defective export in escherichia coli caused by dsba …jb.asm.org/content/179/10/3260.full.pdf ·...
TRANSCRIPT

JOURNAL OF BACTERIOLOGY,0021-9193/97/$04.0010
May 1997, p. 3260–3269 Vol. 179, No. 10
Copyright © 1997, American Society for Microbiology
Defective Export in Escherichia coli Caused by DsbA9-PhoAHybrid Proteins Whose DsbA9 Domain Cannot Fold into
a Conformation Resistant to Periplasmic ProteasesAGNES GUIGUENO, PASCAL BELIN, AND PAUL L. BOQUET*
Departement d’Ingenierie et d’Etudes des Proteines, CEA, CE Saclay,91191 Gif-sur-Yvette Cedex, France
Received 6 December 1996/Accepted 12 March 1997
The disulfide bond-forming factor DsbA and the alkaline phosphatase are stable in the Escherichia coliperiplasmic space and can be overproduced without significant perturbation of the cell’s physiology. Bycontrast, DsbA*-PhoA hybrid proteins resulting from TnphoA insertions into different regions of a plasmid-borne dsbA gene could become toxic (lethal) to bacteria. Toxicity was concomitant with an impairment of somestep of the export mechanism and depended on at least three parameters, i.e., (i) the rate of expression of thehybrid protein, (ii) the ability of the amino-terminal DsbA* domain of the hybrid protein to fold into aprotease-resistant conformation in the periplasmic space, and (iii) the activity of the DegP periplasmicprotease. Even under viable conditions of low expression, DsbA* folding-deficient hybrid proteins accumulatedmore than the folding-proficient ones in the insoluble material and this was aggravated in a strain lacking theDegP protease. When production was more elevated, the folding-deficient hybrid proteins became lethal, butonly in strains lacking the DegP activity, while the folding-proficient ones were not. Under conditions of veryhigh production by degP1 or degP strains, both types of hybrid proteins accumulated as insoluble preproteins.Meanwhile, the export machinery was dramatically handicapped and the cells lost viability. However, thefolding-deficient hybrid proteins had a higher killing efficiency than the folding-proficient ones. Free DsbA*-truncated polypeptides, although not toxic, were processed more slowly when they could not fold into aprotease-resistant form in the periplasmic space. This provides indications in E. coli for a direct or indirectinfluence of the folding of a protein in the periplasmic environment on export efficiency.
In Escherichia coli, periplasmic proteins are synthesized aspreproteins and most of them are exported by the Sec machin-ery (26, 49, 53). Their export can be initiated by preliminaryinteraction with cytoplasmic chaperones such as SecB, GroES-GroEL, or DnaK-DnaJ in order to maintain them in a trans-location-competent state (9, 31, 32, 47, 51, 60, 61). The pre-cursor protein then interacts with SecA, the ATP-driventranslocase of the cytoplasmic membrane, and the signal pep-tide inserts into the lipid bilayer (30). In the membrane, SecAis in close contact with the transmembrane protein complexSecYEG, and the four proteins constitute the core of thesecretion apparatus (18, 25, 28, 44, 59). SecA drives successivesegments of the exported protein by a back-and-forth move-ment coupled with cycles of ATP hydrolysis (20). Transloca-tion is also assisted by the electric charge of the membrane(19). SecD and SecF, which are also membrane bound butprotrude into the periplasm, facilitate the export (4, 20, 48),possibly by stimulating the release of the processed polypep-tide into the periplasmic space (37). Processing itself isachieved on the periplasmic side of the cytoplasmic membraneby the action of the Lep peptidase (12, 14, 62). The term exportrefers hereafter to this whole mechanism.
Once in the periplasm, the exported proteins must haveacquired a folded tridimensional structure to avoid their deg-radation by the multiple proteases of this compartment (for areview, see reference 10). Folding is assisted by soluble ormembrane-bound enzymes such as the Dsb factors, which areinvolved in disulfide bond formation or reshuffling (5, 6, 29, 40,
41), and probably SurA and FkpA, with peptidyl prolyl cis-transisomerizing activity (42).
It may be asked whether the ability or inability of a secretedprotein to fold rapidly into a compact structure once in theperiplasmic environment could influence the rate at which thispolypeptide is exported across the membrane. We have at-tempted to answer this question by comparing the export ofseveral DsbA9-PhoA hybrid proteins already described (8).They differed only by the lengths and the capacities of theirDsbA9 amino-terminus-containing regions to acquire a folded,protease-resistant conformation in the periplasmic space. Thepositions of the fusion joints in such hybrid proteins, whichresulted from TnphoA insertions into a multicopy plasmid-borne dsbA gene, are schematized in Fig. 1. The plasmidsencoding the amino acid residues 1 to 160 of mature DsbAfused to PhoA [DsbA(1–160)-PhoA] or fusions with longerDsbA9 amino-terminal segments promoted the export to theperiplasmic space of stable hybrid proteins (class I proteins),which possessed disulfide bond-forming and alkaline phospha-tase activities (as deduced from the DsbA1 PhoA1 phenotypeconfered by such plasmids to dsbA phoA strains). By contrast,the hybrid proteins in which DsbA9 was equal to or shorterthan the 1 to 136 segment (class II) were partly proteolyzed inthe periplasmic space of a dsbA1 strain, yielding a 47-kDastable and active form of alkaline phosphatase. In a dsbArecipient strain, the plasmids expressing the DsbA9-PhoA hy-brid proteins of class II did not restore a DsbA activity and theentire protein was digested (8). Therefore, the region of DsbAfrom 136 to 160, which includes the unique (cis) Pro151 of themolecule, carries one or more amino acid residues which areimportant for the folding of the DsbA9 part of the hybrid
* Corresponding author. Phone: (33) 1 69 08 72 90. Fax: (33) 1 69 0890 71. E-mail: [email protected].
3260
on May 26, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

protein into a proteolysis-resistant and enzymatically activeconformation.
We show here that in dsbA1 strains, the unstable hybridproteins of class II, albeit shorter, are more readily retainedthan those of class I within the insoluble cell fraction, a defectaggravated in bacteria lacking the DegP protease. They alsoinhibit the export (or perhaps only the processing step) ofother periplasmic proteins more efficiently than those of classI. The possible direct or indirect consequences of this foldingdefect on the different steps necessary for the export are dis-cussed.
MATERIALS AND METHODS
Strains, plasmids, and media. All of the strains used were derivatives of the E.coli K-12 prototrophic strain K10. Their genotypes and origins are listed in Table1. The characteristics and origins of the plasmids already described or con-structed for the present study are also indicated in Table 1.
Unless otherwise stated, bacteria were grown in LB broth (39), to which therequired antibiotics were added at the final following concentrations: ampicillin,50 mg/ml; kanamycin, 30 mg/ml; chloramphenicol, 20 mg/ml; streptomycin andspectinomycin, 10 mg/ml each; and tetracycline 20 mg/ml. Minimal medium wasaccording to Davis and Mingioli (15) and contained glucose (0.4%) as a carbonsource unless otherwise indicated and thiamine (0.01%). This medium was com-plemented with a mixture of all amino acids (50 mg/ml) except methionine andcysteine for labelling experiments (DMAA).
Recombinant DNA techniques. Transformations and other recombinant DNAtechniques necessary for plasmid constructions were according to the meth-ods described by Maniatis et al. (36). The detailed construction of plasmidspAG2753, pAG2784, pAG2884, and pAG2885 is indicated in Fig. 2 and itslegend.
Directed mutagenesis of the degP and dsbA genes. The 2.8-kb ClaI-NruI frag-ment encoding the DegP protein was subcloned into the ClaI and NruI sites ofpBR322, yielding pAG3145. The Ser2103Ala210 mutation was made according tothe procedure developed by Deng and Nickoloff (16) on pAG3145 with theUnique Site Elimination mutagenesis kit (Pharmacia) following the instructionsof the manufacturer. The ScaI site in the plasmid-borne bla gene (AGTACT)was replaced by a MluI site (ACGCGT) as a consequence of the method used,without change in ampicillin resistance. The ScaI/MluI unique site eliminationselection toggle primer employed was 59p(CTGTGACTGGTGACGCGTCAACAAAGTC)39, and the target mutagenic primer was 59p(CAACCGTGGTAACGCAGGTGGTGCGCTGG)39. The sequence of the degPS210A gene (54) wasverified. Then, we subcloned the EcoRI-NruI fragment carrying the mutateddegP gene into the EcoRI and SmaI sites of the low-copy-number plasmidpHSG575, yielding pAG3183. The EcoRI-NruI fragment carrying the wild-typedegP gene was also subcloned into the low-copy-number plasmid pHSG575 (56)to give pAG3182.
Two consecutive termination codons (TAA TAG) were introduced in place ofthe (466)CAG CAG(471) sequence of the dsbA gene (corresponding to Q137-Q138of mature DsbA) carried by plasmid pPB2216 (7). Mutagenesis on double-stranded plasmid DNA with the U.S.E. mutagenesis kit (Pharmacia) was performedas described above. The ScaI/MluI U.S.E. selection toggle primer used was asdescribed above, and the target mutagenic primer was 59p(CTCTGGTCGCTTAATAGGAAAAAGCTGC)39. The entire sequence of the dsbA1–136 gene ob-tained on pPB2928 was verified. Plasmid pPB2368 [encoding DsbA(1–163)], whichcarried a TAA termination triplet in place of the CAG codon corresponding tothe Q164 residue of mature DsbA, was obtained by this technique. Its construc-tion will be reported in more details in a forthcoming publication.
Enzymatic activities. Alkaline phosphatase activities were measured as previ-ously described (8). NADH oxidase activity was measured according to themethod described by Osborn et al. (46).
Bacterial fractionation for the analysis of DsbA*-PhoA subcellular reparti-tion. Bacteria were grown in LB (ampicillin) broth, and 2-ml samples werecollected at the A600 corresponding to the midexponential phase. One milliliter
was centrifuged and submitted to an osmotic shock according to Neu and Heppel(43). The other milliliter was centrifuged and washed by resuspension in 1 ml of10 mM Tris-HCl (pH 8)–1 mM EDTA. The cells were broken at 0°C by sonicoscillation twice for 2 min at 1,200 J/W/s. Unbroken cells were removed bycentrifugation at 10,000 3 g for 5 min, and the supernatant was spun down for2 h at 50,000 3 g. The pellet constituted the insoluble fraction. Bacteria of strainsSBS2027pAG2884 and SBS2027pAG2885 were grown and collected similarlyeither 1 h after induction (1 mM isopropyl-b-D-thiogalactopyranoside [IPTG]) orfrom uninduced cultures. The material sedimenting at 50,000 3 g was preparedas described above. Bacteria of strain SBS2027pAG2884 (200-ml cultures),grown and induced under the same conditions or uninduced were sedimented,washed in 10 mM Tris-HCl (pH 8.0)–1 mM EDTA, resuspended into the samebuffer, and twice broken by sonic oscillation. DNase (100 mg/ml) and MgCl2 (2mM) were added to the extracts and allowed to stand at 0°C for 15 min.Unbroken bacteria were removed by low-speed centrifugation, and the superna-tants were centrifuged at 50,000 3 g for 2 h. The sedimented material (insolublefraction) was washed and layered on top of a sucrose density gradient as de-scribed elsewhere (46).
SDS-PAGE analysis and immunodetections (Western blots). Sodium dodecylsulfate-polyacrylamide gel electrophoresis (SDS-PAGE) was performed accord-ing to Laemmli (33). The gels contained 7 to 15% acrylamide according to theexperiments. Proteins were transferred to nitrocellulose membranes (Schleicher& Schuell; 0.45 mm) with a semidry Trans-Blot apparatus (Bio-Rad) for 1 hunder 100 V. The transfer buffer contained glycine (39 mM), Tris (48 mM), SDS(0.037%), and methanol (20%). The membranes were then soaked for 2 h withagitation into a 2% bovine serum albumin in phosphate-buffered saline (PBS)buffer (100 mM Na phosphate [pH 8.0], 1.54 M NaCl) for saturation. Antibodyfixations and washings were made on the membranes in PBS plus 0.1% Tween 20(Sigma). Monoclonal anti-PhoA antibodies were diluted 1:10,000 and allowed toreact overnight. After five washes, the membrane was soaked in the presence ofperoxidase-coupled anti-mouse goat antibodies (1:5,000) for 2 h. After five otherwashes, peroxidase was detected by diaminobenzidine tetrahydrochloride (Sig-ma) as a 0.5% solution in 100 mM Tris-HCl (pH 7.6). The relative amounts of
FIG. 1. Positions of the fusion joints in the DsbA9-PhoA hybrid proteins. TheDsbA protein and its leader peptide (LP) are represented. For each hybridprotein, the arrows indicate the positions of each last amino acid residue ofmature DsbA bound to the modified PhoA protein encoded by TnphoA. The twounderlined positions correspond to the hybrid proteins chiefly used in this studyas typical of each class and showing the smallest size difference.
TABLE 1. Characteristics of strains and plasmids used
Strain orplasmid Description Source or
reference
E. coli strainsK10 HfrC relA pit-10 tonA22 spoT1
garB10 ompF627B. Bachmann
SBS2027 Same as K10, but DphoA20 This laboratorySBS2146 Same as K10, but degP::Tn5 This laboratorySBS2597 Same as K10, but DphoA20
recA::camThis study
SBS2603 Same as K10, but DphoA20degP::Tn5
This study
SBS2604 Same as K10, but DphoA20degP::Tn5 recA::cam
This study
PlasmidspBR322 Apr Tcr J. BeckwithpHSG575 Cmr 56pTrc99A Apr PharmaciapPB2325 Apr Tcs::dsbA1–136::TnphoA Kmr This laboratory (8)pPB2326 Apr Tcs::dsbA1–160::TnphoA Kmr This laboratory (8)pAG2694 Apr Tcs::dsbA1–136::phoA (a 4.8-kb
DXhoI deletion on plasmidpPB2325) Kms
This study (Fig. 2)
pAG2753 Cmr dsbA1–136::phoA Kms This study (Fig. 2)pAG2784 Cmr dsbA1–160::phoA Kms This study (Fig. 2)pAG2884 Apr dsbA1–136::phoA Kms This study (Fig. 2)pAG2885 Apr dsbA1–160::phoA Kms This study (Fig. 2)pAG2993 Apr Tcs::dsbA1–160::TnphoA
(a 4.8-kb DXhoI deletion onplasmid pPB2326) Kms
This study (Fig. 2)
pPB2928 Apr dsbA1–136 This studypPB2368 Apr dsbA1–163 This laboratorypAG3145 Apr degP This studypAG3182 Cmr degP This studypAG3183 Cmr degPS210A This studypEMR602 Tcr Cms::groES groEL O. FayetpEMR675 Tcr Cms::secB O. FayetpAPKJ-145 Spr Kmr lacIq dnaK dnaJ O. Fayet
VOL. 179, 1997 DsbA9-PhoA HYBRID PROTEIN FOLDING AND EXPORT 3261
on May 26, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

hybrid proteins on each Western blot were evaluated by densitometric tracerecording with a Bioprobe Densylab system. Photographs were taken with aKodak Wratten 80D blue filter on TMax 100 film.
Determination of amino-terminal sequences of hybrid proteins. Hybrid pro-teins from sucrose gradient-prepared fractions or from crude membrane prep-arations were separated by SDS-PAGE, blotted onto Immobilon membranes(Millipore), and analyzed with an Applied Biosystems 477A gas-phase proteinsequencer and by on-line automated high-pressure liquid chromatography.
Pulse-chase experiments. The labelling medium was DMAA plus 0.4% glu-cose or maltose. Cells were pulse-labelled with 20 mCi of [35S]methionine pluscysteine (Promix) per ml for 15 to 30 s and were chased with unlabelled methi-onine plus cysteine (500-mg/ml each). Samples (0.5 ml) were collected intoprecooled tubes containing 0.5 ml of 10% trichloroacetic acid. After centrifuga-tion and washing by acetone, the pellets were resuspended in 50 mM Tris-HCl(pH 8)–1% SDS–1 mM EDTA and heated at 100°C for 5 min. Antibody pre-cipitation and SDS-PAGE analysis were as described by Ito et al. (27).
RESULTS
Class II DsbA*-PhoA hybrid proteins become toxic to E. coliwhen expressed in a degP strain. Our initial aim was to deter-mine whether the absence of DsbA activity in class II DsbA9-PhoA hybrid proteins could simply be attributed to their highinstability in the periplasmic space. A preliminary pulse-chaseexperiment indicated that DsbA(1–136)-PhoA (the longest hy-brid protein of class II) was cleaved near the fusion joint intostable PhoA and a very unstable DsbA9 fragment (data notshown). The periplasmic endopeptidase DegP (HtrA), whichwas shown to display a restricted specificity (35) and to beresponsible for the release of free PhoA from several PhoA-containing periplasmic hybrid proteins (55), was presumablyinvolved in such degradation. To analyze the role of DegP,each of the 14 pBR322-derived plasmids carrying the dsbA9-TnphoA fusions (Fig. 1) was used to transform bacteria ofstrain SBS2597 (DphoA recA) and of its degP::Tn5 isogenicderivative SBS2604 at 30°C. With SBS2597, the yield of trans-formation by any of the plasmids was comparable to that bypBR322. In SBS2604, transformants were obtained with plas-mids encoding class I hybrid proteins (all with a strong PhoA1
phenotype), but not with plasmids encoding those of class II,which gave only rare colonies (about 104-fold less than ex-pected) with various PhoA activities. One exception was theplasmid encoding the shortest hybrid protein available,DsbA(1–7)-PhoA, which gave a number of transformants com-parable to that obtained with a class I plasmid. Plating onminimal medium or deletion of the XhoI-XhoI DNA fragmentencoding most of the Tn5 genes in TnphoA from the plasmidscould not restore the transformability of SBS2604 by plasmidsof class II. The same results were obtained with degP rec1
recipient strains. The few colonies above mentioned wereshown to correspond to mutations in the plasmids themselves(modifications of the dsbA promoter or deletions in the dsbA9region of the fused genes) and, in some cases, in the chromo-some. In such cases, their own plasmids were not tolerated inthe original degP strain and, once cured from these plasmids,they could still be retransformed efficiently by the fusion plas-mids initially used. These chromosomal mutations have not yetbeen characterized.
It was questionable whether the relative protection affordedby DegP was actually due to its proteolytic activity or to some-thing else, such as, for instance, a chaperone activity in theperiplasmic space as proposed elsewhere (11). We constructedplasmids pAG3182, which encodes DegP, and pAG3183, whichencodes the DegP-modified protein carrying a Ser210-to-Ala210substitution as indicated in Materials and Methods. This pro-tein was shown to have no proteolytic activity (54). Bacteria ofstrain SBS2146 (degP) were transformed with each of theseplasmids, and transformants were assayed for their abilities totolerate the compatible pBR322-derived plasmids encoding
class II hybrid proteins. The presence of pAG3183 did notchange the previous results, while pAG3182 restored the tol-erance of class II fusion plasmids as expected.
Consequently, with the exception of that corresponding toDsbA(1–7)-PhoA, the plasmid-borne fused genes encodingclass II hybrid proteins were toxic to bacteria deprived of DegPproteolytic activity while those encoding class I hybrid proteinswere not.
Relation between the toxicity of DsbA*-PhoA hybrid pro-teins and their rate of synthesis. With the hope of determiningwhether the toxicity of the plasmids correlated with theamount of hybrid protein synthesized, we first subcloned aDNA fragment encoding DsbA(1–136)-PhoA (the longest hy-brid protein of class II) without its promoter and Shine-Dal-garno sequence under the control of the arabinose promoteron vector pBAD24 (23). However, even under conditions thatwere expected to be fully inducing, the plasmid was not toxic(not shown). The reasons for such a tolerance are under in-vestigation.
In other attempts to answer the question, DNA fragmentsencoding DsbA(1–136)-PhoA or DsbA(1–160)-PhoA (the short-est hybrid protein of class I) were subcloned into vectorspHSG575 (56) and pTrc99A (2), which differed by their originof replication (copy number) and by the promoter used totranscribe the fusion (see Materials and Methods). In thepHSG575-derived low-copy-number plasmids pAG2753 [en-coding DsbA(1–136)-PhoA of class II] and pAG2784 [encodingDsbA(1–160)-PhoA of class I], the fusions (Fig. 2) were tran-scribed from the weak dsbA P1 proximal promoter (7). Theywere used to transform bacteria of strain SBS2027 (DphoA)and its degP derivative SBS2603. Both plasmids were toleratedin the two strains, and the yields of transformation were com-parable to those of the vector alone. Accordingly, the globalalkaline phosphatase activity (originating from the hybrid pro-teins themselves and from their PhoA-like degradation prod-ucts) was lower in SBS2027 carrying the low-copy-number fu-sion plasmids than in the same strain with the pBR322-derivedfusion plasmids (Table 2). All other class II gene fusions weresimilarly tolerated in this low-copy-number vector.
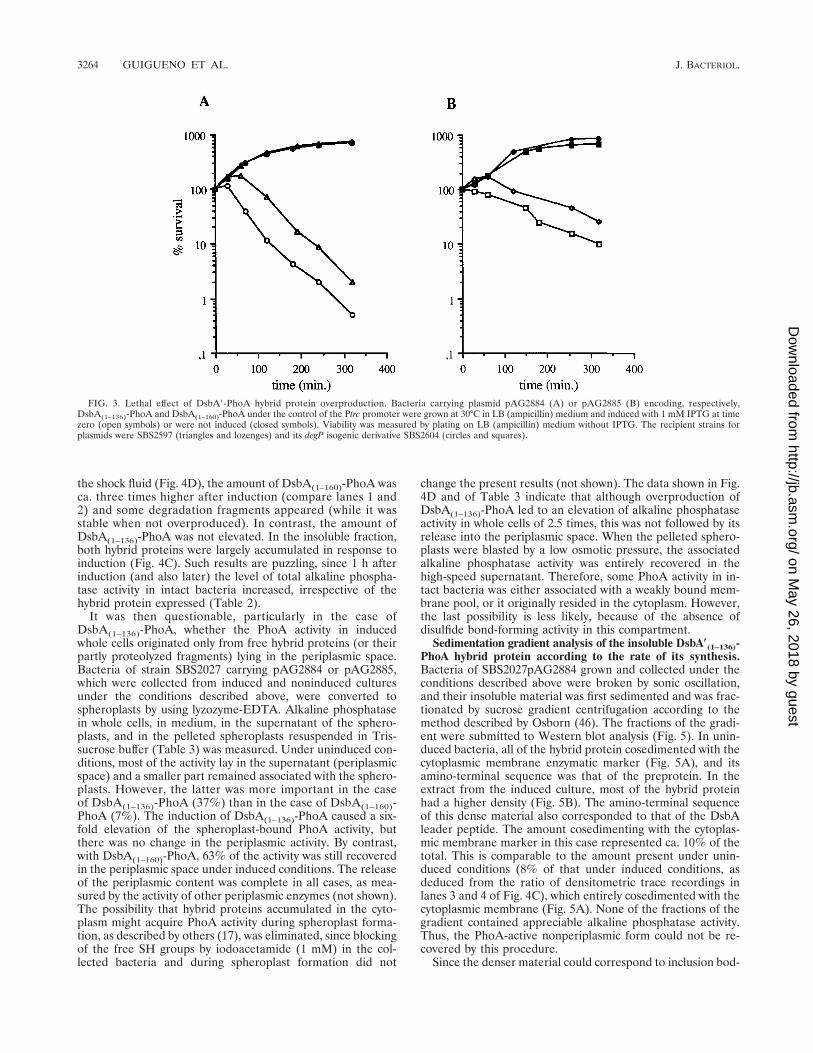
DNA fragments encoding the same DsbA(1–136)-PhoA andDsbA(1–160)-PhoA hybrid proteins were subcloned into thehigh-copy-number vector pTrc99A in such a way that theirtranscription was entirely dependent on the inducible Ptrc (2)strong promoter (Fig. 2). The recombinant plasmids pAG2884and pAG2885 were used to transform bacteria of the isogenicstrains SBS2597 and SBS2604 (degP) in the absence of theinducer IPTG. In all cases, transformants were obtained with anormal yield. In liquid cultures, their viability began to fall 2 hafter induction with 1 mM IPTG (Fig. 3). However, the over-production of the class II hybrid protein (Fig. 3A) was moretoxic than that of the class I (Fig. 3B), and viable counts alsodecreased earlier in the degP strain than in the degP1 strain.The global alkaline phosphatase activity resulting from theexpression of the two different hybrid proteins (and/or of thefree PhoA fragments) increased after induction, even after afew hours, although bacteria lost their viability (Table 2).There was a ca. 20% difference between the activities originat-ing from the two different hybrid proteins. Reducing the con-centration of the inducer on LB plates to 60 mM IPTG showedthat the growth of SBS2597 (degP1) was less affected than thatof SBS2604 (degP) by the expression of DsbA(1–136)-PhoAfrom pAG2884. Similarly, in SBS2597, the difference in toxicitybetween DsbA(1–136)-PhoA (pAG2884) and DsbA(1–160)-PhoA(pAG2885) was also more salient on LB plates containing 70mM IPTG or in minimal medium with 35 mM IPTG (notshown).
3262 GUIGUENO ET AL. J. BACTERIOL.
on May 26, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

Therefore, DsbA9-PhoA hybrid proteins of both classes be-came toxic, provided that their level of synthesis was elevatedabove a specific threshold. This threshold, however, was lowerwith those of class II and was still more reduced in the absenceof DegP protease activity. Whether a relationship could existbetween viability loss and the accumulation of hybrid proteinin an aberrant subcellular location was investigated.
Subcellular distribution of DsbA*-PhoA hybrid proteins un-der low- and high-expression conditions. The repartition ofDsbA(1–136)-PhoA and of DsbA(1–160)-PhoA in acellular ex-tracts of bacteria was analyzed according to the flow rate oftheir synthesis and to the allelic state of degP. Bacteria ofstrains SBS2027 (degP1) and SBS2603 (degP) carrying the low-copy-number plasmids pAG2753 [encoding DsbA(1–136)-PhoA]and pAG2784 [encoding DsbA(1–160)-PhoA] were grown in LBbroth at 30°C and collected in the mid-exponential phase at thesame A600. One half was submitted to an osmotic shock, andthe other half was washed and sonicated. The shock fluid and
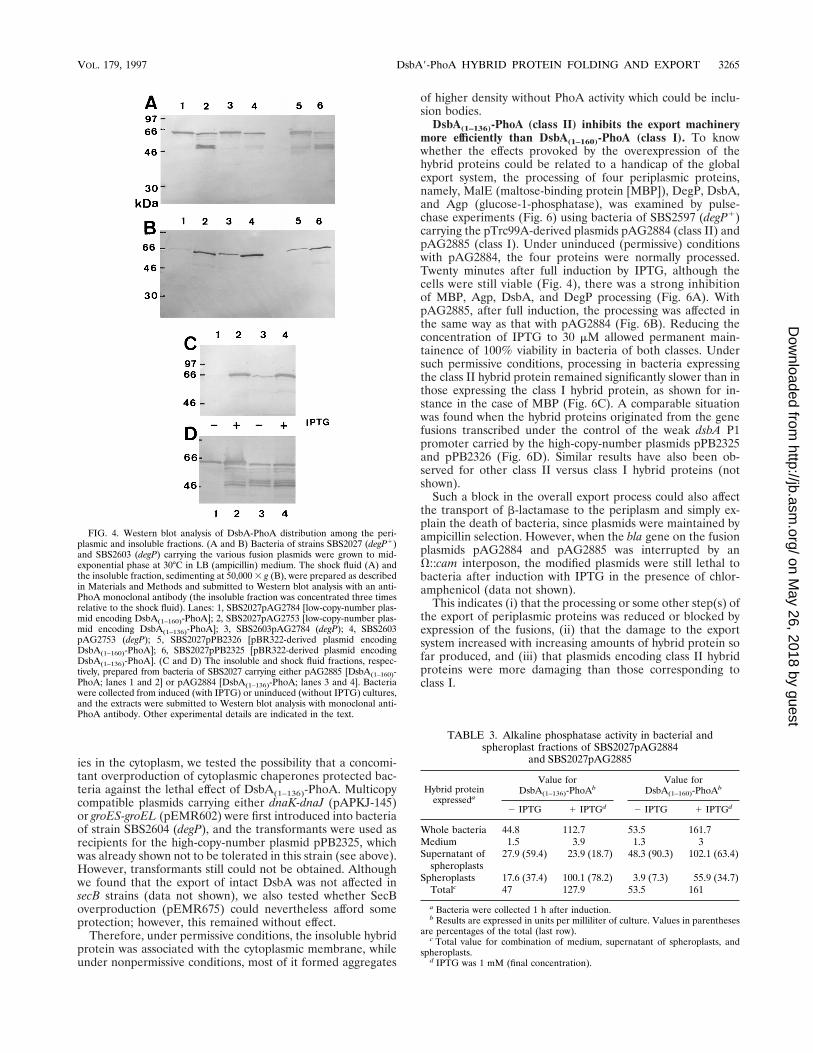
the insoluble material collected by high-speed centrifugation ofthe sonicate (see Materials and Methods) were quantitativelyanalyzed by Western blots with antibodies raised against PhoA(Fig. 4). A similar experiment with bacteria of strain SBS2027(degP1) carrying the pBR322-derived plasmids pPB2325[DsbA(1–136)-PhoA] and pPB2326 [DsbA(1–160)-PhoA] was per-formed. In the shock fluid of SBS2027pAG2753 (degP1), therewas four times less DsbA(1–136)-PhoA than DsbA(1–160)-PhoA,as judged by density recordings (Fig. 4A, lanes 2 and 1), andthe former was largely degraded into proteolyzed anti-PhoA-reacting fragments. The amount of such fragments was greatlyreduced in the degP strain (lane 4); however, the hybrid proteinwas still four times less abundant than DsbA(1–160)-PhoA inthis strain (lane 3). There was also a similar difference inabundance between the two intact different hybrid proteinsproduced in the degP1 strain from the pBR322-derived plas-mids (lanes 6 and 5). The insoluble material contained unex-pectedly large amounts of hybrid proteins. However, contraryto the situation in the periplasmic space, more DsbA(1–136)-PhoA than DsbA(1–160)-PhoA was accumulated in this fraction(Fig. 4B), i.e., 10 times more in SBS2027 (compare lanes 2 and1), 5 times more in SBS2603 (lanes 4 and 3), and 3 times morein SBS2027 carrying the pBR322-derived plasmids (lanes 6 and5). The inactivation of degP caused a two to three times greateraccumulation of both hybrid proteins (compare lanes 4 and 2and also lanes 3 and 1). A similar abnormal repartition be-tween the periplasmic and insoluble compartments was ob-served with all of the other class II hybrid proteins tested, whileother hybrid proteins belonging to class I behaved as DsbA(1–160)-PhoA (data not shown).
At first glance, it could possibly be concluded, taking intoaccount the presence of PhoA-containing degradation prod-ucts in the periplasm of degP1 strains (lanes 2 and 6 of Fig. 4Aand B), that more DsbA(1–136)-PhoA than DsbA(1–160)-PhoAwas made. The overall rates of synthesis of the two hybridproteins were compared by pulse-labelling bacteria of SBS2027carrying pPB2325 or pPB2326 for either 30 or 45 s. Autora-diograms of electrophoresis from total cell extracts immuno-precipitated with anti-PhoA antiserum did not show detectableamounts of degradation products after such short labellingperiods, and the density scanning of the hybrid protein bandson the autoradiograms indicated no difference (not shown).Consequently, the difference in the subcellular repartition ofthe two species originated from a different behavior at a post-translational step.
This repartition was also analyzed 1 h after the induction ofSBS2027 carrying pAG2884 or pAG2885 (Ptrc promoter) by 1mM IPTG, i.e., when bacteria were still 100% viable (Fig. 3),and in the uninduced bacteria collected at the same A600. In
FIG. 2. Construction of recombinant plasmids encoding DsbA(1–136)-PhoAand DsbA(1–160)-PhoA expressed at different levels. The boxed areas representDNA fragments originating from the pBR322-derived plasmids carrying theTnphoA fusions into dsbA (8). The filled and the darkly hatched areas corre-spond to dsbA fragments, and the lightly hatched areas correspond to the phoAregion of TnphoA. The open areas correspond to Tn5 genes. Restriction sitesinclude BamHI (B), BspHI (Bs), ClaI (C), HindIII (H), NcoI (Nc), PstI (P), andXhoI (X). (A) The pBR322-derived plasmid pPB2325, encoding DsbA(1–136)-PhoA under the control of the dsbA P1 promoter, was deleted from the XhoI-XhoI fragment which specifies the Kmr and the transposase genes of Tn5(pAG2694). The BspHI-BspHI fragment carrying the fusion (but lacking thedsbA P1 promoter) was subcloned from pAG2694 into the compatible NcoI siteof vector pTrc99A, yielding the high-copy-number plasmid pAG2884, in whichthe fusion was under the control of the strong inducible Ptrc promoter. TheBamHI-PstI fragment of pAG2694 was subcloned between the BamHI and PstIsites of the low-copy-number vector pHSG575, yielding pAG2753. (B) The XhoI-XhoI fragment was first deleted from plasmid pPB2326 encoding DsbA(1–160)-PhoA as described above (pAG2993). The BspHI-BspHI fragment of pAG2993was subcloned into the NcoI site of pTrc99A to give pAG2885, which is homol-ogous to pAG2884. The construction of the pHSG575-derived plasmid pAG2784was obtained by replacing the BamHI-XhoI fragment of pAG2753 (panel A) withthe homologous fragment taken from pAG2993.
TABLE 2. Alkaline phosphatase activity in cultures ofbacteria carrying dsbA-phoA fusion plasmids
Hybrid protein encodedAlkaline phosphatase activitya
DsbA(1–136)-PhoA DsbA(1–160)-PhoA
Plasmid typepBR322 derivative 566 695pHSG575 derivative 216 201pTrc99A derivative (uninduced) 205 249pTrc99A derivative induced for:
1 h 486 6563 h 686 9935 h 1,029 1,242
a Values are units per milligram of total bacterial protein.
VOL. 179, 1997 DsbA9-PhoA HYBRID PROTEIN FOLDING AND EXPORT 3263
on May 26, 2018 by guest
http://jb.asm.org/
Dow
nloaded from
the shock fluid (Fig. 4D), the amount of DsbA(1–160)-PhoA wasca. three times higher after induction (compare lanes 1 and2) and some degradation fragments appeared (while it wasstable when not overproduced). In contrast, the amount ofDsbA(1–136)-PhoA was not elevated. In the insoluble fraction,both hybrid proteins were largely accumulated in response toinduction (Fig. 4C). Such results are puzzling, since 1 h afterinduction (and also later) the level of total alkaline phospha-tase activity in intact bacteria increased, irrespective of thehybrid protein expressed (Table 2).
It was then questionable, particularly in the case ofDsbA(1–136)-PhoA, whether the PhoA activity in inducedwhole cells originated only from free hybrid proteins (or theirpartly proteolyzed fragments) lying in the periplasmic space.Bacteria of strain SBS2027 carrying pAG2884 or pAG2885,which were collected from induced and noninduced culturesunder the conditions described above, were converted tospheroplasts by using lyzozyme-EDTA. Alkaline phosphatasein whole cells, in medium, in the supernatant of the sphero-plasts, and in the pelleted spheroplasts resuspended in Tris-sucrose buffer (Table 3) was measured. Under uninduced con-ditions, most of the activity lay in the supernatant (periplasmicspace) and a smaller part remained associated with the sphero-plasts. However, the latter was more important in the caseof DsbA(1–136)-PhoA (37%) than in the case of DsbA(1–160)-PhoA (7%). The induction of DsbA(1–136)-PhoA caused a six-fold elevation of the spheroplast-bound PhoA activity, butthere was no change in the periplasmic activity. By contrast,with DsbA(1–160)-PhoA, 63% of the activity was still recoveredin the periplasmic space under induced conditions. The releaseof the periplasmic content was complete in all cases, as mea-sured by the activity of other periplasmic enzymes (not shown).The possibility that hybrid proteins accumulated in the cyto-plasm might acquire PhoA activity during spheroplast forma-tion, as described by others (17), was eliminated, since blockingof the free SH groups by iodoacetamide (1 mM) in the col-lected bacteria and during spheroplast formation did not
change the present results (not shown). The data shown in Fig.4D and of Table 3 indicate that although overproduction ofDsbA(1–136)-PhoA led to an elevation of alkaline phosphataseactivity in whole cells of 2.5 times, this was not followed by itsrelease into the periplasmic space. When the pelleted sphero-plasts were blasted by a low osmotic pressure, the associatedalkaline phosphatase activity was entirely recovered in thehigh-speed supernatant. Therefore, some PhoA activity in in-tact bacteria was either associated with a weakly bound mem-brane pool, or it originally resided in the cytoplasm. However,the last possibility is less likely, because of the absence ofdisulfide bond-forming activity in this compartment.
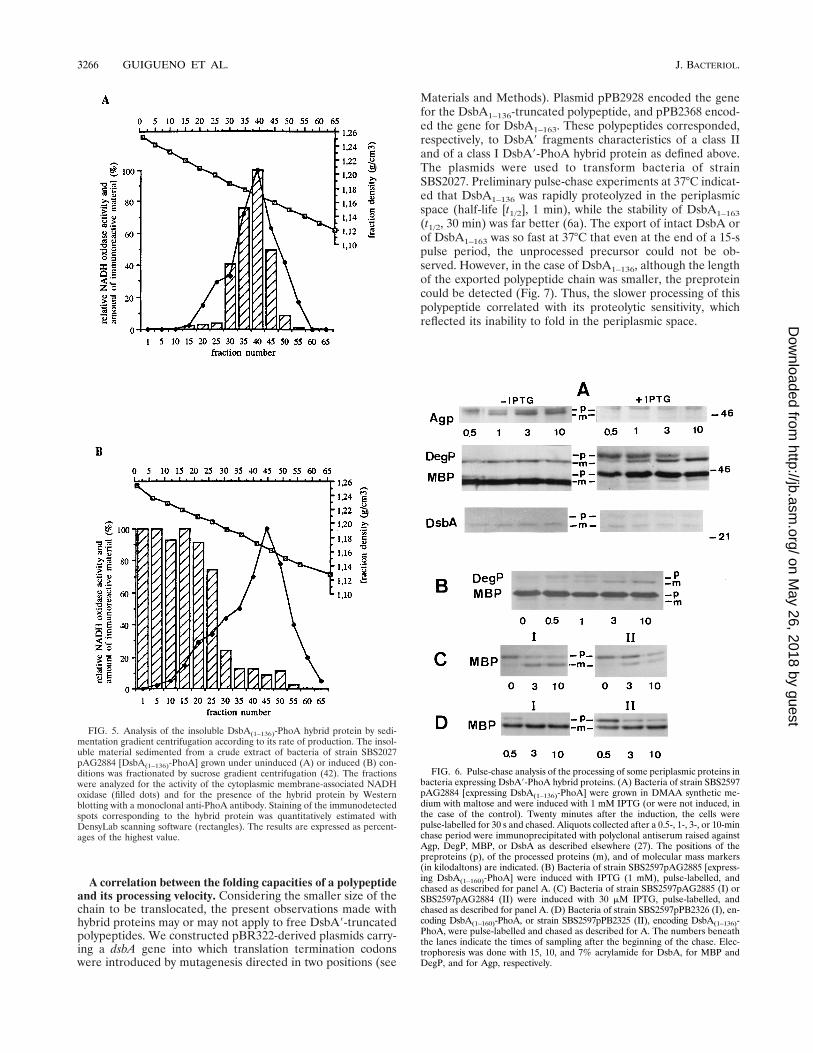
Sedimentation gradient analysis of the insoluble DsbA*(1–136)-PhoA hybrid protein according to the rate of its synthesis.Bacteria of SBS2027pAG2884 grown and collected under theconditions described above were broken by sonic oscillation,and their insoluble material was first sedimented and was frac-tionated by sucrose gradient centrifugation according to themethod described by Osborn (46). The fractions of the gradi-ent were submitted to Western blot analysis (Fig. 5). In unin-duced bacteria, all of the hybrid protein cosedimented with thecytoplasmic membrane enzymatic marker (Fig. 5A), and itsamino-terminal sequence was that of the preprotein. In theextract from the induced culture, most of the hybrid proteinhad a higher density (Fig. 5B). The amino-terminal sequenceof this dense material also corresponded to that of the DsbAleader peptide. The amount cosedimenting with the cytoplas-mic membrane marker in this case represented ca. 10% of thetotal. This is comparable to the amount present under unin-duced conditions (8% of that under induced conditions, asdeduced from the ratio of densitometric trace recordings inlanes 3 and 4 of Fig. 4C), which entirely cosedimented with thecytoplasmic membrane (Fig. 5A). None of the fractions of thegradient contained appreciable alkaline phosphatase activity.Thus, the PhoA-active nonperiplasmic form could not be re-covered by this procedure.
Since the denser material could correspond to inclusion bod-
FIG. 3. Lethal effect of DsbA9-PhoA hybrid protein overproduction. Bacteria carrying plasmid pAG2884 (A) or pAG2885 (B) encoding, respectively,DsbA(1–136)-PhoA and DsbA(1–160)-PhoA under the control of the Ptrc promoter were grown at 30°C in LB (ampicillin) medium and induced with 1 mM IPTG at timezero (open symbols) or were not induced (closed symbols). Viability was measured by plating on LB (ampicillin) medium without IPTG. The recipient strains forplasmids were SBS2597 (triangles and lozenges) and its degP isogenic derivative SBS2604 (circles and squares).
3264 GUIGUENO ET AL. J. BACTERIOL.
on May 26, 2018 by guest
http://jb.asm.org/
Dow
nloaded from
ies in the cytoplasm, we tested the possibility that a concomi-tant overproduction of cytoplasmic chaperones protected bac-teria against the lethal effect of DsbA(1–136)-PhoA. Multicopycompatible plasmids carrying either dnaK-dnaJ (pAPKJ-145)or groES-groEL (pEMR602) were first introduced into bacteriaof strain SBS2604 (degP), and the transformants were used asrecipients for the high-copy-number plasmid pPB2325, whichwas already shown not to be tolerated in this strain (see above).However, transformants still could not be obtained. Althoughwe found that the export of intact DsbA was not affected insecB strains (data not shown), we also tested whether SecBoverproduction (pEMR675) could nevertheless afford someprotection; however, this remained without effect.
Therefore, under permissive conditions, the insoluble hybridprotein was associated with the cytoplasmic membrane, whileunder nonpermissive conditions, most of it formed aggregates
of higher density without PhoA activity which could be inclu-sion bodies.
DsbA(1–136)-PhoA (class II) inhibits the export machinerymore efficiently than DsbA(1–160)-PhoA (class I). To knowwhether the effects provoked by the overexpression of thehybrid proteins could be related to a handicap of the globalexport system, the processing of four periplasmic proteins,namely, MalE (maltose-binding protein [MBP]), DegP, DsbA,and Agp (glucose-1-phosphatase), was examined by pulse-chase experiments (Fig. 6) using bacteria of SBS2597 (degP1)carrying the pTrc99A-derived plasmids pAG2884 (class II) andpAG2885 (class I). Under uninduced (permissive) conditionswith pAG2884, the four proteins were normally processed.Twenty minutes after full induction by IPTG, although thecells were still viable (Fig. 4), there was a strong inhibitionof MBP, Agp, DsbA, and DegP processing (Fig. 6A). WithpAG2885, after full induction, the processing was affected inthe same way as that with pAG2884 (Fig. 6B). Reducing theconcentration of IPTG to 30 mM allowed permanent main-tainence of 100% viability in bacteria of both classes. Undersuch permissive conditions, processing in bacteria expressingthe class II hybrid protein remained significantly slower than inthose expressing the class I hybrid protein, as shown for in-stance in the case of MBP (Fig. 6C). A comparable situationwas found when the hybrid proteins originated from the genefusions transcribed under the control of the weak dsbA P1promoter carried by the high-copy-number plasmids pPB2325and pPB2326 (Fig. 6D). Similar results have also been ob-served for other class II versus class I hybrid proteins (notshown).
Such a block in the overall export process could also affectthe transport of b-lactamase to the periplasm and simply ex-plain the death of bacteria, since plasmids were maintained byampicillin selection. However, when the bla gene on the fusionplasmids pAG2884 and pAG2885 was interrupted by anV::cam interposon, the modified plasmids were still lethal tobacteria after induction with IPTG in the presence of chlor-amphenicol (data not shown).
This indicates (i) that the processing or some other step(s) ofthe export of periplasmic proteins was reduced or blocked byexpression of the fusions, (ii) that the damage to the exportsystem increased with increasing amounts of hybrid protein sofar produced, and (iii) that plasmids encoding class II hybridproteins were more damaging than those corresponding toclass I.
FIG. 4. Western blot analysis of DsbA-PhoA distribution among the peri-plasmic and insoluble fractions. (A and B) Bacteria of strains SBS2027 (degP1)and SBS2603 (degP) carrying the various fusion plasmids were grown to mid-exponential phase at 30°C in LB (ampicillin) medium. The shock fluid (A) andthe insoluble fraction, sedimenting at 50,000 3 g (B), were prepared as describedin Materials and Methods and submitted to Western blot analysis with an anti-PhoA monoclonal antibody (the insoluble fraction was concentrated three timesrelative to the shock fluid). Lanes: 1, SBS2027pAG2784 [low-copy-number plas-mid encoding DsbA(1–160)-PhoA]; 2, SBS2027pAG2753 [low-copy-number plas-mid encoding DsbA(1–136)-PhoA]; 3, SBS2603pAG2784 (degP); 4, SBS2603pAG2753 (degP); 5, SBS2027pPB2326 [pBR322-derived plasmid encodingDsbA(1–160)-PhoA]; 6, SBS2027pPB2325 [pBR322-derived plasmid encodingDsbA(1–136)-PhoA]. (C and D) The insoluble and shock fluid fractions, respec-tively, prepared from bacteria of SBS2027 carrying either pAG2885 [DsbA(1–160)-PhoA; lanes 1 and 2] or pAG2884 [DsbA(1–136)-PhoA; lanes 3 and 4]. Bacteriawere collected from induced (with IPTG) or uninduced (without IPTG) cultures,and the extracts were submitted to Western blot analysis with monoclonal anti-PhoA antibody. Other experimental details are indicated in the text.
TABLE 3. Alkaline phosphatase activity in bacterial andspheroplast fractions of SBS2027pAG2884
and SBS2027pAG2885
Hybrid proteinexpresseda
Value forDsbA(1–136)-PhoAb
Value forDsbA(1–160)-PhoAb
2 IPTG 1 IPTGd 2 IPTG 1 IPTGd
Whole bacteria 44.8 112.7 53.5 161.7Medium 1.5 3.9 1.3 3Supernatant of
spheroplasts27.9 (59.4) 23.9 (18.7) 48.3 (90.3) 102.1 (63.4)
Spheroplasts 17.6 (37.4) 100.1 (78.2) 3.9 (7.3) 55.9 (34.7)Totalc 47 127.9 53.5 161
a Bacteria were collected 1 h after induction.b Results are expressed in units per milliliter of culture. Values in parentheses
are percentages of the total (last row).c Total value for combination of medium, supernatant of spheroplasts, and
spheroplasts.d IPTG was 1 mM (final concentration).
VOL. 179, 1997 DsbA9-PhoA HYBRID PROTEIN FOLDING AND EXPORT 3265
on May 26, 2018 by guest
http://jb.asm.org/
Dow
nloaded from
A correlation between the folding capacities of a polypeptideand its processing velocity. Considering the smaller size of thechain to be translocated, the present observations made withhybrid proteins may or may not apply to free DsbA9-truncatedpolypeptides. We constructed pBR322-derived plasmids carry-ing a dsbA gene into which translation termination codonswere introduced by mutagenesis directed in two positions (see
Materials and Methods). Plasmid pPB2928 encoded the genefor the DsbA1–136-truncated polypeptide, and pPB2368 encod-ed the gene for DsbA1–163. These polypeptides corresponded,respectively, to DsbA9 fragments characteristics of a class IIand of a class I DsbA9-PhoA hybrid protein as defined above.The plasmids were used to transform bacteria of strainSBS2027. Preliminary pulse-chase experiments at 37°C indicat-ed that DsbA1–136 was rapidly proteolyzed in the periplasmicspace (half-life [t1/2], 1 min), while the stability of DsbA1–163
(t1/2, 30 min) was far better (6a). The export of intact DsbA orof DsbA1–163 was so fast at 37°C that even at the end of a 15-spulse period, the unprocessed precursor could not be ob-served. However, in the case of DsbA1–136, although the lengthof the exported polypeptide chain was smaller, the preproteincould be detected (Fig. 7). Thus, the slower processing of thispolypeptide correlated with its proteolytic sensitivity, whichreflected its inability to fold in the periplasmic space.
FIG. 5. Analysis of the insoluble DsbA(1–136)-PhoA hybrid protein by sedi-mentation gradient centrifugation according to its rate of production. The insol-uble material sedimented from a crude extract of bacteria of strain SBS2027pAG2884 [DsbA(1–136)-PhoA] grown under uninduced (A) or induced (B) con-ditions was fractionated by sucrose gradient centrifugation (42). The fractionswere analyzed for the activity of the cytoplasmic membrane-associated NADHoxidase (filled dots) and for the presence of the hybrid protein by Westernblotting with a monoclonal anti-PhoA antibody. Staining of the immunodetectedspots corresponding to the hybrid protein was quantitatively estimated withDensyLab scanning software (rectangles). The results are expressed as percent-ages of the highest value.
FIG. 6. Pulse-chase analysis of the processing of some periplasmic proteins inbacteria expressing DsbA9-PhoA hybrid proteins. (A) Bacteria of strain SBS2597pAG2884 [expressing DsbA(1–136)-PhoA] were grown in DMAA synthetic me-dium with maltose and were induced with 1 mM IPTG (or were not induced, inthe case of the control). Twenty minutes after the induction, the cells werepulse-labelled for 30 s and chased. Aliquots collected after a 0.5-, 1-, 3-, or 10-minchase period were immunoprecipitated with polyclonal antiserum raised againstAgp, DegP, MBP, or DsbA as described elsewhere (27). The positions of thepreproteins (p), of the processed proteins (m), and of molecular mass markers(in kilodaltons) are indicated. (B) Bacteria of strain SBS2597pAG2885 [express-ing DsbA(1–160)-PhoA] were induced with IPTG (1 mM), pulse-labelled, andchased as described for panel A. (C) Bacteria of strain SBS2597pAG2885 (I) orSBS2597pAG2884 (II) were induced with 30 mM IPTG, pulse-labelled, andchased as described for panel A. (D) Bacteria of strain SBS2597pPB2326 (I), en-coding DsbA(1–160)-PhoA, or strain SBS2597pPB2325 (II), encoding DsbA(1–136)-PhoA, were pulse-labelled and chased as described for A. The numbers beneaththe lanes indicate the times of sampling after the beginning of the chase. Elec-trophoresis was done with 15, 10, and 7% acrylamide for DsbA, for MBP andDegP, and for Agp, respectively.
3266 GUIGUENO ET AL. J. BACTERIOL.
on May 26, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

DISCUSSION
The efficient production of a protein in the periplasmicspace of E. coli, in addition to a functional Sec machinery,requires some intrinsic constraints. These are essentially anadequate signal peptide in the preprotein (24) and a primarystructure allowing both its transfer across the membrane andits folding into a protease-insensitive compact form in theperiplasmic environment. This study is concerned with thepossible influence of the folding capacity of a protein in theperiplasmic space on the efficiency of its export to this com-partment. The model chosen consists in a set of DsbA9-PhoAhybrid proteins originating from plasmid-borne TnphoA inser-tions distributed over the whole length of the dsbA gene. All ofthe transcribed hybrid proteins carry the same efficient DsbAleader peptide and the same small linker region and PhoAprotein specified by TnphoA. They all display alkaline phos-phatase activity once disulfide bonds are formed in the peri-plasmic space by DsbA of chromosomal origin. They differ bythe lengths of their amino-terminal DsbA9 regions and by theability or inability of this only part of the hybrid protein to foldinto a proteolysis-resistant configuration in the periplasmicspace. The folding of this region requires the presence of a keydomain located between amino acid residues 136 and 160 ofDsbA (8). This defines two classes: that of the DsbA9 foldablehybrid proteins (class I) and that of the unfoldable ones (classII). The properties, here reported, of each of the two repre-sentative hybrid proteins DsbA(1–136)-PhoA (class II) andDsbA(1–160)-PhoA (class I), which are most closely related bytheir sizes, have been verified to extend to other members ofthe corresponding class.
The present observations show that although their two con-stituting parts are proteins or protein fragments of periplasmicorigin, DsbA9-PhoA hybrid proteins can less efficiently betransferred to the periplasmic space than each of the twooriginal whole proteins in a wild-type strain. Their productionabove a given threshold leads to cell death, and this toxicitycorrelates with their accumulation in the insoluble acellularfraction (mostly as inclusion bodies) and with a collapse of theexport capacity for other periplasmic proteins. Such abnormal-ities are more readily provoked by an unfoldable hybrid pro-tein than by a foldable one. At a low rate of synthesis, a classII hybrid protein is more retained in the cytoplasmic mem-brane as a precursor protein than a member of class I (Fig. 4C,lanes 1 and 3 and Fig. 5A). Increasing its rate of synthesis alsomore dramatically affects the export of other periplasmic pro-teins (such as MBP, for instance) than does a class I hybridprotein (Fig. 6). Meanwhile, its insoluble pool, but not itsperiplasmic soluble pool, is elevated (Fig. 4D). It is also more
toxic than a class I representative in terms of killing efficiency.This is consistent with a reduced export ability of class II hybridproteins compared to that of class I and with a jamming of theexport machinery more efficiently provoked by expression ofthe hybrid proteins which carry an unfoldable DsbA9 amino-terminal part. The destruction of the DegP periplasmic pro-teolytic activity stimulates the accumulation of hybrid proteinin the insoluble fraction and lowers the threshold of the toxicrate of synthesis. Once more, this shows the existence of arelationship between the persistence in the periplasmic spaceof an unfolded region of the molecule and an inhibition of theglobal export phenomenon. Whether this reflects an indirecteffect or simply the lack of clearance of the export apparatus ata final stage remains to be elucidated.
Mutations in dsbA or dsbC (41), which are presumed toprovoke an elevation of the concentration of unfolded proteinsin the periplasmic space, have been shown to stimulate thesynthesis of the alternate transcription initiation factor sE (50).The sE-dependent stress-inducible regulon is also stimulatedby the overproduction or the misfolding of some outer mem-brane proteins or by their unbalanced production (38, 52). More-over, mutations in surA or in fkpA, which encode periplasmicpeptidyl prolyl isomerases, also elevate the level of transcrip-tion of sE and that of DegP, a member of the sE regulon (34,42). Transcription of degP is also stimulated by the overpro-duction of a lipoprotein, NlpE, through the CpxR regulatorand its cognate cytoplasmic membrane sensor CpxA (13). It ispossible that one or more components involved in the exportmechanism are encoded by genes belonging either to the sE
regulon or to the Cpx regulon. However, such regulations havenot yet been described for the Sec proteins. Whether class IIrather than class I hybrid proteins can stimulate the sE regulonor the transcription of degP in response to a CpxA-CpxR signaltransfer is presently under investigation. The presence of anunfolded protein in the periplasmic space could also perhapsreduce the transcription of some gene necessary for export bya different, still unknown mechanism.
The accumulation of hybrid proteins in the insoluble frac-tion and the general inhibition of the export were observedwithin the first hour following an induction of their transcrip-tion from a Ptrc promoter (Fig. 4 and 6). This can hardly beaccounted for by the progressive dilution of a protein involvedin export in response to an arrest of its synthesis, unless thisprotein has a rapid turnover. However, apart from a fast deg-radation of overexpressed and uncomplexed SecY (57), littleinformation about the turnover of such proteins is available.The relative export handicap of class II hybrid proteins and itsconsequences (export jamming and toxicity) could also simplyresult from a more direct interaction with one of the proteinsinvolved in exportation.
First, a peculiar structure of the prehybrid protein mightprevent the polypeptide from adopting in the cytoplasm theconformation required for the translocation by the Sec ma-chinery (3). Although other proteins could be involved, over-synthesis of the heat shock cytoplasmic chaperones GroES-GroEL or DnaJ-DnaK does not restore the viability of degPbacteria carrying pBR322-derived plasmids encoding class IIhybrid proteins. However, such chaperones in excess areknown to prevent the accumulation of unprocessed forms ofperiplasmic or outer membrane proteins under different ex-port-defective conditions (9, 32, 47, 60, 61) and can also sub-stitute for SecB (1).
Second, the hybrid proteins might perhaps inhibit translo-cation itself. The polypeptide under translocation adopts, atleast transiently and locally, a linear conformation (3). This isimposed by the stepwise outward progression of the chain
FIG. 7. Comparison of processing between an unstable and a more-stableDsbA truncated polypeptide. Bacteria of strain SBS2451 (dsbA) carrying pPB2928(encoding DsbA(1–136)) or pPB2368 (encoding DsbA(1–163)) were either pretreat-ed with 3 mM sodium azide (45) 5 min before pulse-labelling (15 s) to allowaccumulation of the preprotein (az) or were pulse-labelled for the same timewithout pretreatment (15 s). p, preprotein; m, mature protein.
VOL. 179, 1997 DsbA9-PhoA HYBRID PROTEIN FOLDING AND EXPORT 3267
on May 26, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

resulting from its successive ATP-driven cycles of interactionwith SecA (18, 20) and probably also by the configuration ofthe SecY, -E, and -G proteins acting as a drawplate. Thisexposes each portion of the chain to the close vicinity of theSec proteins. Because the entire primary sequence of any ofthe class II hybrid proteins is also contained within that of theshortest member of class I, the greater export defect of class IIhybrid proteins cannot be explained by such undesirable inter-actions.
Third, they could behave differently at a late step or ata posttranslocational step. Whether the cleavage of the signalpeptide takes place before or after translocation (58) isachieved may vary according to translocation velocity, differ-ences in the leader peptidase recognition site (21), and possiblyother intrinsic or extrinsic parameters. If the first mechanism islikely to prevail in the case of large proteins, it is questionablewhether the ability of the polypeptide to fold rapidly in theperiplasm can stimulate the last steps of the export. Alterna-tively, its lack of folding very likely results in the exposition ofnormally internal hydrophobic regions. Such regions mightdirectly interact with cytoplasmic membrane components andinhibit the release of the extruded chains. The exact functionsof SecD (22) and SecF are still unclear. These two proteinsstimulate the export of MBP, ribose-binding protein (RBP),and OmpA, and their absence causes a cold-sensitive pheno-type (48). While not necessary for the ATP-dependent trans-location, they are required for the DmH-dependent transloca-tion of MBP (4). Matsuyama et al. (37) suggested that SecDcould be involved in the coupled folding-and-release of pro-cessed proteins. It is thus possible that the inability for foldingof the DsbA9 domain in class II hybrid proteins inhibits thereleasing activity of SecD. As a matter of fact, a fraction of theinsoluble DsbA9-PhoA hybrid protein pool remains attachedto the spheroplasts and possesses alkaline phosphatase activity(Table 3). This means that such molecules very likely lie on theperiplasmic side of the cytoplasmic membrane. This fraction ismuch more important in the case of a DsbA9-folding-reluctanthybrid protein. However, such a retention in the membranecould also directly result from an inhibition of the Lep pepti-dase activity if a coupling between folding and processing doesexist. The inactivation of DegP protease activity enhances thedamage provoked by the expression of a DsbA9-PhoA hybridprotein, while it increases the pool of insoluble precursor, atleast under permissive conditions (Fig. 4). If the processing ofclass II hybrid proteins is more specifically inhibited, then in awild-type strain, the cleavage of the unfolded amino-terminalDsbA9 region of the chain under translocation by DegP mighthelp restore a situation similar to that which normally followsthe processing by the Lep peptidase. Our results exclude pro-tection by a possible chaperone activity of DegP.
Taken together, the present data suggest that the rapid fold-ing of a protein in the periplasmic space, or more likely at theperiplasmic side of the cytoplasmic membrane, exerts a directsynergistic effect on its own export by activation of a late stepof this export. Whether the activities of SecD or of the Leppeptidase are indeed inhibited by persistent unfolded regionsin a protein just translocated remains to be investigated.
ACKNOWLEDGMENTS
We thank J. Beckwith and J. Pogliano for their gift of anti-(MBP/DegP) antiserum and O. Fayet for kindly providing us with groES-groEL dnaJ-dnaK plasmids and secB-carrying plasmids. We thank C.Kumamoto for sending a secB::Tn5 strain.
The attribution by the C.E.A. of a C.F.R. fellowship to A. Guiguenois acknowledged.
REFERENCES
1. Altman, E., C. A. Kumamoto, and S. Emr. 1991. Heat shock proteins cansubstitute for SecB function during protein export in Escherichia coli. EMBOJ. 10:239–245.
2. Amann, E., B. Ochs, and K.-J. Abel. 1988. Tightly regulated tac promotervectors useful for the expression of unfused and fused proteins in Escherichiacoli. Gene 69:301–315.
3. Arkowitz, R. A., J. C. Joly, and W. Wickner. 1993. Translocation can drive theunfolding of a preprotein domain. EMBO J. 12:243–253.
4. Arkowitz, R. A., and W. Wickner. 1994. SecD and SecF are required for theproton electochemical gradient stimulation of preprotein translocation.EMBO J. 13:954–963.
5. Bardwell, J. C. A., K. McGovern, and J. Beckwith. 1991. Identification of aprotein required for disulfide bond formation in vivo. Cell 67:581–589.
6. Bardwell, J. C. A., J. O. Lee, G. Jander, N. Martin, D. Belin, and J. Beckwith.1993. A pathway for disulfide bond formation in vivo. Proc. Natl. Acad. Sci.USA 90:1038–1042.
6a.Belin, P., A. Guigueno, and P. L. Boquet. Unpublished data.7. Belin, P., and P. L. Boquet. 1994. The Escherichia coli dsbA gene is partly
transcribed from the promoter of a weakly expressed upstream gene. Mi-crobiology 140:3337–3348.
8. Belin, P., E. Quemeneur, and P. L. Boquet. 1994. A pleıotropic acid phos-phatase-deficient mutant of Escherichia coli shows premature termination inthe dsbA gene. Use of dsbA::phoA fusions to localize a structurally importantdomain in DsbA. Mol. Gen. Genet. 242:23–32.
9. Berges, H., E. Joseph-Liauzun, and O. Fayet. 1996. Combined effects of thesignal sequence and the major chaperone proteins on the export of humancytokines in Escherichia coli. Appl. Environ. Microbiol. 62:55–60.
10. Cook, R. A. 1988. Periplasmic proteases of Escherichia coli. Crit. Rev. Bio-technol. 8:159–175.
11. Cosma, C. L., and T. J. Silhavy. 1996. Overproduction of DegP proteasesuppresses the toxicity conferred by a processing-defective porin in a prote-olysis-independent fashion, p. 63. In Abstracts of papers presented at the1996 Meeting on Molecular Chaperones and the Heat-Shock Response.Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y.
12. Dalbey, R. E., and W. Wickner. 1985. Leader peptidase catalyzes the releaseof exported proteins from the outer surface of the Escherichia coli plasmamembrane. J. Biol. Chem. 260:15925–15931.
13. Danese, P. N., W. B. Snyder, C. L. Cosma, L. J. B. Davis, and T. J. Silhavy.1995. The Cpx two-component signal transduction pathway of Escherichiacoli regulates transcription of the gene specifying the stress-inducibleperiplasmic protease DegP. Genes Dev. 9:387–398.
14. Date, T., and W. Wickner. 1981. Isolation of the Escherichia coli leaderpeptidase gene and effects of leader peptidase overproduction in vivo. Proc.Natl. Acad. Sci. USA 78:6106–6110.
15. Davis, B. D., and E. S. Mingioli. 1950. Mutants of Escherichia coli requiringmethionine or vitamin B12. J. Bacteriol. 60:17–28.
16. Deng, W. P., and J. A. Nickoloff. 1992. Site-directed mutagenesis of virtuallyany plasmid by eliminating a unique site. Anal. Biochem. 200:81–88.
17. Derman, A. I., and J. Beckwith. 1995. Escherichia coli alkaline phosphataselocalized to the cytoplasm slowly acquire enzymatic activity in cells whosegrowth has been suspended: a caution for gene fusion studies. J. Bacteriol.177:3734–3770.
18. Douville, K., A. Price, J. Eichler, A. Economou, and W. Wickner. 1995.SecYEG and SecA are the stoichiometric components of preprotein trans-locase. J. Biol. Chem. 270:20106–20111.
19. Driessen, A. J. M. 1992. Precursor protein translocation by the Esche-richia coli translocase is directed by the proton motive force. EMBO J.11:847–853.
20. Economou, A., J. A. Pogliano, J. Beckwith, D. B. Oliver, and W. Wickner.1995. SecA membrane cycling at SecYEG is driven by distinct ATP bindingand hydrolysis events and is regulated by SecD and SecF. Cell 83:1171–1181.
21. Fikes, J. D., and J. Bassford, Jr. 1987. Export of unprocessed precursormaltose-binding protein to the periplasm of Escherichia coli. J. Bacteriol.169:2352–2359.
22. Gardel, C., S. Benson, J. Hunt, S. Michaelis, and J. Beckwith. 1987. SecD,a new gene involved in protein export in Escherichia coli. J. Bacteriol.169:1286–1290.
23. Guzman, L.-M., D. Belin, M. J. Carson, and J. Beckwith. 1995. Tight regu-lation, modulation, and high-level expression by vectors containing the arab-inose PBAD promoter. J. Bacteriol. 177:4121–4130.
24. Hikita, C., and S. Mizushima. 1992. Effects of total hydrophobicity andlength of the hydrophobic domain of a signal peptide on in vitro translocationefficiency. J. Biol. Chem. 267:4882–4888.
25. Ito, K. 1992. SecY and integral membrane components of the Escherichiacoli protein translocation system. Mol. Microbiol. 6:2423–2428.
26. Ito, K. 1996. The major pathways of protein translocation across membranes.Genes Cells 1:337–346.
27. Ito, K., P. J. Bassford, Jr., and J. Beckwith. 1981. Protein localization in E.coli: is there a common step in the secretion of periplasmic and outermembrane proteins? Cell 24:707–717.
28. Joly, J. C., and W. Wickner. 1993. The SecA and SecY subunits of translo-
3268 GUIGUENO ET AL. J. BACTERIOL.
on May 26, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

case are the nearest neighbors of a translocating preprotein, shielding it fromphospholipids. EMBO J. 12:255–263.
29. Kamitani, S., Y. Akiyama, and K. Ito. 1992. Identification and characterisa-tion of an Escherichia coli gene required for the formation of correctly foldedalkaline phosphatase, a periplasmic enzyme. EMBO J. 11:57–62.
30. Keller, R. C. A., D. ten Berge, N. Nouwen, M. M. E. Snel, J. Tommassen, D.Marsh, and B. de Kruijff. 1996. Mode of insertion of the signal sequence ofa bacterial precursor protein into phospholipid bilayers as revealed by cys-teine-based site-directed spectroscopy. Biochemistry 35:3063–3071.
31. Kumamoto, C. A. 1989. Escherichia coli SecB protein associates with ex-ported protein precursors in vivo. Proc. Natl. Acad. Sci. USA 86:5320–5324.
32. Kusukawa, N., T. Yura, C. Ueguchi, Y. Akiyama, and K. Ito. 1989. Effects ofmutations in heat-shock genes groES and groEL on protein export in Esch-erichia coli. EMBO J. 8:3517–3521.
33. Laemmli, U. K. 1972. Cleavage of structural proteins during the assembly ofthe head of bacteriophage T4. Nature 227:680–685.
34. Lazar, S. W., and R. Kolter. 1996. SurA assits the folding of Escherichia coliouter membrane proteins. J. Bacteriol. 178:1770–1773.
35. Lipinska, B., M. Zylicz, and C. Georgopoulos. 1990. The HtrA (DegP)protein, essential for Escherichia coli survival at high temperatures, is anendopeptidase. J. Bacteriol. 172:1791–1797.
36. Maniatis, T., E. F. Fritsch, and J. Sambrook. 1982. Molecular cloning: alaboratory manual. Cold Spring Harbor Laboratory, Cold Spring Harbor,N.Y.
37. Matsuyama, S., Y. Fujita, and S. Mizushima. 1993. SecD is involved in therelease of translocated secretory proteins from the cytoplasmic membrane ofEscherichia coli. EMBO J. 12:265–270.
38. Mecsas, J., P. E. Rouviere, J. W. Erickson, T. J. Donohue, and C. A. Gross.1993. The activity of sE, an Escherichia coli heat-inducible s factor, ismodulated by expression of outer membrane proteins. Genes Dev. 7:2618–2628.
39. Miller, J. H. 1972. Experiments in molecular genetics. Cold Spring HarborLaboratory, Cold Spring Harbor, N.Y.
40. Missiakas, D., C. Georgopoulos, and S. Raina. 1993. Identification andcharacterization of the Escherichia coli gene dsbB, whose product is involvedin the formation of disulfide bonds in vivo. Proc. Natl. Acad. Sci. USA90:7084–7088.
41. Missiakas, D., C. Georgopoulos, and S. Raina. 1994. The Escherichia colidsbC (xprA) gene encodes a periplasmic protein involved in disulfide bondformation. EMBO J. 13:2013–2020.
42. Missiakas, D., J. M. Betton, and S. Raina. 1996. New components of proteinfolding in extracytoplasmic compartements of Escherichia coli SurA, FkpAand Skp/OmpH. Mol. Microbiol. 21:871–884.
43. Neu, H. C., and L. A. Heppel. 1965. The release of enzymes from Escherichiacoli by osmotic shock and during the formation of spheroplasts. J. Biol.Chem. 240:3685–3692.
44. Oliver, D. B. 1993. SecA protein: autoregulated ATPase catalysing prepro-tein insertion and translocation across the Escherichia coli inner membrane.Mol. Microbiol. 7:159–165.
45. Oliver, D. B., R. J. Cabelli, K. M. Dolan, and G. P. Jarosik. 1990. Azide-resistant mutants of Escherichia coli alter the SecA protein, an azide-sensi-
tive component of the protein export machinery. Proc. Natl. Acad. Sci. USA87:8227–8231.
46. Osborn, M. J., J. E. Gander, E. Parisi, and J. Carson. 1972. Mechanism ofassembly of the outer membrane of Salmonella typhimurium. J. Biol. Chem.247:3962–3972.
47. Phillips, G. J., and T. J. Silhavy. 1990. Heat-shock proteins DnaK andGroEL facilitate export of LacZ hybrid proteins in E. coli. Nature 344:882–884.
48. Pogliano, J., and J. Beckwith. 1994. SecD and SecF facilitate protein exportin Escherichia coli. EMBO J. 13:554–561.
49. Pugsley, A. P. 1993. The complete general secretory pathway in gram-neg-ative bacteria. Microbiol. Rev. 57:50–108.
50. Raina, S., D. Missiakas, and C. Geogopoulos. 1995. The rpoE gene encodingthe sE (s24) heat shock sigma factor of Escherichia coli. EMBO J. 14:1043–1055.
51. Randall, L. L., and S. J. S. Hardy. 1995. High selectivity with low specificity:how SecB has solved the paradox of chaperone binding. Trends Biochem.Sci. 20:65–69.
52. Rouviere, P. E., and A. De Las Penas. 1995. rpoE, the gene encoding thesecond heat shock sigma factor, sE, in Escherichia coli. EMBO J. 14:1032–1042.
53. Schatz, P. J., and J. Beckwith. 1990. Genetic analysis of protein export inEscherichia coli. Annu. Rev. Genet. 24:215–248.
54. Skorko-Glonek, J., A. Wawrzynow, K. Krewski, K. Kurpierz and B. Lipinska.1995. Site-directed mutagenesis of the HtrA (DegP) serine protease, whoseproteolytic activity is indispensable for Escherichia coli survival at elevatedtemperatures. Gene 163:47–52.
55. Strauch, K. L., and J. Beckwith. 1988. An Escherichia coli mutation prevent-ing degradation of abnormal periplasmic proteins. Proc. Natl. Acad. Sci.USA 85:1576–1580.
56. Takeshita, S., M. Sato, M. Toba, W. Masahashi, and T. Hashimoto-Gotoh.1987. High-copy-number and low-copy-number plasmid vectors for lacZa-complementation and chloramphenicol or kanamycin-resistance selection.Gene 61:63–74.
57. Taura, T., T. Baba, Y. Akiyama, and K. Ito. 1993. Determinants of thequantity of the stable SecY complex in the Escherichia coli cell. J. Bacteriol.175:7771–7775.
58. Uegushi, C., and K. Ito. 1990. Escherichia coli sec mutants accumulate aprocessed immature form of maltose-binding protein (MBP), a late-phaseintermediate in MBP export. J. Bacteriol. 172:5643–5649.
59. Wickner, W., M. L. Leonard, and A. Economou. 1995. On the road totranslocase. Cold Spring Harbor Symp. Quant. Biol. 60:285–290.
60. Wild, J., E. Altman, T. Yura, and C. A. Gross. 1992. DnaK and DnaJ heatshock proteins participate in protein export in Escherichia coli. Genes Dev.6:1165–1172.
61. Wild, J., P. Rossmeissl, W. A. Walter, and C. A. Gross. 1996. Involvement ofthe DnaK-DnaJ-GrpE chaperone team in protein secretion in Escherichiacoli. J. Bacteriol. 178:3608–3613.
62. Zwizinski, C., and W. Wickner. 1980. Purification and characterization ofleader (signal) peptidase from Escherichia coli. J. Biol. Chem. 255:7973–7977.
VOL. 179, 1997 DsbA9-PhoA HYBRID PROTEIN FOLDING AND EXPORT 3269
on May 26, 2018 by guest
http://jb.asm.org/
Dow
nloaded from