dermomyotome development and myotome...
TRANSCRIPT

Windner, et al.
1
Dermomyotome development and myotome patterning require Tbx6
inhibition of embryonic myogenesis
Stefanie E. Windner1,2, Nathan C. Bird1, Sara Elizabeth Patterson1, Rosemarie Doris1,
and Stephen Henri Devoto1*
1Department of Biology, Wesleyan University, Middletown, CT 06459, U.S.A.
phone: (860) 685-3461, Fax: (860) 685-3279.
2 Division of Zoology and Functional Anatomy, Department of Organismic Biology,
University of Salzburg, Salzburg, Austria
Contact: Stephen Devoto
Phone: 860.685.3461
Fax: 860.685.3279
Running title: tbx6 and the specification of the dermomyotome

Windner, et al.
2
SUMMARY
The progenitors of tissue-restricted stem cells must be prevented from undergoing
terminal differentiation during embryogenesis, even while cells around them respond to signaling
and differentiate. Paraxial mesoderm specific progenitors are contained within the epithelial
region of the somite called the dermomyotome. T-box containing transcription factors regulate
many aspects of mesoderm fate including somite patterning. We report that the zebrafish tbx6
ortholog is the fss gene, previously called tbx24. Zebrafish tbx6 is required to prevent premature
differentiation of central dermomyotome precursors. Using transgenic zebrafish expressing
inducible tbx6, we found that the role of tbx6 in dermomyotome specification is not coincident
with its role in segmentation. Our results demonstrate for the first time that a gene required for
segmentation is also required for the maintenance of mesoderm progenitor cell identity.
HIGHLIGHTS
• The zebrafish fused somites gene is the tbx6 orthologue.
• Fss/Tbx6 prevents premature myogenic differentiation.
• The role of Fss/Tbx6 in segmentation is distinct from its role in dermomyotome.

Windner, et al.
3
INTRODUCTION Vertebrate trunk and limb skeletal muscle derives from the paraxial mesoderm, which
also gives rise to the dermis and skeleton of the axial trunk. The subdivision of the paraxial
mesoderm into the precursors to these cell types occurs during the segmentation period of
embryogenesis, with the appearance of the myotome, the dermomyotome, and the sclerotome.
The dermomyotome contains precursors to the embryonic myotome, and to post-embryonic
muscle growth and repair.
In avians and mammals, the earliest myotome cells differentiate after their incorporation
into somites, and at about the same time as the appearance of the dermomyotome [Kahane, 2007
#2899;Denetclaw, 1997 #1535;Venters, 1999 #2855]. In contrast, in most teleosts primary
myotome cells differentiate prior to somite formation and prior to the appearance of the
dermomyotome [Bone, 1978 #442]. In both amniotes and teleosts, signals from tissues
surrounding the somite, including the notochord, spinal cord, and surface ectoderm trigger
myogenic differentiation.
Much less is known about the initial development of the dermomyotome itself, or about
the mechanisms which ensure that the dermomyotome is maintained during early embryogenesis.
In zebrafish, dermomyotome precursors can be first identified in the anterior portion of recently
formed somites [Stellabotte, 2007 #2882;Hollway, 2007 #2883]. These anterior border cells
migrate to the lateral surface of the somite, forming a layer of Pax7-expressing cells on the
surface of the myotome, as in other vertebrates [Devoto, 2006 #2737].
The fused somites (fss) gene is required for the expression of anterior half-somite identity
and the formation of somites throughout the trunk and tail. fss is a member of the tbx gene
family, which encode proteins with a conserved DNA binding motiv known as the T-box.
Several tbx family members are important in embryonic mesoderm development. In mice,

Windner, et al.
4
eomesodermin is required for the initial formation of the mesoderm [Showell, 2004 #3086],
brachyury is required for notochord and posterior mesoderm development [Showell, 2004
#3086], tbx6 is required for paraxial mesoderm development as well as somite patterning
[Chapman, 1998 #2962;White, 2003 #2961], and tbx18 is required for the maintenance of the
anterior identity in newly formed somites [Bussen, 2004 #3076]. Tbx proteins are sequence-
specific DNA binding proteins which can serve as transcriptional repressors or as activators
[Wardle, 2008 #3089].
We identify the fss gene as the zebrafish ortholog of tbx6, and demonstrate that in
addition to its requirement for segmentation, it also is required for the development of part of the
dermomyotome and myotome. Using transgenic fish with a heat-shock inducible tbx6 gene, in
the fss/tbx6 mutant background, we demonstrate that the domain of paraxial mesoderm in which
Tbx6 can rescue dermomyotome is broader than the domain of in which Tbx6 can rescue
segmentation. We propose that fss/tbx6 is necessary for the prevention premature differentiation
of dermomyotome precursors into post-mitotic muscle fibers.

Windner, et al.
5
RESULTS
Identification of the zebrafish tbx6 orthologue
We have re-examined the homology of the fss gene to other members of the tbx gene
family. fss was initially named tbx24 [Nikaido, 2002 #2310], in part because few other teleost
tbx gene sequences were available for comparison and the diversity of the tbx6 subfamily was
not apparent, and in part because there was a previously cloned zebrafish gene which had been
named tbx6 [Hug, 1997 #3092], and in part because no other teleost tbx6 gene sequences were
available for comparison. With further sequences available, it is clear that the Fss amino acid
sequence is most homologous in its T-domain to the Tbx6 subfamily, nesting alongside Tbx6
proteins of other teleosts, amphibians, lizard, and mammal (Figure 1, see Figure S1 for the
cladogram of the entire tree, and Table S1 for list of gene accession numbers). The two nearest
subgroups, Mga and Tbx16, also contain zebrafish genes nested with those of other species. The
originally named zebrafish Tbx6 falls within the Tbx16 subgroup. Thus, we propose to rename
fss/tbx24 to fss/tbx6, and to rename the zebrafish tbx6 to tbx6r.
The chromosomal region containing the zebrafish fss gene has numerous genes with
paralogs of the same genes present near the tbx6 gene in mouse (34 genes), human (69 genes),
and medaka (188 genes) [Catchen, 2009 #3066]. Using the same parameters, the zebrafish gene
previously named tbx6 is has no other genes in common with tbx6 in mouse or human. We
conclude from this analysis of synteny that the zebrafish orthologue of tbx6 is the fss gene.
Tbx6 is required for development of the central dermomyotome
To examine the role of Tbx6 in dermomyotome development, we examined the
expression of pax3, one of the earliest markers of the dermomyotome [Bober, 1994
#3007;Groves, 2005 #2510]. pax3 is expressed in wild type embryos within cells on the anterior

Windner, et al.
6
and central surface of recently formed somites, and in the dorsal neural tube (Figure 2A). The
central domain of early pax3 mRNA expression is less distinct in embryos mutant for fss/tbx6,
while expression in dorsal and ventral domains is unaffected (Figure 2B). The reduction in pax3
mRNA in the central domain is more distinct in the posterior trunk and tail than in the anterior
trunk (data not shown). Pax7 antibody labeling confirms the existence of two separate
dermomoytome subpopulations during early somite patterning. In the posterior trunk and tail,
the dorsal/ventral domain is labeled by Pax7 antibody earlier and independently of tbx6/fss
function, while the central domain is identifiable later and is dependent on fss/tbx6 (Figures 2C
and 2D). The paraxial mesoderm equivalent to somites 1 to 7 do not show any deficit in the
dermomyotome in fss/tbx6 mutant embryos. In these somites, the central domain of Pax7
expression develops at a similar rate in wild type and mutant embryos (Figure 2E and 2F).
Expression of myogenic regulatory factors (MRFs) is more widespread in fss/tbx6
mutants. In wild type embryos, MRF genes are expressed segmentally, in only the posterior of
recently formed somites. We could not detect Myogenin-negative cells in the paraxial mesoderm
of fss/tbx6 mutant embryos, suggesting that dermomyotome precursors differentiate prematurely
into primary myotome cells (Figures 2G and 2H).
The deficit in the central dermomyotome in mutant embryos at the end of the
segmentation period (24h) remains regionally restricted and quite pronounced; the central Pax7+
dermomyotome cells are absent in all of the posterior trunk and tail (Figures 3A and 3B),
beginning in a region ranging from the equivalent of somite 6 to somite 8 of wild type siblings
(n=25). Within the affected region, the loss of Pax7+ cells in the dermomyotome correlates with
a significantly higher proportion of myogenic (Mef2+) nuclei [Shinin, 2006 #3010] (Figure 3C, p
≤ 0.0002). The dorsal/ventral dermomyotome cells are present in mutants in approximately

Windner, et al.
7
normal numbers (Figures 3D and 3E).
Tbx6 is required for myotome patterning
The premature differentiation of dermomyotome progenitors in fss/tbx6 mutant embryos
leads to long term defects in myotome patterning. At 24h, the wild type zebrafish myotome
consists of deep fast muscle fibers, covered laterally by a continuous monolayer of slow muscle
fibers [Waterman, 1969 #330]. This slow muscle monolayer extends from the lateral surface of
the myotome to the notochord, at the position of the future horizontal myoseptum, and separates
the fast muscle fibers into a dorsal and a ventral group (Figures 4A-C). In mutant embryos, the
myotome is dramatically disrupted in the same region where dermomyotome cells have
prematurely differentiated into muscle fibers, the continuity of the slow muscle monolayer is
broken, and the central, medial-most slow fibers are not in contact with the superficial slow
fibers (Figures 4F-H). Moreover, there are numerous fast muscle fibers superficial to the slow
fibers in the central region. These superficial fast fibers can span several segment lengths, have
very large cross sectional diameters, and have an unusually large number of nuclei, with several
present in transverse sections (arrows, Figures 4D and I). Despite the disrupted myotome
patterning, central slow and superficial fast muscle fibers in mutants express Engrailed, like
muscle pioneers and lateral fast in wild type embryos (Figures 4E and J). [Hatta, 1991
#1789;Wolff, 2003 #2321].
At 24h, fss/tbx6 mutant embryos have fewer than half the number of dermomyotome cells
per section through the posterior trunk (Figure 4K). In wild type embryos, the number of Pax7+
dermomyotome cells decreases from 24h to 48h, as some of the central domain cells differentiate
into muscle fibers. In mutant embryos, the number of dermomyotome cells recovers to near wild
type levels by 48h, and they are distributed throughout the dorsal ventral extent of the myotome

Windner, et al.
8
(Figure 4L). This suggests the possibility that the dermomyotome is able to regenerate under
certain circumstances.
The increased number of myonuclei in mutant embryos at 24h (Figure 3C) does not lead
to an increased number of muscle fibers, but rather to an increased number of nuclei per fiber in
the ectopic fast fibers. (Figure 4M). However, fss/tbx6 mutants show delayed hyperplastic
growth: at 48h there are fewer fast muscle fibers in mutants compared to wild type siblings. We
presume this reflects the reduced number of myogenic precursor cells, with a 24h delay. At no
stage do we see a difference in the number of slow muscle fibers, which at these early
developmental stages are not derived from the same population of cells that give rise to the
dermomyotome [Devoto, 1996 #2564] (data not shown). Unusually large fast muscle fibers
remain in the central domain of the myotome of fss/tbx6 mutants at 72h, and even later into
larval stages (data not shown), but new fibers, developing on the outside of the embryonic fast
myotome [Rowlerson, 2001 #1842], have diameters similar to those in wild type (Figure 4N).
Differential rescue of dermomyotome and segmentation by inducible Tbx6
Regular oscillations in the expression of genes such as her1 in the presomitic mesoderm
are an important part of the segmentation process. Tbx6 is required for the maintenance of these
oscillations as cells become part of the anterior presomitic mesoderm [Nikaido, 2002 #2310].
This has led to the suggestion that fss/tbx6 may act at the somite determination front, which in
zebrafish is 4-5 somite lengths posterior to the most recently formed somite [Sawada, 2001
#2829;Holley, 2002 #2309].
To test whether Tbx6 is required in this same zone of paraxial mesoderm for
dermomyotome, we created transgenic fish expressing inducible a heat shock inducible tbx6
gene. We assayed the early response to Tbx6 induction by examining the expression of ripply1

Windner, et al.
9
and fgf8, which are expressed in a Tbx6-dependent manner in the anterior of newly formed
somites, and the expression of myoD, whose expression is restricted from the anterior of newly
formed somites in a Tbx6-dependent manner. We found that a pulse of Tbx6-myc during the
segmentation period restored expression of ripply1 and fgf8 in a partly segmented manner, and
restored gaps in the myoD expression domain (Figure 5).
The induced expression of Tbx6 also restored proliferation within the somite of mutant
embryos. In wild type embryos during the segmentation period, most proliferative cells are on
the surface of the somite, with the highest number at the dorsal and ventral extremes [Barresi,
2001 #1972;Stellabotte, 2007 #2882]. The loss of fss/tbx6 has little effect on proliferation at the
dorsal and ventral extremes, but it leads to the complete absence of proliferative cells in the
central region (Figure 6A). Pulsed expression of Tbx6 restored proliferative cells in the central
region of the somite, consistent with a rescue of the central dermomyotome (Figure 6A). Pulsed
expression of Tbx6 rescued the number of dermomyotome cells in mutant embryos. In the in the
region of the trunk with maximal numbers of induced Pax7+ cells, the rescue was quantitatively
to the levels found in fss/tbx6 heterozygous embryos (Figure 6B).
To further characterize the temporal requirement for fss/tbx6, we induced a pulse of Tbx6
expression at varying times during the segmentation period, and monitored both segmentation
and the development of the dermomyotome. We found that a Tbx6 pulse rescued the formation
of 1 to 3 remarkably normal somites (Figure 6C), along with an 8-10 somite length region of
rescued central dermomyotome. The best somite was consistently formed about 6 somite-
lengths posterior to the somite which would have been forming at the time of heat shock. For
example, if we heat shocked a clutch in which the fss heterozygous embryos were at the 10S
stage, we consistently observed that fss-/-;hsp70:tbx6 embryos formed at least one complete

Windner, et al.
10
segment in the region of the embryo equivalent to somite 15 to 16 (Figures 6C-F). This spatial
pattern is consistent with a role for Tbx6 in the somite determination front. Pax7+
dermomyotome cells in the central region of the somite were rescued from a region which
extended from 1-2 somite lengths posterior to the rescued segment, to about 8 segment lengths
anterior to it (Fig. 4c-g). We conclude that the role of Tbx6 in segmentation is at least partially
distinct from the role of Tbx6 in dermomyotome development.

Windner, et al.
11
DISCUSSION Our work demonstrates that fss/tbx6 is required for the initial development of the central
portion of the dermomyotome in the posterior trunk and tail. The requirement for fss/tbx6 in
central dermoyotome development is separate from the requirement for fss/tbx6 in segmentation,
as a pulse of fss/tbx6 can rescue dermomyotome without rescuing segmentation.
Regionalization of the paraxial mesoderm
Our work subdivides the zebrafish dermomyotome into at least two different populations.
In the anterior 6-8 somites, of the dermomyotome develops independently of tbx6 function,
while in the remainder of the trunk and in the tail, the dermomyotome can be subdivided into a
tbx6-independent dorsal and ventral dermomyotome, and a tbx6-dependent central
dermomyotome. The segmentation process in the anterior 6-8 somites is genetically distinct from
the segmentation process in the more posterior somites [Holley, 2006 #2817;Holley, 2007
#2917]. In fact, fss is the only gene whose mutant does not show a regionally restricted
segmentation deficit. Thus it was a surprise to find that the dermomyotome defect in fss is
restricted to somites posterior to somites 6-8. This suggests that dermomyotome development,
like the segmentation process, is genetically distinct in somites 6-8.
The dermomyotome posterior to somite 7 can be subdivided into a dorsal/ventral domain
developing independently of tbx6, and a central domain dependent on tbx6. Whether this
genetically based subdivision indicates a separate origin or fate of these two populations of
dermomyotome cells remains to be determined.
•The Tbx6 requirement for dermomyotome
As paraxial mesoderm cells mature prior to becoming incorporated into somites, they
become competent to differentiate into muscle, and many of them do so in response to signals

Windner, et al.
12
from surrounding tissues. The known myogenic regulatory signals are derived from notochord,
surface ectoderm, neural tube, and the pronephros; these signals are distributed uniformly along
the anterior-posterior axis. We propose that Tbx6 acts as a segmentally distributed inhibitor of
primary myogenesis in the anterior presomitic mesoderm, and that this inhibition is required for
the development of the dermomyotome (Figure 7). In this model, one role of Tbx6 is to protect
the anterior border cells from the wide-spread growth factors which would otherwise trigger their
differentiation into primary myotome.
Uniform expression of Tbx6 does not prevent myogenesis in the posterior of newly
formed somites, suggesting that Tbx6 is not sufficient for this effect. Previous work in zebrafish
and xenopus indicates that Tbx6 is transformed from an activator to a repressor of transcription,
by the Ripply protein. Ripply is a Tbx6-interacting protein which recruits members of the
Groucho family of Histone de-acetylases, and converts Tbx6 from an activator to a repressor. In
zebrafish, Ripply is expressed in the anterior of future somites; raising the possibility that Ripply
is a necessary co-factor for the effect of Tbx6 in dermomyotome development.
Evolutionary Divergence of tbx6
Orthologous genes in zebrafish and mouse usually carry out similar functions, reflecting
the role of the gene in the last common ancester of these two species. Thus, it is surprising that
the loss of tbx6 function causes distinct phenotypes in mouse and zebrafish. Wherease in
zebrafish it leads to the loss of segmentation and the central dermomyotome, in mouse it leads to
the development of an enlarged tailbud, the transformation of paraxial mesoderm into neural
tube, and the loss of laterality in the node [Hadjantonakis, 2008 #3096; Chapman, 1998 #2962].
The zebrafish spt/tbx16 gene is expressed earlier in the maturation of the presomitic
mesoderm, and when mutant leads to the development of an enlarged tailbud and the loss of

Windner, et al.
13
laterality in zebrafish equivalent of the node [Ho, 1990 #1452; Gourronc, 2007 #3095],
phenotypes with similarity to the mouse tbx6 knockout phenotypes. Although the tbx16 gene is
present in non-mammalian tetrapods, it has apparently been lost in mammals (Figure 1). If tbx6
in mammals has acquired the combined functions of the tbx6 and tbx16 genes, then the loss of
tbx6 would lead to a more severe phenotype in mouse than in zebrafish. Consistent with this
interpretation, partial loss of tbx6 in mouse has similar consequences to the complete loss of tbx6
in zebrafish—a disruption of segmentation. In particular, partial loss of tbx6 leads to the loss of
anterior somite compartment identity, as indicated by the expression of tbx18 [White, 2003
#2961], and the appearance of fused ribs and vertebrae.
The development of the dermomyotome has not been examined in mouse embryos
mutant for tbx6.

Windner, et al.
14
FIGURES AND LEGENDS
Figure 1. Cladogram of Tbx6, Tbx16 and MGA members of the Tbx protein family.
Tree is an internal branch taken from a neighbor-joining tree of the Tbx family of proteins based solely on their T-Box domain. Numbers represent bootstrap values (100 replications, values >50 shown). Sequence alignment using ClustalX 2.0 [Larkin, 2007 #3069], and tree building using SeaView 4.2.12 [Gouy, 2010 #3094]. Note that zebrafish Fss/tbx6 (red) nests within the tbx6 clade.

Windner, et al.
15
Figure 2. Tbx6 prevents myogenic differentiation of central dermomyotome precursors during segmentation stages. (A, B) pax3 expression in heterozygote (A) and fss/tbx6 mutant (B) at 15-16S stage (insets show
higher magnification). Segmented pax3 expression in the central trunk is disrupted in mutant embryos.
(C, D) Pax7 (green) and MF20 (red, Myosin Heavy Chain) expression in heterozygote (c) and fss/tbx6 (d) embryos at 20S stage (insets show higher magnification). In the posterior trunk Pax7 is initialized in cells of the dorsal and ventral dermomyotome; central Pax7 expression is

Windner, et al.
16
delayed in heterozygote (C), but absent in fss/tbx6 mutant (D). (E-H) Pax7 (green), Myogenin (red, Mgn), and MF20 (white) expression in the anterior trunk (E,
G) and on optical horizontal sections through the posterior trunk (F, H) of heterozygote (E, F) and fss/tbx6 mutant (G, H) at 13-14S stage. Heterozygote embryos express all three markers within distinct compartments of the somites (dashed lines in F). In fss/tbx6 mutants, Pax7 expressing central dermomyotome cells are restricted to the anterior trunk; the central posterior trunk shows more cells expressing Myogenin.
Scale bars: 50 µm (A, E, F), 100 µm (C).
Figure 3. Tbx6 is required for the central dermomyotome of the posterior trunk and tail.
(A, B) Tracing of Pax7+ dermomyotome nuclei in representative 24h wt (A) and fss/tbx6 (B) embryos. Note that the central dermomyotome deficit/defect in fss/tbx6 mutants starts at trunk levels correlating to somite 7 in wt (dashed line). Insets show Pax7+ (green) and myogenic Mef2+ (red) cells at anal vent level.
(C) Graph showing average of Pax7+, Mef2+ and differentiating (Pax7+/Mef2+) cells (±SEM) on sections at anal vent level in heterozygote and fss/tbx6 mutant.
(D, E) Cross-sections and graphs showing dorsal-ventral (DV) distribution of dermomyotome (average of Pax7+ cells ± SEM) and % differentiating dermomyotome (Pax7+/Mef2+) in heterozygote and fss/tbx6 mutant (green, Pax7; red, Mef2; blue, Hoechst 33258).
Scale bars: 100 µm (a), 50 µm (d).
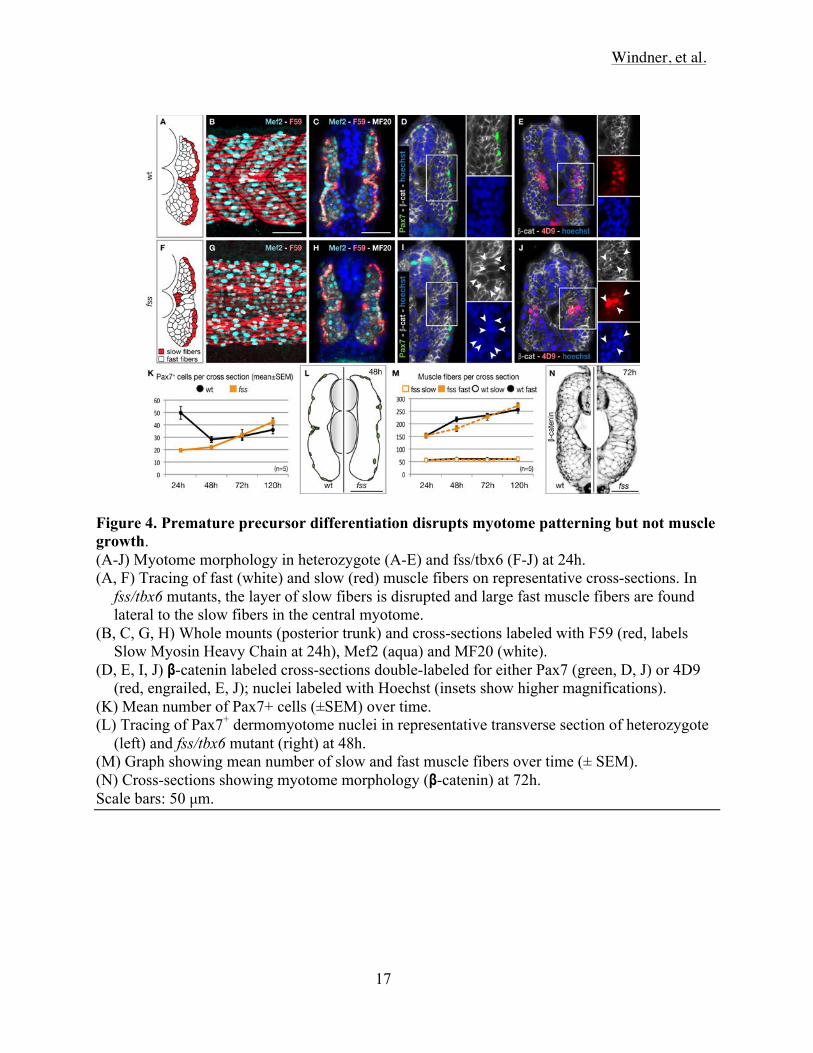
Windner, et al.
17
Figure 4. Premature precursor differentiation disrupts myotome patterning but not muscle growth. (A-J) Myotome morphology in heterozygote (A-E) and fss/tbx6 (F-J) at 24h. (A, F) Tracing of fast (white) and slow (red) muscle fibers on representative cross-sections. In
fss/tbx6 mutants, the layer of slow fibers is disrupted and large fast muscle fibers are found lateral to the slow fibers in the central myotome.
(B, C, G, H) Whole mounts (posterior trunk) and cross-sections labeled with F59 (red, labels Slow Myosin Heavy Chain at 24h), Mef2 (aqua) and MF20 (white).
(D, E, I, J) β-catenin labeled cross-sections double-labeled for either Pax7 (green, D, J) or 4D9 (red, engrailed, E, J); nuclei labeled with Hoechst (insets show higher magnifications).
(K) Mean number of Pax7+ cells (±SEM) over time. (L) Tracing of Pax7+ dermomyotome nuclei in representative transverse section of heterozygote
(left) and fss/tbx6 mutant (right) at 48h. (M) Graph showing mean number of slow and fast muscle fibers over time (± SEM). (N) Cross-sections showing myotome morphology (β-catenin) at 72h. Scale bars: 50 µm.

Windner, et al.
18
Figure 5. A pulse of Tbx6 expression in fss/tbx6 mutants induces segmental expression of ripply1 and fgf8, and locally represses myod. Panels showing expression of genes expressed specifically in the anterior (ripply1, fgf8) and
posterior (myod) somite compartments in non-tg and tg(hsp70:tbx6) heterozygote and fss/tbx6 mutant. Embryos were heat shocked at 9-10S stage, fixed at 13-14S (myod) and 15-16S stage (ripply1, fgf8), and flat-mounted for documentation.
Scale bar: 50 µm.

Windner, et al.
19
Figure 6. Induced Tbx6 expression differentially rescues central dermomyotome and segmentation. (A) Distribution of proliferative (pH3+) cells in non-tg and tg(hsp70:tbx6) heterozygote and
fss/tbx6 mutant embryos 8 hours after heat-shocking at 8S. Data are presented as mean ± SEM.
(B) Graph showing number of Pax7+ cells in non-tg and tg(hsp70:tbx6) heterozygote and fss/tbx6 mutant embryos at 10 hours after heat-shocking at 10S.
(C) Schematic showing the location of rescued dermomyotome (light green), segments (dark green) in individual embryos after heat shocks at various times (orange). Note that a Tbx6 pulse rescues the formation of 1-3 somite boundaries and central dermomyotome over a more extensive 8-10 somite lengths.
(D-F) Rescue of dermomyotome and segmentation in fss/tbx6 mutant tg(hsp70:tbx6) visualized using F59 (red) and MF20 (white) (D) and Pax7 (green) and Mef2 (white) (E) labeling, and DIC imaging (F). Specimen stages are 5d (D, F) and 24h (E), rescued segments are numbered according to the corresponding region in wild-type siblings.
Scale bars: 100 µm (D, E), 50 µm (F)

Windner, et al.
20
Figure 7. Schematic representation of Tbx6 function.

Windner, et al.
21
EXPERIMENTAL PROCEDURES
In situ hybridization and immunocytochemistry. RNA in situ hybridization and immunohistochemistry was carried out as previously
described [Barresi, 2000 #1988]. Embryos in Figures 1e and f are shown anterior to the top. All other embryos are shown dorsal to the top, and for whole mount labeling, anterior to the left. Imaging of whole mount labeling was with a Zeiss LSM510 confocal microscope, individual optical sections were flattened for each image, except for Figure 1e and f, which show single optical sections. Black and white were inverted in Fig. 3n for clarity of presentation. Tracings of individual labeled nuclei (Figures 2a, 2b, and 3f) were done on computer projections.
Zebrafish, transgenesis, and heat shock. All experiments were done on zebrafish from the Wesleyan University strain of wild type
fish, and the te314a allele of fss/tbx6, which behaves as a null [Nikaido, 2002 #2310]. Embryos were cared for using standard procedures [Westerfield, 1995 #745] .
The full open reading frame of zebrafish tbx6 was amplified by PCR from a cDNA kindly provided by Scott Holley, with Gateway recombination sites added at each end. The PCR product was recombined into the donor vector pDONR221 to make the plasmid pME-TBX6. pME-TBX6 was recombined with the 5’ entry clone containing the zebrafish hsp70promoter (p5E-hsp70), and with the 3’ entry clone containing 6x myc tag plus the SV40 late poly adenylation signal (p3E-MTpA), and a destination vector with tol2 sites and egfp under the control of the cardiac myosin light chain 2 promoter (pDestTol2CG2). The resulting plasmid (RAD510) is hsp70-tbx6-myc.
We also recombined pME-TBX6 with p5E-hsp70, a 3’ entry clone containing cherry (p3Emcherry), and (pDestTol2CG2. This resulting plasmid (RAD521) is hsp70-tbx6-cherry.
Single cell fss/tbx6 mutant embryos were injected with hsp70-tbx6-myc or hsp70-tbx6-cherry and tol2 mRNA, and grown to adulthood. A founder was identified on the basis of cardiac eGFP expression in its offspring. These offspring were raised to adulthood and outcrossed to fss/tbx6 homozygous mutants. All crosses generated 50% transgenic embryos, suggesting the presence of only one transgene insertion. All of the shown experiments were done on the v8 allele of tg(hsp70:tbx6-myc), but we obtained qualitatively similar resuts using tg(hsp70:tbx6-cherry), the allele we used is v7.
Embryos were heat shocked at the indicated time by transferring them in small mesh-bottom wells to embryo medium pre-warmed to 37º for one hour.