deoxynivalenol induced mouse skin cell proliferation and inflammation via mapk pathway
TRANSCRIPT

1
2
3Q1
4
567
8
910111213
14151617181920
38
3940
41
42
43Q12
44
45
46
47
48Q2
49
50
51
52
53
54
Toxicology and Applied Pharmacology xxx (2014) xxx–xxx
YTAAP-13117; No. of pages: 12; 4C:
Contents lists available at ScienceDirect
Toxicology and Applied Pharmacology
j ourna l homepage: www.e lsev ie r .com/ locate /ytaap
Deoxynivalenol induced mouse skin cell proliferation and inflammationvia MAPK pathway
OFSakshi Mishra a,b, Anurag Tripathi a, Bhushan P. Chaudhari c, Premendra D. Dwivedi a,
Haushila P. Pandey b, Mukul Das a,⁎a Food Drug and Chemical Toxicology, CSIR-Indian Institute of Toxicology Research (CSIR-IITR), P.O. Box No. 80, Mahatma Gandhi Marg, Lucknow 226 001, Indiab Department of Biochemistry, Banaras Hindu University (BHU), Varanasi, Indiac Pathology Laboratory, CSIR-Indian Institute of Toxicology Research (CSIR-IITR), Mahatma Gandhi Marg, PO Box 80, Lucknow 226001, Uttar Pradesh, India
OAbbreviations: DON, deoxynivalenol; DMBA, 7,12MAPK, mitogen-activated protein kinase; NF-κB, nucletetradecanoyl phorbol-13-acetate; ODC, ornithine decarbo⁎ Corresponding author. Fax: +91 522 2628227.
E-mail addresses: [email protected], mditrc@hot
http://dx.doi.org/10.1016/j.taap.2014.06.0030041-008X/© 2014 Published by Elsevier Inc.
Please cite this article as: Mishra, S., et al., DAppl. Pharmacol. (2014), http://dx.doi.org/1
Ra b s t r a c t
a r t i c l e i n f o21
22
23
24
25
26
27
28
29
30
31
32
Article history:Received 6 January 2014Revised 5 June 2014Accepted 6 June 2014Available online xxxx
Keywords:MycotoxinsDeoxynivalenolSkin cell proliferationSkin inflammationCytokinesPeyer's patches
33
34
35
36
37
ECTED P
Several toxicological manifestations of deoxynivalenol (DON), a mycotoxin, are well documented; however,dermal toxicity is not yet explored. The effect of topical application of DON to mice was studied using markersof skin proliferation, inflammation and tumor promotion. Single topical application of DON (84–672 nmol/mouse)significantly enhanceddermal hyperplasia and skin edema.DON(336and672 nmol) caused significant enhancementin [3H]-thymidine uptake in DNA along with increased myeloperoxidase and ornithine decarboxylase activities,suggesting tissue inflammation and cell proliferation. Furthermore, DON (168 nmol) caused enhancedexpression of RAS, and phosphorylation of PI3K/Akt, ERK, JNK and p38 MAPKs. DON exposure also showedactivation of transcription factors, c-fos, c-jun and NF-κB along with phosphorylation of IkBα. Enhanced phos-phorylation of NF-κB by DON caused over expression of target proteins, COX-2, cyclin D1 and iNOS in skin.Though a single topical application of DMBA followed by twice weekly application of DON (84 and 168 nmol)showed no tumorigenesis after 24 weeks, however, histopathological studies suggested hyperplasia of theepidermis and hypertrophy of hair follicles. Interestingly, intestine was also found to be affected as enlargedPeyer's patches were observed, suggesting inflammatory effects which were supported by elevation of inflammatorycytokines after 24 weeks of topical application of DON. These results suggest that DON induced cell proliferation inmouse skin is through the activationofMAPKsignalingpathway involving transcription factorsNFκBandAP-1, furtherleading to transcriptional activation of downstream target proteins c-fos, c-jun, cyclin D1, iNOS and COX-2 whichmight be responsible for its inflammatory potential.
© 2014 Published by Elsevier Inc.
R55
56
57
58
59
60
61
62
63
64
65
66
67
UNCO
R
Introduction
Mycotoxins are toxic secondary metabolites produced by numerousspecies of fungi (Bennett and Klich, 2003). Deoxynivalenol (DON) is oneof the most frequently occurring Fusarium mycotoxins that predomi-nantly contaminate wheat, maize and barley in temperate regions(CAST, 2003; Schothorst and Van Egmond, 2004). DON ingested athigher doses causes nausea, vomiting, and diarrhea; while at lowerdose it exhibits weight loss and food refusal (Rotter et al., 1996). Dueto the toxic effects of DON, its presence in food commodities is gainingglobal attention. DON is not only the most commonly detected tricho-thecene in cereal grains, but has been found at exceedingly high levels(Almeida et al., 2012; Canady et al., 2001; Edwards et al., 2011; Lee
68
69
70
71
72
73
-dimethylbenz[α]anthracene;ar factor-kappaB; TPA, 12-O-xylase; MPO, myeloperoxidase.
mail.com (M. Das).
eoxynivalenol induced mouse0.1016/j.taap.2014.06.003
et al., 2011; Mishra et al., 2013; Van Der Fels-Klerx et al., 2012).Considering the toxicological data on DON, Joint FAO/WHO ExpertCommittee on Food Additives (JECFA, 2010) has recommended aPMTDI (provisional maximum tolerable daily intake) of 1 μg/kg bw.
There are a few reports on DON showing gastrointestinal andimmune-toxicity in humans (Li et al., 1999; Pestka and Smolinski,2005). An acute outbreak of DON intoxication was reported in theKashmir valley, India affecting about 50,000 persons followingconsumption of contaminated wheat flour (Bhat et al., 1989). Theincidence of esophageal cancer in China and Africa has been linked toDON contamination; however, no clear cut evidence has beendemonstrated (Hsia et al., 1988; Luo et al., 1990). Although DONlacks the ability to cause gene mutation, it is certain that DON notonly alters chromosome aberration, DNA damage and repair butcan also make mammalian cells transform into malignant ones,which suggests that DON has the potential to be a genotoxic carcinogen(Ma and Guo, 2008). However, IARC (1993) has categorized DON ingroup 3, “not classifiable as to its carcinogenicity to humans” becauseof lack of sufficient evidences particularly related to in vivo studies. In
skin cell proliferation and inflammation via MAPK pathway, Toxicol.

TE
74
75
76
77
78
79
80Q3
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140Q4
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
175
176
177
178
179
180
181
182
183
184
185
186
187
188
189
190
191
192
193
194
195
196
2 S. Mishra et al. / Toxicology and Applied Pharmacology xxx (2014) xxx–xxx
UNCO
RREC
a 2-year study, dietary exposure to DON showed no incidence of tumorsin B6C3F1mice (Iverson et al., 1995). However, a high incidence of lungadenocarcinoma and dysplasia of glandular stomach was observed inNIH mice orally exposed to DON after 24 weeks (Huang et al., 2004).
In the recent past WHO has highlighted the need for toxicologicalstudies on mycotoxins through dermal exposure as limited data isavailable following dermal exposure risk to mycotoxins. This is ofimportance in developing countries situated in tropical region; manuallabor employed during pre- and post-harvest stages of agriculturalproduction is vulnerable to mycotoxin exposure through dermal route.In this context our prior studies have revealed that dermal exposure tosomemycotoxins viz aflatoxin B1 (AFB1), patulin, citrinin, and ochratoxinA (OTA) caused skin toxicity including tumor formation (Kumar et al.,2011, 2012; Rastogi et al., 2006; Saxena et al., 2009, 2011). In a recentstudy, the transdermal kinetics of AFB1, OTA, fumonisin B1, citrinin,zearalenone and T-2 toxin were quantitatively evaluated indicating thatmycotoxins can penetrate through the skin, which may lead to toxicmanifestations (Boonenet al., 2012). Due to thewidespread contaminationof DON, it is likely that humans involved in agricultural practices may berepeatedly exposed to DON through dermal route.
A correlation between stimulation of cell proliferation, inflamma-tion and tumor promotion is well established as continuous inductionof cell proliferationmay lead to cell transformation and tumor develop-ment (Poirier, 2004). Studies have shown that skin application oftumor-promoting agent results in inflammatory responses, such as de-velopment of edema, hyperplasia, and induction of proinflammatorycytokines, induction of dermal ornithine decarboxylase (ODC) andcyclooxygenase-2 (COX-2) protein expression, as well as activation ofNF-κB (Chun et al., 2002; Katiyar et al., 1995, 1996; Seo et al., 2002).Activation of mitogen-activated protein kinases (MAPKs)/NF-κB path-ways has been shown to be involved in tumor growth and development(Afaq et al., 2003; Shishodia et al., 2003).
The present study was designed to gain a better understanding ofthe cellular events leading to DON mediated in vivo dermal toxicityand more particularly to investigate whether DON causes induction ofcell proliferation via activation of MAPK pathway which may furtherlead to tumor formation.
Materials and methods
Chemicals. Deoxynivalenol (DON), dithiothreitol (DTT), phenylmethyl-sulphonyl fluoride (PMSF), and diphenylamine (DPA) 7,12-dimethylbenz[α]anthracene (DMBA), 12-O-tetradecanoyl phorbol-13-acetate (TPA),phenylmethylsulfonyl fluoride (PMSF), 2-mercaptoethanol, acrylam-ide, protease inhibitor cocktail set-I, ethylenediamine tetraacetic acid(EDTA) disodium salt, Tris buffer, Triton X-100, bovine serum albumin(BSA), and HRP conjugated β-actin were obtained from SigmaChemicals Co. (St. Louis, MO). [14C]-Ornithine (specific activity,56 mCi/mol) and [3H]-thymidine (specific activity, 1.0 mCi/ml) werepurchased from Amersham Biosciences (Chicago, IL). Rabbit polyclonalantibodies against phospho-ERK1/2, ERK1/2, phospho-p38, phospho-JNK, anti-p-AKT, anti-c-jun, anti-c-fos, anti-p-NFкB, anti-p-IκBα, anti-Cox-2 and anti-cyclin-D1were procured from Santa Cruz Biotechnology(Santa Cruz, CA). Horseradish peroxidase (HRP) conjugated goat anti-rabbit IgG secondary antibody was obtained from Bangalore Genei(Bangalore, India). Anti-p-PI3K, anti-RAS and cytometric bead arraykit was purchased from BD Biosciences (San Diego, CA). All otherchemicals used were of the highest purity commercially available.
Animals and ethics statement. Six to seven week old female Swiss albinomice (20 ± 3 g), derived from the animal breeding colony of CSIR-Indian Institute of Toxicology Research (CSIR-IITR), Lucknow, wereacclimatized under standard laboratory conditions and given acommercial pellet diet (Ashirwad Industries, Chandigarh, India) andwater ad libitum. Animals were housed in plastic cages having ricehusk as bedding and maintained in a controlled atmosphere of 12 h
Please cite this article as: Mishra, S., et al., Deoxynivalenol induced mouseAppl. Pharmacol. (2014), http://dx.doi.org/10.1016/j.taap.2014.06.003
dark/light cycle, 22 ± 2 °C temperature and 50–60% humidity as perrules laid down by the Animal Welfare Committee of CSIR-IITR. All theexperiments involving animals were approved by the Institutional Ani-mal Ethics Committee (IAEC) via approval # ITRC/IAEC/30/11-22/12,CSIR-IITR. Mice were shaved with an electric clipper (Oster, WI, USA)one week prior to the beginning of the experiment. Mice showing nosigns of hair growth or in resting phase of hair cycle were used forfurther experiments. Animals were sacrificed by cervical dislocation asper CSIR-IITR guidelines.
D P
RO
OF
Edema and dermal hyperplasia. To assess the effect of DON on skinedema, animals were distributed randomly into six groups, having fivemice per group. The mice of 1st group received a single topical applica-tion of 0.2 ml acetone to serve as a control, while animals of 2nd, 3rd,4th and the 5th groups received a single topical application of 84, 168,336, and 672 nmol DON in 0.2 ml acetone, respectively. The 6th groupreceived topical application of TPA (4 nmol) and served as positive con-trol. The topical doses of DON (84 nmol and 168 nmol) were chosenfrom the earlier study (Lambert et al., 1995). Following 24 h and 48 hof treatment, mice were sacrificed and 1 cm diameter punches of skinfrom the vehicle, and DON treated and TPA-treated animals were re-moved, made free of fat and weighed quickly. After drying for 24 h at50 °C, the skin punches were reweighed, and the loss of water contentwas determined (Saleem et al., 2004). The difference in the amount ofwater gain between the control (vehicle treated) andDONor TPA treatedgroups represented the extent of edema induced by DON. For thehyperplasia study, skin was immediately removed, washed in coldnormal saline solution, fixed in 10% buffered formalin and embeddedin paraffin after processing. Sections of 5.0 μm thickness were cut andstained with hematoxylin and eosin for microscopic examination.
[3H]-thymidine incorporation assay. For estimation of cell proliferation,female Swiss albino mice of the same age were used as described inthe earlier section. Mice were divided into 6 groups of 5 mice each:(i) acetone (0.2 ml) topical application, as vehicle control; (ii) TPA(4 nmol) was topically applied as positive control; and (iii) DON(84, 168, 336, 672 nmol/mouse). The animals of all the groupswere given i.p. injection of [3H]-thymidine 2 h prior to sacrifice.Animals were sacrificed by cervical dislocation and skin was excised.Dermal DNA was isolated and assessment of incorporation of [3H]-thymidine into DNA was carried out according to the method ofGupta and Mehrotra (1992).
Myeloperoxidase assay. Myeloperoxidase (MPO) was determined as amarker of inflammation in the skin. The activity of MPO was assessedin skin samples obtained from the mice. Mice were divided into 5groups of 5 animals each. Mice were topically treated with 84, 168,336, and 672 nmol of DON following 24 h treatment. MPO wasmeasured as a marker of inflammation in skin homogenate samplesfollowing the procedure of Bradley et al. (1982). Briefly, the skinsamples were homogenized in 50 mM potassium phosphate buffer,pH 6.0, containing 0.5% hexadecyltrimethylammonium bromidefollowed by sonication of the homogenate at 4 °C for 10 s bursts(three times). Further the samples were centrifuged at 13,000 ×g for10 min at 4 °C and the resulting supernatants were used for MPO esti-mation.MPOactivity in the supernatant (0.1ml)was assayed bymixingwith 50 mMphosphate buffer (2.9 ml) pH 6.0, containing 0.167 mg/mlo-dianisidine dihydrochloride and 0.0005% hydrogen peroxide. Thechange in absorbance resulting from decomposition of H2O2 in thepresence of o-dianisidine was measured using Power Wave XS2 platereader (Biotek, Winooski, VT) at 460 nm. The activity was calculatedusing an extinction coefficient of 178 mM−1 cm−1, and the data areexpressed as the mean MPO U/mg protein.
skin cell proliferation and inflammation via MAPK pathway, Toxicol.

T
197
198
199
200
201
202
203
204
205
206
207
208
209
210
211
212
213
214
215
216
217
218
219
220
221
222
223
224
225
226
227
228
229
230
231
232
233
234
235
236
237
238
239
240
241
242
243
244
245
246
247
248
249
250
251
252
253
254
255
256
257
258
259
260
261
262
263
264
265
266
267
268
269
270
271
272
273
274
275
276
277
278
279
280
281
282
283
284
285
286
287
288
289
290
291
292
293
294
295
296
297
298
299
300
301
302
303
304
305
306
307
308
309
310
311
312
313
314
315
3S. Mishra et al. / Toxicology and Applied Pharmacology xxx (2014) xxx–xxx
UNCO
RREC
Ornithine decarboxylase (ODC) enzyme assay. Studies with the mouseskin model have shown an excellent correlation between the inductionof ODC activity and the tumor-promoting ability of a variety ofsubstances (Ahmad et al., 2001; Einspahr et al., 2003). To study thetime dependent effect of DON on cutaneous ODC activity, the micewere divided into ten groups containing 5 animals each: (i) untreated,served as control (ii) acetone (0.2 ml) topical application, as vehiclecontrol; (iii) TPA (4 nmol) was topically applied to serve as positivecontrol; and (iv) DON (336 nmol/mouse). The animalswere euthanizedafter 2, 4, 6 and 8 h of TPA and DON. For dose dependent study, animalswere randomly divided into 8 groups having 5mice each: (i) untreated,(ii) acetone (0.2 ml) topical application as vehicle control; (iii) TPA(4 nmol) was topically applied as positive control; and (iii) DON(84, 168, 336, 672 nmol/mouse) was topically applied. The animalswere euthanized after 4 h by cervical dislocation following therules laid down by the Animal Welfare Committee of IITR. The skinwas dissected out and cytosolic ODC activity was assayed using themethod of Verma et al. (1979). The specific activity was expressedas pmol 14CO2 released/h/mg protein.
To study the ODC expression at the protein level in epidermis, ani-mals were treated at different time points. Following DON treatment(168 nmol), the animals were sacrificed, dermal lysates were preparedand protein contentwas determined by BCAmethod. Equal loadingwasconfirmed by stripping the immunoblot and reprobing it for β-actin.
Preparation of whole cell extract. Female Swiss albino mice were topicallytreated with acetone or DON (168 nmol) on the dorsal skin andsacrificed by cervical dislocation after 1, 3, 6, 12, 24, or 48 h. The skinwas excised and fat was removed on ice. The isolated fat free skin tissuefrom DON treated and acetone treated animals was rinsed with normalsaline and homogenizedwith anUltra Turrax Polytron (Janke & Kunkel,IKA-Labortechnik, Staufen, Germany) in ice cold-RIPA buffer (50 mMTris pH: 7.5, 150 mM NaCl, 1% Triton ×100, 0.5% sodium deoxycholate,0.1% SDS, 0.2 mM PMSF) with 1×freshly added protease inhibitorcocktail. The cell lysate was kept on ice for 10 min and centrifuged at16,000 ×g for 20min at 4 °C. The clear supernatant (whole cell extract)was used for the detection of phosphorylated proteins.
Preparation of nuclear extracts. Female Swiss albino mice were topicallytreated with acetone or DON (168 nmol) on the dorsal skin andsacrificed by cervical dislocation at 1, 3, 6, 12, 24, or 48 h. Skin was ex-cised and fat was removed on ice. The isolated fat scraped skin tissuewas rinsed with normal saline, and homogenized in ice cold hypotonicbuffer A [10 mM HEPES (pH 7.8), 10 mM KCl, 2 mM MgCl2, 1 mMDTT, 0.1 mM EDTA and 0.1 mM phenylmethylsulfonylfluoride(PMSF)]. An 80 μl of 10% Nonidet P-40 (NP-40) solution was added tohomogenate, incubated on ice for 10min and subsequently centrifugedfor 2 min at 14800 ×g. The supernatant was collected as cytosolic frac-tion. The nuclei pellet was washed with 500 μl of buffer A plus 40 μl of10% NP-40, centrifuged and resuspended in 200 μl of buffer C [50 mMHEPES (pH 7.8), 50 mM KCl, 300 mM NaCl, 0.1 mM EDTA, 1 mM DTT,0.1 mM PMSF and 20% glycerol]. This mixture was centrifuged for 5min at 14800 ×g and the supernatant containing nuclear proteins wascollected and stored at−80 °C after determination of proteinconcentrations.
Western blotting. An aliquot of whole cell lysate and nuclear extractcontaining 50 μg of protein was boiled in a 6×sodium dodecyl sulfate(SDS) sample-loading buffer for 5 min before electrophoresis on a 10%SDS-polyacrylamide gel. Proteins in the SDS-polyacrylamide gel wereelectro-transferred to PVDF membrane and the blots were blockedwith 5% nonfat drymilk PBST buffer (phosphate buffered saline containing0.1%Tween-20) for1hat roomtemperature (RT). Theblockedmembraneswere probed with ODC, phospho-ERK, phospho-p38, phospho-JNK, COX2,cyclin D1, c-fos, c-jun, p-IκBα, and phospho-NFκB (nuclear extract)phospho-AKT antibodies at dilutions indicated by the manufacturer
Please cite this article as: Mishra, S., et al., Deoxynivalenol induced mouseAppl. Pharmacol. (2014), http://dx.doi.org/10.1016/j.taap.2014.06.003
followed by incubation with HRP-conjugated secondary antibody.Further bound secondary antibody was detected by enhancedchemiluminescence using Amersham ECL western blotting detectionreagent kit as described in the manufacturer's protocol (Amersham,Fairfield, CT). All the blots were stripped and reprobed with β-actinto ensure equal loading of protein.
ED P
RO
OF
Evaluation of skin tumor promoting potential of DON. For tumor studies,6–7 weeks old female Swiss albino mice (20 ± 3 g) were randomlydivided into 6 groups having 10 animals each and were housed inseparate cages. The following experimental protocol was adopted:
Group 1: Vehicle control; (topical application of 0.2 ml acetone treatedtwice weekly).
Group2: Positive control; single topical application of DMBA(120nmolin 0.2 ml acetone) followed by twice weekly application ofTPA (4 nmol in 0.2 ml acetone).
Group 3: DON as a tumor promoter at 84 nmol; single topical applica-tion of DMBA (120 nmol in 0.2 ml acetone) followed bytwiceweekly application of DON (84 nmol in 0.2ml acetone).
Group 4: DON as a tumor promoter at 168 nmol; single topicalapplication of DMBA (120 nmol in 0.2 ml acetone)followed by twice weekly application of DON (168 nmolin 0.2 ml acetone).
Group 5: A single topical application of acetone (0.2 ml/mouse) followedby twice-weekly application of DON (84 nmol/0.2 ml acetone)after a week.
Group 6: A single topical application of acetone (0.2 ml/mouse) followedby twice-weekly application of DON (168 nmol/0.2 ml acetone)after a week.
The topical doses of DON(84 nmol and168 nmol)were chosen fromthe earlier study (Lambert et al., 1995). The skin tumor formation wasrecorded twice weekly for 24 weeks and tumors greater than 1 mm indiameter were included in the cumulative total only if they persistedfor 2 weeks or more. Cumulative tumors were defined as the totalnumber of tumors observed on each mouse during the experimentalduration.
Histopathological processing. The animals were sacrificed by cervicaldislocation at the termination of the experiment. Skin and intestinewere immediately removed and intestinewas flushedwith cold normalsaline solution. Both the tissueswere fixed in 10% buffered formalin andembedded in paraffin after processing. Sections of 5.0 μm thicknesswere cut and stained with hematoxylin and eosin for microscopicexamination.
Cytokine analysis. Serum was collected before the animals weresacrificed by cervical dislocation at the termination of the experiment(24 week protocol) and stored at−80 °C until use. Thawed sampleswere used for the estimation of IFN-γ, TNF-α, IL-6, IL-1, IL-17, IL-5 andIL-4 cytokines with cytometric bead array kit (BD Biosciences, SanJose, CA). Samples were prepared for cytokine analysis as directed bythe kit manufacturer and analyzed on the same day. CB FCAP evaluatedthe level of cytokines present in the sample on the basis of standardcurve obtained for each cytokine.
Statistical analysis. All the results were expressed as the mean ± SE.Differences between groups were analyzed using one-way ANOVAwith Bonferroni intergroup comparison tests from GraphPad Prism-5.0(GraphPad Prism software, San Diego, CA). A value of P b 0.05 wasconsidered as statistically significant.
skin cell proliferation and inflammation via MAPK pathway, Toxicol.
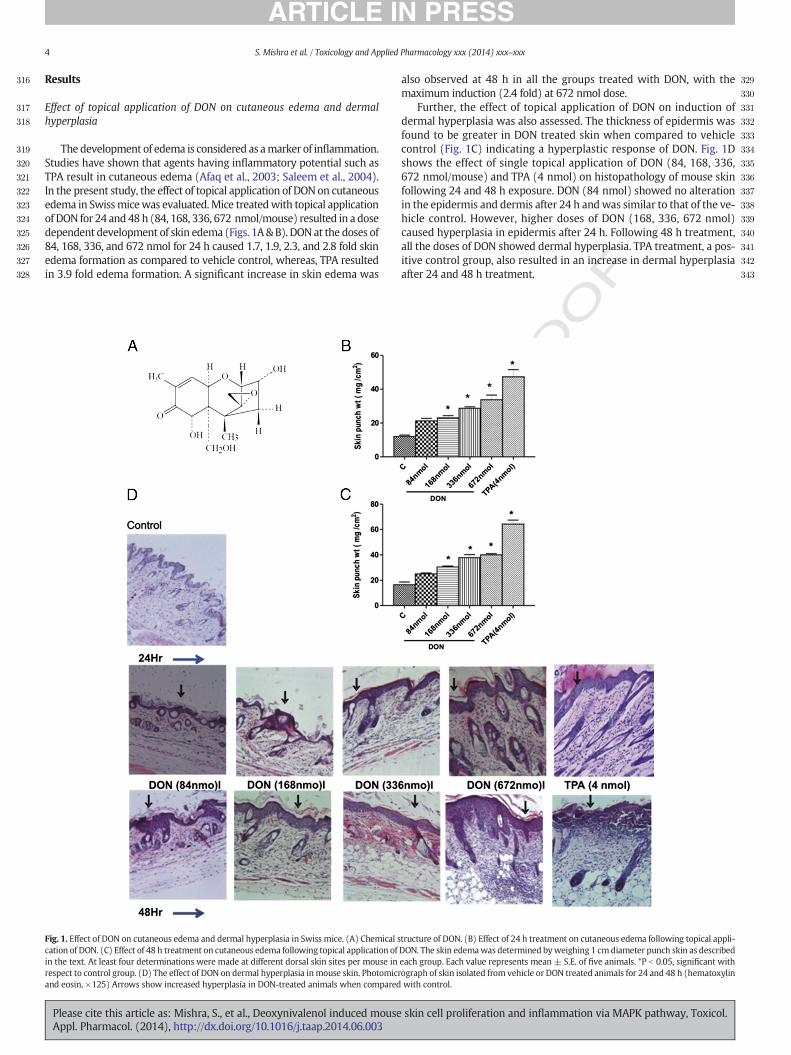
316
317
318
319
320
321
322
323
324
325
326
327
328
329
330
331
332
333
334
335
336
337
338
339
340
341
342
343
4 S. Mishra et al. / Toxicology and Applied Pharmacology xxx (2014) xxx–xxx
Results
Effect of topical application of DON on cutaneous edema and dermalhyperplasia
The development of edema is considered as amarker of inflammation.Studies have shown that agents having inflammatory potential such asTPA result in cutaneous edema (Afaq et al., 2003; Saleem et al., 2004).In the present study, the effect of topical application of DON on cutaneousedema in Swissmicewas evaluated.Mice treatedwith topical applicationof DON for 24 and 48h (84, 168, 336, 672 nmol/mouse) resulted in a dosedependent development of skin edema (Figs. 1A&B). DONat the doses of84, 168, 336, and 672 nmol for 24 h caused 1.7, 1.9, 2.3, and 2.8 fold skinedema formation as compared to vehicle control, whereas, TPA resultedin 3.9 fold edema formation. A significant increase in skin edema was
UNCO
RRECT
Fig. 1. Effect of DON on cutaneous edema and dermal hyperplasia in Swiss mice. (A) Chemical scation of DON. (C) Effect of 48 h treatment on cutaneous edema following topical application ofin the text. At least four determinations were made at different dorsal skin sites per mouse inrespect to control group. (D) The effect of DON on dermal hyperplasia inmouse skin. Photomicrand eosin, ×125) Arrows show increased hyperplasia in DON-treated animals when compared
Please cite this article as: Mishra, S., et al., Deoxynivalenol induced mouseAppl. Pharmacol. (2014), http://dx.doi.org/10.1016/j.taap.2014.06.003
also observed at 48 h in all the groups treated with DON, with themaximum induction (2.4 fold) at 672 nmol dose.
Further, the effect of topical application of DON on induction ofdermal hyperplasia was also assessed. The thickness of epidermis wasfound to be greater in DON treated skin when compared to vehiclecontrol (Fig. 1C) indicating a hyperplastic response of DON. Fig. 1Dshows the effect of single topical application of DON (84, 168, 336,672 nmol/mouse) and TPA (4 nmol) on histopathology of mouse skinfollowing 24 and 48 h exposure. DON (84 nmol) showed no alterationin the epidermis and dermis after 24 h andwas similar to that of the ve-hicle control. However, higher doses of DON (168, 336, 672 nmol)caused hyperplasia in epidermis after 24 h. Following 48 h treatment,all the doses of DON showed dermal hyperplasia. TPA treatment, a pos-itive control group, also resulted in an increase in dermal hyperplasiaafter 24 and 48 h treatment.
ED P
RO
OF
tructure of DON. (B) Effect of 24 h treatment on cutaneous edema following topical appli-DON. The skin edemawas determined byweighing 1 cmdiameter punch skin as describedeach group. Each value represents mean ± S.E. of five animals. *P b 0.05, significant withograph of skin isolated from vehicle or DON treated animals for 24 and 48 h (hematoxylinwith control.
skin cell proliferation and inflammation via MAPK pathway, Toxicol.
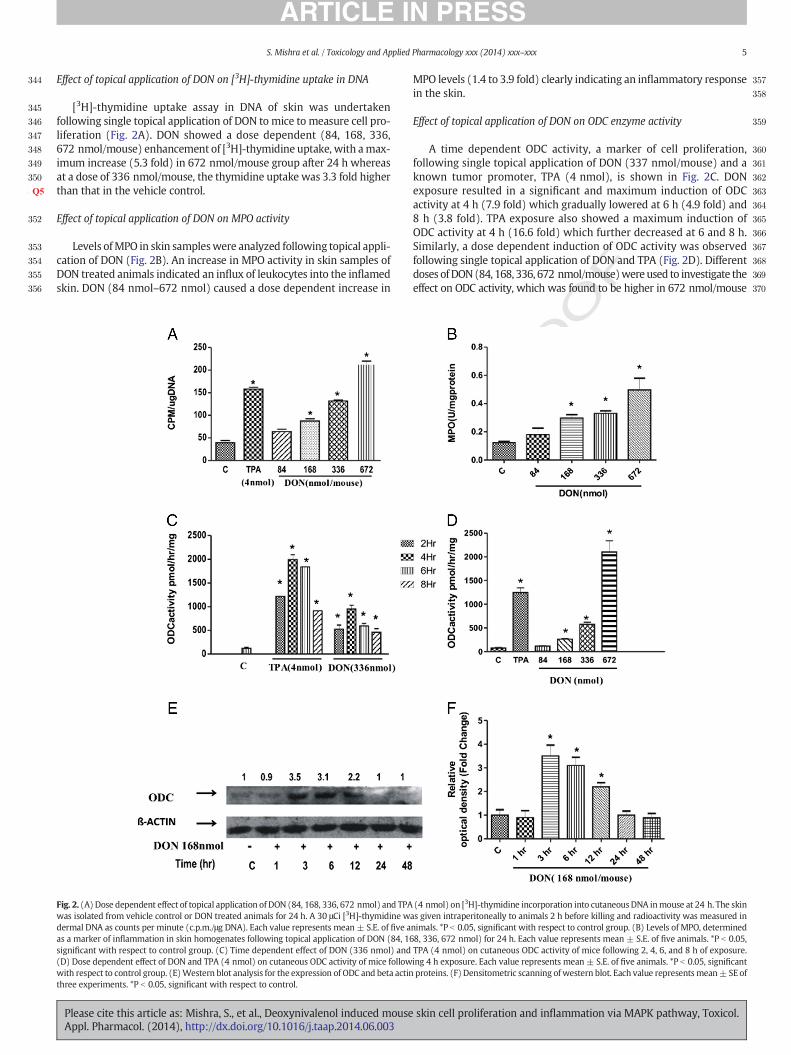
344
345
346
347
348
349
350
351Q5
352
353
354
355
356
357
358
359
360
361
362
363
364
365
366
367
368
369
370
5S. Mishra et al. / Toxicology and Applied Pharmacology xxx (2014) xxx–xxx
Effect of topical application of DON on [3H]-thymidine uptake in DNA
[3H]-thymidine uptake assay in DNA of skin was undertakenfollowing single topical application of DON to mice to measure cell pro-liferation (Fig. 2A). DON showed a dose dependent (84, 168, 336,672 nmol/mouse) enhancement of [3H]-thymidine uptake, with amax-imum increase (5.3 fold) in 672 nmol/mouse group after 24 h whereasat a dose of 336 nmol/mouse, the thymidine uptake was 3.3 fold higherthan that in the vehicle control.
Effect of topical application of DON on MPO activity
Levels ofMPO in skin sampleswere analyzed following topical appli-cation of DON (Fig. 2B). An increase in MPO activity in skin samples ofDON treated animals indicated an influx of leukocytes into the inflamedskin. DON (84 nmol–672 nmol) caused a dose dependent increase in
UNCO
RRECT
Fig. 2. (A) Dose dependent effect of topical application of DON (84, 168, 336, 672 nmol) and TPAwas isolated from vehicle control or DON treated animals for 24 h. A 30 μCi [3H]-thymidine wdermal DNA as counts per minute (c.p.m./μg DNA). Each value represents mean ± S.E. of five aas a marker of inflammation in skin homogenates following topical application of DON (84, 16significant with respect to control group. (C) Time dependent effect of DON (336 nmol) and(D) Dose dependent effect of DON and TPA (4 nmol) on cutaneous ODC activity of mice followwith respect to control group. (E)Western blot analysis for the expression of ODC and beta actinthree experiments. *P b 0.05, significant with respect to control.
Please cite this article as: Mishra, S., et al., Deoxynivalenol induced mouseAppl. Pharmacol. (2014), http://dx.doi.org/10.1016/j.taap.2014.06.003
F
MPO levels (1.4 to 3.9 fold) clearly indicating an inflammatory responsein the skin.
Effect of topical application of DON on ODC enzyme activity
A time dependent ODC activity, a marker of cell proliferation,following single topical application of DON (337 nmol/mouse) and aknown tumor promoter, TPA (4 nmol), is shown in Fig. 2C. DONexposure resulted in a significant and maximum induction of ODCactivity at 4 h (7.9 fold) which gradually lowered at 6 h (4.9 fold) and8 h (3.8 fold). TPA exposure also showed a maximum induction ofODC activity at 4 h (16.6 fold) which further decreased at 6 and 8 h.Similarly, a dose dependent induction of ODC activity was observedfollowing single topical application of DON and TPA (Fig. 2D). Differentdoses ofDON(84, 168, 336, 672 nmol/mouse)wereused to investigate theeffect on ODC activity, which was found to be higher in 672 nmol/mouse
ED P
RO
O
(4 nmol) on [3H]-thymidine incorporation into cutaneousDNA inmouse at 24 h. The skinas given intraperitoneally to animals 2 h before killing and radioactivity was measured innimals. *P b 0.05, significant with respect to control group. (B) Levels of MPO, determined8, 336, 672 nmol) for 24 h. Each value represents mean ± S.E. of five animals. *P b 0.05,TPA (4 nmol) on cutaneous ODC activity of mice following 2, 4, 6, and 8 h of exposure.ing 4 h exposure. Each value represents mean ± S.E. of five animals. *P b 0.05, significantproteins. (F) Densitometric scanning of western blot. Each value representsmean± SE of
skin cell proliferation and inflammation via MAPK pathway, Toxicol.
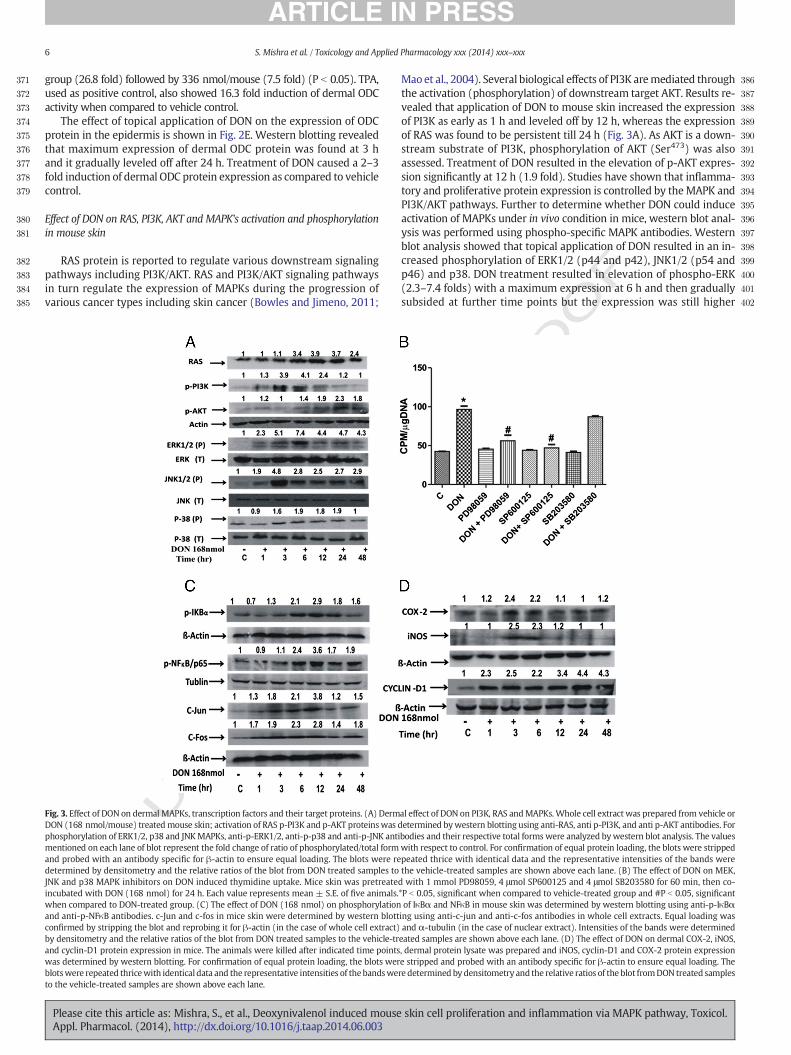
371
372
373
374
375
376
377
378
379
380
381
382
383
384
385
386
387
388
389
390
391
392
393
394
395
396
397
398
399
400
401
402
6 S. Mishra et al. / Toxicology and Applied Pharmacology xxx (2014) xxx–xxx
group (26.8 fold) followed by 336 nmol/mouse (7.5 fold) (P b 0.05). TPA,used as positive control, also showed 16.3 fold induction of dermal ODCactivity when compared to vehicle control.
The effect of topical application of DON on the expression of ODCprotein in the epidermis is shown in Fig. 2E. Western blotting revealedthat maximum expression of dermal ODC protein was found at 3 hand it gradually leveled off after 24 h. Treatment of DON caused a 2–3fold induction of dermal ODC protein expression as compared to vehiclecontrol.
Effect of DON on RAS, PI3K, AKT and MAPK's activation and phosphorylationin mouse skin
RAS protein is reported to regulate various downstream signalingpathways including PI3K/AKT. RAS and PI3K/AKT signaling pathwaysin turn regulate the expression of MAPKs during the progression ofvarious cancer types including skin cancer (Bowles and Jimeno, 2011;
UNCO
RRECT
Fig. 3. Effect of DON on dermal MAPKs, transcription factors and their target proteins. (A) DermDON (168 nmol/mouse) treatedmouse skin; activation of RAS p-PI3K and p-AKT proteinswas dphosphorylation of ERK1/2, p38 and JNKMAPKs, anti-p-ERK1/2, anti-p-p38 and anti-p-JNK antimentioned on each lane of blot represent the fold change of ratio of phosphorylated/total formand probed with an antibody specific for β-actin to ensure equal loading. The blots were redetermined by densitometry and the relative ratios of the blot from DON treated samples toJNK and p38 MAPK inhibitors on DON induced thymidine uptake. Mice skin was pretreateincubated with DON (168 nmol) for 24 h. Each value represents mean ± S.E. of five animals.when compared to DON-treated group. (C) The effect of DON (168 nmol) on phosphorylationand anti-p-NFκB antibodies. c-Jun and c-fos in mice skin were determined by western blottconfirmed by stripping the blot and reprobing it for β-actin (in the case of whole cell extract)by densitometry and the relative ratios of the blot from DON treated samples to the vehicle-trand cyclin-D1 protein expression in mice. The animals were killed after indicated time pointswas determined by western blotting. For confirmation of equal protein loading, the blots werblotswere repeated thricewith identical data and the representative intensities of thebandsweto the vehicle-treated samples are shown above each lane.
Please cite this article as: Mishra, S., et al., Deoxynivalenol induced mouseAppl. Pharmacol. (2014), http://dx.doi.org/10.1016/j.taap.2014.06.003
F
Maoet al., 2004). Several biological effects of PI3K aremediated throughthe activation (phosphorylation) of downstream target AKT. Results re-vealed that application of DON to mouse skin increased the expressionof PI3K as early as 1 h and leveled off by 12 h, whereas the expressionof RAS was found to be persistent till 24 h (Fig. 3A). As AKT is a down-stream substrate of PI3K, phosphorylation of AKT (Ser473) was alsoassessed. Treatment of DON resulted in the elevation of p-AKT expres-sion significantly at 12 h (1.9 fold). Studies have shown that inflamma-tory and proliferative protein expression is controlled by theMAPK andPI3K/AKT pathways. Further to determine whether DON could induceactivation of MAPKs under in vivo condition in mice, western blot anal-ysis was performed using phospho-specific MAPK antibodies. Westernblot analysis showed that topical application of DON resulted in an in-creased phosphorylation of ERK1/2 (p44 and p42), JNK1/2 (p54 andp46) and p38. DON treatment resulted in elevation of phospho-ERK(2.3–7.4 folds) with a maximum expression at 6 h and then graduallysubsided at further time points but the expression was still higher
ED P
RO
O
al effect of DON on PI3K, RAS andMAPKs. Whole cell extract was prepared from vehicle oretermined bywestern blotting using anti-RAS, anti p-PI3K, and anti p-AKT antibodies. Forbodies and their respective total forms were analyzed bywestern blot analysis. The valueswith respect to control. For confirmation of equal protein loading, the blots were strippedpeated thrice with identical data and the representative intensities of the bands werethe vehicle-treated samples are shown above each lane. (B) The effect of DON on MEK,d with 1 mmol PD98059, 4 μmol SP600125 and 4 μmol SB203580 for 60 min, then co-*P b 0.05, significant when compared to vehicle-treated group and #P b 0.05, significantof IκBα and NFκB in mouse skin was determined by western blotting using anti-p-IκBαing using anti-c-jun and anti-c-fos antibodies in whole cell extracts. Equal loading wasand α-tubulin (in the case of nuclear extract). Intensities of the bands were determinedeated samples are shown above each lane. (D) The effect of DON on dermal COX-2, iNOS,, dermal protein lysate was prepared and iNOS, cyclin-D1 and COX-2 protein expressione stripped and probed with an antibody specific for β-actin to ensure equal loading. There determined by densitometry and the relative ratios of the blot fromDON treated samples
skin cell proliferation and inflammation via MAPK pathway, Toxicol.

T
403Q6
404
405
406
407
408
409
410
411
412
413
414
415
416
417
418
419
420
421
422
423
424
425
426Q7
427
428
429
430
431
432
433
434
435
436
437Q8
438
439
440
441
442
443
444
445
446
447
448
449
450
451
452
453
454
455
456
457
458
459
460
461
462
463
464
465
466
467
468
469
470
471
472
473
474
475
476
477
478
479
480
481
482
483
484
485
486
487
488
489
490
491
492
493
494
495
496
497
498
499
500
501
502
503
504
505
506
507
508
509
510
511
512
513
514
515
516
517
518
519
7S. Mishra et al. / Toxicology and Applied Pharmacology xxx (2014) xxx–xxx
UNCO
RREC
than that in the control. However, the expression of phospho-JNK wasenhanced (1.9–4.8 fold) at an early time point of 3 h following DONexposure, which decreased to 2.9 fold that persisted for 48 h. In caseof phospho-p38, an increased expression (0.9–1.9 fold) was foundfrom 1 to 48 h following DON exposure. There was no effect of DONon the total amount of ERK1/2, JNK1/2 and p38 proteins.
MAPK inhibitors suppressed DON induced thymidine uptake
To provide further evidence that MAPKs are involved in DON in-duced cell proliferation, different inhibitors of MAPK pathwayswere topically applied to mice, 1 h prior to DON treatment for 24 hand thymidine uptake in skin DNA was measured. As shown inFig. 3B, pre-treatment with SB203580, a p38 inhibitor, caused only9% reduction of thymidine uptake as compared to the group treatedwith DON alone. On the contrary, PD98059 (a MEK inhibitor ofERK) and SP600125 (JNK inhibitor) caused a significant reductionof thymidine uptake (41 and 51%, respectively), confirming the roleof MAPK pathway in DON induced cell proliferation.
Effect of topical application of DON on phosphorylation of NF-κB and IκBαas well as activation of c-jun and c-fos proteins
Several studies have reported thatMAPKs andAKTplay a critical rolein the activation of NF-κB by phosphorylating NF-κB and IkBαmolecules as well as activating c-jun and c-fos, the subunits of AP-1transcription factor. NF-κB is also involved in the regulation of iNOSand COX-2 expression; and is sequestered in the cytoplasm following itsassociationwith the inhibitory binding protein, IκBα. The phosphorylation,ubiquitination and subsequent degradation of IκBα release NF-κB, whichenters into the nucleus where it induces the expression of iNOS andCOX-2 (Surh and Kundu, 2005). Hence, phosphorylation of NF-κB, andIκBα and expression of c-jun and c-fos proteins were further studiedfollowing topical application of DON. Phosphorylation of IκBα afterDON treatmentwas observed as early as 6 h (2.1 fold) and remained ac-tivated in the skin of mice till 24 h (1.8 fold), whereas phosphorylationof NF-κB (Ser536) was observed at 3 h (1.1 fold) which remained overexpressed till 48 h (1.9 fold) with the maximum induction at 12 h(3.6 fold) (Fig. 3C). Being the downstream target protein of JNK andone of the subunits of AP-1, c-jun was found to be induced after 1 h ofDON exposure and remained enhanced up to 12 h (3.8 fold), however,c-fos protein was over expressed at 1 h (1.7 fold) and remained en-hanced up to 48 h.
DON induced activation of COX-2, iNOS and cyclin-D1 protein expressions
There is evidence that NF-κB regulates a wide variety of genes thatencode proteinswhich are involved in inflammation and carcinogenesis(Baldwin, 2001; Luque and Gelinas, 1997; Rayet and Gelinas, 1999). Ac-tivation of NF-κB can up-regulate the expression of proinflammatorycytokines and inflammatory gene products, such as COX-2 and iNOS(Baldwin, 2001; Rayet and Gelinas, 1999). NF-κB is also regarded as akey regulator of cell proliferation through its direct or indirect effectson cell cycle regulation including its effects on cyclin D1 (Guttridgeet al., 1999). Most of these genes have been shown to be up-regulatedin human cancers, suggesting the role of NF-κB in the development ofcancers including skin cancer (Guttridge et al., 1999). COX-2 and iNOSare important enzymes involved in mediating the inflammation andcell proliferation, whereas over expression of cyclin-D1 induces exces-sive dermal cell proliferation (Herschman, 1994; Saleem et al., 2004).Therefore the effect of topical application of DON on dermal iNOS andCOX-2 protein expression was undertaken. Topical application of DONto mouse skin resulted in an increase in epidermal COX-2 protein ex-pression, which reached to a maximum level at 3 h (2.4-fold) whencompared to control and then declined to basal levels at 6 h. Similarly,the expression of iNOS peaked at 3 h following DON treatment and
Please cite this article as: Mishra, S., et al., Deoxynivalenol induced mouseAppl. Pharmacol. (2014), http://dx.doi.org/10.1016/j.taap.2014.06.003
further declined to basal level at 6 h post DON treatment (Fig. 3D).Cyclin-D1 protein expression was also found to be over expressed(2.3–4.3-fold) during 1–48 h of DON treatment.
PRO
OF
Evaluation of skin tumor promoting property of DON
As observed in the previous sections that single topical application ofDON (84–672 nmol) could induce the cell proliferative markers as wellas inflammatory markers in mouse skin, it was of importance to evalu-ate the skin tumor promoting property of DON inmice. Doses of 84–168nmol of DONwere chosen for assessing skin tumor promoting propertyas significant enhancement of cell proliferativemarkerswas observed atthe tested doses. A single topical application of DMBA as tumor initiatorfollowed by twice-weekly application of TPA as a tumor promoter re-sulted in the development of skin papillomas. However, no tumorswere detected in animals given either single initiating dose of DMBAfollowed by twice weekly application of DON (84 nmol or 168 nmol/mouse), or vehicle control group treated with acetone twice weekly ora single dose of acetone followed by twice weekly topical applicationof DON (84 nmol or 168 nmol/mouse). The first incidence of tumori-genesis in the positive control group treated with DMBA/TPA occurredin the 7th week. The cumulative number of tumors in the DMBA/TPAtreated group was 59 after 24 weeks of exposure (Fig. 4A). The weeklydata of percent mice with tumors during the treatment schedule ispresented in Fig. 4B. A hundred percent of the mice developed tumorsin DMBA/TPA treated group by the 16th week of treatment (Fig. 4B).
EDHistopathological analysis of skin and intestine exposed to DON
Fig. 4C depicts the histopathological features of skin isolated fromvehicle control, DMBA/DON (84 nmol or 168 nmol/mouse) treatedmice. Control mice showed normal appearance of epidermis, dermisand hypodermis of skin. The underlying muscle layer was also visiblein the control animals. In the positive control group, where animalswere treated with DMBA/TPA, characteristic skin lesions includingpapillomatous growth, comprising of squamous cell carcinoma andpearl formation were observed. The skin also showed hyperkeratosisalongwith hyperplasia of squamous cells protruding out of the epidermis(data not shown). The skin of mice treated with both the doses of DON(84 nmol and 168 nmol/mouse) in DMBA/DON treated group apparentlydemonstrated an increase in adipose tissue, infiltration of inflammatorycells along with extensive inflammation in the hair follicle, hypertrophyof sebaceous gland as well as characteristic inflammation in the skin.
Intestine is one of the primary target tissues of DON toxicity (Pestkaet al., 2004). Peyer's patches are considered to be the immune sensors ofintestine and have been shown to be enhanced in number as well as insize during intestinal toxicity. Fig. 4D shows morphological changes inthe intestine of DON treated animals with enlarged Peyer's patches ascompared to vehicle control. Histopathological study revealed lym-phoid hyperplasia as well as submucosal lymphoid hyperplasia alongwith destruction of submucosal villi in the intestine of animals topicallytreated with DON (84 nmol or 168 nmol/mouse) (Fig. 4E).
Effect of MPO levels on skin and intestine
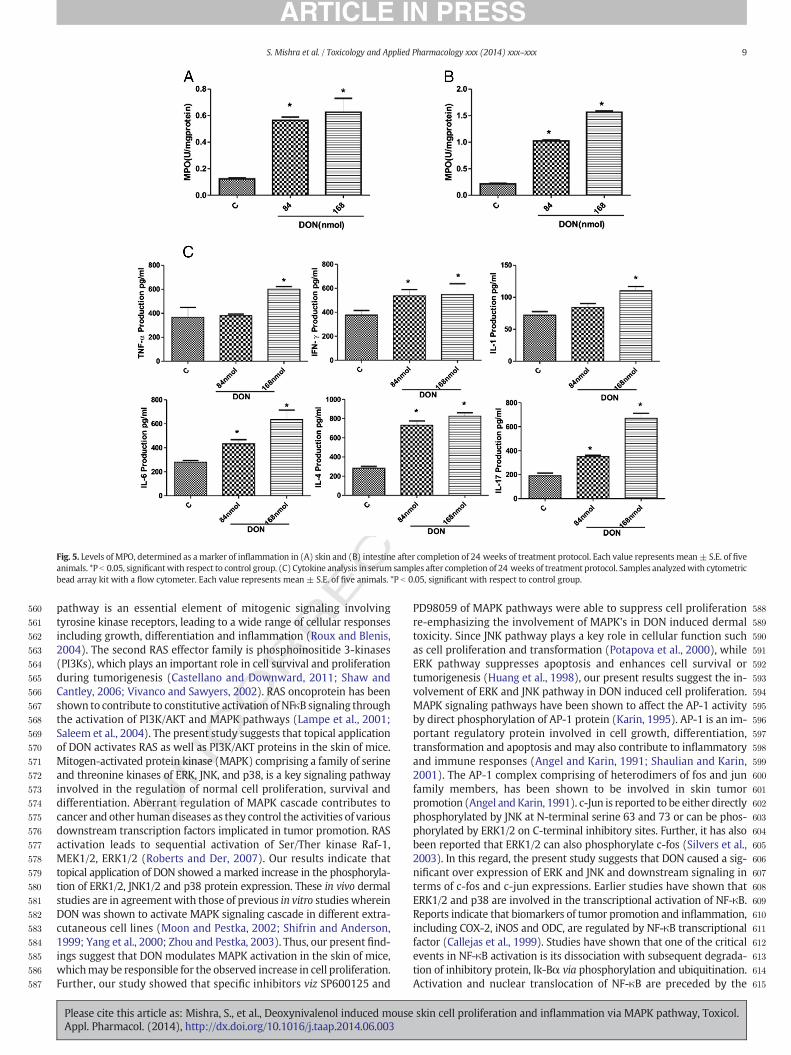
Neutrophil infiltration plays a central role in the inflammation and isconsidered to be a major cause of tissue damage. MPO, a marker en-zyme of infiltrating leukocytes (macrophages and neutrophils) is usedas a parameter for various inflammatory processes in target tissues in-cluding the skin (Bradley et al., 1982). Twice weekly topical applicationof DON for 24 weeks to DMBA initiated animals resulted in significantenhancement ofMPO levels not only in the skin but also in the intestine(Figs. 5A and B).
skin cell proliferation and inflammation via MAPK pathway, Toxicol.
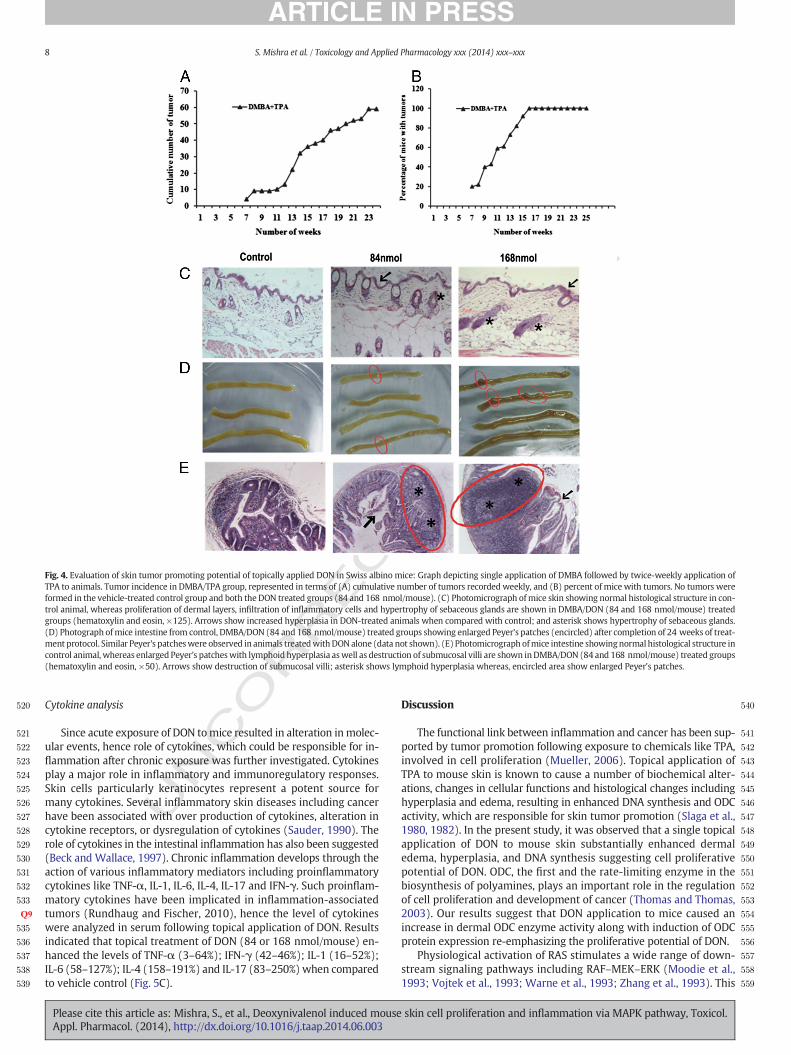
RRECTED P
RO
OF
520
521
522
523
524
525
526
527
528
529
530
531
532
533
534Q9
535
536
537
538
539
540
541
542
543
544
545
546
547
548
549
550
551
552
553
554
555
556
557
558
559
Fig. 4. Evaluation of skin tumor promoting potential of topically applied DON in Swiss albino mice: Graph depicting single application of DMBA followed by twice-weekly application ofTPA to animals. Tumor incidence in DMBA/TPA group, represented in terms of (A) cumulative number of tumors recorded weekly, and (B) percent of mice with tumors. No tumors wereformed in the vehicle-treated control group and both the DON treated groups (84 and 168 nmol/mouse). (C) Photomicrograph ofmice skin showing normal histological structure in con-trol animal, whereas proliferation of dermal layers, infiltration of inflammatory cells and hypertrophy of sebaceous glands are shown in DMBA/DON (84 and 168 nmol/mouse) treatedgroups (hematoxylin and eosin, ×125). Arrows show increased hyperplasia in DON-treated animals when compared with control; and asterisk shows hypertrophy of sebaceous glands.(D) Photograph ofmice intestine from control, DMBA/DON (84 and 168 nmol/mouse) treated groups showing enlarged Peyer's patches (encircled) after completion of 24 weeks of treat-ment protocol. Similar Peyer's patcheswere observed in animals treatedwithDONalone (data not shown). (E) Photomicrograph ofmice intestine showingnormal histological structure incontrol animal, whereas enlarged Peyer's patcheswith lymphoid hyperplasia aswell as destruction of submucosal villi are shown inDMBA/DON (84 and 168 nmol/mouse) treated groups(hematoxylin and eosin, ×50). Arrows show destruction of submucosal villi; asterisk shows lymphoid hyperplasia whereas, encircled area show enlarged Peyer's patches.
8 S. Mishra et al. / Toxicology and Applied Pharmacology xxx (2014) xxx–xxx
UNCOCytokine analysis
Since acute exposure of DON tomice resulted in alteration inmolec-ular events, hence role of cytokines, which could be responsible for in-flammation after chronic exposure was further investigated. Cytokinesplay a major role in inflammatory and immunoregulatory responses.Skin cells particularly keratinocytes represent a potent source formany cytokines. Several inflammatory skin diseases including cancerhave been associated with over production of cytokines, alteration incytokine receptors, or dysregulation of cytokines (Sauder, 1990). Therole of cytokines in the intestinal inflammation has also been suggested(Beck and Wallace, 1997). Chronic inflammation develops through theaction of various inflammatory mediators including proinflammatorycytokines like TNF-α, IL-1, IL-6, IL-4, IL-17 and IFN-γ. Such proinflam-matory cytokines have been implicated in inflammation-associatedtumors (Rundhaug and Fischer, 2010), hence the level of cytokineswere analyzed in serum following topical application of DON. Resultsindicated that topical treatment of DON (84 or 168 nmol/mouse) en-hanced the levels of TNF-α (3–64%); IFN-γ (42–46%); IL-1 (16–52%);IL-6 (58–127%); IL-4 (158–191%) and IL-17 (83–250%) when comparedto vehicle control (Fig. 5C).
Please cite this article as: Mishra, S., et al., Deoxynivalenol induced mouseAppl. Pharmacol. (2014), http://dx.doi.org/10.1016/j.taap.2014.06.003
Discussion
The functional link between inflammation and cancer has been sup-ported by tumor promotion following exposure to chemicals like TPA,involved in cell proliferation (Mueller, 2006). Topical application ofTPA to mouse skin is known to cause a number of biochemical alter-ations, changes in cellular functions and histological changes includinghyperplasia and edema, resulting in enhanced DNA synthesis and ODCactivity, which are responsible for skin tumor promotion (Slaga et al.,1980, 1982). In the present study, it was observed that a single topicalapplication of DON to mouse skin substantially enhanced dermaledema, hyperplasia, and DNA synthesis suggesting cell proliferativepotential of DON. ODC, the first and the rate-limiting enzyme in thebiosynthesis of polyamines, plays an important role in the regulationof cell proliferation and development of cancer (Thomas and Thomas,2003). Our results suggest that DON application to mice caused anincrease in dermal ODC enzyme activity along with induction of ODCprotein expression re-emphasizing the proliferative potential of DON.
Physiological activation of RAS stimulates a wide range of down-stream signaling pathways including RAF–MEK–ERK (Moodie et al.,1993; Vojtek et al., 1993; Warne et al., 1993; Zhang et al., 1993). This
skin cell proliferation and inflammation via MAPK pathway, Toxicol.
CTED P
RO
OF
560
561
562
563
564
565
566
567
568
569
570
571
572
573
574
575
576
577
578
579
580
581
582
583
584
585
586
587
588
589
590
591
592
593
594
595
596
597
598
599
600
601
602
603
604
605
606
607
608
609
610
611
612
613
614
615
Fig. 5. Levels of MPO, determined as a marker of inflammation in (A) skin and (B) intestine after completion of 24 weeks of treatment protocol. Each value represents mean± S.E. of fiveanimals. *P b 0.05, significantwith respect to control group. (C) Cytokine analysis in serum samples after completion of 24 weeks of treatment protocol. Samples analyzedwith cytometricbead array kit with a flow cytometer. Each value represents mean ± S.E. of five animals. *P b 0.05, significant with respect to control group.
9S. Mishra et al. / Toxicology and Applied Pharmacology xxx (2014) xxx–xxx
UNCO
RREpathway is an essential element of mitogenic signaling involving
tyrosine kinase receptors, leading to a wide range of cellular responsesincluding growth, differentiation and inflammation (Roux and Blenis,2004). The second RAS effector family is phosphoinositide 3-kinases(PI3Ks), which plays an important role in cell survival and proliferationduring tumorigenesis (Castellano and Downward, 2011; Shaw andCantley, 2006; Vivanco and Sawyers, 2002). RAS oncoprotein has beenshown to contribute to constitutive activation of NFκB signaling throughthe activation of PI3K/AKT and MAPK pathways (Lampe et al., 2001;Saleem et al., 2004). The present study suggests that topical applicationof DON activates RAS as well as PI3K/AKT proteins in the skin of mice.Mitogen-activated protein kinase (MAPK) comprising a family of serineand threonine kinases of ERK, JNK, and p38, is a key signaling pathwayinvolved in the regulation of normal cell proliferation, survival anddifferentiation. Aberrant regulation of MAPK cascade contributes tocancer and other humandiseases as they control the activities of variousdownstream transcription factors implicated in tumor promotion. RASactivation leads to sequential activation of Ser/Ther kinase Raf-1,MEK1/2, ERK1/2 (Roberts and Der, 2007). Our results indicate thattopical application of DON showed a marked increase in the phosphoryla-tion of ERK1/2, JNK1/2 and p38 protein expression. These in vivo dermalstudies are in agreementwith those of previous in vitro studies whereinDON was shown to activate MAPK signaling cascade in different extra-cutaneous cell lines (Moon and Pestka, 2002; Shifrin and Anderson,1999; Yang et al., 2000; Zhou and Pestka, 2003). Thus, our present find-ings suggest that DON modulates MAPK activation in the skin of mice,whichmay be responsible for the observed increase in cell proliferation.Further, our study showed that specific inhibitors viz SP600125 and
Please cite this article as: Mishra, S., et al., Deoxynivalenol induced mouseAppl. Pharmacol. (2014), http://dx.doi.org/10.1016/j.taap.2014.06.003
PD98059 of MAPK pathways were able to suppress cell proliferationre-emphasizing the involvement of MAPK's in DON induced dermaltoxicity. Since JNK pathway plays a key role in cellular function suchas cell proliferation and transformation (Potapova et al., 2000), whileERK pathway suppresses apoptosis and enhances cell survival ortumorigenesis (Huang et al., 1998), our present results suggest the in-volvement of ERK and JNK pathway in DON induced cell proliferation.MAPK signaling pathways have been shown to affect the AP-1 activityby direct phosphorylation of AP-1 protein (Karin, 1995). AP-1 is an im-portant regulatory protein involved in cell growth, differentiation,transformation and apoptosis and may also contribute to inflammatoryand immune responses (Angel and Karin, 1991; Shaulian and Karin,2001). The AP-1 complex comprising of heterodimers of fos and junfamily members, has been shown to be involved in skin tumorpromotion (Angel and Karin, 1991). c-Jun is reported to be either directlyphosphorylated by JNK at N-terminal serine 63 and 73 or can be phos-phorylated by ERK1/2 on C-terminal inhibitory sites. Further, it has alsobeen reported that ERK1/2 can also phosphorylate c-fos (Silvers et al.,2003). In this regard, the present study suggests that DON caused a sig-nificant over expression of ERK and JNK and downstream signaling interms of c-fos and c-jun expressions. Earlier studies have shown thatERK1/2 and p38 are involved in the transcriptional activation of NF-κB.Reports indicate that biomarkers of tumor promotion and inflammation,including COX-2, iNOS and ODC, are regulated by NF-κB transcriptionalfactor (Callejas et al., 1999). Studies have shown that one of the criticalevents in NF-κB activation is its dissociation with subsequent degrada-tion of inhibitory protein, Ik-Bα via phosphorylation and ubiquitination.Activation and nuclear translocation of NF-κB are preceded by the
skin cell proliferation and inflammation via MAPK pathway, Toxicol.

T
616
617
618
619
620
621
622
623
624
625
626
627
628
629
630
631
632
633
634
635
636
637
638
639
640
641
642
643
644
645
646
647
648
649
650
651
652
653
654
655
656
657
658
659
660
661
662
663
664
665
666
667
668
669
670
671
672
673
674
675
676
677
678
679
680
681
682
683
684
685
686
687
688
689
10 S. Mishra et al. / Toxicology and Applied Pharmacology xxx (2014) xxx–xxx
C
phosphorylation and proteolytic degradation of Ik-Bα (Israel, 1995).Evidence suggests that proteins from NF-κB family have an importantregulatory role in inflammation, cell proliferation and oncogenesis andthat they control the expression of several growth factors, oncogenesand tumor suppressor genes (Thanos and Maniatis, 1995). The presentstudy demonstrates that topical application of DON to mice resulted inthe activation and nuclear translocation of NF-κB along with thephosphorylation of Ik-Bα in the skin indicating the involvement ofNF-κB protein in dermal toxicity of DON.
The transcription factors, NF-κB and AP-1 enter the nucleus wherethey bind to the promoter region of various target genes includingcyclin-D1 and COX-2 (Surh and Kundu, 2005). Our present study sug-gests that application of DON caused induction of iNOS and COX-2 pro-teins in the skin, thereby indicating the inflammatory property of DON.Earlier studies suggest that RAS-mediated tumorigenesis depends onsignaling pathways that act preferentially through cyclin D1. Cyclin D1expression and the activity of its associated kinase have been shownto be up-regulated in keratinocytes in response to oncogenic RAS(Peters, 1994). Studies in the present investigation suggest that DONup-regulated the expression of cyclin D1 protein, which may be due tothe activation of MAPK cascade and thereby resulting in activation ofthe cyclin D1 promoter region of AP1.
Since all the short-term markers of cell proliferation andinflammation including ODC, [3H]-thymidine uptake, hyperplasia,elevated expression of MAPKs, transcription factors NF-κB, c-jun,c-fos as well as proteins cyclin-D1, iNOS and COX-2 were found tobe elevated following single topical application of DON, it was necessaryto investigate the tumor promoting potential of DON. In a two-stagemouse skin tumorigenesis protocol, twice weekly topical applicationof DON (84 and 168 nmol/mouse) toDMBA initiatedmice did not resultin the development of tumors. However, histopathological studiessuggest characteristic hypertrophy of sebaceous gland and infiltrationof inflammatory cells in the skin indicating dermal lesions in DMBA/DON treated groups. Further, it has been observed for the first timethat long term topical treatment of DON showed enlargement of Peyer'spatches in the intestine when compared to vehicle control animals.Earlier studies have shown that there is a rapid absorption of DON in
UNCO
RRE
Fig. 6. Mechanism of cell proliferation and inflammati
Please cite this article as: Mishra, S., et al., Deoxynivalenol induced mouseAppl. Pharmacol. (2014), http://dx.doi.org/10.1016/j.taap.2014.06.003
ED P
RO
OF
the upper gastrointestinal tract, suggesting intestine as the primarytarget organ of DON toxicity (Goyarts and Da¨nicke, 2006). Previousstudies have reported that dietary vomitoxin exposure enhances IgA+andCD4+cells and T-cell proliferation and/or differentiation in Peyer'spatch (Pestka et al., 1990). Furthermore, organs related to immunesystem appear to be another target of DON (Pestka, 2008), whichmight be the reason for the observed enlargement in Peyer's patchesinduced by topical application of DON thereby causing enhancedinflammatory cytokines; TNF-α, IFN-γ, IL-6, IL-1, IL-5, and IL-4. Tumorpromoters are also known to induce TNF-α gene expression in theirtarget organs, thereby enhancing the production of TNF-α, whichactivates IL-1 (Fujiki et al., 1997; Lee et al., 1993). Proinflammatorycytokines such as IL-1, IL-6, IL-17 and IFN play important roles in theinflammation. Our present studies indicate that MPO levels wereincreased in the skin and intestine of mice topically treated with DONsuggesting infiltration of inflammatory cells, which may be one of thereasons for the observed increase in Peyer's patches. In this regard,studies have shown that as the inflammation progresses various typesof leukocytes, lymphocytes and other inflammatory cells are activatedand attracted to the inflamed site by a signaling network involving anumber of growth factors, cytokines, and chemokines (Lu et al., 2006).Even though DON did not act as a tumor promoter at the doses tested,however, it showed induction of cell proliferation aswell as inflammationin the epidermis, which are supposed to be the key factors for tumor pro-motion. Although the results of cytokine induction by topical applicationof DON suggest its inflammatory potential, nonetheless, it can be arguedthat the concentration of DONmay not be at the threshold levels to elicittumor promotion at the tested doses (84 and 168 nmol). Our presentstudy is in agreementwith that of Lambert et al. (1995) where no tumorswere produced in any group promotedwithmultiple applications of DONat the dose of 25 and 50 μg/animal in mice. The fact that DON could notact as a tumor promoter at the doses 84–168 nmol may be due to thefact that TPA, the most powerful tumor promoting agent, at a very lowdose of 0.16 nmol caused an increase in DNA synthesis and mitoticactivity but does not have promoting action, suggesting that cellularevents other than cell proliferation or hyperplasia are required to enablethe expression of the neoplastic phenotype (Raick, 1974). Thus, it is
on induced by topical application of DON to mice.
skin cell proliferation and inflammation via MAPK pathway, Toxicol.

T
690
691
692
693
694
695
696
697
698
699Q10
700
701
702
703
704Q11
705
706
707
708
709
710
711
712
713
714
715
716
717
718719720721722723724725726727728729730731732733734735736737738739740741742743744745746747748749750751752753754755
756757758759760761762763764765766767768769770771772773774775776777778779780781782783784785786787788789790791792793794795796797798799800801802803804805806807808809810811812813814815816817818819820821822823824825826827828829830831832833834835836837838839840841
11S. Mishra et al. / Toxicology and Applied Pharmacology xxx (2014) xxx–xxx
UNCO
RREC
clear that thoughDONmaynot act as a tumor promoter at the testeddose(84 and 168 nmol), it is quite likely that prolonged exposure to DON at ahigher dosemay induce tumor promotion and cause further toxicologicalmanifestations.
In conclusion our result suggests that topical application of DON tomice results in increased cell proliferation, DNA synthesis, andinflammationwhich aremediated through PI3K/AKT andMAPK signalingpathways involving transcription factor NFκB and AP-1, further leading totranscriptional activation of downstream target proteins c-fos, c-jun,cyclin D1, iNOS and cox-2 (Fig. 6). The present findings are of relevancetowards the health risks of population where dermal exposure mayoccur especially in agricultural fields.
Conflicts of interest
The authors declare that they have no conflicts of interest.
Uncited references
Degen, 2011Derijard et al., 1994Hanahan and Weinberg, 2000Hibi et al., 1993WHO Food Additives Series: 47, 1998
Acknowledgments
We are grateful to the director of our institute for his keen interest inthis present study. One of us (S.M.) is thankful to the University GrantCommission (UGC), New Delhi for the award of Senior ResearchFellowship. Financial assistance from the CSIR network project no.NWP-17 is duly acknowledged. This manuscript is IITR communication# 3218.
References
Afaq, F., Adhami, V.M., Ahmad, N., Mukhtar, H., 2003. Suppression of UVB-induced phos-phorylation ofmitogen-activated protein kinases and nuclear factor kappa B by greentea polyphenol in SKH-1 hairless mice. Oncogene 22, 9254–9264.
Ahmad, N., Gilliam, A.C., Katiyar, S.K., O'Brien, T.G., Mukhtar, H., 2001. A definitive role ofornithine decarboxylase in photocarcinogenesis. Am. J. Pathol. 159, 885–892.
Almeida, M.I., Almeida, N.G., Carvalho, K.L., Gonçalves, G.A., Silva, C.N., Santos, E.A., Garcia,J.C., Vargas, E.A., 2012. Co-occurrence of aflatoxins B1, B2, G1 and G2, ochratoxin A,zearalenone, deoxynivalenol, and citreoviridin in rice in Brazil. Food Addit. Contam.Part A: Chem. Anal. Control Expo. Risk Assess. 294, 694–703.
Angel, P., Karin, M., 1991. The role of Jun, Fos and the AP-1 complex in cell-proliferationand transformation. Biochim. Biophys. Acta 1072, 129–157.
Baldwin, A.S., 2001. Control of oncogenesis and cancer therapy resistance by thetranscription factor NF-κB. J. Clin. Invest. 107, 241–246.
Beck, P.L., Wallace, J.L., 1997. Cytokines in inflammatory bowel disease. Mediat. Inflamm.2, 95–103.
Bennett, J.W., Klich, M., 2003. Mycotoxins. Clin. Microbiol. Rev. 497–516.Bhat, R.V., Sashidhar, R.B., Ramakrishna, Y., Munshi, K.L., 1989. Outbreak of trichothecene
mycotoxicoses associated with consumption of mould damaged wheat products inKashmir valley, India. Lancet I, 35–37.
Boonen, J., Malysheva, S.V., Taevernier, L., Diana, Di, Mavungu, J., DeSaeger, S.,Despiegeleer, B., 2012. Human skin penetration of selected model mycotoxins.Toxicology 301, 21–32.
Bowles, D.W., Jimeno, A., 2011. New phosphatidylinositol 3-kinase inhibitors for cancer.Expert. Opin. Investig. Drugs 20, 507–518.
Bradley, P.P., Priebat, D.A., Christensen, R.D., Rothstein, G., 1982. Measurement of cutaneousinflammation: estimation of neutrophil content with an enzyme marker. J. Investig.Dermatol. 78, 206–209.
Callejas, N.A., Casado,M., Bosca, L., Martin-Sanz, P., 1999. Requirement of nuclear factor κBfor the constitutive expression of nitric oxide synthase-2 and cyclooxygenase-2 in rattrophoblasts. J. Cell Sci. 18, 3147–3155.
Canady, R.A., Coker, R.D., Rgan, S.K., Krska, R., Kuiper-Goodman, T., Olsen, M., Pestka, J.J.,Resnik, S., Schlatter, J., 2001. Deoxynivalenol. Safety evaluation of certain mycotoxinsin food. Fifty-sixth report of the Joint FAO/WHO Expert Committee on Food Additives.WHO Food Additives Series, 47. International Programme on Chemical Safety—WorldHealth Organization, Geneva, pp. 420–555.
CAST, 2003. Potential economic costs of mycotoxins in the United States. Mycotoxins:Risks in Plant, Animal, and Human Systems. Task Force Report No. 139. Council forAgricultural Science and Technology, Ames, IA, pp. 136–142.
Please cite this article as: Mishra, S., et al., Deoxynivalenol induced mouseAppl. Pharmacol. (2014), http://dx.doi.org/10.1016/j.taap.2014.06.003
ED P
RO
OF
Castellano, E., Downward, J., 2011. Role of RAS in the regulation of PI 3-kinase. Curr. Top.Microbiol. Immunol. 346, 143–169.
Chun, K.S., Kang, J.Y., Kim, O.H., Kang, H., Surh, Y.J., 2002. Effects of yakuchinone A andyakuchinone B on the phorbol ester-induced expression of COX-2 and iNOS andactivation of NF-κB in mouse skin. J. Environ. Pathol. Toxicol. Oncol. 21, 131–139.
Degen, G.H., 2011. Tools for investigating workplace-related risks from mycotoxinexposure. World Mycotoxin J. 4, 315–327.
Derijard, B., Hibi, M.,Wu, I.H., Barrett, T., Su, B., Deng, T., Karin, M., Davis, R.J., 1994. JNK1: aprotein kinase stimulated by UV light and Ha-RAS that binds and phosphorylates thec-Jun activation domain. Cell 76, 1025–1037.
Edwards, S.G., Dickin, E.T., MacDonald, S., Buttler, D., Hazel, C.M., Patel, S., Scudamore, K.A.,2011. Distribution of Fusarium mycotoxins in UK wheat mill fractions. Food Addit.Contam. Part A: Chem. Anal. Control Expo. Risk Assess. 28 (12), 1694–1704.
Einspahr, J.G., Bowden, G.T., Alberts, D.S., 2003. Skin cancer chemoprevention: strategiesto save our skin. Recent Results Cancer Res. 163, 151–164.
Fujiki, H., Sueoka, E., Komori, A., Suganuma, M., 1997. Tumor promotion and TNF-a geneexpression by the okadaic acid class tumor promoters. Environ. Carcinog. Ecotoxicol.Rev. 15, 1–40.
Goyarts, T., Da¨nicke, S., 2006. Bioavailability of the Fusarium toxin deoxynivalenol (DON)from naturally contaminated wheat for the pig. Toxicol. Lett. 163, 171–182.
Gupta, K.P., Mehrotra, N.K., 1992. Status of ornithine decarboxylase activity and DNAsynthesis in mancozeb-exposed mouse skin. Carcinogenesis 13, 131–133.
Guttridge, D.C., Albanese, C., Reuther, J.Y., Pestell, R.G., Baldwin, A.S., 1999. NF-nB controlscell growth and differentiation through transcriptional regulation of cyclin D1. Mol.Cell. Biol. 19, 5785–5799.
Hanahan, D., Weinberg, R.A., 2000. The hallmarks of cancer. Cell 100, 57–70.Herschman, H.R., 1994. Regulation of prostaglandin synthase-1 and prostaglandin
synthase-2. Cancer Metastasis Rev. 13, 241–256.Hibi, M., Lin, A., Smeal, T., Minden, A., Karin, M., 1993. Identification of an oncoprotein-
and UV-responsive protein kinase that binds and potentiates the c-jun activationdomain. Genes Dev. 7, 2135–2148.
Hsia, C.C., Wu, J.L., Lu, X.Q., Li, Y.S., 1988. Natural occurrence and clastogenic effects ofnivalenol, deoxynivalenol, 3-acetyl-deoxynivalenol, 15-acetyl-deoxynivalenol, andzearalenone in corn from a high-risk area of esophageal cancer. Cancer Detect.Prev. 13, 79–86.
Huang, C., Ma, W.Y., Young, M.R., Colburn, N., Dong, Z., 1998. Shortage of mitogen-activated protein kinase is responsible for resistance to AP-1 transactivation andtransformation in mouse JB6 cells. Proc. Natl. Acad. Sci. U. S. A. 95, 156–161.
Huang, X.H., Zhang, X.H., Li, Y.H., Wang, J.L., Yan, X., Xing, L.X., Wang, F.R., 2004. Carcino-genic effects of sterigmatocystin and deoxynivalenol in NIH mice. Chin. J. Oncol. 26(12), 705–708.
IARC (International Agency for Research on Cancer), 1993. Monographs on the evaluationof carcinogenic risks to humans; some naturally occurring substances, food items andconstituents. Heterocyclic Aromatic Amines and Mycotoxins, International Agencyfor Research on Cancer, World Health Organization, Lyon, France. 56, pp. 397–444.
Israel, A., 1995. A role for phosphorylation and degradation in the control of NF-kappa Bactivity. Trends Genet. 11, 203–205.
Iverson, F., Armstrong, C., Nera, E., Truelove, J., Fernie, S., Scott, P., Stapley, R., Hayward, S.,Gunner, S., 1995. Chronic feeding study of deoxynivalenol in B6C3F1male and femalemice. Teratog. Carcinog. Mutagen. 15, 283–306.
JECFA, 2010. Joint Food and Agriculture Organization/World Health Organization ExpertCommittee on Food Additives. Joint FAO/WHO Expert Committee on Food AdditivesSeventy-second Meeting, Rome, 16–25 February 2010: Summary and Conclusions(http://www.who.int/foodsafety/chem/summary72_rev.pdf).
Karin, M., 1995. The regulation of AP-1 activity by mitogen-activated protein kinases. J.Biol. Chem. 270, 16483–16486.
Katiyar, S.K., Rupp, C.O., Korman, N.J., Agarwal, R., Mukhtar, H., 1995. Inhibition of 12-O-tetradecanoylphorbol-13-acetate and other skin tumor-promoter-caused inductionof epidermal interleukin-1 alpha mRNA and protein expression in SENCAR mice bygreen tea polyphenols. J. Investig. Dermatol. 105, 394–398.
Katiyar, S.K., Agarwal, R., Mukhtar, H., 1996. Inhibition of tumor promotion in SENCARmouse skin by ethanol extract of Zingiber officinale rhizome. Cancer Res. 56,1023–1030.
Kumar, R., Dwivedi, P.D., Dhawan, A., Das, M., Ansari, K.M., 2011. Citrinin generatedreactive oxygen species cause cell cycle arrest leading to apoptosis via the intrinsicmitochondrial pathway in mouse skin. Toxicol. Sci. 122, 557–566.
Kumar, R., Ansari, K.M., Chaudhari, B.P., Dhawan, A., Dwivedi, P.D., Jain, S.K., Das, M., 2012.Topical application of ochratoxin A causes DNA damage and tumor initiation inmouse skin. PLoS One 7, e47280.
Lambert, L.A., Hines, F.A., Eppley, R.M., 1995. Lack of initiation and promotion potential ofdeoxynivalenol for skin tumorigenesis in Sencar mice. Food Chem. Toxicol. 33,217–222.
Lampe, E., Heinrich, M., Walczak, H., Kalthoff, H., 2001. CD95 and TRAIL receptor-mediated activation of protein kinase C and NF-kappaB contributes to apoptosisresistance in ductal pancreatic adenocarcinoma cells. Oncogene 20, 4258–4269.
Lee, W.Y., Fischer, S.M., Butler, A.P., Locniskar, M.F., 1993. Modulation of interleukin-1amRNA expression in mouse epidermis by tumor promoters and antagonists. Mol.Carcinog. 7, 26–35.
Lee, T., Lee, S.H., Lee, S.H., Shin, J.Y., Yun, J.C., Lee, Y.W., Ryu, J.G., 2011. Occurrence ofFusarium mycotoxins in rice and its milling by-products in Korea. J. Food Prot. 74,1169–1174.
Li, F.Q., Luo, X.Y., Yoshizawa, T., 1999. Mycotoxins (trichothecenes, zearalenone andfumonisins) in cereals associated with human red-mold intoxications stored since1989 and 1991 in China. Nat. Toxins 7, 93–97.
Lu, H., Ouyang, W., Huang, C., 2006. Inflammation, a key event in cancer development.Mol. Cancer Res. 4, 221–233.
skin cell proliferation and inflammation via MAPK pathway, Toxicol.

T
842843844845846847848849850851852853854855856857858859860861862863864865866867868869870871872873874875876877878879880881882883884885886887888889890891892893894895896897898899900
901902903904905906907908909910911912913914915916917918919920921922923924925926927928929930931932933934935936937938939940941942943944945946947948949950951952953954955956957958959
12 S. Mishra et al. / Toxicology and Applied Pharmacology xxx (2014) xxx–xxx
RREC
Luo, Y., Yoshizawa, T., Katayama, T., 1990. Comparative study on the natural occurrence ofFusariummycotoxins (trichothecenes and zearalenone) in corn and wheat from highand low-risk areas for esophageal cancer in China. Appl. Environ. Microbiol. 56,3723–3726.
Luque, I., Gelinas, C., 1997. Rel/NF-κB and IkB factors in oncogenesis. Semin. Cancer Biol. 8,103–111.
Ma, Y.Y., Guo, H.W., 2008. Mini-review of studies on the carcinogenicity of deoxynivalenol.Environ. Toxicol. Pharmacol. 25, 1–9.
Mao, J.H., To, M.D., Losada, J.P., Wu, D., Rosario, R.D., Balmain, A., 2004. Mutually exclusivemutations of the Pten and RAS pathways in skin tumor progression. Genes Dev. 18,1800–1805.
Mishra, S., Ansari, K.M., Dwivedi, P.D., Pandey, H.P., Das, M., 2013. Occurrence ofdeoxynivalenol in cereals and exposure risk assessment in Indian population. FoodControl 30, 549–555.
Moodie, S.A., Willumsen, B.M., Weber, M.J., Wolfman, A., 1993. Complexes of RAS.GTPwith Raf-1 and mitogen-activated protein kinase kinase. Science 260, 1658–1661.
Moon, Y., Pestka, J.J., 2002. Vomitoxin-induced cyclooxygenase-2 gene expression inmacrophages mediated by activation of ERK and p38 but not JNK mitogen activatedprotein kinases. Toxicol. Sci. 69, 373–382.
Mueller, M., 2006. Inflammation in epithelial skin tumours: old stories and new ideas.Eur. J. Cancer 42, 735–744.
Pestka, J.J., 2008. Mechanisms of deoxynivalenol-induced gene expression and apoptosis.Food Addit. Contam. 25, 1128–1140.
Pestka, J.J., Smolinski, A.T., 2005. Deoxynivalenol toxicity and potential effects on humans.J. Toxicol. Environ. Health B 8, 39–69.
Pestka, J.J., Dong, W., Warner, R.L., Rasooly, L., Bond, G.S., 1990. Effect of dietary adminis-tration of the trichothecene vomitoxin (deoxynivalenol) on IgA and IgG secretion byPeyer's patch and splenic lymphocytes. Food Chem. Toxicol. 28, 693–699.
Pestka, J.J., Zhou, H.R., Moon, Y., Chung, Y.J., 2004. Cellular and molecular mechanisms forimmune modulation by deoxynivalenol and other trichothecenes: unraveling aparadox. Toxicol. Lett. 15, 61–73.
Peters, G., 1994. The D-type cyclins and their role in tumorigenesis. J. Cell Sci. 18, 89–96.Poirier, M.C., 2004. Chemical-induced DNA damage and human cancer risk. Nat. Rev.
Cancer 4, 630–637.Potapova, O., Gorospe, M., Bost, F., Dean, N.M., Gaarde, W.A., Mercola, D., Holbrook, N.J.,
2000. c-Jun N-terminal kinase is essential for growth of human T98G glioblastomacells. J. Biol. Chem. 275, 24767–24775.
Raick, A.N., 1974. Cell proliferation and promoting action in skin carcinogenesis. CancerRes. 34, 920–926.
Rastogi, S., Dogra, R.K.S., Khanna, S.K., Das, M., 2006. Skin tumorigenic potential of aflatoxinB1 in mice. Food Chem. Toxicol. 44, 670–677.
Rayet, B., Gelinas, C., 1999. Aberrant rel/nfκb genes and activity in human cancer.Oncogene 18, 6938–6947.
Roberts, P.J., Der, C.J., 2007. Targeting the RAF–MEK–ERK mitogen-activated proteinkinase cascade for the treatment of cancer. Oncogene 26, 3291–3310.
Rotter, B.A., Prelusky, D.B., Pestka, J.J., 1996. Toxicology of deoxynivalenol (vomitoxin). J.Toxicol. Environ. Health 48, 1–34.
Roux, P.P., Blenis, J., 2004. ERK and p38 MAPK activated protein kinases: a family of pro-tein kinases with diverse biological functions. Microbiol. Mol. Biol. Rev. 68, 320–344.
Saleem, M., Afaq, F., Adhami, V.M., Mukhtar, H., 2004. Lupeol modulates NF-kappaB andPI3K/Akt pathways and inhibits skin cancer in CD-1 mice. Oncogene 23, 5203–5214.
Sauder, D.N., 1990. The role of epidermal cytokines in inflammatory skin diseases. J.Investig. Dermatol. 95, 27–28.
Saxena, N., Ansari, K.M., Kumar, R., Dhawan, A., Dwivedi, P.D., Das, M., 2009. Patulincauses DNA damage leading to cell cycle arrest and apoptosis though modulation ofBax, p53 and p21/WAF1 proteins in skin of mice. Toxicol. Appl. Pharmacol. 234, 192–201.
Saxena, N., Ansari, K.M., Kumar, R., Chaudhari, B.P., Dwivedi, P.D., Das, M., 2011. Role ofmitogen activated protein kinases in skin tumorigenicity of patulin. Toxicol. Appl.Pharmacol. 257, 264–271.
UNCO
Please cite this article as: Mishra, S., et al., Deoxynivalenol induced mouseAppl. Pharmacol. (2014), http://dx.doi.org/10.1016/j.taap.2014.06.003
ED P
RO
OF
Schothorst, R., Van Egmond, H., 2004. Report from SCOOP task 3.2.10 “collection ofoccurrence data of Fusarium toxins in food and assessment of dietary intake by thepopulation of EU member states” subtask: trichothecenes. Toxicol. Lett. 153,133–143.
Seo, H.J., Park, K.K., Han, S.S., Chung,W.Y., Son, M.W., Kim, W.B., Surh, Y.J., 2002. Inhibitoryeffects of the standardized extract (DA-9601) of Artemisia asiatica Nakai on phorbolester-induced ornithine decarboxylase activity, papilloma formation, cyclooxygenase-2expression, inducible nitric oxide synthase expression and nuclear transcriptionfactor kappa B activation in mouse skin. Int. J. Cancer 100, 456–462.
Shaulian, E., Karin, M., 2001. AP-1 in cell proliferation and survival. Oncogene 20,2390–2400.
Shaw, R.J., Cantley, L.C., 2006. RAS, PI(3)K and mTOR signalling controls tumour cellgrowth. Nature 441, 424–430.
Shifrin, V.I., Anderson, P., 1999. Trichothecene mycotoxins trigger a ribotoxic stressresponse that activates c-Jun N-terminal kinase and p38 mitogen-activated proteinkinase and induces apoptosis. J. Biol. Chem. 274, 13985–13992.
Shishodia, S., Majumdar, S., Banerjee, S., Aggarwal, B.B., 2003. Ursolic acid inhibits nuclearfactor-kappaB activation induced by carcinogenic agents through suppression ofIkappaBalpha kinase and p65 phosphorylation: correlation with down-regulation ofcyclooxygenase 2, matrix metalloproteinase 9, and cyclin D1. Cancer Res. 63,4375–4383.
Silvers, A.L., Bachelor, M.A., Bowden, G.T., 2003. The role of JNK and p38MAPK activities inUVA-induced signaling pathways leading to AP-1 activation and c-Fos expression 1.Neoplasia 5, 319–329.
Slaga, T.J., Fischer, S.M., Nelson, K., Gleason, G.L., 1980. Studies on the mechanism of skintumor promotion: evidence for several stages in promotion. Proc. Natl. Acad. Sci. U. S. A.77, 3659–3663.
Slaga, T.J., Fischer, S.M., Weeks, C.E., Klein-Szanto, A.J.P., Reiners, J., 1982. Studies on themechanisms involved in multistage carcinogenesis in mouse skin. J. Cell. Biochem.18, 99–119.
Surh, Y.J., Kundu, J.K., 2005. Signal transduction network leading to COX-2 induction: aroad map in search of cancer chemopreventives. Arch. Pharm. Res. 28, 1–15.
Thanos, D., Maniatis, T., 1995. NF-kappa B: a lesson in family values. Cell 80, 529–532.Thomas, T., Thomas, T.J., 2003. Polyamine metabolism and cancer. J. Cell. Mol. Med. 7,
113–126.Van Der Fels-Klerx, H.J., Klemsdal, S., Hietaniemic, V., Lindblad, M., Ioannou-Kakouri, E.,
Van Asselta, E.D., 2012. Mycotoxin contamination of cereal grain commodities inrelation to climate in North West Europe. Food Addit. Contam. Part A: Chem. Anal.Control Expo. Risk Assess. 29, 1581–1592.
Verma, A.K., Shapas, B.G., Rice, H.M., Boutwell, R.K., 1979. Correlation of the inhibition byretinoid of tumor promoter-induced mouse epidermal ornithine decarboxylaseactivity and skin tumor promotion. Cancer Res. 39, 419–425.
Vivanco, I., Sawyers, C.L., 2002. The phosphatidylinositol 3-kinase AKT pathway in humancancer. Nat. Rev. Cancer 2, 489–501.
Vojtek, A.B.., Hollenberg, S.M., Cooper, J.A., 1993. Mammalian RAS interacts directly withthe serine/threonine kinase Raf. Cell 74, 205–214.
Warne, P.H., Viciana, P.R., Downward, J., 1993. Direct interaction of RAS and the amino-terminal region of Raf-1 in vitro. Nature 364, 352–355.
WHO Food Additives Series: 47, 1998. Safety evaluation of certain mycotoxins in food.Prepared by 49th Meeting of JECFA. WHO, Geneva.
Yang, G.H., Jarvis, B.B., Chung, Y.J., Pestka, J.J., 2000. Apoptosis induction by the satratoxinsand other trichothecene mycotoxins: relationship to ERK, p38 MAPK and SAPK/JNKactivation. Toxicol. Appl. Pharmacol. 164, 149–160.
Zhang, X.F., Settleman, J., Kyriakis, J.M., Takeuchi-Suzuki, E., Elledge, S.J., Marshall,M.S., Bruder, J.T., Rapp, U.R., Avruch, J., 1993. Normal and oncogenic p21RASproteins bind to the amino-terminal regulatory domain of c-Raf-1. Nature364, 308–313.
Zhou, H.R., Pestka, J.J., 2003. Deoxynivalenol-induced apoptosis mediated by p38 MAPK-dependent p53 gene induction in RAW 264.7 macrophages. Toxicol. 72, 330.
skin cell proliferation and inflammation via MAPK pathway, Toxicol.