demonstration of extensive chromatin cleavage in transplanted
TRANSCRIPT

American journal of Pathology, Vol. 142, No. 3, March 1993Copyright C) Amenican Societyfor Intvestigative Pathology
Demonstration of Extensive Chromatin Cleavagein Transplanted Morris Hepatoma 7777 Tissue:Apoptosis or Necrosis?
Kazunori Fukuda,* Masamichi Kojiro,* andJen-Fu ChiutFrom the First Department of Pathology, * Kurume UniversitySchool ofMedicine, Kurume 830, Japan, and DepartmentofBiochemistry,t University of Vermont, College ofMedicine, Burlington, Vermont
CeU death may occur by either of two mecha-nisms: necrosis or apoptosis (programmed ceUldeath). In this paper, we demonstrate extensivechromatin cleavage into oligonucleosome-lengthfragments (DNA ladder) in transplanted Morrishepatoma 7777 tissue, which is suggestive ofthestimulation of an endogenous endonucleaseactivity previously found to be involved in theprocess ofapoptosis. The existence ofmany ap-optotic ceUs, which are morphologically charac-terized by condensed cytoplasm and basophilicnuclearfragments, were also seen in this tissue.In vivo and in vitro experiments were designedto further differentiate the morphological andbiochemicalfeatures of necrosis and apoptosisin liver and hepatoma cells. Liver tissue under-going ischemic necrosis showed a distinct DNAladder pattern without demonstrating the mor-phology of apoptosis, indicating that chromatincleavage into oligonucleosomal-length fragmentsis not confined to apoptotic ceU death, at least inliver cells. In in vitro-cultured McA-RH7777 cells,however, DNA ladder pattern was detected onlyin ceUs showing characteristic morphology ofapoptosis. From these two criteria (Le., charac-teristic morphology and DNA ladder), it wasstrongly suggested that the apoptotic process ishighly activated in the transplanted 7777 tissue.Based on the results obtained from in vitroexperiments, it was suggested that tumor apop-tosis may represent a residual attempt at auto-regulation within the expanding tumor popula-tion and/or may resultfrom mild cellular injuriessuch as hypoxia, nutrient deficiency, or otherunknown noxiousfactor(s). We also showed evi-dence that apoptosis is inducible in hepatoma
ceUs in vitro by a wide range of mild injuries orstimuli. (AmJ Pathol 1993, 142:935-946)
It has been widely accepted that there is a markeddiscrepancy between the rate of enlargement ofmany malignant tumors and the rate of proliferationof their component cells. This was attributed to spon-taneous cell death.1 There are two major distinctmodes of death in eukaryotic cells: necrosis andapoptosis.23 These two processes differ both mor-phologically and biochemically. Necrosis, which usu-ally involves groups of contiguous cells, is an uncon-trolled destructive phenomenon induced by a varietyof non-physiological agents (i.e., ischemia, toxicchemicals, infectious organelles, etc.) that disturbenergy-producing pathways and create an osmoticimbalance in the cell, leading to swelling of cyto-plasm and irreversible failure of cellular structures,caused by lysosomal enzymes.4- Apoptosis, on theother hand, is the term for programmed cell death,which plays an important role in controlled deletion ofcells during metamorphosis, differentiation, hor-mone-dependent atrophy, and normal cell turnoverand is regulated by signal transduction-coupledevents.23 Recent accumulating evidence hasexpanded the definition of apoptosis, and it is nowgenerally accepted that apoptosis is not alwaysphysiological.78 Many kinds of injurious agents areshown to be able to induce apoptosis in many kindsof cell lineages.9` The mechanisms that permitcells to die in this stereotyped fashion are now themajor concern in this field.
Apoptosis is characterized morphologically bycell shrinkage and hyperchromatic nuclear frag-ments3 and biochemically by chromatin cleavageinto nucleosomal oligomers.12-14 In this paper, we
Supported by U. S. Public Health Service grants CA42965 andCA25098 awarded by the National Cancer Institute.Accepted for publication August 26, 1992.Address reprint requests to Dr. Jen-Fu Chiu, Department of Bio-
chemistry, University of Vermont College of Medicine, Burlington, VT05405.
935

936 Fukuda et alAJP March 1993, Vol. 142, No. 3
demonstrate extensive chromatin cleavage into oli-gonucleosome-length DNA fragments in Morrishepatoma 7777 tissue transplanted in the thigh mus-cle of rats. Although it is generally accepted that thisDNA fragmentation, known as a "DNA ladder", is acharacteristic biochemical indicator for apopto-sis,12-14 the possibility that the DNA ladder mightalso be implicated in the process of hepatomanecrosis still has not been excluded. This possibilityseems to be further strengthened by the followingfacts. Earlier work has shown that a Ca2+-dependentendonuclease exists in nuclei of many types ofcells including liver cells.15'16 It is also well recog-nized that the process of coagulative necrosis, whichis a common feature of death in injured liver cells, isclosely associated with the accumulation of cytosolicCa2 , which may result in the activation ofvarious Ca2+-dependent enzymes including endo-nucleases.4 17 Stimulation of endogenous endonu-clease activity in liver cells is indeed shown in sev-eral types of cell injury.18 So the question is nowopen as to whether the DNA fragmentation intonucleosomal oligomers observed in 7777 tissue isdue to apoptosis or coagulative necrosis. The exper-iments described in this report were designed toanswer these questions and to elucidate the possi-ble mechanisms for cell death in 7777 hepatomatissue.
Materials and Methods
Tumor Cells and Cell Culture
Transplantable Morris hepatoma 7777 cells weregrown in the thigh muscle of Buffalo rats. The McA-RH7777 cell line, derived from the Morris hepatoma7777, was obtained from the American Type CultureCollection (Rockville, MD) and grown in Dulbecco'smodified Eagle's medium supplemented with 15%heat-inactivated fetal bovine serum and antibiotics(100 pg/ml of streptomycin and 100 units/ml of pen-icillin) at 37 C and 5% CO2. Cells were seeded at 1x 105/ml and cultured for 24 hours before treatment.The serum-free synthetic medium (SFM) used in theexperiments of serum starvation was RPMI 1640supplemented with 3 x 10-8 mol/L sodium seleniteand other trace elements according to the procedureof Nakabayashi et al.19 For experiments investigatingthe effect of calcium depletion on cell death in cul-ture cells, a number of agents were tested underboth calcium-sufficient and calcium-deficientconditions. For calcium-deficient conditions, cellswere cultured in Ca2+-free Dulbecco's modified
Eagle's medium containing 15% dialyzed fetalbovine serum (GIBCO Laboratories, Grand Island,NY).
In Vivo Induction of Liver Cell NecrosisMale Wistar rats weighing approximately 150 g (6weeks) were used for the following experiments thatinduce liver cell necrosis in vivo. The experimentswere performed according to the institution's guide-lines for the care and use of laboratory animals inresearch.
Ischemic Necrosis
Under ether anesthesia, the peritoneal cavity wasopened and blood vessels for the left lateral lobe ofthe liver were ligated. After closure of the peritonealcavity, the rats were maintained in a cage with feed-ing ad libitum. Treated rats were killed (0.5,1, 2, 4, 8,12, 24, and 48 hours after the treatment), and liversamples were removed.
Acute Carbon Tetrachloride (CC14) Hepatotoxicity
Rats were injected intraperitoneally with CCI4(1.25 ml/kg of body weight), mixed with an equalvolume of olive oil. Animals were killed at 6, 12, 24,36, 48, 72, and 96 hours after the injection, and livertissue samples were removed. Liver tissue samplesfor biochemical analysis were immediately frozenand stored at -80 C. Small slices were processed forlight microscopy after fixation with 10% buffered for-malin and embedding in paraffin. The degree of cel-lular injury was assessed histologically after hema-toxylin and eosin (H&E) staining.
In Vitro Induction of Cell Death inHepatoma CellsMonolayer cells were treated with the following inju-rious agents at various dosages to induce cell death;Ca2+ ionophore A23187, ethanol, ricin lectin, cyclo-heximide, and ultraviolet (UV) exposure. The specificbiological activity of A23187 (Sigma Chemical Co.,St. Louis, MO) is to create Ca2+ channels andthereby to overcome the permeability barrier repre-sented by the cellular membrane with the resultantinflux and intracytoplasmic accumulation of Ca2+that leads to cell death.20 A23187 was dissolved indimethylsulfoxide to make a 25-mmol/L stock solu-tion and stored at -20 C. Ricin lectin (EY Laborato-ries, Inc., San Mateo, CA) and cycloheximide (Sig-ma) have inhibitory effects on protein synthesis. ForUV exposure, cells were cultured in a tissue culture

Apoptosis of Rat Hepatoma Cells 937AJP MIarcb 1993, Vol. 142, No. 3
dish of 10 cm diameter (Falcon 3003, Becton-Dick-inson, Lincoln Park, NJ) with 9 ml of medium, and themonolayer culture cells were exposed to variousdoses of UV using the Stratalinker UV crosslinker(Stratagene, La Jolla, CA), which has a built-in detec-tor to ensure consistent UV energy for eachexperiment. Irradiated cultures were subsequentlyincubated under standard conditions.
DNA Isolation and Agarose GelElectrophoresisDNA was isolated according to the methoddescribed by Sambrook et al21 with minormodifications. In brief, tissue and cells were directlyhomogenized in DNA extraction buffer (10 mmol/LTris-HCI, pH 8.0, 100 mmol/L ethylenediaminetet-raacetic acid, 0.5% sodium dodecyl sulfate, 100pg/ml RNAse A) and incubated for 1 hour at 37 Cfollowed by incubation for 3 hours (for culture cells)or 16 hours (for tissue) at 50 C in the presence of 100pg/ml of proteinase K. The DNA was then extractedwith phenol/chloroform and precipitated in ethanol.Pellets were resuspended in T10E1 (10 mmol/L Tris-HCI, pH 8.0/1 mmol/L ethylenediaminetetraaceticacid), and the DNA concentration was determinedfrom the absorbance at 260 nm. Each DNA sample(20 pg) was electrophoresed through a 1.6% agar-ose gel containing 1 pg/ml of ethidium bromide. DNAbands were visualized with a UV transilluminator andphotographed.
Results
Morphological Signs of Cell Death byApoptosis in Transplanted MorrisHepatoma 7777 TissueScattered single cell death was frequently detectedin the 7777 tissue. Under light microscopy, thesedead cells showed the following histological featuresthat are characteristic of apoptosis (Figure 1A): 1)acidophilic cytoplasm and nuclear fragmentsappearing as discrete basophilic masses and 2) theabsence of inflammatory reaction. Several mitoticcells were also observed in the vicinity of apoptoticcells, indicating that stimulation of tumor growth andapoptosis were concurrent and that ischemia and/ornutritional deficiency are not the major cause for thescattered single cell death in the 7777 hepatomatissue. Apoptotic and mitotic cells were scored bycounting 10,000 cells in random fields of thehepatoma tissue. Apoptotic cells and mitotic cellsaccounted for 7.6% and 4.3% of the total cell num-
ber, respectively. Together with the frequent appear-ance of a cluster of apoptotic cells, the higher inci-dence of apoptotic cells than mitotic cells maysuggest the accumulation of dead cells in tumor tis-sue, probably due to immaturity of drainage vesselsand/or a lack of phagocytes. The tissue also con-tained regions showing cell death of contiguoustumor cells whose histological distribution is verysimilar to that of ischemic necrosis (Figure 1 B).Although apoptosis was originally characterized assingle cell death, the cells in these regions alsoshowed fragmented and condensed nuclei, whichare usually not observed in necrotic cells. Feulgenstaining clearly demonstrated condensed chromatinin fragmented nuclei of these dead cells in 7777tissue (Figure 1C). These findings strongly suggestthat the apoptotic process is implicated in the celldeath of both scattered and contiguous distributionin 7777 hepatoma tissue. Taking these contiguouscell deaths into account, cells undergoing apoptoticcell death were estimated to occupy around 20 to30% of total cell number in the 7777 hepatomatissue.
Demonstration of DNA Ladder in 7777Tissue
Agarose gel electrophoresis revealed a characteris-tic pattern of DNA fragments in transplanted 7777tissue. The DNA fragments consisted of multimers of180 to 190 base pairs, consistent with internucleo-somal cleavage of chromatin DNA by an endonu-clease (Figure 1 D, lane 2). DNA isolated from normalrat liver was detected only in the high-molecular-weight region (Figure 1D, lane 1).
DNA Fragmentation in Liver TissueInduced By Ischemic Necrosis or CC14HepatotoxicityAfter ligation of both the hepatic artery and the portalvein, the tissue of the ischemic lobe showed thehistological pattern of coagulative necrosis (Figure2A). Both cytoplasm and nuclei showed decreasedstainability with hematoxylin and eosin. Fragmenta-tion or condensation of the cytoplasm and nucleiwere not observed. DNA fragmentation was not sig-nificant in liver tissue taken at 30 minutes and at 1hour after ligation. The characteristic DNA ladderpattern was observed in the liver taken 2 hours afterthe ligation in which the tissue seemed to be at anearly stage of ischemic necrosis. At later time points,the steps in the ladder became smaller due to pro-gressive DNA hydrolysis. However, even at the latest

938 Fukuda et alAJP March 1993, Vol. 142, No. 3
M12
...U..... ,
Figure 1. Apoptotic features X'n transplanted Morris hepatoma 7777 tissue. A: Many scattered apoptotic cells (arrows) are present. These cells showacidophilic cytoplasm with condensed andfragmented nuclear materials. No inflammatory cell infiltration is seen. Several mitotic cells are observedin close vicinity to apoptotic cells (H&E, X 281). B: Regions showing cell death ofcontiguous tumor cells arefrequently observed in which histologicaldistribution is very similar to that of ischemic necrosis (H&E, X 70). C: Condensation andfragmentation of nuclear materials are clearly demon-strated in dead cells by Feulgen staining (Feulgen stain, x 562). D: Analysis ofDNAs isolatedfrom normal rat liver (lane 1) and transplanted Morrishepatoma 7777 tissue (lane 2: with 1.6% agarose gel electrophoresis. Note extensive DNAfragmentation of multi-nucleosomal units (DNA ladder) inlane 2, suggesting the process of apoptosis. Lane M contains DNA molecuilar weight markers.
Apoptosis of Rat Hepatoma Cells 939AJP March 1993, Vol. 142, No. 3
M 1 2 3 4 5 6 7 8 9
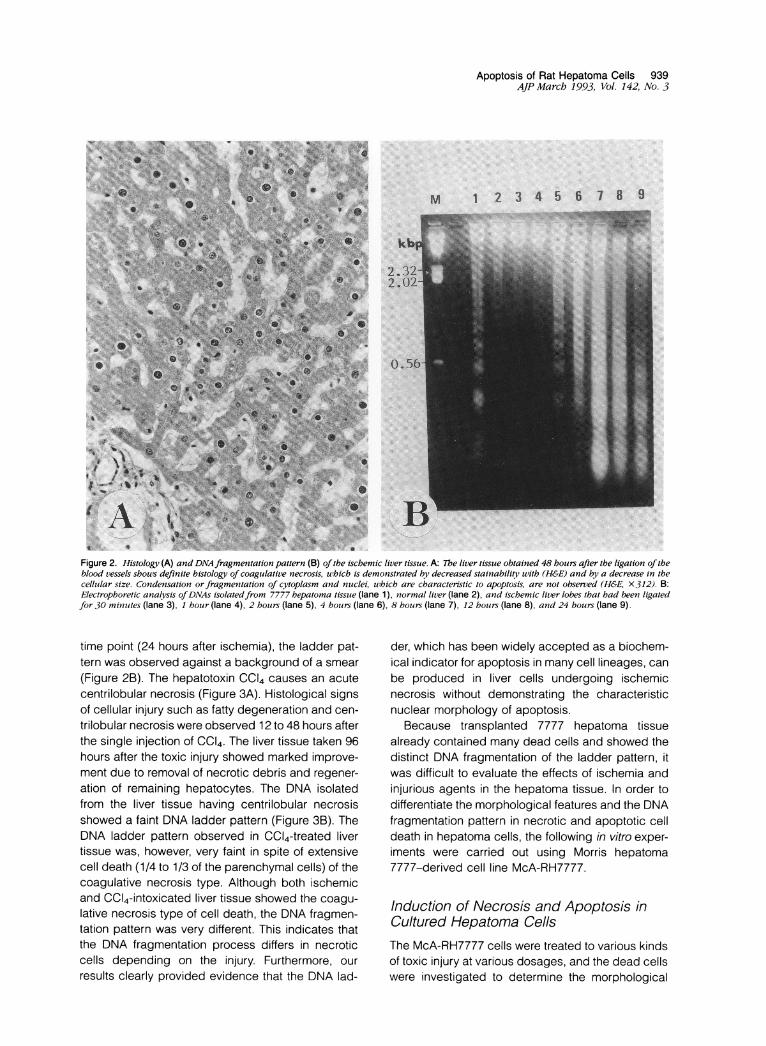
Figure 2. Histology (A) and DNA fragmentation pattern (B) ofthe ischemic liver tissue. A: Tbe liver tissue obtained 48 hours after the ligation oftheblood vessels shouws definite histology of coagulative necrosis, uhich is demonstrated by decreased stainability with (H&E) and by a decrease in thecelltular size. Condensation or fragmenitation of cytoplasm and ntuclei, which are characteristic to apoptosis, are not observed (H&E, X312). B:Electrophoretic analysis ofDNAs isolatedfrom 7777 bepatoma tissue (lane 1), normal liver (lane 2), and iscbemic liver lobes that had beetn ligatedfor 30 minutites (lane 3), 1 houir (lane 4), 2 hours (lane 5), 4 houirs (lane 6), 8 hours (lane 7), 12 hours (lane 8), and 24 hours (lane 9).
time point (24 hours after ischemia), the ladder pat-tern was observed against a background of a smear(Figure 2B). The hepatotoxin CC14 causes an acutecentrilobular necrosis (Figure 3A). Histological signsof cellular injury such as fatty degeneration and cen-trilobular necrosis were observed 12 to 48 hours afterthe single injection of CCI4. The liver tissue taken 96hours after the toxic injury showed marked improve-ment due to removal of necrotic debris and regener-ation of remaining hepatocytes. The DNA isolatedfrom the liver tissue having centrilobular necrosisshowed a faint DNA ladder pattern (Figure 3B). TheDNA ladder pattern observed in CC14-treated livertissue was, however, very faint in spite of extensivecell death (1/4 to 1/3 of the parenchymal cells) of thecoagulative necrosis type. Although both ischemicand CC14-intoxicated liver tissue showed the coagu-lative necrosis type of cell death, the DNA fragmen-tation pattern was very different. This indicates thatthe DNA fragmentation process differs in necroticcells depending on the injury. Furthermore, ourresults clearly provided evidence that the DNA lad-
der, which has been widely accepted as a biochem-ical indicator for apoptosis in many cell lineages, canbe produced in liver cells undergoing ischemicnecrosis without demonstrating the characteristicnuclear morphology of apoptosis.
Because transplanted 7777 hepatoma tissuealready contained many dead cells and showed thedistinct DNA fragmentation of the ladder pattern, itwas difficult to evaluate the effects of ischemia andinjurious agents in the hepatoma tissue. In order todifferentiate the morphological features and the DNAfragmentation pattern in necrotic and apoptotic celldeath in hepatoma cells, the following in vitro exper-iments were carried out using Morris hepatoma7777-derived cell line McA-RH7777.
Induction of Necrosis and Apoptosis inCultured Hepatoma CellsThe McA-RH7777 cells were treated to various kindsof toxic injury at various dosages, and the dead cellswere investigated to determine the morphological
O.
IA.0
..........

940 Fukuda et alAJP March 1993, Vol. 142, No. 3
1 2 3 4 5.. ...
bp-1353-10784872
-603
-310
i194
Figure 3. Histology (A) and DNA fragmentation pattern (B) of the liver tissue after a single dose of CC14 (1.25 mllkg of body weight). A: Histologvof the liver tissue taketn 24 hours after a sintgle dose of CC14, shou'ing centrilobular coaguilative necrosis andfatty degenzerationz (H&E, x 156). B:Electrophoretic analysis ofDNAs isolatedfrom the liver tissue taken 6 hoturs (lane 1), 12 hbours (lane 2), 24 hours (lane 3), 36 hozurs (lane 4), anid48 hourNs (lane 5) after a sinigle dose of CC14.
changes and the pattern of DNA fragmentation. Ap-optosis and necrosis were morphologically quitedifferent. Apoptotic cells were easily identifiable onH&E-stained samples by cell shrinkage with con-densed and fragmented nuclei. The criteria that gov-erned the identification of necrotic cells includedcytoplasmic swelling and loss of nuclear basophilia,eventually giving rise to karyolysis. Cells in the finalstages of necrosis underwent cell lysis, in direct con-trast to what takes place during apoptosis. Analysisof DNA from necrotic cells revealed no specificcleavage of the DNA, indicating random fragmenta-tion of the DNA.
From these two criteria, hepatoma cells wereshown to have two major pathways in the process ofcell death in spite of a variety of methods used toinitiate injurious action. One is direct cell death thatwas induced shortly after severe cellular damage.Another type is indirect cell death in which the cellsdie several hours after initiating injury. The latencyperiod from initiating injury to cell death variesaccording to the severity and type of injury. Directcell death showed findings characteristic of necro-
sis, and indirect cell death showed characteristicfeatures of apoptosis. For example, direct cell deathwas induced in cultured McA-RH7777 cells by a highdosage of ethanol (more than 10%), UV exposure(more than 3 x 103 mJ/cm2), or Ca2l ionophoreA23187 (more than 20 pmol/L). Figure 4 shows themorphology and the DNA fragmentation pattern indead cells induced by Ca2+ ionophore A23187.When the McA-RH7777 cells were treated withA23187 at concentrations of more than 20 pmol/L,almost all cells died within a few hours. High doses ofUV irradiation also induced necrotic cell death in thehepatoma cells: cells irradiated with 3 x 103 mJ/cm2of UV died within a few hours. These dead cells werecharacterized by swelling of the cytoplasm. Neithernuclear condensation nor fragmentation wasobserved (Figures 4, B and C, and 5A). A DNA lad-der was not demonstrated in this process (Figures4D and 5D, lane UV 3,000 mJ). On the other hand,mild cellular injuries such as low dosage of UV expo-sure (10 to 500 mJ), ricin lectin, and cycloheximideshowed indirect cell death in which cells died 12hours to 3 days after the initial exposure. The dead

Apoptosis of Rat Hepatoma Cells 9414/P March 1993, Vol. 142, No. 3
Caliwpinre A23187 ( jt )
bp
1353-1.07181-872-
60-
310-
Dcell
(J
Figure 4. Cell death in McA-RH7777 cells induced by the Ca2+ ionophore A23187. A: Phase-contrast photomicrograph of a control co/ftoire ofMcA-RH7777 in medioim conitainzinig 155% fetal bovine.senrm (X 140). B: Phase-conitrastphotomicrograph ofMcA-RH7777cells treated with 20,ymol/Lof Ca2+ ioniophore A23187Jbr 1 booir. Dead cells detachedfrom the soibstratoim show' swelling of the cells, which soggests cell death bhi necrosis(X 140). C: H&E5 histology) (of dcead cells obseried in (B) (X 562). D: Electrophoretic analysis ofDAAs isolatedfromn McA-RH7777 cells treated uithivarioos concentrations of Ca2+ ioioophore A23187 for 24 booirs. Cell viability, is expressed as the percentage of cells that excltoded trjpan bl/ne.
SO
C
AN,i
io" ;..'A .r
p.f: PI1-i z,-,':.:,w,07,
MrAwi_-s, W, If;!i,l
!;=FWfim"IF".Ts r, .A`
IIA I"' L -::
IF Mlllrwl(lW': .,.s
Am.

942 Fukuda et alAJP March 1993, Vol. 142, No. 3
...
..'I.,W",0~~~' q'P_ 4_i _ 4
..
c .0.Rwi:1
UV exposure ( m joules )0 1 5 10 2 50 IOD50X)1(O3ox
bp
1353.1078872
310
Figure 5. Morphologicalfeatures of UV-mediated cell death. A: Phase-contrast photomicrograph ofMcA-RH7777 cells treated with 3 x 10U3 mJ cm2of UVexposure. 7he photo was taken 3 hours after the exposure. Swelling ofcytoplasm in floating cells is obvious, indicating cell death by necrosis.B: Phase-contrast photomicrograph ofMcA-RH7777 cells treated with 50 mJ1cm2 of UV exposure. 7Te photo was taken 24 hours after the exposure.Shrinkage andfragmentation ofdead cells are obvious, indicating cell death by apoptosis. C: H&E stain of cells obtainedfrom (B). Many apoptoticcells (some arrowed), which contain condensed nuclearfragments, are observed. Irregular aggregation ofchromatin is also observed in many nuclei(H&E, x 431). D: Electrophoretic analysis ofDNAs isolatedfrom McA-RH7777 cells exposed with various dosages ofUV (O to 3 x 103 mJlcm2). 7Tecells were collected 24 hours after the exposure, andDNAs were isolated and analyzed with 1.6% agarosegel electrophoresis. DNA ladder was observedin cells treated with 10 to 500 mJ/cm2 of UV but not in cells treated with a high dose (3 x 103 mJlcm2) of UV
cells showed condensed and fragmented nuclei withcondensed cytoplasm, which fulfilled the morpho-logical criteria of apoptosis (Figure 5, B and C). Adistinct DNA ladder was observed in these cells,supporting the activation of an endonuclease (Figure5D). Elimination of Ca2l from the culture medium didnot affect the appearance of apoptotic cells (data notshown). The protein synthesis inhibitor cyclohexim-ide did not inhibit the process of apoptosis, butcycloheximide itself induced apoptosis in McA-RH7777 cells (data not shown).
Induction of Apoptosis in McA-RH7777Cells by Serum DeprivationApoptosis was also induced by deprivation of trophicfactors from culture media. In serum-free culturemedia, McA-RH7777 cells showed extensive celldeath by way of apoptosis 2 to 3 days after serumdeprivation (Figure 6). However, cultured cells werenot diminished after being in SFM for long periods(more than 2 months with changing medium at 2-day
intervals in the same culture flask), suggesting thatthe cell death was counterbalanced by mitosis inSFM culture and that stimulation of mitosis and ap-optosis are concurrent in this system.
Apoptosis Observed in Confluent CultureApoptotic cells are rarely observed when the cul-tured cells are sparse. However, many apoptoticcells were observed in high-cell-density culture (con-fluent stage) (Figure 7). Eosinophilic globules, oftencontaining nuclear remnants (apoptotic bodies),were frequently found within the cytoplasm of intacthepatoma cells, suggesting phagocytosis of deadcells by intact cells. The appearance of apoptoticcells in the confluent stage might be due in part tonutritional deficiency and accumulation of toxicmetabolites in the medium. However, frequentmedium changes did not abrogate the appearanceof apoptotic cells, and several mitotic cells were fre-quently observed in the same culture, indicating thatstimulation of cell growth and death were concurrent

Apoptosis of Rat Hepatoma Cells 943A/PMarch 1993, Vol. 142, No. 3
W*S *r 1 2
bp1353
^ 31078-^ t - i ~~~~~~872-
l ~~~~~~~~~~~~~~~~~603-_
D~~~~l~~~~~~~~~~~~~~~~~. 31
%~; * " ~~~~~~~~~~~~~~~~194-
Figure 6. Demonistration ofapoptosis in McA-RH7777 cells culttired in SFM. Phase-contrast photomicrograph (A) and H&Estain (B) ofMcA-RH7777cells ctnltzured in SFMfor 3 days. Shrinkage anzdfragmentation of dead cells are demonstrated in (A), anid nnticlearfragmentationi uwith condentsedchronmatini is clearly observed in many apoptotic cells (somie arrowed) in (B) (A X 106, B x 212). C: Electrophoretic anlalysis ofDNAs isolatedfromcontrol cl(ltiure in mediumni conitaininlg 150% fetal bovine serun (lane 1) anid cells clultlured ini SEM for i da)s (lane 2).
4.
Figure 7. Histological evidence of apoptosis in McA-RH7777 cells at
confluency. Apoptosis is characterized by cell sh-Inkage and
fragmentation. Apoptotic bodies are seen within thecytoplasm ofadja-
cenit hepatomna cells.s Several mitotic cells are also observed in close
vicinity to apoptotic cclls (H&E. X 206)
in the confluent culture of McA-RH7777 cells. This
suggests the possibility that the apoptotic process is
actively induced to maintain homeostasis of cellular
density by counterbalancing mitoses in the expand-
ing tumor population.
Discussion
The focus of molecular oncology is likely to remain on
the control of proliferation. However, there is now
compelling evidence that the other side of the equa-
tion, the rate of cell death, must also be
considered.22 Cell death may occur by necrosis or
through the specific process of apoptosis. Apop
tosis is now widely recognized as a common mech-anism for physiological cell deletion and plays a roleopposite to that of mitosis in cell population kinetics.7Association of tumor suppressor gene p53 with apo-ptosis has recently been reported, suggesting aclose relationship between apoptosis and carcino-genesis.23A number of the steps leading to apoptosis in
immature thymocytes and lymphoma cells are welldefined.12-14,24 These include Ca2+ influx into thecell and endonuclease activation in the nuclei thatproduces a characteristic DNA fragmentation (DNAladder) of multi-nucleosomal length, due to cleavageat the internucleosomal linker region of the chromatinDNA. Apoptotic cells are recognized at the lightmicroscopic level by the characteristic morphologyof cell shrinkage with condensed and fragmentednuclei, which are then phagocytosed by macro-phages or parenchymal cells.2 3,7 From these crite-ria, it is now obvious that the apoptotic process ishighly activated in the Morris hepatoma 7777 tissuetransplanted into muscle tissue. Distinct DNA ladderpatterns were also demonstrated in the liver cells atearly stages of ischemic necrosis. Although thesteps in the DNA ladder became smaller due to pro-gressive DNA hydrolysis, the ladder pattern wasobserved against a background of a smear even at24 hours after ischemia. The DNA ladder was faintlyobserved as well in the CCI4-treated liver showingcentrilobular coagulative necrosis. The degree of theladder pattern was, however, very weak in spite ofcell death of more than 1/4 of the parenchymal cells.

944 Fukuda et alAJP March 1993, Vol. 142, No. 3
Differences in the DNA fragmentation pattern inthese two liver lesions showing coagulative necrosisindicate that the processes of DNA fragmentationmay be altered in necrotic cells depending on thecause of injury. Although Arends et al14 reported thatthe structural changes in the nucleus in apoptoticthymocytes are the direct result of a selectivenuclease activation within dying cells and althoughDNA fragmentation into oligonucleosomal units hasbeen widely accepted as a characteristic biochem-ical indicator for apoptosis12 14, the DNA ladder pat-tern does not seem to be confined to only apoptoticcell death in liver cells.Many studies have shown that alterations in
intracellular Ca2+ homeostasis can be criticallyinvolved in cytotoxicity.24 One prevalent hypothesisis that a sustained elevation of cytosolic Ca2+ con-
centration can stimulate Ca2+-dependent degener-ative enzymes, such as proteases, phospholipases,and nucleases, resulting in irreversible cell damagethat leads to cell death.4 6,17 McConkey et alreported that Ca2+ ionophore can induce a DNAladder in thymocytes and suggested that Ca2+ influxis important for the internucleosomal DNA digest.24This may not be true of all cell types. Cell deathinduced by Ca2+ ionophore in cultured McA-RH7777 cells was shown to be necrosis, judgingfrom the morphology and DNA fragmentationpattern. Increased level of intracellular Ca2+ byCa2+ ionophore does not seem to be sufficient toproduce characteristic DNA ladder formation andthe morphology of apoptosis in hepatoma cells.
It has recently been suggested that changes inchromatin structure may play a critical role in theinduction of DNA cleavage in the early phases ofapoptosis.25 Rat liver nuclei have been known tohave endogenous nucleases of the micrococcalnuclease and pancreatic DNAse type.15 The formerpreferentially attacks linker DNA between nucleo-somes, giving rise to DNA fragmentation of oligonu-cleosomal units. The latter type of nuclease pro-
duces single-strand cuts in nucleosomal core andinternucleosomal DNA to give fragments that are
regular multimers of 10 to 11 nucleotides, which mayresult in a smear pattern of DNA fragmentation inagarose gel electrophoresis. Morris hepatoma 7777cells have been reported to have low nuclear endo-nuclease activity.15 Thus, the activation of internu-cleosomal DNA cleavage in 7777 cells is a finelyregulated active process and is not merely a result ofCa2+ influx due to cell damage or death.
The apoptotic process is involved in many physi-ological and pathological processes in liver tissue,such as normal cell turnover, involution of hyperplas-
tic liver, and regression of preneoplastic cells.26-30The cellular process of apoptosis is, however, largelyunknown. De novo protein synthesis is reported to benecessary for the induction of apoptosis in severalcell lineages.9 13 However, conflicting results arealso reported by many investigators.10' 1131'32 War-ing has shown that cycloheximide and the transcrip-tional inhibitor actinomycin D have no effect ongliotoxin-induced apoptosis of macrophages andthat these agents are capable of enhancing apopto-sis of these cells.31 Cycloheximide did not inhibitapoptosis induced by hyperthermia in a Burkitt's lym-phoma cell line.11 All these recent findings questionthe validity of the widely held view that active proteinsynthesis is an invariable prerequisite for the execu-tion of apoptosis. In this paper, we demonstrated thatthe cell death in cultured hepatoma cells can bedivided into two major pathways, direct and indirectcell death. Direct cell death, which is inducible bysevere cellular damage, demonstrated features ofnecrosis. On the other hand, indirect cell death,which is inducible by mild cellular injury or elimina-tion of trophic factors, showed the characteristic fea-tures of apoptosis. Similar dose-dependent induc-tion of apoptosis by a range of injurious agents werealso demonstrated in several hematopoietic celllines.10 Lennon et al reported that cells sufferingminor injury have the capacity to activate an inter-nally programmed death mechanism, whereas celldeath induced by a greater injury may take the formof necrosis.10 The characteristic DNA ladder andmorphological signs of apoptosis were reproduciblydemonstrated in McA-RH7777 cells by mild doses ofUV exposure. This in vitro experimental model seemsto be useful in the elucidation of the process of apo-ptosis in hepatoma cells.
In the earliest study of apoptosis, Kerr demon-strated the induction of apoptosis in normal liver cellsby deprivation of the portal blood supply.33 The SFMthat was used in our experiment has been shown toretain liver function and cellular activity for long peri-ods of time in many established hepatoma celllines.19 Therefore, it is interesting that the McA-RH7777 cells can proliferate, but that many cells dieby way of apoptosis at the same time in SFM. Thissuggests that McA-RH7777 cells are highly suscep-tible to apoptosis in the absence of serum growthfactors and that cell mitosis and apoptosis are finelyregulated in this experimental model.
Even in the presence of serum, many apoptoticcells were demonstrated when the cell densitybecame high (confluent stage). It might be possiblethat nutritional deficiency and accumulation of toxicmetabolites are involved in the process of apoptotic

Apoptosis of Rat Hepatoma Cells 945AJP March 1993, Vol. 142, No. 3
cell death in confluent culture. However, frequentmedium change could not inhibit the appearance ofapoptotic cells, and mitotic cells were frequentlyobserved in the same culture, suggesting that thestimulation of mitosis and apoptosis are concurrentin confluent cultures of McA-RH7777 cells. Cell andtissue growth is regulated through a complex inter-play of stimulatory and inhibitory signals. It is pro-posed that proliferation and apoptosis together com-prise the regulation of cell growth.34 The apoptoticprocess seems to be activated to regulate homeo-stasis in cell-population kinetics at the confluentstage by counterbalancing mitoses. It is also sug-gested that a disturbance in oxygen and nutritionalsupply may be one of the triggering factors in theinduction of apoptosis in 7777 tissue.
In conclusion, the cell death observed in trans-planted 7777 hepatoma tissue is mainly apoptotic innature. Based on the results obtained from in vitroexperiments, it was suggested that tumor apoptosismay represent a residual attempt at autoregulationwithin the expanding tumor population and/or mayresult from mild cellular injuries such as hypoxia,nutritional deficiency, or other unknown noxiousfactor(s).
The induction of apoptosis in cancer cells is one ofthe major concerns in recent studies in the develop-ment of cancer therapy.8,922 A number of studieshave explored the attractive idea that tumor cellscould be eliminated by artificially triggering celldeath through apoptosis.22'35 Cell-mediated cytotox-icity involves induction of at least some features ofapoptosis in the targets.36 The cell death that isinduced by irradiation, hyperthermia, and a range ofcytotoxic drugs including many used in cancer ther-apy has been found to be apoptosis.8'9'11 Clarifica-tion of the biochemical pathways involved in the acti-vation of apoptosis should lead to fundamentaladvances in the treatment of cancer. Therefore, thiscell line seems to be useful as an in vivo and in vitroexperimental model for the elucidation of apoptosisin hepatoma cells.
AcknowledgmentWe wish to acknowledge Dr. Roy Korson for criticalreview of this manuscript.
References1. Cooper EH, Bedford AJ, Kenny TE: Cell death in normal
and malignant tissues. Adv Cancer Res 1975, 21:59-120
2. Wyllie AH, Kerr JFR, Currie AR: Cell death: the signifi-cance of apoptosis. Int Rev Cytol 1980, 68:251-306
3. Searle J, Kerr JFR, Bishop CJ: Necrosis and apoptosis:distinct modes of cell death with fundamentally differ-ent significance. Pathol Annu 1982, 17:229-259
4. Farber JL, Chien KR, Mittnacht S Jr: The pathogenesisof irreversible cell injury in ischemia. Am J Pathol 1981,102:271-281
5. Farber JL: Biology of disease: membrane injury andcalcium homeostasis in the pathogenesis of coagula-tive necrosis. Lab Invest 1982, 47:114-123
6. Popper H: Hepatocellular degeneration and death. TheLiver: Biology and Pathobiology. Edited by Arias IM,Jakoby WB, Popper H, Schachter D, Shafritz DA, NewYork, Raven Press, 1988, pp 1087-1103
7. Kerr JFR, Harmon BV: Definition and incidence of ap-optosis: an historical perspective. Apoptosis: theMolecular Basis of Cell Death. Edited by Tomein LD,Cope FO, New York, Cold Spring Harbor LaboratoryPress, 1991, pp 5-29
8. Dive C, Hickman JA: Drug-target interactions: only thefirst step in commitment to a programmed cell death?Br J Cancer 1991, 64:192-196
9. Barry MA, Behnke CA, Eastman A: Activation of pro-grammed cell death (apoptosis) by cisplatin, other anti-cancer drugs, toxins and hyperthermia. Biochem Phar-macol 1990, 40:2353-2362
10. Lennon SV, Martin SJ, Cotter TG: Dose-dependentinduction of apoptosis in human tumor cell lines bywidely diverging stimuli. Cell Prolif 1991, 24:203-214
11. Takano YS, Harmon BV, Kerr JFR: Apoptosis inducedby mild hyperthermia in human and murine tumor celllines: a study using electron microscopy and DNA gelelectrophoresis. J Pathol 1991, 163:329-336
12. Cohen JJ, Duke RC: Glucocorticoid activation of a cal-cium-dependent endonuclease in thymocyte nucleileads to cell death. J Immunol 1984, 132:38-42
13. Wyllie AH, Morris RG, Smith AL, Dunlop D: Chromatincleavage in apoptosis: association with condensedchromatin morphology and dependence on macromo-lecular synthesis. J Pathol 1984, 142:67-77
14. Arends MJ, Morris RG, Wyllie AH: Apoptosis: the role ofthe endonuclease. Am J Pathol 1990, 136:593-608
15. Vanderbilt JN, Bloom KS, Anderson JN: Endogenousnuclease: properties and effects on transcribed genesin chromatin. J Biol Chem 1982, 257:13009-13017
16. Jones DP, McConkey DJ, Nicotera P, Orrenius S: Cal-cium-activated DNA fragmentation in rat liver nuclei. JBiol Chem 1989, 264:6398-6403
17. Thomas CE, Reed DJ: Current status of calcium inhepatocellular injury. Hepatology 1989, 10:375-384
18. Ray SD, Sorge CL, Tavacoli A, Rancy JL, Corcoran GB:Extensive alteration of genomic DNA and rise innuclear Ca2+ in vivo early after hepatotoxic acetami-nophen overdose in mice. Biological Reactive Interme-diates, vol IV. Edited by Witmer CM, New York, PlenumPress, 1990, pp 699-705

946 Fukuda et alAJP March 1993, Vol. 142, No. 3
19. Nakabayashi H, Taketa K, Yamane T, Miyazaki M, Miy-ano K, Sato J: Phenotypical stability of a humanhepatoma cell line, HuH-7, in long term culture withchemically defined medium. Jpn J Cancer Res 1985,75:151-158
20. Pressman BS: Biological applications of ionophores.Annu Rev Biochem 1976, 45:501-530
21. Sambrook J, Fritsch EF, Maniatis T: Molecular Cloning:a Laboratory Manual. New York, Cold Spring HarborLaboratory Press, 1989, pp 9.16-9.21
22. Williams GT: Programmed cell death: apoptosis andoncogenesis. Cell 1991, 65:1097-1098
23. Yonish-Rouach E, Resnitzky D, Lotem J, Sachs L, Kim-chi A, Oren M: Wild-type p53 induces apoptosis ofmyeloid leukemic cells that is inhibited by interleukin-6.Nature 1991, 352:345-347
24. McConkey DJ, Hartzell P, Nicotera P, Orrenious S: Cal-cium-activated DNA fragmentation kills immaturethymocytes. FASEB J 1989, 3:1843-1849
25. Alnemri ES, Litwack G: Activation of internucleosomalDNA cleavage in human CEM lymphocytes by gluco-corticoid and novobiocin: evidence for a non-Ca2+-requiring mechanism(s). J Biol Chem 1990, 265:17323-17333
26. Columbano A, Ledda-Columbano GM, Rao PM,Rajalakshmi S, Sarma DSR: Occurrence of cell death(apoptosis) in preneoplastic and neoplastic liver cells;a sequential study. Am J Pathol 1984, 116:441-446
27. Bursch W, Lauer B, Timmermann-Trosiener I, Barthel G,Schuppler J, Schulte-Hermann R: Controlled death(apoptosis) of normal and putative preneoplastic cellsin rat liver following withdrawal of tumor promoters.Carcinogenesis 1984, 5:453-458
28. Bursch W, Taper HS, Lauer B, Schulte-Hermann R:Quantitative histological and histochemical studies onthe occurrence and stages of controlled cell death(apoptosis) during regression of rat liver hyperplasia.Virchows Archiv [B] 1985, 50:153-166
29. Searle J, Harmon BV, Bishop CJ, Kerr JFR: The signif-icance of cell death by apoptosis in hepatobiliarydisease. J Gastroenterol Hepatol 1987, 2:77-96
30. Schulte-Hermann R, Timmermann-Trosiener I, BarthelG, Bursch W: DNA synthesis, apoptosis, and pheno-typic expression as determinants of growth of alteredfoci in rat liver during phenobarbital promotion. CancerRes 1990, 50:5127-5135
31. Waring P: DNA fragmentation induced in macrophagesby gliotoxin does not require protein synthesis and ispreceded by raised IP3 levels. J Biol Chem 1990,265:14476-14480
32. Collins RJ, Harmon BV, Souvlis T, Pope JH, Kerr JFR:Effects of cycloheximide on B-chronic lymphocytic leu-kaemic and normal lymphocytes in vitro: induction ofapoptosis. Br J Cancer 1991, 64:518-522
33. Kerr JFR: Shrinkage necrosis: a distinct mode of cellardeath. J Pathol 1971, 105:13-20
34. Rotello RJ, Lieberman RC, Purchio AF, GerschensonLE: Coordinated regulation of apoptosis and cell pro-liferation by transforming growth factor ,B1 in cultureduterine epithelial cells. Proc Natl Acad Sci USA 1991,88:3412-3415
35. Kyprianou N, English HF, Isaacs JT: Programmed celldeath during regression of PC-82 human prostate can-cer following androgen ablation. Cancer Res 1990,50:3748-3753
36. Duvall E, Wyllie AH: Death and the cell. Immunol Today1986, 7:115-119