPa intersects the boundary lineat reference Gs (Gs-ref), which was 61 mmol m-2 s-1 for REW> 50% and 46 mmol m-2 s-1 for REW < 50%. The slope of theresponse of Gs to lnD was 27 and 22 mmol m-2 s-1 ln(kPa)-1 forREW > 50% and for REW < 50%, respectively (parameter min Equation 5). The exponential saturation curves of the formy = a(1 - e-bx) were used to fit the relationships between E and D.The maximum transpiration rate (parameter a of the fitted curve)was 1.06 mmol m-2 s-1 for REW > 50% and 0.82 mmol m-2s-1 forREW < 50%.
Influence of leaf and root hydraulic conductances on stomatal conductance 987
© 2009 Blackwell Publishing Ltd, Plant, Cell and Environment, 32, 980–991

several conifer seedlings that root regeneration is inhibitedby more than 50% when pre-dawn Y drops below -0.5 MPa(Zou, Sands & Sun 2000).
It is possible that the progressive reduction of soil watercontent could have decreased the conductance at the soil toroot interface, and consequently decreased the apparentKroot (Passioura 1988). However, because the Y at zero tran-spiration was not different than measured pre-dawn Y, soilY was probably in equilibrium with tree Y and Kroot repre-sented the soil to root hydraulic conductance (Cohen et al.1987).
Above-ground partitioning ofhydraulic resistance
Needles constituted the major part of the above groundhydraulic resistance to water flow. Leaf resistances repre-sented more than 35% of the total hydraulic resistances,and more that 75% of the above ground resistance,although leaves constituted less than 3% of the totalhydraulic pathway. Therefore, on a distance basis, leaf con-ductance is the major determinant of Ktree, which may putsome limits on maximum needle length (Zwieniecki et al.
Gs-ref (mmol m–2
s–1
)
40 45 50 55 60–dG
s/d
lnD
(mm
ol m
–2
s–
1ln
kP
a–
1)
35
30
25
20
−dGs/dlnD = 0.57 G
s-ref.−1.7
R2 = 0.93
dGs-ref/dKtree (MPa−1
)
66 68 70 72 74 76 78
−dGs/dlnD = 0.93 d G
s-ref./dK
tree−40
R2 = 0.81
Figure 6. Linear relationship between the sensitivity of canopy conductance to vapour pressure deficit (-dGs/dlnD) and Gs at D = 1 kPa(Gs-ref) and to the stomatal sensitivity to tree hydraulic conductance (dGs-ref/dKtree). The dashed line (slope = 0.6) represents the theoreticalslope between stomatal conductance at D = 1 kPa and stomatal sensitivity to D that is consistent with the role of stomata in regulatingminimum Yleaf (Oren et al. 1999).
−dG
s/d
lnD
( mm
ol m
–2 s
–1
lnkP
a–
1 )
36
32
28
24
20
REW (%)
0 20 40 60 80 100
Gs-r
ef
(mm
ol m
–2
s–
1 )
35
40
45
50
55
60
r 2 = 0.91
r 2 = 0.81
Ktree (mmol m−2
s−1
MPa−1
)
0.60 0.65 0.70 0.75 0.80
r 2 = 0.92
r 2 = 0.84
Kroot or Kleaf (mmol m−2
s−1
MPa−1
)
1.0 1.2 1.4 2.0 2.4 2.8
r 2 = 0.79
r 2 = 0.89
root leaf
r 2 = 0.76
r 2 = 0.61
root leaf
Figure 7. Relationships between the sensitivity of canopy conductance to vapour pressure deficit (-dGs/dlnD) and between Gs atD = 1 kPa (Gs-ref) to the relative extractable water (REW) and to tree hydraulic conductance (Ktree) and its two main components Kroot
and Kleaf. The function y = a(1 - e-bx) was used to fit the relationships between Gs-ref and REW, as well as between Gs-ref and Kleaf and-dGs/dlnD and Kleaf. The function y = aebx was used to fit the relationships between Gs-ref and Kroot and between -dGs/dlnD and Kroot. Allother fitted curves were linear.
988 J.-C. Domec et al.
© 2009 Blackwell Publishing Ltd, Plant, Cell and Environment, 32, 980–991

2006) and maximum tree height (Woodruff et al. 2007).Some of the variations observed in Ktree between the begin-ning and the end of the measurement period were related toa decrease in Kleaf as needles senesced, and to an increaseas new foliage became hydroactive (Neumann 1987). Thedecline in Kleaf at the peak of the growing season from Mayto July was not induced by decreasing Yleaf, but rather by theexpression of previous-year leaf senescence and a decline inKleaf-previous. Decrease in Kleaf because of leaf senescence hasbeen reported in angiosperm species but not in conifers(Salleo et al. 2000; Brodribb & Holbrook 2003). Because ofleaf expansion from May to September, the increase in Kleaf
was the consequence of current-year needle developmentthat increased hydraulic efficiency (Zwieniecki et al. 2006).The concurrent increase in current-year LAI and inKleaf-current increased the overall leaf hydraulic efficiencyin mid-summer, which limited the effect of the decrease inKroot on Ktree. Many studies assume a constant LAI or con-stant leaf area to sapwood area ratio in the calculationof Ktree. Omitting such variation would affect the interpre-tation of the relationship between Ktree and stomatalconductance.
Stomatal regulation in response to soilwater availability
Confirming our second hypothesis, the onset of the reduc-tion in Gs-ref was tied to the decrease in soil water contentbelow 50% REW. As pre-dawn water potential declined inresponse to decreasing REW, Gs-ref. declined in order toreduce transpiration at a time of declining water availabil-ity. The threshold of 40–50% for REW, beyond which Gs
was reduced, was previously reported in a large number oftree species and soil types (Bréda et al. 2006; Granier et al.2007). The seasonal decline in Gs-ref was strongly relatedto the seasonal decline in Ktree (a 25% reduction in Ktree
induced a nearly 30% reduction in Gs-ref) indicative of ahydraulic effect on stomata regulation. Ktree acted inconcert with stomata to limit water loss by maintainingpre-dawn and midday Yleaf above -0.8 MPa and above-2.2 MPa, respectively. For loblolly pine, those Yleaf repre-sent critical values that would induce 80% loss of Kroot andloss of leaf turgor (Hacke et al. 2000). Furthermore, theresponse of Gs to D was closely correlated with theresponse of Gs-ref. to Ktree (Fig. 6), which is indicative of astrong coordination between the liquid- and gas-phase(Sellin & Kupper 2005), and consistent with the view thatthe stomatal response to D represents a feedbackresponse of Gs to changes in Ktree (Sperry & Pockman1993; Meinzer 2002).
Our results also show that the sensitivity of Gs to D wasmostly attributable to the variation in Gs-ref, which is againconsistent with the isohydric regulation of water potential(Oren et al. 1999).The effect of Ktree on the sensitivity of Gs toD was rather surprising (Fig. 7). Although Gs-ref decreasedsharply with decreasing Ktree, the decrease in the sensitivityof Gs to D became asymptotic as Ktree continued to decrease.The very low sensitivity of Gs to D under severe water stress
(REW < 20%) was probably because stomata were nearlyfully closed and transpiration was mostly cuticular.
Co-ordination between Gs-ref and Kleaf, andbetween Gs-ref and Kroot as soil dried
Confirming our last hypothesis, Gs-ref was driven by changein Kleaf once soil moisture was above 50%, and by Kroot whenREW was kept below 50%. This soil moisture threshold forstomatal closure was reached when Kroot started to drop andthus when the root resistance had become an effective com-petitor for water with the above-ground portion of the tree(Domec et al. 2006). The strong effect of declining Kleaf onGs-ref may be explained by the coupled effect of the diurnaldynamics of gs, Kleaf and Yleaf (Fig. 4; Brodribb et al. 2003;Woodruff et al. 2007), to the production of new and veryhydraulically efficient needles. Many models have been pro-posed to describe water movement within trees and topredict tree transpiration with varying degrees of success(Katul et al. 2003). As suggested by Bohrer et al. (2005) andChuang et al. (2006), a stomatal model can be coupled witha model of the hydraulic system of a tree to provide inde-pendent time series of transpiration and advance our under-standing of stomatal functioning under various soilmoisture and atmospheric demand. Our study providesempirical data that can be used in such theoretical hydro-dynamic models to track midday water potentials in thedifferent parts of the tree system and see which of thesewater potentials is more effective in affecting gs. Suchmodels should be able to predict whether the magnitude ofseasonal water depletion and refilling of root and leaf xylemconstituted a signal involved in stomatal regulation toprevent Yleaf from declining to values that could provokeembolism in stems where embolism reversal may not be asefficient as in roots (Salleo et al. 2000; Domec et al. 2004).
In conclusion, we feel that a more complete view of treeadaptation to the environment can emerge from a descrip-tion of hydraulic architecture of plant organs. The linkagesbetween stomatal behaviour and Ktree shown in this studyforce us to reconsider the ecological implications of species-specific strategies with regard to embolism and stomatalopening. In a future with higher temperatures and evapo-transpiration, trees might face hydraulic limitations inregions of increased frequency and intensity of drought(Luo et al. 2001; Granier et al. 2007). The extreme droughtevent that occurred in the southern United States during2007 highlighted the need to understand the key processesthat may allow trees and stands to overcome such severewater shortages (McDowell et al. 2008). We conclude that agiven increase in D and a decrease in soil moisture inresponse to such climate change will reduce Ktree, which inturn may impose a series of constraints on the water andcarbon economy of the plant (Irvine et al. 2004; Noormetset al. 2009). Finally, more work needs to be done to deter-mine the main drivers of Ktree in the root to leaf continuum,and how seasonal variation in LAI and leaf phenology ofdifferent needle cohorts affect Gs-ref and stomatal sensitivityto D.
Influence of leaf and root hydraulic conductances on stomatal conductance 989
© 2009 Blackwell Publishing Ltd, Plant, Cell and Environment, 32, 980–991

ACKNOWLEDGMENTS
This work was supported by the USDA Forest ServiceSouthern Global Change Program cooperative agreement04-CA-11330147-238.The Weyerhaeuser Company is grate-fully acknowledged for facilitating access to the study siteand offering logistical assistance.
REFERENCES
Addington R.N., Mitchell R.J., Oren R. & Donovan L.A. (2004)Stomatal sensitivity to vapor pressure deficit and its relationshipto hydraulic conductance in Pinus palustris. Tree Physiology 24,561–569.
Bohrer G., Mourad H., Laursen T.A., Drewry D., Avissar R., PoggiD., Oren R. & Katul G.G. (2005) Finite-Element Tree CrownHydrodynamics model (FETCH) using porous media flowwithin branching elements – a new representation of tree hydro-dynamics. Water Resource Research 41, W11404. doi:10.1029/2005WR004181.
Bréda N., Huc R., Granier A. & Dreyer E. (2006) Temperate foresttrees and stands under severe drought: a review of ecophysi-ological responses, adaptation processes and long-term conse-quences. Annals of Forest Science 63, 625–644.
Brodribb T.J. & Holbrook N.M. (2003) Changes in leaf hydraulicconductance during leaf shedding in seasonally dry tropicalforest. New Phytologist 158, 295–303.
Brodribb T.J., Holbrook N.M., Edwards E.J. & Gutiérrez M.V.(2003) Relations between stomatal closure, leaf turgor andxylem vulnerability in eight tropical dry forest trees. Plant, Cell& Environment 26, 443–450.
Bucci S.J., Scholtz F.G., Goldstein G., Meinzer F.C. & Sternberg L.(2003) Dynamic changes in hydraulic conductivity in petioles oftwo savanna tree species: factors and mechanisms contributingto the refilling of embolized vessels. Plant, Cell & Environment26, 1633–1645.
Chuang Y.-L., Oren R., Bertozzi A.L., Phillips N. & Katul G.G.(2006) The porous media model for the hydraulic system of aconifer tree: linking sap flux data to transpiration rate. EcologicalModeling 191, 447–468.
Cochard H., Coll L., Le Roux X. & Améglio T. (2002) Unravelingthe effects of plant hydraulics on stomatal closure during waterstress in walnut. Plant Physiology 128, 282–290.
Cohen Y., Moreshet S. & Fuchs M. (1987) Changes in hydraulicconductance of citrus trees following a reduction in wetted soilvolume. Plant, Cell & Environment 10, 53–57.
Collins D.B.G. & Bras R.L. (2007) Plant rooting strategies in water-limited ecosystems. Water Resources Research 43, 64–70.
Cruiziat P., Cochard H. & Ameglio T. (2002) Hydraulic architec-ture of trees: main concepts and results. Annals of Forest Science7, 723–752.
Domec J.-C. & Gartner B.L. (2002) Age and position-related inhydraulic versus mechanical dysfunction of xylem: inferring thedesign criteria for Douglas-fir wood structure.Tree Physiology 22,91–104.
Domec J.-C. & Pruyn M.L. (2008) Bole girdling affects metabolicproperties and root, trunk and branch hydraulics of young pon-derosa pine trees. Tree Physiology 28, 1493–1504.
Domec J.-C., Warren J.M., Meinzer F.C., Brooks J.R. & CoulombeR. (2004) Native root xylem embolism and stomatal closure instands of Douglas-fir and ponderosa pine: mitigation by hydrau-lic redistribution. Oecologia 141, 7–16.
Domec J.-C., Pruyn M.L. & Gartner B.L. (2005) Axial and radialprofiles in conductivities, water storage and native embolism intrunks of young and old growth ponderosa pine trees. Plant, Cell& Environment 28, 1103–1113.
Domec J.-C., Scholz F.G., Bucci S.J., Meinzer F.C., Goldstein G. &Villalobos Vega R. (2006) Diurnal and seasonal variation in rootxylem embolism in neotropical savanna woody species: impacton stomatal control of plant water status. Plant, Cell & Environ-ment 29, 26–35.
Ewers B.E. & Oren R. (2000) Analysis of assumptions and errors inthe calculation of stomatal conductance from sap-flow measure-ments. Tree Physiology 20, 579–589.
Ewers B.E., Oren R. & Sperry J.S. (2000) Influence of nutrientversus water supply on hydraulic architecture and water balancein Pinus taeda. Plant, Cell & Environment 23, 1055–1066.
Ewers B.E., Gower S.T., Bond-Lamberty B. & Wang C.K. (2005)Effects of stand age and tree species on canopy transpiration andaverage stomatal conductance of boreal forests. Plant, Cell &Environment 28, 660–678.
Franks P.J. & Farquhar G.D. (1999) A relationship between humid-ity response, growth form and photosynthetic operating point inC3 plants. Plant, Cell & Environment 22, 1337–1349.
Granier A. (1987) Sap flow measurement in Douglas fir stems usinga new thermal method. Annales Des Sciences Forestieres 44, 1–14.
Granier A., Loustau D. & Bréda N. (2000) A generic model offorest canopy conductance dependent on climate, soil wateravailability and leaf area index. Annales Des Sciences Forestieres57, 755–765.
Granier A., Reichstein M., Bréda N., et al. (2007) Evidence for soilwater control on carbon and water dynamics in European forestsduring the extremely dry year: 2003. Agricultural and ForestMeteorology 143, 123–145.
Hacke U.G., Sperry J.S., Ewers B.E., Ellsworth D.S., Schafer K.V.R.& Oren R. (2000) Influence of soil porosity on water use in Pinustaeda. Oecologia 124, 495–505.
Iiames J.S., Congalton R., Andrew P. & Timothy L. (2008) Valida-tion of an integrated estimation of loblolly pine (Pinus taeda L.)leaf area index using two indirect optical methods in the south-eastern United States. Southern Journal of Applied Forestry 32,101–110.
Irvine J., Perks M.P., Magnani F. & Grace J. (1998) The response ofPinus sylvestris to drought: stomatal control of transpiration andhydraulic conductance. Tree Physiology 18, 393–402.
Irvine J., Law B.E., Kurpius M.R., Anthoni P.M., Moore D. &Schwarz P.A. (2004) Age-related changes in ecosystem structureand function and effects on water and carbon exchange inponderosa pine. Tree Physiology 24, 753–763.
James S., Clearwater M.J., Meinzer F.C. & Goldstein G. (2002) Heatdissipation sensors of variable length for the measurement ofsapflow in trees with deep sap wood.Tree Physiology 22, 277–283.
Jones H.G. (1992) Plants and Microclimate: A QuantitativeApproach to Environmental Plant Physiology. CambridgeUniversity Press, Cambridge, UK.
Katul G., Leuning L. & Oren R. (2003) Relationship between planthydraulic and biochemical properties derived from a steady-state coupled water and carbon transport model. Plant, Cell &Environment 26, 339–350.
Kim H.-S., Oren R. & Hinckley T.M. (2008) Actual and potentialtranspiration and carbon assimilation in an irrigated poplar plan-tation. Tree Physiology 28, 559–577.
Loustau D., Domec J.C. & Bosc A. (1998) Interpreting the varia-tions in xylem sap flux density within the trunk of maritime pine(Pinus pinaster Ait.): application of a model for calculating waterflows at tree and stand levels. Annales Des Sciences Forestieres55, 29–46.
Luo Y., Medlyn B., Hui D., Ellsworth D., Reynolds J. & Katul G.(2001) Gross primary productivity in Duke Forest: modelingsynthesis of CO2 experiment and eddy-flux data. EcologicalApplications 11, 239–252.
McDowell N.G., White S. & Pockman W.T. (2008) Transpiration
990 J.-C. Domec et al.
© 2009 Blackwell Publishing Ltd, Plant, Cell and Environment, 32, 980–991

and stomatal conductance across a steep climate gradient in thesouthern Rocky Mountains. Ecohydrology 1, 193–204.
Magnani F. & Borghetti M. (1995) Interpretation of seasonalchanges of xylem embolism and plant hydraulic resistance inFagus sylvatica. Plant, Cell & Environment 18, 689–696.
Maier C.A., Palmroth S. & Ward E. (2008) Branch growth and gasexchange in 13-year-old loblolly pine (Pinus taeda) trees inresponse to elevated carbon dioxide concentration and fertiliza-tion. Tree Physiology 28, 1093–1106.
Maseda P.H. & Fernández R.J. (2006) Stay wet or else: three waysin which plants can adjust hydraulically to their environment.Journal of Experimental Botany 57, 3963–3977.
Meinzer F.C. (2002) Co-ordination of vapour and liquid phasewater transport properties in plants. Plant, Cell & Environment25, 265–274.
Meinzer F.C., Goldstein G., Jackson P., Holbrook N.M., GutierrezM.V. & Cavelier J. (1995) Environmental and physiological regu-lation of transpiration in tropical forest gap species: the influenceof boundary layer and hydraulic properties. Oecologia 101, 514–522.
Monteith J.L. (1995) A reinterpretation of stomatal responses tohumidity. Plant, Cell & Environment 18, 357–364.
Mott K. & Buckley T. (1998) Stomatal heterogeneity. Journal ofExperimental Botany 49, 407–417.
Motulsky H. & Christopoulos A. (2004) Fitting Models to BiologicalData Using Linear and Nonlinear Regression:A Practical Guide toCurve Fitting. Oxford University Press, New York, NY, USA.
Nardini A., Salleo S., Trifillo P. & Lo Gullo M.A. (2003) Waterrelations and hydraulic characteristics of three woody speciesco-occurring in the same habitat. Annals of Forest Science 60,297–305.
Neumann P.M. (1987) Sequential leaf senescence and correlativelycontrolled increases in xylem flow resistance. Plant Physiology83, 941–944.
Noormets A., Gavazzi M., McNulty S., Sun G., Domec J.-C., King J.& Chen J. (2009) Response of carbon fluxes to drought in acoastal plain loblolly pine forest. Global Change Biology.doi:10.1111/j.1365-2486.2009.01928.x.
Oishi A.C., Oren R. & Stoy P.C. (2008) Estimating components offorest evapotranspiration: a footprint approach for scaling sapflux measurements. Agricultural Forest and Meteorology 148,719–732.
Oren R., Phillips N., Katul G.G., Ewers B.E. & Pataki D.E. (1998)Scaling xylem sap flux and soil water balance and calculatingvariance. Annales des Sciences Forestieres 55, 191–216.
Oren R., Sperry J.S., Katul G.G., Pataki D.E., Ewers B.E., Phillips N.& Schäfer K.V.R. (1999) Survey and synthesis of intra- andinterspecific variation in stomatal sensitivity to vapour pressuredeficit. Plant, Cell & Environment 22, 1515–1526.
Passioura J.B. (1988) Water transport in and to roots. AnnualReview of Plant Physiology and Plant Molecular Biology 39,245–265.
Phillips N., Nagchaudhuri A., Oren R. & Katul G. (1997) Timeconstant for water transport in loblolly pine trees estimated fromtime series of evaporative demand and stem sapflow. Trees 11,412–419.
Phillips N., Bond B.J., McDowell N.G. & Ryan M. (2002) Canopyand hydraulic conductance in young, mature and old Douglas-firtrees. Tree Physiology 22, 205–211.
Richter H. (1997) Water relations of plants in the field: some com-ments on the measurement of selected parameters. Journal ofExperimental Botany 48, 1–7.
Rosner S., Klein A., Wimmer R. & Karlsson B. (2006) Extraction offeatures from ultrasound acoustic emissions: a tool to assess thehydraulic vulnerability of Norway spruce trunkwood? New Phy-tologist 171, 106–116.
Rosner S., Klein A., Muller U. & Karlsson B. (2008) Tradeoffsbetween hydraulic and mechanical stress responses of matureNorway spruce trunk wood. Tree Physiology 28, 1179–1188.
Rundel P.W. & Yoder B.J. (1998) Ecophysiology of Pinus. InEcology and biogeography of Pinus (ed. D.M. Richardson) pp.296–323. Cambridge University Press, Cambridge, UK.
Sack L. & Holbrook N.M. (2006) Leaf hydraulics. Annual Reviewof Plant Biology 57, 361–381.
Sack L., Cowan P.D., Jaikumar N. & Holbrook N.M. (2003) The‘hydrology’ of leaves: co-ordination of structure and function intemperate woody species. Plant, Cell & Environment 26, 1343–1356.
Salleo S., Nardini A., Pitt F. & Lo Gullo M.A. (2000) Xylem cavi-tation and hydraulic control of stomatal conductance in laurel(Laurus nobilis L.). Plant, Cell & Environment 23, 71–79.
Schenk H.J. & Jackson R.B. (2005) Mapping the global distributionof deep roots in relation to climate and soil characteristics. Geo-derma 126, 129–140.
Sellin A. & Kupper P. (2005) Variation in leaf conductance of silverbirch: effects of irradiance, vapour pressure deficit, and leafwater status and position within a crown. Forest Ecology andManagement 206, 153–166.
Siau J.F. (1984) Transport Processes in Wood. Springer-Verlag,Berlin, Germany.
Sperry J.S. & Pockman W.T. (1993) Limitation to transpiration byhydraulic conductance and xylem cavitation in Betula occiden-talis. Plant, Cell & Environment 16, 279–287.
Sperry J.S., Hacke U.G., Oren R. & Comstock J.P. (2002) Waterdeficits and hydraulic limits to leaf water supply. Plant, Cell &Environment 25, 2251–2263.
Steudle E. (2000) Water uptake by plant roots: an integration ofviews. Plant & Soil 226, 45–56.
Thérézien M., Palmroth S., Brady R. & Oren R. (2007) Estimationof light interception properties of conifer shoots by an improvedphotographic method and a 3D model of shoot structure. TreePhysiology 27, 1375–1387.
Ward E.J., Oren R., Sigurdsson B.D., Jarvis P.G. & Linder S. (2008)Fertilization effects on mean stomatal conductance are mediatedthrough changes in the hydraulic attributes of mature Norwayspruce trees. Tree Physiology 28, 579–596.
Warren J.M., Meinzer F.C., Brooks J.R. & Domec J.-C. (2005) Ver-tical stratification of soil water storage and release dynamics inPacific Northwest Coniferous Forests. Agricultural and ForestMeteorology 130, 39–58.
Whitehead D. (1998) Regulation of stomatal conductance andtranspiration in forest canopies. Tree Physiology 18, 633–644.
Woodruff D.R., McCulloh K.A., Warren J.M., Meinzer F.C. &Lachenbruch B. (2007) Impacts of tree height on leaf hydraulicarchitecture and stomatal control in Douglas-fir. Plant, Cell &Environment 30, 559–569.
Zou C., Sands R. & Sun O. (2000) Physiological responses ofradiate pine roots to soil strength and soil water deficit. TreePhysiology 20, 1205–1207.
Zwieniecki M.A., Hutyra L., Thompson M.V. & Holbrook N.M.(2000) Dynamic changes in petiole specific conductivity in redmaple (Acer rubrum L.), tulip tree (Liriodendron tulipifera L.)and northern fox grape (Vitis labrusca L.). Plant, Cell & Envi-ronment 23, 407–414.
Zwieniecki M.A., Stone H.A., Leigh A., Boyce K. & HolbrookN.M. (2006) Hydraulic design of pine needles: one-dimensionaloptimization for single-vein leaves. Plant, Cell & Environment29, 803–809.
Received 25 November 2008; received in revised form 5 March 2009;accepted for publication 15 March 2009
Influence of leaf and root hydraulic conductances on stomatal conductance 991
© 2009 Blackwell Publishing Ltd, Plant, Cell and Environment, 32, 980–991